1. Introduction
Although ethanol is currently blended with commercial gasoline in many countries worldwide and is produced from various renewable resources, intrinsic physiochemical properties make n-butanol (butanol hereafter) more suitable than ethanol for use in motor engines due to its energy content (which is comparable to gasoline), lower corrosiveness, and safer management [
1]. As a result, butanol is one of the most promising alternatives to ethanol as a fuel blendstock for transportation. Biobutanol could be produced from renewable lignocellulosic biomass resources through the well-known acetone-butanol-ethanol (ABE) fermentation. However, the renewable biobutanol production process still faces various challenges. In particular, the production of butanol and other biochemicals is limited by microbe intolerance to butanol itself and the combined effects with other endproducts. In addition, tolerance to by-products of the biomass hydrolysis process is especially significant for biorefinery purposes when lignocellulosic biomass is used as the feedstock. Recent developments in genetic tools have enabled the engineering of some Clostridium species to mitigate some of the limitations and improve butanol production.
This study provides a comprehensive perspective on strain tolerance related to general chemical stresses, considering the oxygen stress (butanol-producing clostridia are strictly anaerobic), fermentation endproducts, as well as the inhibitors from lignocellulosic biomass processing, with a special focus on butanol production and improvement through metabolic engineering approaches. Unlike other studies that focus on the single inhibition effect of inhibitors [
1,
2,
3,
4,
5], we classify inhibition mechanisms according to specific types of cell stress, such as self-product inhibition, oxidative stress, and other chemical stresses. We also summarize a state-of-the-art and our own perspective on recent advances in the chemical tolerance of Clostridium strains and other microorganisms for biomanufacturing broadly.
2. Types of Inhibitors
2.1. Fatty Acids as Inhibitors
depicts the primary metabolic pathways for ABE production in solventogenic clostridia. During the first stage of fermentation, namely acidogenesis, the sugars are quickly converted into acetic and butyric acids with rapid cell growth. High concentrations of acids can lead to an early stop of a fermentation if uncontrolled fermentations reach pH lower than ~5. This phenomenon is called “acid crash” [
6]. Although fatty acids (acetate and butyrate) are intermediates during ABE fermentation which are usually re-assimilated for solvent production in the late stage [
7,
8], the high concentrations of acids are highly inhibitory and lead to cell death. If the cells can accomplish metabolic switch from acetogenesis to solventogenesis successfully before pH reaches too low, or an appropriate pH control strategy is implemented, “acid crash” can usually be avoided.
Acetate production and tolerance has been extensively studied in acetogenic clostridia, such as
C. ljungdahlii,
C. autoethanogenum,
C. ragsdalei,
C. coskatii [
9],
C. thermoaceticum,
C. thermoautotrophicum [
7,
10],
C. drakei,
C. scatologenes, &
C. carboxidivorans [
11]. Acetogenic
Clostridium can convert syngas (CO, CO
2/H
2) into acetate (and/or ethanol) through the Wood–Ljungdahl pathway, which has attracted tremendous attention recently [
9]. Although those species are not solventogenic, they closely resemble and share some metabolic pathways with solventogenic clostridia. For example, many molecular chaperones (or heat and chemical shock proteins) are upregulated in the acidogenic
Clostridium AWRP [
12]. In solventogenic clostridial fermentation, such as with
Clostridium acetobutylicum ATCC 824, chaperone-like genes such as
phaP, as well as others such as
pncB and
cfa demonstrated elevated expression levels in the presence of acetic and other inhibitors [
13]. Endogenous acetate and butyrate are usually not significant inhibitors, apart from their impart on pH, due to their typically low concentration. However, acetate inhibition for fermentation could become noteworthy under certain conditions, such as when biomass is pretreated with acetic acid or when acetylated biomass undergoes hydrolysis, resulting in substantial amounts of exogenous acetate.
Solventogenic clostridia species produce other organic acids, like formate and lactate, but in minimal quantities and typically only during the initial phase of fermentation. Although present in small amounts, formate can add to the cumulative stress caused by acetic and butyric acids. It is believed that many enzymes involved in managing this stress likely use zinc as a cofactor. Wu et al. observed an increase in formic, acetic, and butyric acids tolerance after the addition of zinc in a fermentation with
C. acetobutylicum [
14], which resulted in a higher butyric acid peak production and a higher butanol titer.
2.2. Fermentation Endproducts as Inhibitors
Acetone production is almost inevitable during butanol production through the ABE pathway, because acetone is a coproduct for the acid reassimilation (). High acetone concentration can also inhibit cell growth and butanol production. Recently, rewiring the pathway for converting acetone into isopropanol by inclusion of a secondary alcohol dehydrogenase gene (
sadh) in the ABE strain has been implemented by various researchers [
8,
15,
16,
17]. This often resulted in slight reduction in the production of butanol and total solvents, due to the limited reducing power availability (one additional NADH is consumed for isopropanol production) and also the toxicity of isopropanol. The metabolically engineered
C. tyrobutyricum Δcat1::adhE2 for butanol production does not produce acetone, as it does not contain the
ctfAB-based acid re-assimilation pathway [
18]. Interestingly, recently, a new wild type isolate
Thermoanaerobacterium thermosaccharolyticum TG57 was reported, which is capable of converting microcrystalline cellulose directly to produce butanol without acetone as a coproduct [
19].
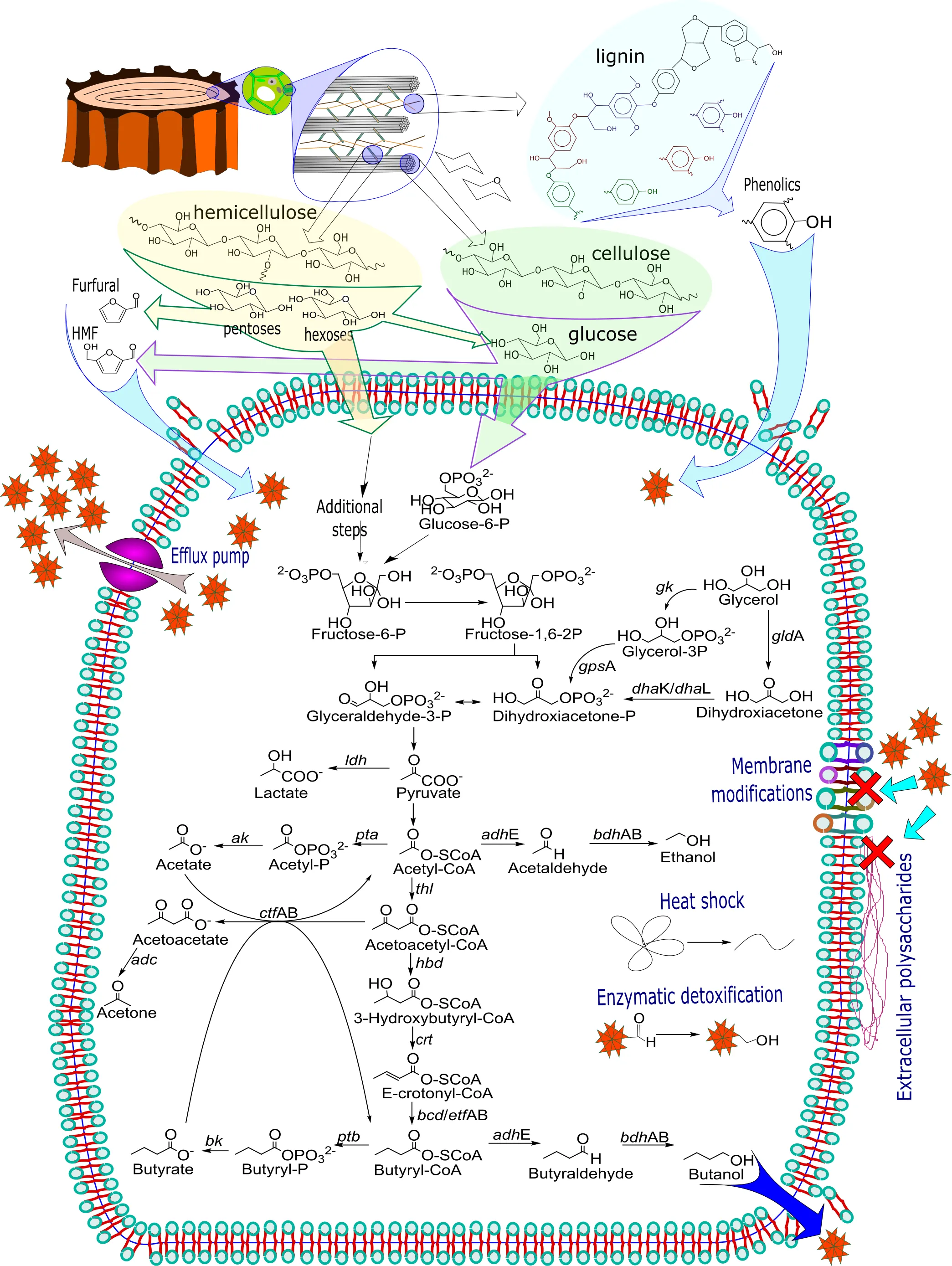
. Butanol production from lignocellulosic biomass through the acetone-butanol-ethanol (ABE) pathway and tolerance mechanisms. In the top of the figure, the lignocellulosic material transformation including formation on inhibitory by-products is described. ABE metabolic pathway is shown in the inner of the cell membrane, as well as the enzymatic tolerance mechanisms. Interchange of substrate products and inhibitors is described through the cell membrane, as well as the membrane-related tolerance mechanisms. Abbreviations: <em>gk</em> glycerol kinase, <em>gps</em>A glycerol-3-phosphate dehydrogenase, <em>gld</em>A glycerol dehydrogenase, <em>dha</em>K/<em>dha</em>L dihydroxyacetone kinase, <em>ldh</em> lactate dehydrogenase, <em>ak</em> acetate kinase, <em>pta</em> phosphotransacetylase, <em>adh</em>E acetyl dehydrogenase, <em>bdh</em>AB butyryl dehydrogenase, <em>thl</em> thiolase, <em>ctf</em>AB acetoacetyl-CoA:acetate/butyrate CoA, <em>adc</em> acetoacetate descaboxylase, <em>hbd</em> 3-hydroxybutyryl-CoA dehydrogenase, <em>crt</em> short-chain-enoyl-CoA hydratase, <em>bcd</em>/<em>etf</em>AB Electron Transferring Flavoprotein (<em>etf</em>) and butyryl-CoA dehydrogenase (<em>bcd</em>), <em>bk</em> butyrate kinase, <em>ptb</em> phosphotransbutyrylase.
Aliphatic alcohols, including butanol, isopropanol, and ethanol, exhibit similar interactions with microbial cells, although butanol is the most toxic among the three. The reduced toxicity of isopropanol in comparison to butanol is not solely attributable to its shorter aliphatic chain, but is also due to its branched and bulkier structure, which impedes its intercalation into the cell membrane components [
20]. Previous studies have reported enhanced isopropanol tolerance through chemical mutagenesis and selection [
21] or the isolation of tolerant strains [
22]. In contrast to butanol, ethanol is significantly less toxic, and its tolerance is not considered a key factor in ABE strains, although ethanol stress is crucial in ethanologenic strains which can produce ethanol to much higher levels. A metabolomic and proteomic analysis of ethanologenic
C. thermocellum has revealed that increasing ethanol concentration leads to a reduction in glutamic acid, accumulation of sugar phosphates, and inhibition of glycolysis [
23]. Evolutionary engineering has been used to develop tolerant strains of
C. thermocellum and
C. phytofermentans [
24,
25].
2.3. Inhibitors Derived from Biomass Processing
Cellulose is the most abundant polysaccharide found on earth and its natural function in plants is structural rather than energetic. However, lignocellulosic wastes are considered valuable feedstock due to their availability as byproducts of forestry and agricultural production, their low cost, and their lack of competition with food production.
Despite the abundance of cellulose, there is usually no natural butanol production pathway in cellulolytic strains, and overall their degradation of cellulose is not very efficient. Nevertheless, some cellulolytic strains have been genetically engineered to produce butanol and
iso-butanol. For example,
C. cellulovorans was engineered to produce 3.47 g/L butanol through the introduction of adhE1 for butanol production, and
ctfAB and
adc for acid reassimilation, and evolutionary engineering for tolerance enhancement [
26]. Additionally,
C. thermocellum and
C. cellulolyticum were engineered to produce 5.4 and 0.66 g/L iso-butanol, respectively, through the introduction of the hybrid keto acid pathway:
alsS/
ahaS,
kari,
dhaD,
kivD, and
adh [
27,
28]. Although these strains hold promise for consolidated bioprocessing for biobutanol production, they are currently not commercially competitive.
T. thermosaccharolyticum TG57 is the only natural cellulolytic butanologenic bacterium identified thus far, however, the maximum butanol production achieved was only 1.93 g/L [
19].
Alternatively, the lignocellulosic biomass could be first converted into sugars via physical-chemical pretreatment and enzymatic hydrolysis. Typically, the lignocellulosic material requires a pretreatment stage to remove lignin (which may involve the utilization of acid, alkali, oxidant, solvent and/or ionic liquids) and make the cellulose accessible to chemicals or enzymes. [
29,
30].
The acid-mediated process is the most extensively studied and successful pretreatment method thus far. The acid process is optimized to create mild conditions during the preparation of material or hydrolysis of glycosidic bonds in hydrolysate, although it generates some by-products. Even small concentrations of these compounds can have a negative impact on the growth of solventogenic
Clostridium, which reduces production efficiency [
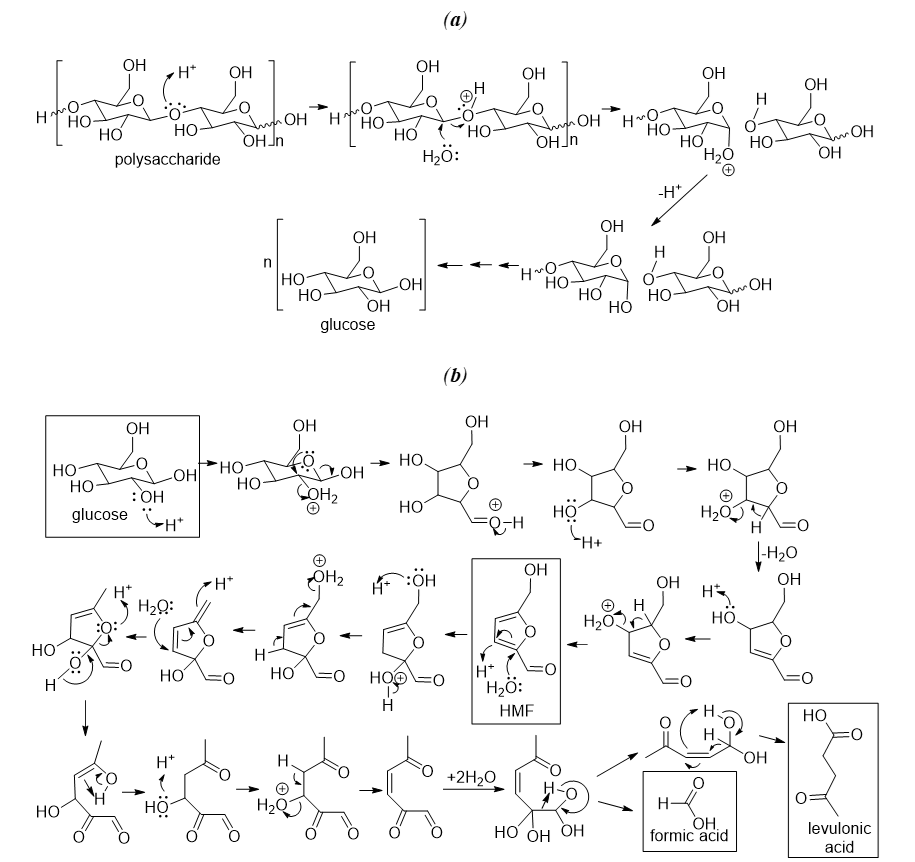
31]. (a) illustrates the hydrolysis mechanism of polysaccharides into simple sugars. (b) explains the conversion mechanism of glucose into hydroxymethylfurfural (HMF) and then into formic acid and levulinic acid. Pentoses can also be converted into furfural, similar to the first step shown in b.
Furan aldehydes, such as furfural and HMF, react with amines, including DNA bases, due to their aldehyde group. The presence of allopurinol has been shown to increase tolerance to furan aldehydes in
C. beijerinckii NCIMB 8052 [
32] indicating that increased purine salvage and DNA repairing may be the key mechanism. Furan aldehyde tolerance in C
. beijerinckii BA101 and
C. saccharoperbutylacetonicum N1–4 has been reported to be approximately 3 g/L [
33,
34].
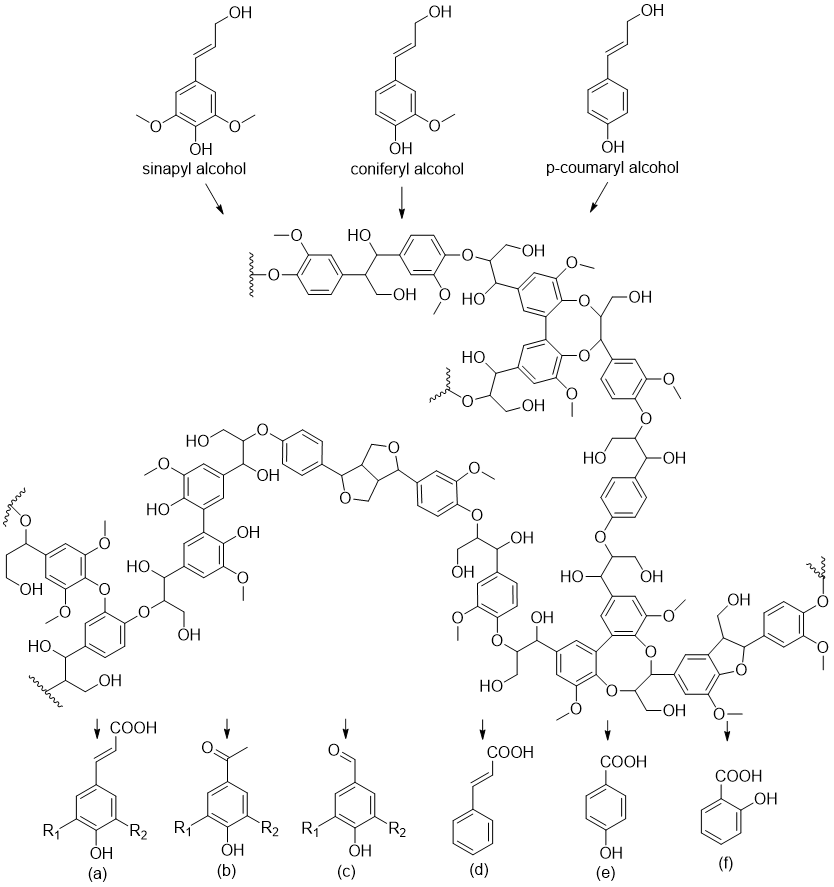
Lignin is a complex polyphenolic polymer that is crosslinked with structural polysaccharides. It is synthesized from sinapyl, coniferyl, and p-coumaryl alcohols. depicts the precursors, a fragment of the polymeric structure, and some water-soluble byproducts of acid or alkali pretreatment of biomass. Some of the compounds generated during pretreatment include
trans-ferulic acid, acetovanillon, vanillin, syringaldehyde, 4-hydroxyacetophenone,
p-coumaric acid, sinapic acid, cinnamic acid, 4-hydroxybenzoic acid, and salicylic acid [
35].
Phenolic compounds, in general, exhibit stronger inhibition of clostridial fermentations than furan aldehydes. In
C. beijerinckii, the severity of inhibition follows the order of
p-coumaric acid > ferulic acid >
p-hydroxybenzoic acid > vanillic acid > syringaldehyde > vanillin [
36], whereas in
C. sacharoperbutylacetonicum N1–4, p-coumaric acid is followed by syringaldehyde and then ferulic acid [
34]. Additionally, the absence of the methoxy group in
p-coumaric acid, compared to ferulic acid, enhances the inhibitory effect, suggesting that electron-inducing groups on the aromatic ring reduce inhibition. Studies on the electric charge transferred with methyl viologen and the measurement of the concentration of NAD
+/NADH cofactors suggest that the disruption of the electron transfer chain is a primary mechanism for phenolic inhibitors [
37]. This mechanism also explains the observed decrease in activities of NADH-dependent (or associated) enzymes, such as acetyl-CoA acetyltransferase, β-hydroxybutyryl-CoA dehydrogenase, butyryl-CoA dehydrogenase, butyraldehyde dehydrogenase, and butanol dehydrogenase [
38]. Furthermore, the reduced electron capture ability by elements in lignin fragment structures may explain the decreased toxicity.
. Mechanism for the polysaccharide’s hydrolysis. (<b>a</b>) cellulose hydrolysis into glucose. (<b>b</b>) glucose transformation into hydroxymethylfurfural (HMF) and then into levulinic acid and formic acid.
. Lignin, precursors and derivatives. (a) R<sub>1</sub> = R<sub>2</sub> = –H: coumaric acid; R<sub>1</sub> = –OCH<sub>3</sub>, R<sub>2</sub> = H: ferulic acid; R<sub>1</sub> = R<sub>2</sub> = –OCH<sub>3</sub>: sinapic acid. (b) R<sub>1</sub> = R<sub>2</sub> = –H: 4-hydroxyacetophenone; R<sub>1</sub> = –OCH<sub>3</sub>, R<sub>2</sub> = H: acetovanillone. (c) R<sub>1</sub> = R<sub>2</sub> = H: 4-hydroxybenzaldehyde; R<sub>1 </sub>= –OCH<sub>3</sub>, R<sub>2</sub> = H: vanillin; R<sub>1</sub> = R<sub>2</sub> = –OCH<sub>3</sub>: syringaldehyde. (d) cinnamic acid. (e) 4-hydroxybenzoic acid. (f) salicylic acid.
strains are strict anaerobic bacteria. In the butanol production pathway, the hydrogenation of crotonyl-CoA to butyryl-CoA, catalyzed by the butyryl-CoA dehydrogenase/electron transferring flavoprotein (
bcd/
etfAB) complex, is difficult to express in recombinant systems (presumably due to oxygen sensitive) [
39]. In addition, oxidative conditions negatively affect the reducing power, which is crucial for alcohol production. The redox-responsive repressor (
rex) plays a critical role in regulating oxidative stress and the NADH/NAD
+ ratio, which affects butanol production [
40]. NAD kinase (
nadK), which synthesizes NADP
+ from NAD
+, also plays an important role in the oxidative stress [
41,
42]. It has been demonstrated that, both
dnaK from extremophile
Deinococcus wulumuqiensis R12 and native
dnaK can increase oxidative tolerance (also related to furfural inhibition) and butanol production in
C. acetobutylicum ATCC824 [
43].
The introduction of the trehalose pathway into
C. tyrobutyricum CCTCC W428 and the overexpression of transglutaminase
mtg have been shown to reduce acid and oxidative stress, resulting in increased solvent production [
42,
44]. The expression of
rprA2,
dsr, and rubredoxin (Rd) in a recombinant
nroR operon also increased oxidative tolerance in
C. acetobutylicum [
45].
C. acetobutylicum YM1 is a natural oxygen-tolerant strain that possesses various enzymes responsible for oxygen scavenging, including superoxide dismutase (SOD), catalase, and NADH/NADPH oxidases [
46]. Other endogenous genes related to oxygen tolerance in
Clostridium strains include desulfoferrodoxin (
dfx), peroxide repressor (PerR) and its homologous, and
rbr3A and
rbr3B [
47].
Various aldehydes in the biomass hydrolysates, such as vanillin and syringaldehyde, are also associated with oxidative stress [
48]. Aldehydes have an unclear relationship with reactive oxygen species, and also act as mild oxidant agents [
49]. The expression of the glutathione pathway (
gshA or
gshB genes) in
C. acetobutylicum has been shown to increase oxygen and butanol tolerance and solvent robustness, possibly related to the intermediate aldehyde compounds during solvent production, such as acetaldehyde and butyraldehyde [
50].
Co-cultures of
Clostridium strains and other microorganisms have demonstrated improved oxygen tolerance and butanol production. For instance, the co-culture of
C. acetobutylicum TSH1/Bacillus cereus TSH2 [
51,
52],
C. beijerinckii NCIMB
8052/
B. cereus CGMCC 1.895 [
53], and
C. acetobutylicum/
Saccharomyces cerevisiae [
54] have been successful in increasing production or conferring specific features by utilizing the metabolic pathways of the individual strains synergistically. Particularly, Mai et al. [
53] reported an enhanced capability of the co-culture to utilize starch, whereas Luo et al. [
54] showed that co-cultivation can enhance nutrient provision, such as amino acids, as well as increase intracellular NADH production through glucose competition. Moreover, a facultative anaerobic partner in the co-culture can consume remaining levels of oxygen in the media and aid in maintaining anaerobic conditions. Co-cultures featuring facultative anaerobic or aerobic bacteria will utilize oxygen as they interact, creating an environment conducive to anaerobic conditions, thereby supporting the growth of
Clostridium [
55].
2.5. Other Inhibitors
Elevated sodium concentration has been shown to hinder both the pentose phosphate and glycolytic pathways. However, studies have indicated that the production of solvents is not significantly affected by high sodium concentration up to 15 g/L NaCl [
56]. In contrast, when the total salinity ranges from 3%–6%, the production of butanol and butyrate in
C. acetobutylicum is altered to favor acetone production [
57].
3. Techniques for Tolerance Improvement
3.1. Evolutionary Engineering and Random Mutagenesis
Evolutionary engineering is a widely used technique to increase the tolerance of strains to different compounds or carbon sources. This technique involves gradual adaptation of the strain to live in the toxic compound or to consume the carbon source by adaptative subculturing in media containing increasing concentrations of the desired target [
58,
59]. Mutations in genes related to tolerance are generated during this process, and mutants with faster growth are selected. Comparison of genome changes between the engineered and parental strains can provide information about genes related to tolerance [
60].
Clostridium strains have shown an increase in tolerance and production using this technique, although results are unpredictable, and the resulting strains are harder to engineer due to their ticker and/or harder cell envelope.
Random mutagenesis employs physical or chemical treatments to induce mutations.
N-methyl-
N'-nitro-
N-nitrosoguanidine (NTG) is frequently used for this purpose. Following mutation, strains are screened for enhanced tolerance by exposing them to toxic levels of inhibitory chemicals or by assessing their tolerance improvement [
21]. For example, in
C. ragsdalei, ethanol production via syngas fermentation increased eightfold through chemical induced mutagenesis [
61].
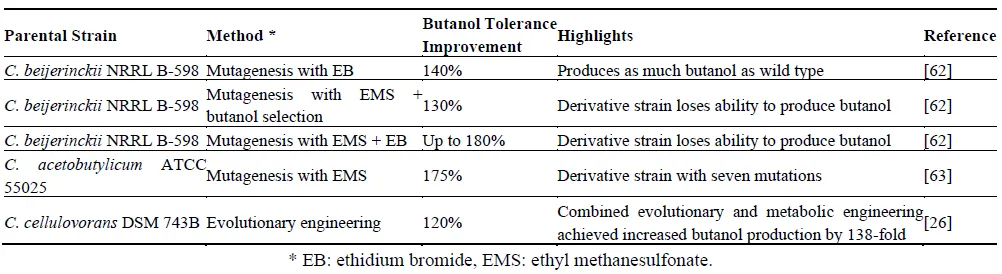
. Selected solventogenic clostridial strains improved through random mutagenesis and/or evolutionary engineering.
outlines various strains that have been enhanced through random mutagenesis and/or evolutionary engineering. Frequently, strains engineered through these approaches lose their capability for solvent production when selection pressures are applied solely for increased tolerance [
62]. This typically results from the strains adjusting their metabolic pathways to reduce stress from elevated butanol levels, often by rerouting metabolic flux towards alternative end products. Despite these setbacks in butanol production, such experimental approaches remain valuable for deciphering the roles of specific genes in tolerance. By comparing sequences of parent and modified strains or analyzing their transcriptional activities, researchers can pinpoint genes that bolster tolerance without adversely affecting butanol production. Then, techniques like genome shuffling [
64], individual selection, or gene function-based selection [
64] can be applied to integrate these beneficial tolerance genes from the modified strains into the original solvent-producing strains, thereby preserving or even improving their ability to produce solvents.
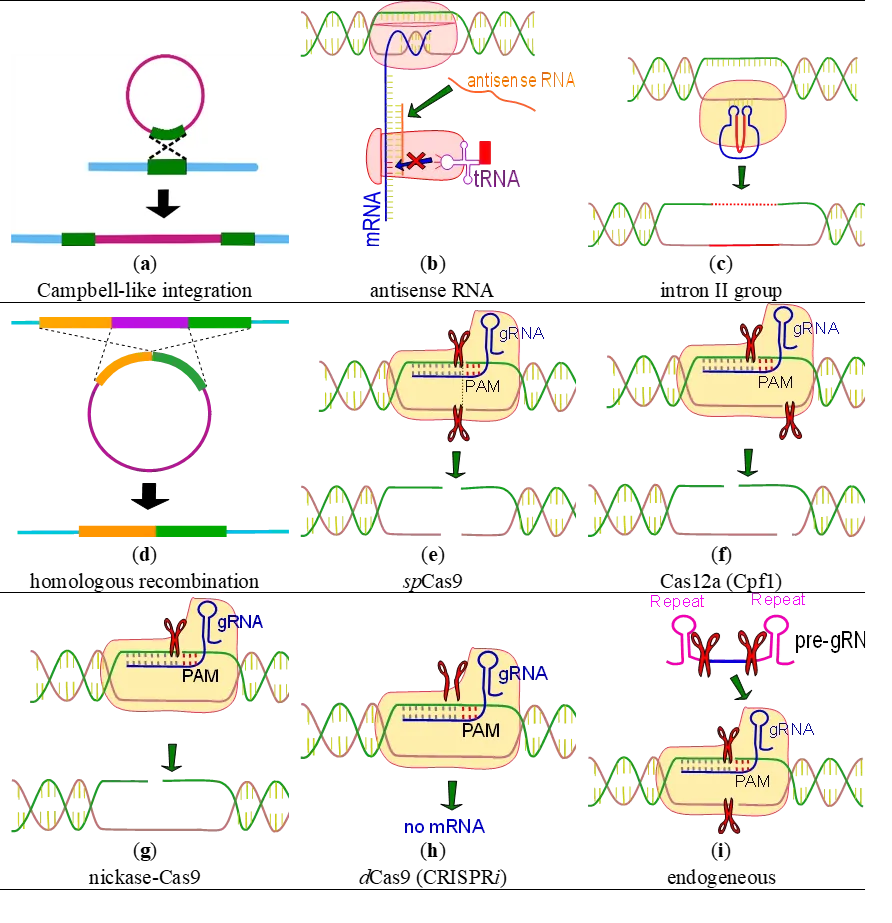
3.2. Gene Repression, Activation and Editing
Clostridium, a genus with gram-positive, spore-forming, and anaerobic characteristics, poses challenges for genetic engineering despite its significant medical and biotechnological importance. Recent advances in genetic engineering tools have enabled a better understanding and enhancement of mechanisms related to tolerance and robustness (). The application of genetic engineering in
Clostridium is relatively recent. The first plasmid was cloned into
C. acetobutylicum ATCC824 in 1992 [
65], followed by the development of Campbell-like integration (single crossover homologous recombination) mutants in the same decade [
66]. In the 2000s, antisense RNA was utilized to block gene expression, and counter-selection markers were employed for double crossover homologous recombination. Additionally, group II intron technologies, such as Targetron® (Sigma-Aldrich), were introduced [
66]. Although these group II introns represent significant advances in terms of effectiveness and work time, they operate based on gene disruption rather than deletion, which can sometimes result in residual or unexpected activity.
Recent studies have presented various alternatives for efficient, selective, markerless, and clean gene editing, inactivation, or repression using different versions of the CRISPR system. CRISPR contains nucleases that can be programmed to specifically target DNA sequences. There are five main alternatives: nickase-Cas9 (nCas9), spCRISPR-Cas9, CRISPR-Cpf1, CRISPRi, and the endogenous CRISPR system. provides a summary of these systems.
The
nCas9 was first introduced as a tool in genetic engineering of Clostridium. Given that double-stranded DNA breaks can be lethal in many bacteria, the nickase version of the Cas9 protein creates a single-stranded break, which is then repaired via homologous recombination [
67]. In a subsequent development, CRISPR-Cas9 from Streptococcus pyogenes (spCRISPR-Cas9), under an inducible promoter, was used to circumvent the lethality associated with double-stranded DNA breaks. Here, DNA templates facilitate homologous recombination, and the Cas9 gene serves as a selection tool by eliminating non-mutant strains [
68]. Another innovative approach involves a fusion of cytidine deaminase with nickase-Cas9D10A and uracil DNA glycosylase inhibitor, leading to gene inactivation through the interconversion of “CG” to “TA” and vice versa. This method, while not achieving a clean deletion, allows for the construction of small plasmids for strains that are challenging to transform [
69].
CRISPR-Cpf1 (also known as Cas12a) is a system similar to CRISPR-Cas9 but with distinct features. Cpf1 autonomously matures pre-crRNA, guided by the single-stranded mature crRNA, and cleaves target DNA with a 5-nt staggered cut. Particularly, the CRISPR-Cpf1 system from
Acidaminococcus sp. possesses the protospacer-adjacent motif (PAM) sequence, “TTTN”, which is particularly advantageous for AT-rich organisms like
Clostridium [
70]. This technique has been employed to make multiplex gene editing in
C. beijerinckii [
71].
. Genetic tools in Clostridium. (<b>a</b>) Single crossover plasmid integration event. (<b>b</b>) Antisense <em>m</em>RNA blocks gene expression. (<b>c</b>) Intron II disrupts a gene, and thus eliminates or repress its expression. (<b>d</b>) double-crossover based homologous recombination. (<b>e</b>) double-strand (ds) blunt ended DNA breakage by Cas9. (<b>f</b>) ds-sticky ended DNA breakage by Cpf1. (<b>g</b>) single-strand (ss) DNA breakage by <em>n</em>Cas9. (<b>h</b>) nuclease deficient (broken scissors) Cas binds DNA interfering with expression. (<b>i</b>) Precursor of guide RNA is expressed from plasmid, using cell machinery to generate mature <em>g</em>RNA.
CRISPR
i employs a ‘broken scissors’ principle, with dead Cas9 (dCas9) being a typical example. This nuclease-deficient version of spCas9 binds to target sequences without cutting, effectively blocking gene expression [
72]. Tested in various
Clostridium species [
73,
74], CRISPRi offers adjustable gene downregulation, making it a valuable tool for the partial repression of essential genes [
75], and the concurrent repression of multiple genes [
76]. CRISPRa, on the other hand, involves a nuclease-deficient Cas protein fused with an activation domain for gene upregulation [
77]. While not yet reported in
Clostridium, it holds potential as an alternative to plasmid-based gene overexpression.
Endogenous CRISPR systems, developed in
C. pasteurianum and
C. tyrobutyricum, are noteworthy, especially for strains difficult to transform [
18,
78]. These systems achieve high transformation efficiency due to the absence of Cas protein in plasmids. In endogenous CRISPR, a guide sequence is placed between two repeat sequences, and the bacterial machinery converts it into mature guide RNA. Then, the genome editing process utilizes a similar principle to the one previously described wherein homologous DNA arms on the plasmid enable homologous recombination. Additionally, the Cas component acts as a selective tool, targeting and eliminating strains that have not undergone mutation.
Argonautes are emerging as potential next-generation genetic tools. These proteins, present across all life domains, are known for their roles in silencing and cleaving nucleotides. Most well-characterized nucleolytic argonautes require high temperatures (>65 °C) for significant activity, limiting their applicability in genome engineering. Recent research, however, has identified
Clostridium butyricum Argonaute (CbAgo) with nucleolytic activity at 37 °C. Utilizing small interfering DNA guides (siDNA), CbAgo can be reprogrammed to break both single and double-stranded DNA [
79], thus possessing a great potential as an innovative genome editing tool. The main advantage of the argonaute based system is the absence of a PAM sequence requirement.
summarizes a list of genetically engineered solventogenic Clostridium strains, modified utilizing the genetic techniques as described above. These genetic methodologies facilitate the targeted selection and manipulation of genes aimed at simultaneously boosting tolerance and solvent production.
. Selected solventogenic clostridial strains improved by genetic engineering.
4. Genetic Mechanisms of Tolerance and Relevant Strain Engineering Efforts
4.1. Extracellular Polysaccharides
illustrates known cellular mechanisms for managing chemical stress. Biofilm, a natural mechanism that imparts robustness, consists of extracellular polymeric substances (EPS) such as polysaccharides, proteins, lipids, nucleic acids, and other macromolecules. These EPS components shield cells from environmental stressors and are among the most effective strategies for enhancing tolerance.
Biofilm plays a crucial role in cell immobilization. Biofilm reactors use a supporting material that allows cells to grow and produce biofilm. The EPS then forms a microenvironment with lower concentrations of inhibitors. Various materials, including tygon rings [
87], brick pieces [
88], activated carbon, silk, cotton, polyester [
89], and bagasse [
90], have been tested, showing a 4–7 times increase in productivity compared to planktonic cells [
89]. In batch fermentations, biofilm formation is crucial during immobilization by adsorption. Various materials, including cotton, cotton towels, linen, bamboo fiber, silk, and surface modifications with polyethylenimide and stearic acids, have been explored [
91,
92]. Some of these materials reduce the mobility of the polymeric substances and cells, thereby enhancing attachment. For example, a greater surface area (smaller particles) and cationization of the surface have been shown to promote biofilm formation [
91,
92].
The composition of the biofilm also impacts tolerance. While the EPS is primarily composed of polysaccharides, the presence of proteins in the biofilm can significantly address specific stressors. For instance, the heterologous system of Tilapia metallothionein (
OmpC-TMT) increases butanol production and oxidative tolerance in
E. coli harboring a
Clostridium pathway [
93]. Metallothioneins, known for their cysteine richness and heavy metal binding properties, counteract oxidative stress [
94]. Additionally, other sulfur compounds can protect against oxidative stress. Nitrogen-containing protein residues also contribute to reducing acid stress.
4.2. Metabolic Switch
The transition from acidogenesis to solventogenesis in
Clostridium species is governed by several genes. In
C. acetobutylicum ATCC 824,
spo0A acts as a master regulator, influencing the expression of numerous genes, including those for sporulation and the shift from acidogenesis to solventogenesis [
95], such as
adc, and the
sol operon (
adhE,
ctfA &
ctfB) [
96].
solR has been identified as a repressor of the
sol operon [
97]. A positive feedback loop involving
spo0A and its phosphorylated form (
spo0A-P) is mediated by multiple sigma factors [
95]. Histidine kinase genes related to
spo0A also influence butanol tolerance. For instance, CLOSTRON inactivation (group II intron) of the cac3319 gene from
C. acetobutylicum ATCC 55025 enhances butanol production and tolerance [
98], while overexpression of SMB_G1518-1519 reduces the tolerance [
99].
4.3. Heat Shock Proteins
Proteins may misfold under various stress conditions, including thermal and chemical stress, leading to potential inactivation and degradation. This can disrupt protein homeostasis and result in protein aggregation [
100]. Heat shock proteins act as molecular chaperones, unfolding proteins to maintain active enzymes and homeostasis.
The
groESL gene, sometimes referred to as
groE, is a widespread heat shock protein in bacteria. It consists of
groEL and
groES subunits within the same operon. The
groEL subunit is a cylinder composed of three domains: an equatorial domain housing the nucleotide pocket, an apical domain presenting a hydrophobic surface for binding non-native polypeptides,
groES, and an intermediate domain connecting the other two domains. The
groES subunit is a single, seven-membered ring, extending a hydrophobic loop to form a molecular “lid” for the central cavity [
101]. Native overexpression of
C. acetobutylicum groESL has been shown to increase butanol tolerance by 45% [
102], and similar results have been obtained with exogenous analogs. For instance,
groESL from
Deinococcus wulumuqiensis R12 and
C. acetobutylicum ATCC824 improves tolerance to butanol, furfural, oxidative, and acid stress [
43]. Expression of
groESL from
Thermoanaerobacter tengcongensis in
C. acetobutylicum enhances growth by fourfold in media containing 25% corn cob hydrolysates and increases butanol production from 0 to 0.7 g/L [
103]. Conversely,
groESL from
Pseudomonas putida did not improve tolerance in
C. acetobutylicum but enhanced thermal tolerance in
E. coli [
103].
Overexpression of the tandem genes
rbr3A and
rbr3B, which encode the heat shock protein Hsp21, has been shown to increase tolerance in
C. acetobutylicum to various stressors including cold shock, sodium, acid, oxidative stress, and butanol [
104]. Similarly,
grpE and
htpG overexpression enhance butanol tolerance by 25% and 56%, respectively [
102]. Transcriptomic analysis reveals upregulation of
grpE,
dnaK,
dnaJ,
groESL, and
htpG in
C. acetobutylicum under butanol and butyrate stress [
105], and similar findings were observed in
C. tyrobutyricum under butyrate stress. Notably, overexpression of
groESL and
htpG significantly improves tolerance of
C. tyrobutyricum to butyric acid, whereas overexpression of
dnaK and
dnaJ exhibited negative effects [
106].
4.4. Efflux Pumps
Efflux pumps, membrane proteins that actively transport chemicals, are well-known for their role in antibiotic resistance. Recent studies have also highlighted their ability to pump out small molecules. In the RND (Resistance-Nodulation-Division) superfamily, particularly the hydrophobic-amphiphilic efflux (HAE-1) subfamily in Gram-negative bacteria, efflux pumps are a focus of study [
107,
108]. RND pumps consist of an inner membrane protein responsible for extrusion, an outer pore, and an accessory lipoprotein for stabilization within the peptidoglycan [
109,
110]. The extrusion process is driven by a proton flux from the intermembrane space.
Various efflux pumps, including
Pseudomonas putida mexEF-oprN,
P. putida ttgABC,
Alcanivorax borkumensis YP_692684,
E. coli acrAB-tolC, and
P. putida srpABC, have shown potential activity against butanol and other small organic chemicals [
111]. The
acrABC pump, in particular, has been the subject of extensive study, with engineering of the
acrB extruder element to enhance specificity for butanol through selection from a random mutagenesis library [
112].
Efflux pumps are complex systems with varying levels of specificity for substrates and intricate regulation. The overexpression of efflux pumps can lead to toxicity, likely due to limitations in the expression of other membrane proteins or alterations in membrane composition. It is essential to find a balance where expression minimizes both the toxicity of the inhibitor and the pump itself [
113]. Notably, certain
P. putida strains exhibit high tolerance to butanol, making their efflux pumps of particular interest. Natural promoters of the
srpABC system elevate expression levels in the presence of various organic solvents, including aromatics, aliphatics, and alcohols. Exposure to 3 mM butanol, for instance, results in a 6.6-fold increase in expression [
114]. Furthermore, efflux pumps from the
srpABC system and
srpB alone have been shown to enhance butanol tolerance by 20%–35% in
E. coli, at butanol concentrations of 0.5%–1% [
115]. Expression of
srpB in
C. saccharoperbutylacetonicum not only enhances butanol tolerance but also increases tolerance to furan aldehydes and phenolics derived from biomass hydrolysates [
83].
4.5. Cell Envelope
Clostridium species are Gram-positive bacteria. The composition and thickness of their peptidoglycan, along with specific phospholipid elements such as phosphatidic heads and tail characteristics (e.g., saturation, trans-unsaturation rather than cis, compact steric configuration, long carbon chains, and cyclic bonds), contribute to their tolerance (). A more robust and thicker membrane increases resilience but can also make the strain more challenging to engineer. Membrane composition affects its fluidity and permeability.
The cis-trans isomerase (
cti) from
P. aeruginosa, for example, has been found to increase tolerance in
E. coli [
116]. Cyclopropane fatty acid (
cfa) from various strains, including
C. acetobutylicum, is also related to tolerance [
117]. Interestingly,
C. beijerinckii eliminates unsaturated fatty acids under alcohol stress [
118], while
C. thermocellum produces longer fatty acids [
119], attributed to higher Van der Waals forces, which increase with surface area and with linear, rigid structures. Cis bonds create a disruption in chain alignment, increasing free volume.
. Membrane compositions. Changes in phospholipid chains or tails, peptidoglycan cell walls or other components of the cell envelope, and its influence in the thickness and permeability of toxins.
also produce plasmalogen, lipids with a vinyl ether linkage opposite to the ester () [
117]. In
E. coli, several genes related to fatty acid composition, such as
fabA,
fabB,
fabD (and mutated versions),
fabF,
fabG,
fabH,
fabI,
fabZ, and
feoA, have been studied, with variants of
fabA,
fabD, and
fabH showing increased alcohol tolerance [
115]. In
C. acetobutylicum, changes in cardiolipin and glycerol acetals of plasmenylethanolamine and plasmenyl-
N-monomethylethanolamine were observed, along with a decrease in phosphatidylglycerol and the sum of phosphatidylethanolamine and phosphatidyl-
N-monomethylethanolamine [
120]. This trend seems linked to a shift from unsaturated to saturated fatty acids. Similarly,
C. pasteurianum shows increases in cardiolipin and corresponding plasmalogen [
121].
Genes associated with peptidoglycan biosynthesis, such as
glmM,
murE,
murF,
amiB,
ftsW,
ddlB, and
ftsQ, are linked to butanol tolerance in some strains [
117]. Furthermore, proline addition and enhancement of proline biosynthesis (via proABC overexpression) stabilize protein structures, maintain cell membrane functions, scavenge intracellular reactive oxygen species, and lower DNA melting points, thereby improving tolerance to lignocellulosic inhibitors [
31]. The integration of proline into cellular processes enhances the cell's resilience against environmental stressors, thus contributing to overall robustness.
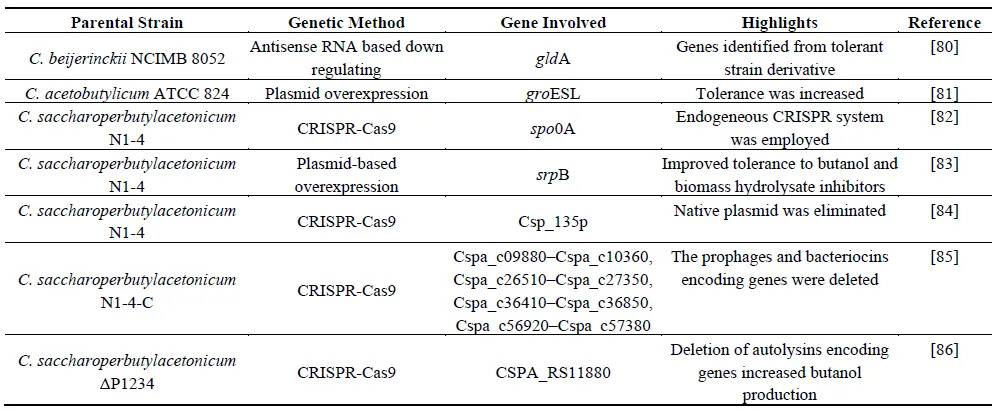
4.6. Cell Lysis
Historically, between 1974–1992, several autolysins were isolated from
C. acetobutylicum P262, such as
lit-1, and strains deficient in
lit-1 [
122,
123,
124], as well as from
C. acetobutylicum ATCC 824 [
125], and
C. saccharoperbutylacetonicum [
126,
127]. The deletion of three genes (SMB_G2359, SMB_G3117 [
128], and CA_C0554 [
129]) in
C. acetobutylicum ATCC 824 resulted in improved growth, stability, and production yield. Autolysis, a process essential for sporulation, detracts from biomass at the end of fermentation, which is not desirable for production efficiency. Recently, we systematically evaluated the role of autolysin genes within the
C. saccharoperbutylacetonicum genome related to cell autolysis and further developed more stable strains for enhanced and more stable butanol production by deleting the significant autolysin genes, thus providing an essential reference for developing robust strains for enhanced biofuel and biochemical production [
86].
4.7. Enzymatic Detoxification of Inhibitory Compounds
Various toxic compounds can be metabolized or transformed into less harmful chemical species by
Clostridium species. For example,
C. beijerinckii and
C. saccharoperbutylacetonicum can convert furan aldehydes (furfural and HMF) into their corresponding alcohols [
34,
130].
C. beijerinckii also degrades 4-hydroxybenzaldehyde and
p-coumaric acid through mechanisms not yet fully understood [
130].
Clostridium formicoaceticum oxidizes aromatic aldehydes like 4-hydroxybenzaldehyde to 4-hydroxybenzoate via a constitutive aldehyde oxidoreductase [
131]. Other
Clostridium species reduce
p-coumaric acid to
p-hydroxyhydrocinnamic acid or convert it to 4-vinylphenol and subsequently to 4-ethylphenol through decarboxylation and reduction [
132].
4.8. Stress Response Mechanisms
There are numerous genes and enzymes with unclear or poorly understood functions and mechanisms. Bioinformatic comparison of DNA and amino acid sequences suggest functions for genes or proteins based on sequence homologies [
133]. Omics technologies, aimed at the universal detection of genes (genomics), mRNA (transcriptomics), proteins (proteomics), and metabolites (metabolomics), are also employed to identify stress response mechanisms [
134]. Proteomic and transcriptomic analyses of gene regulation under various stress conditions or at different growth and metabolic stages provide powerful strategies to identify new genes related to tolerance. In
C. acetobutylicum, 29 genes were identified as upregulated in response to butanol and butyrate stress, implicating protein folding, riboflavin biosynthesis, histidine biosynthesis, ferredoxin hydrogenase, pentose & glucoronate interconversion, fatty acid metabolism, and purine metabolism. Seven genes were downregulated [
105]. Additionally, 9 genes are upregulated by butanol, acetate, and butyrate:
lonA,
hrcA-grpE-dnaK,
groESL,
ctsR-yacHI-clpC,
hsp90,
hsp18,
htrA,
CAP0102, and
aad-ctfAB, most of which are chaperones [
135]. CAC1915-CAC1944, a putative phage protein, upregulates butanol and acetate production,
adhE2,
ldh, and amino acids biosynthesis genes. Furthermore, K and Fe uptake, Na efflux, His biosynthesis, and
thlB operon are upregulated in the presence of acetate and butyrate [
135].
5. Conclusions and Prospectives
Traditional methods such as evolutionary engineering and mutagenesis for tolerance improvement have limitations, and recent advances in genetically engineered strains have surpassed the production limits achieved with older techniques. The enhancement of genetic tools not only applies existing knowledge about tolerance-related genes but also broadens our understanding of these mechanisms. As more tolerant strains typically feature tougher membranes and greater stability, the development of genetic tools is a dynamic field facing new challenges with each advancement.
The interplay between different types of chemical stress suggests that strategies targeting general robustness might be more effective than those focusing on a single stressor. However, a sole focus on tolerance can sometimes fail to achieve higher production, which is often the primary objective. Tolerance mechanisms typically consume cellular resources, including reducing power, energy, and biomass, and occupy membrane space. Therefore, balancing tolerance mechanisms with production is likely beneficial. Any strategy employed should aim to increase both productivity and tolerance.
Understanding tolerance is only one aspect of the complexity of cellular functions. Genes related to production and tolerance, and their effects on other cellular functions, need careful study to develop a robust strategy that increases butanol productivity.
Author Contributions
Conceptualization, Y.W. and P.J-B; Writing – Original Draft Preparation, P.J-B, Writing – Review & Editing, Y.W., T.W., S.W., W.L, Supervision, Y.W., D.B., Y.f.W., L-E.G-B., W.L, Project Administration, Y.W, Funding Acquisition, Y.W.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This work was supported by the US Department of Energy’s Office of Energy Efficiency and Renewable Energy under Award DE-EE0008483 (Co-Optima), the Agriculture and Food Research Initiative Competitive Grant no. 2018-67021-27715 from the USDA National Institute of Food and Agriculture (NIFA), the US National Aeronautics & Space Administration (NASA) under Award No. 80NSSC19M0195 (sub-awarded through Prairie View A&M University), the USDA-NIFA Hatch project (ALA014-1017025), and the Alabama Agricultural Experiment Station.
Declaration of competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Patakova, P, Kolek, J, Sedlar, K, Koscova, P, Branska, B, Kupkova, K, et al. Comparative analysis of high butanol tolerance and production in clostridia.
Biotech. Adv. 2018,
36, 721–738.
[Google Scholar]
-
2.
Liu, S, Qureshi, N, Hughes, S.R. Progress and perspectives on improving butanol tolerance.
World J. Microb. Biotech. 2017,
33, 51.
[Google Scholar]
-
3.
Wang S, Sun X, Yuan Q. Strategies for enhancing microbial tolerance to inhibitors for biofuel production: A review.
Bioresour. Tech. 2018,
258, 302–309.
[Google Scholar]
-
4.
Amiri H, Karimi K. Pretreatment and Hydrolysis of Lignocellulosic Wastes for Butanol Production: Challenges and Perspectives.
Bioresour. Tech. 2018,
270, 702–721.
[Google Scholar]
-
5.
Kim D. Physico-chemical conversion of lignocellulose: Inhibitor effects and detoxification strategies: A mini review.
Molecules 2018,
23, 309.
[Google Scholar]
-
6.
Maddox I, Steiner E, Hirsch S, Wessner S, Gutierrez N, Gapes J, et al. The Cause of "Acid Crash" and "Acidogenic Fermentations" During the Batch Acetone-Butanol-Ethanol(ABE-) Fermentation Process.
J. Mol. Microbiol. Biotech. 2000,
2, 95–100.
[Google Scholar]
-
7.
Reed W, Keller F, Kite F, Bogdan M, Ganoung J. Development of increased acetic acid tolerance in anaerobic homoacetogens through induced mutagenesis and continuous selection.
Enzyme Microb. Tech. 1987,
9, 117–120.
[Google Scholar]
-
8.
Wang P. Engineering Clostridium
saccharoperbutylacetonicum for enhanced isopropanol-butanolethanol (IBE)
production from lignocellulosic biomass through acetic acid pretreatment. Dissertation for
Doctoral Degree; Auburn University: Auburn, AL USA; 2018.
-
9.
Bengelsdorf FR, Poehlein A, Linder S, Erz C, Hummel T, Hoffmeister S, et al. Industrial acetogenic biocatalysts: a comparative metabolic and genomic analysis.
Front. Microbiol. 2016,
7, 1036.
[Google Scholar]
-
10.
Schwartz RD, Keller FA. Acetic acid production by Clostridium thermoaceticum in pH-controlled batch fermentations at acidic pH.
Appl. Environ. Microbiol. 1982,
43, 1385–1392.
[Google Scholar]
-
11.
Gößner AS, Picardal F, Tanner RS, Drake HL. Carbon metabolism of the moderately acid-tolerant acetogen Clostridium drakei isolated from peat.
FEMS Microbiol. Lett. 2008,
287, 236–242.
[Google Scholar]
-
12.
Kwon SJ, Lee J, Lee HS. Metabolic changes of the acetogen Clostridium sp. AWRP through adaptation to acetate challenge.
Front. Microbiol. 2022,
13, 982442.
[Google Scholar]
-
13.
Hwang JH, Kim HJ, Kim S, Lee Y, Shin Y, Choi S, et al. Positive effect of phasin in biohydrogen production of non polyhydroxybutyrate-producing Clostridium acetobutylicum ATCC 824.
Bioresour. Tech. 2024,
395, 130355.
[Google Scholar]
-
14.
Wu Y-D, Xue C, Chen L-J, Yuan W-J, Bai F-W. Improvements of metabolites tolerance in Clostridium acetobutylicum by micronutrient zinc supplementation.
Biotech. Bioproc. E. 2016,
21, 60–67.
[Google Scholar]
-
15.
Bankar SB, Jurgens G, Survase SA, Ojamo H, Granström T. Genetic engineering of Clostridium acetobutylicum to enhance isopropanol-butanol-ethanol production with an integrated DNA-technology approach.
Renewable Energy 2015,
83, 1076–1083.
[Google Scholar]
-
16.
Dai Z, Dong H, Zhu Y, Zhang Y, Li Y, Ma Y. Introducing a single secondary alcohol dehydrogenase into butanol-tolerant Clostridium acetobutylicum Rh8 switches ABE fermentation to high level IBE fermentation.
Biotech. Biofuels 2012,
5, 44.
[Google Scholar]
-
17.
Lee J, Jang Y-S, Choi SJ, Im JA, Song H, Cho JH, et al. Metabolic engineering of Clostridium acetobutylicum ATCC 824 for isopropanol-butanol-ethanol fermentation.
Appl. Environ. Microbiol. 2012,
78, 1416–1423.
[Google Scholar]
-
18.
Zhang J, Zong W, Hong W, Zhang Z-T, Wang Y. Exploiting endogenous CRISPR-Cas system for multiplex genome editing in Clostridium tyrobutyricum and engineer the strain for high-level butanol production.
Metab. Eng. 2018,
47, 49–59.
[Google Scholar]
-
19.
Li T, Zhang C, Yang K-L, He J. Unique genetic cassettes in a Thermoanaerobacterium contribute to simultaneous conversion of cellulose and monosugars into butanol.
Sci. Adv. 2018,
4, e1701475.
[Google Scholar]
-
20.
Jiménez-Bonilla P, Wang Y. In situ biobutanol recovery from clostridial fermentations: a critical review.
Crit. Rev. Biotech. 2018,
38, 469–482.
[Google Scholar]
-
21.
De Gérando HM, Fayolle-Guichard F, Rudant L, Millah S, Monot F, Ferreira NL, et al. Improving isopropanol tolerance and production of Clostridium beijerinckii DSM 6423 by random mutagenesis and genome shuffling.
Appl. Microbiol. Biotech. 2016,
100, 5427–5436.
[Google Scholar]
-
22.
Youn SH, Lee KM, Kim K-Y, Lee S-M, Woo HM, Um Y. Effective isopropanol–butanol (IB) fermentation with high butanol content using a newly isolated Clostridium sp. A1424.
Biotech. Biofuels 2016,
9, 230.
[Google Scholar]
-
23.
Yang S, Giannone RJ, Dice L, Yang ZK, Engle NL, Tschaplinski TJ, et al. Clostridium thermocellum ATCC27405 transcriptomic, metabolomic and proteomic profiles after ethanol stress.
BMC Genom. 2012,
13, 336.
[Google Scholar]
-
24.
Tolonen AC, Zuroff TR, Mohandass R, Boutard M, Cerisy T, Curtis WR. Physiology, genomics, and pathway engineering of an ethanol-tolerant strain of Clostridium phytofermentans.
Appl. Environ. Microbiol. 2015,
81, 5440–5448.
[Google Scholar]
-
25.
Zhu Y, Yang ST. Adaptation of Clostridiumtyrobutyricum for Enhanced Tolerance to Butyric Acid in a Fibrous‐Bed Bioreactor.
Biotech. Progr. 2003,
19, 365–372.
[Google Scholar]
-
26.
Wen Z, Ledesma-Amaro R, Lin J, Jiang Y, Yang S. Improved n-butanol production from Clostridium cellulovorans by integrated metabolic and evolutionary engineering.
Appl. Environ. Microbiol. 2019,
85, e02560-18.
[Google Scholar]
-
27.
Lin PP, Mi L, Morioka AH, Yoshino KM, Konishi S, Xu SC, et al. Consolidated bioprocessing of cellulose to isobutanol using Clostridium thermocellum.
Metab. Eng. 2015,
31, 44–52.
[Google Scholar]
-
28.
Higashide W, Li Y, Yang Y, Liao JC. Metabolic engineering of Clostridium cellulolyticum for production of isobutanol from cellulose.
Appl. Environ. Microbiol. 2011,
77, 2727–2733.
[Google Scholar]
-
29.
Singh S, Cheng G, Sathitsuksanoh N, Wu D, Varanasi P, George A, et al. Comparison of different biomass pretreatment techniques and their impact on chemistry and structure.
Front. Energy Res. 2015,
2, 62.
[Google Scholar]
-
30.
Mankar AR, Pandey A, Modak A, Pant KJBT. Pretreatment of lignocellulosic biomass: A review on recent advances.
Bioresour. Technol. 2021,
334, 125235.
[Google Scholar]
-
31.
Liao Z, Guo X, Hu J, Suo Y, Fu H, Wang J. The significance of proline on lignocellulose-derived inhibitors tolerance in Clostridium acetobutylicum ATCC 824.
Bioresour. Tech. 2019,
272, 561–569.
[Google Scholar]
-
32.
Ujor V, Agu CV, Gopalan V, Ezeji TC. Allopurinol-mediated lignocellulose-derived microbial inhibitor tolerance by Clostridium beijerinckii during acetone–butanol–ethanol (ABE) fermentation.
Appl. Microbiol. Biotech. 2015,
99, 3729–3740.
[Google Scholar]
-
33.
Ezeji T, Qureshi N, Blaschek HP. Butanol production from agricultural residues: impact of degradation products on Clostridium beijerinckii growth and butanol fermentation.
Biotech. Bioeng. 2007,
97, 1460–1469.
[Google Scholar]
-
34.
Yao D, Dong S, Wang P, Chen T, Wang J, Yue Z-B, Wang Y. Robustness of Clostridium saccharoperbutylacetonicum for acetone-butanol-ethanol production: Effects of lignocellulosic sugars and inhibitors.
Fuel 2017,
208, 549–557.
[Google Scholar]
-
35.
Liu J, Lin Q, Chai X, Luo Y, Guo T. Enhanced phenolic compounds tolerance response of Clostridium beijerinckii NCIMB 8052 by inactivation of Cbei_3304.
Microb. Cell Fact. 2018,
17, 35.
[Google Scholar]
-
36.
Cho DH, Lee YJ, Um Y, Sang B-I, Kim YH. Detoxification of model phenolic compounds in lignocellulosic hydrolysates with peroxidase for butanol production from Clostridium beijerinckii.
Appl. Microbiol. Biotech. 2009,
83, 1035–1043.
[Google Scholar]
-
37.
Liu J, Liu Z, Chai X, Luo Y, Guo T, Ying H. Regulation of ρ-coumaric acid tolerance in Clostridium beijerinckii by disturbing the intracellular electron transport chain.
Process Biochem. 2018,
68, 43–52.
[Google Scholar]
-
38.
Chen W-H, Zeng Y-R. Mathematical model to appraise the inhibitory effect of phenolic compounds derived from lignin for biobutanol production.
Bioresour. Tech. 2018,
261, 44–51.
[Google Scholar]
-
39.
Lan EI, Liao JC. ATP drives direct photosynthetic production of 1-butanol in cyanobacteria. Proc. Natl. Acad. Sci.
2012 109,
6018–6023, .
[Google Scholar]
-
40.
Zhang L, Nie X, Ravcheev DA, Rodionov DA, Sheng J, Gu Y, et al. Redox-responsive repressor Rex modulates alcohol production and oxidative stress tolerance in Clostridium acetobutylicum.
J. Bacteriol. 2014,
196, 3949–3963.
[Google Scholar]
-
41.
Wang C, Xin F, Kong X, Zhao J, Dong W, Zhang W, et al. Enhanced isopropanol–butanol–ethanol mixture production through manipulation of intracellular NAD (P) H level in the recombinant Clostridium acetobutylicum XY16.
Biotech. Biofuels 2018,
11, 12.
[Google Scholar]
-
42.
Wu Q, Zhu L, Xu Q, Huang H, Jiang L, Yang S-T. Tailoring the Oxidative Stress Tolerance of Clostridium tyrobutyricum CCTCC W428 by Introducing Trehalose Biosynthetic Capability.
J. Agric. Food Chem. 2017,
65, 8892–8901.
[Google Scholar]
-
43.
Liao Z, Zhang Y, Luo S, Suo Y, Zhang S, Wang J. Improving cellular robustness and butanol titers of Clostridium acetobutylicum ATCC824 by introducing heat shock proteins from an extremophilic bacterium.
J. Biotech. 2017,
252, 1–10.
[Google Scholar]
-
44.
Tao W, Dong H, Zhang Y, Cai Z, Li Y. Introducing transglutaminase with its precursor region into Clostridium acetobutylicum improves its tolerance to oxidative stress and solvent production.
Process Biochem. 2015,
50, 111–118.
[Google Scholar]
-
45.
Kawasaki S, Sakai Y, Takahashi T, Suzuki I, Niimura Y. O2 and reactive oxygen species detoxification complex, composed of O2-responsive NADH: rubredoxin oxidoreductase-flavoprotein A2-desulfoferrodoxin operon enzymes, rubperoxin, and rubredoxin, in Clostridium acetobutylicum.
Appl. Environ. Microbiol. 2009,
75, 1021–1029.
[Google Scholar]
-
46.
Al-Shorgani NKN, Kalil MS, Yusoff WMW, Hamid AA. Biobutanol production by a new aerotolerant strain of Clostridium acetobutylicum YM1 under aerobic conditions.
Fuel 2015,
158, 855–863.
[Google Scholar]
-
47.
Hillmann F, Fischer RJ, Saint‐Prix F, Girbal L, Bahl H. PerR acts as a switch for oxygen tolerance in the strict anaerobe Clostridium acetobutylicum.
Mol. Microbiol. 2008,
68, 848–860.
[Google Scholar]
-
48.
Liu Z-Y, Yao X-Q, Zhang Q, Liu Z, Wang Z-J, Zhang Y-Y, et al. Modulation of the acetone/butanol ratio during fermentation of corn stover-derived hydrolysate by Clostridium beijerinckii strain NCIMB 8052.
Appl. Environ. Microbiol. 2017,
83, e03386–03316.
[Google Scholar]
-
49.
Kang M-K, Nielsen J. Biobased production of alkanes and alkenes through metabolic engineering of microorganisms.
J. Indust. Microbiol. Biotech. 2017,
44, 613–622.
[Google Scholar]
-
50.
Zhu L, Dong H, Zhang Y, Li Y. Engineering the robustness of Clostridium acetobutylicum by introducing glutathione biosynthetic capability.
Metab. Eng. 2011,
13, 426–434.
[Google Scholar]
-
51.
Lin Z, Liu H, Yan X, Zhou Y, Cheng K, Zhang J. High-efficiency acetone-butanol-ethanol production and recovery in non-strict anaerobic gas-stripping fed-batch fermentation.
Appl. Microbiol. Biotech. 2017,
101, 8029–8039.
[Google Scholar]
-
52.
Wu P, Wang G, Wang G, Børresen BT, Liu H, Zhang J. Butanol production under microaerobic conditions with a symbiotic system of Clostridium acetobutylicum and Bacillus cereus.
Microb. Cell Fact. 2016,
15, 8.
[Google Scholar]
-
53.
Mai S, Wang G, Wu P, Gu C, Liu H, Zhang J, et al. Interactions between Bacillus cereus CGMCC 1.895 and Clostridium beijerinckii NCIMB 8052 in coculture for butanol production under nonanaerobic conditions.
Biotech. Appl. Biochem. 2017,
64, 719–726.
[Google Scholar]
-
54.
Luo H, Ge L, Zhang J, Zhao Y, Ding J, Li Z, et al. Enhancing butanol production under the stress environments of co-culturing Clostridium acetobutylicum/Saccharomyces cerevisiae integrated with exogenous butyrate addition.
PloS ONE 2015,
10, e0141160.
[Google Scholar]
-
55.
Du Y, Zou W, Zhang K, Ye G, Yang J. Advances and applications of Clostridium co-culture systems in biotechnology.
Front. Microbiol. 2020,
11, 560223.
[Google Scholar]
-
56.
Zhao, X, Condruz, S, Chen, J, Jolicoeur, M. A quantitative metabolomics study of high sodium response in Clostridium acetobutylicum ATCC 824 acetone-butanol-ethanol (ABE) fermentation.
Sci. Rep. 2016,
6, 28307.
[Google Scholar]
-
57.
Zabihi R, Mowla D, Karimi G. Production of biosolvents and acids by salinity-adapted strain of Clostridium acetobutylicum: Effects of salt and molasses concentrations.
J. Serb. Chem. Soc. 2018,
83, 411–423.
[Google Scholar]
-
58.
Lee JW, Na D, Park JM, Lee J, Choi S, Lee SY. Systems metabolic engineering of microorganisms for natural and non-natural chemicals.
Nat. Chem. Biol. 2012,
8, 536.
[Google Scholar]
-
59.
Liu W, Zhang X-Z, Zhang Z, Zhang Y-HP. Engineering of Clostridium phytofermentans endoglucanase Cel5A for improved thermostability.
Appl. Environ. Microbiol. 2010,
76, 4914–4917.
[Google Scholar]
-
60.
Sandoval-Espinola WJ, Makwana ST, Chinn MS, Thon MR, Azcárate-Peril MA, Bruno-Barcena JM. Comparative phenotypic analysis and genome sequence of Clostridium beijerinckii SA-1, an offspring of NCIMB 8052.
Microbiology 2013,
159, 2558–2570.
[Google Scholar]
-
61.
Patankar S, Dudhane A, Paradh A, Patil S. Improved bioethanol production using genome-shuffled Clostridium ragsdalei (DSM 15248) strains through syngas fermentation.
Biofuels 2018,
12, 81–89.
[Google Scholar]
-
62.
Vasylkivska M, Branska B, Sedlar K, Jureckova K, Provaznik I, Patakova P. Phenotypic and genomic analysis of Clostridium beijerinckii NRRL B-598 mutants with increased butanol tolerance.
Front. Bioeng. Biotech. 2020,
8, 598392.
[Google Scholar]
-
63.
Jain, M.K, Beacom, D, Datta, R. Mutant strain of C.
acetobutylicum and process for making butanol. US Patent 5,192,673. May 9th, 1993.
-
64.
Gao X, Zhao H, Zhang G, He K, Jin Y. Genome shuffling of Clostridium acetobutylicum CICC 8012 for improved production of acetone–butanol–ethanol (ABE).
Curr. Microbiol. 2012,
65, 128–132.
[Google Scholar]
-
65.
Mermelstein LD, Welker NE, Bennett GN, Papoutsakis ET. Expression of cloned homologous fermentative genes in Clostridium acetobutylicum ATCC 824.
Nat. Biotech. 1992,
10, 190.
[Google Scholar]
-
66.
Moon HG, Jang Y-S, Cho C, Lee J, Binkley R, Lee SY. One hundred years of clostridial butanol fermentation.
FEMS Microbiol. Lett. 2016,
363, fnw001.
[Google Scholar]
-
67.
Xu T, Li Y, Shi Z, Hemme CL, Li Y, Zhu Y, et al. Efficient genome editing in Clostridium cellulolyticum via CRISPR-Cas9 nickase.
Appl. Environ. Microbiol. 2015,
81, 4423–4431.
[Google Scholar]
-
68.
Wang Y, Zhang Z-T, Seo S-O, Lynn P, Lu T, Jin Y-S, et al. Bacterial genome editing with CRISPR-Cas9: deletion, integration, single nucleotide modification, and desirable “clean” mutant selection in Clostridium beijerinckii as an example.
ACS Synth. Biol. 2016,
5, 721–732.
[Google Scholar]
-
69.
Li Q, Seys FM, Minton NP, Yang J, Jiang Y, Jiang W, et al. CRISPR‐Cas9D10A nickase‐assisted base editing in solvent producer Clostridium beijerinckii.
Biotech. Bioeng. 2019,
113, 1475–1483.
[Google Scholar]
-
70.
Zhang J, Hong W, Zong W, Wang P, Wang Y. Markerless genome editing in Clostridium beijerinckii using the CRISPR-Cpf1 system.
J. Biotech. 2018,
284, 27–30.
[Google Scholar]
-
71.
Patinios C, de Vries ST, Diallo M, Lanza L, Verbrugge PL, López-Contreras AM, et al. Multiplex genome engineering in Clostridium beijerinckii NCIMB 8052 using CRISPR-Cas12a.
Sci. Rep. 2023,
13, 10153.
[Google Scholar]
-
72.
Wang Y, Zhang ZT, Seo SO, Lynn P, Lu T, Jin YS, et al. Gene transcription repression in Clostridium beijerinckii using CRISPR‐dCas9.
Biotech. Bioeng. 2016,
113, 2739–2743.
[Google Scholar]
-
73.
Woolston BM, Emerson DF, Currie DH, Stephanopoulos G. Rediverting carbon flux in Clostridium ljungdahlii using CRISPR interference (CRISPRi).
Metab. Eng. 2018,
48, 243–253.
[Google Scholar]
-
74.
Rostain W, Zaplana T, Boutard M, Baum C, Tabuteau S, Sanitha M, et al. Tuning of Gene Expression in Clostridium phytofermentans Using Synthetic Promoters and CRISPRi.
ACS Synth. Biol. 2022,
11, 4077–4088.
[Google Scholar]
-
75.
Liu X, Gallay C, Kjos M, Domenech A, Slager J, van Kessel SP, et al. High‐throughput CRISPRi phenotyping identifies new essential genes in Streptococcus pneumoniae.
Mol. Syst. Biol. 2017,
13, 931.
[Google Scholar]
-
76.
Joseph RC, Sandoval NR. Single and multiplexed gene repression in solventogenic Clostridium via Cas12a-based CRISPR interference.
Synth. Syst. Biotech. 2023,
8, 148–156.
[Google Scholar]
-
77.
La Russa MF, Qi LS. The new state of the art: Cas9 for gene activation and repression.
Mol. Cell. Biol. 2015,
35, 3800–3809.
[Google Scholar]
-
78.
Pyne ME, Bruder MR, Moo-Young M, Chung DA, Chou CP. Harnessing heterologous and endogenous CRISPR-Cas machineries for efficient markerless genome editing in Clostridium.
Sci. Rep. 2016,
6, 25666.
[Google Scholar]
-
79.
Hegge JW, Swarts DC, Chandradoss SD, Cui TJ, Kneppers J, Jinek M, et al. DNA-guided DNA cleavage at moderate temperatures by Clostridium butyricum Argonaute.
Nucleic Acid Res. 2019,
47, 5809–5821.
[Google Scholar]
-
80.
Liyanage H, Young M, Kashket E. Butanol tolerance of Clostridium beijerinckii NCIMB 8052 associated with down-regulation of gldA by antisense RNA.
J. Mol. Microbiol. Biotech. 2000,
2, 87–93.
[Google Scholar]
-
81.
Tomas CA, Beamish J, Papoutsakis ET. Transcriptional analysis of butanol stress and tolerance in Clostridium acetobutylicum.
J. Bacteriol. 2004, ,
186, 2006–2018.
[Google Scholar]
-
82.
Atmadjaja AN, Holby V, Harding AJ, Krabben P, Smith HK, Jenkinson ER. CRISPR-Cas, a highly effective tool for genome editing in Clostridium saccharoperbutylacetonicum N1-4 (HMT).
FEMS Microbiol. Lett. 2019,
366, fnz059.
[Google Scholar]
-
83.
Jiménez-Bonilla P, Zhang J, Wang Y, Blersch D, de-Bashan L-E, Guo L, et al. Enhancing the tolerance of Clostridium saccharoperbutylacetonicum to lignocellulosic-biomass-derived inhibitors for efficient biobutanol production by overexpressing efflux pumps genes from Pseudomonas putida.
Bioresour. Tech. 2020,
312, 123532.
[Google Scholar]
-
84.
Gu Y, Feng J, Zhang Z-T, Wang S, Guo L, Wang Y, et al. Curing the endogenous megaplasmid in Clostridium saccharoperbutylacetonicum N1-4 (HMT) using CRISPR-Cas9 and preliminary investigation of the role of the plasmid for the strain metabolism.
Fuel 2019,
236, 1559–1566.
[Google Scholar]
-
85.
Feng J, Zhang J, Wang P, Jimenez-Bonilla P, Gu Y, Zhou J, et al. Renewable Fatty Acid Ester Production in Clostridium.
BioRxiv 2020,
0, 014746. doi:10.1101/2020.03.29.014746.
[Google Scholar]
-
86.
Jiménez-Bonilla P, Feng J, Wang S, Zhang J, Wang Y, Blersch D, et al. Identification and Investigation of Autolysin Genes in Clostridium saccharoperbutylacetonicum Strain N1-4 for Enhanced Biobutanol Production.
Appl. Environ. Microbiol. 2021,
87, e02442–02420.
[Google Scholar]
-
87.
Raganati F, Procentese A, Olivieri G, Russo ME, Gotz P, Salatino P, et al. Butanol production by Clostridium acetobutylicum in a series of packed bed biofilm reactors.
Chem. Eng. Sci. 2016,
152, 678–688.
[Google Scholar]
-
88.
Qureshi N, Karcher P, Cotta M, Blaschek HP. High-productivity continuous biofilm reactor for butanol production.
Appl. Biochem. Biotech. 2004,
114, 713–721.
[Google Scholar]
-
89.
Liu D, Chen Y, Ding F-Y, Zhao T, Wu J-L, Guo T, et al. Biobutanol production in a Clostridium acetobutylicum biofilm reactor integrated with simultaneous product recovery by adsorption.
Biotech. Biofuels 2014,
7, 5.
[Google Scholar]
-
90.
Zhou W, Liu J, Fan S, Xiao Z, Qiu B, Wang Y, et al. Biofilm immobilization of Clostridium acetobutylicum on particulate carriers for acetone-butanol-ethanol (ABE) production.
Biores. Tech. Rep. 2018,
3, 211–217.
[Google Scholar]
-
91.
Zhuang W, Liu X, Yang J, Wu J, Zhou J, Chen Y, et al. Immobilization of Clostridium acetobutylicum onto natural textiles and its fermentation properties.
Microb. Biotech. 2017,
10, 502–512.
[Google Scholar]
-
92.
Zhuang W, Yang J, Wu J, Liu D, Zhou J, Chen Y, et al. Extracellular polymer substances and the heterogeneity of Clostridium acetobutylicum biofilm induced tolerance to acetic acid and butanol.
RSC Adv. 2016,
6, 33695–33704.
[Google Scholar]
-
93.
Chin W-C, Lin K-H, Liu C-C, Tsuge K, Huang C-C. Improved n-butanol production via co-expression of membrane-targeted tilapia metallothionein and the clostridial metabolic pathway in Escherichia coli.
BMC Biotech. 2017,
17, 36.
[Google Scholar]
-
94.
Ruttkay-Nedecky B, Nejdl L, Gumulec J, Zitka O, Masarik M, Eckschlager T, et al. The role of metallothionein in oxidative stress.
Int. J. Mol. Sci. 2013,
14, 6044–6066.
[Google Scholar]
-
95.
Liao C, Seo S-O, Celik V, Liu H, Kong W, Wang Y, et al. Integrated, systems metabolic picture of acetone-butanol-ethanol fermentation by Clostridium acetobutylicum.
Proc. Natl. Acad. Sci. 2015,
112, 8505–8510.
[Google Scholar]
-
96.
Fischer RJ, Helms J, Dürre P. Cloning, sequencing, and molecular analysis of the sol operon of Clostridium acetobutylicum, a chromosomal locus involved in solventogenesis.
J. Bacteriol. 1993,
175, 6959–6969.
[Google Scholar]
-
97.
Lee SY, Park JH, Jang SH, Nielsen LK, Kim J, Jung KS. Fermentative butanol production by Clostridia.
Biotech. Bioeng. 2008,
101, 209–228.
[Google Scholar]
-
98.
Xu M, Zhao J, Yu L, Tang I-C, Xue C, Yang S-T. Engineering Clostridium acetobutylicum with a histidine kinase knockout for enhanced n-butanol tolerance and production.
Appl. Microbiol. Biotech. 2015,
99, 1011–1022.
[Google Scholar]
-
99.
Jia K, Zhang Y, Li Y. Identification and characterization of two functionally unknown genes involved in butanol tolerance of Clostridium acetobutylicum.
PloS ONE 2012,
7, e38815.
[Google Scholar]
-
100.
Żwirowski S, Kłosowska A, Obuchowski I, Nillegoda NB, Piróg A, Ziętkiewicz S, et al. Hsp70 displaces small heat shock proteins from aggregates to initiate protein refolding.
EMBO J. 2017,
36, 783–796.
[Google Scholar]
-
101.
Horwich AL, Farr GW, Fenton WA. GroEL−GroES-mediated protein folding.
Chem. Rev. 2006,
106, 1917–1930.
[Google Scholar]
-
102.
Mann MS, Dragovic Z, Schirrmacher G, Lütke-Eversloh T. Over-expression of stress protein-encoding genes helps Clostridium acetobutylicum to rapidly adapt to butanol stress.
Biotech. Lett. 2012,
34, 1643–1649.
[Google Scholar]
-
103.
Luan G, Dong H, Zhang T, Lin Z, Zhang Y, Li Y, et al. Engineering cellular robustness of microbes by introducing the GroESL chaperonins from extremophilic bacteria.
J. Biotech. 2014,
178, 38–40.
[Google Scholar]
-
104.
Hillmann F, Fischer R-J, Bahl H. The rubrerythrin-like protein Hsp21 of Clostridium acetobutylicum is a general stress protein.
Arch. Microbiol. 2006,
185, 270.
[Google Scholar]
-
105.
Wang Q, Venkataramanan KP, Huang H, Papoutsakis ET, Wu CH. Transcription factors and genetic circuits orchestrating the complex, multilayered response of Clostridium acetobutylicum to butanol and butyrate stress.
BMC Syst. Biol. 2013,
7, 120.
[Google Scholar]
-
106.
Suo Y, Luo S, Zhang Y, Liao Z, Wang J. Enhanced butyric acid tolerance and production by Class I heat shock protein-overproducing Clostridium tyrobutyricum ATCC 25755.
J. Indust. Microbiol. Biotech. 2017,
44, 1145–1156.
[Google Scholar]
-
107.
Anes J, McCusker MP, Fanning S, Martins M. The ins and outs of RND efflux pumps in Escherichia coli.
Front. Microbiol. 2015,
6, 587.
[Google Scholar]
-
108.
Nikaido H. RND transporters in the living world.
Res. Microbiol. 2018,
169, 363–371.
[Google Scholar]
-
109.
Ramos J-L, Sol Cuenca M, Molina-Santiago C, Segura A, Duque E, Gómez-García MR, Udaondo Z, Roca A. Mechanisms of solvent resistance mediated by interplay of cellular factors in Pseudomonas putida.
FEMS Microbiol. Rev. 2015,
39, 555–566.
[Google Scholar]
-
110.
Nikaido H, Takatsuka Y. Mechanisms of RND multidrug efflux pumps.
BBA Proteins Proteom. 2009,
1794, 769–781.
[Google Scholar]
-
111.
Dunlop MJ, Dossani ZY, Szmidt HL, Chu HC, Lee TS, Keasling JD, et al. Engineering microbial biofuel tolerance and export using efflux pumps.
Mol. Syst. Biol. 2011,
7, 487.
[Google Scholar]
-
112.
Fisher MA, Boyarskiy S, Yamada MR, Kong N, Bauer S, Tullman-Ercek D. Enhancing tolerance to short-chain alcohols by engineering the Escherichia coli AcrB efflux pump to secrete the non-native substrate n-butanol.
ACS Synth. Biol. 2013,
3, 30–40.
[Google Scholar]
-
113.
Turner WJ, Dunlop MJ. Trade-offs in improving biofuel tolerance using combinations of efflux pumps.
ACS Synth. Biol. 2014,
4, 1056–1063.
[Google Scholar]
-
114.
Kieboom J, Dennis JJ, Zylstra GJ, De Bont JA. Active Efflux of Organic Solvents byPseudomonas putida S12 Is Induced by Solvents.
J. Bacteriol. 1998,
180, 6769–6772.
[Google Scholar]
-
115.
Bui LM, Lee JY, Geraldi A, Rahman Z, Lee JH, Kim SC. Improved n-butanol tolerance in Escherichia coli by controlling membrane related functions.
J. Biotech. 2015,
204, 33–44.
[Google Scholar]
-
116.
Tan Z, Yoon JM, Nielsen DR, Shanks JV, Jarboe LR. Membrane engineering via trans unsaturated fatty acids production improves Escherichia coli robustness and production of biorenewables.
Metab. Eng. 2016,
35, 105–113.
[Google Scholar]
-
117.
Sandoval NR, Papoutsakis ET. Engineering membrane and cell-wall programs for tolerance to toxic chemicals: Beyond solo genes.
Curr. Opin. Microbiol. 2016,
33, 56–66.
[Google Scholar]
-
118.
Huffer S, Clark ME, Ning JC, Blanch HW, Clark DS. The role of alcohols in growth, lipid composition, and membrane fluidity of yeast, bacteria, and archaea.
Appl. Environ. Microbiol. 2011,
77, 6400–6408.
[Google Scholar]
-
119.
Timmons MD, Knutson BL, Nokes SE, Strobel HJ, Lynn BC. Analysis of composition and structure of Clostridium thermocellum membranes from wild-type and ethanol-adapted strains.
Appl. Microbiol. Biotech. 2009,
82, 929–939.
[Google Scholar]
-
120.
Tian B, Guan Z, Goldfine H. An ethanolamine-phosphate modified glycolipid in Clostridium acetobutylicum that responds to membrane stress. BBA-Mol.
Cell Biol. Lipids 2013,
1831, 1185–1190.
[Google Scholar]
-
121.
Kolek J, Patáková P, Melzoch K, Sigler K, Řezanka T. Changes in membrane plasmalogens of Clostridium pasteurianum during butanol fermentation as determined by lipidomic analysis.
PLoS ONE 2015,
10, e0122058.
[Google Scholar]
-
122.
Van Der Westhuizen A, Jones DT, Woods DR. Autolytic activity and butanol tolerance of Clostridium acetobutylicum.
Appl. Environ. Microbiol. 1982,
44, 1277–1281.
[Google Scholar]
-
123.
Webster JR, Reid SJ, Jones DT, Woods DR. Purification and characterization of an autolysin from Clostridium acetobutylicum.
Appl. Environ. Microbiol. 1981,
41, 371–374.
[Google Scholar]
-
124.
Allcock ER, Reid SJ, Jones DT, Woods DR. Autolytic activity and an autolysis-deficient mutant of Clostridium acetobutylicum. Appl. Environ. Microbiol. ,
1981, 42. 929–935
[Google Scholar]
-
125.
Croux C, Canard B, Goma G, Soucaille P. Autolysis of Clostridium acetobutylicum ATCC 824.
Microbiology 1992,
138, 861–869.
[Google Scholar]
-
126.
Yoshino S, Ogata S, Hayashida S. Some properties of autolysin of Clostridium saccharoperbutylacetonicum.
Agr. Biol. Chem. 1982,
46, 1243–1248.
[Google Scholar]
-
127.
Ogata S, Hongo M. Lysis induced by sodium ion and its relation to lytic enzyme systems in Clostridium saccharoperbutylacetonicum.
Microbiology 1974,
81, 315–323.
[Google Scholar]
-
128.
Yang L, Bao G, Zhu Y, Dong H, Zhang Y, Li Y. Discovery of a novel gene involved in autolysis of Clostridium cells.
Protein Cell 2013,
4, 467–474.
[Google Scholar]
-
129.
Liu Z, Qiao K, Tian L, Zhang Q, Liu Z-Y, Li F-L. Spontaneous large-scale autolysis in Clostridium acetobutylicum contributes to generation of more spores.
Front. Microbiol. 2015,
6, 950.
[Google Scholar]
-
130.
Zhang Y. Detoxification of Lignocellulose-derived
Microbial Inhibitory Compounds by Clostridium beijerinckii NCIMB 8052 during Acetone-Butanol-Ethanol
Fermentation. Dissertation for Doctoral Degree; The Ohio State University:
Columbus, OH 43210, USA; 2013.
-
131.
Frank C, Schwarz U, Matthies C, Drake HL. Metabolism of aromatic aldehydes as cosubstrates by the acetogen Clostridium formicoaceticum.
Arch. Microbiol. 1998,
170, 427–434.
[Google Scholar]
-
132.
Chamkha M, Garcia J-L, Labat M. Metabolism of cinnamic acids by some Clostridiales and emendation of the descriptions of Clostridium aerotolerans, Clostridium celerecrescens and Clostridium xylanolyticum.
Int. J. Systemat. Evolutionary Microbiol. 2001,
51, 2105–2111.
[Google Scholar]
-
133.
Schmid, R, Schmidt-Dannert, C. Biotechnollogy. An ilustrated primer; Wiley-VCH: Weinheim, Germany; 2016.
-
134.
Horgan RP, Kenny LC. ‘Omic’technologies: genomics, transcriptomics, proteomics and metabolomics.
Obstetrician Gynaecologist 2011,
13, 189–195.
[Google Scholar]
-
135.
Alsaker KV, Paredes C, Papoutsakis, E.T. Metabolite stress and tolerance in the production of biofuels and chemicals: gene‐expression‐based systems analysis of butanol, butyrate, and acetate stresses in the anaerobe Clostridium acetobutylicum.
Biotech. Bioeng. 2010,
105, 1131–1147.
[Google Scholar]
 Shangjun Wang
1
Tyler Whitfield
1
David Blersch
1
Yifen Wang
1
Luz-Estela Gonzalez-de-Bashan
3,4,5
Wei Luo
6
Yi Wang
1,7,8,*
Shangjun Wang
1
Tyler Whitfield
1
David Blersch
1
Yifen Wang
1
Luz-Estela Gonzalez-de-Bashan
3,4,5
Wei Luo
6
Yi Wang
1,7,8,*