1. Introduction
Biomanufacturing is a type of manufacturing that utilizes biological systems (e.g., living microorganisms, resting cells, plants, animals, cells, tissues, enzymes, cascade enzymes (i.e., enzyme cocktails, one-pot multi-enzyme system), or
in vitro synthetic enzymatic systems) to produce value-added products for use in the agricultural, food, energy, material, and pharmaceutical industries [
1,
2,
3]. Its products may also be isolated from natural sources, such as blood, cultures of microbes, animal cells or organs, or plants grown in specialized equipment or dedicated cultivation environments [
4,
5]. The enzymes, cells, tissues, animals, or plants used may be natural or modified by protein engineering, genetic engineering, metabolic engineering, and synthetic biology [
2,
3,
6,
7].
To benchmark the revolutions from Industry 1.0 to Industry 4.0 [
8,
9,
10], biomanufacturing was proposed to be classified to four generations in terms of respective product types (e.g., primary metabolite, secondary metabolite, biomacromolecule, tissue, etc.), production technologies (e.g., solid state fermentation, anaerobic liquid fermentation, aerobic submerged fermentation, cell cultures, enzymes, etc.), and research technologies (e.g., isolation of new microorganisms, mutated microorganisms, recombinant DNA technology, stem cells, directed evolution of enzymes, etc.) [
3]. Industrial biomanufacturing (i.e., Biomanufacturing 1.0) started from acetone-butanol-ethanol (ABE) fermentation with foci on the production of primary metabolites (e.g., ethanol, citric acid, acetone, butanol, amino acids, etc.) by using mono-culture fermentation [
11,
12,
13]. Biomanufacturing 2.0 started from penicillin fermentation in World War II with foci on the production of secondary metabolites (e.g., penicillin, streptomycin) by using a dedicated microorganism mutant and aerobic submerged liquid fermentation [
12,
14,
15,
16]. Biomanufacturing 3.0 started in 1970s with foci on the production of large-size biomolecules proteins and enzymes (e.g., erythropoietin, insulin, growth hormone, amylase, DNA polymerase, restriction enzymes) by using recombinant DNA technology and advanced cell culture [
17,
18,
19,
20,
21]. In this century, several most important challenges of humankind, such as, food security [
22,
23], climate change [
24,
25], energy security and sustainability [
26,
27], as well as the energy, food, and water nexus [
28,
29,
30,
31] motivate to develop disruptive biomanufacturing platforms (called Biomanufacturing 4.0).
The great needs always motivate to develop new tools and disciplines. For example, the First World War accelerated the industrialization of the Haber ammonia synthesis [
32] and the biomanufacturing of ABE fermentation for German and Great Britain, respectively [
20,
33]. Disruptive innovations can drive rapid and adaptive change in terms of new market and value network, and eventually disrupt an existing market and displace established market leaders and alliances [
34,
35]. Such rare innovations are often being driven by paradigm-shifting concept or theory, novel research tools, and game-changing production methods, as evidenced in Industrial Revolutions [
36].
According to a dimension—price and mass basis of targeted products as well as their market sizes, biomanufacturing can be classified to biocommodities and high-value products [
37]. Biocommodities are biobased bulk chemicals with low selling prices ranging from less than 10 Chinese Yuan (CNY) to 150 CNY per kilogram, such as biofuels (e.g., ethanol, butanol, hydrogen), sweeteners (e.g., sucrose, high fructose corn syrup), organic acids (e.g. lactic acid, succinic acid), and so on [
37,
38]. Also, the biotechnology for the production of biocommodities was often named as White Biotechnology or industrial biotechnology [
39]. This is distinct from Red Biotechnology, which is used for the production of high-value medical products whose prices range from thousands to billions of CNY per kilogram, such as protein drugs, antibodies, chiral compounds, taxol, antibiotics, and so on [
37,
40]. A mass basis market size of a single biocommodity often exceed that of a pharmaceutical product by approximately 10 orders of magnitude [
37]. White biotechnology and red biotechnology are completely different at multiple levels, including economic driving force, importance of feedstock price, processing cost, capital investment, scale of application, and feedstock availability [
37].
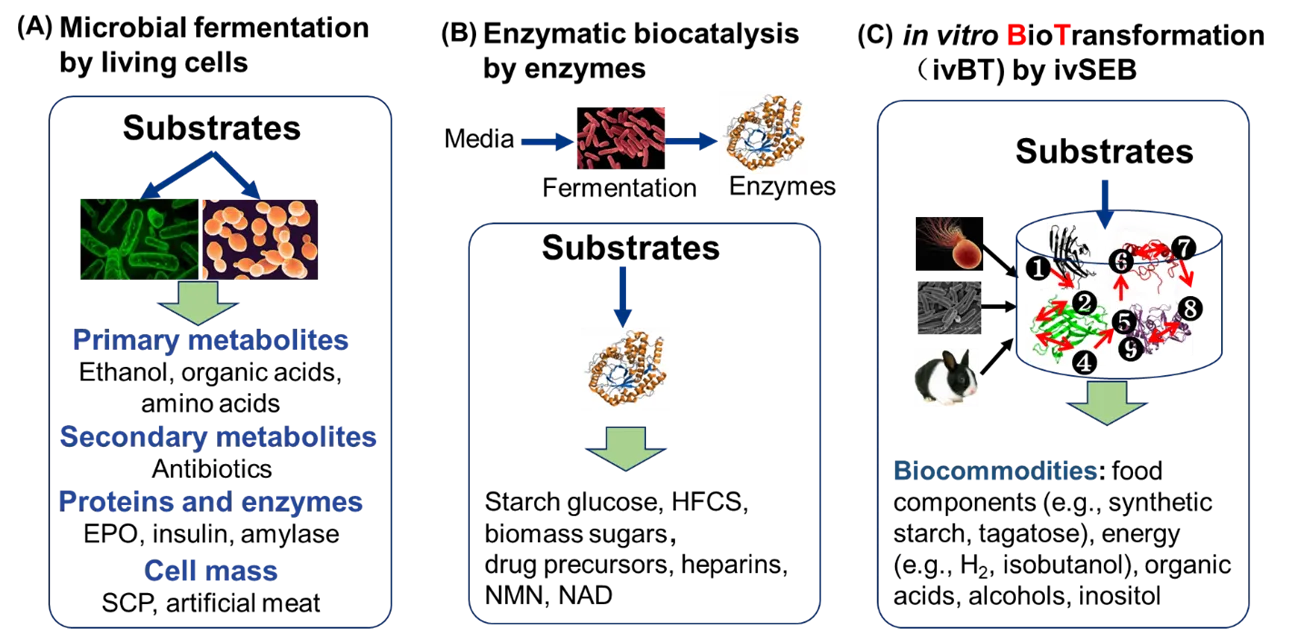
According to another dimension—biocatalysts used, here we suggest that biomanufacturing are classified to three platforms: microbial fermentation by cells [
15], enzymatic biocatalysis [
7], and
in vitro biotransformation (ivBT) by
in vitro synthetic enzymatic system (ivSEB) () [
3,
38,
41,
42,
43,
44]. Microbial fermentation can produce more than 10,000 commercial products, including primary metabolites (e.g., ethanol, butanol, amino acids, organic acids, etc.), secondary metabolites (e.g., antibiotics), proteins and enzymes (e.g., erythropoietin, insulin, industrial enzymes (e.g., α-amylase, glucoamylase, glucose isomerase, cellulase, proteinase, phytase, etc.), and cell mass (e.g., single cell protein (SCP), artificial meat) [
15]. Enzymatic biocatalysis with enzymes made from microbial fermentation can convert feedstock to bulk biochemicals (e.g., glucose hydrolyzed from starch, biomass sugars (glucose and xylose) hydrolyzed from lignocellulosic biomass, high-fructose corn syrup (HFCS)) and high-value products (e.g., drug precursors, nicotinamide dinucleotide (NAD), nicotinamide mononucleotide (NMN), heparins) [
7,
45]. The third platform is ivBT mediated by ivSEB that is independent of basic functions of living cells, such as self-duplication and basic metabolisms [
38,
41]. ivBT has achieved several important biochemical reactions that microorganisms cannot do before. For example, natural or engineered anaerobic hydrogen-producing microorganisms cannot produce more than four molecules of hydrogen per mole of glucose, called the Thauer limit [
46]. It means that only one-third of chemical energy of glucose is converted to hydrogen while two-thirds are converted to acetate. In contrast, the ivSEB can generate 12 moles of hydrogen from one glucose unit of starch and water [
47,
48]. A second example is the
in vitro biotransformation of cellulose to starch, both of which are macromolecules that cannot cross the membrane of living cells. Natural cellulolytic microorganisms can hydrolyze cellulose, assimilate soluble sugars, and accumulate intracellular glycogan (i.e., animal starch) [
49]. However, such ex vitro cellulose hydrolysis and in vivo starch synthesis are isolated by a cellular membrane, leading to very low product yield and slow reaction rate [
49,
50]. In contrast, an ivSEB has been assembled to achieve high-yield and low-cost biotransformation of cellulose to starch [
50,
51].
In this review, we attempt to define the concepts of ivBT as a biomanufacturing platform and ivSEB as an ultra-high-efficiency new biocatalyst that is different from fundamental research tools (such as
in vitro enzymology, cell-free protein synthesis), revisit ivBT’s design principle, present its representative examples for indepth understanding, highlights its biomanufacturing advantages as compared to the other two biomanufacturing platforms, as well as present its remaining challenges and opportunities. We strongly believe that ivBT mediated by ivSEB would be a disrupt biomanufacturing platform, especially for biocommodities, opening markets worth tens of trillions of CNY, such as the carbon-neutral renewable energy system [
30,
52], artificial food and feed [
50,
51,
53], alcohols [
54], healthy sweeteners [
55,
56,
57], organic acids, amino acids, and so on.
. Biomanufacturing platform classification based on biocatalysts used, microbial fermentation by whole cells (A), enzymatic biocatalysis by enzymes (B), and in vitro biotransformation (ivBT) by in vitro synthetic enzymatic biosystems (ivSEB) (C).
2. ivBT and ivSEB
Humankind is confronting great challenges for the coming sustainability revolution [
30,
58]. We believe that great needs of renewable energy system and food security result in the birth of the third biomanufacturing platform—ivBT, which would meet these two great needs better than living whole-cells.
2.1. Definitions
ivBT mediated by ivSEB is the biomanufacturing of biocommodities by the
in vitro reconstruction of artificial (nonnatural) metabolic pathways with numerous natural enzymes, artificial enzymes, (biomimetic or natural) coenzymes, or (artificial) membrane or organelles. Its design philosophy are (1) reductionism, that is, an intellectual and philosophical position that interprets complexity of living cells as a sum of their parts (e.g., enzymes, coenzymes, and membranes) and (2) need-directed integrated innovation, that is, the most important and urgent needs motivate to conduct intensive R&D innovations by integrating multi-disciplinary experts.
2.2. Design Principles
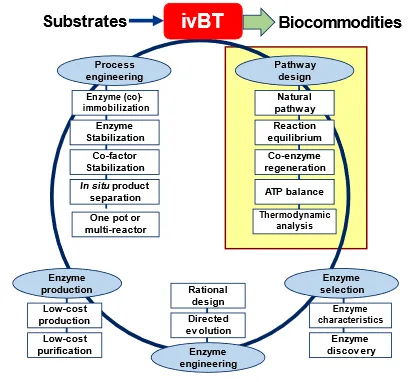
In 2010, Prof. Zhang proposed the development cycle of ivBT which can convert substrates to biocommodities [
38,
41] (). It is composed of five parts: (i) pathway design, (ii) enzyme selection, (iii) enzyme engineering, (iv) enzyme production, and (v) process engineering [
38]. The whole ivBT processing can be improved in an iterative manner, requiring an integrated innovation from many specialists in different areas. Systems engineering principles should be applied for setting priorities for improvements of ivBT, a detailed understanding of the costs and tech improvement potentials of each step [
38].
The design of an
in vitro synthetic enzymatic pathway does not rely on cellular metabolism and bioenergetics, surpassing living whole-cells [
41]. This pathway is usually designed based on natural metabolic pathways with some necessary modifications. First, it is important to design the coenzyme-balanced pathways because it is pretty costly to regenerate coenzymes (e.g., ATP and reduced NAD(P)H)
in vitro. What is even better, it might be better to design coenzyme-free pathways (e.g., starch-to-rare sugars cases). Second, it is important to check reaction equilibria for each reaction, ensuring the rapid reaction rates, no deadly rate-limiting steps, and high product titers. For example, very low reaction equilibria of the CO
2-to-formate reaction and the formate-to-formaldehyde reaction powered by NADH, being 0.003 and 8.5 × 10−10, respectively, results in very low titers of the desired products. It is preferred that the consolidation of a series of reversible reactions with the last irreversible reactions leads to high product yields, for example, the inositol-synthesis case [
59,
60]. Third, it is economically prohibitive to regenerate
in vitro ATP by the consumption of costly substrates, such as ATP, creatine phosphate, phosphoenolpyruvate, pyruvate [
38,
61]. In contrast, these
in vitro ATP regeneration methods are widely used in cascade enzyme biocatalysis (CEB) [
61,
62]. Careful pathway design is highly recommended to ensure ATP balanced. For example, the generation of ATP number from one molecule of glucose to two molecules of pyruvate can range from zero to four, depending on the pathway chosen [
61]. If net ATP is accumulated in the overall process, careful addition of ATPase (F0 part), phosphatase or arsenate would dissipate some ATP [
63] or hydrolyze high-energy phosphate bond-containing metabolites [
64]. If a large amount of ATP is needed as input, the cell-free oxidative phosphorylation may be a future choice [
65]. Fourth, it is essential to keep NAD(P) consumption and NAD(P)H supply balanced. NAD is preferred to NADP because the former is less costly and more stable. When net NAD(P)H is accumulated in the
in vitro biosystem, excess reducing powers can be removed by addition of hydrogenase [
66], cogeneration of electricity through enzymatic fuel cells [
67] or oxidation to water by water-forming NADH oxidase [
68]. If net NAD(P)H input is required, NADH can be regenerated by hydrogenase with H2 [
69], by electricity [
70], other NAD(P)H donors, such as formate [
71], methanol [
72], glucose [
72], propanol [
73], phosphite [
74], glucose-6-phosphate by one enzyme [
75] or through the modified pentose phosphate pathway [
76].
. The development cycle of ivBT for biomanufacturing, modified from Ref. [
20].
In principle, enzymes are chosen based on its chemical reaction and substrate specificity. In addition, enzymes with high substrate selectivity, high catalytic efficiency, low product inhibition, and high stability are highly desired. Although enzymes are well-known for their high selectivity, some enzymes with some promiscuous activities may not be good for ivBT. For example, most phosphatases usually have low substrate specificities [
57,
77,
78]. In the starch-to-mannose case, the discovery and engineering of high-selectivity mannose 6-phosphatase is essential to high product yields [
79]. Because different enzymes may have different optima for pH, temperature, and ionic strength, and may require different metal ions (e.g., Mg
2+, Mn
2+) as activators, trade-offs have to been made among them [
80,
81]. Therefore, it might be preferable that as many enzymes as possible be obtained from the same microorganism because they may have similar optimal working conditions, and they may form enzyme complexes to allow metabolite channeling [
82,
83,
84,
85].
Furthermore, natural enzymes can be engineered by rational design, directed evolution, and their combination [
86,
87,
88]. Several properties are important for ivBT, such as specific activity [
57,
89,
90], thermostability [
60,
89,
91,
92,
93,
94,
95], substrate selectivity [
55,
57], coenzyme preference from NADP to NAD to NMN to other biomimics [
44,
96,
97,
98,
99,
100], optimal pH shift [
101,
102,
103,
104], etc.
Costs of enzymes in terms of CNY/unit or CNY/kg dry enzyme are strongly related to whether industrial biomanufacturing is economically viable or not [
3,
105]. Most academic researchers might have wrong impressions about it, mislead by the difficulty in the preparation of milligram-level recombinant enzymes in their labs or very costly commercial biological science enzymes purchased by labs. Indeed, large-scale industrial enzyme production costs (e.g., α-amylase, protease, cellulase, phytase) were as low as 100 CNY per kg of dry protein weight [
3,
20,
105]. The price ratio of industrial enzymes to commercial biological science enzymes in terms of mass could be in a range of 1,000,000 to 1,000,000,000 [
51,
105]. More details how to make ultra-low-cost enzymes is discussed in Section 4.
Lastly, process engineering includes enzyme immobilization [
106], multi-enzyme co-immobilization [
107], enzyme stabilization, coenzyme stabilization [
108],
in situ product separation [
109], reactor engineering (one pot or multi-pot cascade, microreactor, continuous stirred tank reactor or plug flow reactor) [
110,
111], and so on. Besides widely-known enzyme- and coenzyme-immobilization techniques, it was found that a simple mixture of multiple enzymes at high concentrations could greatly co-stabilize their lifetimes possibly due to macromolecular crowding effects [
112,
113,
114].
in situ product removal could be very important for some products whose reaction equilibria is low (i.e., a low product titer of fructose 1,6-bisphosphate [
80]) or whose product inhibition is strong (e.g., the starch-to-mannose case).
In situ removal of the desired product facilitates high product yield and volumetric productivity [
47,
115]. In most cases, it is preferred and simple to consolidate the whole pathway in one reactor. But it might have special advantages to separate cascade reactors in multiple reactors or apply a continuous-flow microreactor for special biomanufacturing advantages [
110,
111].
2.3. Comparison with Other Seemingly-like Technologies
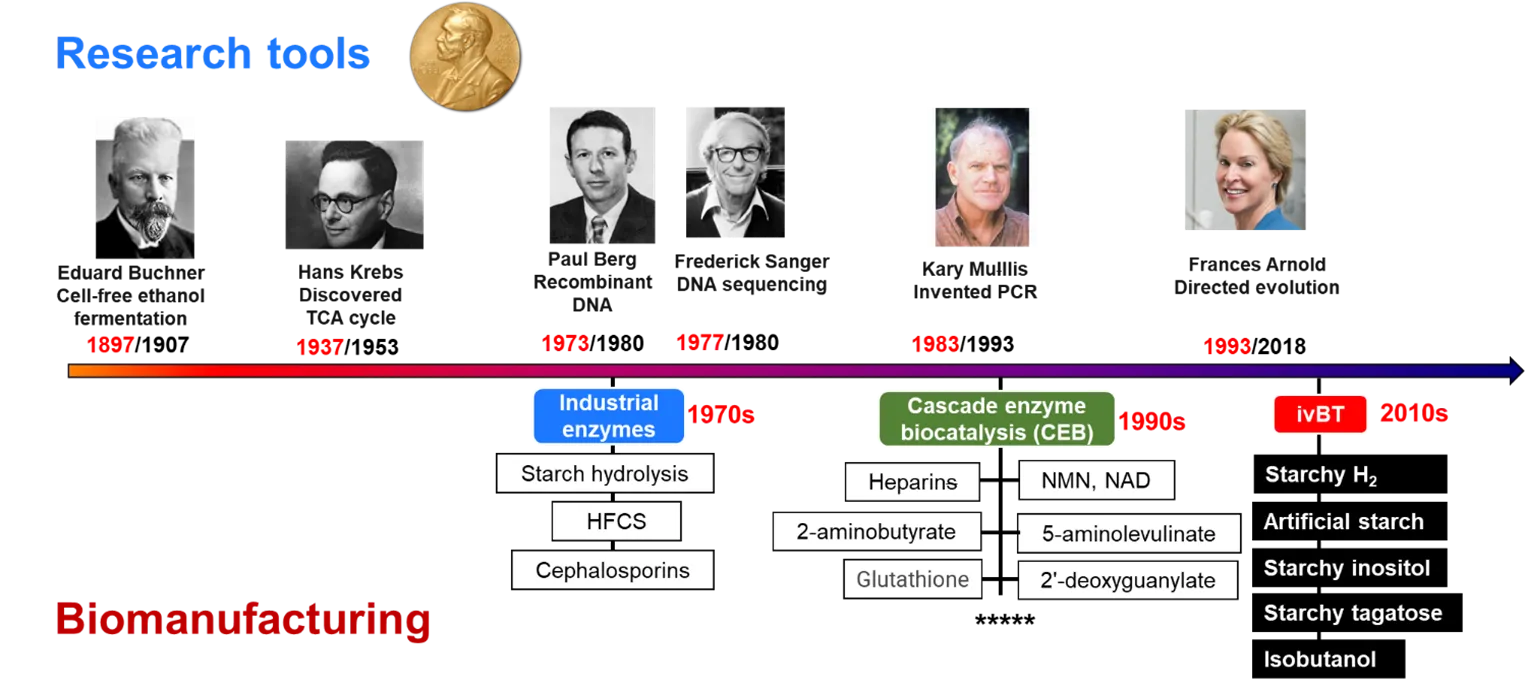
shows the key milestones of enzyme-based fundamental research and industrial biomanufacturing. Biochemistry starts from the discovery of cell-free ethanol fermentation by Eduard Buchner in 1897 [
116], leading to the Nobel Prize Chemistry 1907. Later, more and more scientists conducted fundamental research in biochemistry and enzymology. For example, Otto Meyerhof won the Nobel Prize in Physiology in 1922 for his elucidation of the glycolytic pathway [
117] and Hans Krebs won Nobel Prize of Chemistry in 1953 for his discovery of the tricarboxylic acid cycle [
118,
119]. Paul Bern became a Nobel laureate of Chemistry in 1980 by using enzymes to split genes and ligate recombinant DNA fragments [
120]. In 1983, Kary Mulllis conceptualized and validated the PCR technology by using DNA polymerase, winning the Nobel Prize of Chemistry in 1993 [
121]. Recently, Frances Arnold won the Nobel Prize of Chemistry in 2018 for her contributions to directed evolution of enzymes [
88,
122,
123]. Now
in vitro reconstitution of natural or artificial enzymatic pathways is a widely-used tool to help understand natural or synthetic pathways
in vivo. For example, Khosla and his coworkers investigated the kinetics of the fatty acid synthesis by using the reconstitution of the purified
E. coli fatty acid synthase components [
124]. Liu and his co-workers investigated the synthesis of farnesene by using purified enzymes [
125]. Adams and his coworkers demonstrated the production of 3-hydroxypropionic acid from hydrogen and carbon dioxide by using
P. furiosus cell-free extract [
126]. Liao and his coworkers tested the non-oxidative glycolysis design by using purified enzyme components [
127]. But the goal of these
in vitro pathway reconstitution is completely different from the biomanufacturing goal of ivBT proposed here.
. Key milestones of in vitro enzyme-based fundamental research and industrial biomanufacturing. Year in red represented the time that seminal research paper was published; year in black represented the time that Nobel prize was awarded.
Biomanufacturing mediated by enzymes has a shorter history than microbial fermentation. Biocatalysis starts from the use of one enzyme [
6,
105,
128]. Invertase may be the first immobilized enzyme used commercially for the production of Golden Syrup (a hydrolytic product of sucrose) by Tate & Lyle in World War II. Industrial process for L-amino acid production by soluble aminoacylase was developed in 1954. Tanabe Seiyaku Co. (Japan) started the industrial production of L-methionine by using immobilized aminoacylase in a packed bed reactor in 1969. Clinton Corn Processing Company (USA) was the first to produce fructose corn syrup by glucose isomerase in 1967. Currently, immobilized glucose isomerase by Novozymes is packed into columns for bioconversion of glucose into fructose. Now, annual world production of high fructose corn syrup exceeds 20 million metric tons.
Over time, biocatalysis has evolved to cascade enzyme biocatalysis (CEB) or multi-enzyme one pot for making fine chemicals due to several advantages, such as fewer unit operations, smaller reactor volume, higher volumetric and space-time yields, shorter cycle times, and less waste generated [
129,
130,
131,
132]. At the beginning, this technology may be developed to address NAD(P)H regeneration for dehydrogenase-mediated biocatalysis, especially for the synthesis of high-value chiral compounds in the pharmaceutical industry [
66,
133]. NAD(P)H is usually generated by using a pair of a hydrogen-donor substrate and a single enzyme, including formate/formate dehydrogenase [
134], glucose/glucose dehydrogenase [
135], glucose-6-phosphate/glucose-6-phosphate dehydrogenase [
75], dihydrogen/hydrogenase [
136], and phosphite/phosphite dehydrogenase [
137]. Similarly, this strategy includes ATP-based biocatalysis [
61,
138,
139]. In the organic chemistry field, the synthesis of monosaccharides, activated monosaccharides, oligosaccharides, and glycopeptides by using multi-enzyme one pot has been intensively investigated [
140,
141,
142,
143,
144,
145,
146]. Until now, up to several hundred of small molecule active pharmaceutical ingredients (APIs) have been manufactured in the pharmaceutical industry [
132].
ivBT might be regarded as a further development of CEB with more enzymes or coenzymes. However, we would like to urge to distinct ivBT from CEB, systems biocatalysis [
147,
148] or synthetic biochemistry [
149,
150], by considering many aspects ().
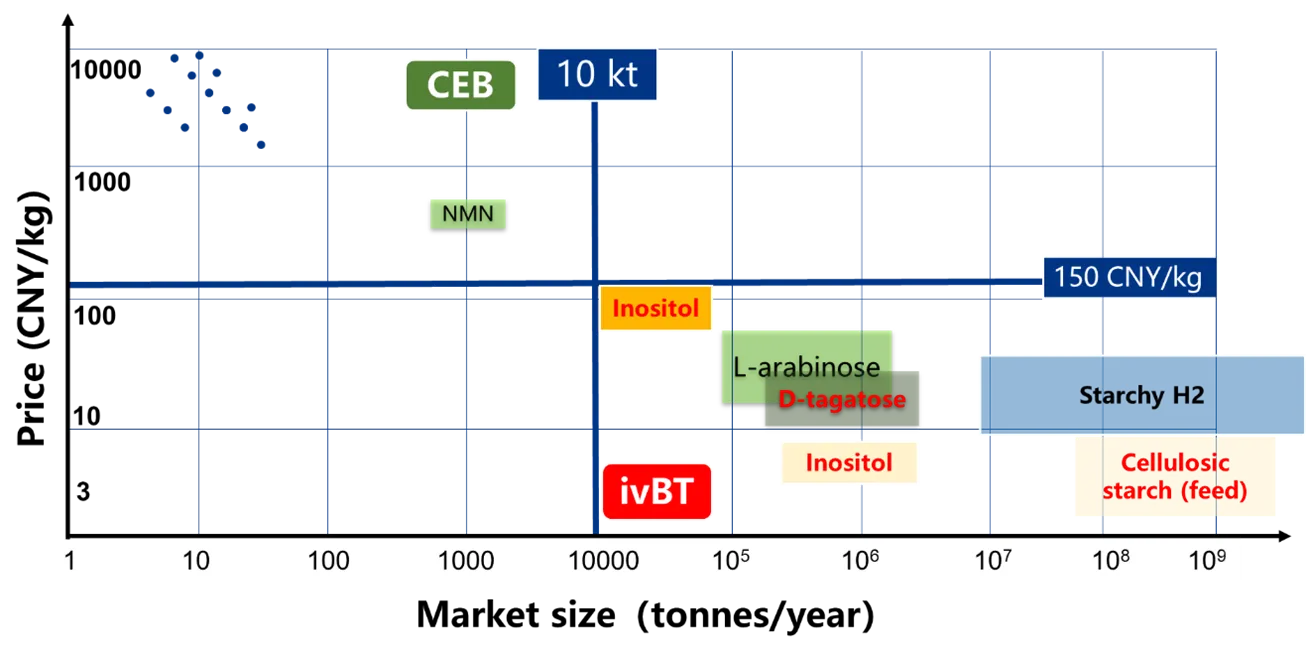
First, the biomanufacturing goals of ivBT and CEB are different ( and ). The former aims at biomanufacturing of biocommodities whose minimum mass is 10,000 metric tons, whose market size is greater than 500 million CNY, some of which could be greater than one billion metric tons. The latter aims at production of fine chemicals whose typical mass ranges from 100 kg to 100 metric tons and whose market size ceiling could be up to 500 million CNY and whose average market size could be approximately tens of millions CNY [
132]. The representative products of ivBT are energy-, food/feed-, and material-related, and their number could be small, in a range of hundreds. In contrast, the representative products of CEB are drug precursors, natural products, and fine chemicals, the number of these is large, greater than 10,000 ().
Second, the pathway design principles of ivBT and CEB are different (). ivBT pathways are redesigned based on artificial enzymatic pathways and artificial electric transfer chains (ETCs). CEB is mostly designed based on one-enzyme catalysis plus related coenzyme recycling or a part of natural pathways.
Third, the biocatalytic part requirements of ivBT and CEB are different (). The former requires low-cost and ultra-stable enzymes (plus enzyme immobilization), as well as low-cost biomimetic coenzymes and engineered dehydrogenases [
108]. As a result, the ratio of substrate cost to product price could be greater than 50%, or even 80–90%. The latter can use natural mesophilic enzymes or rarely used immobilized enzymes plus natural coenzymes [
151]. Due to high product price, the ratio of substrate cost to product price could be around 20% or lower [
152].
Fourth, the biomanufacturing processes of ivBT and CEB are different (). The former prefers a long reaction time (e.g., weeks or even months) for decreasing the enzyme-related biomanufacturing costs [
153]. The latter is usually operated in a batch mode, lasting hours or days only [
154].
Also, it is worth mentioning that cell-free protein synthesis (CFPS) based on numerous cell lysates from bacteria (e.g.
E. coli), mammalian, plant, insect, and so on [
155,
156], purified recombinant components (e.g., PURExpress) [
157], cell-free metabolic engineering [
158] or cell-free biomanufacturing [
159] is far different from ivBT (). According to dictionaries,
in vitro is defined as “outside the living body and in an artificial environment” or “(of a biological process) made to occur in a laboratory vessel or other controlled experimental environment rather than within a living organism or natural setting”. Cell-free is defined as without living cell by cell lysis and its synthesis began with fundamental research (i.e., rapid synthesis of proteins within hours [
156] and optimization of
in vitro natural pathways for in living microorganisms [
160,
161]). Now it might be a special (small-size) biomanufacturing [
162], such as urgent vaccines [
163], antibody-drug conjugates (ADCs) with high quality and incorporation of nonnatural amino acids [
164,
165], toxin (e.g., botulinum toxin) [
166,
167], RNA products [
168], and so on. Clearly, CFPS or cell-free metabolic engineering is far different from ivBT in many aspects from aimed products, catalysts and applications.
. Schematic comparison of the product market sizes biomanufactured by ivBT and cascade enzyme biocatalysis (CEB).
. Comparison of ivSEB, cascade enzyme biocatalysis, and cell-free protein biosynthesis.
The most important three criteria for biomanufacturing are TRY, Titer in terms of g/L, Rate (volumetric productivity) in terms of g/L/h, and Yield in terms of g/g. Among TRY, there is no doubt that the product yield or energy efficiency is the most important for biomanufacturing of a biocommodity because the substrate costs usually account for more than a half of the costs of the desired product () [
37,
38]. The largest biomanufacturing advantage of ivBT is its nearly theoretical yield of the target product [
48,
59,
60,
67,
169,
170,
171,
172] because most enzymes have nearly 100% chemical selectivity and there is neither synthesis of cells nor production of other products, all of which waste the substrate. ivBT has one inherent biomanufacturing advantage: very low energy consumption. When ivSEB is designed, it should have balanced NAD(H) regeneration and consumption. As a result, most ivSEBs need neither aeration, nor vigorous stirring, nor cooling, i.e., they have very low energy consumption.
The second biomanufacturing advantage is high volumetric productivity possibly due to a lack of cellular membrane and high volumetric enzyme loadings. Typically, enzyme-based biocatalysis usually has one-to-two orders of magnitude faster than microbial fermentation [
160,
163,
173], wherein microbes containing thousands of intracellular enzymes have only a few enzymes responsible for the production of the desired product.
ivBET is an open biosystem without competing pathways, complicated regulations of protein synthesis, nor cell self-duplication [
41,
155]. Taking a relatively simple pathway involving six cascade biochemical reactions where each step has five choices (genes or enzymes), ivBES would have 30 combinations since each enzyme in a layer can be easily exchanged by another enzyme. Microbes may have 5
6 = 625 combinations because each layer is linked with other layers. Furthermore, microbes are far more complicated by the possibility that reaction rate at each layer involves regulation in terms of strength of promoter at the gene level, at the level of mRNA stability, at the level of protein translation, at the level of protein delivery to the site of activity in the cell, and at the level of protein interactions with other proteins and coenzymes [
41]. As a result, ivBT featuring easy process operation and optimization has better biomanufacturing robustness than microbes [
158,
170,
174,
175].
Biochemically, enzymes can tolerate toxic components better than microbes [
76,
150,
176,
177]. For example, natural enzymes can tolerate very high concentrations of alcohols [
176], at least one order of magnitude higher than microbes [
178]. Enzymes can be easily engineered to tolerate very high concentrations of organic solvents [
122]. An ivSEB exhibited to work well in the presence of complicated toxic inhibitors in the hydrolysate from biomass pretreatment [
76], while yeast cannot grow. Therefore, ivSEB is believed to make high-titer toxic compounds better than microbes [
54].
3. ivBT Examples
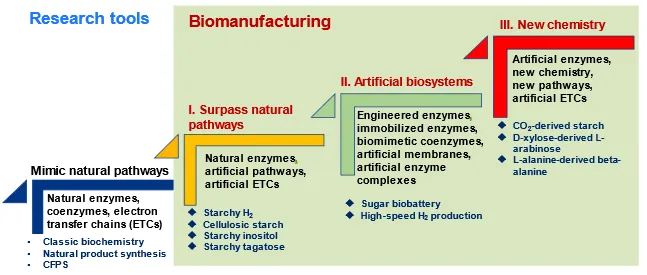
Although ivBT mediated by ivSEB has a pretty short history, it has passed through three phases in terms of science behind it (). The first phase is exceeding natural pathways by the reconstitution of natural enzymes, natural coenzymes and natural electron transfer chains, for example, high-yield hydrogen production from starch and water [
47], production of inositol and rare sugars from starch [
60,
179,
180,
181], the directed bioconversion of cellulose to starch [
50]. The second phase is artificial enzymatic biosystems with new man-made parts (e.g., biomimetic coenzymes, artificial membranes, artificial enzyme complexes, artificial ETCs) and engineered parts (e.g., engineered enzymes, immobilized enzymes). The representative example of Phase II are sugary biobattery equipped with a Nafion membrane and an electron mediator [
67], high-speed enzymatic hydrogen production featuring an artificial ETC and immobilized coenzyme and enzyme complexes [
108]. The third phase is new chemistry featuring new chemical reactions catalyzed by artificial enzymes and new pathways, for example, starch biosynthesis from CO
2 with a nonnatural enzyme [
53], L-arabinose biosynthesis from D-xylose with a nonnatural enzyme pentose 4-epimerase [
182], beta-alanine synthesis from L-alaine by cascade amino decarboxylases [
183].
Here we present several examples with their potential market sizes from small to large to help readers understand the ivBT and ivSEB concepts, technology development history and its future applications.
. The phase development of enzyme-based biocatalysis as compared to the fundamental research of biochemistry and enzymology.
-Inositol (called inositol later) is a six-carbon alcohol featuring its carbon-carbon ring [
59,
60]. Inositol is a member of the vitamin B (VB8), it has been widely used in the drug [
184], food [
59], and feed industries [
60]. Traditional inositol production is based on the isolation of phytate from corn kernels and rice brans. This method suffers from limited feedstock supplies (i.e., one kg of inositol is made from the steep liquor of 2000 kg of corn kernels), costly feedstock, awful odor pollution, serious phosphorous pollution, and complicated separation of feedstock and product, resulting in high price tags of inositol and limited supplies. Current inositol market size is approximately 15,000 metric tons and its potential market size could be up to one million metric tons as predicted.
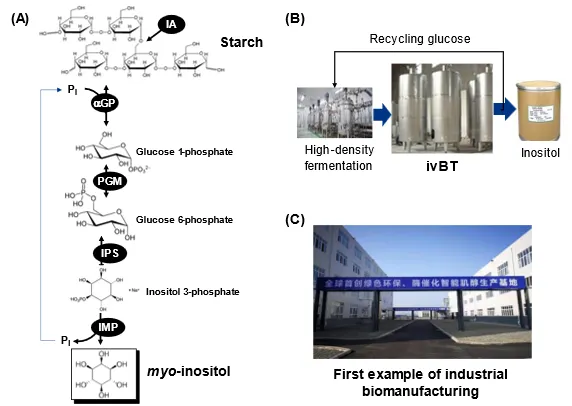
Inositol is the first bioproduct industrially produced by ivBT. Prof. Zhang generated the idea of the starch-to-inositol synthetic enzymatic pathway in 2013 and identified all thermophilic enzymes from Kyoto Encyclopedia of Genes and Genomes (KEGG) database suitable for biomanufacturing. shows the artificial enzymatic pathway comprised of (i) α-glucan phosphorylase (αGP, EC 2.4.1.1), which produces glucose 1-phosphate from starch and phosphate; (ii) phosphoglucomutase (PGM, EC 5.4.2.2); (iii) inositol 3-phosphate synthase (IPS, EC 5.5.1.4); and (iv) inositol monophosphatase (IMP, EC 3.1.3.25), wherein phosphate is recycled between Reactions 1 and Reaction 4 in one vessel. To increase its yield, branched starch can be hydrolyzed to linear amylodextrin by isoamylase [
185]. Also, one glucose of maltose can be further utilized by 4-glucanotransferase [
48], the residual glucose can be phosphorylated in the presence of polyphosphate catalyzed by polyphosphate glucose kinase [
89,
186]. Prof. Zhang utilized two enzymes—αGP and PGM to generate glucose 6-phosphate from starch and phosphate without ATP. The energy required for glucose phosphorylation comes from α-1,4 glycosidic bond energy of starch; PGM rapidly catalyzes glucose 1-phosphate to glucose 6-phosphate with a negative Gibbs free energy change (∆G° = −7.4 kJ/mol). Furthermore, the Gibbs free energies of the carbon-carbon cycloisomerization catalyzed by IPS and the dephosphorylation catalyzed by IMP [
187] are −55.2 and −20.7 kJ/mol, respectively (A), suggesting that both reactions are spontaneous, thermodynamically favorable, and highly irreversible. The consolidation of the reversible reactions and the irreversible reactions have an overall Gibbs energy of −80.1 kJ/mol, driving the overall reaction toward completeness. Later Prof. Zhang and Dr. You filed the first patent application in 2015 [
181].
To speed up its industrial biomanufacturing, Prof. Zhang organized a Chinese team including scientists and engineers to address remaining industrialization challenges (e.g., low-cost enzyme production, simple enzyme purification, optimization of enzyme ratios, inositol purification, and so on). The team developed high-density fermentation of
Escherichia coli overexpressing all thermophilic enzyme building blocks and purified them by heat treatment. As compared to microbial fermentation by engineered microorganisms [
188,
189], this two-step bioprocessing (B) includes enzyme production by microbial fermentation and ivBT, each of which step has the sole goal. The unutilized sugars in ivBT can be recycled back to microbial fermentation, resulting in very high substrate utilization efficiency and low pollutants generated. In 2016, a Chinese company acquiring this technology built a factory that can produce up to 10,000 metric tons of inositol yearly and is the world largest inositol maker (C). This ivBT has been successfully operated on 60,000-L bioreactors. Over years, Prof. Zhang’s group continues improving this technology by developing low-cost enzyme co-immobilization techniques [
190] and decreased its biomanufacturing costs greatly [
153].
. Schematic presentation of the in vitro starch-to-inositol pathway (A), wherein enzymes are αGP (α-glucan phosphorylase), PGM (phosphoglucomutase), (iii) IPS (inositol 3-phosphate synthase), (iv) IMP (inositol monophosphatase), and IA (isoamylase); the concept of two-step biomanufacturing as well as product separation and glucose reuse (B); and image of the first large-scale inositol factory (C).
Several months later, another Japanese group led by Prof. Atomi, who is an expert in biochemistry in hyperthermophiles, published the starch-to-inositol paper independently [
179], whose submission date was just three months later than Prof. Zhang’s inositol paper [
60]. It suggests that two independent groups thought of the same product and same pathway, like the case of the telephone patent applications. Interestingly, both Profs. Zhang and Atomi selected exactly three enzymes (i.e., PGM, IPS and IMP) from the same microorganisms in the four-enzyme pathway, wherein each enzyme could be chosen from up to thousands of gene sources in open databases. It implies that it is common for experts to design the same pathway and find out the same enzymes from public information (e.g., publications, database) for the same purpose.
Later, the similar inositol-producing pathways
in vitro and
in vivo have been designed and validated to make inositol from sucrose, cellulose, xylose, glucose and so on [
59,
171,
188,
189,
191,
192]. Now inositol is being produced by ivBT and microbial fermentation in China. We predict that microbial fermentation could not be economically competitive to compete with ivBT by considering biomanufacturing rules.
3.2. Rare Sugars (Healthy Sweeteners)
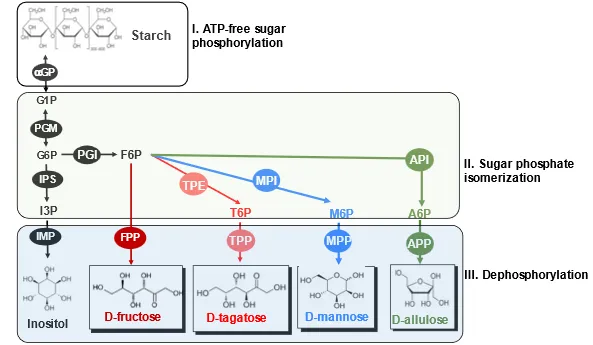
Rare sugars are monosaccharides with a limited availability in nature and some of them have special healthy functions [
193,
194]. In 2004, Prof. Izumori proposed a complete strategy for the biosynthesis of rare sugars by using a newly-discovered enzyme monosaccharides (e.g., D-tagatose 3-epimerase) plus known enzymes—aldose isomerase, aldose reductase, and oxidoreductase enzymes [
193,
195]. The Izumori methodology enables to make all rare sugars from cheap substrates. However, this strategy has several weaknesses: (1) a series of equilibrium reactions, requiring the use of simulated moving bed (SMB) chromatography to separate the desired product and substrate; (2) the possible addition of costly NAD(P), resulting in high biomanufacturing costs; and (3) limited and costly supplies of some feedstocks (e.g., D-galactose). Among 24 hexoses, D-tagatose, D-allulose, and D-mannose are of special interests.
D-tagatose is a natural sweetener and its sweetness is the nearly same as that of sucrose with only one third of its calories [
194,
196]. It has many physiological benefits, such as low calorie, low glycemic index, anti-caries, anti-oxidation, prebiotics, improvement of intestinal function, enhancement of immune [
194]. In 2000 US Food and Drug Administration (FDA) approved it as generally recognized as safe (GRAS) [
196]. The scientists and companies around the world have worked on its industrial biomanufacturing for long. According to Izumoring, D-tagatose is made through D-galactose isomerization from lactose [
197,
198]. The isomerization of galactose to tagatose is an equilibrium reaction so that chromatography is required to separate the two monomeric sugars. Prof. Sun, who finished her training from the Izumori group, came back to a newly-established research institute—Tianjin Institute of Industrial Biotechnology (TIB), Chinese Academy of Sciences (CAS) in 2009, and began to work on low-cost biomanufacturing of tagatose with the TIB Director Ma. They published several papers of D-tagatose 3-epimerase mining [
199,
200,
201,
202] and obtained several patents of tagatose production [
203,
204]. However, D-tagatose was a relatively costly sweetener to produce due to costly and limited feedstock lactose and complicated separation [
205,
206].
In 2014, Prof. Zhang who founded Cell Free Bioinnovations Inc. (CFB) started to investigate new methods to produce D-tagatose from less costly and more abundant feedstock D-glucose instead of D-galactose because he knew the great potentials of rare sugars from TIB scientists. He hypothesized to mine and create an artificial enzyme glucose 4-epimerase from a natural enzyme, such as UDP-glucose 4-epimerase as early as 2014. In 2015, he recruited a new CFB employee Dr. Danial Wichelecki to work on the tagatose project. Soon, Dr. Wichelecki presented his postdoc research results—the discovery of a novel natural enzyme—
Agrobacterium tumefaciens tagatose 6-phosphate epimerase interconverting tagatose 6-phosphate and fructose 6-phosphate [
207]. Therefore, Zhang and Wichelecki together conceptualized the starch-to-tagatose pathway () by combining (1) the starch-to-inositol pathway, (2) tagatose 6-phosphate epimerase, and (3) highly-selective phosphatase [
77,
208], whose function is like IMP in the starch-to-inositol pathway [
60]. This pathway design was partially inspired by the starch-to-fructose pathway [
44,
209] ().
Director Ma and Prof. Sun at TIB worked on tagatose as early as 2009 [
199,
200,
201,
202]. A little later, TIB’s starch-to-tagatose patent disclosure [
210] was filed based on the starch-to-inositol patent [
181] and fructose-to-tagatose patent [
208]. It has three modules: (1) the ATP-free generation of sugar phosphates from starch [
47,
81,
181], (2) the isomerization and epimerization to D-tagatose 6-phosphate [
207,
208], and (3) the dephosphorylation of D-tagatose 6-phosphate to tagatose [
77,
208]. Later, they improved this technology by using five whole
E. coli cells expressing thermophilic enzymes [
211] and used the GRAS bacterium
Bacillus subtilis co-expressing five enzymes in a host [
212,
213]. The starch-to-tagatose synthesis was demonstrated to be catalyzed by immobilized enzyme mixture [
56].
A little later, another Chinese group Prof. Jiang, who also worked on tagatose for long, characterized new enzymes
Dictyoglomus turgidum α-glucan phosphorylase [
214] and
Caldilinea aerophila fructose 6-phosphate 4-epimerase [
215], and used them to produce tagatose from maltodextrin, whose pathway is the same as the patent applications [
180,
210]. They also co-expressed five enzymes in one
E. coli host and used the heat-treated whole cells to produce tagatose from starch [
216].
D-Allulose (also called D-psicose) is another low-calorie rare sugar and is the C-3 epimerization product of fructose [
217,
218]. It has 70% relative sweetness and only 10% of the calories of sucrose, and it is considered an ideal substitute for high-fructose corn syrup [
219]. FDA approved it as GRAS food additive in numerous foods and dietary supplements for enhancing gel strength, decreasing the oxidation extent, and improving flavor during food processing [
220]. Also, it has distinctive physicochemical properties and physiological functions, such as antihyperlipidemic effects [
219], antihyperglycemic effects [
221,
222], anti-inflammatory effects [
223] and so on [
57]. D-allulose is the most popular rare sugar because it can be synthesized from D-fructose catalyzed by allulose 3-epimerase [
193]. However, its epimerization suffers from an unfavorable equilibrium, i.e., it requires multiple times of SMB for achieving high product yields. Similar to the cases of inositol by two independent groups, and of tagatose by three independent groups, three unrelated companies and one academic group independently conceptualized the same
in vitro enzymatic pathway for the production of D-allulose from starch (). In 2014, Korean CJ company filed the first starch-to-allulose patent application [
224]. Two years later, within a half month two American companies Bonumose [
225] and Greenlight Biosciences [
226] filed the following starch-to-allulose pathway disclosures. However, their disclosures did not release key detailed information, such as enzyme sources, enzyme characteristics, optimal experimental conditions, product titer, and so on [
225,
226]. In 2021, Dr. You at TIB published the first academic paper to demonstrate high-yield production of D-allulose from starch [
57] with enough experimental details. It is noted that Dr. You is the first author of the starch-to-inositol research paper [
60].
D-mannose, a 2-epimer of D-glucose, is another rare sugar that can be found in certain fruits and vegetables. In human blood, it is found to be approximately a hundred times less abundant than glucose in human blood [
227]. It is widely used to prevent urinary tract infections or bladder inflammation from infections. Recent studies demonstrated that supraphysiological levels of D-mannose inhibit tumor growth [
228], stimulate regulatory T cell differentiation [
229], and suppresses macrophage IL-1β production [
227]. According to the Izumori methodology, it can be produced from D-fructose catalyzed by isomerases and also suffers from a low equilibrium constant. Inspired by the ATP-free sugar phosphorylation, sugar phosphate isomerization and dephosphorylation, an
in vitro starch-to-mannose pathway was designed and demonstrated [
230].
Clearly, the consolidation of the ATP-free sugar phosphorylation, sugar phosphate isomerization and dephosphorylation () would be widely adopted to produce numerous rare sugars with higher yields and overcome the equilibrium limitation of the Izumori methodology. The success of industrial starch-to-inositol biomanufacturing would encourage the industrial biomanufacturing of rare sugars soon.
. Schematic presentation of the in vitro starch-to-rare sugars pathways as compared to the starch-to-inositol pathway. Enzymes are αGP, PGM, IPS, IMP, PGI, PGI (phosphoglucose isomerase), FPP (fructose 6-phosphatase), TPE (tagatose 6-phosphate 4-epimerase), TPP (tagatose 6-phosphatase), MPI (mannose 6-phosphate isomerase), MPP (mannose 6-phosphatase), API (allulose 6-phosphate isomerase), and APP (allulose 6-phosphatase).
1,3-propanediol (1,3-PDO) is a colorless viscous liquid, which can be used as monomer in the production of polymers such as polytrimethylene terephthalate [
231]. It can be produced from glucose [
232] or glycerol [
233] by microbial fermentation. Scientists at DuPont and Genencor metabolically engineered
E. coli to produce 1,3-PDO from glucose on an industrial scale [
234]. However, its yields based on glycerol are approximately 0.60 mol 1,3-PDO/mol glycerol [
233,
235,
236]. This relatively low yields are due to the synthesis of cell mass, the generation of extra reducing power by branching glycerol utilization pathways under micro-aerobic conditions, and the production of undesired side-products [
42].
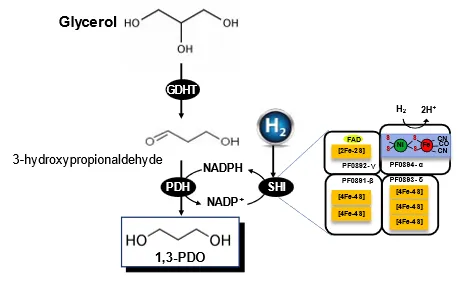
Prof. Zeng and his coworkers developed a straightforward
in vitro pathway for converting glycerol to 1,3-PDO under strictly anaerobic condition () [
237]. This pathway includes two modules: (i) the generation of 1,3-PDO from glycerol catalyzed by glycerol dehydratase (GDHT) and NADPH-dependent propanediol dehydrogenase (PDH) at a cost of one NADPH consumption per molecule; and (ii) NADPH regeneration from H2 by soluble hydrogenase I (SHI). As a result, one mole of 1,3-PDO was produced from one mole of glycerol and one mole of H2. 1,3-PDO yield of approximately 0.95 mol/mol has been accomplished [
237].
. Schematic presentation of the
in vitro synthetic enzymatic pathway for the production of 1,3-propanediol from glycerol, modified from Ref. [
237]. Enzymes are GDHT (glycerol dehydratase), PDH (NADPH-dependent propanediol dehydrogenase), and SHI (soluble hydrogenase I) that is four-subunit cytoplasmic hydrogenase from
P. furious.
Isobutanol, a branched C4 alcohol, is a better biofuel than ethanol, has limited miscibility with water, and is completely miscible with gasoline [
238]. Isobutanol’s branching gives it a better octane number than
N-butanol. It can be produced through the Ehrlich pathway [
239,
240]. Heterologous pathway for isobutanol production from carbohydrates have been introduced to a number of microorganisms, such as
E. coli [
238],
Bacillus subtilis [
241],
Corynebacterium glutamicum [
242],
S. cerevisiae [
243]. However, its hydrophobic effect of the long-chain alcohol is highly toxic to cellular membranes, resulting on low isobutanol titers (e.g., 1–2%) [
244].
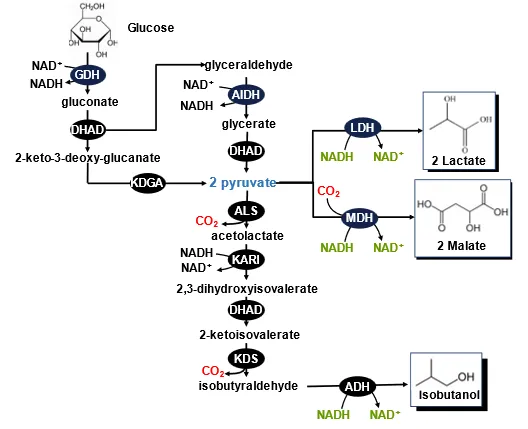
Prof. Sieber and his coworkers designed an
in vitro ATP-free synthetic enzymatic pathway for isobutanol production from glucose () [
54]. This pathway contains two modules: (i) the generation of two pyruvate from glucose mediated by four enzymes (note: this is an ATP-free pathway), and (ii) the production of one isobutanol from two pyruvate. Therefore, one glucose can produce one isobutanol, releasing two CO
2 and one water. This pathway is much shorter than the natural
N-butanol pathway featuring neither ATP nor CoA. The proof-of-concept experiment obtained a 53% isobutanol yield even in the presence of 4% isobutanol [
54], suggesting that ivSEB can tolerate organic solvents far better than cellular membranes.
. Schematic presentation of the
in vitro ATP-free synthetic enzymatic pathways for the production of isobutanol [
54] and lactate [
245]. The enzymes are GDH (glucose dehydrogenase), DHAD (dihydroxy acid dehydratase), AIDH (glyceraldehyde dehydrogenase), KDGA (2-keto-3-desoxygluconate aldolase), ALS (acetolactate synthase), KARI (ketol-acid reductor-isomerase), KDC (2-ketoacid decarboxylase), ALD (alcohol dehydrogenase), LDH (lactate dehydrogenase), and MDH (malate dehydrogenase) [
246].
A combination of this shortest glucose-to-pyruvate pathway and NADH-consuming lactate dehydrogenase enabled to produce L-lactate with a yield of 90% [
245]. Similarly, the replacement of lactate dehydrogenase with decarboxylating malate dehydrogenase could produce two molecules of malate from one glucose and two CO
2; a combination of this glucose-to-pyruvate pathway and NADH-consuming L-alanine dehydrogenase may produce a theoretical yield of L-alanine. This five-enzyme pathway could be better than another
in vitro ATP-self-balanced pathway for malate production from starch [
172].
3.5. Artificial Starch
Starch is a natural polysaccharide that is used as an energy storage compound for plants, animals (more exactly, glycogen, animal starch), and microorganisms. Plant starch include amylose (a nearly linear glucan linked by α-1,4-glycosidic bonds) and amylopectin (a branched molecule in which the branch points consist of α-1,6 glycosidic bonds while the linear portions of the branches are made up of α-1,4 bond as in amylose). Human beings can digest starch but cannot cellulose. Human civilization started with 10,000-year ago agriculture that produced starch-rich grains as human food. To feed the increasing world population, modern agriculture has consumed approximately 70% freshwater withdrawal, all available lands, and huge amounts of fertilizers to make 2.76 billion metric tons of grains per year [
28,
29].
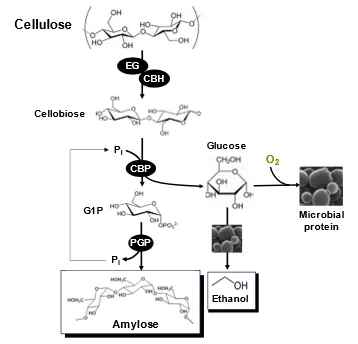
A huge amount of nonfood lignocellulose (i.e., ~200 billion metric tons) is produced yearly, a small fraction of which is used as animal feed, materials, and burning fuels. Cost-effective biotransformation of cellulose to starch would revolutionize 10,000-year agriculture. Prof. Zhang designed a simple
in vitro coenzyme-free pathway to transform cellulose to amylose () [
50]. This pathway includes three parts: (1) partial hydrolysis of cellulose to cellobiose by endoglucanase and cellobiohydrolase, (2) the ATP-free generation of glucose 1-phosphate catalyzed by cellobiose phosphorylase, and (3) amylose synthesis from glucose 1-phosphate catalyzed by potato α-glucan phosphorylase, wherein phosphate is recycled between two enzymes. To avoid wasting one substrate of cellobiose, yeast cells can ferment into ethanol under anaerobic conditions [
50]. Alternatively, glucose can be used to make microbial protein (i.e., another key protein source as food and feed) under aerobic conditions [
51]. This breakthrough was highlighted as a news article entitled “Could wood feed the world” in Science magazine [
247]. Frances Arnold commented it as “it’s a simple but nice idea”. Profs. Arnold and Tyner doubts “whether the process is economically feasible overall is the big question” [
247]. However, Prof. Zhang believe that it would be economically feasible to biotransform beta-1,4-glycosidic bond-linked cellulose to α-1,4-glycosidic bond-linked starch because it is 100% energy-conservative, requiring neither coenzyme nor energy input nor sugar loss by considering coproduction of microbial protein or ethanol, neither costly conezymes nor harsh reaction conditions. It was speculated that US government disliked this breakthrough and was unwilling to sponsor it because it could impair the food weapon that the US government often used.
. Schematic presentation of the
in vitro coenzyme-free synthetic enzymatic pathway for the production of synthetic amylose, ethanol by anaerobic ethanol-producing microorganism [
50], and microbial protein by aerobic fermentation [
51]. The enzymes are EG, endoglucanase; CBH, cellobiohydrolase; CBP, cellobiose phosphorylase; and PGP, potato α-glucan phosphorylase.
Cost-effective and highly-efficient biotransformation of starch from agricultural residues, such as corn stover and wheat straw could easily double current agricultural food/feed output. Prof. Zhang believed that he can address this economical challenge with intensive R&D efforts [
51]. Zhang and his coworkers spent five years on making advances, such as (1) up to one million-fold cost reduction in enzyme parts used for ivBES, (2) high glucan digestibility and high starch yield of pretreated corn stover, and (3) synthesis of amylose and amylopectin tailored to different diet needs [
51]. Also, Prof. Zhang found out a novel way to enzymatically transform D-xylose to L-Arabinose (a healthy natural sweeteners with more than 50 billion CNY) so to increase economical viability of new biorefining of agricultural residues to new food/feed sources. Trillions of CNY market size of new food/feed from agricultural residues could revolutionize agriculture and reshape the bioeconomy, while maintaining biodiversity, minimizing agriculture’s environmental footprint, and conserving fresh water [
29,
248]. This biotransformation would not only promote the cultivation of plants chosen for rapid growth rather than those optimized for starch-rich seed production, but it would also efficiently utilize marginal land for the production of the biomass required to meet the increasing needs of biofuels and biochemicals [
29,
249,
250]. In addition, perennial cellulosic plants and dedicated bioenergy crops can grow on low-quality or marginal land, and require low input, such as fertilizers, herbicides, pesticides, and water [
29].
Alternatively, artificial photosynthesis of starch from CO
2 and solar energy is hypothesized to surpass plant photosynthesis [
251]. Plants utilize intermittent non-point insolation to biologically fix CO
2 to biomass. However, plant photosynthesis has pretty low solar energy-to-chemical energy conversion efficiencies (e.g., ~0.2–0.3%, global average) and consumes a large amount of water. Such low energy efficiencies are mainly attributed to three factors: (i) a narrow light absorption spectrum by chlorophyll, (ii) relatively low efficiencies of carbohydrate synthesis and unmatched reaction rates between fast light-harvesting reactions and slow dark chemical synthesis reactions, and (iii) carbohydrate losses due to the respiration of living plants. Prof. Zhang hypothesized that the integration of solar cells, water electrolysis, and ivBES may surpass these limitations in plants and make water-insoluble amylose from CO
2 and hydrogen based on the design principles of ivBT, knowledge in the literature, and thermodynamics analysis [
29,
251].
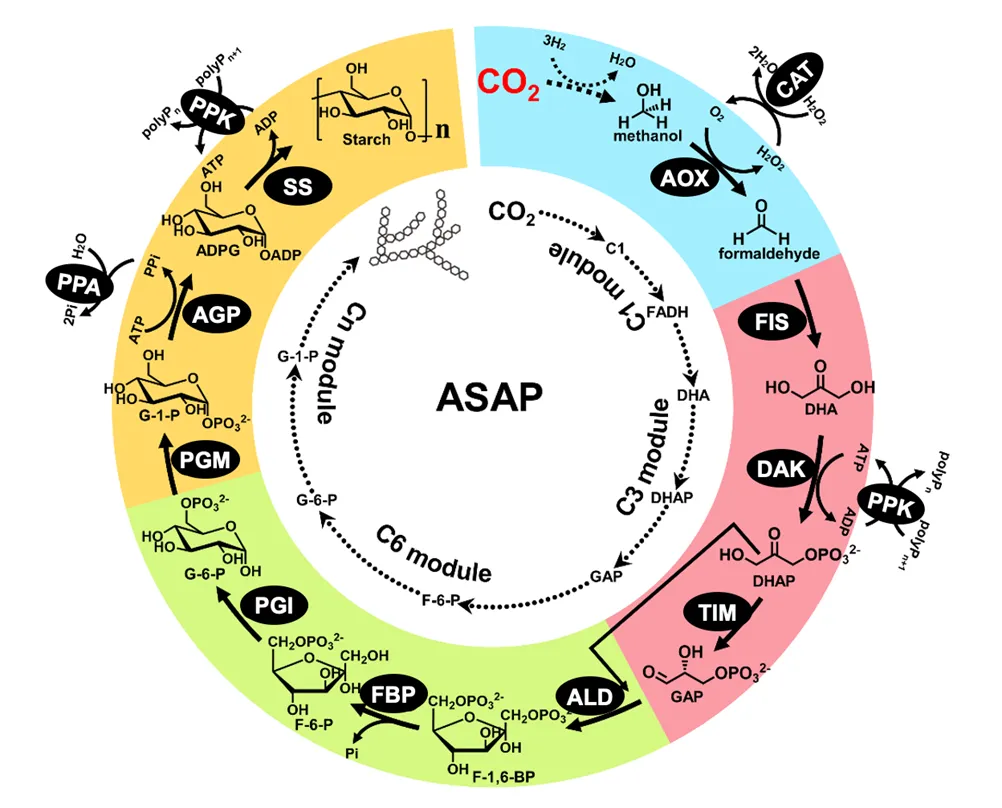
In 2021, TIB Director Ma led a large integrated team to validate the complete biosynthesis of starch from CO
2 and hydrogen made from solar cells for the first time [
53]. They designed an
in vitro synthetic enzymatic pathway, named as artificial starch anabolic pathway (ASAP) (). The C1 module is a hybrid of chemoenzymatic conversion from CO
2 to formaldehyde, the C3 module is the formation of glyceraldehyde 3-phosphate and dihydroxyacetone phosphate, the C6 module is glucose 6-phosphate synthesis, and Cn module is for starch biosynthesis. After intensive optimization and engineering of three rate-limiting enzymes, this ivSEB can make starch at a volumetric productivity of 22 nanomoles of CO
2 per minute per milligram of total catalyst, ~8.5-time of that of maize. This achievement has been selected in numerous awards, such as the National 13th Five-Year Plan Scientific and Technological Innovation Exhibition, China’s Top Ten Scientific Advances in 2021, China’s Top Ten Scientific Progress News in 2021, and so on. In 2023, the same team demonstrated further improvement in the biosynthesis of sugars from CO
2 and hydrogen [
55].
Clearly, food security challenge could be addressed soon if we addressed remaining technical challenges of ivBT timely [
29].
. Schematic presentation of the in vitro methanol-to-starch enzymatic pathway [
53]. Central enzymes are AOX (alcohol oxidase), FIS (formolase), DAK (dihydroxyacetone kinase, TIM (triose phosphate isomerase), ALD (fructose-bisphosphate aldolase), FBP (fructose bisphosphatase), PGI (phosphoglucose isomerase), PGM (phosphoglucomutase), AGP (ADP-glucose pyrophosphorylase), SS (starch synthase) and three supplementary enzymes are CAT (catalase), PPK (polyphosphate kinase), and PPA (pyrophosphatase).
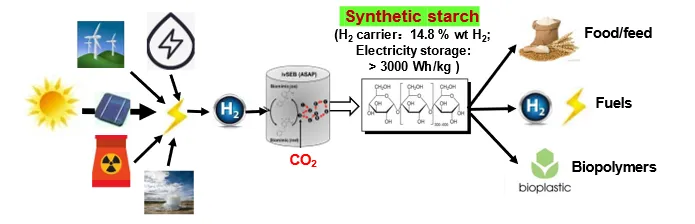
Hydrogen is the most promising future energy carrier (i.e., a second energy that is produced from primary energies), especially for the distributed users (e.g., heavy-duty trucks, aircrafts, remote areas, etc.). Now global hydrogen production is approximately 70 million metric tons, most of which are made by fossil fuels. It is anticipated that its production would increase to 300 million metric tons in 2030 and most of them will be produced from renewable energy. The future energy system based on hydrogen as an energy carrier is named as the hydrogen economy [
252,
253]. However, the hydrogen economy has four technical challenges, e.g., green cost-competitive distributed hydrogen production methods, low-cost high-density hydrogen carrier, an affordable infrastructure for the distribution and storage of hydrogen, as well as affordable and long-lasting hydrogen fuel cell systems [
254,
255]. If the hydrogen economy came true, the green hydrogen market size would be tens of trillions of CNY and greater than starch as food/feed [
256].
Natural dark-fermentation microorganisms produce hydrogen with a maximum yield of 4 H2 per glucose, plus two acetates, that is, the Thauer limit [
41,
257]. In theory, the complete oxidation of glucose with water as an oxidant may produce up to 12 hydrogen per glucose [
253,
258]. However, chemical catalysts produced green hydrogen with a far lower than this theoretical yield plus a lot of impurities [
259]. In nature, no microorganism evolved to make 12 hydrogen per glucose due to multiple reasons. If an anaerobic microorganism may generate 12 reduced coenzymes (e.g., NADPH) per glucose under strictly anaerobic conditions, the complete biotransformation to hydrogen by hydrogenase is endothermic so that it cannot generate any ATP to support microorganism’s basic metabolism. If this microorganism split a small fraction of 12 NADPH (e.g., 1–2) to generate a pH gradient across cellular membrane for ATP generation, such pH gradient would be made through oxidative phosphorylation, which was in conflict of oxygen-sensitive hydrogenase. Current microorganism cannot handle hydrogen production by oxygen-sensitive hydrogenase under microaerobic conditions. Clearly, it is why no engineered microorganisms can break the Thauer limit [
260,
261].
The ivBT concept was began with
in vitro enzymatic hydrogen production [
38,
252]. In March 2006, Prof. Zhang conceptualized an ATP-free synthetic enzymatic pathway to produce high-yield hydrogen from starch and water. This pathway includes four modules: (1) ATP-free generation of glucose 6-phosphate from starch and phosphate (this was later used in the starch-to-inositol and -rare sugars pathways); (2) the generation of two NADPH and ribulose 5-phosphate from glucose 6-phosphate mediated by two dehydrogenases; (3) the regeneration of glucose 6-phosphate from ribulose 5-phosphate; and (4) hydrogen generation from NADPH mediated by soluble hydrogenase I (SHI). The consolidation of these four modules lead to the stoichiometric reaction (Equation 1).
The proof-of-concept experiment was conducted by using 13 enzymes from five different sources (e.g., bacterium, yeast, plant, animal, and archaea), most of which were purchased from Sigma [
47]. In this seminal publication [
47], its significance was clearly written as “
The unique features, such as mild reaction conditions, high hydrogen yields, likely low production costs ($2/kg H2), and a high energy-density carrier starch (14.8 H2-based mass%), provide great potential for mobile applications. With technology improvements and integration with fuel cells, this technology also solves the challenges associated with hydrogen storage, distribution, and infrastructure in the hydrogen economy.”
Great potentials of the hydrogen economy up to tens of trillions of CNY and this out-of-the-box solution as envisioned in 2007 drove Zhang’s group to spend 15 years on this important area [
252]. His lab’s key advances are summarized as below. (1) To address enzyme stability and high-cost of purchased enzymes, all of them with recombinant ultra-stable enzymes produced by E. coli [
108] except recombinant SHI expressed in
Thermococcus kodakarensis KOD1 [
69] have been replaced. Now all of them can work at 80 °C for more than one week [
108]. (2) Zhang’s team expanded sugar sources from starch to cellodextrins, sucrose, xylooligosaccharides, xylose, and glucose [
262,
263,
264,
265]. (3) To completely utilize all glucose units of starch as a high-density hydrogen carrier [
48], Zhang’s team mined and engineered thermophilic enzymes (e.g., 4-glucanotransferase for the utilization of maltose [
266], PPGK for the ATP-free activation of glucose [
89,
186], and isoamylase for debranching starch [
185]). (4) Zhang’s team increased volumetric productivity of hydrogen by 1000-fold by using kinetic modeling to identify the rate-limiting steps [
169,
262], optimization of enzyme loadings and the use of high-activity enzymes [
108]. (5) Zhang’s team constructed artificial electron transfer chains (ETCs) (B–E) for decreasing coenzyme cost and increasing reaction rates. The addition of an electron mediator benzyl viologen, neutral red, or methyl viologen as a bridge between NADPH and SHI catalyzed by NADPH rubredoxin oxidoreductase greatly increased the hydrogen productivity due to great decreases in the activation energy of the rate-limiting step [
173]. (6) Zhang’s team conducted the coenzyme engineering for two dehydrogenases of ivSEB from NADP to NAD to a biomimetic coenzyme NMN [
44,
90,
96,
98,
267]. Further, Prof. Zhang hypothesize that the use of abiotic electron mediators and biomimetic coenzymes accompanied with engineered dehydrogenases (E) could help achieve both goals of low cost and fast reaction rate [
42].
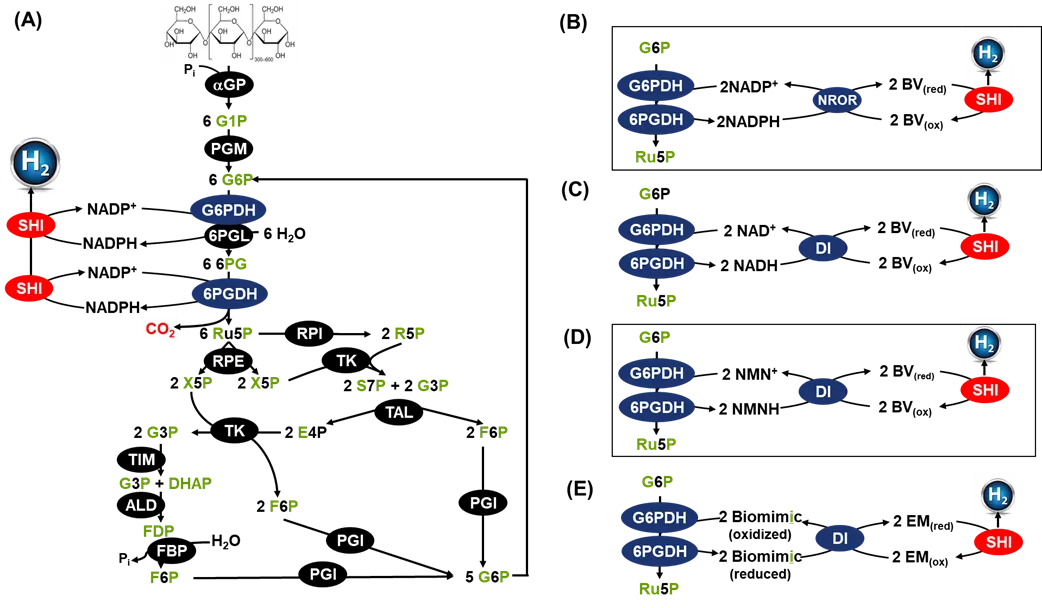
. Schematic presentation of the
in vitro synthetic enzymatic pathways for the production of high-yield hydrogen from starch and water (
A) [
47,
48,
108] and four artificial ETCs converting reduced power to hydrogen (
B–
E). The enzymes are αGP, PGM, G6PDH (glucose 6-phosphate dehydrogenase), 6PGL (6-phosphogluconolactonase), 6PGDH (6-phosphogluconate dehydrogenase), RPI (ribose 5-phosphate isomerase), RPE (ribulose-5-phosphate 3-epimerase), TK (transketolase), TAL (transaldolase), TIM, (triose phosphate isomerase), ALD (fructose-bisphosphate aldolase), FBP (fructose bisphosphatase), PGI, SHI, NROR (NADPH rubredoxin oxidoreductase), and DI (diaphorase). The metabolites are: g1p, glucose-1-phosphate; g6p, glucose-6-phosphate; ru5p, ribulose 5-phosphate; x5p, xylulose 5-phosphate; r5p, ribose 5-phosphate; s7p, sedoheptulose 7-phosphate; g3p, glyceraldehyde 3-phosphate; e4p, erythrose 4-phosphate; dhap, dihydroxacetone phosphate; fdp, fructose-1,6-diphosphate; f6p, fructose 6-phosphate.
Equation (1) catalyzed by ivSEB is a very rare entropy-driven chemical reaction under normal temperature and pressure conditions that can convert ambient thermal energy without another lower-temperature heat source to produce a usable energy – hydrogen. This is not against the Laws of Thermodynamics. The thermodynamic mechanism of this chemical reaction is that based on the entropy-increasing (phase transition) chemical reaction principle, the ivSEB can transform the environmental heat energy (low-grade enthalpy) and continuously produce high-grade chemical energy hydrogen energy without significant temperature gradient; by means of cascade release of the chemical energy of carbohydrate molecules, the ivSEB can split the chemical bonds of water molecules to produce high-purity hydrogen under normal temperature and pressure conditions (). Prof. Zhang hypothesized that a combination of the endothermic hydrogen-producing reaction, the exothermic hydrogen-fuel cell, and the high-energy-efficiency motor would lead to build the energy conversion system with the highest chemical-to-mechanical energy efficiency in the human history, laying a solid scientific and technological foundation for the future vehicle power system. Further, Prof. Zhang propose that starch would be a high-density hydrogen storage compound, whose hydrogen storage densities of starch-water slurry and dry starch are as high as 8.33% and 14.8% (weight/weight ratio), respectively. This new energy system based on starch as a high-density hydrogen storage compound would systematically solve the challenges of its production, storage, transportation, and application in the hydrogen economy [
52].
In a word, great potentials of the hydrogen economy and unique features of the starch-to-hydrogen technology as a holy grail of the hydrogen economy is motivating more scientists and engineers to continue improving this technology.
4. Advances in ivBT’s Supporting Technologies
Until now there is the only one commercial product – inositol produced by ivBT. A US company Bonumose claimed to complete annual 1000-tonnes tagatose production factory. To accelerate the biomanufacturing of ivBT, we highlight some recent advances to support ivBT.
4.1. Collection of Ultra-stable Enzymes
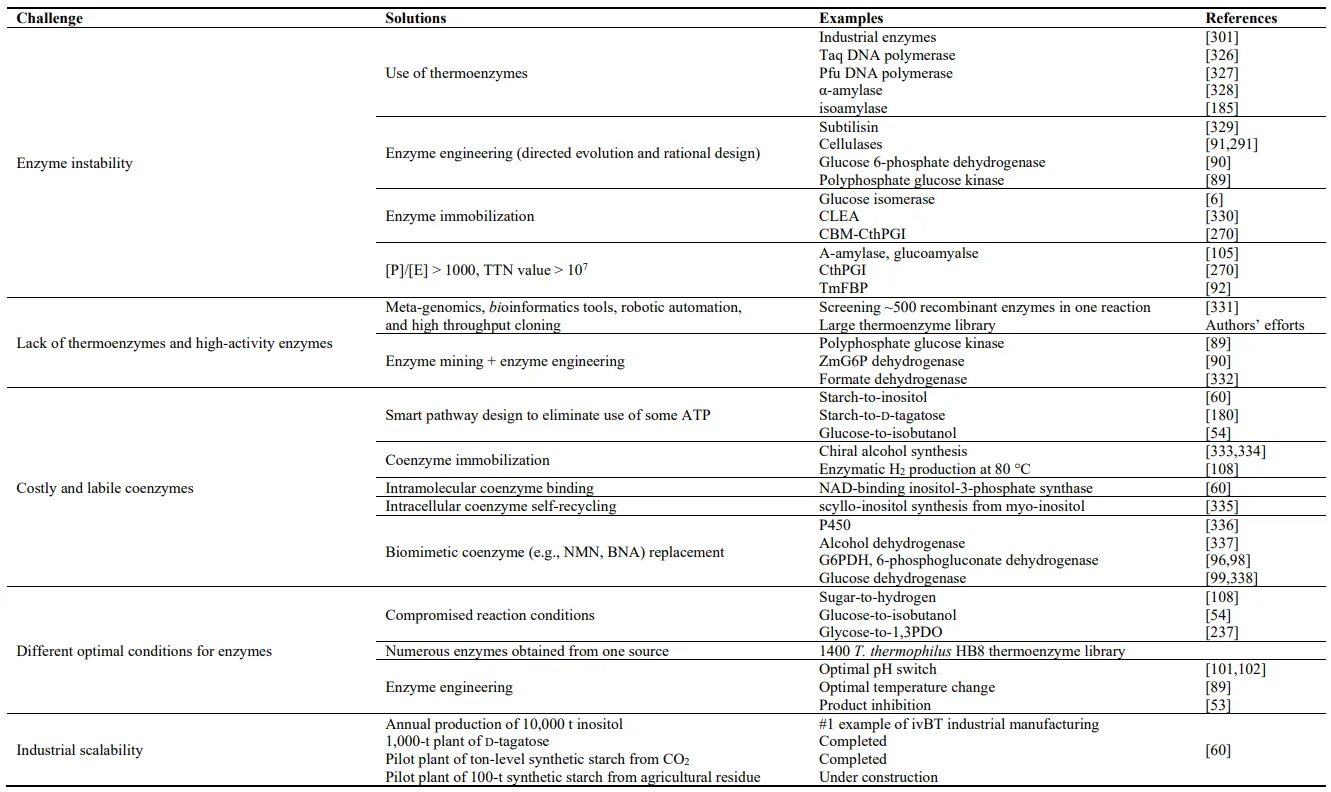
ivBT usually prefers the use of ultra-stable enzymes from thermophilic enzymes or more stable mutants obtained by enzyme engineering because ivSEBs typically last for a long time from weeks to months (). In contrast, CEB often prefers the use of mesophilic enzymes because (1) enzyme costs account for a small fraction of high prices of products (), (2) most of products or their intermediates are unstable at elevated temperature, and (3) enzyme choices from mesophilic organisms are far more than from thermophilic microorganisms.
Building a large library of ultra-stable enzymes is the foundation of ivBT. It is preferred to discover ultra-stable enzymes from thermophilic and hyper-thermophilic microorganisms whose growth temperatures vary from 55 to up to 121 °C [
268]. Several websites have provided good collections for characterized enzymes and putative enzyme sources, for example, BRENDA, Uniport, Genbank, the KEGG, and protein data bank (PDB). According to our experience, we prefer cloning putative enzymes from some thermophilic microorganisms whose optimal growth temperature ranges from 55 °C to nearly 100 °C and whose physiology and microbial ecology are widely diverse. Such representative thermophilic microorganisms are
Hungateiclostridium thermocellum (its old name
Clostridium thermocellum) [
269,
270],
Thermoanaerobacterium saccharolyticum [
271],
Geobacillus stearothermophilus [
272],
Acidithiobacillus ferrooxidans [
273,
274],
Archaeoglobus fulgidus [
275,
276],
Methanocaldococcus jannaschii [
277,
278],
Sulfolobus tokodaii [
185,
279],
Pyrococcus furiosus [
280,
281],
Thermotoga maritima [
75,
92,
179,
282,
283],
Thermus thermophilus [
284,
285],
Thermococcus kodakarensis [
69,
286,
287,
288,
289], and so on.
Alternatively, ultra-stable enzymes can be engineered by directed evolution, when targeted enzymes are available only from mesophilic source. For example, the most thermostable polyphosphate glucose kinase in nature was discovered from a modest thermophilic bacterium
Thermobifida fusca [
186], but it was not stable enough at 70 °C. Via directed evolution, the best mutant had a 7200-fold longer half-life at 55 °C than the wild-type PPGK [
89]. Directed evolution without knowing protein structures has been used to increase thermostability of numerous mesophilic enzymes, such as esterase of
B. subtilis [
290], beta-glucosidase of
Paenibacillus polymyxa [
91], endoglucanase of
Clostridium phytofermentans [
291], and so on.
Recent advances, especially ΑFold and machine learning, greatly decrease technical challenges to increase thermostability by rational design [
292,
293]. Before, the semi-rational design based on consensus design was widely used to enhance enzyme stability [
294,
295]. For example, the most thermostable sucrose phosphorylase with an optima temperature of 58 °C was discovered from
Bifidobacterium adolescentis [
296]. Later, the half-life time of this enzyme mutant was increased to 62 h at 60 °C by using the semi-rational design strategy [
297]. With highly accurate protein structure prediction by ΑFold [
298], machine learning [
132,
299,
300] is playing more and more important role by the introduction of non-covalent interactions and covalent bonds, increase of proline and/or decrease in glycine, reinforcement of subunit-subunit interactions, hydrogen bond, salt bridge, and hydrophobic interaction, filling the hydrophobic cavity core, decreasing surface hydrophobicity, truncating loop, introduction of glycosylation sites, truncation and cyclization, and so on [
93,
95].
4.2. Low-cost Production of Enzymes
Low-cost enzymes are prerequisite for the biomanufacturing success of ivBT. Now a large number of enzymes have been produced by bacteria, fungal, yeast, and plant platforms [
105,
301]. Among them, biomanufacturing costs of secretory enzymes are as low as 100 CNY per kg of dry weight enzyme, for example, α-amylase and subtilisin produced by
Bacillus sp. [
302,
303,
304], cellulase and hemocellulase produced by
Trichoderma reesei [
305,
306], phytase by
Pichia pastoris [
307,
308], glucoamylase, proteinase, and beta-glucosidase by
Aspergillus spp. [
309].
Production costs of recombinant intracellular enzymes is a little higher than secretory enzymes and its production cost are as low as approximately 250–300 CNY per kg dry weight enzyme on large scales [
3]. Such cost estimates have been validated independently in several enzyme-producing companies in China. Intracellular expression of recombinant proteins can be produced in high-cell density fermentation of mesophilic bacteria
E. coli or
B. subtilis that grow in low-cost industrial media [
306,
310,
311]. So many different strategies have been developed to increase soluble expression of targeted proteins, for example, optimizations of fermentation conditions (e.g., inducer concentration, timing, temperature, etc.) [
312], fusion proteins [
313,
314], expression plasmids and hosts, codon optimization [
75], rare-codon introduction [
315], co-expression of chaperones [
316], directed evolution [
317], and so on. Recently, we succeeded in transferring the
E. coli T7 expression system into a
B. subtilis host, which can overexpress some inclusion body-forming enzymes better than
E. coli [
318].
In addition of enzyme production, low-cost protein purification methods have been developed for intracellular recombinant enzymes (), such as heat precipitation for thermostable enzymes [
319,
320], ammonia precipitation [
114], one-step purification and immobilization [
186,
270,
321], carrier-free enzyme complex purification and co-immobilization [
322], so on. Among them, simple and less-costly heat precipitation may be the most appealing for the purification of thermostable enzymes [
319,
320,
323,
324]. Prof. Honda and his coworkers attempted to co-expressing up to nine thermoenzymes in one
E. coli cell [
325]. After heat precipitation, they harvested the thermoenzyme cocktail suitable for ivBT [
325]. By using the similar strategy, they prepared the cell lysate containing thermophilic enzyme cocktails to make CoA
in vitro [
282].
. The technical challenges of ivBT and their respective solutions and supportive examples.
Now the most challenging task of the overexpression of recombinant oxygen-sensitive enzymes include hydrogenases [
339,
340,
341,
342], nitrogenases [
340,
343], formate dehydrogenase [
332,
344,
345,
346], and so on. They have to be overexpressed under anaerobic conditions. Anaerobic cell culture has much lower cell yields and its high-cell density fermentation is difficult to achieve [
347]. Taking SH1 of Pyroccus furiosus [
348] as an example, its expression and purification from its wild-type host was costly and technically challenging because of low cell yields and short exponential and stationary phases [
347]. Two hundred liters of the wild-type
P. furiosus cell culture produced about 7500 U of purified SHI [
347]. Later, Prof. Adam developed the engineered
P. furiosus that can over-express His-tagged SHI by 100-fold [
349]. Later, they improved its active expression level by 40% with increased expression of maturation genes [
350]. To avoid fermenting
P. furious at nearly boiling temperature, Dr. Song and Adams succeeded in expressing active SHI in E. coli by co-expression of thirteen
P. furiosus genes (i.e., four structural genes encoding the hydrogenase and nine encoding maturation proteins) [
351]. Recently, we developed another alternative to express
P. furiosus SH1 in a hyperthermophilic archaeon
T. kodakarensis [
69]. The engineered microorganism had more than 1200-fold enhancement in the hydrogenase activity of the cell lysate and approximately 9000 U of 12-His-tagged were purified from five liters of the cell culture [
69]. In a word, future R&D of hydrogenase production could focus on high-density fermentation, over-expression of ultra-high-activity thermostable hydrogenase, large-scale fermentation, as well as artificial hydrogenases [
352,
353,
354,
355].
4.3. Enzyme Immobilization
Enzyme immobilization technologies have been developed for more than a half century. A variety of techniques include physical adsorption, cross-linked enzyme aggregates, covalent binding to support structures, such as microspheres, nanogels, nanocrystals, metal-organic frameworks (MOFs) [
356,
357], covalent organic frameworks (COFs) [
358,
359], hydrogen-bonded organic frameworks (HOFs) [
357], and so on [
106,
107,
360,
361]. Immobilized enzyme has clear biomanufacturing advantages: prolonging the lifetime of enzyme, decreasing biocatalyst cost, and simplifying the biocatalyst/product separation. The most successful example of enzyme immobilization is glucose isomerase (Sweetzyme IT) made by Novozymes [
6]. This enzyme can make up to 15,000 kg HFCS (F42, i.e., 42% fructose) per kg of immobilized enzyme at ~55 °C, that is [P]/[E] = 6300:1. The enzyme cost is as low as 20 CNY per metric ton of F42 (dry weight) and less than 0.5% of the product price.
A combination of low-cost production of thermostable enzymes and immobilization can lead to ultra-stable enzyme parts suitable for ivBT. For example, we developed a simple one-step purification and immobilization of cellulose-binding module (CBM)-tagged thermostable enzymes on solid cellulosic material [
270]. It was found that the total (turn-over number (TTN) of immobilized CBM-phosphoglucose isomerase from
Clostridium thermocellum was as high as 1.1 × 10
9 at 60 °C [
270], that is, [P]/[E] = 300,000 kg product per kg of immobilized enzyme.
Multi-enzyme co-immobilization is another practical choice. Enzyme components can be randomly distributed [
362,
363] and positionally assembled [
363] on solid supports. The enhanced reaction rates among co-immobilization of cascade enzymes have been observed for several cases [
83,
364,
365], but direct cross linking of these enzymes to solid supports may lead to the activity loss . For biomanufacturing of inositol, we developed a biomimetic mineralized microcapsules containing the four-enzyme cocktail (), whose lumped enzyme half-life of the microcapsules was 55.5 h at 70 °C, 5.9 folds of that of the non-immobilized four-enzyme cocktail [
153]. A fed-batch of the substrate can lead to an inositol titer of up to 210 g/L and its biomanufacturing cost was decreased to a half [
153]. Most enzyme immobilization techniques seem to stay on the stage of trial-and-error. It is highly expected to develop low-cost general enzyme co-immobilization suitable for most enzyme parts of ivSEB.
4.4. Artificial Enzyme Complexes
Constructing artificial enzyme complexes could bring multiple benefits, such as increase the volumetric rate mainly due to metabolite channeling effects, protecting vulnerable enzymes or metabolites in defined microenvironments, sequestering toxic, labile or volatile intermediates [
41,
83,
84,
366].
There are numerous methods to construct enzyme complexes. (1) The simplest way to obtain multi-function enzymes is the creation of fusion proteins, in which two or more cascade enzymes are combined by a linker to form a multi-functional single polypeptide [
367,
368,
369]. Similar effects have been reported in other fusion enzymatic systems [
369,
370,
371,
372,
373]. However, the misfolding of large multi-domain protein often happens and their apparent activities often decrease [
374], resulting in a large uncertainty of this strategy. (2) Scaffolding molecules, includes proteins and nucleic acids, can recruit enzymes to form multienzyme complexes [
84,
375,
376]. The recruited enzymes usually fused with a relatively small protein tag without obvious activity loss. Inspired by cellulosomes [
377], a synthetic scaffold containing different cohesins from different microorganisms is used for the assembly of three metabolic enzymes [
84]: triosephosphate isomerase, aldolase, and fructose 1,6-bisphosphatase are engineered to have a dockerin at their C-terminals. These three dockerin-containing enzymes can be self-assembled into a static trifunctional enzyme complex through the interaction with a mini-scaffold protein consisting of three different matching cohesins. The synthetic metabolon showed more than one order of magnitude enhancements in reaction rates compared to the non-complexed enzyme mixture [
84]. Recently, many polypeptide interaction pairs, such as SpyCatcher with SpyTag [
378], RIAD with RIDD [
379], and PDZ with PDZlig [
380], have been reported for the assembly of enzyme complex. (3) Both of DNA and RNA can be easily designed to fold into various structures
in vitro, forming simple structures such as sheets to very more complicated structures such as tubes and capsules [
375,
376]. It is more convenient to
in vitro synthesize oligonucleotide as DNA or RNA scaffolds by using automate oligonucleotide synthesis machines than protein scaffolds. However, the cost of DNA and RNA synthesis is far more expensive than that of protein scaffolds, which may impair their applications on a large scale, especially for the production of biocommodities. Using nucleic acids for the construction of enzyme complex have been described by a recent review [
381]. Recently, Prof. You developed a method for carrier-free immobilization of multienzyme complex for converting starch to inositol. Through the utilization of polypeptide interactions (SpyCatcher/SpyTag and dockerin/cohesin) and enzyme component self-oligomerization, the multi-enzyme complex could form significant precipitation, which could be recycled easily by simple centrifugation [
322].
4.5. NAD(P) Engineering
Living cells use NAD and NADP as electron carriers of catabolism and anabolism, respectively. To construct ivBT, NAD is a preferred carrier to NADP due to better stability and lower cost [
44]. Protein engineering for NAD(P) preference has been implemented for long. A number of examples include from NADP to NAD [
97,
173,
382,
383,
384,
385,
386,
387,
388], from NAD to NADP [
389,
390,
391,
392,
393], and relaxed or broadened the cofactor specificity [
90,
337,
394,
395,
396,
397]. To further enhance its stability, NAD can be conjugated with dehydrogenases by chemical bonds, whereas its total-turn-over number could be as high as 100,000 at 80 °C [
108].
Despite advances in NAD(P) engineering and immobilization, high cost of coenzymes NAD(P) may be the last obstacle to industrial biomanufacturing of biocommodities by ivBT. ivBT and CEB have different requirement for coenzymes, the latter can use natural NAD or NADP for the biosynthesis of high-value products. The use of less-costly and more stable biomimetic nicotinamide coenzymes (BNCs) is perquisite of industrial biomanufacturing of biocommodities, such as isobutanol, 1,3-propanediol, organic acids, and hydrogen [
44,
398,
399]. Many BNCs, such as nicotinamide mononucleotide (NMN), 1-benzyl-3-carbamoyl-pyridinium (BCP), 1-buta-3-carbamoyl-pyridinium (BuCP), 1-phenylethylnicotinamide, 1-(3-phenylpropyl) nicotinamide, nicotinamide flucytosine dinucleotide, nicotinamide cytosine dinucleotide, carba-NAD(P), and so on, have been investigated to replace NAD(P) for the hydride transfer [
400,
401,
402,
403,
404].
It is worth mentioning that a significant fraction of wild-type flavin-containing oxidoreductases can utilize small-size biomimetic coenzymes (e.g., NMN and BCP), such as wild-type old yellow enzyme (OYE) [
405,
406], water-forming NADH oxidase [
407], and P450 mutants [
336]. The catalytic efficiency of these OYEs on these BNCs could be better than those on natural coenzymes [
399,
408].
Most wild-type dehydrogenases that did not contain flavin rarely utilize small-size BNCs, such as NMN and BCP. The activity of horse liver alcohol dehydrogenase on BCP may be the first report [
409]. However, this discovery was in question because the wild-type enzyme may contain a small amount of NAD [
410]. The Sieber group (another group working on ivBT for long) engineered glucose dehydrogenase from
Sulfolobus solfataricus with two mutation sites, which exhibits a ~10-fold activity increase on BCP
+ over the wild-type enzyme [
68]. Later, this enzyme was further improved with a 160-fold enhancement by the same group [
99].
Because few NAD-dependent wild-type dehydrogenases have been reported to have very low activities on NMN [
411,
412], the Scott group applied rational design to
P. furiosus alcohol dehydrogenase and obtained a two-amino acid mutant that can generate NMNH from NMN
+ [
408]. Later, Li and his coworkers applied rational design to glucose dehydrogenase with a 107-fold coenzyme specificity switch toward NMN
+ over NADP
+ [
413],
E. coli glutathione reductase [
414], and
Pseudomonas stutzeri phosphite dehydrogenase [
414].
In contrast to rational design, directed evolution is a powerful tool to modify enzymes’ substrate preference without detailed understanding of their catalytic mechanisms. To address coenzyme stability and cost issue of ivBT [
44], the Zhang group also has worked on this area for long. First, they conducted coenzyme engineering to change coenzyme preference from NADP to NAD for two dehydrogenases (i.e., glucose 6-phosphate dehydrogenase [
90,
108] and 6-phosphogluconate dehydrogenase [
97]). Second, their coenzymes were changed to NMN by semi-rational design and directed evolution [
96]. To facilitate high-throughput screening of dehydrogenases on NMN without noises from inherent NAD, they developed a convenient Petri-dish plate-based method, involving heat lysis of cell colonies on plates, colony filter paper reprinting, colony washings, a colorimetric enzyme-coupled assay, and digital imaging [
96]. The
T. maritima 6-phosphogluconate dehydrogenase mutant on NMN
+ was improved by a 50-fold enhancement in catalytic efficiency and had a specific activity of 17.7 U/mg on NMN
+, comparable to that of the wild-type 6PGDH on its natural coenzyme NADP [
96]. Recently, the
Zymomonas mobilis glucose-6-phosphate dehydrogenase was engineered to work on NMN [
98]. Until now, the high-yield biohydrogen pathway could be implemented by using NMN rather than NAD and NADP, implying a great cut in its production cost. (Note: Because NMN is a hot nutritional compound, its manufacturing cost has been decreased to 500 CNY per kg in China.)
To address the coenzyme stability issue for ivBT, Prof. Sieber suggested another way—the use of a very stable biomimetic coenzyme carba-NADP+, which has a 33-times longer half-life time of NADP at 50 °C [
338]. Many wild-type NAD(P)-dependent dehydrogenases can use this coenzyme without protein engineering [
338].
4.6. Addition of Organelles
Although the key idea of ivBT is to simplify complicated living cells for implementing biomanufacturing purpose, it is not surprising that ivSEBs would evolve to more complicated for more applications. For example, membrane-bound thylakoids in chloroplasts are the photosynthetic organelles of green algae and plants, producing NADPH and ATP for its cellular activities. Prof. Zhu and his coworkers added thylakoid membranes as a green engine to drive a five-enzyme cascade that converted acetate to poly(3-hydroxybutyrate) (PHB). The molar conversion efficiency of carbon of 80.0% and the light-energy conversion efficiency of 3.04% were achieved [
415]. Similarly, another work coupled thylakoid membranes with an ATP- and NADPH-dependent unnatural CO
2 fixation pathway constituted by 16 enzymes and achieved the production of glycolate from CO
2 and light energy [
416]. These results show great promise of using thylakoid membranes, with good mechanical stability, cost-effective extraction and purification, and the ability to co-regenerate ATP and NADPH driven by light. Furthermore, there are opportunities of coupling enzymes with other organelles, such as mitochondria, peroxisome, or lysosome for reaction compartmentalization.
4.7. System Optimization and Modeling
Experimental conditions of ivSEB, such as temperature, pH, optimal metal ions, and enzyme components can be optimized for the high yield and fast reaction rate with one variable changed followed by another as conducted elsewhere [
80].
Mathematical modeling can be conducted for ivSEBs at multiple levels from molecules to modules to systems [
366]. The relative simplicity of ivSEB compared to whole cells makes them far easier to simulate processes and predict optimal enzyme ratios [
44,
417]. Taking enzymatic hydrogen as an example which has the starting point of ivBT, Prof. Zhang and his coworker [
262] developed a nonlinear kinetic model to analyze rate-limiting steps, suggesting that under the same unit loading conditions hydrogenase and two dehydrogenases were responsible for catalyzing rate-limiting steps [
262]. Furthermore, they fit the model with experimental data by using a genetic algorithm, identified the most important enzymes by a global sensitivity analysis, used optimizing enzyme ratios to increase model by volumetric productivity of hydrogen by more than 10-fold [
169]. Later, Prof. Zeng and his coworker [
418] further improved this
in silico model by using a genetic algorithm to solve a multi-objective optimization. One set of solutions predicted to increase the reaction rate to 355 mmol/L/h by nearly another 10-fold [
418]. Experimental data demonstrated the fastest biohydrogen rate of more than 500 mmol H
2/L/h by a factor of 1000 [
108].
5. Opportunities
Here we briefly discuss opportunities of the supporting technologies for ivBT.
First, it is needed to further develop ultra-low cost production of recombinant enzymes. Industrial production costs of some secretory enzymes (e.g., protease, α-amylase, cellulase, glucoamylase, phytase, etc.) are as low as 100 CNY per kg of enzyme (dry weight) and of intracellular recombinant enzymes are as low as 250 CNY per kg of enzyme [
3,
51]. We expect that it is possible to make the least costly recombinant enzymes by transgenic plants with anticipated production costs of as low as 50 CNY per kg of dry protein weight [
3,
419,
420,
421]. The low-cost production of oxygen-sensitive enzyme complexes (e.g., hydrogenase, nitrogenase, CO
2 reductase) is still challenging on a large scale.
Second,
in vitro regeneration of reduced biomimetic coenzymes replacing current enzymatic or chemical regeneration is more and more important because the use of electricity eliminates the need for a sacrificial electron donor and results in the potentially low cost, ease of separation and improved system simplicity [
422]. The electrochemical NAD(P)H regeneration suffer from high NAD costs due to the formation of inactive dimers and 1,6-NAD(P)H, high overpotential, low faradaic efficiency [
423]. This challenge may be partially addressed by the electrode surface modification [
424], the use of redox mediators, such as viologen-based redox hydrogel [
425] and redox polymer [
426]. It is worth noting that the best solution may develop biomimetic coenzymes that cannot form a dimer when they are reduced by electricity plus coenzyme engineering of NAD-dependent dehydrogenases.
Third, it is essentially important to regenerate ATP
in vitro, especially for ATP-intensive biotransformation, such as the CO
2-to-starch biosynthesis [
53]. Although substrate phosphorylation for ATP regeneration has been widely used for
in vitro enzymatic biocatalysis [
61], it is not suitable for some applications. Inspired by ATP synthase-based photophosphorylation and oxidative phosphorylation [
427], it is appealing to synthesize ATP by using generate a proton gradient across an artificial membrane embedded ATP synthase. One of pioneering efforts was the synthesis of ATP by using
F1F0-ATPase powered by proton transmembrane gradient to drive ATP-synthase-involved phosphorylation [
428,
429,
430]. In the past decade, considerable efforts have been devoted to the biomimetic assembly of
F1F0-ATPase in artificial membrane structures, including liposome, polymersome, microsphere/microcapsule-supported lipid bilayers, layer-by-layer assembled multilayer-supported lipid bilayer, and planar solid-supported lipid bilayers [
431]. It is hypothesized that developing a novel electro-phosphorylation method that uses electricity to generate proton gradient across a dual-functional membrane and ATP synthesis by a
F1F0-ATPase embedded the artificial membrane. If this technology is developed, more ivBT applications are expected. In 2023, Prof. Erb designed a minimal electrobiological 3–4 enzyme module that allows direct regeneration of ATP from electricity. This breakthrough opens a great door to the industrialization of ivBT for cost-competitive production of more products [
432].
Fourth, it is a great opportunity to carry out ivBT in continuous flow microreactors on the scale of submillimeters to submicrometers due to better process control, high quality data with integrated sensors, better mass phenomena for spatial and temporal control of laminar flow [
433,
434]. Much higher volumetric productivity can be achieved due to very large enzyme to substrate ratio and large surface/volume ratio to eliminate diffusion-related limitations [
435,
436].
Lastly, there are so many opportunities in ivBT, such as the development of biomimetic CoA so that CoA-dependent enzymes can be used in ivSEB, artificial electron transfer chains (ETCs) for fast reaction rate and Faraday efficiency [
108], artificial membranes embedded with membrane enzymes [
431], organelles [
415], and so on.
6. Conclusions
Now humankind has such great needs as the sustainable development, the carbon-neutral renewable energy system, and food security to feed more than 8 billion people. This new biomanufacturing platform ivBT features theoretical product yields, high energy efficiencies, and potential low-biomanufacturing costs. To partially address food security, the economically viable biomanufacturing of ivBT of agriculture residues to edible food/feed would come into being within this decade (). Furthermore, the partial land switch from the cultivation of annual grain crops to the cultivation of perennial crops and next biorefining could address food security, decrease fresh water withdrawal, upside down carbon-emitting modern agriculture to carbon farming, maintain better biodiversity, and so on [
28]. Large-scale implementation of CO
2-to-starch artificial photosynthesis could lead to the starch-centered carbon-neutral energy system (), which would solve challenges, such as electricity and hydrogen storage challenges, CO
2 utilization, fresh water conservation, and maintenance of a small closed ecosystem for human survival in emergency situations [
29,
30]. We believe that ivBT is becoming a disruptive biomanufacturing platform to address several of the most important challenges in the sustainability revolution.
. Schematic presentation of the starch-centered carbon-neutral economy [
29,
52], where starch is a high-density hydrogen carrier, an electricity storage compound, food/feed source and a bioplastic component. In it, ivBT would play a central role in artificial photosynthesis for starch synthesis [
53] and reverse artificial photosynthesis for production of hydrogen [
47,
252] or electricity [
30,
52,
67].
Acknowledgments
This manuscript cannot be published with the full support of TIB, CAS. This study was jointly funded by the National Key Research and Development Program of China (2022YFA0912300), Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project, China (TSBICIP-CXRC-067), Major Project of Haihe Laboratory of Synthetic Biology (22HHSWSS00015), and National Natural Science Foundation of China (32271544).
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This research was funded by the National Key Research and Development Program of China grant number 2022YFA0912300, Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (China) grant number TSBI-CIP-CXRC-067, Major Project of Haihe Laboratory of Synthetic Biology (China) grant number 22HHSWSS00015, and National Natural Science Foundation of China grant number 32271544.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests.
References
-
1.
Clomburg JM, Crumbley AM, Gonzalez R. Industrial biomanufacturing: The future of chemical production.
Science 2017,
355, aag0804.
[Google Scholar]
-
2.
Scown CD. Prospects for carbon-negative biomanufacturing.
Trends Biotechnol. 2022,
40, 1415–1424.
[Google Scholar]
-
3.
Zhang Y-HP, Sun J, Ma Y. Biomanufacturing: history and perspective.
J. Ind. Microbiol. Biotechnol. 2017,
44, 773–784.
[Google Scholar]
-
4.
Sugii S, Wong CYQ, Lwin AKO, Chew LJM. Alternative fat: redefining adipocytes for biomanufacturing cultivated meat.
Trends Biotechnol. 2023,
41, 686–700.
[Google Scholar]
-
5.
Gervasi T, Pellizzeri V, Calabrese G, Di Bella G, Cicero N, Dugo G. Production of single cell protein (SCP) from food and agricultural waste by using
Saccharomyces cerevisiae.
Nat. Prod. Res. 2018,
32, 648–653.
[Google Scholar]
-
6.
Vasic-Racki D. History of industrial biotransformations—Dreams and realities. In Industrial Biotransformations; Wiley-VCH: Weinheim, Germany, 2006; pp. 1–37.
-
7.
Bornscheuer UT, Huisman GW, Kazlauskas RJ, Lutz S, Moore JC, Robins K. Engineering the third wave of biocatalysis.
Nature 2012,
485, 185–194.
[Google Scholar]
-
8.
Pereira N, Lima AC, Lanceros-Mendez S, Martins P. Magnetoelectrics: Three Centuries of Research Heading towards the 4.0 Industrial Revolution.
Materials 2020,
13, 4033.
[Google Scholar]
-
9.
Lake F. From industry 4.0 to lab 4.0.
Biotechniques 2019,
66, 247.
[Google Scholar]
-
10.
Schwab K. The Fourth Industrial Revolution; World Economic Forum: Geneva, Switzerland, 2016.
-
11.
Awang GM, Jones GA, Ingledew WM. The acetone-butanol-ethanol fermentation.
Crit. Rev. Microbiol. 1988,
15, S33–S67.
[Google Scholar]
-
12.
Buchholz K, Collins J. The roots—a short history of industrial microbiology and biotechnology.
Appl. Microbiol. Biotechnol. 2013,
97, 3747–3762.
[Google Scholar]
-
13.
Ezeji TC, Qureshi N, Blaschek HP. Bioproduction of butanol from biomass: from genes to bioreactors.
Curr. Opin. Biotechnol. 2007,
18, 220–227.
[Google Scholar]
-
14.
Zhang Y-HPJ. Remembering Professor Daniel IC Wang’s contribution to biorefining and my opinions in perspective of biorefining.
Syn. Biol. J. 2021,
2, 497–508.
[Google Scholar]
-
15.
Demain AL. Microbial biotechnology.
Trends Biotechnol. 2000,
18, 26–31.
[Google Scholar]
-
16.
Demain AL, Newcomb M, Wu JHD. Cellulase, clostridia, and ethanol.
Microbiol. Mol. Biol. Rev. 2005,
69, 124–154.
[Google Scholar]
-
17.
Loenen WA, Dryden DT, Raleigh EA, Wilson GG, Murray NE. Highlights of the DNA cutters: a short history of the restriction enzymes.
Nucleic Acids Res. 2014,
42, 3–19.
[Google Scholar]
-
18.
Heckmann CM, Paradisi F. Looking Back: A Short History of the Discovery of Enzymes and How They Became Powerful Chemical Tools.
ChemCatChem 2020,
12, 6082–6102.
[Google Scholar]
-
19.
Lee SH, Yoon KH. A Century of Progress in Diabetes Care with Insulin: A History of Innovations and Foundation for the Future.
Diabetes Metab. J. 2021,
45, 629–640.
[Google Scholar]
-
20.
Demain AL. The business of biotechnology.
Ind. Biotechnol. 2007,
3, 269–283.
[Google Scholar]
-
21.
Afeyan NB, Cooney CL. Professor Daniel I.C. Wang: A legacy of education, innovation, publication, and leadership.
Biotechnol. Bioeng. 2006,
95, 206–217.
[Google Scholar]
-
22.
The World Economic Forum Water Initiative. Water Security: The Water-food-energy-climate Nexus; Island Press: Washington, DC, USA, 2011.
-
23.
-
24.
Wang F, Harindintwali JD, Yuan Z, Wang M, Wang F, Li S, et al. Technologies and perspectives for achieving carbon neutrality.
Innovation 2021,
2, 100180.
[Google Scholar]
-
25.
Zhao C, Liu B, Piao S, Wang X, Lobell DB, Huang Y, et al. Temperature increase reduces global yields of major crops in four independent estimates.
Proc. Natl. Acad. Sci. USA 2017,
114, 9326–9331.
[Google Scholar]
-
26.
Lynd LR, Beckham GT, Guss AM, Jayakody LN, Karp EM, Maranas C, et al. Toward low-cost biological and hybrid biological/catalytic conversion of cellulosic biomass to fuels.
Energy Environ. Sci. 2022,
15, 938–990.
[Google Scholar]
-
27.
Sartbaeva A, Kuznetsov VL, Wells SA, Edwards PP. Hydrogen nexus in a sustainable energy future.
Energy Environ. Sci. 2008,
1, 79–85.
[Google Scholar]
-
28.
Chen H-G, Zhang Y-HP. New biorefineries and sustainable agriculture: Increased food, biofuels, and ecosystem security.
Renew. Sust. Energy Rev. 2015,
47, 117–132.
[Google Scholar]
-
29.
Zhang Y-HP. Next generation biorefineries will solve the food, biofuels, and environmental trilemma in the energy-food-water nexus.
Energy Sci. Eng. 2013,
1, 27–41.
[Google Scholar]
-
30.
Zhang Y-HP, Huang W-D. Constructing the electricity-carbohydrate-hydrogen cycle for a sustainability revolution.
Trends Biotechnol. 2012,
30, 301–306.
[Google Scholar]
-
31.
Bazilian M, Rogner H, Howells M, Hermann S, Arent D, Gielen D, et al. Considering the energy, water and food nexus: Towards an integrated modelling approach.
Energy Policy 2011,
39, 7896–7906.
[Google Scholar]
-
32.
Stern F. Fritz Haber: Flawed Greatness of Person and Country.
Angew. Chem. Int. Ed. 2012,
51, 50–56.
[Google Scholar]
-
33.
Demain AL. Pickles, pectin, and penicillin.
Annu. Rev. Microbiol. 2004,
58, 1–42.
[Google Scholar]
-
34.
Gottschalk U, Brorson K, Shukla AA. The need for innovation in biomanufacturing.
Nat. Biotechnol. 2012,
30, 489–492.
[Google Scholar]
-
35.
Gartland KMA, Gartland JS. Opportunities in biotechnology.
J. Biotechnol. 2018,
282, 38–45.
[Google Scholar]
-
36.
National Academies of Sciences, Engineering, and Medicine; Policy and Global Affairs; Government-University-Industry Research Roundtable. The Fourth Industrial Revolution: Proceedings of a Workshop—in Brief; National Academies Press: Washington, DC, USA; 2017.
-
37.
Lynd LR, Wyman CE, Gerngross TU. Biocommodity engineering.
Biotechnol. Prog. 1999,
15, 777–793.
[Google Scholar]
-
38.
Zhang Y-HP. Production of biocommodities and bioelectricity by cell-free synthetic enzymatic pathway biotransformations: Challenges and opportunities.
Biotechnol. Bioeng. 2010,
105, 663–677.
[Google Scholar]
-
39.
Frazzetto G. White biotechnology.
EMBO Rep. 2003,
4, 835–837.
[Google Scholar]
-
40.
Bauer MW. Distinguishing Red and Green Biotechnology: Cultivation Effects of the Elite Press.
Int. J. Public Opin. Res. 2005,
17, 63–89.
[Google Scholar]
-
41.
Zhang Y-HP. Simpler is better: high-yield and potential low-cost biofuels production through cell-free synthetic pathway biotransformation (SyPaB).
ACS Catal. 2011,
1, 998–1009.
[Google Scholar]
-
42.
Zhang Y-HP. Production of biofuels and biochemicals by in vitro synthetic biosystems: Opportunities and challenges.
Biotechnol. Adv. 2015,
33, 1467–1483.
[Google Scholar]
-
43.
Zhang Y-HP, Myung S, You C, Zhu ZG, Rollin J. Toward low-cost biomanufacturing through cell-free synthetic biology: bottom-up design.
J. Mater. Chem. 2011,
21, 18877–18886.
[Google Scholar]
-
44.
Rollin JA, Tam W, Zhang Y-HP. New biotechnology paradigm: cell-free biosystems for biomanufacturing.
Green Chem. 2013,
15, 1708–1719.
[Google Scholar]
-
45.
Lovelock SL, Crawshaw R, Basler S, Levy C, Baker D, Hilvert D, et al. The road to fully programmable protein catalysis.
Nature 2022,
606, 49–58.
[Google Scholar]
-
46.
Thauer K, Jungermann K, Decker K. Energy conservation in chemotrophic anaerobic bacteria.
Bacteriol. Rev. 1977,
41, 100–180.
[Google Scholar]
-
47.
Zhang Y-HP, Evans BR, Mielenz JR, Hopkins RC, Adams MWW. High-yield hydrogen production from starch and water by a synthetic enzymatic pathway.
PLoS One 2007,
2, e456.
[Google Scholar]
-
48.
Kim J-E, Kim E-J, Chen H, Wu C-H, Adams MWW, Zhang Y-HP. Advanced water splitting for green hydrogen gas production through complete oxidation of starch by in vitro metabolic engineering.
Metab. Eng. 2017,
44, 246–252.
[Google Scholar]
-
49.
Desvaux M, Guedon E, Petitdemange H. Cellulose catabolism by Clostridium cellulolyticum growing in batch culture on defined medium.
Appl. Environ. Microbiol. 2000,
66, 2461–2470.
[Google Scholar]
-
50.
You C, Chen H, Myung S, Sathitsuksanoh N, Ma H, Zhang X-Z, et al. Enzymatic transformation of nonfood biomass to starch.
Proc. Natl. Acad. Sci. USA 2013,
110, 7182–7187.
[Google Scholar]
-
51.
Xu X, Zhang W, You C, Fan C, Ji W, Park J-T, et al. Biosynthesis of artificial starch and microbial protein from agricultural residue.
Sci. Bull. 2023,
68, 214–223.
[Google Scholar]
-
52.
Song Y, Wu R, Wei X, Shi T, Li Y, You C, et al. Advances in a new energy system based on electricity-hydrogen-carbohydrate cycle.
Chin. J. Biotechnol. 2022,
38, 4081–4100.
[Google Scholar]
-
53.
Cai T, Sun H, Qiao J, Zhu L, Zhang F, Zhang J, et al. Cell-free chemoenzymatic starch synthesis from carbon dioxide.
Science 2021,
373, 1523–1527.
[Google Scholar]
-
54.
Guterl J-K, Garbe D, Carsten J, Steffler F, Sommer B, Reiße S, et al. Cell-free metabolic engineering—Production of chemicals via minimized reaction cascades.
ChemSusChem 2012,
5, 2165–2172.
[Google Scholar]
-
55.
Yang J, Song W, Cai T, Wang Y, Zhang X, Wang W, et al. De novo artificial synthesis of hexoses from carbon dioxide. Sci. Bull. 2023, in press. doi: 10.1016/j.scib.2023.08.023.
-
56.
Han P, Wang X, Li Y, Wu H, Shi T, Shi J. Synthesis of a healthy sweetener D-tagatose from starch catalyzed by semiartificial cell factories.
J. Agri. Food Chem. 2023,
71, 3813–3820.
[Google Scholar]
-
57.
Li Y, Shi T, Han P, You C. Thermodynamics-driven production of value-added D-allulose from inexpensive starch by an in vitro enzymatic synthetic biosystem.
ACS Catal. 2021,
11, 5088–5099.
[Google Scholar]
-
58.
Graham AE, Ledesma-Amaro R. The microbial food revolution.
Nat. Commun. 2023,
14, 2231.
[Google Scholar]
-
59.
Zhong C, You C, Wei P, Zhang Y-HP. Thermal cycling cascade biocatalysis of myo-inositol synthesis from sucrose.
ACS Catal. 2017,
7, 5992–5999.
[Google Scholar]
-
60.
You C, Shi T, Li Y, Han P, Zhou X, Zhang Y-HP. An in vitro synthetic biology platform for the industrial biomanufacturing of myo-inositol from starch.
Biotechnol. Bioeng. 2017,
114, 1855–1864.
[Google Scholar]
-
61.
Chen H-G, Zhang Y-HPJ. Enzymatic regeneration and conservation of ATP: challenges and opportunities.
Crit. Rev. Biotechnol. 2021,
41, 16–33.
[Google Scholar]
-
62.
Andexer JN, Richter M. Emerging enzymes for ATP regeneration in biocatalytic processes.
ChemBioChem 2015,
16, 380–386.
[Google Scholar]
-
63.
Welch P, Scopes RK. Studies on cell-free metabolism: Ethanol production by a yeast glycolytic system reconstituted from purified enzymes.
J. Biotechnol. 1985,
2, 257–273.
[Google Scholar]
-
64.
Allain EJ. Cell-free ethanol production: the future of fuel ethanol?
J. Chem. Technol. Biotechnol. 2007,
82, 117–120.
[Google Scholar]
-
65.
Jewett MC, Calhoun KA, Voloshin A, Wuu JJ, Swartz JR. An integrated cell-free metabolic platform for protein production and synthetic biology.
Mol. Syst. Biol. 2008,
4, 220.
[Google Scholar]
-
66.
Wichmann R, Vasic-Racki D. Cofactor regeneration at the lab scale.
Adv. Biochem. Eng. Biotechnol. 2005,
92, 225–260.
[Google Scholar]
-
67.
Zhu Z-G, Kin Tam T, Sun F, You C, Zhang Y-HP. A high-energy-density sugar biobattery based on a synthetic enzymatic pathway.
Nat. Commun. 2014,
5, 3026.
[Google Scholar]
-
68.
Nowak C, Pick A, Lommes P, Sieber V. Enzymatic reduction of nicotinamide biomimetic cofactors using an engineered glucose dehydrogenase: providing a regeneration system for artificial cofactors.
ACS Catal. 2017,
7, 5202–5208.
[Google Scholar]
-
69.
Song Y, Liu M, Xie L, You C, Sun J, Zhang Y-HPJ. A recombinant 12-His tagged Pyrococcus furiosus soluble [NiFe]-hydrogenase I overexpressed in
Thermococcus kodakarensis KOD1 facilitates hydrogen-powered in vitro NADH regeneration.
Biotechnol. J. 2019,
14, e1800301.
[Google Scholar]
-
70.
Anne A, Bourdillon C, Daninos S, Moiroux J. Can the combination of electrochemical regeneration of NAD+, selectivity of L-a-amino-acid dehydrogenase, and reductive amination of a-keto-acid be applied to the inversion of configuration of a L-a-amino-acid?
Biotechnol. Bioeng. 1999,
64, 101–107.
[Google Scholar]
-
71.
Tishkov VI, Popov VO. Protein engineering of formate dehydrogenase.
Biomol. Eng. 2006,
23, 89–110.
[Google Scholar]
-
72.
Wandrey C. Biochemical reaction engineering for redox reactions.
Chem. Rec. 2004,
4, 254–265.
[Google Scholar]
-
73.
Inoue K, Makino Y, Itoh N. Purification and characterization of a novel alcohol dehydrogenase from
Leifsonia sp. strain S749: a promising biocatalyst for an asymmetric hydrogen transfer bioreduction.
Appl. Environ. Microbiol. 2005,
71, 3633–3641.
[Google Scholar]
-
74.
Johannes TW, Woodyer RD, Zhao H. Directed evolution of a thermostable phosphite dehydrogenase for NAD(P)H regeneration.
Appl. Environ. Microbiol. 2005,
71, 5728–5734.
[Google Scholar]
-
75.
Wang Y, Zhang Y-HP. Overexpression and simple purification of the
Thermotoga maritima 6-phosphogluconate dehydrogenase in
Escherichia coli and its application for NADPH regeneration.
Microb. Cell Fact. 2009,
8, 30.
[Google Scholar]
-
76.
Wang Y, Huang W, Sathitsuksanoh N, Zhu Z, Zhang Y-HP. Biohydrogenation from biomass sugar mediated by in vitro synthetic enzymatic pathways.
Chem. Biol. 2011,
18, 372–380.
[Google Scholar]
-
77.
Huang H, Pandya C, Liu C, Al-Obaidi NF, Wang M, Zheng L, et al. Panoramic view of a superfamily of phosphatases through substrate profiling.
Proc. Natl. Acad. Sci. USA 2015,
112, E1974–E1983.
[Google Scholar]
-
78.
Verhees CH, Akerboom J, Schiltz E, de Vos WM, van der Oost J. Molecular and biochemical characterization of a distinct type of fructose-1,6-bisphosphatase from
Pyrococcus furiosus.
J. Bacteriol. 2002,
184, 3401–3405.
[Google Scholar]
-
79.
Tian C, Yang J, Liu C, Chen P, Zhang T, Men Y, et al. Engineering substrate specificity of HAD phosphatases and multienzyme systems development for the thermodynamic-driven manufacturing sugars.
Nat. Commun. 2022,
13, 3582.
[Google Scholar]
-
80.
Wang W, Liu M, You C, Li Z, Zhang Y-HP. ATP-free biosynthesis of a high-energy phosphate metabolite fructose 1,6-diphosphate by in vitro metabolic engineering.
Metab. Eng. 2017,
42, 168–174.
[Google Scholar]
-
81.
Zhou W, You C, Ma H, Ma Y, Zhang Y-HP. One-pot biosynthesis of high-concentration α-glucose 1-phosphate from starch by sequential addition of three hyperthermophilic enzymes.
J. Agric. Food Chem. 2016,
64, 1777–1783.
[Google Scholar]
-
82.
Srivastava DK, Bernhard SA. Metabolite transfer via enzyme-enzyme complexes.
Science 1986,
234, 1081–1086.
[Google Scholar]
-
83.
Zhang Y-HP. Substrate channeling and enzyme complexes for biotechnological applications.
Biotechnol. Adv. 2011,
29, 715–725.
[Google Scholar]
-
84.
You C, Myung S, Zhang Y-HP. Facilitated substrate channeling in a self-assembled trifunctional enzyme complex.
Angew. Chem. Int. Ed. 2012,
51, 8787–8790.
[Google Scholar]
-
85.
Miles EW, Rhee S, Davies DR. The molecular basis of substrate channeling.
J. Biol. Chem. 1999,
274, 12193–12196.
[Google Scholar]
-
86.
Zhu Z, Song H, Wang Y, Zhang Y-HPJ. Protein engineering for electrochemical biosensors.
Curr. Opin. Biotechnol. 2022,
76, 102751.
[Google Scholar]
-
87.
Chen K, Arnold FH. Engineering new catalytic activities in enzymes.
Nat. Catal. 2020,
3, 203–213.
[Google Scholar]
-
88.
Arnold FH. Innovation by evolution: Bringing new chemistry to life (Nobel lecture).
Angew. Chem. Int. Ed. 2019,
58, 14420–14426.
[Google Scholar]
-
89.
Zhou W, Huang R, Zhu Z, Zhang Y-HPJ. Coevolution of both thermostability and activity of polyphosphate glucokinase from
Thermobifida fusca YX.
Appl. Environ. Microbiol. 2018,
84, e01224–e01218.
[Google Scholar]
-
90.
Huang R, Chen H, Zhou W, Ma C, Zhang Y-HP. Engineering a thermostable highly active glucose 6-phosphate dehydrogenase and its application to hydrogen production in vitro.
Appl. Microbiol. Biotechnol. 2018,
102, 3203–3215.
[Google Scholar]
-
91.
Liu W, Hong J, Bevan DR, Zhang Y-HP. Fast identification of thermostable beta-glucosidase mutants on cellobiose by a novel combinatorial selection/screening approach.
Biotechnol. Bioeng. 2009,
103, 1087–1094.
[Google Scholar]
-
92.
Myung S, Wang YR, Zhang Y-HP. Fructose-1,6-bisphosphatase from a hyper-thermophilic bacterium
Thermotoga maritima: Characterization, metabolite stability and its implications.
Proc. Biochem. 2010,
45, 1882–1887.
[Google Scholar]
-
93.
Xu Z, Cen YK, Zou SP, Xue YP, Zheng YG. Recent advances in the improvement of enzyme thermostability by structure modification.
Crit. Rev. Biotechnol. 2020,
40, 83–98.
[Google Scholar]
-
94.
Wheeler LC, Lim SA, Marqusee S, Harms MJ. The thermostability and specificity of ancient proteins.
Curr. Opin. Struct. Biol. 2016,
38, 37–43.
[Google Scholar]
-
95.
Nezhad NG, Rahman R, Normi YM, Oslan SN, Shariff FM, Leow TC. Thermostability engineering of industrial enzymes through structure modification.
Appl. Microbiol. Biotechnol. 2022,
106, 4845–4866.
[Google Scholar]
-
96.
Huang R, Chen H, Upp DM, Lewis JC, Zhang Y-HPJ. A high-throughput method for directed evolution of NAD(P)
+-dependent dehydrogenases for the reduction of biomimetic nicotinamide analogues.
ACS Catal. 2019,
9, 11709–11719.
[Google Scholar]
-
97.
Huang R, Chen H, Zhong C, Kim JE, Zhang Y-HP. High-throughput screening of coenzyme preference change of thermophilic 6-phosphogluconate dehydrogenase from NADP
+ to NAD
+.
Sci. Rep. 2016,
6, 32644.
[Google Scholar]
-
98.
Meng D, Liu M, Su H, Song H, Chen L, Li Q, et al. Coenzyme engineering of glucose-6-phosphate dehydrogenase on a nicotinamide-based biomimic and its application as a glucose biosensor.
ACS Catal. 2023,
13, 1983–1998.
[Google Scholar]
-
99.
Zachos I, Güner S, Essert A, Lommes P, Sieber V. Boosting artificial nicotinamide cofactor systems.
Chem. Commun. 2022,
58, 11945–11948.
[Google Scholar]
-
100.
Zachos I, Genth R, Sutiono S, Marczynski M, Lieleg O, Sieber V. Hot Flows: Evolving an Archaeal Glucose Dehydrogenase for Ultrastable Carba-NADP
+ Using Microfluidics at Elevated Temperatures.
ACS Catal. 2022,
12, 1841–1846.
[Google Scholar]
-
101.
Ma CL, Wu RR, Huang R, Jiang WX, You C, Zhu LL, et al. Directed evolution of a 6-phosphogluconate dehydrogenase for operating an enzymatic fuel cell at lowered anodic pHs.
J. Electroanal. Chem. 2019,
851, 113444.
[Google Scholar]
-
102.
Ma CL, Liu MX, You C, Zhu ZG. Engineering a diaphorase via directed evolution for enzymatic biofuel cell application.
Biores. Bioproc. 2020,
7, 1–11.
[Google Scholar]
-
103.
Li H, Du H, Wang X, Gao P, Liu Y, Lin W. Remarks on computational method for identifying acid and alkaline enzymes.
Curr. Pharm. Des. 2020,
26, 3105–3114.
[Google Scholar]
-
104.
Sakoda H, Imanaka T. Cloning and sequencing of the gene coding for alcohol dehydrogenase of Bacillus stearothermophilus and rational shift of the optimum pH.
J. Bacteriol. 1992,
174, 1397–1402.
[Google Scholar]
-
105.
Schäfer T, Borchert TW, Nielsen VS, Skagerlind P, Gibson K, Wenger K, et al. Industrial enzymes.
Adv. Biochem. Eng. Biotechnol. 2007,
105, 59–131.
[Google Scholar]
-
106.
Liese A, Hilterhaus L. Evaluation of immobilized enzymes for industrial applications.
Chem. Soc. Rev. 2013,
42, 6236–6249.
[Google Scholar]
-
107.
Arana-Peña S, Carballares D, Morellon-Sterlling R, Berenguer-Murcia Á, Alcántara AR, Rodrigues RC, et al. Enzyme co-immobilization: Always the biocatalyst designers’ choice…or not?
Biotechnol. Adv. 2021,
51, 107584.
[Google Scholar]
-
108.
Kim E-J, Kim J-E, Zhang Y-HPJ. Ultra-rapid rates of water splitting for biohydrogen gas production through in vitro artificial enzymatic pathways.
Energy Environ. Sci. 2018,
11, 2064–2072.
[Google Scholar]
-
109.
Woodley JM, Bisschops M, Straathof AJJ, Ottens M. Future directions for in-situ product removal (ISPR).
J. Chem. Technol. Biotechnol. 2008,
83, 121–123.
[Google Scholar]
-
110.
Deng X, Fan M, Wu M, Zhang X, Cheng Y, Xia J, et al. Continuous-flow enzymatic synthesis of chiral lactones in a three-dimensional microfluidic reactor. Chin. Chem. Lett. 2023. doi:10.1016/j.cclet.2023.108684.
-
111.
Li H-P, You Z-N, Liu Y-Y, Zheng G-W, Gong H, Mo Y, et al. Continuous-flow microreactor-enhanced clean NAD
+ regeneration for biosynthesis of 7-oxo-lithocholic acid.
ACS Sust. Chem. Eng. 2022,
10, 456–463.
[Google Scholar]
-
112.
Myung S, Zhang Y-HP. Non-complexed four cascade enzyme mixture: simple purification and synergetic co-stabilization.
PLoS ONE 2013,
8, e61500.
[Google Scholar]
-
113.
Ellis RJ. Macromolecular crowding: obvious but underappreciated.
Trends Biochem. Sci. 2001,
26, 597–604.
[Google Scholar]
-
114.
Minton AP. The Influence of Macromolecular Crowding and Macromolecular Confinement on Biochemical Reactions in Physiological Media.
J. Biol. Chem. 2001,
276, 10577–10580.
[Google Scholar]
-
115.
Freeman A, Woodley JM, Lilly MD. In Situ Product Removal as a Tool for Bioprocessing.
Nat. Biotechnol. 1993,
11, 1007–1012.
[Google Scholar]
-
116.
Buchner E. Alkoholische Gärung ohne Hefezellen (Vorläufige Mitteilung).
Berichte der Deutschen Chemischen Gesellschaft 1897,
30, 117–124.
[Google Scholar]
-
117.
Kresge N, Simoni RD, Hill RL. Otto Fritz Meyerhof and the Elucidation of the Glycolytic Pathway.
J. Biol. Chem. 2005,
280, e3.
[Google Scholar]
-
118.
Krebs HA, Eggleston LV. Metabolism of Acetoacetic Acid in Animal Tissues.
Nature 1944,
154, 209–210.
[Google Scholar]
-
119.
Krebs HA. The discovery of carbon dioxide fixation in mammalian tissues.
Mol. Cell. Biochem. 1974,
5, 79–94.
[Google Scholar]
-
120.
Berg P, Mertz JE. Personal reflections on the origins and emergence of recombinant DNA technology.
Genetics 2010,
184, 9–17.
[Google Scholar]
-
121.
Bartlett JM, Stirling D. A short history of the polymerase chain reaction.
Methods Mol. Biol. 2003,
226, 3–6.
[Google Scholar]
-
122.
Chen K, Arnold FH. Turning the activity of an enzyme for unusual environments: sequential random mutagenesis of subtilisin E for catalysis in dimethylformamide.
Proc. Natl. Acad. Sci. USA 1993,
90, 5618–5622.
[Google Scholar]
-
123.
Arnold FH. Directed evolution: Bringing new chemistry to life.
Angew. Chem. Int. Ed. 2018,
57, 4143–4148.
[Google Scholar]
-
124.
Yu X, Liu T, Zhu F, Khosla C. In vitro reconstitution and steady-state analysis of the fatty acid synthase from
Escherichia coli.
Proc. Nat. Acad. Sci. USA 2011,
108, 18643–18648.
[Google Scholar]
-
125.
Zhu F, Zhong X, Hu M, Lu L, Deng Z, Liu T. In vitro reconstitution of mevalonate pathway and targeted engineering of farnesene overproduction in
Escherichia coli.
Biotechnol. Bioeng. 2014,
111, 1396–1405.
[Google Scholar]
-
126.
Keller MW, Schut GJ, Lipscomb GL, Menon AL, Iwuchukwu IJ, Leuko TT, et al. Exploiting microbial hyperthermophilicity to produce an industrial chemical, using hydrogen and carbon dioxide.
Proc. Nat. Acad. Sci. USA 2013,
110, 5840–5845.
[Google Scholar]
-
127.
Bogorad IW, Lin T-S, Liao JC. Synthetic non-oxidative glycolysis enables complete carbon conservation.
Nature 2013,
502, 693–697.
[Google Scholar]
-
128.
Michels P, Rosazza J. The evolution of microbial transformations for industrial applications.
SIM News 2009,
2009, 36–52.
[Google Scholar]
-
129.
Fu J, Yang YR, Johnson-Buck A, Liu M, Liu Y, Walter NG, et al. Multi-enzyme complexes on DNA scaffolds capable of substrate channelling with an artificial swinging arm.
Nat. Nanotechnol. 2014,
9, 531–536.
[Google Scholar]
-
130.
Lin J-L, Palomec L, Wheeldon I.
Design and Analysis of Enhanced Catalysis in Scaffolded Multi-Enzyme Cascade
Reactions. ACS Catal. 2014, 4, 505–511.
-
131.
France SP, Hepworth LJ, Turner NJ, Flitsch SL. Constructing biocatalytic cascades: In vitro and in vivo approaches to de novo multi-enzyme pathways.
ACS Catal. 2017,
7, 710–724.
[Google Scholar]
-
132.
Woodley JM. Accelerating the implementation of biocatalysis in industry.
Appl. Microbiol. Biotechnol. 2019,
103, 4733–4739.
[Google Scholar]
-
133.
Wildeman SMAD, Sonke T, Schoemaker HE, May O. Biocatalytic reductions: From lab curiosity to “first choice”.
Acc. Chem. Res. 2007,
40, 1260–1266.
[Google Scholar]
-
134.
Bozic M, Pricelius S, Guebitz GM, Kokol V. Enzymatic reduction of complex redox dyes using NADH-dependent reductase from
Bacillus subtilis coupled with cofactor regeneration.
Appl. Microbiol. Biotechnol. 2010,
85, 563–571.
[Google Scholar]
-
135.
Xu Z, Jing K, Liu Y, Cen P. High-level expression of recombinant glucose dehydrogenase and its application in NADPH regeneration.
J. Ind. Microbiol. Biotechnol. 2007,
34, 83–90.
[Google Scholar]
-
136.
Mertens R, Liese A. Biotechnological applications of hydrogenases.
Curr. Opin. Biotechnol. 2004,
15, 343–348.
[Google Scholar]
-
137.
Johannes TW, Woodyer RD, Zhao H. Efficient regeneration of NADPH using an engineered phosphite dehydrogenase.
Biotechnol. Bioeng. 2007,
96, 18–26.
[Google Scholar]
-
138.
Nam KY, Struck DK, Holtzapple MT. ATP regeneration by thermostable ATP synthase.
Biotechnol. Bioeng. 1996,
51, 305–316.
[Google Scholar]
-
139.
Resnick SM, Zehnder AJ. In vitro ATP regeneration from polyphosphate and AMP by polyphosphate:AMP phosphotransferase and adenylate kinase from
Acinetobacter johnsonii 210A.
Appl. Environ. Microbiol. 2000,
66, 2045–2051.
[Google Scholar]
-
140.
Franke D, Machajewski T, Hsu C-C, Wong C-H. One-pot synthesis of L-fructose using coupled multienzyme systems based on rhamnulose-1-phosphate aldolase.
J. Org. Chem. 2003,
68, 6828–6831.
[Google Scholar]
-
141.
Schoevaart R, van Rantwijk F, Sheldon RA. A four-step enzymatic cascade for the one-pot synthesis of non-natural carbohydrates from glycerol.
J. Org. Chem. 2000,
65, 6940–6943.
[Google Scholar]
-
142.
Huang K-T, Wu B-C, Lin C-C, Luo S-C, Chen C, Wong C-H, et al. Multi-enzyme one-pot strategy for the synthesis of sialyl Lewis X-containing PSGL-1 glycopeptide.
Carbohydr. Res. 2006,
341, 2151–2155.
[Google Scholar]
-
143.
Zhang J, Shao J, Kowal P, Wang PG. Enzymatic Synthesis of Oligosaccharides; Wiley-VCH: Weinheim, Germany, 2005.
-
144.
Fessner W-D, Helaine V. Biocatalytic synthesis of hydroxylated natural products using aldolases and related enzymes.
Curr. Opin. Biotechnol. 2001,
12, 574–586.
[Google Scholar]
-
145.
Fessner W-D. Enzyme mediated C–C bond formation.
Curr. Opin. Chem. Biol. 1998,
2, 85–97.
[Google Scholar]
-
146.
Endo T, Koizumi S. Large-scale production of oligosaccharides using engineered bacteria.
Curr. Opin. Struct. Biol. 2000,
10, 536–541.
[Google Scholar]
-
147.
Fessner W-D. Systems biocatalysis: development and engineering of cell-free “artificial metabolisms” for preparative multi-enzymatic synthesis.
New Biotechnol. 2015,
32, 658–664.
[Google Scholar]
-
148.
Tessaro D, Pollegioni L, Piubelli L, D’Arrigo P, Servi S. Systems biocatalysis: An artificial metabolism for interconversion of functional groups.
ACS Catal. 2015,
5, 1604–1608.
[Google Scholar]
-
149.
Opgenorth PH, Korman TP, Bowie JU. A synthetic biochemistry molecular purge valve module that maintains redox balance.
Nat. Commun. 2014,
5, 4113.
[Google Scholar]
-
150.
Korman TP, Opgenorth PH, Bowie JU. A synthetic biochemistry platform for cell free production of monoterpenes from glucose.
Nat. Commun. 2017,
8, 15526.
[Google Scholar]
-
151.
Koeller KM, Wong C-H. Enzymes for chemical synthesis.
Nature 2001,
409, 232–240.
[Google Scholar]
-
152.
Ayala-Aguilera CC, Valero T, Lorente-Macías Á, Baillache DJ, Croke S, Unciti-Broceta A. Small molecule kinase inhibitor drugs (1995–2021): medical indication, pharmacology, and synthesis.
J. Med. Chem. 2022,
65, 1047–1131.
[Google Scholar]
-
153.
Han P, You C, Li Y, Shi T, Wu H, Zhang Y-HPJ. High-titer production of myo-inositol by a co-immobilized four-enzyme cocktail in biomimetic mineralized microcapsules.
Chem. Eng. J. 2023,
461, 141946.
[Google Scholar]
-
154.
Teixeira CSS, Sousa SF. Current status of the use of multifunctional enzymes as anti-cancer drug targets.
Pharmaceutics 2021,
14, 10.
[Google Scholar]
-
155.
Swartz JR. Cell-free bioprocessing.
Chem. Eng. Prog. 2013,
2013, 40–45.
[Google Scholar]
-
156.
Carlson ED, Gan R, Hodgman CE, Jewett MC. Cell-free protein synthesis: Applications come of age.
Biotechnol. Adv. 2012,
30, 1185–1194.
[Google Scholar]
-
157.
Shimizu Y, Inoue A, Tomari Y, Suzuki T, Yokogawa T, Nishikawa K, et al. Cell-free translation reconstituted with purified components.
Nat. Biotechnol. 2001,
19, 751–755.
[Google Scholar]
-
158.
Dudley QM, Karim AS, Jewett MC. Cell-free metabolic engineering: Biomanufacturing beyond the cell.
Biotechnol. J. 2015,
10, 69–82.
[Google Scholar]
-
159.
Rasor BJ, Vögeli B, Landwehr GM, Bogart JW, Karim AS, Jewett MC. Toward sustainable, cell-free biomanufacturing.
Curr. Opin. Biotechnol. 2021,
69, 136–144.
[Google Scholar]
-
160.
Karim AS, Jewett MC. A cell-free framework for rapid biosynthetic pathway prototyping and enzyme discovery.
Metab. Eng. 2016,
36, 116–126.
[Google Scholar]
-
161.
Harris DC, Jewett MC. Cell-free biology: exploiting the interface between synthetic biology and synthetic chemistry.
Curr. Opin. Biotechnol. 2012,
12, 672–678.
[Google Scholar]
-
162.
Chiba CH, Knirsch MC, Azzoni AR, Moreira AR, Stephano MA. Cell-free protein synthesis: advances on production process for biopharmaceuticals and immunobiological products.
Biotechniques 2021,
70, 126–133.
[Google Scholar]
-
163.
Pardee K, Slomovic S, Nguyen Peter Q, Lee Jeong W, Donghia N, Burrill D, et al. Portable, on-demand biomolecular manufacturing.
Cell 2016,
167, 248–259.
[Google Scholar]
-
164.
Stamatis C, Farid SS. Process economics evaluation of cell-free synthesis for the commercial manufacture of antibody drug conjugates.
Biotechnol. J. 2021,
16, e2000238.
[Google Scholar]
-
165.
Stech M, Rakotoarinoro N, Teichmann T, Zemella A, Thoring L, Kubick S. Synthesis of fluorescently labeled antibodies using non-canonical amino acids in eukaryotic cell-free systems.
Methods Mol. Biol. 2021,
2305, 175–190.
[Google Scholar]
-
166.
Lüddecke T, Paas A, Talmann L, Kirchhoff KN, von Reumont BM, Billion A, et al. A Spider Toxin Exemplifies the Promises and Pitfalls of Cell-Free Protein Production for Venom Biodiscovery.
Toxins 2021,
13, 575.
[Google Scholar]
-
167.
Ramm F, Jack L, Kaser D, Schloßhauer JL, Zemella A, Kubick S. Cell-Free Systems Enable the Production of AB(5) Toxins for Diagnostic Applications.
Toxins 2022,
14, 233.
[Google Scholar]
-
168.
Pe’ery T, Mathews MB. Synthesis and purification of single-stranded RNA for use in experiments with PKR and in cell-free translation systems.
Methods 1997,
11, 371–381.
[Google Scholar]
-
169.
Rollin JA, Martin del Campo J, Myung S, Sun F, You C, Bakovic A, et al. High-yield hydrogen production from biomass by in vitro metabolic engineering: Mixed sugars coutilization and kinetic modeling.
Proc. Natl. Acad. Sci. USA 2015,
112, 4964–4969.
[Google Scholar]
-
170.
Opgenorth PH, Korman TP, Bowie JU. A synthetic biochemistry module for production of bio-based chemicals from glucose.
Nat. Chem. Biol. 2016,
12, 393–395.
[Google Scholar]
-
171.
Cheng K, Zheng W, Chen H, Zhang Y-HPJ. Upgrade of wood sugar D-xylose to a value-added nutraceutical by in vitro metabolic engineering.
Metab. Eng. 2019,
52, 1–8.
[Google Scholar]
-
172.
Shi T, Liu S, Zhang Y-HPJ. CO
2 fixation for malate synthesis energized by starch via in vitro metabolic engineering.
Metab. Eng. 2019,
55, 152–160.
[Google Scholar]
-
173.
Kim E-J, Adams M, Wu C-H, Zhang Y-HP. Exceptionally high rates of biological hydrogen production by biomimetic in vitro synthetic enzymatic pathways.
Chemistry 2016,
22, 16047–16051.
[Google Scholar]
-
174.
Opgenorth PH, Korman TP, Iancu L, Bowie JU. A molecular rheostat maintains ATP levels to drive a synthetic biochemistry system.
Nat. Chem. Biol. 2017,
13, 938–942.
[Google Scholar]
-
175.
Hold C, Billerbeck S, Panke S. Forward design of a complex enzyme cascade reaction.
Nat. Commun. 2016,
7, 12971.
[Google Scholar]
-
176.
Holtzapple MT, Cognata M, Shu Y, Hendrickson C. Inhibition of Trichoderma reesei cellulase by sugars and solvents.
Biotechnol. Bioeng. 1990,
36, 275–287.
[Google Scholar]
-
177.
Serdakowski A, Dordick J. Enzyme activation for organic solvents made easy.
Trends Biotechnol. 2008,
26, 48–54.
[Google Scholar]
-
178.
Garcia V, Pakkila J, Ojamo H, Muurinen E, Keiski RL. Challenges in biobutanol production: How to improve the efficiency?
Renew. Sustain. Energy Rev. 2011,
15, 964–980.
[Google Scholar]
-
179.
Fujisawa T, Fujinaga S, Atomi H. An in vitro enzyme system for the production of myo-inositol from starch.
Appl. Environ. Microbiol. 2017,
83, e00550–e00517.
[Google Scholar]
-
180.
Wichelecki DJ, Zhang YHP. Enzymatic synthesis of D-tagatose. World Patent WO2017059278A1, 2015.
-
181.
Zhang Y-HP, You C. Inositol preparation method. China Patent CN106148425B, 2015.
-
182.
Zhang Y-HP, Zhou W. D-xylulose 4-epimerase, mutants and applications. World Patent WO2021135796A1, 2019.
-
183.
Zhan S, Li Y, Li Y, Cui X, Zhong J-J, Zhang Y-HPJ. Aminomutation catalyzed by CO2 self-sufficient cascade amino acid decarboxylases. bioRxiv 2023. doi: 10.1101/2023.08.12.552924.
-
184.
Colodny L, Hoffman RL. Inositol—clinical applications for exogenous use.
Altern. Med. Rev. 1998,
3, 432–447.
[Google Scholar]
-
185.
Cheng K, Zhang F, Sun F-F, Zhu Z-G, Chen H-G, Zhang Y-HP. Doubling power output of starch biobattery treated by the most thermostable isoamylase from an archaeon
Sulfolobus tokodaii.
Sci. Rep. 2015,
5, 13184 .
[Google Scholar]
-
186.
Liao HH, Myung S, Zhang Y-HP. One-step purification and immobilization of thermophilic polyphosphate glucokinase from
Thermobifida fusca YX: glucose-6-phosphate generation without ATP.
Appl. Microbiol. Biotechnol. 2012,
93, 1109–1117.
[Google Scholar]
-
187.
Flamholz A, Noor E, Bar-Even A, Milo R. eQuilibrator—the biochemical thermodynamics calculator.
Nucl. Acids Res. 2012,
40, D770–D775.
[Google Scholar]
-
188.
You R, Wang L, Shi C, Chen H, Zhang S, Hu M, et al. Efficient production of myo-inositol in Escherichia coli through metabolic engineering.
Microb. Cell Fact. 2020,
19, 109.
[Google Scholar]
-
189.
Tang E, Shen X, Wang J, Sun X, Yuan Q. Synergetic utilization of glucose and glycerol for efficient myo-inositol biosynthesis.
Biotechnol. Bioeng. 2020,
117, 1247–1252.
[Google Scholar]
-
190.
Han P, Zhou X, You C. Efficient multi-enzymes imobilized on Porous microspheres for producing inositol From starch.
Front. Bioeng. Biotechnol. 2020,
8, 380.
[Google Scholar]
-
191.
Lu Y, Wang L, Teng F, Zhang J, Hu M, Tao Y. Production of myo-inositol from glucose by a novel trienzymatic cascade of polyphosphate glucokinase, inositol 1-phosphate synthase and inositol monophosphatase.
Enzyme Microb. Technol. 2018,
112, 1–5.
[Google Scholar]
-
192.
Meng D, Wei X, Zhang Y-HPJ, Zhu Z, You C, Ma Y. Stoichiometric conversion of cellulosic biomass by in vitro synthetic enzymatic biosystems for biomanufacturing.
ACS Catal. 2018,
8, 9550–9559.
[Google Scholar]
-
193.
Granström TB, Takata G, Tokuda M, Izumori K. Izumoring: A novel and complete strategy for bioproduction of rare sugars.
J. Biosci. Bioeng. 2004,
97, 89–94.
[Google Scholar]
-
194.
Zhang W, Zhang T, Jiang B, Mu W. Enzymatic approaches to rare sugar production.
Biotechnol. Adv. 2017,
35, 267–274.
[Google Scholar]
-
195.
Izumori K. Izumoring: a strategy for bioproduction of all hexoses.
J. Biotechnol. 2006,
124, 717–722.
[Google Scholar]
-
196.
Levin GV. Tagatose, the new GRAS sweetener and health product.
J. Med. Food 2002,
5, 23–36.
[Google Scholar]
-
197.
Cheetham PSJ, Wootton AN. Bioconversion of D-galactose into D-tagatose.
Enzyme Microb. Technol. 1993,
15, 105–108.
[Google Scholar]
-
198.
Rhimi M, Aghajari N, Juy M, Chouayekh H, Maguin E, Haser R, et al. Rational design of
Bacillus stearothermophilus US100 L-arabinose isomerase: Potential applications for D-tagatose production.
Biochim. 2009,
91, 650–653.
[Google Scholar]
-
199.
Men Y, Zhu Y, Zhang L, Kang Z, Izumori K, Sun Y, et al. Enzymatic conversion of D-galactose to D-tagatose: Cloning, overexpression and characterization of L-arabinose isomerase from
Pediococcus pentosaceus PC-5.
Microbiol. Res. 2014,
169, 171–178.
[Google Scholar]
-
200.
Chan HC, Zhu Y, Hu Y, Ko TP, Huang CH, Ren F, et al. Crystal structures of D-psicose 3-epimerase from
Clostridium cellulolyticum H10 and its complex with ketohexose sugars.
Protein Cell 2012,
3, 123–131.
[Google Scholar]
-
201.
Men Y, Zhu Y, Guan Y, Zhang T, Izumori K, Sun Y. Screening of food-grade microorganisms for biotransformation of D-tagatose and cloning and expression of L-arabinose isomerase.
Sheng Wu Gong Cheng Xue Bao 2012,
28, 592–601.
[Google Scholar]
-
202.
Li Y, Zhu Y, Liu A, Sun Y. Identification and characterization of a novel L-arabinose isomerase from
Anoxybacillus flavithermus useful in D-tagatose production.
Extremophiles 2011,
15, 441–450.
[Google Scholar]
-
203.
Sun Y, Zhu Y. Nucleotide sequence of Clostridium D-tagatose 3-epimerase and application thereof. China Patent CN102373230A, 2010.
-
204.
Sun Y, Zhu Y. Nucleotide sequence of D-tagatose-3-epimerase (DTE) of Ruminococcus sp. and use thereof. China Patent CN103131721B, 2011.
-
205.
Bosshart A, Hee CS, Bechtold M, Schirmer T, Panke S. Directed divergent evolution of a thermostable D-tagatose epimerase towards improved activity for two hexose substrates.
ChemBioChem 2015,
16, 592–601.
[Google Scholar]
-
206.
Oh H-J, Kim H-J, Oh D-K. Increase in D-tagatose Production Rate by Site-directed Mutagenesis of L-arabinose Isomerase from
Geobacillus thermodenitrificans.
Biotechnol. Lett. 2006,
28, 145–149.
[Google Scholar]
-
207.
Wichelecki DJ, Vetting MW, Chou L, Al-Obaidi N, Bouvier JT, Almo SC, et al. ATP-binding cassette (ABC) transport system solute-binding protein-guided identification of novel D-altritol and galactitol catabolic pathways in
Agrobacterium tumefaciens C58.
J. Biol. Chem. 2015,
290, 28963–28976.
[Google Scholar]
-
208.
Oh D-K, Hong S-H, Lee S-H. Aldolase, aldolase mutant, and method and composition for producing tagatose by using same. WO2015016544 A1. US Patent 2015.
-
209.
Moradian A, Benner SA. A biomimetic biotechnological process for converting starch to fructose: thermodynamic and evolutionary considerations in applied enzymology.
J. Am. Chem. Soc. 1992,
114, 6980–6987.
[Google Scholar]
-
210.
Ma Y, Sun Y. The preparation method of tagatose. China Patent CN106399427B, 2016.
-
211.
Ma Y, Sun Y, Yang J, Li Y. Method for preparing tagatose through whole-cell catalysis. China Patent CN 107988286B, 2017.
-
212.
Ma Y, Shi T, Li Y, Han P, Li Y. Bacillus subtilis gene engineering bacteria for producing tagatose and method for preparing tagatose. CN112342179B. 2021.
-
213.
Ma Y, Sun Y, Yang J, Li Y. Engineering strain for producing tagatose, and construction method and application thereof. China Patent CN109666620B. 2019.
-
214.
Dai Y, Zhang T, Jiang B, Mu W, Chen J, Hassanin HA.
Dictyoglomus turgidum DSM 6724 α-Glucan Phosphorylase: Characterization and Its Application in Multi-enzyme Cascade Reaction for D-Tagatose Production.
Appl. Biochem. Biotechnol. 2021,
193, 3719–3731.
[Google Scholar]
-
215.
Dai Y, Zhang J, Zhang T, Chen J, Hassanin HA, Jiang B. Characteristics of a fructose 6-phosphate 4-epimerase from
Caldilinea aerophila DSM 14535 and its application for biosynthesis of tagatose.
Enzyme Microb. Technol. 2020,
139, 109594.
[Google Scholar]
-
216.
Dai Y, Li C, Zheng L, Jiang B, Zhang T, Chen J. Enhanced biosynthesis of D-tagatose from maltodextrin through modular pathway engineering of recombinant
Escherichia coli.
Biochem. Eng. J. 2022,
178, 108303.
[Google Scholar]
-
217.
Zhang W, Yu S, Zhang T, Jiang B, Mu W. Recent advances in D-allulose: Physiological functionalities, applications, and biological production.
Trends Food Sci. Technol. 2016,
54, 127–137.
[Google Scholar]
-
218.
Jiang S, Xiao W, Zhu X, Yang P, Zheng Z, Lu S, et al. Review on D-Allulose: In vivo Metabolism, Catalytic Mechanism, Engineering Strain Construction, Bio-Production Technology.
Front. Bioeng. Biotechnol. 2020,
8, 26.
[Google Scholar]
-
219.
Matsuo T, Suzuki H, Hashiguchi M, Izumori K. D-psicose is a rare sugar that provides no energy to growing rats.
J. Nutr. Sci. Vitaminol. 2002,
48, 77–80.
[Google Scholar]
-
220.
Zeng Y, Zhang X, Guan Y, Sun Y. Characteristics and antioxidant activity of Maillard reaction products from psicose-lysine and fructose-lysine model systems.
J. Food Sci. 2011,
76, C398–C403.
[Google Scholar]
-
221.
Hayashi N, Iida T, Yamada T, Okuma K, Takehara I, Yamamoto T, et al. Study on the postprandial blood glucose suppression effect of D-psicose in borderline diabetes and the safety of long-term ingestion by normal human subjects.
Biosci. Biotechnol. Biochem. 2010,
74, 510–519.
[Google Scholar]
-
222.
Chung MY, Oh DK, Lee KW. Hypoglycemic health benefits of D-psicose.
J. Agric. Food Chem. 2012,
60, 863–869.
[Google Scholar]
-
223.
Moller DE, Berger JP. Role of PPARs in the regulation of obesity-related insulin sensitivity and inflammation.
Int. J. Obes. Relat. Metab. Disord. 2003,
27, S17–S21.
[Google Scholar]
-
224.
Yang S, Cho HK, Lee YM, Kim SB, Cho SJ. Thermostable fructose-6-phosphate-3-epimerase and a method for producing allulose using the same. US Patent 10907182, 2017.
-
225.
Wichelecki DJ, Rogers E. Enzymatic production of hexoses. World Patent WO2018169957A1, Patent 2017.
-
226.
Maceachran D, Cunningham DS, Blake WJ, Moura ME. Cell-free production of sugars. US Patent US20180320210A1, 2017.
-
227.
Torretta S, Scagliola A, Ricci L, Mainini F, Di Marco S, Cuccovillo I, et al. D-mannose suppresses macrophage IL-1β production.
Nat. Commun. 2020,
11, 6343.
[Google Scholar]
-
228.
Gonzalez PS, O’Prey J, Cardaci S, Barthet VJA, Sakamaki JI, Beaumatin F, et al. Mannose impairs tumour growth and enhances chemotherapy.
Nature 2018,
563, 719–723.
[Google Scholar]
-
229.
Zhang D, Chia C, Jiao X, Jin W, Kasagi S, Wu R, et al. D-mannose induces regulatory T cells and suppresses immunopathology.
Nat. Med. 2017,
23, 1036–1045.
[Google Scholar]
-
230.
Tian C, Yang J, Li Y, Zhang T, Li J, Ren C, et al. Artificially designed routes for the conversion of starch to value-added mannosyl compounds through coupling in vitro and in vivo metabolic engineering strategies.
Metab. Eng. 2020,
61, 215–224.
[Google Scholar]
-
231.
Liu H, Xu Y, Zheng Z, Liu D. 1,3-Propanediol and its copolymers: research, development and industrialization.
Biotechnol. J. 2010,
5, 1137–1148.
[Google Scholar]
-
232.
Nakamura CE, Whited GM. Metabolic engineering for the microbial production of 1,3-propanediol.
Curr. Opin. Biotech. 2003,
14, 454–459.
[Google Scholar]
-
233.
Dietz D, Zeng A-P. Efficient production of 1,3-propanediol from fermentation of crude glycerol with mixed cultures in a simple medium.
Bioproc. Biosyst. Eng. 2014,
37, 225–233.
[Google Scholar]
-
234.
Sabra W, Groeger C, Zeng AP. Microbial cell factories for diol production.
Adv. Biochem. Eng. Biotechnol. 2016,
155, 165–197.
[Google Scholar]
-
235.
Celińska E. Debottlenecking the 1,3-propanediol pathway by metabolic engineering.
Biotechnol. Adv. 2010,
28, 519–530.
[Google Scholar]
-
236.
Zeng A-P, Sabra W. Microbial production of diols as platform chemicals: Recent progresses.
Curr. Opin. Biotechnol. 2011,
22, 749–757.
[Google Scholar]
-
237.
Rieckenberg F, Ardao I, Rujananon R, Zeng A-P. Cell-free synthesis of 1,3-propanediol from glycerol with a high yield.
Eng. Life Sci. 2014,
14, 380–386.
[Google Scholar]
-
238.
Atsumi S, Hanai T, Liao JC. Non-fermentative pathways for synthesis of branched-chain higher alcohols as biofuels.
Nature 2008,
451, 86–89.
[Google Scholar]
-
239.
Hazelwood LA, Daran J-M, van Maris AJA, Pronk JT, Dickinson JR. The Ehrlich Pathway for Fusel Alcohol Production: a Century of Research on
Saccharomyces cerevisiae Metabolism.
Appl. Environ. Microbiol. 2008,
74, 2259–2266.
[Google Scholar]
-
240.
Peralta-Yahya PP, Zhang F, del Cardayre SB, Keasling JD. Microbial engineering for the production of advanced biofuels.
Nature 2012,
488, 320–328.
[Google Scholar]
-
241.
Li S, Wen J, Jia X. Engineering
Bacillus subtilis for isobutanol production by heterologous Ehrlich pathway construction and the biosynthetic 2-ketoisovalerate precursor pathway overexpression.
Appl. Microbiol. Biotechnol. 2011,
91, 577–589.
[Google Scholar]
-
242.
Smith K, Cho K-M, Liao J. Engineering
Corynebacterium glutamicum for isobutanol production.
Appl. Microbiol. Biotechnol. 2010,
87, 1045–1055.
[Google Scholar]
-
243.
Chen X, Nielsen K, Borodina I, Kielland-Brandt M, Karhumaa K. Increased isobutanol production in
Saccharomyces cerevisiae by overexpression of genes in valine metabolism.
Biotechnol. Biofuels 2011,
4, 21.
[Google Scholar]
-
244.
Atsumi S, Wu T-Y, Machado IMP, Huang W-C, Chen P-Y, Pellegrini M, et al. Evolution, genomic analysis, and reconstruction of isobutanol tolerance in
Escherichia coli.
Mol. Syst. Biol. 2010,
6, 449.
[Google Scholar]
-
245.
Xie L, Wei X, Zhou X, Meng D, Zhou R, Zhang Y-HPJ, et al. Conversion of D-glucose to L-lactate via pyruvate by an optimized cell-free enzymatic biosystem containing minimized reactions.
Syn. Syst. Biotechnol. 2018,
3, 204–210.
[Google Scholar]
-
246.
Ye X, Honda K, Morimoto Y, Okano K, Ohtake H. Direct conversion of glucose to malate by synthetic metabolic engineering.
J. Biotechnol. 2013,
164, 34–40.
[Google Scholar]
-
247.
-
248.
Somerville C, Youngs H, Taylor C, Davis SC, Long SP. Feedstocks for Lignocellulosic Biofuels.
Science 2010,
329, 790–792.
[Google Scholar]
-
249.
Sheppard AW, Gillespie I, Hirsch M, Begley C. Biosecurity and sustainability within the growing global bioeconomy.
Curr. Opin. Environ. Sustain. 2011,
3, 4–10.
[Google Scholar]
-
250.
Casillas CE, Kammen DM. The Energy-Poverty-Climate Nexus.
Science 2010,
330, 1181–1182.
[Google Scholar]
-
251.
Zhang Y-HP, You C, Chen H, Feng R. Surpassing photosynthesis: High-efficiency and scalable CO2 utilization through artificial photosynthesis. In Recent Advances in Post-Combustion CO2 Capture Chemistry; ACS Publications: Washington, DC, USA, 2012; Volume 1097; pp. 275–292.
-
252.
Zhang Y-HP. A sweet out-of-the-box solution to the hydrogen economy: is the sugar-powered car science fiction?
Energy Environ. Sci. 2009,
2, 272–282.
[Google Scholar]
-
253.
Harnisch F, Morejón MC. Hydrogen from Water is more than a Fuel: Hydrogenations and Hydrodeoxygenations for a Biobased Economy.
Chem. Rec. 2021,
21, 2277–2289.
[Google Scholar]
-
254.
Dou Y, Sun L, Ren J, Dong L. Chapter 10—Opportunities and Future Challenges in Hydrogen Economy for Sustainable Development. In Hydrogen Economy; Academic Press Cambridge, MA, USA, 2017; pp. 277–305.
-
255.
Tarascon J-M. Towards Sustainable and Renewable Systems for Electrochemical Energy Storage.
ChemSusChem 2008,
1, 777–779.
[Google Scholar]
-
256.
Zhang Y-HP. What is vital (and not vital) to advance economically-competitive biofuels production.
Proc. Biochem. 2011,
46, 2091–2110.
[Google Scholar]
-
257.
Thauer RK, Kaster AK, Seedorf H, Buckel W, Hedderich R. Methanogenic archaea: ecologically relevant differences in energy conservation.
Nat. Rev. Microbiol. 2008,
6, 579–591.
[Google Scholar]
-
258.
Chheda J, Huber G, Dumesic J. Liquid-phase catalytic processing of biomass-derived oxygenated hydrocarbons to fuels and chemicals.
Angew. Chem. Int. Ed. 2007,
46, 7164–7183.
[Google Scholar]
-
259.
Huber GW, Shabaker JW, Dumesic JA. Raney Ni-Sn catalyst for H2 production from biomass-derived hydrocarbons.
Science 2003,
300, 2075–2077.
[Google Scholar]
-
260.
Maeda T, Sanchez-Torres V, Wood TK. Hydrogen production by recombinant
Escherichia coli strains.
Microb. Biotechnol. 2012,
5, 214–225.
[Google Scholar]
-
261.
Maeda T, Sanchez-Torres V, Wood TK. Metabolic engineering to enhance bacterial hydrogen production.
Microb. Biotechnol. 2008,
1, 30–39.
[Google Scholar]
-
262.
Ye X, Wang Y, Hopkins RC, Adams MWW, Evans BR, Mielenz JR, et al. Spontaneous high-yield production of hydrogen from cellulosic materials and water catalyzed by enzyme cocktails.
ChemSusChem 2009,
2, 149–152.
[Google Scholar]
-
263.
Myung S, Rollin J, You C, Sun F, Chandrayan S, Adams MWW, Zhang Y-HP. In vitro metabolic engineering of hydrogen production at theoretical yield from sucrose.
Metab. Eng. 2014,
24, 70–77.
[Google Scholar]
-
264.
Rollin JA, Ye XH, Martin dCJS, Adams MWW, Zhang Y-HP. Novel hydrogen detection apparatus along with bioreactor Systems. In Bioreactor Engineering Research and Industrial Applications II; Springer: Berlin/Heidelberg, 2016.
-
265.
Martín del Campo JS, Rollin J, Myung S, Chun Y, Chandrayan S, Patiño R, et al. High-Yield Production of Dihydrogen from Xylose by Using a Synthetic Enzyme Cascade in a Cell-Free System.
Angew. Chem. Int. Ed. 2013,
52, 4587–4590.
[Google Scholar]
-
266.
Berezina OV, Zverlov VV, Lunina NA, Chekanovskaya LA, Dubinina EN, Liebl W, et al. Gene and properties of thermostable 4-α-glucanotransferase of
Thermotoga neapolitana.
Mol. Biol. 1999,
33, 801–806.
[Google Scholar]
-
267.
Chen H, Huang R, Kim E-J, Zhang Y-HPJ. Building a thermostable metabolon for facilitating coenzyme transport and in vitro hydrogen production at elevated temperature.
ChemSusChem 2018,
11, 3020–3030.
[Google Scholar]
-
268.
Takai K, Nakamura K, Toki T, Tsunogai U, Miyazaki M, Miyazaki J, et al. Cell proliferation at 122 °C and isotopically heavy CH
4 production by a hyperthermophilic methanogen under high-pressure cultivation.
Proc. Natl. Acad. Sci. USA 2008,
105, 10949–10954.
[Google Scholar]
-
269.
Alexander JK. Purification and specificity of cellobiose phosphorylase from
Clostridium thermocellum.
J. Biol. Chem. 1968,
243, 2899–2904.
[Google Scholar]
-
270.
Myung S, Zhang X-Z, Zhang Y-HP. Ultra-stable phosphoglucose isomerase through immobilization of cellulose-binding module-tagged thermophilic enzyme on low-cost high-capacity cellulosic adsorbent.
Biotechnol. Prog. 2011,
27, 969–975.
[Google Scholar]
-
271.
Verhaeghe T, Aerts D, Diricks M, Soetaert W, Desmet T. The quest for a thermostable sucrose phosphorylase reveals sucrose 6′-phosphate phosphorylase as a novel specificity.
Appl. Microbiol. Biotechnol. 2014,
98, 7027–7037.
[Google Scholar]
-
272.
Nazina T, Tourova T, Poltaraus A, Novikova E, Grigoryan A, Ivanova A, et al. Taxonomic study of aerobic thermophilic bacilli: descriptions of
Geobacillus subterraneus gen. nov., sp. nov. and
Geobacillus uzenensis sp. nov. from petroleum reservoirs and transfer of
Bacillus stearothermophilus,
Bacillus thermocatenulatus,
Bacillus thermoleovorans,
Bacillus kaustophilus
, Bacillus thermoglucosidasius and Bacillus thermodenitrificans to Geobacillus as the new combinations
G. stearothermophilus,
G. thermocatenulatus,
G. thermoleovorans,
G. kaustophilus,
G. thermoglucosidasius and
G. thermodenitrificans.
Int. J. Syst. Evol. Microbiol. 2001,
51, 433–446.
[Google Scholar]
-
273.
Zhang S, Yan L, Xing W, Chen P, Zhang Y, Wang W.
Acidithiobacillus ferrooxidans and its potential application.
Extremophiles 2018,
22, 563–579.
[Google Scholar]
-
274.
Quatrini R, Johnson DB.
Acidithiobacillus ferrooxidans.
Trends Microbiol. 2019,
27, 282–283.
[Google Scholar]
-
275.
Klenk H-P, Clayton RA, Tomb J-F, White O, Nelson KE, Ketchum KA, et al. The complete genome sequence of the hyperthermophilic, sulphate-reducing archaeon
Archaeoglobus fulgidus.
Nature 1997,
390, 364–370.
[Google Scholar]
-
276.
Chen L, Zhou C, Yang H, Roberts MF. Inositol-1-phosphate synthase from
Archaeoglobus fulgidus is a class II aldolase.
Biochemistry 2000,
39, 12415–12423.
[Google Scholar]
-
277.
Wang Y, Xu H, White RH. β-alanine biosynthesis in
Methanocaldococcus jannaschii.
J. Bacteriol. 2014,
196, 2869–2875.
[Google Scholar]
-
278.
Graham DE, Kyrpides N, Anderson IJ, Overbeek R, Whitman WB. Genome of
Methanocaldococcus (
Methanococcus)
jannaschii.
Methods Enzymol. 2001,
330, 40–123.
[Google Scholar]
-
279.
Kawarabayasi Y, Hino Y, Horikawa H, Jin-no K, Takahashi M, Sekine M, et al. Complete genome sequence of an aerobic thermoacidophilic crenarchaeon,
Sulfolobus tokodaii strain 7.
DNA Res. 2001,
8, 123–140.
[Google Scholar]
-
280.
Schut GJ, Lipscomb GL, Han Y, Notey JS, Kelly RM, Adams MMW. The order thermococcales and the family Thermococcaceae. In The Prokaryotes: Other Major Lineages of Bacteria and The Archaea; Springer: Berlin/Heidelberg, Germany, 2014; pp. 363–383.
-
281.
Kengen SWM.
Pyrococcus furiosus, 30 years on.
Microb. Biotechnol. 2017,
10, 1441–1444.
[Google Scholar]
-
282.
Suryatin Alim G, Iwatani T, Okano K, Kitani S, Honda K. In vitro production of coenzyme A using thermophilic enzymes.
Appl. Environ. Microbiol. 2021,
87, e0054121.
[Google Scholar]
-
283.
Nelson KE, Clayton RA, Gill SR, Gwinn ML, Dodson RJ, Haft DH, et al. Evidence for lateral gene transfer between Archaea and Bacteria from genome sequence of
Thermotoga maritima.
Nature 1999,
399, 323–329.
[Google Scholar]
-
284.
Cava F, Hidalgo A, Berenguer J.
Thermus thermophilus as biological model.
Extremophiles 2009,
13, 213–231.
[Google Scholar]
-
285.
Lioliou E, Pantazaki A, Kyriakidis D.
Thermus thermophilus genome analysis: benefits and implications.
Microb. Cell Fact. 2004,
3, 5.
[Google Scholar]
-
286.
Aono R, Sato T, Imanaka T, Atomi H. A pentose bisphosphate pathway for nucleoside degradation in Archaea.
Nat. Chem. Biol. 2015,
11, 355–360.
[Google Scholar]
-
287.
Rashid N, Aslam M. An overview of 25 years of research on
Thermococcus kodakarensis, a genetically versatile model organism for archaeal research.
Folia Microbiol. 2020,
65, 67–78.
[Google Scholar]
-
288.
Santangelo TJ, Cubonová Lu, Reeve JN.
Thermococcus kodakarensis genetics: TK1827-encoded beta-glycosidase, new positive-selection protocol, and targeted and repetitive deletion technology.
Appl. Environ. Microbiol. 2010,
76, 1044–1052.
[Google Scholar]
-
289.
Farkas JA, Picking JW, Santangelo TJ. Genetic techniques for the archaea.
Annu. Rev. Genet. 2013,
47, 539–561.
[Google Scholar]
-
290.
Given L, Gershenson A, Freskgard P-O, Arnold FH. Directed evolution of a thermostable esterase.
Proc. Natl. Acad. Sci. USA 1998,
95, 12809–12813.
[Google Scholar]
-
291.
Liu W, Zhang X-Z, Zhang Z-M, Zhang Y-HP. Engineering of
Clostridium phytofermentans endoglucanase Cel5A for improved thermostability.
Appl. Environ. Microbiol. 2010,
76, 4914–4917.
[Google Scholar]
-
292.
Wu Z, Kan SBJ, Lewis RD, Wittmann BJ, Arnold FH. Machine learning-assisted directed protein evolution with combinatorial libraries.
Proc. Natl. Acad. Sci. USA 2019,
116, 8852–8858.
[Google Scholar]
-
293.
Yang KK, Wu Z, Arnold FH. Machine-learning-guided directed evolution for protein engineering.
Nat. Methods 2019,
16, 687–694.
[Google Scholar]
-
294.
Porebski BT, Buckle AM. Consensus protein design.
Protein Eng. Des. Sel. 2016,
29, 245–251.
[Google Scholar]
-
295.
Goldenzweig A, Fleishman SJ. Principles of protein stability and their application in computational design.
Annu. Rev. Biochem. 2018,
87, 105–129.
[Google Scholar]
-
296.
Cerdobbel A, Desmet T, De Winter K, Maertens J, Soetaert W. Increasing the thermostability of sucrose phosphorylase by multipoint covalent immobilization.
J. Biotechnol. 2010,
150, 125–130.
[Google Scholar]
-
297.
Cerdobbel A, De Winter K, Aerts D, Kuipers R, Joosten HJ, Soetaert W, et al. Increasing the thermostability of sucrose phosphorylase by a combination of sequence- and structure-based mutagenesis.
Protein Eng. Des. Sel. 2011,
24, 829–834.
[Google Scholar]
-
298.
Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, et al. Highly accurate protein structure prediction with ΑFold.
Nature 2021,
596, 583–589.
[Google Scholar]
-
299.
Li G, Qin Y, Fontaine NT, Ng Fuk Chong M, Maria-Solano MA, Feixas F, et al. Machine Learning Enables Selection of Epistatic Enzyme Mutants for Stability Against Unfolding and Detrimental Aggregation.
Chembiochem 2021,
22, 904–914.
[Google Scholar]
-
300.
Siedhoff NE, Schwaneberg U, Davari MD. Machine learning-assisted enzyme engineering.
Methods Enzymol. 2020,
643, 281–315.
[Google Scholar]
-
301.
Demain AL, Vaishnav P. Production of recombinant proteins by microbes and higher organisms.
Biotechnol. Adv. 2009,
27, 297–306.
[Google Scholar]
-
302.
Su Y, Liu C, Fang H, Zhang D.
Bacillus subtilis: a universal cell factory for industry, agriculture, biomaterials and medicine.
Microb. Cell Fact. 2020,
19, 173.
[Google Scholar]
-
303.
Ngalimat MS, Yahaya RSR, Baharudin MMA, Yaminudin SM, Karim M, Ahmad SA, et al. A review on the biotechnological applications of the operational group
Bacillus amyloliquefaciens.
Microorganisms 2021,
9, 614.
[Google Scholar]
-
304.
Liu S, Wang J, Zhu Z, Shi T, Zhang Y-HPJ. Efficient secretory production of large-size heterologous enzymes in Bacillus subtilis: A secretory partner and directed evolution.
Biotechnol. Bioeng. 2020,
117, 2957–2968.
[Google Scholar]
-
305.
Zhang Y-HP, Himmel M, Mielenz JR. Outlook for cellulase improvement: Screening and selection strategies.
Biotechnol. Adv. 2006,
24, 452–481.
[Google Scholar]
-
306.
Tangnu SK, Blanch HW, Wilke CR. Enhanced production of cellulase, hemicellulase, and β-glucosidase by
Trichoderma reesei (Rut C-30).
Biotechnol. Bioeng. 1981,
23, 1837–1849.
[Google Scholar]
-
307.
Spohner SC, Müller H, Quitmann H, Czermak P. Expression of enzymes for the usage in food and feed industry with
Pichia pastoris.
J. Biotechnol. 2015,
202, 118–134.
[Google Scholar]
-
308.
Niu C, Yang P, Luo H, Huang H, Wang Y, Yao B. Engineering of Yersinia Phytases to Improve Pepsin and Trypsin Resistance and Thermostability and Application Potential in the Food and Feed Industry.
J. Agric. Food Chem. 2017,
65, 7337–7344.
[Google Scholar]
-
309.
Ueno S, Miyama M, Ohashi Y, Izumiya M, Kusaka I. Secretory enzyme production and conidiation of Aspergillus oryzae in submerged liquid culture.
Appl. Microbiol. Biotechnol. 1987,
26, 273–276.
[Google Scholar]
-
310.
Barnard G, Henderson G, Srinivasan S, Gerngross T. High level recombinant protein expression in
Ralstonia eutropha using T7 RNA polymerase based amplification.
Protein Expr. Purif. 2004,
38, 264–271.
[Google Scholar]
-
311.
Hartley JL. Cloning technologies for protein expression and purification.
Curr. Opin. Biotechnol. 2006,
17, 359–366.
[Google Scholar]
-
312.
Galloway CA, Sowden MP, Smith HC. Increasing the yield of soluble recombinant protein expressed in
E. coli by induction during late log phase.
Biotechniques 2003,
34, 524–526.
[Google Scholar]
-
313.
Kaur J, Kumar A, Kaur J. Strategies for optimization of heterologous protein expression in
E. coli: Roadblocks and reinforcements.
Int. J. Biol. Macromol. 2018,
106, 803–822.
[Google Scholar]
-
314.
Ki MR, Pack SP. Fusion tags to enhance heterologous protein expression.
Appl. Microbiol. Biotechnol. 2020,
104, 2411–2425.
[Google Scholar]
-
315.
Zhong C, Wei P, Zhang Y-HP. Enhancing functional expression of codon-optimized heterologous enzymes in
Escherichia coli BL21(DE3) by selective introduction of synonymous rare codons.
Biotechnol. Bioeng. 2017,
114, 1054–1064.
[Google Scholar]
-
316.
Martínez-Alonso M, García-Fruitós E, Ferrer-Miralles N, Rinas U, Villaverde A. Side effects of chaperone gene co-expression in recombinant protein production.
Microb. Cell Fact. 2010,
9, 64.
[Google Scholar]
-
317.
Lin Z, Thorsen T, Arnold FH. Functional expression of horseradish peroxidase in E. coli by directed evolution.
Biotechnol. Prog. 2000,
16, 467–471.
[Google Scholar]
-
318.
Ye J, Li YJ, Bai YQ, Zhang T, Jiang W, Shi T, et al. A facile and robust T7-promoter-based high-expression of heterologous proteins in
Bacillus subtilis.
Biores. Bioproc. 2022,
9, 1–12.
[Google Scholar]
-
319.
Sun FF, Zhang XZ, Myung S, Zhang Y-HP. Thermophilic
Thermotoga maritima ribose-5-phosphate isomerase RpiB: Optimized heat treatment purification and basic characterization.
Protein Expr. Purif. 2012,
82, 302–307.
[Google Scholar]
-
320.
Ye X, Honda K, Sakai T, Okano K, Omasa T, Hirota R, et al. Synthetic metabolic engineering—a novel, simple technology for designing a chimeric metabolic pathway.
Microb. Cell Fact. 2012,
11, 120.
[Google Scholar]
-
321.
You C, Zhang Y-HP. Self-assembly of synthetic metabolons through synthetic protein scaffolds: one-step purification, co-immobilization, and substrate channeling.
ACS Syn. Biol. 2013,
2, 102–110.
[Google Scholar]
-
322.
Liu M, Song Y, Zhang Y-HPJ, You C. Carrier-free immobilization of multi-enzyme complex facilitates in vitro synthetic enzymatic biosystem for biomanufacturing.
ChemSusChem 2023,
16, e202202153.
[Google Scholar]
-
323.
Krutsakorn B, Imagawa T, Honda K, Okano K, Ohtake H. Construction of an in vitro bypassed pyruvate decarboxylation pathway using thermostable enzyme modules and its application to N-acetylglutamate production.
Microb. Cell Fact. 2013,
12, 91.
[Google Scholar]
-
324.
Krutsakorn B, Honda K, Ye X, Imagawa T, Bei X, Okano K, et al. In vitro production of N-butanol from glucose.
Metab. Eng. 2013,
20, 84–91.
[Google Scholar]
-
325.
Ninh PH, Honda K, Sakai T, Okano K, Ohtake H. Assembly and Multiple Gene Expression of Thermophilic Enzymes in
Escherichia coli for In Vitro Metabolic Engineering.
Biotechnol. Bioeng. 2015,
112, 189–196.
[Google Scholar]
-
326.
Potapov V, Ong JL. Examining sources of error in PCR by single-molecule sequencing.
PLoS ONE 2017,
12, e0169774.
[Google Scholar]
-
327.
Sankar PS, Citartan M, Siti AA, Skryabin BV, Rozhdestvensky TS, Khor GH, et al. A simple method for in-house Pfu DNA polymerase purification for high-fidelity PCR amplification.
Iran J. Microbiol. 2019,
11, 181–186.
[Google Scholar]
-
328.
Mielenz JR.
Bacillus stearothermophilus contains a plasmid-borne gene for α-amylase.
Proc. Natl. Acad. Sci. USA 1983,
80, 5975–5979.
[Google Scholar]
-
329.
Zhao H, Arnold FH. Directed evolution converts subtilisin E into a functional equivalent of thermitase.
Protein Eng. 1999,
12, 47–53.
[Google Scholar]
-
330.
Cao L, Langen Lv, Sheldon RA. Immobilised enzymes: carrier-bound or carrier-free?
Curr. Opin. Biotechnol. 2003,
14, 387–394.
[Google Scholar]
-
331.
Li H, Sethuraman N, Stadheim TA, Zha D, Prinz B, Ballew N, et al. Optimization of humanized IgGs in glycoengineered
Pichia pastoris.
Nat. Biotechnol. 2006,
24, 210–215.
[Google Scholar]
-
332.
Schwarz FM, Schuchmann K, Müller V. Hydrogenation of CO
2 at ambient pressure catalyzed by a highly active thermostable biocatalyst.
Biotechnol. Biofuels 2018,
11, 237.
[Google Scholar]
-
333.
Liu W, Wang P. Cofactor regeneration for sustainable enzymatic biosynthesis.
Biotechnol. Adv. 2007,
25, 369–384.
[Google Scholar]
-
334.
Kazandjian R, Klibanov A. Regioselective oxidation of phenols catalyzed by polyphenol oxidase in chloroform.
J. Am. Chem. Soc. 1985,
107, 5448–5450.
[Google Scholar]
-
335.
Li Y, Liu S, You C. Permeabilized
Escherichia coli whole cells containing co-expressed two thermophilic enzymes facilitate the synthesis of scyllo-inositol from myo-inositol.
Biotechnol. J. 2020,
15, 1900191.
[Google Scholar]
-
336.
Ryan JD, Fish RH, Clark DS. Engineering cytochrome P450 enzymes for improved activity towards biomimetic 1,4-NADH cofactors.
ChemBioChem 2008,
9, 2579–2582.
[Google Scholar]
-
337.
Campbell E, Wheeldon IR, Banta S. Broadening the cofactor specificity of a thermostable alcohol dehydrogenase using rational protein design introduces novel kinetic transient behavior.
Biotechnol. Bioeng. 2010,
107, 763–774.
[Google Scholar]
-
338.
Zachos I, Döring M, Tafertshofer G, Simon RC, Sieber V. carba nicotinamide adenine dinucleotide phosphate: robust cofactor for redox niocatalysis.
Angew. Chem. Int. Ed. 2021,
60, 14701–14706.
[Google Scholar]
-
339.
Ma K, Adams MW. Hydrogenases I and II from
Pyrococcus furiosus.
Methods Enzymol. 2001,
331, 208–216.
[Google Scholar]
-
340.
Stiefel EI, George GN. Ferredoxins, hydrogenases, and nitrogenases: Metal-sulfide proteins. In Bioinorganic Chemistry; University Science Books: Melville, NY, USA, 1994; pp. 365–453.
-
341.
Schoelmerich MC, Müller V. Energy-converting hydrogenases: the link between H
2 metabolism and energy conservation.
Cell Mol. Life Sci. 2020,
77, 1461–1481.
[Google Scholar]
-
342.
Peters JW, Schut GJ, Boyd ES, Mulder DW, Shepard EM, Broderick JB, et al. [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation.
Biochim. Biophys. Acta 2015,
1853, 1350–1369.
[Google Scholar]
-
343.
DosSantos PC, Dean DR, Hu Y, Ribbe MW. Formation and insertion of the nitrogenase iron-molybdenum cofactor.
Chem. Rev. 2004,
104, 1159–1174.
[Google Scholar]
-
344.
Schuchmann K, Müller V. Direct and Reversible Hydrogenation of CO
2 to Formate by a Bacterial Carbon Dioxide Reductase.
Science 2013,
342, 1382–1385.
[Google Scholar]
-
345.
Dietrich HM, Righetto RD, Kumar A, Wietrzynski W, Trischler R, Schuller SK, et al. Membrane-anchored HDCR nanowires drive hydrogen-powered CO
2 fixation.
Nature 2022,
607, 823–830.
[Google Scholar]
-
346.
Leo F, Schwarz FM, Schuchmann K, Müller V. Capture of carbon dioxide and hydrogen by engineered
Escherichia coli: hydrogen-dependent CO
2 reduction to formate.
Appl. Microbiol. Biotechnol. 2021,
105, 5861–5872.
[Google Scholar]
-
347.
Rieckenberg F, Götz K, Hilterhaus L, Liese A, Zeng A-P. Strategies for reliable and improved large-scale production of
Pyrococcus furiosus with integrated purification of hydrogenase I.
Bioproc. Biosys. Eng. 2014,
37, 2475–2482.
[Google Scholar]
-
348.
Ma K, Schicho RN, Kelly RM, Adams MWW. Hydrogenase of the hyperthermophile
Pyrococcus furiosus is an elemental sulfur reductase or sulfhydrogenase: Evidence for a sulfur-reducing hydrogenase ancestor.
Proc. Natl. Acad. Sci. USA 1993,
90, 5341–5344.
[Google Scholar]
-
349.
Chandrayan SK, McTernan PM, Hopkins RC, Sun JS, Jenney FE, Adams MWW. Engineering hyperthermophilic Archaeon Pyrococcus furiosus to overproduce Its cytoplasmic NiFe -hydrogenase.
J. Biol. Chem. 2012,
287, 3257–3264.
[Google Scholar]
-
350.
Wu C-H, Ponir CA, Haja DK, Adams MWW. Improved production of the NiFe-hydrogenase from
Pyrococcus furiosus by increased expression of maturation genes.
Protein Eng. Des. Sel. 2018,
31, 337–344.
[Google Scholar]
-
351.
Sun J, Hopkins RC, Jenney FE, McTernan PM, Adams MWW. Heterologous expression and maturation of an NADP-dependent [NiFe]-hydrogenase: a key enzyme in biofuel production.
PLoS ONE 2010,
5, e10526.
[Google Scholar]
-
352.
Onoda A, Hayashi T. Artificial hydrogenase: biomimetic approaches controlling active molecular catalysts.
Curr. Opin. Chem. Biol. 2015,
25, 133–140.
[Google Scholar]
-
353.
Pan H-J, Huang G, Wodrich MD, Tirani FF, Ataka K, Shima S, et al. A catalytically active [Mn]-hydrogenase incorporating a non-native metal cofactor.
Nat. Chem. 2019,
11, 669–675.
[Google Scholar]
-
354.
Schaupp S, Arriaza-Gallardo FJ, Pan HJ, Kahnt J, Angelidou G, Paczia N, et al. In Vitro Biosynthesis of the [Fe]-Hydrogenase Cofactor Verifies the Proposed Biosynthetic Precursors.
Angew. Chem. Int. Ed. 2022,
61, e202200994.
[Google Scholar]
-
355.
Zhang Y, Muhammad F, Wei H. Inorganic enzyme mimics.
ChemBioChem 2021,
22, 1496–1498.
[Google Scholar]
-
356.
Ye N, Kou X, Shen J, Huang S, Chen G, Ouyang G. Metal-organic frameworks: A new platform for enzyme immobilization.
ChemBioChem 2020,
21, 2585–2590.
[Google Scholar]
-
357.
Liang W, Wied P, Carraro F, Sumby CJ, Nidetzky B, Tsung CK, et al. Metal-organic framework-based enzyme biocomposites.
Chem. Rev. 2021,
121, 1077–1129.
[Google Scholar]
-
358.
Oliveira FL, de SFA, de Castro AM, Alves de Souza ROM, Esteves PM, Gonçalves RSB. Enzyme Immobilization in Covalent Organic Frameworks: Strategies and Applications in Biocatalysis.
Chempluschem 2020,
85, 2051–2066.
[Google Scholar]
-
359.
Sicard C. In situ enzyme immobilization by covalent organic frameworks.
Angew. Chem. Int. Ed. 2023,
135, e202213405.
[Google Scholar]
-
360.
Sheldon RA, van Pelt S. Enzyme immobilisation in biocatalysis: why, what and how.
Chem. Soc. Rev. 2013,
42, 6223–6235.
[Google Scholar]
-
361.
Ansorge-Schumacher MB, Thum O. Immobilised lipases in the cosmetics industry.
Chem. Soc. Rev. 2013,
42, 6475–6490.
[Google Scholar]
-
362.
Betancor L, Berne C, Luckarift HR, Spain JC. Coimmobilization of a redox enzyme and a cofactor regeneration system.
Chem. Commun. 2006,
34, 3640–3642.
[Google Scholar]
-
363.
El-Zahab B, Jia H, Wang P. Enabling multienzyme biocatalysis using nanoporous materials.
Biotechnol. Bioeng. 2004,
87, 178–183.
[Google Scholar]
-
364.
Mateo C, Chmura A, Rustler S, van Rantwijk F, Stolz A, Sheldon RA. Synthesis of enantiomerically pure (S)-mandelic acid using an oxynitrilase-nitrilase bienzymatic cascade: a nitrilase surprisingly shows nitrile hydratase activity.
Tetrahedron: Asymmetry 2006,
17, 320–323.
[Google Scholar]
-
365.
Myung S, You C, Zhang Y-HP. Recyclable cellulose-containing magnetic nanoparticles: immobilization of cellulose-binding module-tagged proteins and synthetic metabolon featuring substrate channeling.
J. Mater. Chem. B 2013,
1, 4419–4427.
[Google Scholar]
-
366.
Jandt U, You C, Zhang Y-HP, Zeng AP. Compartmentalization and Metabolic Channeling for Multienzymatic Biosynthesis: Practical Strategies and Modeling Approaches.
Adv. Biochem. Eng. Biotechnol. 2013,
137, 1–25.
[Google Scholar]
-
367.
Conrado RJ, Varner JD, DeLisa MP. Engineering the spatial organization of metabolic enzymes: Mimicking nature’s synergy.
Curr. Opin. Biotechnol. 2008,
19, 492–499.
[Google Scholar]
-
368.
Bulow L, Ljungcrantz P, Mosbach K. Preparation of a soluble bifunctional enzyme by gene fusion.
Nat. Biotechnol. 1985,
3, 821–823.
[Google Scholar]
-
369.
Liu X, Hou H, Li Y, Yang S, Lin H, Chen H. Fusion of cellobiose phosphorylase and potato α-glucan phosphorylase facilitates substrate channeling for enzymatic conversion of cellobiose to starch.
Prep. Biochem. Biotechnol. 2022,
52, 611–617.
[Google Scholar]
-
370.
Riedel K, Bronnenmeier K. Intramolecular synergism in an engineered exo-endo-1,4-b-glucanase fusion protein.
Mol. Microbiol. 1998,
28, 767–775.
[Google Scholar]
-
371.
Orita I, Sakamoto N, Kato N, Yurimoto H, Sakai Y. Bifunctional enzyme fusion of 3-hexulose-6-phosphate synthase and 6-phospho-3-hexuloisomerase.
Appl. Microbiol. Biotechnol. 2007,
76, 439–445.
[Google Scholar]
-
372.
Agapakis C, Ducat D, Boyle P, Wintermute E, Way J, Silver P. Insulation of a synthetic hydrogen metabolism circuit in bacteria.
J. Biol. Eng. 2010,
4, 3.
[Google Scholar]
-
373.
Meynial-Salles I, Forchhammer N, Croux C, Girbal L, Soucaille P. Evolution of a Saccharomyces cerevisiae metabolic pathway in
Escherichia coli.
Metab. Eng. 2007,
9, 152–159.
[Google Scholar]
-
374.
Chang H-C, Kaiser CM, Hartl FU, Barral JM. De novo Folding of GFP Fusion Proteins: High Efficiency in Eukaryotes but Not in Bacteria.
J. Mol. Biol. 2005,
353, 397–409.
[Google Scholar]
-
375.
Aldaye FA, Palmer AL, Sleiman HF. Assembling Materials with DNA as the Guide.
Science 2008,
321, 1795–1799.
[Google Scholar]
-
376.
Wei B, Dai M, Yin P. Complex shapes self-assembled from single-stranded DNA tiles.
Nature 2012,
485, 623–626.
[Google Scholar]
-
377.
Bayer EA, Morag E, Lamed R. The cellulosome--a treasure-trove for biotechnology.
Trends Biotechnol. 1994,
12, 379–386.
[Google Scholar]
-
378.
Reddington SC, Howarth M. Secrets of a covalent interaction for biomaterials and biotechnology: SpyTag and SpyCatcher.
Curr. Opin. Chem. Biol. 2015,
29, 94–99.
[Google Scholar]
-
379.
Kang W, Ma T, Liu M, Qu J, Liu Z, Zhang H, et al. Modular enzyme assembly for enhanced cascade biocatalysis and metabolic flux.
Nat. Commun. 2019,
10, 4248.
[Google Scholar]
-
380.
Gao X, Yang S, Zhao C, Ren Y, Wei D. Artificial multienzyme supramolecular device: Highly ordered self-assembly of oligomeric enzymes in vitro and in vivo.
Angew. Chem. Int. Ed. 2014,
126, 14251–14254.
[Google Scholar]
-
381.
Zhu G, Song P, Wu J, Luo M, Chen Z, Chen T. Application of nucleic acid frameworks in the construction of nanostructures and cascade biocatalysts: Recent progress and perspective.
Front. Bioeng. Biotechnol. 2021,
9, 792489.
[Google Scholar]
-
382.
Zhang L, Ahvazi B, Szittner R, Vrielink A, Meighen E. Change of nucleotide specificity and enhancement of catalytic efficiency in single point mutants of
Vibrio harveyi aldehyde dehydrogenase.
Biochemistry 1999,
38, 11440–11447.
[Google Scholar]
-
383.
Yaoi T, Miyazaki K, Oshima T, Komukai Y, Go M. Conversion of the coenzyme specificity of isocitrate dehydrogenase by module replacement.
J. Biochem. 1996,
119, 1014–1018.
[Google Scholar]
-
384.
Bastian S, Liu X, Meyerowitz JT, Snow CD, Chen MMY, Arnold FH. Engineered ketol-acid reductoisomerase and alcohol dehydrogenase enable anaerobic 2-methylpropan-1-ol production at theoretical yield in
Escherichia coli.
Metab. Eng. 2011,
13, 345–352.
[Google Scholar]
-
385.
Rosell A, Valencia E, Ochoa WF, Fita I, Pares X, Farres J. Complete Reversal of Coenzyme Specificity by Concerted Mutation of Three Consecutive Residues in Alcohol Dehydrogenase.
J. Biol. Chem. 2003,
278, 40573–40580.
[Google Scholar]
-
386.
Döhr O, Paine MJI, Friedberg T, Roberts GCK, Wolf CR. Engineering of a functional human NADH-dependent cytochrome P450 system.
Proc. Nat. Acad. Sci. USA 2001,
98, 81–86.
[Google Scholar]
-
387.
Banta S, Swanson BA, Wu S, Jarnagin A, Anderson S. Alteration of the specificity of the cofactor-binding pocket of Corynebacterium 2,5-diketo- D-gluconic acid reductase A.
Protein Eng. Des. Sel. 2002,
15, 131–140.
[Google Scholar]
-
388.
Banta S, Swanson BA, Wu S, Jarnagin A, Anderson S. Optimizing an artificial metabolic pathway: Engineering the cofactor specificity of Corynebacterium 2,5-Diketo- D-gluconic acid reductase for use in vitamin C biosynthesis.
Biochemistry 2002,
41, 6226–6236.
[Google Scholar]
-
389.
Bocanegra JA, Scrutton NS, Perham RN. Creation of an NADP-dependent pyruvate dehydrogenase multienzyme complex by protein engineering.
Biochemistry 1993,
32, 2737–2740.
[Google Scholar]
-
390.
Mittl PRE, Berry A, Scrutton NS, Perham RN, Schulz GE. Structural differences between wild-type NADP-dependent glutathione reductase from
Escherichia coli and a redesigned NAD-dependent mutant.
J. Mol. Biol. 1993,
231, 191–195.
[Google Scholar]
-
391.
Steen IH, Lien T, Madsen MS, Birkeland N-K. Identification of cofactor discrimination sites in NAD-isocitrate dehydrogenase from
Pyrococcus furiosus.
Arch. Microbiol. 2002,
178, 297–300.
[Google Scholar]
-
392.
Watanabe S, Kodaki T, Makino K. Complete Reversal of Coenzyme Specificity of Xylitol Dehydrogenase and Increase of Thermostability by the Introduction of Structural Zinc.
J. Biol.Chem. 2005,
280, 10340–10349.
[Google Scholar]
-
393.
Glykys DJ, Banta S. Metabolic control analysis of an enzymatic biofuel cell.
Biotechnol. Bioeng. 2009,
102, 1624–1635.
[Google Scholar]
-
394.
Woodyer RD, van der Donk WA, Zhao H. Relaxing the nicotinamide cofactor specificity of phosphite dehydrogenase by rational design.
Biochemistry 2003,
42, 11604–11614.
[Google Scholar]
-
395.
Wiegert T, Sahm H, Sprenger GA. The Substitution of a Single Amino Acid Residue (Ser-116 → Asp) Alters NADP-containing Glucose-Fructose Oxidoreductase of
Zymomonas mobilis into a Glucose Dehydrogenase with Dual Coenzyme Specificity.
J. Biol. Chem. 1997,
272, 13126–13133.
[Google Scholar]
-
396.
Katzberg M, Skorupa-Parachin N, Gorwa-Grauslund M-F, Bertau M. Engineering Cofactor Preference of Ketone Reducing Biocatalysts: A Mutagenesis Study on a γ-Diketone Reductase from the Yeast
Saccharomyces cerevisiae Serving as an Example.
Int. J. Mol. Sci. 2010,
11, 1735–1758.
[Google Scholar]
-
397.
Sanli G, Banta S, Anderson S, Blaber M. Structural alteration of cofactor specificity in
Corynebacterium 2,5-diketo- D-gluconic acid reductase.
Protein Eng. 2004,
13, 504–512.
[Google Scholar]
-
398.
Paul CE, Arends IWCE, Hollmann F. Is simpler better? Synthetic nicotinamide cofactor analogues for redox chemistry.
ACS Catal. 2014,
4, 788–797.
[Google Scholar]
-
399.
Knaus T, Paul CE, Levy CW, de Vries S, Mutti FG, Hollmann F, et al. Better than Nature: Nicotinamide Biomimetics That Outperform Natural Coenzymes.
J. Am. Chem. Soc. 2016,
138, 1033–1039.
[Google Scholar]
-
400.
Zachos I, Nowak C, Sieber V. Biomimetic cofactors and methods for their recycling.
Curr. Opin. Chem. Biol. 2019,
49, 59–66.
[Google Scholar]
-
401.
Nowak C, Pick A, Csepei LI, Sieber V. Characterization of Biomimetic Cofactors According to Stability, Redox Potentials, and Enzymatic Conversion by NADH Oxidase from
Lactobacillus pentosus.
ChemBioChem 2017,
18, 1944–1949.
[Google Scholar]
-
402.
Ji D, Wang L, Hou S, Liu W, Wang J, Wang Q, et al. Creation of bioorthogonal redox systems depending on nicotinamide flucytosine dinucleotide.
J. Am. Chem. Soc. 2011,
133, 20857–20862.
[Google Scholar]
-
403.
Wang X, Feng Y, Guo X, Wang Q, Ning S, Li Q, et al. Creating enzymes and self-sufficient cells for biosynthesis of the non-natural cofactor nicotinamide cytosine dinucleotide.
Nat. Commun. 2021,
12, 2116.
[Google Scholar]
-
404.
Guarneri A, van Berkel WJ, Paul CE. Alternative coenzymes for biocatalysis.
Curr. Opin. Biotechnol. 2019,
60, 63–71.
[Google Scholar]
-
405.
Toogood HS, Scrutton NS. Discovery, characterisation, engineering and applications of ene reductases for industrial biocatalysis.
ACS Catal. 2019,
8, 3532–3549.
[Google Scholar]
-
406.
Paul CE, Gargiulo S, Opperman DJ, Lavandera I, Gotor-Fernández V, Gotor V, et al. Mimicking nature: synthetic nicotinamide cofactors for C=C bioreduction using enoate reductases.
Org. Lett. 2013,
15, 180–183.
[Google Scholar]
-
407.
Nowak C, Beer B, Pick A, Roth T, Lommes P, Sieber V. A water-forming NADH oxidase from Lactobacillus pentosus suitable for the regeneration of synthetic biomimetic cofactors.
Front. Microbiol. 2015,
6, 957.
[Google Scholar]
-
408.
Campbell E, Meredith M, Minteer SD, Banta S. Enzymatic biofuel cells utilizing a biomimetic cofactor.
Chem. Commun. 2012,
48, 1898–1900.
[Google Scholar]
-
409.
Lo HC, Fish RH. Biomimetic NAD
+ models for tandem cofactor regeneration, horse liver alcohol dehydrogenase recognition of 1,4-NADH derivatives, and chiral synthesis.
Angew. Chem. Int. Ed. 2002,
41, 478–481.
[Google Scholar]
-
410.
Paul CE, Hollmann F. A survey of synthetic nicotinamide cofactors in enzymatic processes.
Appl. Microbiol. Biotechnol. 2016,
100, 4773–4778.
[Google Scholar]
-
411.
Fisher HF, McGregor LL. The ability of reduced nicotinamide mononucleotide to function as a hydrogen donor in the glutamic dehydrogenase reaction.
Biochem. Biophys. Res. Commun. 1969,
34, 627–632.
[Google Scholar]
-
412.
Sicsic S, Durand P, Langrene S, Goffic FL. Activity of NMN
+, nicotinamide ribose and analogs in alcohol oxidation promoted by horse-liver alcohol dehydrogenase.
Eur. J. Biochem. 1986,
155, 403–407.
[Google Scholar]
-
413.
Black WB, Zhang L, Mak WS, Maxel S, Cui Y, King E, et al. Engineering a nicotinamide mononucleotide redox cofactor system for biocatalysis.
Nat. Chem. Biol. 2020,
16, 87–94.
[Google Scholar]
-
414.
Zhang L, King E, Black WB, Heckmann CM, Wolder A, Cui Y, et al. Directed evolution of phosphite dehydrogenase to cycle noncanonical redox cofactors via universal growth selection platform.
Nat. Commun. 2022,
13, 5021.
[Google Scholar]
-
415.
Li F, Wei X, Zhang L, Liu C, You C, Zhu Z. Installing a green engine to drive an enzyme cascade: A light-powered In vitro biosystem for poly(3-hydroxybutyrate) synthesis.
Angew. Chem. Int. Ed. 2022,
61, e202111054.
[Google Scholar]
-
416.
Miller TE, Beneyton T, Schwander T, Diehl C, Girault M, McLean R, et al. Light-powered CO
2 fixation in a chloroplast mimic with natural and synthetic parts.
Science 2020,
368, 649–654.
[Google Scholar]
-
417.
Zhong C, Wei P, Zhang Y-HP. A kinetic model of one-pot rapid biotransformation of cellobiose from sucrose catalyzed by three thermophilic enzymes.
Chem. Eng. Sci. 2017,
161, 159–166.
[Google Scholar]
-
418.
Ardao I, Zeng A-P. In silico evaluation of a complex multi-enzymatic system using one-pot and modular approaches: Application to the high-yield production of hydrogen from a synthetic metabolic pathway.
Chem. Eng. Sci. 2013,
87, 183–193.
[Google Scholar]
-
419.
Yamamoto T, Hoshikawa K, Ezura K, Okazawa R, Fujita S, Takaoka M, et al. Improvement of the transient expression system for production of recombinant proteins in plants.
Sci. Rep. 2018,
8, 4755.
[Google Scholar]
-
420.
Krenek P, Samajova O, Luptovciak I, Doskocilova A, Komis G, Samaj J. Transient plant transformation mediated by Agrobacterium tumefaciens: Principles, methods and applications.
Biotechnol. Adv. 2015,
33, 1024–1042.
[Google Scholar]
-
421.
Dugdale B, Mortimer CL, Kato M, James TA, Harding RM, Dale JL. Design and construction of an in-plant activation cassette for transgene expression and recombinant protein production in plants.
Nat. Protoc. 2014,
9, 1010–1027.
[Google Scholar]
-
422.
Wang X, Saba T, Yiu HHP, Howe RF, Anderson JA, Shi J. Cofactor NAD(P)H Regeneration Inspired by Heterogeneous Pathways.
Chem 2017,
2, 621–654.
[Google Scholar]
-
423.
Ali I, Khan T, Omanovic S. Direct electrochemical regeneration of the cofactor NADH on bare Ti, Ni, Co and Cd electrodes: The influence of electrode potential and electrode material.
J. Mol. Catal. A Chem. 2014,
387, 86–91.
[Google Scholar]
-
424.
Morello G, Megarity CF, Armstrong FA. The power of electrified nanoconfinement for energising, controlling and observing long enzyme cascades.
Nat. Commun. 2021,
12, 340.
[Google Scholar]
-
425.
Castañeda-Losada L, Adam D, Paczia N, Buesen D, Steffler F, Sieber V, et al. Bioelectrocatalytic Cofactor Regeneration Coupled to CO
2 Fixation in a Redox-Active Hydrogel for Stereoselective C−C Bond Formation.
Angew. Chem. Int. Ed. 2021,
60, 21056–21061.
[Google Scholar]
-
426.
Wu R, Ma C, Zhu Z. Enzymatic electrosynthesis as an emerging electrochemical synthesis platform.
Curr. Opin. Electrochem. 2020,
19, 1–7.
[Google Scholar]
-
427.
Simonis W, Urbach W. Photophosphorylation in vivo.
Annu. Rev. Plant Physiol. 1973,
24, 89–114.
[Google Scholar]
-
428.
Gutierrez-Sanz O, Marques M, Pereira IAC, De Lacey AL, Lubitz W, Rudiger O. Orientation and Function of a Membrane-Bound Enzyme Monitored by Electrochemical Surface-Enhanced Infrared Absorption Spectroscopy.
J. Phys. Chem. Lett. 2013,
4, 2794–2798.
[Google Scholar]
-
429.
Gutierrez-Sanz O, Tapia C, Marques MC, Zacarias S, Velez M, Pereira IAC, et al. Induction of a Proton Gradient across a Gold-Supported Biomimetic Membrane by Electroenzymatic H-2 Oxidation.
Angew. Chem. Int. Ed. 2015,
54, 2684–2687.
[Google Scholar]
-
430.
Gutierrez-Sanz O, Natale P, Marquez I, Marques MC, Zacarias S, Pita M, et al. H-2-Fueled ATP Synthesis on an Electrode: Mimicking Cellular Respiration.
Angew. Chem.-Int. Ed. 2016,
55, 6216–6220.
[Google Scholar]
-
431.
Jia Y, Li J. Reconstitution of FoF1-ATPase-based biomimetic systems.
Nat. Rev. Chem. 2019,
3, 361–374.
[Google Scholar]
-
432.
Luo S, Adam D, Giaveri S, Barthel S, Cestellos-Blanco S, Hege D, et al. ATP production from electricity with a new-to-nature electrobiological module.
Joule 2023,
7, 1745–1758.
[Google Scholar]
-
433.
Wu Z-Q, Li Z-Q, Li J-Y, Gu J, Xia X-H. Contribution of convection and diffusion to the cascade reaction kinetics of β-galactosidase/glucose oxidase confined in a microchannel.
Phys. Chem. Chem. Phys. 2016,
18, 14460–14465.
[Google Scholar]
-
434.
Marques MPC, Szita N. Bioprocess microfluidics: applying microfluidic devices for bioprocessing.
Curr. Opin. Chem. Eng. 2017,
18, 61–68.
[Google Scholar]
-
435.
Miyazaki M, Maeda H. Microchannel enzyme reactors and their applications for processing.
Trends Biotechnol. 2006,
24, 463–470.
[Google Scholar]
-
436.
Giannakopoulou A, Gkantzou E, Polydera A, Stamatis H. Multienzymatic nanoassemblies: Recent progress and applications.
Trends Biotechnol. 2020,
38, 202–216.
[Google Scholar]
 Zhiguang Zhu
1,2,3
Chun You
2,3
Lingling Zhang
1,2,3
Kuanqing Liu
2,3
Zhiguang Zhu
1,2,3
Chun You
2,3
Lingling Zhang
1,2,3
Kuanqing Liu
2,3