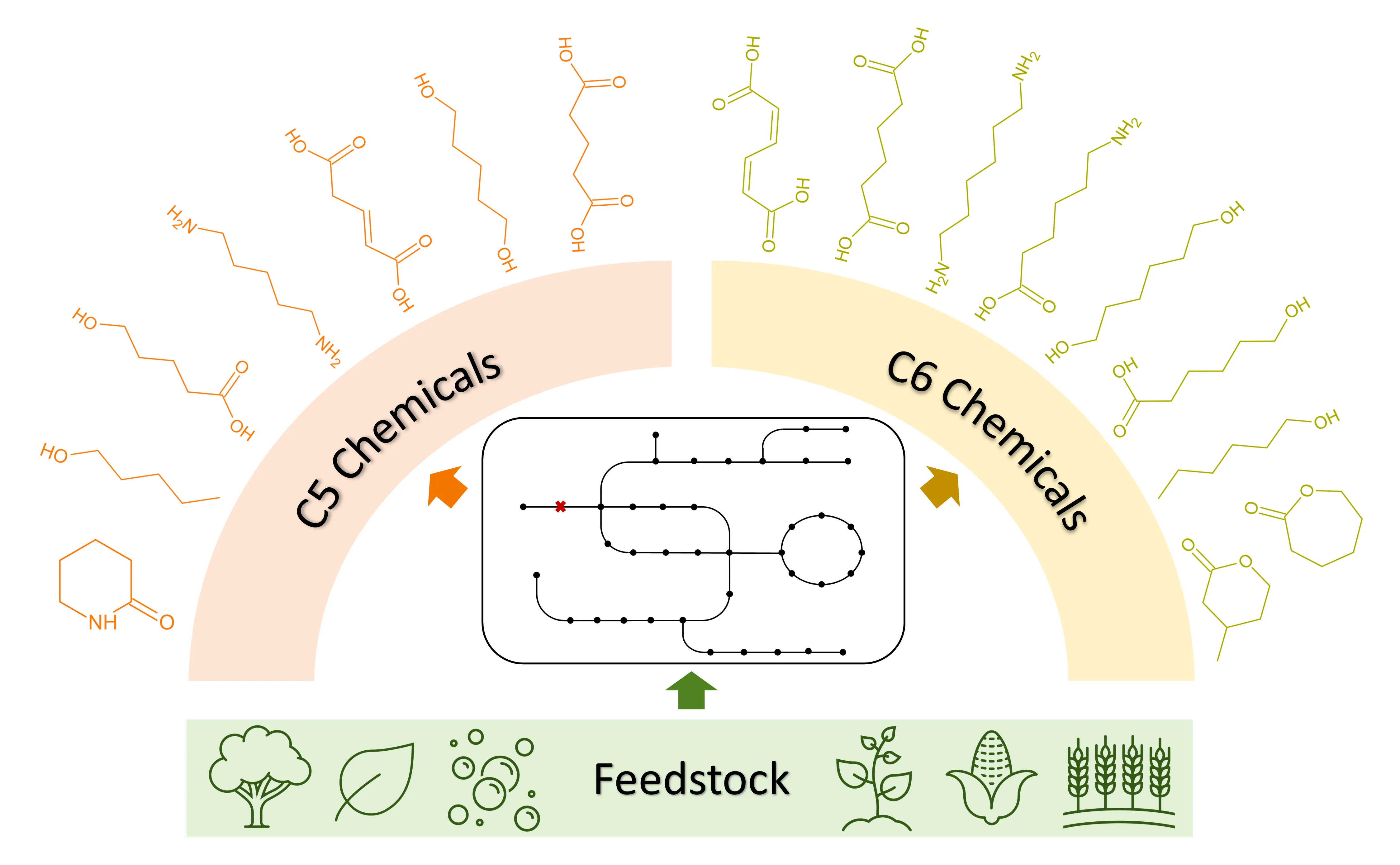
1. Introduction
Microorganisms are natural reservoirs for a wide variety of metabolites, many of which are important compounds such as amino acids, vitamins, alcohols, organic acids, and antibiotics, with broad applications in our daily lives. Metabolic engineering enables the design and rational modification of natural microorganisms, empowering them with the capability to efficiently biosynthesize these naturally occurring molecules. Furthermore, the development of synthetic biology makes it possible to reconstitute and fine-tune non-native biosynthesis pathways in a heterologous microbial chassis or even construct artificial pathways for the production of non-natural molecules.
While most of the microbial-derived small-molecule products that have reached the commercial stage are natural metabolites synthesized through their native metabolic pathways, such as ethanol, lactic acid, succinic acid, citric acid, amino acids and antibiotics, some pioneering companies have attempted and succeeded in the industrial scale microbial production of non-native and non-natural molecules such as 1,3-propanediol (1,3-PDO), 1,4-butanediol (BDO) and isobutanol. 1,3-PDO is a platform chemical with primary applications in the synthesis of polyesters, polyethers, and polyurethanes [
1,
2]. It is naturally produced only from glycerol by certain microbial species including
Klebsiella,
Citrobacter,
Enterobacter, and
Clostridium, while
Escherichia coli lacks the 1,3-PDO biosynthesis pathway [
3]. Moreover, no naturally occurring microorganisms have been reported to produce 1,3-PDO from glucose. In 2000, scientists from DuPont and Genencor introduced an artificial pathway into
E. coli to convert glucose to glycerol and then incorporated a foreign 1,3-PDO biosynthesis pathway [
1]. Following extensive optimizations in the production strain and process, economically viable bioprocesses were successfully developed in 2006, using starch sugar as the raw material [
4]. This milestone marked a starting point for microbial production of non-natural commodity chemicals through engineered artificial pathways. Following this success, Genomatica and Gevo commercialized the biomanufacturing of 1,4-BDO and isobutanol, respectively, through non-natural biosynthesis pathways [
5,
6]. Other well-known examples with the potential for commercialization include 1,2-propanediol, 1-butanol, 2,3-butanediol, 1,2,4-butanetriol, cadaverine, etc. [
7,
8,
9,
10,
11].
It's evident that the majority of these non-natural chemicals were primarily confined to three- and four-carbon (C3 and C4) molecules. This limitation arises from the fact that the abundant C2-C5 metabolites such as acetyl-CoA, pyruvate, PEP, oxaloacetate, succinyl-CoA, and α-ketoglutarate within the host cells can readily serve as biosynthetic precursors to the C3 and C4 molecules. In contrast, biosynthesis of certain C5 and C6 chemicals encounters challenges due to the lack of suitable native precursors.
In recent years, significant efforts have been made to develop microbial synthesis approaches for their production, because C5 and C6 chemicals have broad applications, especially in the production of polymers, such as nylon 5, nylon 5,6, nylon 6, nylon 6,6 and poly-caprolactone (PCL). The global polymer market was valued at $17.1 billion in 2022 and is expected to grow 3.5% annually from 2023–2030 [
12]. Currently, almost all the C5 and C6 chemicals are manufactured through chemical synthesis processes using petroleum-derived feedstocks such as benzene, cyclohexanol, and cyclohexanone. The chemical processes involve multiple toxic reactants, intermediates, and catalysts, and result in significant emissions of greenhouse gases such as nitrogen oxides. For instance, the chemical synthesis of caprolactam, a precursor for nylon 6, relies on the hydrogenation of benzene to cyclohexane, and then subsequent oxidation. This manufacturing route results in the emission of nitrous oxide, carbon dioxide, and sulfur dioxide, in addition to various non-methane volatile organic compounds. Nitrous oxide emissions alone are 10–15 million metric tons of CO
2-equivalent [
13]. Particularly, the limited availability of C5 feedstock from petroleum makes chemical synthesis complicated and expensive.
To alleviate the dependence on fossil fuels and pave the way to a greener future, scientists and engineers have made substantial efforts to explore the bio-based production of these chemicals, utilizing renewable starting materials such as glucose, glycerol, xylose, lignin, and syngas. Glucose, sourced from plant-based origins such as potato, wheat, and maize starch [
14,
15], and glycerol, a byproduct of biodiesel synthesis, offer readily available and eco-friendly carbon sources [
16]. Using these carbon sources, microbial cell factories have been engineered to produce a variety of C5 and C6 chemicals. Due to the frequent involvement of decarboxylation reactions resulting in the loss of one carbon, biosynthesis of C5 and C6 molecules typically requires C6 and C7 precursors. For example, lysine, a C6 amino acid, and its decarboxylation products are often employed as the precursor to many non-natural C5 chemicals. Given the lack of native C7 precursors in microbial cells, biosynthesis of C6 molecules is even more challenging. To address this limitation, efforts have been made by engineering novel, non-natural biosynthesis pathways involving chain extension (+1) reactions, condensation reactions, and ring-fission reactions of aromatic compound catechol to create the six-carbon backbone. Recently, Genomatica and Aquafil announced the pre-commercial production for nylon-6 derived from plant sugars [
17]. Additionally, Toray is currently upscaling its production of plant-based adipic acid (AA) in preparation for commercialization [
18]. As the development of bio-based chemicals matures, these innovations are anticipated to account for an increased percentage of chemicals produced worldwide.
In this review, we focus on the progress in the microbial production of the non-natural C5 chemicals () including 1-pentanol, 1,5-pentanediol (1,5-PDO), cadaverine, δ -valerolactam, glutaric acid (GA), glutaconic acid, and 5-hydroxyvaleric acid (5-HV), as well as the non-natural C6 chemicals () including
cis,
cis-muconic acid (MA), AA, 1,6-hexamethylenediamine (HMDA), 6-aminocaproic acid (ACA), β-methyl-δ-valerolactone (MVL), 1-hexanol, ε-caprolactone (CL), 6-hydroxyhexanoic acid (HHA), and 1,6-hexanediol (1,6-HDO) with an emphasis on strategies employing renewable carbon sources.
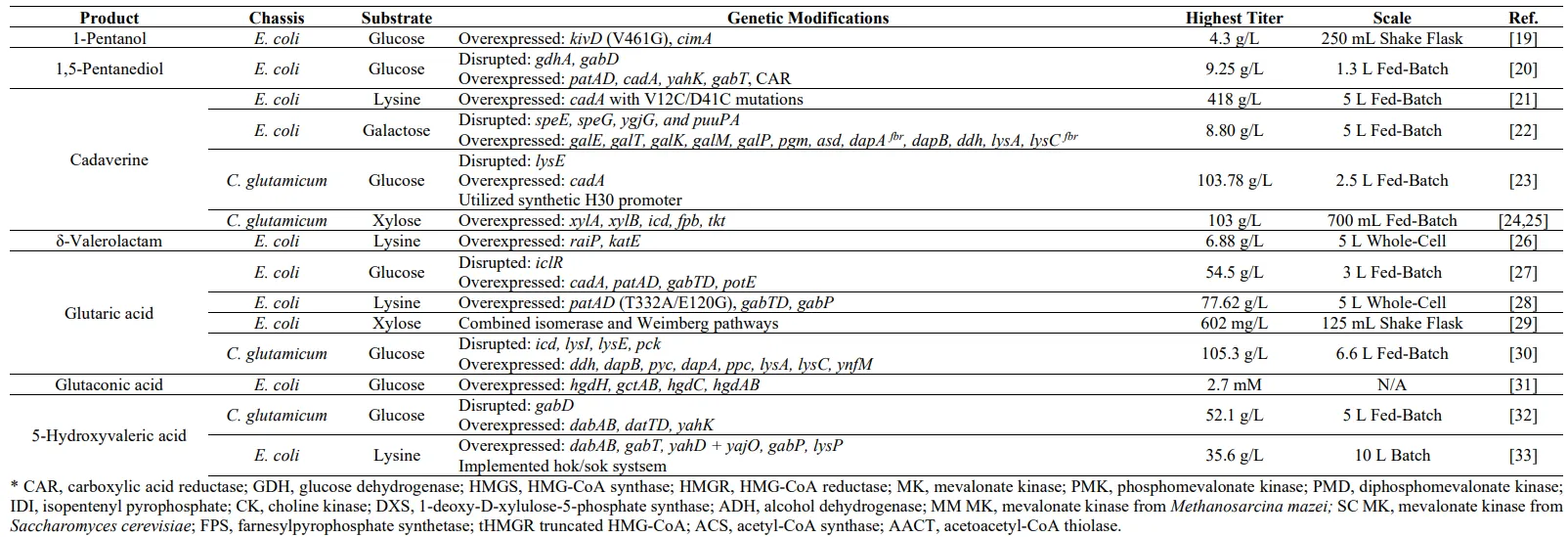
. Best-performing metabolic engineering strategies for the microbial production C5 chemicals with diverse chassis and substrates.
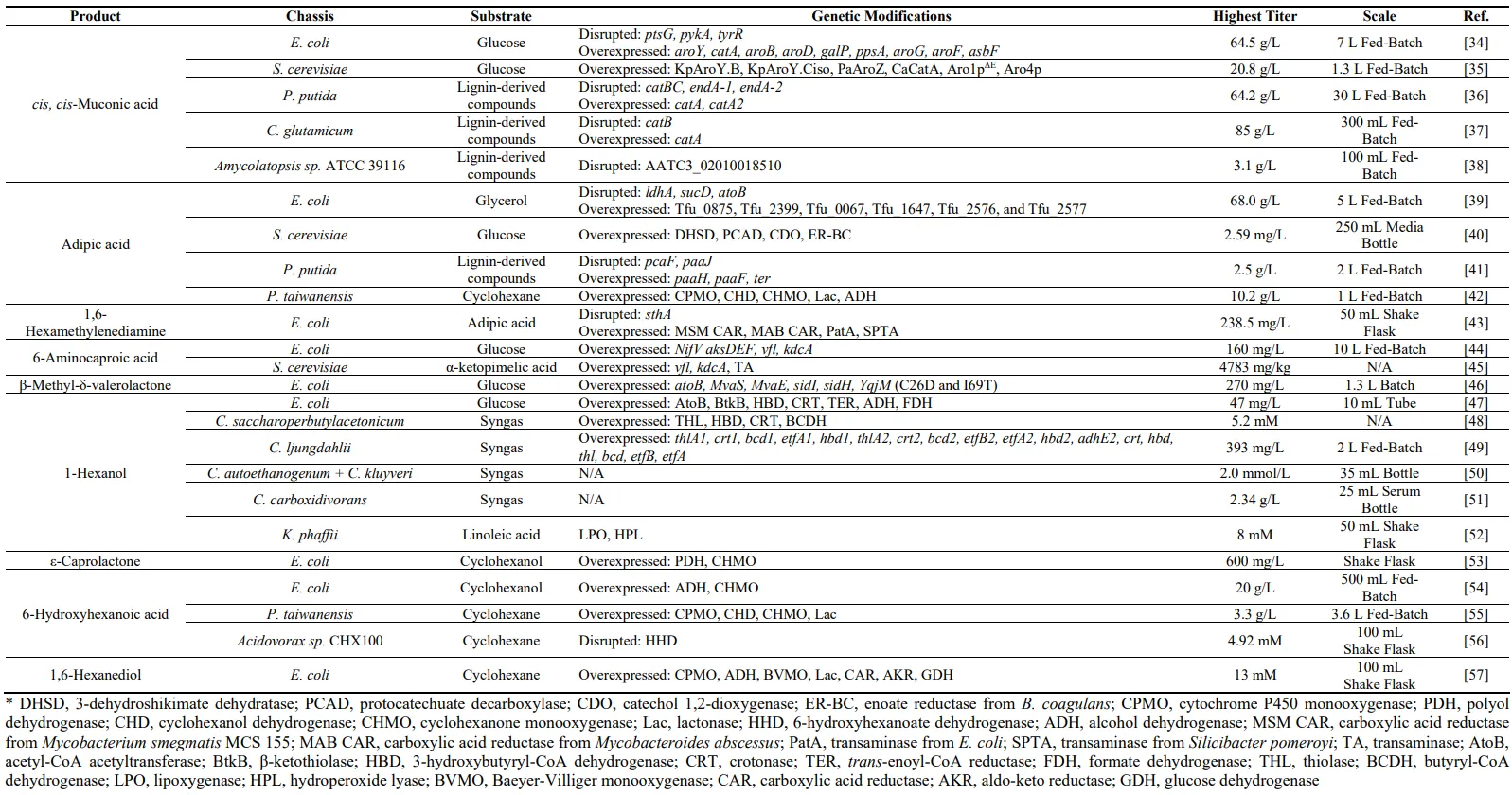
. Best performing metabolic engineering strategies for the microbial production C6 chemicals with diverse chassis and substrates.
2. C5 Chemicals
2.1. 1-Pentanol
1-Pentanol is a straight-chain C5 alcohol commonly used as a coating solvent or in fragrances, while it also has potential as a fuel alternative [
19]. It is considered a semi-natural alcohol, as only trace amounts are generated in yeast fermentations. Recent studies have reported the microbial production of 1-pentanol in
E. coli through extending the 2-ketobutyrate (2-KB) pathway [
58]. This pathway utilizes the chain elongation cycle mediated by the broad-substrate LeuABCD in the leucine biosynthesis and can be employed to produce C5-C7 alcohols that lack a natural biosynthetic pathway, such as 3-methyl-1-butanol and 3-methyl-1-pentanol [
58]. However, the wild-type LeuABCD was unable to act on 2-KB for 1-pentanol production. To address this limitation, Zhang et al. introduced a leucine-feedback insensitive mutation into LeuA (G462D). Along with the action of 2-ketoisovalerate decarboxylase (KIVD) from
Lactococcus lactis and alcohol dehydrogenase (ADH6) from
Saccharomyces cerevisiae, they achieved the biosynthesis of 494 mg/L of 1-pentanol. Subsequent protein engineering of KIVD led to the identification of a more efficient mutant V461A/M538A, resulting in an increase in 1-pentanol to 750 mg/L [
59].
Building on this foundation, Chen et al. attempted to further enhance the 1-pentanol production by specifically focusing on the effects of the residue V461 on KIVD activity. A saturated mutagenesis study of this residue revealed that the V461G mutant exhibited the highest activity and selectivity towards the 2-ketocaproate, the precursor of 1-pentanol. This improvement may be attributed to the extra space created within the binding pocket, allowing for a looser docking of the substrate. Notably, this particular mutant also showed a reduced catalytic efficiency for 2-KB and 2-ketovalerate, which decreased the production of the shorter-chain alcohols, 1-propanol, and 1-butanol, while increasing the production of 1-pentanol. Additional improvement was achieved by synergistically overexpressing the
E. coli native threonine pathway alongside the
CimA gene from
Methanococcus jannaschii, whose catalytic activity was enhanced through directed evolution to bolster the citramalate pathway, ultimately resulting in an increased supply of 2-KB. Moreover, the elevated supply of acetyl-CoA, the iterative addition unit, by acetate feeding further directed the 2-ketoacid flux into the elongation cycle, thus enhancing the 1-pentanol productivity. As a result of these combined efforts, a final titer of 4.3 g/L was achieved [
19].
2.2. 1,5-Pentanediol (1,5-PDO)
1,5-PDO is a commodity chemical widely recognized for its industrial application as a plasticizer and its role in the production of polyester and other diol compounds [
60]. Despite its high industrial value, the production and market size of 1,5-PDO has been constrained due to its complex manufacturing process and lack of readily available C5 feedstock in petroleum. 1,5-PDO is a non-natural chemical that lacks a natural biosynthesis pathway which is a great challenge for its microbial production [
60]. However, metabolic engineering and synthetic biology allow the construction of non-natural pathways in microorganisms for their production ().
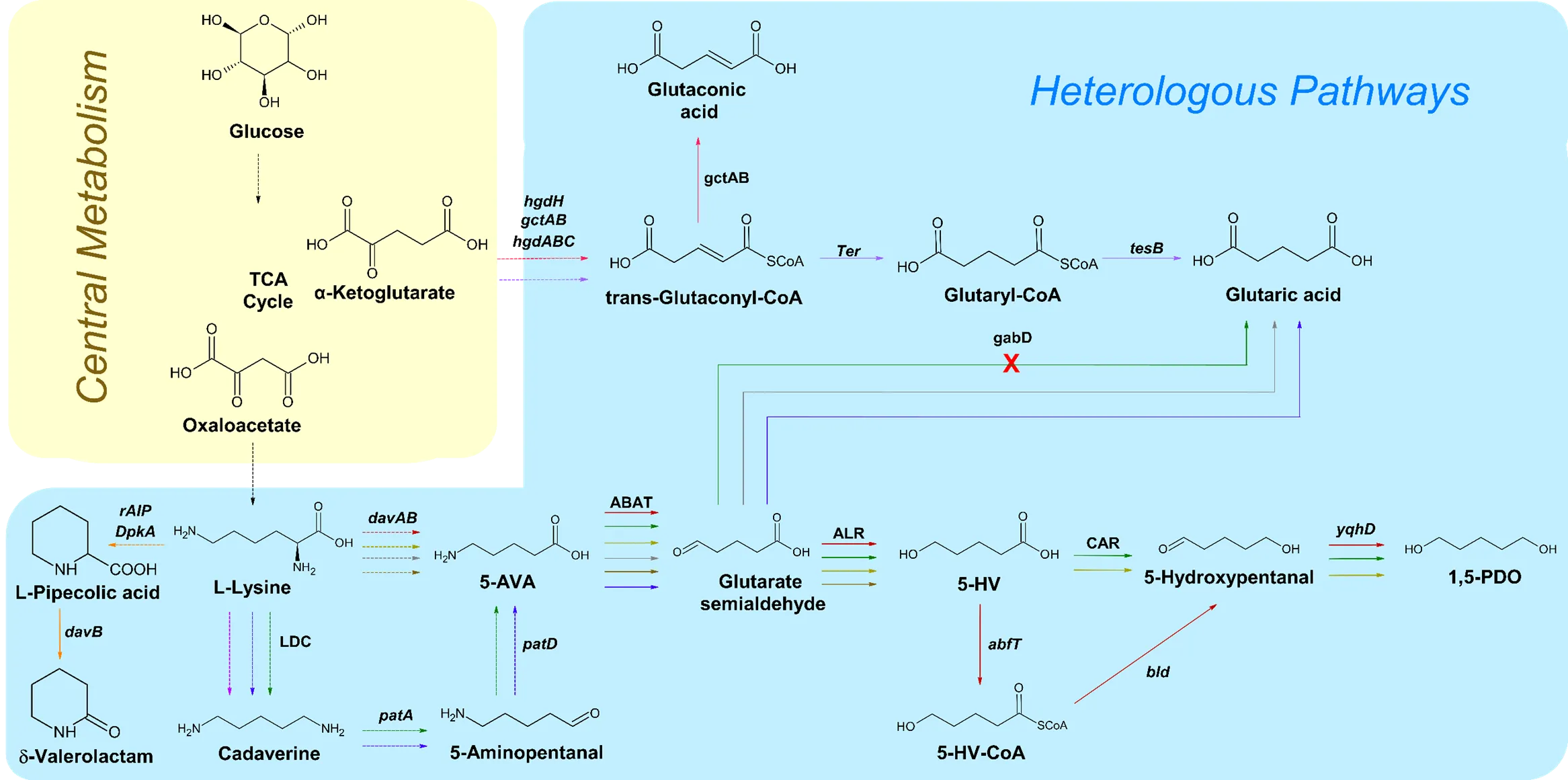
. An overall schematic of the biosynthetic pathways that extend off of the TCA cycle for the production of C5 chemicals. Solid arrows signify direct steps. Dashed arrows represent multiple steps. The red pathway corresponds to the reference [
60]. The green pathway coincides with the reference [
20]. Light olive pathway correlates to reference [
61]. The fuchsia pathway is shown in reference [
21,
22,
23,
24,
25,
62–75]. The orange pathway can be found in reference [
26,
76]. The grey pathway is highlighted in the reference [
30,
77,
78,
79,
80,
81,
82,
83,
84,
85,
86]. Blue pathway belongs to reference [
27,
28,
87]. The purple pathway corresponds to the reference [
88]. The pink pathway correlates with the reference [
31]. Brown pathway signifies the reaction in reference [
32,
33]. Black arrows represent steps utilized by multiple pathways. LDC, lysine decarboxylase; ABAT, 4-aminobutyrate transaminase; ALR, aldehyde reductase; CAR, carboxylic acid reductase; 5-AVA, 5-aminovaleric acid; 5-HV, 5-hydroxyvaleric acid; 5-HV-CoA, 5-hydroxyvaleric acid CoA; 1,5-PDO, 1,5-pentanediol.
As of now, the microbial production of 1,5-PDO via metabolic engineering has only been studied in
E. coli as the host microorganism. For example, Cen et al. created a lysine-overproducing
E. coli strain by expressing
lysC,
dapA, and
ddh, which are all genes that encode for rate-limiting enzymes in the biosynthesis pathway. Building upon the lysine-producing chassis, the
davB and
davA genes from
Pseudomonas putida were introduced to allow for an efficient conversion from lysine to 5-AVA as well as an alcohol dehydrogenase encoded by
yqhD. Furthermore, the performance of 4-aminobutyrate transaminases (ABAT) encoded by gabT genes from different organisms including
E. coli,
P. putida, and
Pseudomonas stutzeri were evaluated, among which GabT from
E. coli produced the highest amount of 5-hydroxyvalerate (5-HV). During the conversion from 5-AVA to 5-HV, a high accumulation of GA was observed. Therefore, Cen et al. included the use of two types of transporters, which are encoded by
cgl0841 from
C. glutamicum and
gabP from
E. coli into the plasmid construction to reduce the byproduct accumulation. As this article used the 5-HV-CoA-based pathway, Cen et al. screened different CoA-transferases from different sources, with the
abfT gene from
Clostridium aminobutyricum producing the highest titer. Ultimately, the final strain showed a considerable amount of 5-HV and lysine accumulation and a low yield of the final product, 1,5-PDO, of less than 1 g/L. This result suggests there is a huge metabolic imbalance present between all three modules, which could be due to the lack of NADPH available for the entire pathway [
60].
The same group of researchers continued to optimize this reaction and found a new pathway using cadaverine, which was found to be more energy efficient. As a result, in addition to the
gabT gene and alcohol dehydrogenase used to synthesize 5-HV, this study required the involvement of a putrescine transaminase (
patA), 4-aminobutyraldehyde dehydrogenase (
patD), and an endogenous lysine decarboxylase. In
E. coli, there are two lysine decarboxylases, encoded by
cadA and
ldcC, while the former was chosen for its greater stability.
CadA and
patAD were co-expressed along with
gabT and
yqhD or
yahK as the alcohol dehydrogenase. Unlike the previous study, where
yqhD was used in the final strain, this study saw
yahK producing a higher titer of 1,5-PDO. Cen et al. also made an adjustment in this study by using the CAR-based pathway from 5-HV to 1,5-PDO, therefore, different CAR enzymes were screened to test its efficiency for the production of 5-hydroxypentanal. Ultimately, the CAR from
M. marinum produced the highest 1,5-PDO titer. Another addition is the knock-out of the
gdhA and
gabD genes, which are involved in a separate GA synthesis pathway, in an attempt to reduce its accumulation. The strain consisting of these changes resulted in an overall titer of 9.25 g/L of 1,5-PDO, which is significantly higher than the previous study but is still considered low on the industry scale. Furthermore, a high accumulation of byproducts still remained as well, therefore, this engineered pathway needs to be further optimized and tested [
20].
In a separate study conducted by Wang et al., they explored the charged amino acid catabolic pathway to synthesize various diols, including 1,3-PDO, 1,4-BDO, and 1,5-PDO. However, for the purpose of this discussion, the focus will be primarily on the last compound. Starting with glucose as the carbon source to produce lysine, Wang et al. applied a similar strategy as discussed in prior studies. They inserted a synthetic AMV pathway by introducing
davAB from
P. putida,
gabT, and
yqhD from native
E. coli. Additionally, they employed a CAR-based pathway for the conversion of ω-hydroxy acids (ω-HA) to diols. This pathway specifically consisted of the CAR enzyme from
M. marinum and phosphopantetheine transferase (
sfp) from
B. subtilis. The study revealed that the titer of 1,5-PDO increased exponentially only when lysine was exogenously supplied to the cell culture, highlighting the bottleneck limitation imposed by lysine supply on the pathway. To address this issue, Wang et al. co-expressed
lysC and
dapA, to enhance the lysine degradation pathway. However, even after deleting the
iclR gene in an effort to maintain the precursor supply of oxaloacetate, the highest titer reached only 0.97 g/L [
61].
2.3. Cadaverine
Cadaverine, or 1,5-diaminopentane is a straight-chain diamine that has shown to have many commercial applications, notably in bio-based materials, and is seen in chelating agents [
89]. Cadaverine is naturally formed by bacterial decarboxylation of lysine that occurs during putrefaction of animal tissues. There is a growing interest in enhancing cadaverine production using microbial engineering methods with a focus on renewable carbon sources () [
11].
2.3.1. Cadaverine Production in
E. coli
In
E. coli, the majority of studies focused on the bioconversion of lysine to cadaverine instead of a direct fermentative approach from glucose in order to avoid limitations of precursor availability and cell growth [
89]. There are two main lysine decarboxylases (LDC) that can achieve this conversion: CadA and LdcC, both of which originate from
E. coli. When compared, CadA has a higher catalytic activity than LdcC, thus the former is more frequently used to produce cadaverine. Hong et al. worked to improve the enzymatic stability and activity of the CadA by inserting a combination of mutations: F14C, K44C, L7M, and N8G. This mutated strain produced an overall titer of 157 g/L of cadaverine [
62]. Another study by Kou et al. focused on enhancing the thermal and pH stability of CadA by overexpressing a T88S mutant to achieve an increase in titer to 198 g/L [
63]. Xi et al. also attempted to accomplish this by introducing additional mutations, K477R, E445Q, and F102V, along with T88S. However, this combinatorial mutagenesis approach led to a slight decrease in concentration of 160.7 g/L compared to the previous study [
64]. Gao et al. discovered a combination of different mutations, V12C and D41C, using rational engineering on the CadA. In addition to optimizing the pH conditions during fermentation, this strain achieved the highest cadaverine titer at 418 g/L [
21].
Additionally, Ma et al. co-expressed the CadA gene with CadB, a lysine and cadaverine antiporter fused with a signal sequence PelB, leading to a production titer of 221 g/L [
65]. Huang et al. further built upon this system by deleting the genes responsible for the accumulation of byproducts, including
speE,
speG,
ygjC, and
puuA. However, this knockout strain only produced 32.1 g/L of cadaverine [
66]. An additional effort was made by Moon et al. on modification of the ATP regeneration system within the pathway. They overexpressed
cadA, along with a polyphosphate kinase (
ppk), and pyridoxal kinase (
pdxY), to assist in the conversion of lysine. They further optimized the cell system by introducing hexadecyltrimethylammonium bromide (CTAB) to increase the permeability of the cell membrane. The implementation of both these strategies resulted in the production of 1 M of cadaverine, with a 100% conversion rate from lysine [
67]. Xue et al. achieved a titer of 83.2 g/L by utilizing a similar strategy of co-expressing
cadA with only
pdxY [
68]. In another study, Wang et al. introduced lysine decarboxylase (LDC) from
Hafnia alvei with the mutation, E583G into
E. coli. This heterologous expression saw a decent concentration of 63.9 g/L [
69].
Noh et al. conducted an experiment involving the engineering of a synthetic sRNA scaffold. Using the cadaverine biosynthesis pathway to test their approach, they identified and used 67 genes to construct the strongest scaffold. To determine the effect of the modifications on carbon metabolism, this strategy utilized direct fermentation from glucose. As a result, 13.7 g/L of cadaverine was produced following fed-batch fermentation [
70]. As for the study performed by Wei et al., their experiment focused on altering the fermentation process by using a CO
2 reconversion approach to control pH levels, which reduced cellular damage and led to a titer of 208.2 g/L [
71].
Instead of using glucose as a carbon source, Kwak et al. utilized galactose as the carbon source to produce cadaverine in
E. coli. To metabolize galactose, Kwak et al. engineered the Leloir pathway by overexpressing the genes involved,
galE,
galT,
galK,
galM,
galP, and
pgm. Additionally, they modified the carbon flux to increase the supply of lysine by inserting
asd,
dapA fbr,
dapB,
ddh,
lysA, and
lysC fbr into the chromosome. Disruption of the cadaverine degradation pathway was also attempted by knocking out
speE,
speG,
ygjG, and
puuPA, resulting in a titer of 8.80 g/L [
22].
2.3.2. Cadaverine Production in
C. glutamicum
Corynebacterium glutamicum has shown tremendous success in constructing engineered microbial cells in the past, making it an excellent candidate as the host organism for cadaverine production. Kind et al. utilized a previously engineered
C. glutamicum strain, LYS-12, that hyper-produces lysine as a precursor. In this strain, Kind et al. opted to introduce the constitutive gene,
ldcC from
E. coli, as the lysine decarboxylase, along with a major facilitator permease,
Cg2893, to enhance cellular transport. They additionally deleted the lysine exporter gene,
lysE, to maintain the precursor supply, and removed
NCgl1469, a gene involved in the production of by-product
N-acetyl-diaminopentane. This modified strain resulted in a titer of 88 g/L following fed-batch fermentation using glucose as the starting substrate [
72].
Similarly, the direct fermentation approach was utilized by Li et al., who employed a heterologous co-expression system involving
cadB from
E. coli and the LDC gene from
Hafnia alvei, producing 2.75 g/L of cadaverine [
73]. In another study, Kim et al. used a strong synthetic H30 promoter to express the
ldcC gene from
E. coli, where it significantly increased the activity of lysine decarboxylase. Coupled with the deletion of
lysE, this approach achieved one of the highest titers from glycose in
C. glutamicum, 103.78 g/L of cadaverine [
23].
Kobayashi et al. utilized a different approach by constructing a system with an oxygen-responsive switch that could transition the carbon flux from glycolysis to the pentose phosphate pathway (PPP). This was accomplished through the replacement of the glucose 6-phosphate isomerase (
pgi) promoter with the promoter of lactate dehydrogenase (
ldhA). Furthermore, they deleted the lactate pathway through the disruption of
ldhA, menaquinone oxidoreductase (
pqo), acetate kinase (
ack), and phosphotransacetylase (
pta). However, to improve the supply of oxaloacetate, a precursor of lysine, they introduced a mutation in isocitrate dehydrogenase to reduce its activity. They also introduced a P328S mutation into pyruvate carboxylase (
pyc) to increase its enzymatic activity. Finally, a
lysC-T311I mutant was introduced into the strain to increase lysine by eliminating the feedback inhibition. The final strain produced 0.74 g/L of cadaverine [
74].
In the studies by Buschke et al., xylose was used as the carbon source. They constructed a mutant strain by expressing
xylA and
xylB genes from
E. coli, that converts xylose into xylulose 5-phosphate to be used as a metabolic intermediate [
24]. Using this recombinant strain, Buschke et al. overexpressed
icd, along with fructose bisphosphatase (
fbp), and transketolase (
tkt), enzymes involved in the production of lysine from a carbon source. This strain consumed xylose to achieve 103 g/L of cadaverine, a high titer seen in
C. glutamicum [
25]. Mao et al. also employed xylose as their substrate, but their method centered on the introduction of Beta-xylosidase BSU17580 from
B. subtilis into the strain, facilitated by a PorH anchor protein. This enzyme played a crucial role in generating the xylose supply through the conversion of xylo-oligosaccharides. Along with the overexpression of the
xylA and LDC genes, Imao et al. achieved a yield of 11.6 mM of cadaverine [
75].
2.4. δ-Valerolactam
δ-valerolactam plays a pivotal role in the synthesis of bioplastics, which can subsequently be utilized in the manufacturing of various items and goods [
76]. This class of products has garnered increased attention due to its reliance on non-petroleum-based sources and its exceptional durability, with predictions suggesting that bioplastics production may exceed 2.43 million tons in the near future [
26]. While there are various other compounds capable of synthesizing biopolyamides, δ-valerolactam has been shown to be a potential candidate for large-scale bioplastics production. However, currently, there are limited methods for δ-valerolactam production, achieved either through plant extraction or the chemical conversion of 5-AVA. Therefore, engineered microorganisms have become a focal point, although there have only been a handful of studies reporting on this compound ().
While there have been relatively few studies on the synthesis of δ-valerolactam via metabolic engineering, Xu et al. proposed a novel pathway that included L-pipecolic acid (L-PA) as the intermediate. In this study, Xu et al. uses
E. coli as the host organism to first introduce the
DavB genes from
P. putida to convert L-PA to δ-valerolactam, which produced an initial titer of 90.3 mg/L. Further modifications involved co-expressing DavB with multiple proteins including an apoptosis-inducing protein (
rAIP) from
Scomber japonicus,
Bacillus subtilis glucose dehydrogenase (GDH),
P. putida Δ-piperideine-2-carboxylae reductase (
DpkA), and Oryza sativa lysine permease (
LysP). These changes allowed for δ-valerolactam to be produced directly from L-lysine, resulting in a final titer of 242 mg/L of δ-valerolactam [
76]. In a separate study, Cheng et al. built upon the information from this previous experiment and further optimized the pathway to co-produce δ-valerolactam and 5-AVA. While Cheng et al. also introduced the
RaiP from
S. japonicus, they also expressed a catalase gene,
katE. They supplemented the process with H
2O
2 and adjusted the pH level to the optimal value of 9.0. With these modifications, this strain obtained a significantly improved titer of 6.88 g/L [
26].
2.5. Glutaric Acid (GA)
GA, also known as 1,5-pentanedioic acid, is a dicarboxylic acid that is an important component for the production of polyester and polyamides, which are essential for the manufacturing of fabric and textiles, amongst other materials [
87]. Typically, GA is synthesized via a chemical process using potassium cyanide to catalyze the ring-opening reaction of butyrolactone or using other toxic petroleum-based materials [
30,
90]. Apparently, some of the main issues associated with this approach are the high costs as well as the environmental burdens. Therefore, there is a preference for transitioning to efficient biobased approaches over traditional chemical methods.
2.5.1. GA Production in
E. coli
E. coli is a widely used host microorganism for metabolic engineering due to its fast growth and capacity to express heterologous genes, enabling the rapid construction of efficient microbial systems for producing desired compounds. GA is a compound derived from lysine and 5-AVA, whose pathways require the introduction of heterologous enzymes from different organisms. Two separate studies conducted by Park et al. and Adkins et al. first focused on the production of 5-AVA and then the conversion of 5-AVA to GA in
E. coli (). Both studies synthesized 5-AVA from L-lysine by introducing the davAB genes from
P. putida into an
E. coli, which encodes for the δ-aminovaleramidase and lysine 2-monooxygenase, respectively. The
gabTD genes from
P. putida, which encode for 5-AVA aminotransferase and GA semialdehyde dehydrogenase, were also co-expressed to catalyze the conversion of 5-AVA to GA. Adkins et al. further deleted the
cadA and
ldcC genes, which were involved in a separate lysine degradation pathway to reduce the formation of the byproduct, cadaverine. With similar modifications, Adkins et al. and Park et al. obtained a final titer of 0.82 and 1.7 g/L, respectively [
77,
78].
In a study conducted by Li et al., their approach closely resembled the biosynthesis of 1,5-PDO via the cadaverine pathway, as previously explored by Cen et al [
59]. In this approach,
E. coli native genes
cadA,
patAD, and
gabTD were incorporated to facilitate the synthesis of GA. Furthermore, to reduce the accumulation of cadaverine, Li et al. overexpressed PotE, a bi-directional cadaverine transporter. The study also focused on boosting the supply of lysine by enhancing the precursor oxaloacetate, achieved through the deletion of
iclR. These modifications resulted in minimal byproduct formation, culminating in a final GA titer of 54.5 g/L [
27]. Wang et al. also utilized the cadaverine pathway in their study, albeit with slight variations. They introduced the
patAD genes from
Klebsiella pneumoniae and
gabTD from
Pseudomonas fluorescens into their pathway. Wang et al. also discovered the
patA gene was a rate-limiting enzyme in the 5-AVA synthesis reaction, to which they improved its efficiency via a hydrophobic scanning strategy, where certain hydrophilic residues within the catalytic binding pocket were replaced with a hydrophobic residue. The introduction of a T332A/E120G
patA mutant into the strain resulted in an enhancement of 5-AVA production. The transporter gene,
gabP, was also overexpressed to reduce the accumulation of 5-AVA. This particular strain achieved an impressive titer of GA, reaching 77.62 g/L [
28].
Hong et al. improved the production of GA via the 5-AVA pathway by coupling the overexpression of the
gabTD genes with the Nox gene from
Lactobacillus sanfranciscensis that encodes for an NADPH oxidase. In doing so, it reduced the need for NAD
+ as a cofactor and allowed the overall production of GA to reach 282 mM [
79]. In another study, they inserted the
gabTD genes from
B. subtilis into
E. coli, where they obtained 191 μmol of GA after five cycles of whole-cell bioconversion [
80]. Yang et al. attempted to improve this experiment by using polyvinyl alcohol and polyethylene glycol (PVA-PEG) hydrogel to entrap and immobilize the entire system. With this addition, the production of GA from 5-AVA increased to 995.2 mM [
81]. The same group published another study focusing on the involvement of α-ketoglutaric acid, where they introduced glutamate oxidase (GOX) from
Streptomyces mobaraensis to lower the demand of α-KG by regenerating its supply, with the addition of a catalase to further enhance the system. Despite these changes, the titer of GA lowered to 468.5 mM [
82].
The whole-cell biocatalysts approach was also explored by Wang et al., who used
E. coli to build a synthetic microbial consortium. In this study, they used two engineered strains to express the
davAB and
gabTD genes separatly, as they realized that these two genes had a negative interaction when placed in a single cell strain. This approach resulted in the production of 43.8 g/L [
83].
In a separate study, Wang et al. established a new pathway by using a ketoacid chain elongation strategy in
E. coli. They extend the TCA cycle intermediate α-ketoglutarate to form α-ketoadipate (α-KA) by introducing three enzymes from
S. cerevisiae: homocitrate synthase, homoaconitase, and homoisocitrate dehydrogenase. After the formation of α-KA, they expressed a α-ketoacid decarboxylase, KivD from
L. lactis, and a semialdehyde dehydrogenase,
gabD from
P. putida, to catalyze the conversion to GA. To optimize this alternative pathway, Wang et al. used CRISPR to silence
sucAB genes from the TCA cycle, in order to increase the supply of the precursor, α-ketoglutarate. This novel pathway resulted in a GA titer of 420 mg/L [
76]. The same research group explored the GA biosynthesis using xylose as a carbon source instead of glucose. They also investigated different metabolic pathways to produce GA and found that a combination of the xylose isomerase and Weimberg pathways yielded the highest titer of 602 mg/L [
29].
Yu et al. proposed an alternative strategy to produce GA in
E. coli, by utilizing α-ketoglutarate in a reported glutaconate biosynthesis pathway. They first formed trans-glutaconyl-CoA by expressing
hgdH,
gctAB, and
hgdC from
A. fermentans, and
hgdAB from
C. symbiosum. This compound can then be converted to form GA using either trans-enoyl-CoA reductase from
E. gracilis (egTer) or
T. denticola (tdTer), along with the
E. coli thioesterase enzymes (
TesB). Using this pathway, the best strain obtained 3.8 mg/L of GA with egTer, although it accumulated a higher amount of glutaconate with 27.7 mg/L [
88].
Another study by Zhao et al. explored the production of GA in
E. coli through the
Thermobifida fusca native reverse adipate degradation pathway (RADP), with the overexpression of five enzymes: Tfu_0875, Tfu_2399, Tfu_0067, Tfu_1647, and Tfu_2576-7. Using this pathway, Zhao et al. attempted to use both acetyl-CoA and malonyl-CoA as precursors for GA. They increased the supply of malonyl-CoA by using the antibiotic, cerulenin, to inhibit a fatty acid synthesis pathway. Additionally, to increase acetyl-CoA, genes encoding for L-lactate dehydrogenase (
ldhA), acetyl-CoA acetyltransferase (
atoB), and formate C-acetyltransferase I (
pflB) were deleted to disrupt competitive pathways and reduce byproduct formation. The resulting yield of GA from this engineered
E. coli strain was approximately 4.8 g/L [
91]. Years later, another study was published extending upon this established pathway. As the previous experiment found a limitation with the supply of malonyl-CoA, Sui et al. introduced the
matB and
matC genes from
Clover rhizobia to convert malonic acid to malonyl-CoA. The addition of malonic acid into this pathway increased the overall titer to 6.3 g/L [
92].
2.5.2. GA Production in
C. glutamicum
C. glutamicum has shown to be an excellent microorganism for the synthesis of amino acids such as lysine and glutamate via fermentation [
87]. As GA biosynthesis requires the catabolism of lysine, the engineering of lysine producing
C. glutamicum was thought to be a promising approach. Rohles et al. conducted a study in
C. glutamicum using strategies similar to those of Park et al. Deletion of the
lysE gene that encodes for the lysine exporter reduced the amount of lysine excretion, resulting in the production of a total 7 g/L of GA [
84]. In a subsequent study, Rohles et al. further refined their approach by using native
gabTD genes and overexpressing a 5-AVA importer,
NCg10464, which would re-import 5-AVA back into the cell to maintain the supply for GA production. Amplifying the transport protein significantly improved the final titer of GA after fed-batch fermentation to 90 g/L, with little to no presence of any known byproduct [
85].
Kim et al. adopted a different strategy, where they used a synthetic promoter H30 to express the
davAB and
gabTD genes to enhance the enzyme activity. Additionally, Kim et al. fused the
davB gene with an N-terminal His6-tag, with the intention of improving its solubility and expression within the cell. As a result, this incorporation led to 24.5 g/L of GA to be produced [
86]. Han et al. continued to optimize the engineered
C. glutamicum strain by overexpressing or deleting 11 different genes (
icd,
ddh,
dapB,
pyc,
dapA,
ppc,
lysA,
lysI,
lysE,
lysC, and
pck) that were found to increase the supply of lysine as the precursor. Additionally, they introduced the gene
ynfM, which encodes for a GA exporter, into the strain, producing a final titer of 105.3 g/L of GA [
30].
Perez-Garcia et al. developed a new pathway that does not require oxygen to produce GA. For this, they employed the use of the transaminase-oxidoreductase pathway to form 5-AVA by heterologous expression of the
patAD and
ldcC genes from
E. coli. To reduce byproduct formation and increase the production of 5-AVA as a precursor, they deleted the transcriptional repressor (
sugR), lactose dehydrogenase (
ldh), N-acetyltransferase (
snA), and diamine exporter (
cgmA). In this study, Perez-Garcia examined the
gabTD genes from three different
Pseudomonas species to produce GA, with the gene from
P. stutzeri performing the best. Additionally, the glutamate biosynthesis pathway was eliminated by deleting the
gdh gene. The final modified
C. glutamicum strain produced 25.2 g/L of GA through fed-batch fermentation [
87].
2.6. Glutaconic Acid
Glutaconic acid is an unsaturated dicarboxylic acid that can be reduced to synthesize GA. As such, it has similar applications, being a precursor to polyamides and other biodegradable polymers [
31]. Currently, the only natural biosynthetic pathway is found in anaerobic organisms such as
Acidaminococcus fermentans and
Clostridium symbiosum, which are capable of removing the ammonia from glutamate to convert into glutaconic acid [
9]. However, since these microbes cannot produce glutaconic acid on an industrial scale, alternative microbial production methods have been developed ().
In prior studies, it has been shown that anaerobic bacteria are capable of converting glutamate to glutaconate in fermentation conditions. From this information, Djurdjevic et al. attempted to modify
E. coli to produce glutaconate by introducing enzymes from those bacteria. Specifically, they heterologously introduced six genes: 2-hydroxyglutarate dehydrogenase (
hgdH), glutaconate CoA-transferase (
gctAB), 2-hydroxyglutaryl-CoA dehydratase (
hgdC) from
A. fermentans, and 2-hydroxyglutaryl-CoA dehydratase (
hgdAB) from
C. symbiosum. When this modified strain was cultivated in an anaerobic medium to replicate the conditions of an anaerobic bacteria, it yielded a glutaconate concentration of approximately 2.7 mM [
31].
Sun et al. conducted another study in which they developed a novel pathway in
E. coli using the estradiol ring catechol degradation pathway. Through this pathway, catechol can be catabolized to ketoacid, 4-oxalocrotonate, which can then be converted to glutaconic acid. Sun et al. first started this experiment by introducing the genes directly involved in this reaction:
C23O and
DmpI from
P. putida, and
DmpC from
R. eutropha. Once 4-oxalocrotonate was reached, KivD from
L. lactis and
gabD from
P. putida are capable of converting it into glutaconic acid. To synthesize catechol, Sun et al. used salicylic acid as the precursor, which required the expression of
nahG to catalyze this bioconversion. Furthermore, the
entC and
pchB genes were also expressed in
E. coli to help produce salicylic acid from glucose. This modified strain resulted in a titer of 35.3 mg/L of glutaconic acid [
93].
2.7. 5-Hydroxyvaleric Acid
5-hydroxyvaleric acid (5-HV), also referred to as 5-hydroxypentanoic acid, is a versatile compound with a wide range of applications. It is used in the production of various materials, such polyurethanes and polyhydroxyalkanoates (PHA) [
32]. In addition, 5-HV serves as a precursor to 1,5-pentanediol. Similar to GA, the current chemical approach primarily relies on furfural derivatives, involving costly substrates and catalysts, and is a highly resource-intensive process [
33]. As of recently, the interest in PHAs have been rising due to its applicability in packaging and in the biomedical industry. Given that 5-HV is a major building block for the material, the interest in its production has risen significantly as well.
In a previous study by Rohles et al., they established an innovative pathway for synthesizing 5-AVA, a valuable precursor to 5-HV, by incorporating the
davAB and
davTD genes from
P. putida into
C. glutamicum. In this study, Sohn et al. attempts to increase the production of 5-HV by building onto this established 5-AVA pathway, while making slight modifications such as introducing an appropriate aldehyde reductase that would efficiently convert glutarate semialdehyde to 5-HV (). In the pathway found by Rohles et al., the absence of 5-HV demonstrated a clear preference in the system's metabolic flux toward GA. To address this issue, Sohn et al. deleted the
gabD gene to eliminate the GA pathway entirely. Despite this deletion, the overall titer of 5-HV did not improve, indicating that further investigation is needed to understand this metabolic flux. Additionally,
yahK gene from
E. coli was used in the final strain as the aldehyde reductase. The culmination of these modifications resulted in an overall titer of 52.1 g/L of 5-HV following fermentation, however, a significant amount of GA remained present [
32].
In contrast, instead of using an engineered
C. glutamicum strain, Chen et al. utilized
E. coli to produce 5-HV. The main pathway is very similar to Sohl et al. with the implementation of the
davAB gene, however, a key difference is the use of a native enzyme, aminobutyrate aminotransferase encoded by
gabT to convert 5-AVA to glutarate semialdehyde. Additionally, Chen et al. screened
yqhD and
yahK genes from
E. coli and
yajO gene from
P. putida for the best aldehyde reductase activity. Although it was found that the
yqhD gene performed the best, a dual vector system was employed, constructing a secondary promoter with
yajO to overexpress the enzyme activity. Chen et al. also introduced genes that encoded for a 5-aminovalerate exporter and a lysine-specific permease,
gabP and
lysP, respectively, to increase the transport rate of key intermediates into the cell. Furthermore, a hok/sok system was introduced to reduce plasmid instability. Despite these changes differing from Sohl et al., the outcome of this study yielded a lower titer of 5-HV at 35.6 g/L [
33].
3. C6 Chemicals
3.1. cis,cis-Muconic Acid
MA is a pivotal six-carbon platform chemical serving as a precursor for a diverse range of polymers used in consumer plastics such as polyurethane and terephthalate [
94,
95]. Furthermore, MA can be converted into AA, which is a building block for nylon-6,6 synthesis [
44]. However, the petroleum-derived production of AA involves carcinogenic substrates and intermediates such as benzene, cyclohexane, cyclohexanol, and cyclohexanone, which cause environmental concerns [
95]. To date, microbial production of MA has been achieved in various chassis microorganisms through several different pathways. Remarkably, this is the only approach for C6 chemical production via ring-fission oxidation of catechol ().
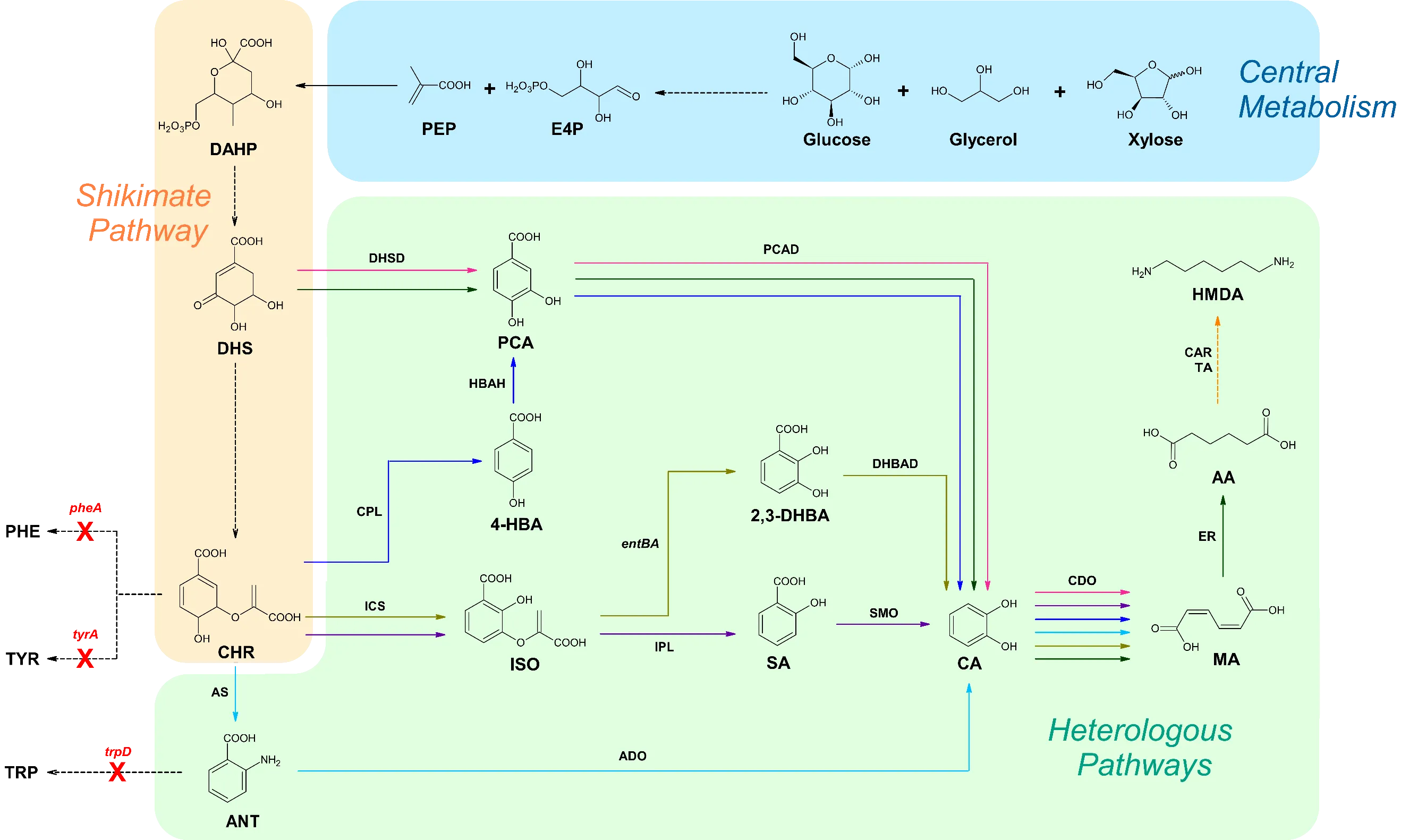
. Overview of metabolic pathways extended from the shikimate pathway. Solid arrows signify direct steps. Dashed arrows represent multiple steps. Red X’s portray genes that are disrupted. Pink pathway corresponds to references [
34,
35,
96,
97,
98,
99,
100,
101,
102]. Purple pathway correlates to reference [
103]. Dark blue pathway corresponds to reference [
104]. Light blue pathway correlates to reference [
105]. Olive pathway corresponds to references [
106,
107]. Dark green pathway correlates to reference [
40]. Orange pathway corresponds to reference [
43]. Black arrows represent steps utilized by multiple pathways. E4P, erythrose-4-phosphate; PEP, phosphoenolpyruvate; DAHP, 3-deoxy-D-arabino heptulosonate-7-phosphate; DHS, 3-dehydroshikimate; CHR, chorismate; PHE, phenylalanine; TYR, tyrosine; TRP, tryptophan; PCA, protocatechuate; 4-HBA, 4-hydroxybenzoic acid; ISO, isochorismate; ANT, anthranilate; 2,3-DHBA, 2,3-dihydroxybenzoic acid; SA, salicylate; CA, catechol; MA,
cis,
cis-muconic acid; AA, adipic acid; HMDA, 1,6- hexamethylenediamine; DHSD, 3-dehydroshikimate dehydratase; PCAD, protocatechuate decarboxylase; CPL, chorismate pyruvate lyase; HBAH, 4-hydroxybenzoic acid hydrolase; ICS, isochorismate synthase; IPL, isochorismate pyruvate lyase; SMO, salicylate 1-monooxygenase; DHBAD, 2,3-dihydroxybenzoic acid decarboxylase; AS, anthranilate synthase; ADO, anthranilate 1,2-dioxygenase; CDO, catechol 1,2-dioxygenase; ER, enoate reductase; CAR, carboxylic acid reductase; TA, transaminase.
3.1.1 MA Production in
E. coli
The earliest reported microbial production of MA was pioneered by Draths and Frost in
E. coli, utilizing glucose as the substrate. The pathway begins with the dehydration of DHS, an intermediate in the shikimate pathway. Subsequently, the conversion of DHS to MA is facilitated by the introduction of
aroZ (encoding DHSD) and
aroY (encoding PCAD) from
K. pneumoniae, as well as
catA (encoding CDO) from
A. calcoaceticus [
96]. Niu et al. optimized this strategy by employing feedback inhibition-insensitive
aroF and integrating
aroZ into the
E. coli chromosome while disrupting
aroE (encoding shikimate dehydrogenase) to prevent the consumption of DHS. These improvements resulted in the production of 36.8 g/L of MA under fed-batch fermentation conditions [
97]. Further enhancements involved the disruption of
ptsG (encoding phosphoenolpyruvate phosphotransferase) and
pykA (encoding pyruvate kinase 2) to increase phosphoenolpyruvate (PEP) availability and the disruption of
tyrR (a tyrosine dependent transcriptional regulator) to prevent the negative transcriptional control of
aroG and
aroF. Moreover,
aroB (encoding DHQ synthase),
aroD (encoding DHQ dehydratase),
galP (encoding D-galactose transporter),
ppsA (encoding PEP synthase),
aroG (encoding DAHP synthase),
aroF (encoding DAHP synthase), and codon-optimized
asbF (encoding DHS dehydratase) were overexpressed, leading to the production of 64.5 g/L of MA through fed-batch fermentation [
34]. In addition to the single culture process, Zhang et al. reported coculture strategies with two separate
E. coli strains, each carrying a portion, or the full MA pathway reported by Draths and Frost. A transporter protein encoded by
shiA was employed to facilitate the cross-membrane transportation of the intermediate DHS. The optimized cocultures utilized glycerol as the substrate and produced 2 g/L with a batch bioreactor [
108]. This group also devised another coculture strategy that utilized two engineered
E. coli strains, one of which utilized glucose while the other utilized xylose, resulting in the production of 4.7 g/L of MA from a mixture of glucose and xylose in a bioreactor [
109]. Fujiwara et al. employed a parallel metabolic pathway engineering approach by simultaneously expressing the Dahms pathway from
Caulobacter crescentus alongside a MA biosynthesis pathway. The primary goal of this strategy was to direct the host organism’s metabolic processes in such a way that glucose was exclusively utilized for MA synthesis, while xylose was allocated for biomass production. For this purpose,
eda,
ppsA,
ppc, and
pck disruptions were incorporated to prevent the intermediates generated through glycolysis from entering the TCA cycle which yielded 4.09 g/L of MA using minimal media [
103].
Instead of diverting DHS from the shikimate pathway to produce MA, Lin et al. utilized chorismate (CHR) as a precursor and introduced
entC (encoding ICS),
pchB (encoding IPL), codon-optimized
nahG (encoding SMO), and
catA (encoding CDO) into an engineered phenylalanine overproducing
E. coli strain QH4 with
pheA and
tyrA disrupted. To further optimize the process,
aroL (encoding shikimate kinase),
ppsA,
tktA (encoding transketolase), and feedback-inhibition resistant
aroG were overexpressed to increase the availability of CHR, leading to the production of 1453.64 mg/L of MA in shake flasks [
110]. Fujiwara et al. explored the expression of an AroC-MenF fusion protein to increase the availability of intermediates while expressing the MA biosynthetic pathway previously established by Lin et al. Fusion proteins enhance reaction efficiency by maintaining enzymes in close proximity to one another, facilitating a smoother transfer of intermediates between their respective enzymes. This strategy achieved a production 4.45 g/L of MA under pH-controlled jar fermenter conditions [
111].
Inspired by the ubiquinone pathway, Sengupta et al. devised the conversion of 4-HBA into MA by expressing
ubiC (encoding CPL),
pobA (encoding HBAH),
aroY, and
catA. To increase the availability of precursors,
ptsH,
ptsI, and
crr, which code for proteins involved in the phosphotransferase system (PTS), and
pykF (pyruvate kinase 1) were deleted from the
E. coli genome. These modifications resulted in the production of 170 mg/L of MA [
104].
Additionally, Sun et al. reported the production of 389.96 mg/L of MA by hybridizing the tryptophan biosynthesis and anthranilate degradation pathways, which was achieved by expressing
trpEG (AS), ADO from
P. aeruginosa, and CDO from
P. putida. The
trpD gene was deleted to prevent the formation of tryptophan, which would otherwise consume anthranilate [
105]. Yan and Zheng groups produced 480 mg/L and 605.18 mg/L of MA, respectively, by connecting 2,3-DHBA biosynthesis with its degradation pathway.
EntCBA, DHBAD from
K. pneumoniae, and CDO from
P. putida were expressed while
entE was knocked out to prevent the consumption of 2,3-DHBA [
106,
107].
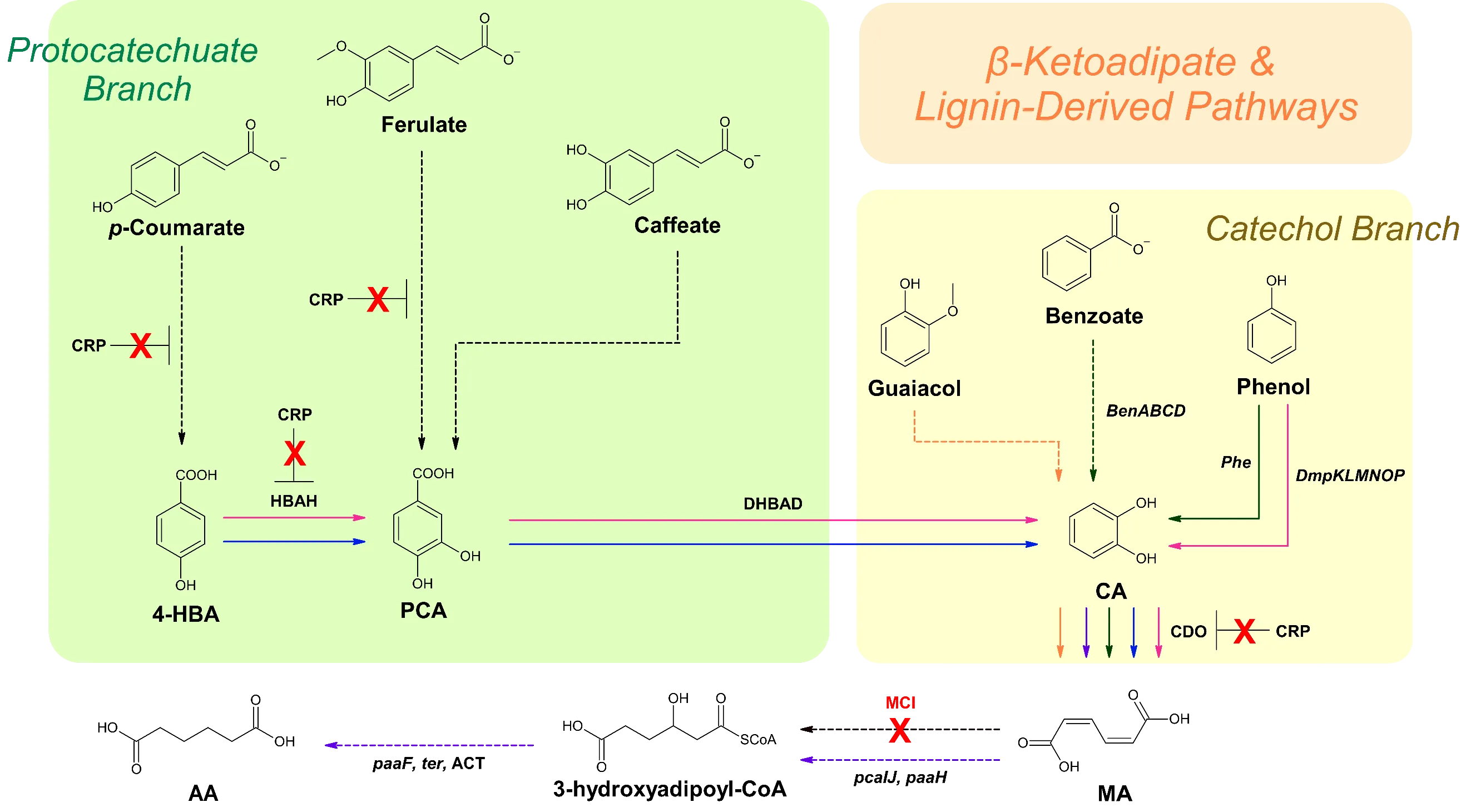
. Schematic of biosynthesis pathways for MA and AA production that utilize lignin-derived feedstocks. Solid arrows signify direct steps. Dashed arrows represent multiple steps. Red X’s portray enzymes that are disrupted. Dark green pathway correlates to reference [
37]. Orange pathway corresponds to reference [
38]. Pink pathway correlates to reference [
112]. Dark blue pathway corresponds to references [
113,
114]. Purple pathway correlates to reference [
41]. Black arrows represent steps utilized by multiple pathways. 4-HBA, 4-hydroxybenzoic acid; PCA, protocatechuate; CA, catechol; MA,
cis,
cis-muconic acid; AA, adipic acid; CRP, catabolite repression protein; HBAH, 4-hydroxybenzoic acid hydrolase; DHBAD, 2,3-dihydroxybenzoic acid; CDO, catechol 1,2-dioxygenase; ACT, adipoyl-CoA thioesterase; MCI, muconate cycloisomerase.
3.1.2. MA Production in
S. cerevisiae
Curran et al. pioneered the production of MA in
S. cerevisiae by introducing codon optimized DHSD from
Podospora anserina, PCAD from
Enterobacter cloacae, and CDO from
Candida albicans, creating the MuA12 strain. Furthermore, this strain harbors
aro3 and
zwf1 knockouts and overexpresses
tkl1 and feedback-resistant
aro4 to increase the flux of precursors. They achieved a production of 141 mg/L of MA in a shake flask culture [
98]. Suastegui et al. further refined the MuA12 strain by incorporating
aroZ from
P. anserina,
aroY from
K. pneumoniae, HQD2 gene (CDO) from
C. albicans, and
tkl1, resulting in 559.5 mg/L of MA [
99]. Leavitt et al. employed an adaptive laboratory evolution and rational metabolic engineering strategy for MA production. An aromatic amino acid-overproducing
S. cerevisiae strain was employed due to the innate ability to synthesize greater amounts of shikimate pathway intermediates. This strain was selected through anti-metabolite selection. Its derivative strain MuA-5.01.1.02+aro1t+scPAD1 produced 2.1 g/L of MA in a fermentor [
100].
Wang et al. engineered the endogenous shikimate pathway in
S. cerevisiae to produce 20.8 g/L of MA using a biosensor-aided genome engineering approach. The MA-responsive biosensor BenM was used to identify cells with enhanced MA production capabilities due to the presence of multiple copies of AroY.B and AroY.Ciso, which encode functional subunits of PCAD from
K. pneumoniae. Restoring
URA3 prototrophy, introducing Aro1p
ΔE, Aro4p, and additional copies of PaAroZ and CaCatA were found to increase MA yields [
35].
Suastegui et al. disrupted Ric1, a transcriptional repressor of several genes in the aromatic amino acid pathway and overexpressed
RKI1 (encoding ribose-5-phosphate isomerase), which promotes the accumulation of E4P and flux redirection into the aromatic amino acid pathway. These modifications in addition to the introduction of a non-native MA biosynthesis pathway resulted in 2.7 g/L of MA [
101]. Liu et al. overexpressed an MA biosynthesis pathway alongside a xylose fermentation pathway to facilitate the production of MA using glucose and xylose as carbon sources. Specifically, the xylose isomerase gene from
Bacteroides valgutus, pentose phosphate pathway genes from
S. cerevisiae,
Aro1 with an
AroE stop codon, and feedback inhibition-resistant
Aro4opt were expressed. The addition of CA to the media increased MA titer to 1286 mg/L, surpassing the yield obtained when culturing with only glucose and xylose [
102].
3.1.3. MA Production in
C. glutamicum
The superior amino acid production capabilities of
C. glutamicum suggest its suitability for the production of MA via the aromatic amino acid pathway [
115]. Becker et al. engineered
C. glutamicum to produce MA by deleting
catB (MCI) and overexpressing
catA. Endogenous
BenABCD and
Phe facilitate the degradation of benzoate and phenol, respectively, enabling the organism to utilize lignin as a carbon source (). When cultivated in the presence of CA, the created MA-2 strain produced the highest reported titer of 85 g/L of MA in 60 h [
37]. Lee et al. introduced a previously established DHS-derived MA biosynthetic pathway in
C. glutamicum and engineered the endogenous shikimate pathway. This was achieved by disrupting
aroE, along with
cg1835 (encoding shikimate dehydrogenase),
pcaGH (encoding PCA dioxygenase alpha/beta subunits), and
catB (encoding MCI) to prevent carbon flow into the TCA cycle. Codon-optimized
aroY and
kpdBD (encoding 4-HBA decarboxylase from
K. pneumoniae) was overexpressed to convert PCA into CA, which was subsequently converted to MA, achieving a production of 54 g/L of MA from a 50 L fed-batch fermentation [
115]. Shin et al. aimed to increase the availability of PEP, which is a precursor in the shikimate pathway. Introducing a Δ
ptsI knockout prevents the PTS system from consuming PEP at the cost of limiting cell growth. To address this limitation,
iolR, which encodes a transcriptional regulator, was inactivated. Additionally,
qsuB,
aroY, and
kpdBD were overexpressed to facilitate the conversion of DHS to MA, resulting in 4.5 g/L of MA [
116].
3.1.4. MA Production by Other Microorganisms
Amycolatopsis sp. ATCC 39116 is capable of the natural production of MA on a micro-scale and favors lignin-based aromatic compounds such as CA, guaiacol, phenol, toluene,
p-coumarate, and benzoate as carbon sources over sugar. Barton et al. deleted AATC3_02010018510, which encodes a putative MCI, to promote the accumulation of MA. Using guaiacol as the feedstock, 3.1 g/L of MA was produced in a fed-batch fermentor [
38].
P. putida KT2440 exhibits native production of MA via the CA branch of the β-ketoadipate pathway (). To optimize MA production, Vardon et al. aimed to funnel both the PCA and CA branch carbon fluxes towards MA production. This was achieved by disrupting
pcaHG and
catBC, which prevents PCA and CA progression through the β-ketoadipate pathway.
aroY,
dmpKLMNOP (encoding phenol hydroxylase), and
asbF were introduced to further accumulate CA. This strategy produced 13.5 g/L of MA after 78.5 h using lignin-derived compounds as the starting materials [
112]. By expressing codon-optimized
AroY and its subunits
EcdB and
EcdD from
E. cloacae, 15.59 g/L of MA was produced in fed-batch conditions supplemented with
p-coumarate and glucose [
113]. Subsequent deletion of the CRP reduced accumulation of the precursor 4-HBA and increased MA production from
p-coumarate by 70% and doubled from ferulate [
114]. Another strategy involved the overexpression of
catA and
catA2 along with
catBC,
endA-1, and
endA-2 (both encoding endonucleases) knockouts, which achieved an MA titer of 64.2 g/L from lignin-derived catechol [
36].
3.2. Adipic Acid
AA, also known as hexanedioic acid, is a derivative of MA with a wide variety of applications in the production of nylon-6,6, polyurethanes, lubricant components, polyester polyols, and food additives [
117]. However, the conventional chemical synthesis of AA involves benzene, which yields harmful byproducts such as cyclohexane, cyclohexanol, and cyclohexanone, which are not only carcinogenic but also pose environmental contamination risks [
117]. To address these concerns, a benzene-free bio-based approach was developed that utilizes glucose as the feedstock [
96].
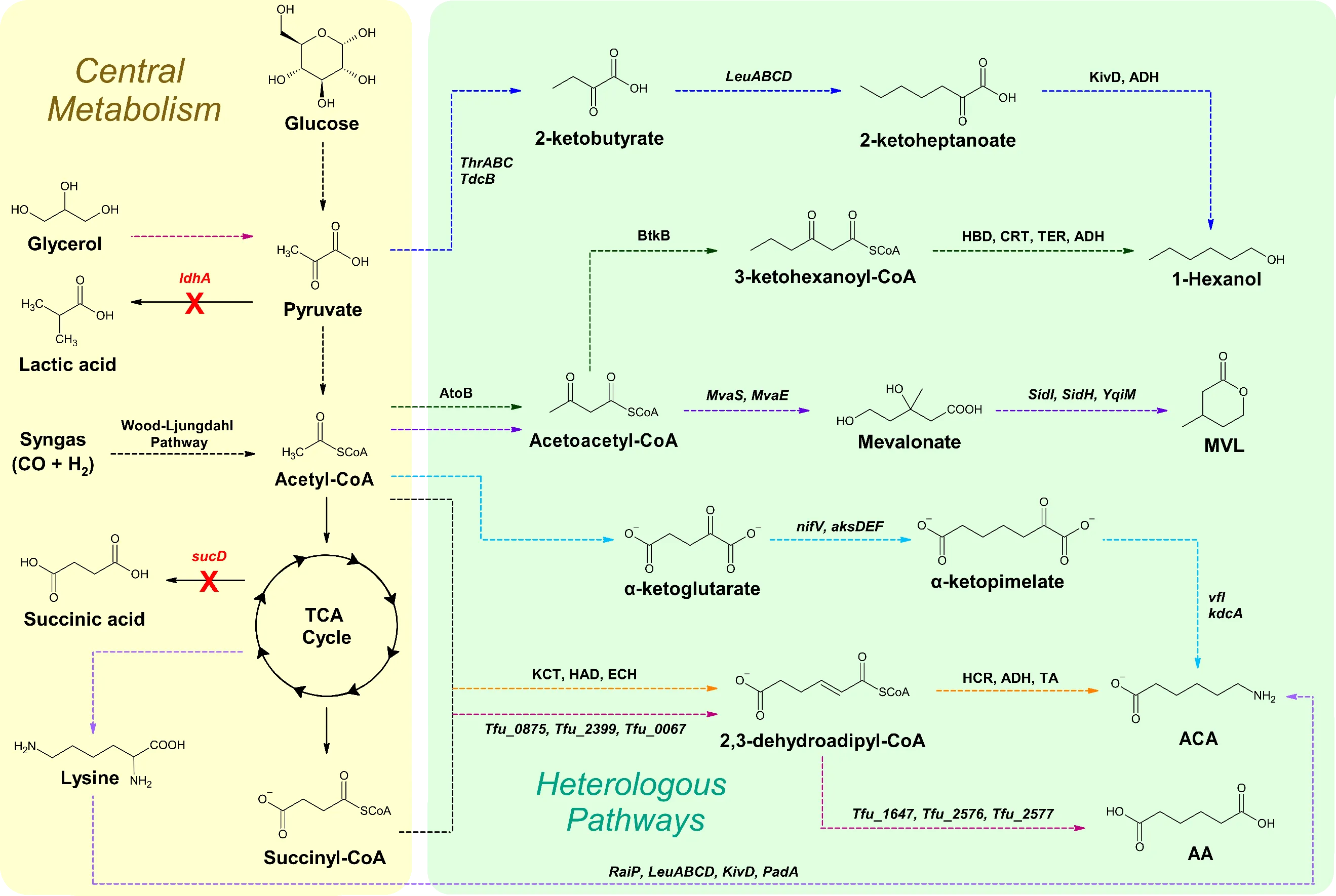
. Overview of biosynthetic pathways extended from the TCA cycle for the production of C6 chemicals. Solid arrows signify direct steps. Dashed arrows represent multiple steps. Red X’s portray genes that are disrupted. Light blue and orange pathways correspond to reference [
44]. Purple pathway correlates to reference [
46]. Pink pathway corresponds to reference [
39]. Light purple pathway correlates to reference [
118]. Dark green pathway corresponds to references [
47,
48,
49,
50]. Dark blue pathway correlates to reference [
59]. Black arrows represent steps utilized by multiple pathways. MVL, β-methyl-δ-valerolactone; ACA, 6-aminocaproic acid; AA, adipic acid; KivD, α-ketoisovalerate decarboxylase; ADH, alcohol/aldehyde dehydrogenase; AtoB, acetyl-CoA acetyltransferase; BktB, β-ketothiolase; HBD, 3-hydroxybutyryl-CoA dehydrogenase; CRT, crotonase; TER,
trans-enoyl-CoA reductase; KCT, β-ketoadipyl-CoA thiolase; HAD, 3-hydroxyadipyl-CoA dehydrogenase; ECH, enoyl-CoA hydratase; HCR, hexenoyl-CoA-reductase; TA, transaminase.
3.2.1. AA Production in
E. coli
Joo et al. explored the capability of ERs from a variety of microorganisms to reduce the double bond(s) in MA and 2-hexenedioic acid to produce AA in
E. coli. Notably, ER from
Bacillus coagulans (ER-BC) had an in vivo substrate conversion rate of 94.3% for MA and 99% for 2-hexenedioic acid after 24 h [
119].
Zhao et al. reconstructed the reverse adipate-degradation pathway from
T. fusca in
E. coli by overexpressing the genes Tfu_0875, Tfu_2399, Tfu_0067, Tfu_1647, Tfu_2576, and Tfu_2577 (). Initially, this yielded 0.3 g/L of AA in suboptimal broth. To remove competing pathways,
ldhA (encoding lactate dehydrogenase),
sucD (encoding succinyl-CoA synthase alpha subunit), and
atoB were deleted. After optimizations, 68.0 g/L of AA was produced using fed-batch fermentation [
39].
3.2.2. AA Production by Various Microorganisms
Raj et al. reported the production of 2.59 mg/L of AA in
S. cerevisiae utilizing glucose as the starting material. An MA biosynthesis pathway was introduced. Subsequently, ER-BC was overexpressed to complete the conversion of MA into AA. However, CDO necessitates oxygen as a substrate while ER-BC requires anaerobic conditions for functional maturation. This challenge was addressed through a three-stage fermentation process.
S. cerevisiae was initially cultivated under aerobic conditions to achieve the desired cell density. Thereafter, anaerobic conditions were created to facilitate the functional formation of ER-BC. Finally, the process reverted to aerobic conditions to complete AA production [
40].
Niu et al. designed an AA biosynthesis pathway that harnessed both endogenous and heterogenous enzymes within
P. putida KT2440. This pathway effectively converted lignin-derived aromatic compounds into either PCA or CA, which were then funneled into the β-ketoadipate pathway. The native enzymes
pcaIJ and ACT, coupled with the overexpression of
paaH,
paaF, and
ter, and the deletion of
pcaF and
paaJ facilitated the production of 2.5 g/L of AA under fermentor-controlled conditions [
41].
Since
Pseudomonas taiwanensis VLB120 possesses high tolerance to highly toxic substrates such as cyclohexane, Bretschneider et al. introduced a heterologous biosynthesis pathway to produce HHA from cyclohexane. Cytochrome P450 monooxygenase (CPMO), cyclohexanol dehydrogenase (CHD), cyclohexanone monooxygenase (CHMO), and lactonase (Lac) from
Acidovorax sp. CHX100 were expressed to produce HHA. Additionally, ADH and aldehyde dehydrogenases (AldDH) from
Acidovorax sp. CHX100 were overexpressed to complete the conversion of HHA to AA, producing 10.2 g/L of AA under fed-batch conditions [
42].
3.3. 1,6-Hexamethylenediamine (HMDA)
HMDA, also named 1,6-hexanediamine or 1,6-diaminohexane, serves as a precursor for the production of nylon-6,6 and various resins [
44,
120]. Beyond its application in nylon production, HMDA is also utilized in the textile and water treatment industries [
120]. So far, some whole-cell bioconversion approaches were reported for its production.
Wang et al. engineered a strategy in which a CAR and TA enzyme cascade is employed to produce HMDA in
E. coli, using AA as the feedstock. This work tested the use of a variety of CARs and TAs, each that utilize their respective substrates. The strain with the highest HMDA titer, DAH86, expressed CARs from
Mycobacterium smegmatis MCS 155 (MSM CAR) and
Mycobacteroides abscessus (MAB CAR), and TAs from
Silicibacter pomeroyi (SPTA) and
E. coli (PatA), resulting in the production of 53.11 mg/L of HMDA. Since CARs are NADPH-dependent enzymes,
sthA, which encodes a transhydrogenase responsible for the conversion of NADH to NADP, was disrupted to increase the availability of NADPH. This modification raised the titer to 238.5 mg/L in a shake-flask culture [
43]. A patent converted (S)-2-amino-6-oxopimelate, an intermediate of the lysine biosynthesis pathway, into HMDA in six steps [
121]. An alternative approach converted acetyl-CoA into HMDA in polyhydroxyalkanoate-accumulating bacteria by utilizing CoA-dependent elongation or analogous enzymes associated with carbon storage pathway [
122].
3.4. 6-Aminocaproic Acid (ACA)
ACA, also known as 6-aminohexanoic acid, serves as a precursor for nylon-6 and is used in the medical field as an antifibrinolytic drug [
123]. The current chemical synthesis of ACA is done by ring fission of caprolactam, a derivative of benzene [
44]. Microbial synthesis approaches were developed through carbon chain elongation cycles.
Turk et al. devised two pathways to produce ACA in
E. coli, utilizing glucose as the substrate. The first synthetic pathway was inspired by the adipyl-CoA route which involves the combination of succinyl-CoA and acetyl-CoA to produce 3-oxoadipyl-CoA which is subjected to further reduction. However, conversion rates for this strategy were found to be below 5%. The second strategy is based on the branched-chain amino acid degradation pathway which occurs in methanogenic archaea, referred to as the α-ketopimelate route. In this pathway, α-ketoacids are decarboxylated into their corresponding aldehydes.
NifV from
Azotobacter vinelandii, codon-optimized
aksD,
aksE, and
aksF from
Methanococcus aeolicus,
vfl from
Vibrio fluvialis, and
kdcA from
Lactococcus lactis were introduced. Due to the oxygen-sensitive nature of
aksE and
aksE, special considerations were made during fed-batch fermentation, ultimately resulting in the production of 160 mg/L of ACA [
44]. An alternative approach yielded 24.12 mg/L of ACA from lysine via a sequence of carbon-chain-extension cycles involving condensation, isomerization, oxidation, and decarboxylation reactions which were catalyzed by the enzymes
LeuABCD and
PadA from
E. coli MG1655,
RaiP from
Scomber japonicas, and KivD from
L. lactis [
118]. A patent reported a yield of 4783 mg/kg of ACA when expressing
vfl from
V. fluvialis,
kdcA from
L. lactis, and TA from
P. aeruginosa in
S. cerevisiae [
45]. Another patent reported a strategy that converts (S)-2-amino-6-oxopimelate, a lysine biosynthesis pathway intermediate, into ACA in four steps [
121].
Additionally, Bretschneider et al. reported a mixed species strategy that allows the ACA production from cyclohexane.
P. taiwanensis strains carrying the upstream cascade converting cyclohexane to 6-hydroxyhexanoic acid (HHA) and
E. coli strains carrying ADH and ω-TA responsible for downstream conversion of HHA into ACA, were co-cultured to circumvent complications when expressing the whole pathway in a single species, which were likely caused by improper folding of enzymes and high metabolic burden. This approach led to an 86% yield and 7.7 U g
−1 CDW of the initial formation rate of ACA [
124].
3.5. β-methyl-δ-valerolactone (MVL)
Polyurethanes (PUs) are compounds with versatile applications in adhesives, sealants, elastomers, and foams [
59]. Like most polymers, PUs are petroleum-derived and non-biodegradable material [
125]. To lessen the environmental impact, efforts have been made to develop alternatives that are biodegradable or derived from renewable materials. MVL is a C6 compound that is used to create precursors for recyclable PUs [
46]. Poly(MVL) is a rubbery polyester that can be produced by ring-opening polymerization of MVL, which is then incorporated into PUs [
126]. MVL can be produced microbially, which is more environmentally friendly than traditional chemical synthesis methods [
46].
Xiong et al. established a total biosynthesis pathway for the production MVL in
E. coli utilizing glucose as the substrate by combining the mevalonate pathway with the fungal siderophore biosynthesis pathway (). Overexpression of endogenous
atoB (encoding acetyl-CoA acetyltransferase), and
MvaS (encoding HMG-CoA synthase) and
MvaE (HMG-CoA reductase) from
Lacticaseibacillus casei lead to the accumulation of 14.6 g/L and 88 g/L of mevalonate in shake flasks and bioreactors, respectively. However, MVL biosynthesis was limited to 270 mg/L due to the less efficient downstream pathway even though the codon-optimized
sidI (acyl-CoA ligase) and
sidH (enoyl-CoA hydratase) from
A. fumigatus in addition to mutant
YqjM with C26D and I69T mutations from
B. subtilis were expressed. Additionally, a semisynthetic approach was developed to produce MVL through biosynthesizing mevalonate followed by chemical conversion of mevalonate to MVL [
46].
3.6. 1-Hexanol
1-Hexanol, a versatile chemical compound, finds broad applications in biofuels, solvents, plasticizers, pesticides, flavorings, and as a building block for chemical synthesis [
49]. 1-Hexanol has a higher cetane number and energy density than short-chain alcohols such as 1-butanol, which make it an attractive option to blend into diesel [
127]. Furthermore, 1-hexanol can be converted into 1-hexene, which can be utilized as a platform chemical for the production of jet fuel [
128]. 1-Hexanol is currently synthesized using petroleum-based routes that require harsh reaction conditions [
129]. To reduce the dependency on fossil fuels and unsustainable processes, efforts have been made to research the production of alternative and renewable fuels.
Zhang et al. reported the extension of the Ehrlich pathway, redirecting 2-keto acid intermediates from amino acid biosynthesis pathways towards alcohol synthesis, yielding 38.4 mg/L of 1-hexanol in
E. coli using glucose as the starting material.
ThrABC facilitates the production of threonine which is subsequently converted into 2-KB by
TdcB. G462D, S139G, and H97A mutations were introduced to LeuA which results in larger binding pockets and allows for chain elongation to occur by
LeuABCD. 2-KB is eventually elongated to form 2-ketoheptanoate. Subsequently, ADH VI from
S. cerevisiae and KivD from
L. lactis with V461A and F381L mutations converts 2-ketoheptanoate to 1-hexanol [
59].
Dekishima et al. expanded a CoA-dependent 1-butanol pathway that converts glucose into 1-hexanol in 9 steps in
E. coli. The 1-butanol pathway involves the expression of AtoB, BtkB, HBD, CRT, TER, and ADH from various organisms. BtkB from
Ralstonia eutropha has the ability to add 2 carbons onto the chain of acetoacetyl-CoA which provides the base for 1-hexanol production. The titer can be raised by overexpressing formate dehydrogenase from
C. boidinii, which increases the availability of NADH, a cofactor required for reducing enzymes. This method produced 47 mg/L of 1-hexanol after 48 h [
47]. Similarly, another strategy utilizes
Clostridium saccharoperbutylacetonicum due to its ability to convert sugars into acids, such as acetate. THL, HBD, CRT, and BCDH with electron-transferring flavoprotein A and B were all overexpressed to enable the elongation of acetyl-CoA, yielding 5.2 mM of 1-hexanol [
48]. Following a similar route, Lauer et al. engineered
Clostridium ljungdahlii since it can utilize gaseous substrates such as CO
2 and H
2 through the Wood-Ljungdahl pathway. The
C. kluyveri genome encodes at least 2 versions of THL, CRT, HBD, and BCHD. Both were grouped into separate operons:
thlA1,
crt1,
bcd1,
etfB1,
etfA1, and
hbd1, as well as
thlA2,
crt2,
bcd2,
etfB2,
etfA2, and
hbd2, which were subsequently integrated into the genome. Additionally,
adhE2 (encoding a bifunctional aldehyde-alcohol dehydrogenase) from
C. acetobutylicum were included to complete the biosynthesis of 1-hexanol. To address the low abundance of enzymes observed,
hbd,
thl,
crt,
bcd,
etfB,
etfA, and
hbd from
C. carboxidivorans were further integrated, producing 393 mg/L of 1-hexanol under fermentation conditions [
49].
Diender et al. explored the usage of a synthetic co-culture of
Clostridium autoethanogenum and
C. kluyveri fueled by syngas.
C. autoethanogenum can convert syngas into ethanol and acetate, which serve as main substrates while
C. kluyveri utilizes its native reverse β-oxidation metabolism for chain elongation. Together, these microbes are capable of producing 2.0 mmol/L/day of 1-hexanol [
50]. Oh et al. optimized the syngas fermentation conditions for the production of 1-hexanol using
C. carboxidivorans P7, attaining a production 2.34 g/L of hexanol [
51].
Hashem et al. reported the production of 8 mM of 1-hexanol in
Komagataella phaffii using linoleic acid as the substrate. Inspired by the synthesis of green leaf volatiles in plants, a lipoxygenase and hydroperoxide lyase from
Pleurotus sapidus and
Medicago truncatula, respectively, was expressed to produce hexanal. Subsequently, an endogenous ADH/aldo-keto reductase converts hexanal to 1-hexanol [
52].
3.7. ε-Caprolactone, 6-Hydroxyhexanoic Acid, and 1,6-Hexanediol
CL, HHA, and HDO hold applications in the production of important polymers such as polycaprolactone and nylon 66. Currently, the microbial synthesis of CL, HHA, and HDO mainly relies on the bioconversion of cyclohexane and cyclohexanol, which are nonrenewable substrates derived from petroleum [
56].
Mallin et al. devised a method to produce 600 mg/L of CL from cyclohexanol in
E. coli. A tailored PDH with a substrate loop from
Rhodobacter sphaeroides and the thermostable scaffold of PDH from
Deinococcus geothermalis converts cyclohexanol into cyclohexanone. CHMO from A. calcoaceticus subsequently produces CL from cyclohexanone. Initially, the PDH mutant suffered from poor expression rates. To remedy this, chaperones were co-expressed and found to increase the expression of PDH 4.9-fold. To increase conversion rates, the redesigned PDH and CHMO were co-immobilized on RelizymeTM HA403 to promote enzymatic stability. While immobilization offered certain advantages in terms of enzyme stability, it resulted in lower conversion rates, achieving only 34% efficiency compared to the 83% conversion rate observed when utilizing free enzymes [
53].
Srinivasamurthy et al. reported the production of HHA in
E. coli utilizing cyclohexanol as the feedstock. ADH and CHMO were overexpressed simultaneously allowing for the recycling of cofactors NADP+/NADPH. ADH catalyzes the conversion of cyclohexanol to cyclohexanone, followed by the conversion of cyclohexanone to CL by CHMO. Product inhibition of CHMO caused by the formation of CL was addressed by the use of lipase CAL-B to hydrolyze CL, leading to the production of 20 g/L of HHA under fed-batch fermentation conditions [
54]. Bretschneider et al. designed a cyclohexane-based HHA biosynthesis pathway in
P. taiwanensis. CPMO, CHD, CHMO, and Lac were overexpressed to produce 3.3 g/L of HHA in a bioreactor [
55]. Following a similar strategy, Salamanca et al. utilized
Acidovorax sp. CHX100, which inherently possesses a cyclohexane degradation pathway. To promote the accumulation of HHA, the gene encoding 6-hydroxyhexanoate dehydrogenase was deleted, resulting in the synthesis of 4.92 mM of HHA in 6 h [
56].
Zhang et al. engineered a 3-module system to synthesize 1,6-HDO in
E. coli using cyclohexane as the feedstock. Each module was expressed in a separate strain to facilitate redox self-sufficiency. Module 1 converts cyclohexane into cyclohexanol via CPMO. Module 2 utilizes ADH, Baeyer-Villiger monooxygenase, and Lac to synthesize cyclohexanone from cyclohexanol. Module 3 completes the synthesis of 1,6-HDO through the usage of CAR, aldo-keto reductase, and glucose dehydrogenase. 13 mM of 1,6-HDO was produced from 30 mM substrate concentration without the accumulation of intermediates [
57].
4. Discussion and Conclusions
C5 and C6 molecules are important and valuable chemicals for the polymer, pharmaceutical, and biofuel industries. As the demand for these compounds continues to rise, research efforts have been dedicated to developing methods for the efficient and low-cost production of these compounds. However, current chemical synthesis strategies lead to adverse environmental repercussions such as the emission of greenhouse gases and pollution, promoting expanded research into more sustainable methods.
Bio-based routes that utilize microbial production of value-added chemicals have great potential to address concerns of climate change and petroleum depletion. This is because these microorganisms can be engineered to convert renewable feedstock, such as glucose, glycerol, xylose, lignin, and syngas into desired compounds. These modifications include: (a) expanding or combining natural metabolic pathways; (b) design and construction of non-natural/artificial pathways; (c) deleting competing pathways to redirect metabolic flux; (d) modifying enzymes to improve substrate scope and/or catalytic efficiency; (e) cultivating co-cultures to reduce the metabolic burden of overexpressing multiple genes; (f) employing synthetic biology tools to optimize native and artificial pathways; (g) optimization of the fermentation process, through combinational approaches.
While the microbial production of C3 and C4 compounds benefits from the abundance of C2-C5 metabolites inherently produced in host cells, the biosynthesis of C5 and C6 chemicals poses challenges due to the lack of natural C5-C7 precursors. Creative solutions to this issue involve chain extension, condensation, and ring fission reactions catalyzed by various enzymes. Chain extension reactions are employed to elongate the carbon backbones of molecules, which expands the range of products that can be biosynthesized from shorter-chain compounds. Condensation reactions involve the combination of two separate molecules into a single molecule, which provides a viable alternative for utilizing short chain chemicals to produce longer carbon backbones. In ring fission reactions, the cyclic structures of aromatic compounds are linearized to serve as carbon-chain backbones for C5 and C6 chemicals. These strategies illustrate the versatility of metabolic engineering approaches in manipulating microbial pathways to produce C5 and C6 compounds from more readily available starting materials, overcoming the limitations imposed by the scarcity of suitable native precursors.
To achieve industrially viable yields, several major factors should be taken into consideration. Certain microbial species may not have the ability to produce functional enzymes from another species. Therefore, selection of a suitable chassis microorganisms is a prerequisite for efficient biosynthesis. Additionally, many pathway enzymes require cofactors and/or coenzymes, which may become scarce when multiple enzymes are co-expressed, leading to reduced reaction rates. To address this limitation, construction of co-enzyme regeneration mechanisms is critical for making full use of the enzyme activity. Moreover, enzymes exhibiting suboptimal catalytic rates can create bottlenecks in the pathway, which limits potential yield. Therefore, selecting well-suited enzymes is essential for the optimization of metabolic pathways. Alternatively, enzymes can be engineered by modifying critical amino acid residues to better accommodate non-native substrates and enhance pathway performance. On the other hand, excessive expression of numerous pathway genes can increase the metabolic burden on the host cells, which may decrease the titer of the desired chemical. Optimization of the enzyme expression level with static or dynamic regulation has become a feasible solution to this issue. Lastly, microorganisms may have different tolerances towards toxic intermediates and products, which require the creation of more resistant microbial chassis through rational genetic modification or directed evolution approaches.
Semi-synthetic routes that utilize a microbial host for a portion of the production steps offer a compelling middle ground between chemical and fully bio-based strategies. Notably, the completely bio-based production of MVL has low yields that can be remedied by finishing the conversion of the intermediate, mevalonate, to MVL through chemical methods [
46]. While these methods are not entirely renewable, they can address the disadvantages of a microbial host, increase yields of the desired compound, and utilize renewable feedstock.
Although certain strategies for producing C5 and C6 chemicals have shown promising titers and yields viable for industrial production, many still require further optimization before they become suitable for commercialization. Nonetheless, these routes promote the development of recycling practices, as these green pathways frequently utilize renewable carbon sources or byproducts from the manufacturing of other products. As the field of synthetic biology continues to advance and refine its techniques, the prospects for renewable and sustainable production of conventional petroleum-derived chemicals may improve significantly, potentially becoming mainstream in the global market.
Acknowledgements
This work is supported by the Biotechnology Program, Department of Engineering Technology, Division of Technology, Cullen College of Engineering, University of Houston.
Author Contributions
A.T. and V.N. drafted the manuscript; A.T., V.N., and Y.L. revised the manuscript; All authors reviewed and approved the manuscript.
Funding
This work was funded by Dr. Lin’s start-up funds from the Department of Engineering Technology, Division of Technology, Cullen College of Engineering, University of Houston, Houston, Texas, United States
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Nakamura CE, Whited GM. Metabolic engineering for the microbial production of 1,3-propanediol.
Curr. Opin. Biotechnol. 2003,
14, 454–459.
[Google Scholar]
-
2.
Harmsen PFH, Hackmann MM, Bos HL. Green building blocks for bio-based plastics.
Biofuels Bioprod. Biorefin. 2014,
8, 306–324.
[Google Scholar]
-
3.
Eriksen D, Li S, Zhao H. Chapter 3—Pathway Engineering as an Enabling Synthetic Biology Tool. In Synthetic Biology; Elsevier Inc: Amsterdam, The Netherlands, 2013; pp. 43–61.
-
4.
Laffend L, Haynie S, Emptage M, Pucci J, Whited G. Process for the biological production of 1,3-propanediol with high titer. US Patent US7504250B2.5, 2019.
-
5.
Burgard A, Burk MJ, Osterhout R, Van Dien S, Yim H. Development of a commercial scale process for production of 1,4-butanediol from sugar.
Curr. Opin. Biotechnol. 2016,
42, 118–125.
[Google Scholar]
-
6.
-
7.
Główka M, Krawczyk T. New Trends and Perspectives in Production of 1,2-Propanediol.
ACS Sustain. Chem. Eng. 2023,
11, 7274–7287.
[Google Scholar]
-
8.
Nawab S, Wang N, Ma X, Huo Y. Genetic engineering of non-native hosts for 1-butanol production and its challenges: A review.
Microbial Cell Fact. 2020,
19, 79.
[Google Scholar]
-
9.
Hakizimana O, Matabaro E, Lee BH. The current strategies and parameters for the enhanced microbial production of 2,3-butanediol.
Biotechnol. Rep. 2020,
25, e00397.
[Google Scholar]
-
10.
Hu S, Gao Q, Wang X, Yang J, Xu N, Chen K, et al. Efficient production of D-1,2,4-butanetriol from D-xylose by engineered
Escherichia coli whole-cell biocatalysts.
Front. Chem. Sci. Eng. 2018,
12, 772–779.
[Google Scholar]
-
11.
Huang Y, Ji X, Ma Z, Łężyk M, Xue Y, Zhao H. Green chemical and biological synthesis of cadaverine: recent development and challenges.
RSC Adv. 2021,
11, 23922–23942.
[Google Scholar]
-
12.
-
13.
Thaore V, Chadwick D, Shah N. Sustainable production of chemical intermediates for nylon manufacture: A techno-economic analysis for renewable production of caprolactone.
Chem. Eng. Res. Des. 2018,
135, 140–152.
[Google Scholar]
-
14.
Hermann BG, Blok K, Patel MK. Producing Bio-Based Bulk Chemicals Using Industrial Biotechnology Saves Energy and Combats Climate Change.
Environ. Sci. Technol. 2007,
41, 7915–7921.
[Google Scholar]
-
15.
Kapar Yilmaz E, Akbayrak A, Bayraç C. An Optimization Study for Laboratory Scale Production of Glucose Syrup from Potato, Wheat and Maize Starch.
Akademik Gıda 2021,
19, 364–372.
[Google Scholar]
-
16.
Lima PJM, da Silva RM, Neto CACG, Gomes e Silva NC, Souza JEdS, Nunes YL, et al. An overview on the conversion of glycerol to value‐added industrial products via chemical and biochemical routes.
Biotechnol. Appl. Biochem. 2022,
69, 2794–2818.
[Google Scholar]
-
17.
-
18.
-
19.
Chen GS, Siao SW, Shen CR. Saturated mutagenesis of ketoisovalerate decarboxylase V461 enabled specific synthesis of 1-pentanol via the ketoacid elongation cycle.
Sci. Rep. 2017,
7, 11284.
[Google Scholar]
-
20.
Cen X, Liu Y, Zhu F, Liu D, Chen Z. Metabolic Engineering of Escherichia coli for high production of 1,5-pentanediol via a cadaverine-derived pathway.
Metab. Eng. 2022,
74, 168–177.
[Google Scholar]
-
21.
Gao S, Zhang A, Ma D, Zhang K, Wang J, Wang X, et al. Enhancing pH stability of lysine decarboxylase via rational engineering and its application in cadaverine industrial production.
Biochem. Eng. J. 2022,
186, 108548.
[Google Scholar]
-
22.
Kwak DH, Lim HG, Yang J, Seo SW, Jung GY. Synthetic redesign of
Escherichia coli for cadaverine production from galactose.
Biotechnol. Biofuels 2017,
10, 20.
[Google Scholar]
-
23.
Kim HT, Baritugo K, Oh YH, Hyun SM, Khang TU, Kang KH, et al. Metabolic Engineering of Corynebacterium glutamicum for the High-Level Production of Cadaverine That Can Be Used for the Synthesis of Biopolyamide 510.
ACS Sus. Chem. Eng. 2018,
6, 5296–5305.
[Google Scholar]
-
24.
Buschke N, Schröder H, Wittmann C. Metabolic engineering of Corynebacterium glutamicum for production of 1,5-diaminopentane from hemicellulose.
Biotechnol. J. 2011,
6, 306–317.
[Google Scholar]
-
25.
Buschke N, Becker J, Schäfer R, Kiefer P, Biedendieck R, Wittmann C. Systems metabolic engineering of xylose-utilizing
Corynebacterium glutamicum for production of 1,5-diaminopentane.
Biotechnol. J. 2013,
8, 557–570.
[Google Scholar]
-
26.
Cheng J, Tu W, Luo Z, Liang L, Gou X, Wang X, et al. Coproduction of 5-Aminovalerate and δ-Valerolactam for the Synthesis of Nylon 5 From L-Lysine in
Escherichia coli.
Front. Bioeng. Biotechnol. 2021,
9, 726126.
[Google Scholar]
-
27.
Li W, Ma L, Shen X, Wang J, Feng Q, Liu L, et al. Targeting metabolic driving and intermediate influx in lysine catabolism for high-level glutarate production.
Nat. Comm. 2019,
10, 3337.
[Google Scholar]
-
28.
Wang J, Gao C, Chen X, Liu L. Engineering the Cad pathway in
Escherichia coli to produce glutarate from l-lysine.
Appl. Microbiol. Biotechnol. 2021,
105, 3587–3599.
[Google Scholar]
-
29.
Wang J, Shen X, Lin Y, Chen Z, Yang Y, Yuan Q, et al. Investigation of the Synergetic Effect of Xylose Metabolic Pathways on the Production of Glutaric Acid.
ACS Syn. Biol. 2018,
7, 24–29.
[Google Scholar]
-
30.
Han T, Kim GB, Lee SY. Glutaric acid production by systems metabolic engineering of an L-lysine–overproducing
Corynebacterium glutamicum.
Proc. Natl. Acad. Sci. USA 2020,
117, 30328–30334.
[Google Scholar]
-
31.
Djurdjevic I, Zelder O, Buckel W. Production of Glutaconic Acid in a Recombinant
Escherichia coli Strain.
Appl. Environ. Microbiol. 2011,
77, 320–322.
[Google Scholar]
-
32.
Sohn YJ, Kang M, Baritugo K, Son J, Kang KH, Ryu M, et al. Fermentative High-Level Production of 5-Hydroxyvaleric Acid by Metabolically Engineered
Corynebacterium glutamicum.
ACS Sustain. Chem. Eng. 2021,
9, 2523–2533.
[Google Scholar]
-
33.
Chen C, Wang T, Ye P, Li N. Metabolic engineering of
Escherichia coli for the efficient production of 5-hydroxyvaleric acid.
Process Biochem. 2023,
130, 625–633.
[Google Scholar]
-
34.
Choi S, Seo S, Park S, Lee H, Song J, Kim J, et al. Cell Factory Design and Culture Process Optimization for Dehydroshikimate Biosynthesis in
Escherichia coli.
Front. Bioeng. Biotechnol. 2019,
7, 241.
[Google Scholar]
-
35.
Wang G, Øzmerih S, Guerreiro R, Meireles AC, Carolas A, Milne N, et al. Improvement of cis,cis-Muconic Acid Production in
Saccharomyces cerevisiae through Biosensor-Aided Genome Engineering.
ACS Syn. Biol. 2020,
9, 634–646.
[Google Scholar]
-
36.
Kohlstedt M, Starck S, Barton N, Stolzenberger J, Selzer M, Mehlmann K, et al. From lignin to nylon: Cascaded chemical and biochemical conversion using metabolically engineered
Pseudomonas putida.
Metab. Eng. 2018,
47, 279–293.
[Google Scholar]
-
37.
Becker J, Kuhl M, Kohlstedt M, Starck S, Wittmann C. Metabolic engineering of
Corynebacterium glutamicum for the production of cis, cis-muconic acid from lignin.
Microbial Cell Fact. 2018,
17, 115.
[Google Scholar]
-
38.
Barton N, Horbal L, Starck S, Kohlstedt M, Luzhetskyy A, Wittmann C. Enabling the valorization of guaiacol-based lignin: Integrated chemical and biochemical production of cis,cis-muconic acid using metabolically engineered
Amycolatopsis sp ATCC 39116.
Metab. Eng. 2018,
45, 200–210.
[Google Scholar]
-
39.
Zhao M, Huang D, Zhang X, Koffas MAG, Zhou J, Deng Y. Metabolic engineering of
Escherichia coli for producing adipic acid through the reverse adipate-degradation pathway.
Metab. Eng. 2018,
47, 254–262.
[Google Scholar]
-
40.
Raj K, Partow S, Correia K, Khusnutdinova AN, Yakunin AF, Mahadevan R. Biocatalytic production of adipic acid from glucose using engineered
Saccharomyces cerevisiae.
Metab. Eng. Comm. 2018,
6, 28–32.
[Google Scholar]
-
41.
Niu W, Willett H, Mueller J, He X, Kramer L, Ma B, et al. Direct biosynthesis of adipic acid from lignin-derived aromatics using engineered
Pseudomonas putida KT2440.
Metab. Eng. 2020,
59, 151–161.
[Google Scholar]
-
42.
Bretschneider L, Heuschkel I, Bühler K, Karande R, Bühler B. Rational orthologous pathway and biochemical process engineering for adipic acid production using
Pseudomonas taiwanensis VLB120.
Metab. Eng. 2022,
70, 206–217.
[Google Scholar]
-
43.
Wang L, Li G, Li A, Deng Y. Directed Synthesis of Biobased 1,6-Diaminohexane from Adipic Acid by Rational Regulation of a Functional Enzyme Cascade in
Escherichia coli.
ACS Sustain. Chem. Eng. 2023,
11, 6011–6020.
[Google Scholar]
-
44.
Turk SCHJ, Kloosterman WP, Ninaber DK, Kolen KPAM, Knutova J, Suir E, et al. Metabolic Engineering toward Sustainable Production of Nylon6.
ACS Syn. Biol. 2016,
5, 65–73.
[Google Scholar]
-
45.
Trefzer AC, De Wildeman SMA, Schurmann M, Raemakers-Franken PC. Preparation of 6-aminocaproic acid from 5-formylvaleric acid. US Patent US20220064679A1.113, 2021.
-
46.
Xiong M, Schneiderman DK, Bates FS, Hillmyer MA, Zhang K. Scalable production of mechanically tunable block polymers from sugar.
Proc. Natl. Acad. Sci. USA 2014,
111, 8357–8362.
[Google Scholar]
-
47.
Dekishima Y, Lan EI, Shen CR, Cho KM, Liao JC. Extending Carbon Chain Length of 1-Butanol Pathway for 1-Hexanol Synthesis from Glucose by Engineered
Escherichia coli.
J. Am. Chem. Soc. 2011,
133, 11399–11401.
[Google Scholar]
-
48.
Wirth S, Dürre P. Investigation of putative genes for the production of medium-chained acids and alcohols in autotrophic acetogenic bacteria.
Metab. Eng. 2021,
66, 296–307296–307.
[Google Scholar]
-
49.
Lauer I, Philipps G, Jennewein S. Metabolic engineering of
Clostridium ljungdahlii for the production of hexanol and butanol from CO
2 and H
2.
Microbial Cell Fact. 2022,
21, 85.
[Google Scholar]
-
50.
Diender M, Stams AJM, Machado de Sousa DZ. Production of medium-chain fatty acids and higher alcohols by a synthetic co-culture grown on carbon monoxide or syngas.
Biotechnol. Biofuels 2016,
9, 82.
[Google Scholar]
-
51.
Oh HJ, Ko JK, Gong G, Lee S, Um Y. Production of Hexanol as the Main Product Through Syngas Fermentation by
Clostridium carboxidivorans P7.
Front. Bioeng. Biotechnol. 2022,
10, 850370.
[Google Scholar]
-
52.
Hashem C, Hochrinner J, Bürgler MB, Rinnofner C, Pichler H, Winkler M. From linoleic acid to hexanal and hexanol by whole cell catalysis with a lipoxygenase, hydroperoxide lyase and reductase cascade in
Komagataella phaffii.
Front. Mol. Biosci. 2022,
9, 965315.
[Google Scholar]
-
53.
Mallin H, Wulf H, Bornscheuer UT. A self-sufficient Baeyer–Villiger biocatalysis system for the synthesis of ɛ-caprolactone from cyclohexanol.
Enzyme Microbial Technol. 2013,
53, 283–287.
[Google Scholar]
-
54.
Srinivasamurthy VST, Böttcher D, Bornscheuer UT. A multi-enzyme cascade reaction for the production of 6-hydroxyhexanoic acid.
Zeitschrift für Naturforschung C. A J. Biosci. 2019,
74, 71–76.
[Google Scholar]
-
55.
Bretschneider L, Heuschkel I, Wegner M, Lindmeyer M, Bühler K, Karande R, et al. Conversion of Cyclohexane to 6-Hydroxyhexanoic Acid Using Recombinant Pseudomonas taiwanensis in a Stirred-Tank Bioreactor.
Front. Catal. 2021,
1, 683248.
[Google Scholar]
-
56.
Salamanca D, Bühler K, Engesser K, Schmid A, Karande R. Whole-cell biocatalysis using the
Acidovorax sp. CHX100 Δ6HX for the production of ω-hydroxycarboxylic acids from cycloalkanes.
New Biotechnol. 2021,
60, 200–206.
[Google Scholar]
-
57.
Zhang Z, Li Q, Wang F, Li R, Yu X, Kang L, et al. One-pot biosynthesis of 1,6-hexanediol from cyclohexane by de novo designed cascade biocatalysis.
Green Chem. 2020,
22, 7476–7483.
[Google Scholar]
-
58.
Cann AF, Liao JC. Pentanol isomer synthesis in engineered microorganisms.
Appl. Microbiol. Biotechnol. 2010,
85, 893–899.
[Google Scholar]
-
59.
Zhang K, Sawaya MR, Eisenberg DS, Liao JC. Expanding metabolism for biosynthesis of nonnatural alcohols.
Proc. Natl. Acad. Sci. USA 2008,
105, 20653–20658.
[Google Scholar]
-
60.
Cen X, Liu Y, Chen B, Liu D, Chen Z. Metabolic Engineering of
Escherichia coli for De Novo Production of 1,5-Pentanediol from Glucose.
ACS Syn. Biol. 2021,
10, 192–203.
[Google Scholar]
-
61.
Wang J, Li C, Zou Y, Yan Y. Bacterial synthesis of C3-C5 diols via extending amino acid catabolism.
Proc. Natl. Acad. Sci. USA 2020,
117, 19159–19167.
[Google Scholar]
-
62.
Hong EY, Lee S, Park BJ, Lee JM, Yun H, Kim B. Simultaneously Enhancing the Stability and Catalytic Activity of Multimeric Lysine Decarboxylase CadA by Engineering Interface Regions for Enzymatic Production of Cadaverine at High Concentration of Lysine.
Biotechnol. J. 2017,
12, 1700278.
[Google Scholar]
-
63.
Kou F, Zhao J, Liu J, Sun C, Guo Y, Tan Z, et al. Enhancement of the thermal and alkaline pH stability of
Escherichia coli lysine decarboxylase for efficient cadaverine production.
Biotechnol. Lett. 2018,
40, 719–727.
[Google Scholar]
-
64.
Xi Y, Ye L, Yu H. Enhanced thermal and alkaline stability of L-lysine decarboxylase CadA by combining directed evolution and computation-guided virtual screening.
Bioresour. Bioproc. 2022,
9, 1–15.
[Google Scholar]
-
65.
Ma W, Cao W, Zhang H, Chen K, Li Y, Ouyang P. Enhanced cadaverine production from L-lysine using recombinant
Escherichia coli co-overexpressing CadA and CadB.
Biotechnol. Lett. 2015,
37, 799–806.
[Google Scholar]
-
66.
Huang C, Ting W, Chen Y, Wu P, Dong C, Huang S, et al. Facilitating the enzymatic conversion of lysine to cadaverine in engineered
Escherichia coli with metabolic regulation by genes deletion.
Biochem. Eng. J. 2020,
156, 107514.
[Google Scholar]
-
67.
Moon Y, Yang SY, Choi TR, Jung H, Song H, Han YH, et al. Enhanced production of cadaverine by the addition of hexadecyltrimethylammonium bromide to whole cell system with regeneration of pyridoxal-5′-phosphate and ATP.
Enzyme Microbial Technol. 2019,
127, 58–64.
[Google Scholar]
-
68.
Xue C, Hsu K, Ting W, Huang S, Lin H, Li S, et al. Efficient biotransformation of l-lysine into cadaverine by strengthening pyridoxal 5’-phosphate-dependent proteins in
Escherichia coli with cold shock treatment.
Biochem. Eng. J. 2020,
161, 107659.
[Google Scholar]
-
69.
Wang C, Zhang K, Zhongjun C, Cai H, Honggui W, Ouyang P. Directed evolution and mutagenesis of lysine decarboxylase from
Hafnia alvei AS1.1009 to improve its activity toward efficient cadaverine production.
Biotechnol. Bioproc. E 2015,
20, 439–446.
[Google Scholar]
-
70.
Noh M, Yoo SM, Yang D, Lee SY. Broad-Spectrum Gene Repression Using Scaffold Engineering of Synthetic sRNAs.
ACS Syn. Biol. 2019,
8, 1452–1461.
[Google Scholar]
-
71.
Wei G, Zhang A, Lu X, He F, Li H, Xu S, et al. An environmentally friendly strategy for cadaverine bio-production: in situ utilization of CO
2 self-released from L-lysine decarboxylation for pH control.
J. CO2 Utiliz. 2020,
37, 278–284.
[Google Scholar]
-
72.
Kind S, Neubauer S, Becker J, Yamamoto M, Völkert M, Abendroth Gv. et al. From zero to hero—Production of bio-based nylon from renewable resources using engineered
Corynebacterium glutamicum.
Metab. Eng. 2014,
25, 113–123.
[Google Scholar]
-
73.
Li M, Li D, Huang Y, Liu M, Wang H, Tang Q, et al. Improving the secretion of cadaverine in
Corynebacterium glutamicum by cadaverine–lysine antiporter.
J. Ind. Microbiol. Biotechnol. 2014,
41, 701–709.
[Google Scholar]
-
74.
Kobayashi S, Kawaguchi H, Shirai T, Ninomiya K, Takahashi K, Kondo A, et al. Automatic Redirection of Carbon Flux between Glycolysis and Pentose Phosphate Pathway Using an Oxygen-Responsive Metabolic Switch in
Corynebacterium glutamicum.
ACS Syn. Biol. 2020,
9, 814–826.
[Google Scholar]
-
75.
Imao K, Konishi R, Kishida M, Hirata Y, Segawa S, Adachi N, et al. 1,5-Diaminopentane production from xylooligosaccharides using metabolically engineered
Corynebacterium glutamicum displaying beta-xylosidase on the cell surface.
Bioresour. Technol. 2017,
245, 1684–1691.
[Google Scholar]
-
76.
Xu Y, Zhou D, Luo R, Yang X, Wang B, Xiong X, et al. Metabolic engineering of
Escherichia coli for polyamides monomer δ-valerolactam production from feedstock lysine.
Appl. Microbiol. Biotechnol. 2020,
104, 9965–9977.
[Google Scholar]
-
77.
Park SJ, Kim EY, Noh W, Park HM, Oh YH, Lee SH, et al. Metabolic engineering of
Escherichia coli for the production of 5-aminovalerate and glutarate as C5 platform chemicals.
Metab. Eng. 2013,
16, 42–47.
[Google Scholar]
-
78.
Adkins J, Jordan J, Nielsen DR. Engineering
Escherichia coli for renewable production of the 5-carbon polyamide building-blocks 5-aminovalerate and glutarate.
Biotechnol. Bioeng. 2013,
110, 1726–1734.
[Google Scholar]
-
79.
Hong Y, Moon Y, Choi T, Jung H, Yang S, Ahn J, et al. Enhanced production of glutaric acid by NADH oxidase and GabD‐reinforced bioconversion from l‐lysine.
Biotechnol. Bioeng. 2019,
116, 333–341.
[Google Scholar]
-
80.
Hong Y, Moon Y, Hong J, No S, Choi T, Jung H, et al. Production of glutaric acid from 5-aminovaleric acid using
Escherichia coli whole cell bio-catalyst overexpressing GabTD from
Bacillus subtilis.
Enzyme Microbial Technol. 2018,
118, 57–65.
[Google Scholar]
-
81.
Yang S, Choi T, Jung H, Park Y, Han Y, Song H, et al. Production of glutaric acid from 5-aminovaleric acid by robust whole-cell immobilized with polyvinyl alcohol and polyethylene glycol.
Enzyme Microbial Technol. 2019,
128, 72–78.
[Google Scholar]
-
82.
Yang S, Choi T, Jung H, Park Y, Han Y, Song H, et al. Development of glutaric acid production consortium system with α-ketoglutaric acid regeneration by glutamate oxidase in
Escherichia coli.
Enzyme Microbial Technol. 2020,
133, 109446.
[Google Scholar]
-
83.
Wang X, Su R, Chen K, Xu S, Feng J, Ouyang P. Engineering a Microbial Consortium Based Whole-Cell System for Efficient Production of Glutarate From L-Lysine.
Front. Microbiol. 2019,
10, 341.
[Google Scholar]
-
84.
Rohles CM, Gießelmann G, Kohlstedt M, Wittmann C, Becker J. Systems metabolic engineering of
Corynebacterium glutamicum for the production of the carbon-5 platform chemicals 5-aminovalerate and glutarate.
Microbial Cell Fact. 2016,
15, 154.
[Google Scholar]
-
85.
Rohles CM, Gläser L, Kohlstedt M, Gießelmann G, Pearson S, del Campo A, et al. A bio-based route to the carbon-5 chemical glutaric acid and to bionylon-6,5 using metabolically engineered
Corynebacterium glutamicum.
Green Chem. 2018,
20, 4662–4674.
[Google Scholar]
-
86.
Kim HT, Khang TU, Baritugo K, Hyun SM, Kang KH, Jung SH, et al. Metabolic engineering of
Corynebacterium glutamicum for the production of glutaric acid, a C5 dicarboxylic acid platform chemical.
Metab. Eng. 2019,
51, 99–109.
[Google Scholar]
-
87.
Pérez-García F, Jorge JMP, Dreyszas A, Risse JM, Wendisch VF. Efficient Production of the Dicarboxylic Acid Glutarate by
Corynebacterium glutamicum via a Novel Synthetic Pathway.
Front. Microbiol. 2018,
9, 2589.
[Google Scholar]
-
88.
Yu J, Xia X, Zhong J, Qian Z. A novel synthetic pathway for glutarate production in recombinant
Escherichia coli.
Proc. Biochem. 2017,
59, 167–171.
[Google Scholar]
-
89.
Chen C, Chen X, Liu L, Wu J, Gao C. Engineering Microorganisms to Produce Bio-Based Monomers: Progress and Challenges.
Fermentation 2023,
9, 137.
[Google Scholar]
-
90.
Wang J, Wu Y, Sun X, Yuan Q, Yan Y. De Novo Biosynthesis of Glutarate via αKeto Acid Carbon Chain Extension and Decarboxylation Pathway in
Escherichia coli.
ACS Syn. Biol. 2017,
6, 1922–1930.
[Google Scholar]
-
91.
Zhao M, Li G, Deng Y. Engineering
Escherichia coli for Glutarate Production as the C5 Platform Backbone.
Appl. Environ. Microbiol. 2018,
84, e00814-18.
[Google Scholar]
-
92.
Sui X, Zhao M, Liu Y, Wang J, Li G, Zhang X, et al. Enhancing glutaric acid production in
Escherichia coli by uptake of malonic acid.
J. Ind. Microbiol. BioTechnol. 2020,
47, 311–318.
[Google Scholar]
-
93.
Sun J, Ren Y, Raza M, Sun X, Yuan Q. Microbial production of glutaconic acid via extradiol ring cleavage of catechol.
J. Chem. Technol. BioTechnol. 2018,
93, 1677–1683.
[Google Scholar]
-
94.
Khalil I, Quintens G, Junkers T, Dusselier M. Muconic acid isomers as platform chemicals and monomers in the biobased economy.
Green Chem. 2020,
22, 1517–1541.
[Google Scholar]
-
95.
Choi S, Lee H, Park E, Lee S, Kim E. Recent Advances in Microbial Production of cis,cis-Muconic Acid.
Biomolecules 2020,
10, 1238.
[Google Scholar]
-
96.
Draths KM, Frost JW. Environmentally compatible synthesis of adipic acid from D-glucose.
J. Am. Chem. Soc. 1994,
116, 399–400.
[Google Scholar]
-
97.
Niu W, Draths KM, Frost JW. Benzene-Free Synthesis of Adipic Acid.
BioTechnol. Progr. 2002,
18, 201–211.
[Google Scholar]
-
98.
Curran KA, Leavitt JM, Karim AS, Alper HS. Metabolic engineering of muconic acid production in
Saccharomyces cerevisiae.
Metab. Eng. 2013,
15, 55–66.
[Google Scholar]
-
99.
Suastegui M, Matthiesen JE, Carraher JM, Hernandez N, Rodriguez Quiroz N, Okerlund A, et al. Titelbild: Combining Metabolic Engineering and Electrocatalysis: Application to the Production of Polyamides from Sugar.
Angewandte Chem. 2016,
128, 2317.
[Google Scholar]
-
100.
Leavitt JM, Wagner JM, Tu CC, Tong A, Liu Y, Alper HS. Biosensor‐Enabled Directed Evolution to Improve Muconic Acid Production in
Saccharomyces cerevisiae.
BioTechnol. J. 2017,
12, 1600687.
[Google Scholar]
-
101.
Suástegui M, Yu Ng C, Chowdhury A, Sun W, Cao M, House E, et al. Multilevel engineering of the upstream module of aromatic amino acid biosynthesis in
Saccharomyces cerevisiae for high production of polymer and drug precursors.
Metab. Eng. 2017,
42, 134–144.
[Google Scholar]
-
102.
Liu T, Peng B, Huang S, Geng A. Recombinant xylose-fermenting yeast construction for the co-production of ethanol and cis,cis-muconic acid from lignocellulosic biomass.
Bioresour. Technol. Rep. 2020,
9, 100395.
[Google Scholar]
-
103.
Fujiwara R, Noda S, Tanaka T, Kondo A. Metabolic engineering of
Escherichia coli for shikimate pathway derivative production from glucose–xylose co-substrate.
Nat. Comm. 2020,
11, 279.
[Google Scholar]
-
104.
Sengupta S, Jonnalagadda S, Goonewardena L, Juturu V. Metabolic Engineering of a Novel Muconic Acid Biosynthesis Pathway via 4-Hydroxybenzoic Acid in
Escherichia coli.
Appl. Environ. Microbiol. 2015,
81, 8037–8043.
[Google Scholar]
-
105.
Sun X, Lin Y, Huang Q, Yuan Q, Yan Y. A Novel Muconic Acid Biosynthesis Approach by Shunting Tryptophan Biosynthesis via Anthranilate.
Appl. Environ. Microbiol. 2013,
79, 4024–4030.
[Google Scholar]
-
106.
Sun X, Lin Y, Yuan Q, Yan Y. Biological Production of Muconic Acid via a Prokaryotic 2,3-Dihydroxybenzoic Acid Decarboxylase.
Chem. Sus. Chem. 2014,
7, 2478–2481.
[Google Scholar]
-
107.
Wang J, Zheng P. Muconic acid production from glucose using enterobactin precursors in
Escherichia coli.
J. Ind. Microbiol. BioTechnol. 2015,
42, 701–709.
[Google Scholar]
-
108.
Zhang H, Li Z, Pereira B, Stephanopoulos G. Engineering
E. coli–
E. coli cocultures for production of muconic acid from glycerol.
Microbial Cell Fact. 2015,
14, 134.
[Google Scholar]
-
109.
Zhang H, Pereira B, Li Z, Stephanopoulos G. Engineering
Escherichia coli coculture systems for the production of biochemical products.
Proc. Natl. Acad. Sci. USA 2015,
112, 8266–8271.
[Google Scholar]
-
110.
Lin Y, Sun X, Yuan Q, Yan Y. Extending shikimate pathway for the production of muconic acid and its precursor salicylic acid in
Escherichia coli.
Metab. Eng. 2014,
23, 62–69.
[Google Scholar]
-
111.
Fujiwara R, Noda S, Tanaka T, Kondo A. Muconic Acid Production Using Gene-Level Fusion Proteins in
Escherichia coli.
ACS Syn. Biol. 2018,
7, 2698–2705.
[Google Scholar]
-
112.
Vardon D, Franden M, Johnson C, Karp E, Guarnieri M, Linger J, et al. Adipic acid production from lignin.
Energy Environ. Sci. 2015,
8, 617–628.
[Google Scholar]
-
113.
Johnson CW, Salvachúa D, Khanna P, Smith H, Peterson DJ, Beckham GT. Enhancing muconic acid production from glucose and lignin-derived aromatic compounds via increased protocatechuate decarboxylase activity.
Metab. Eng. Comm. 2016,
3, 111–119.
[Google Scholar]
-
114.
Johnson CW, Abraham PE, Linger JG, Khanna P, Hettich RL, Beckham GT. Eliminating a global regulator of carbon catabolite repression enhances the conversion of aromatic lignin monomers to muconate in
Pseudomonas putida KT2440.
Metab. Eng. Comm. 2017,
5, 19–25.
[Google Scholar]
-
115.
Lee H, Shin W, Seo S, Choi S, Song J, Kim J, et al. Corynebacterium Cell Factory Design and Culture Process Optimization for Muconic Acid Biosynthesis.
Sci. Rep. 2018,
8, 18041.
[Google Scholar]
-
116.
Shin W, Lee D, Lee SJ, Chun G, Choi S, Kim E, et al. Characterization of a non-phosphotransferase system for cis,cis-muconic acid production in
Corynebacterium glutamicum.
Biochem. Biophys. Res. Comm. 2018,
499, 279–284.
[Google Scholar]
-
117.
Polen T, Spelberg M, Bott M. Toward biotechnological production of adipic acid and precursors from biorenewables.
J. Biotechnol. 2013,
167, 75–84.
[Google Scholar]
-
118.
Cheng J, Hu G, Xu Y, Torrens-Spence MP, Zhou X, Wang D, et al. Production of nonnatural straight-chain amino acid 6-aminocaproate via an artificial iterative carbon-chain-extension cycle.
Metab. Eng. 2019,
55, 23–32.
[Google Scholar]
-
119.
Joo JC, Khusnutdinova AN, Flick R, Kim T, Bornscheuer UT, Yakunin AF, et al. Alkene hydrogenation activity of enoate reductases for an environmentally benign biosynthesis of adipic acid.
Chem. Sci. 2017,
8, 1406–1413.
[Google Scholar]
-
120.
Zhang Q, Xu G, Pizzi A, Lei H, Xi X, Du G. A Green Resin Wood Adhesive from Synthetic Polyamide Crosslinking with Glyoxal.
Polymers 2022,
14, 2819.
[Google Scholar]
-
121.
Lau MK. Methods and microorganisms for the biological synthesis of (S)-2-amino-6-hydroxypimelate, hexamethylenediamine and 6-aminocaproate. US Patent US20150203879A1, 2013.
-
122.
Botes AL, Conradie AE. Methods of producing 6-carbon chemicals via CoA-dependent carbon chain elongation associated with carbon storage. US Patent US9102958B2, 2012.
-
123.
Markowska A, Markowski AR, Jarocka-Karpowicz I. The Importance of 6-Aminohexanoic Acid as a Hydrophobic, Flexible Structural Element.
Int. J. Mol. Sci. 2021,
22, 12122.
[Google Scholar]
-
124.
Bretschneider L, Wegner M, Bühler K, Bühler B, Karande R. One‐pot synthesis of 6‐aminohexanoic acid from cyclohexane using mixed‐species cultures.
Microbial Biotechnol. 2021,
14, 1011–1025.
[Google Scholar]
-
125.
Guptill DM, Brutman JP, Hoye TR. Thermoplastic polyurethanes from β-methyl-δ-valerolactone-derived amidodiol chain extenders.
Polymer 2017,
111, 252–257.
[Google Scholar]
-
126.
Schneiderman DK, Vanderlaan ME, Mannion AM, Panthani TR, Batiste DC, Wang JZ, et al. Chemically Recyclable Biobased Polyurethanes.
ACS Macro Lett. 2016,
5, 515–518.
[Google Scholar]
-
127.
De Poures MV, Sathiyagnanam AP, Rana D, Rajesh Kumar B, Saravanan S. 1-Hexanol as a sustainable biofuel in DI diesel engines and its effect on combustion and emissions under the influence of injection timing and exhaust gas recirculation (EGR).
Appl. Thermal Eng. 2017,
113, 1505–1513.
[Google Scholar]
-
128.
Harvey B, Meylemans H. 1-Hexene: a renewable C6 platform for full-performance jet and diesel fuels.
Green Chem. 2014,
16, 770–776.
[Google Scholar]
-
129.
Falbe J, Bahrmann H, Lipps W, Mayer D, Frey GD. Alcohols, Aliphatic.
Ullmann's Encycl. Ind. Chem. 2003,
2, 19–46.
[Google Scholar]