1. Introduction
Cyanobacteria’s ability to use solar radiation to fix atmospheric CO
2 provides the opportunity for engineering sustainable production of chemicals while simultaneously benefiting from their simple nutrient requirements and innate ability to occupy niches unsuitable for the production of food [
1,
2,
3].
Terpenoids are the largest class of natural products and can be found in all living organisms [
4,
5]. Their structural and functional diversity affords many of the compounds with economic significance in the fragrance, cosmetics, food ingredients, and pharmaceutical industries. Much of our current production of terpenoids relies on traditional chemical synthesis or extraction and isolation from plant material [
6]. Extraction from plants is often based on the use of organic solvents, proving wasteful with low yields of the desired terpenoid from significant amounts of plant biomass. The extraction of diterpenoids such as forskolin, ginkgolides, and triptonide, which are exclusively synthesized in the roots of their host plants, necessitates the excavation and harvesting of root materials [
7,
8,
9,
10]. However, this process invariably destroys the plants, imposing additional adverse ecological costs such as soil erosion. Furthermore, each harvest demands the cultivation of a new generation of plants, leading to an increased demand for arable land.
Biosynthesis of terpenoids is dependent on complex modular processes. In cyanobacteria, the C5 isoprene building blocks are derived from the 2-C-methyl-d-erythritol 4-phosphate pathway (MEP pathway) [
11]. The linear carbon skeletons of terpenoids are derived from the condensation of C5 isopentenyl diphosphate (IDP) and dimethylallyl diphosphate (DMADP). The length of the terpenoid is dependent on the number of C5 building blocks used as starting material for terpene synthases that catalyzes the production of monoterpenoids (C10), sesquiterpenoids (C15), and diterpenoids (C20). In the case of diterpenoids, diterpene synthases (diTPSs) catalyze the formation of multicyclic backbones, while functionalization of the cyclic structures is catalyzed by ER-bound cytochrome P450s (P450s) resulting in stereospecific and regioselective oxygenations [
9,
10,
12,
13,
14].
In prokaryotes, the MEP pathway is the only pathway present for the production of terpenoid precursors. Cyanobacteria already produce terpenoids that are crucial for their photosynthetic machinery. These include the phytol tail of chlorophylls, carotenoids, tocopherols, and phylloquinones [
11,
15] Thus, cyanobacteria have an inherent potential as heterologous hosts for the production and storage of additional terpenoids [
16].
A common strategy to improve terpenoid production is to increase the endogenous supply of precursor metabolites [
17]. Overexpressing rate-limiting enzymes to enhance precursor supply is one commonly used strategy. The first committed step in the MEP pathway is the condensation of pyruvate and glyceraldehyde-3-phosphate (GAP) to yield 1-deoxy-D-xylulose-5-phosphate (DXP) catalyzed by 1-Deoxy-D-xylulose-5-phosphate synthase (DXS) [
18]. DXS is the rate-limiting enzyme to control flux through the MEP pathway [
19], and overexpression of DXS has led to increased titers of terpenes and terpenoids (reviewed in [
20]). Likewise, the enzyme GGDP synthase (GGPDS) which condenses three molecules of IDP and one molecule of DMADP to form geranylgeranyl diphosphate (GGDP, C20), is the entry point leading to the biosynthesis of diterpenoids as well as carotenoids and the phytol tail of chlorophylls. Geranylgeranyl diphosphate synthase (GGDPS) is thus considered a key metabolic engineering target and accumulation of GGDP requires an uninhibited flux towards GGDP and usually a strong “metabolic pull” strategy is used by overexpressing GGDP synthase [
21,
22].
In this study, we focus on the initial steps of the biosynthetic pathway leading to the diterpenoid forskolin. Forskolin is naturally produced and present in the root cork of the plant Plectranthus barbatus. The initial steps in the forskolin pathway involves two terpene synthases (
PbTPS2 and
PbTPS3) [
8]. A previous report showed that expression of
PbTPS2 and
PbTPS3 in
Synechocystis resulted in the formation of 13-R-manoyl oxide (13-R-MO), a precursor in the forskolin pathway [
23]. Here we report successful engineering by increasing carbon flux towards the endogenous metabolite GGDP which resulted in an 11-fold increase in production of 13-R-MO in comparison to the
Synechocystis strain only expressing the two terpene synthases. We also demonstrate that removal of 13-R-MO from the culture medium using a dodecane overlay can potentially triple production, albeit in a different cultivation system.
2. Materials and Methods
2.1. Strains and Media
All cloning was performed using
E. coli NEB 10-β carrying either pDF-trc [
24] or pSL2387 (kind gift from Prof. Himadri Pakrasi) grown in standard LB media supplemented with 50 µg/mL spectinomycin or kanamycin. Conjugation was performed using
E. coli strain HB101 carrying the helper plasmid pRL443 supplemented with 100 µg/mL carbenicillin in standard LB media. Wild type
Synechocystis PCC. 6803 and resulting strains were grown on solid BGH5 media or in liquid BGH11 media (BG5 or BG11 supplemented with 1.5 or 4.75 g/L HEPES, respectively) [
25].
2.2. Culturing Conditions
2.2.1. Starter Cultures
Starter cultures were grown from freezer stocks on plates at 30 °C at approximately 50 µE irradiation using fluorescent lights. Liquid cultures were supplemented by flushing with 3% CO
2 (
v/
v).
2.2.2. Photobioreactor Cultures
Growth curves were performed on Multi Cultivator MC 1000-OD (Photon System Instruments; Drásov, Czech Republic). Starter cultures (20 mL) were inoculated from plates and grown in liquid cultures as described above. Initial 70 mL cultures were inoculated at 0.3 OD
730 in BGH11 and supplemented with 100 µg/mL kanamycin and spectinomycin. Cultures were flushed with 3% CO2 (
v/
v), grown at 30 °C, and in 100 µE of light. Samples were taken every 24 h for 7 day. Kanamycin (100 µg/mL) was supplemented each day. Cultures were induced on day 2 with 1 mM IPTG. Cells were harvested at the end of the run for further analysis.
2.3. Overlay Experiment
Overlay experiments were performed in 40 mL cultures in Erlenmeyer flasks in BGH11 supplemented with 50 mM NaHCO
3 and Na
2CO
3 with or without the addition of 10 mL
N-dodecane. Flasks were grown at 30 °C with constant shaking under 100 µE of light. Initial cultures were inoculated at 0.2 OD
730 and time points were taken every 24 h for 7 day. Cells and the overlay were harvested and stored in glass vials with PTFE lids for quantification of 13-R-MO using GC-MS analysis (Shimadzu, Ballerup, Denmark).
2.4. Cloning and Transformation
PbTPS2,
PbTPS3,
SjGGDPS,
PbGGDPS, and
ScGGDPS were codon optimized and obtained from GenScript. All amplifications were performed using Q5 Polymerase (New England BioLabs; Ipswich, MA, USA). All cloning was performed using HiFi DNA Assembly (New England BioLabs). The TPS construct comprised of
PbTPS2 and
PbTPS3 in an operon driven by the Pcpc560 promoter and integrated into the pSL2387 backbone containing the NSP1 flanking homology regions.
PbTPS2 and
PbTPS3 were tagged with the Strep-II and HA tag respectively. The three GGDPSs were tagged with the HA tag and were driven by the trc promoter located on the pDF-trc backbone. Additionally, the
SjGG/D construct comprised of the
SjGGDPS and a mutant DXS in an operon [
26]. The mutant DXS was generated by overlap extension PCR. All construct sequences used can be found in Supplementary Table S1.
Transformation was performed by either natural transformation for the integrative TPS construct or triparental mating for the replicative GGDPS constructs. Natural transformation was performed as follows. Roughly 10 mL of WT at an OD
730 around 0.8 were spun down and washed once to remove any residual antibiotics. Cells were resuspended in 10 mL BGH11 and 200 µL of the resulting cell suspension were used per construct. Roughly 5 µg of purified plasmid was mixed with 200 µL of cells and incubated for 4 h at RT in 100 µE light. Cells were then plated on BGH5 plates without antibiotics and incubated for 1 day at 30 °C under 30 µE of light. After incubation for a day, the cells were scraped off the BGH5 plates and plated onto BGH5 plates with 25 µg/mL kanamycin. Following the appearance of single colonies, colonies were picked and restreaked onto BGH5 with 50 µg/mL kanamycin. This process was repeated until the strain was fully segregated. Triparental mating was performed as described in [
27].
2.5. Western Blotting
Cell pellets on day 7 of the photobioreactor run were harvested from 10 mL of culture and resuspended in approximately 600 µL of 50 mM Tris-HCl pH 7.5 containing 2X cOmplete EDTA- free protease inhibitor (Roche; Copenhagen, Denmark). Zirconium oxide beads (200 µL, 0.15 mm diameter) were added to the cell suspension and disrupted using a NextAdvance Bullet Blender (Troy, NY, USA). Samples were processed 3 times at power 12 for 5 min each time. The resulting whole-cell lysate was quantified for total protein content using a Pierce BCA Protein Assay Kit (Thermo Scientific; Roskilde, Denmark).
Total protein (10 or 40 µg) was boiled at 90 °C for 5 min in 2% SDS/0.1 M DTT/10% glycerol/0.05 M Tris HCl pH 6.8 and applied onto 12% TGX stain-free gels (Bio-Rad; Copenhagen, Denmark). Gels were run at 250 V for 30–35 min in Tris-glycine-SDS buffer (Bio-Rad). Proteins were transferred onto a Trans-Blot Turbo PVDF membrane for 7 min at 2.5 A using a Trans-Blot Turbo blotting system (Bio-Rad). Membranes were blocked for 1 h at room temperature in 5% skim milk in PBS-T (PBS with 0.05% Tween-20) and washed with PBS-T. Primary anti-HA antibodies (dilution 1:1000) were applied for 1 h at room temperature in 2% skim milk in PBS-T with a. Membranes were washed in PBS-T and secondary swine anti-rabbit HRP conjugated antibodies (Dako; Glostrup, Denmark) (dilution of 1:5000) were applied in 2% skim milk in PBS-T with a for 1 h. Chemiluminescence was detected using Super Signal West Dura substrate (Thermo Scientific) using a ChemiDoc MP imaging system equipped with a cold CCD camera (Bio-Rad).
2.6. GC-MS Analysis
13-R-MO quantification was carried out using the cell pellet obtained from 10 to 20 mL of a day 7 culture from from the photobioreactor run. The pellet was resuspended in 8 mL of 10% methanolic KOH and the samples were incubated at 65 °C for 1 h. Liquid-liquid extraction was performed by the addition of 2 mL of hexane. For quantification of 13-R-MO in the overlay, direct injection of dodecane from the overlay experiments was used.
GC-MS analysis was performed on a Shimadzu GCMS-QP2010 PLUS instrument using a HP-5MS UI column (20 m × 0.18 mm i.d.; 0.18 µm film thickness) (Shimadzu, Ballerup, Denmark). Sampling temperature and time occurred at 250 °C and 1 min in splitless mode. Chromatography was carried out using H
2 as the carrier gas, with a linear velocity of 50 cm/s, and a purge flow of 1 mL/min. The gradient used was as follows: 60 °C for 1 min; 60 °C increasing at a rate of 30 °C/min until reaching 150 °C; 150 °C increasing at a rate of 15 °C/min until reaching 250 °C; 250 °C increasing at a rate of 30 °C/min reaching 280 °C; hold at 280 °C for 3 min. 1 µL of either hexane or dodecane extract was injected into the column. The ion source temperature was set to 250 °C, with spectral recording between 50
m/
z to 400
m/
z. MS acquisition was set to start after 3 min or 8 min for hexane or dodecane extracts respectively.
3. Results
3.1. Strain Design and Construction
A base strain capable of producing 13-R MO was constructed by the introduction of the two terpene synthases,
PbTPS2 and
PbTPS3, from
Plectranthus barbatus [
7] (A). The TPS cassette was introduced into the NSP1 neutral site under the control of the Pcpc560 promoter [
28,
29]. Two steps in the MEP pathway were chosen for engineering in accordance with the “push-pull” strategy, namely DXS and GGDPS. DXS is positioned at the start of the MEP pathway and catalyzes the condensation of two central carbon metabolites, glyceraldehyde-3-phosphate and pyruvate. GGDPS catalyzes the condensation of 4 units of DMADP and IDP in order to form geranylgeranyl diphosphate, the substrate of
PbTPS2 and
PbTPS3 [
11]. A prokaryotic, a yeast, and a plant derived GGDPS was selected for screening:
Synechococcus JA-3-3Ab (
SjGGDPS),
Plectranthus barbatus (
PbGGDPS), and
Saccharomyces cerevisiae (
ScGGDPS).
SjGGDPS was chosen among cyanobacterial GGDPS to represent a prokaryotic enzyme and was previously used to enhance forskolin titers in yeast [
30].
PbGGDPS was chosen as derived from the original host organism of the forskolin biosynthetic pathway. Enzymes from the same biosynthetic pathway are known to be able to form protein-protein interactions that can potentially facilitate substrate channeling [
31,
32,
33,
34].
ScGGDPS was chosen due to its evolutionary divergence from both plants and cyanobacteria, potentially avoiding endogenous feedback regulation that may or may not occur in
Synechocystis. An alignment of all three GGDPSs is shown in Figure S1, where
ScGGDPS is shown to differ drastically from both
SjGGDPS and
PbGGDPS by the presence of two extra sequences inserted at the C-terminus. Mutations from a DXS variant shown to be responsible for the high monoterpenol content in a
Vitis vinifera cultivar were introduced into
E. coli DXS and expressed in an operon with the cyanobacterial
SjGGDPS (resulting in
SjGG/D) [
26]. All resulting constructs were introduced into the replicative pDF-trc under the IPTG inducible Ptrc promoter.
3.2. Growth Characterization and Expression
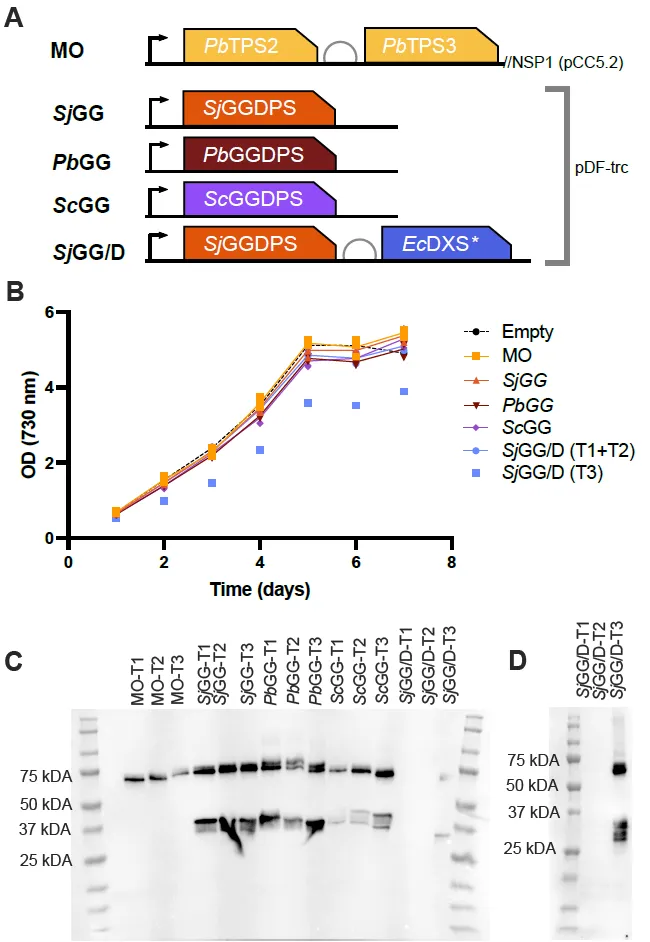
Initial verification of transformants and their possible ability to produce 13-R-MO was carried out by culturing transformants in a photobioreactor for 7 day. Strains were grown in triplicate as described in the Materials and Methods. No large or significant effect on the final cell density between any of the strains was observed (B). One particular transformant carrying the
SjGG/D construct grew slower and reached a lower final OD
730 than the other two strains carrying the same construct. The growth of transformant
SjGG/D T3 is shown in B (light blue diamonds). The final OD
730 of the other two transformants were 5.0 and 5.2, while the final OD
730 of
SjGG/D T3 only reached 3.9. This transformant was later found to be the sole transformant from the
SjGG/D construct capable of producing 13-R-MO.
Cells were harvested after 7-day of growth and were analyzed for protein expression.
PbTPS3 and all three GGDPSs were tagged with HA. The expected sizes for
PbTPS3,
SjGGDPS,
PbGGDPS, and
ScGGDPS are 68.7, 33.0, 32.6, and 39.7 kDa, respectively.
PbTPS2 was tagged with strepII but was not monitored as the antibody did not bind well, while DXS was untagged. Normalization was done by equal loading (10 µg total protein content). All strains expressed the expected proteins. Except for
SjGG/D T1 and T2 (C), appearance of minor potential degradation products was also observed. All
SjGG/D strains were re-run with 40 µg of total protein content to confirm if the absence of the expected bands was due to low expression (D). Strains
SjGG/D T1 and T2 did not express
PbTPS3 or
SjGGDPS In Figure S2, PCR genotyping shows the presence of the
SjGGDPS and
EcDXS gene construct in all three
SjGG/D transformants (T1-T3), suggesting suppression of expression due to metabolic stress. This correlates with the differences in growth seen in the
SjGG/D transformants (B).
. Constructed strains and initial characterization of transformants. (A) The base strain MO was constructed by integration of PbTPS2 and PbTPS3 under the strong Pcpc560 promoter into the NSP1 neutral site. Additional strains were constructed by transforming the base strain with the replicative plasmid pDF-trc containing the Synechococcus sp. JA-3-3Ab GGDPS (SjGG), the Plectranthus barbatus GGDPS (PbGG), the Saccharomyces cerevisiae GGDPS (ScGG), and the Synechococcus sp. JA-3-3Ab GGDPS with an E. coli DXS mutant (SjGG/D). (B) OD730 measurements of strains grown in a photobioreactor. All strains except the empty control strain (n = 1) were grown in triplicate. T1-T3 indicates three independent transformants. SjGG/D transformant 3 (T3) was plotted separately, due to its marked deviation from the other two transformants (T1 + T2). One-way ANOVA of day 7 OD730 values showed no significance. (C) Western blot showing the expression of the HA tagged GGDPSs as well as the HA-tagged PbTPS3. Lanes were normalized by loading 10 µg total protein. (D) Anti-HA Western blot of the SjGG/D transformants (T1–T3) with 40 µg total protein loaded per lane.
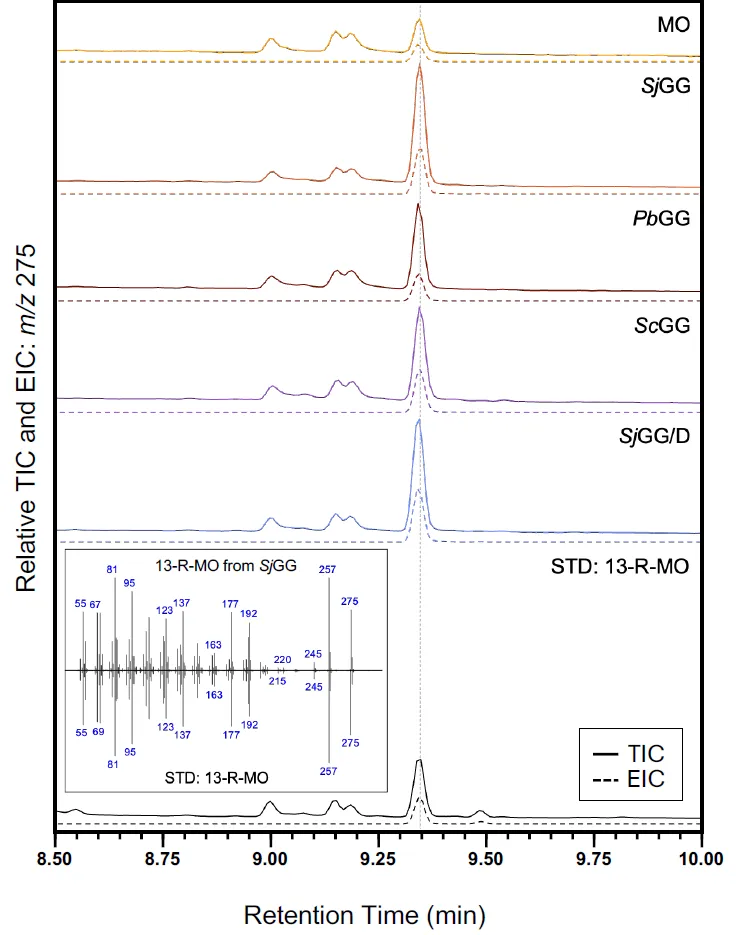
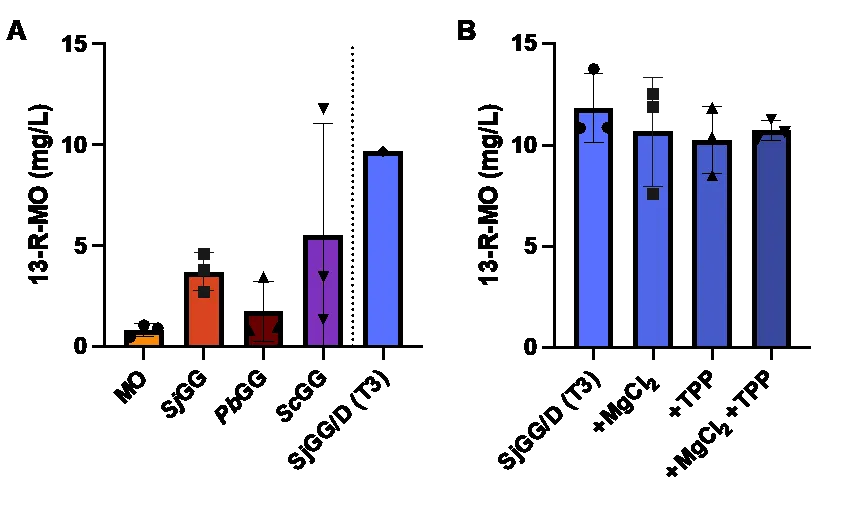
Cells collected from day 7 of the photobioreactor run were extracted and 13-R-MO was quantified by GC-MS (). The base strains (MO), containing
PbTPS2 and
PbTPS3, showed the lowest production of 13-R-MO (A). All other strains, containing GGDPS’, had at least one replicate with greater than 4-fold increase over the average base strain titer. Fold increases ranged from 4.6 to 5.5-fold for
SjGG, from 1-fold to 4.2-fold for
PbGG, from 1.6 to 14-fold for
ScGG, and 11.6 fold for the only
SjGG/D transformant (T3) capable of 13-R-MO production. The highest titers measured was in a
ScGG replicate at 11.8 mg/L, with the second highest measured at 9.7 mg/L in the
SjGG/D T3 replicate. As seen in C, strains carrying the
PbGG and
ScGG construct showed a greater variance in protein expression in comparison with the cyanobacterial GGDPS in strain
SjGG. The
SjGGDPS was therefore used in combination with
EcDXS to create the
SjGG/D strain. The highest titers shift to the
SjGG/D transformant 3 (T3) was grown in triplicate (B).
Due to the toxicity of GGDP, overexpression of a GGDPS in yeast has been reported to result in the secretion of geranylgeraniol or geranyllinalool as detoxification products [
35]. Geranyllinalool is formed by the spontaneous isomerization of geranylgeraniol in an acidic environment. We therefore investigated whether excess flux was directed towards side products. Analysis of 10 mL of supernatant did not detect any geranylgeraniol or geranyllinalool secreted to the media (Figure S3). The single constituent in the chromatograms is the internal standard 1-eicosene.
. Detection of 13-MO by GC–MS analysis. Cell extracts were used to measure 13-R-MO of each engineered strain used in this study by GC-MS and verified using an authentic 13-R-MO standard (bottom). Both the relative total ion chromatograms (TIC) and the extracted ion chromatograms (EIC) at m/z 275 are plotted for each strain and the standard. The mass spectra of the main peaks at around 9.35 min for both SjGG and the standard are also plotted to confirm the correct identical fragmentation patterns.
. 13-R-MO titers from a 7 d photobioreactor experiment. (A) Three transformants of each construct were grown in the photobioreactor and 13-R-MO was quantified on the 7th day. Since PbTPS3 or SjGGDPS was not expressed in SjGG/D T1 and T2, they were excluded from the figure. (B) 13-R-MO titers for SjGG/D (T3), grown in triplicate, supplemented with 400 mM MgCl2, 150 mM TPP, or both. There is no significant difference in production between groups (one-way ANOVA). The titers normalized to OD730 is shown in Figure S4A,B.
Both
PbTPSs and
EcDXS* are dependent on the presence of co-factors to be catalytically active. TPS coordinates Mg
2+ ions while DXS coordinates Mg
2+ and thiamine pyrophosphate (TPP). In
Synechococcus elongatus 7942, production of ethanol was improved by increasing the concentration of essential co-factors in the media [
36]. Accordingly, we supplemented the growth medium for the
SjGG/D (T3) strain with 400 mM MgCl
2, 150 mM TPP, or both. Each experimental condition was carried out in triplicate and the resulting 13-R-MO titers were measured (B). No significant differences were detected between the conditions tested and cofactors are therefore not the limiting factor.
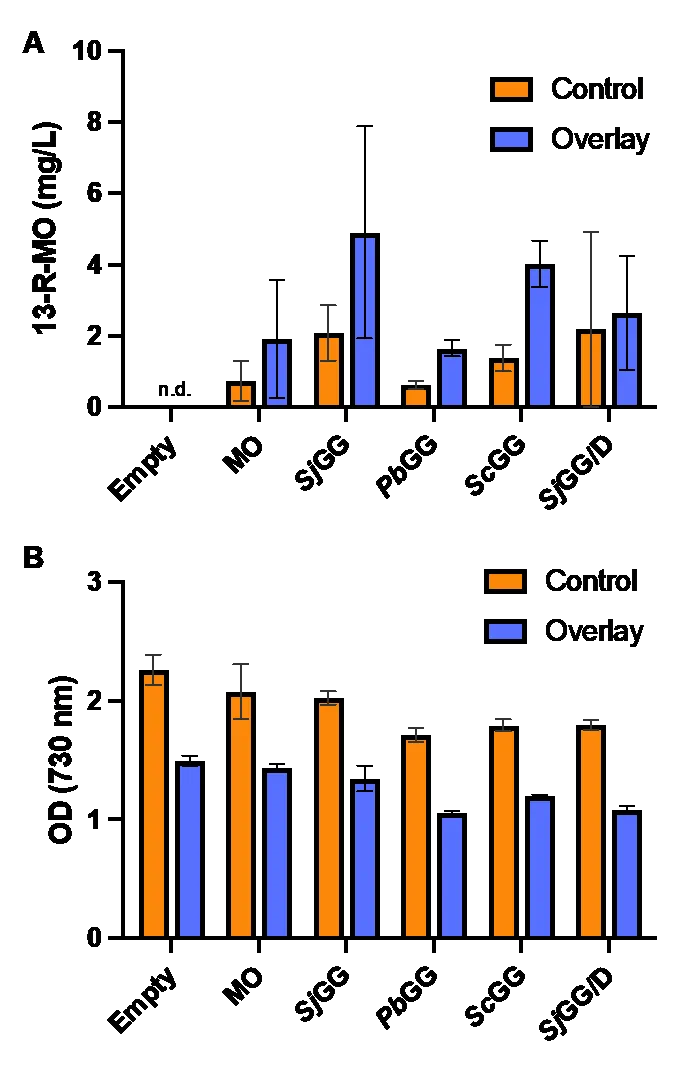
3.5. Adding an Organic Overlay Increases the Production of 13-R-manoyl Oxide
Because 13-R-MO was discovered not to be excreted to the media in any significant amount, the possibility of increasing the production of 13-R-MO by orchestrating an artificial sink for 13-R-MO was examined. In this experimental setup, 40 mL cultures were grown in flasks with and without a 10 mL
N-dodecane overlay. A minor growth inhibition in all strains was observed between the standard and overlay conditions including the “Empty” control strain (B). The introduction of an overlay increased 13-R-MO titers in all conditions (A) by a factor of 2.6 except for the
SjGG/D strain, where no meaningful increase was observed. Cultures without overlay exclusively were found to have 13-R-MO in the cell fraction. Cultures with overlay were quantified solely with the overlay fraction due to negligible amounts in the cell fraction (below quantification limit). When the data are normalized to OD
730 to account for the effects of overlay growth inhibition, normalized titers showed an increase of a factor of 4 for all GGDPS only constructs and an increase of a factor of 9.8 for the the
SjGG/D construct (Figure S4C).
. Shake flask experiment with n-dodecane overlays. (A) Day 7 13-R-MO titers of each culture with and without (i.e. control) N-dodecane overlays. (B) 7-day end point OD730 measurements of strains cultured with and without an N-dodecane overlay.
4. Discussion
The carbon flux through the MEP pathway in the cyanobacterium Synechocystis PCC. 6803 was enhanced by expression of EcDXS
∗ and SjGGDPS. Co-expression of the two diterpenoid synthases PbTPS2 and PbTPS3 from P. barbatus with EcDXS
∗ and SjGGDPS resulted in the production of the diterpenoid 13-R-manoyl oxide at a titer of 9.7 mg/L, an 11-fold increase in titer compared to expression in the base strain. Removal of 13-R-MO from the culture medium by introduction of an n-dodecane overlay increased titers on average 2.6 fold in a separate cultivation system.
4.1. Overexpression of Both DXS and GGDPS Shows Growth Defects
During the course of a 7-day photobioreactor experiment, GGDPS over-expression strains showed no significant growth inhibition compared with the control strain. The single transformant that produced 13-R-MO with the
SjGG/D construct grew much slower compared to the WT (B). It is unlikely that this growth defect is due to altered partitioning of central carbon metabolites to produce 13-R-MO as the resulting 13-R-MO yield was low compared to the carbon equivalents needed to make up for the difference in growth. Instead, toxicity from the 13-R-MO produced may play a role since it is not excreted and thus stored within the cells. Due to its hydrophobic nature, it is plausible that the 13-R-MO produced might partition into the Synechocystis membranes or form droplets and thus perturb the membrane function/fluidity. A previous study on a squalene-producing strain of Synechococcus elongatus PCC 7942 demonstrated squalene droplets forming between distorted thylakoid membranes coupled with a measurable growth defect [
37]. There was also noticeable variation in the protein expression seen by the western blot for
ScGGDPs. While this difference in expression cannot be explained for certain, it could be due to the toxic effects of 13-R-MO.
4.2. Pathway Manipulation and Overlays Increase Titers
The use of an overlay resulted in a 2.6 fold increase in titers when compared to cultures without overlays. The results indicate that removal of 13-R-MO from the cells potentially causes favorable shifts in the reaction equilibrium and/or relieves stress caused by 13-R-MO which in turn increases total amounts of 13-R-MO produced. Our results also show that the use of
n-dodecane as an overlay inhibited growth, however the growth inhibition was not enough to negate the increased production driven by product removal. A previous study demonstrated slight N-dodecane toxicity when used as an overlay for the sequestration of squalene and amorphadiene [
38]. However, the use of hexadecane and tetradecane did not result in growth defects. Comparing the normalized titers between the photobioreactor and flask runs shows that in general the normalized titers in the flasks were much lower (Figure S4C). The source of this discrepancy is likely due to the different culturing conditions, particularly the absence of CO
2 supplementation when grown in flasks. This could suggest that the CO
2 supply is a limiting factor for the production of 13-R-MO. Furthermore, the
SjGG/D was no longer superior to the other constructs under the flask conditions when compared to the photobioreactor conditions.
4.3. Attemted Optimization of 13-R-MO Production
As demonstrated by Velmurugan and Incharoensakdi [
36], the enzymes expressed in this study also use Mg
2+ and TPP as co-factors, but the supplementation of these co-factors did not result in an increase in production. One major difference between our study and Velmurugan and Incharoensakdi [
36] was the length of the culture period and it is possible that if we had extended our culture length, benefits of supplementation of co-factors would have become apparent. Another explanation could be that a portion of our overexpressed enzymes were not properly folded and thus would not benefit from the increased supply of co-factors. DXS is of particular concern as it is known to be prone to aggregation in chloroplasts and needs reactivation to gain activity [
39]. It is unclear whether these pathways are present in cyanobacteria, although homologs can be found. Activation of the refolding pathway could potentially further increase flux through the MEP pathway.
Production of 13-R-MO was also introduced into the model photosynthetic eukaryote green algae
Chlamydomonas reinhardtii resulting in 50 mg/L of 13-R-MO [
40]. The authors used a GGDPS-antibiotic resistance gene-
PbTPS2 fusion construct in a 16:8 light/dark growth cycle with a N-dodecane overlay in order to achieve these titers. Fusion of the terpene synthase with the GGDPS along with the use of a different light regime should be implemented in
Synechocystis in order to see if any further improvements can be made as shown in
C. reinhardtii. A cultivation strategy with a 16:8 light/dark growth cycle was tested with the
Synechocystis strains generated in the current study but it did not improve production (data not shown).
4.4. Perspectives
Manoyl Oxide (13-R-MO) formation is the first committed step in forskolin biosynthesis. This is followed by five oxygenation events on the carbon skeleton catalyzed by three P450s and finally an acetyltransferase completes the biosynthesis of forskolin [
8]. Thus, in future experiments, the MO boosting presented here should be combined with the three P450s and the acetyltransferase. In this context, the P450 enzymes require reducing equivalents such as ferredoxin or NADPH to complete their catalytic cycle. Both reducing equivalents are generated through photosynthesis, making the forskolin pathway an ideal target for heterologous production in cyanobacteria. Previous studies have demonstrated the transfer of electrons from photosystem I (PSI) to cytochrome P450s using endogenous soluble electron donors, eliminating the need for a dedicated reductase [
41,
42,
43,
44,
45,
46]. Among the two photosystems embedded in the thylakoid membranes, PSI stands out as the most stable. Even when subjected to photoinhibitory conditions, it is shielded by alterations in cyclic electron transport around PSI [
47]. Consequently, the light-driven synthesis reliant on electron donation from PSI exhibits remarkable robustness.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/151.
Acknowledgments
We would like to thank Himadri Pakrasi for the plasmid pSL2387.
Author Contributions
L.C.S., P.E.J. and T.G. designed the experiments. L.C.S., N.D., S.L.W., and N.J.B. performed the experiments. L.C.S. analyzed the data and wrote a draft manuscript. L.C.S., P.E.J., N.D., and B.L.M. wrote the final manuscript.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
The authors acknowledge financial support from: (1) Center for Synthetic Biology “bioSYNergy” (UCPH Excellence Program for Interdisciplinary Research), (2) Innovation Fund Denmark (Project no: 12-131834), (3) Novo Nordisk Foundation (NNF13OC0005685; NNF19OC0057634; NNF19OC0054563), (4) VILLUM Foundation (Project no: 13363), (5) VILLUM Center for Plant Plasticity (Project No. VKR023054) and (6) European Research Council Advanced Grant (ERC-2012-ADG_20120314, Project no: 323034).
Declaration of Competing Interest
The authors do not declare any conflicts of interest.
References
-
1.
Nielsen AZ, Mellor SB, Vavitsas K, Wlodarczyk AJ, Gnanasekaran T, Ramos H de Jesus MP, et al. Extending the biosynthetic repertoires of cyanobacteria and chloroplasts.
Plant J. 2016,
87, 87–102.
[Google Scholar]
-
2.
Lassen LM, Zygadlo Nielsen A, Friis Ziersen BE; Gnanasekaran T, Møller BL, Jensen PE. Redirecting photosynthetic electron flow into light-driven synthesis of alternative products including high-value bioactive natural compounds.
ACS Synth. Biol. 2014,
3, 1–12.
[Google Scholar]
-
3.
Sørensen M, Andersen-Ranberg J, Hankamer B, Møller BL. Feature Article: Circular biomanufacturing through harvesting solar energy and CO
2.
Trends Plant Sci. 2022,
27, 655–673.
[Google Scholar]
-
4.
Pichersky E, Raguso RA. Why do plants produce so many terpenoid compounds?
New. Phytol. 2018,
220, 692–702.
[Google Scholar]
-
5.
Cheng AX, Lou YG, Mao YB, Lu S, Wang LJ, Chen XY. Plant terpenoids: biosynthesis and ecological functions.
J. Integr. Plant Biol. 2007,
49, 179–186.
[Google Scholar]
-
6.
Moser S, Pichler H. Identifying and engineering the ideal microbial terpenoid production host.
Appl. Microbiol. Biotechnol. 2019,
103, 5501–5516.
[Google Scholar]
-
7.
Pateraki I, Andersen-Ranberg J, Hamberger Britta, Heskes AM, Martens HJ, Zerbe P, et al. Manoyl oxide (13R), the biosynthetic precursor of forskolin, is synthesized in specialized root cork cells in Coleus forskohlii.
Plant Physiol. 2014,
164, 1222–1236.
[Google Scholar]
-
8.
Pateraki I, Andersen-Ranberg J, Jensen NB, Wubshet SG, Heskes AM, Forman V, et al. Total biosynthesis of the cyclic AMP booster forskolin from Coleus forskohlii.
eLife 2017,
6, e23001.
[Google Scholar]
-
9.
Forman V, Luo D, Lemcke R, Nelson DR. Staerk D, Kampranis S, et al. A gene cluster in Ginkgo biloba encoding for unique multifunctional cytochrome P450s orchestrates key steps in ginkgolide biosynthesis.
Nat. Commun. 2022,
13, 5143.
[Google Scholar]
-
10.
Hansen NL, Kjærulff L, Heck Q, Forman V, Stærk D, Møller BL, et al. Tripterygium wilfordii cytochrome P450s catalyze the key methyl shift and epoxidations in biosynthesis of triptonide.
Nat. Commun. 2022,
13, 5011.
[Google Scholar]
-
11.
Pattanaik B, Lindberg P. Terpenoids and their biosynthesis in cyanobacteria.
Life 2015,
5, 269–293.
[Google Scholar]
-
12.
Bathe U, Tissier A. Cytochrome P450 enzymes: A driving force of plant diterpene diversity.
Phytochemistry 2019,
161, 149–162.
[Google Scholar]
-
13.
Gericke O, Hansen NL, Pedersen GB, Kjaerulff L, Luo D, Staerk D, et al. Nerylneryl diphosphate is the precursor of serrulatane, viscidane and cembrane-type diterpenoids in Eremophila species.
BMC Plant Biol. 2020,
20, 91.
[Google Scholar]
-
14.
Luo D, Callari R, Hamberger B, Wubshet SG, Nielsen MT, Andersen-Ranberg J, et al. Oxidation and cyclization of casbene in the biosynthesis of
Euphorbia factors from mature seeds of Euphorbia lathyris L.
Proc. Natl. Acad. Sci. USA 2016,
113, E5082–E5089.
[Google Scholar]
-
15.
Mills LA, McCormick AJ, Lea-Smith DJ. Current knowledge and recent advances in understanding metabolism of the model cyanobacterium Synechocystis sp. PCC 6803.
Biosci. Rep. 2020,
40, BSR20193325.
[Google Scholar]
-
16.
Rodrigues JS, Lindberg P. Engineering cyanobacteria as Host Organisms for Production of Terpenes and Terpenoids. In Cyanobacteria Biotechnology; Nielsen J, Lee S, Stephanopoulos G, Hudson P, Eds; Wiley: Hoboken, NJ, USA, 2021; pp. 267–300
-
17.
Leonard E, Ajikumar PK, Thayer K, Xiao W-H, Mo JD, Tidor B, et al. Combining metabolic and protein engineering of a terpenoid biosynthetic pathway for overproduction and selectivity control.
Proc. Natl. Acad. Sci. USA 2010,
107, 13654–13659.
[Google Scholar]
-
18.
Tian, S, Wang D, Yang L, Zhang, Z, Liu, L. A systematic review of 1-Deoxy-D-xylulose-5-phosphate synthase in terpenoid biosynthesis in plants.
Plant Growth Regul. 2022,
96, 221–235.
[Google Scholar]
-
19.
Volke DC, Rohwer J, Fischer R, Jennewein S. Investigation of the methylerythritol 4-phosphate pathway for microbial terpenoid production through metabolic control analysis.
Microb. Cell Fact. 2019,
18, 192.
[Google Scholar]
-
20.
Klaus O, Hilgers F, Nakielski A, Hasenklever D, Jaeger K-E, Axmann IM, et al. Engineering phototrophic bacteria for the production of terpenoids.
Curr. Opin. Biotechnol. 2022,
77, 102764.
[Google Scholar]
-
21.
Ajikumar PK, Xiao W-H, Tyo KEJ, Wang Y, Simeon F, Leonard E, et al. Isoprenoid Pathway Optimization for Taxol Precursor Overproduction in
Escherichia coli.
Science 2010,
330, 70–74.
[Google Scholar]
-
22.
Brückner K, Tissier A. High-level diterpene production by transient expression in Nicotiana benthamiana.
Plant Methods 2013,
9, 46.
[Google Scholar]
-
23.
Englund E, Andersen-Ranberg J, Miao R, Hamberger B, Lindberg P. Metabolic Engineering of Synechocystis sp. PCC 6803 for Production of the Plant Diterpenoid Manoyl Oxide.
ACS Synth. Biol. 2015,
4, 1270–1278.
[Google Scholar]
-
24.
Guerrero F, Carbonell V, Cossu M, Correddu D, Jones PR. Ethylene Synthesis and Regulated Expression of Recombinant Protein in Synechocystis sp. PCC 6803.
PLoS ONE 2012,
7, e50470.
[Google Scholar]
-
25.
Stanier RY, Kunisawa R, Mandel M, Cohen-Bazire G. Purification and properties of unicellular blue-green algae (order Chroococcales).
Bacteriol. Rev. 1971,
35, 171–205.
[Google Scholar]
-
26.
Hugueney P, Duchene E, Merdinoglu D. 1-deoxy-D-xylulose 5-phosphate synthase alleles responsible for enhanced terpene biosynthesis. DK2630157T3, 2012.
-
27.
Casella S, Huang F, Mason D, Zhao G-Y, Johnson GN, Mullineaux CW, et al. Dissecting the Native Architecture and Dynamics of Cyanobacterial Photosynthetic Machinery.
Mol. Plant 2017,
10, 1434–1448.
[Google Scholar]
-
28.
Ng AH, Berla BM, Pakrasi HB. Fine-tuning of photoautotrophic protein production by combining promoters and neutral sites in the cyanobacterium Synechocystis sp. strain PCC 6803.
Appl. Environ. Microbiol. 2015,
81, 6857–6863.
[Google Scholar]
-
29.
Zhou J, Zhang H, Meng H, Zhu Y, Bao G, Zhang Y, et al. Discovery of a super-strong promoter enables efficient production of heterologous proteins in cyanobacteria.
Sci. Rep. 2014,
4, 1–6.
[Google Scholar]
-
30.
Forman V, Bjerg-Jensen N, Dyekjær JD, Møller BL, Pateraki I. Engineering of CYP76AH15 can improve activity and specificity towards forskolin biosynthesis in yeast.
Microb. Cell Fact. 2018,
17, 181.
[Google Scholar]
-
31.
Møller BL, Conn EE. The biosynthesis of cyanogenic glucosides in higher plants. Channeling of intermediates in dhurrin biosynthesis by a microsomal system from
Sorghum bicolor (Linn.) Moench.
J. Biol. Chem. 1980,
255, 3049–3056.
[Google Scholar]
-
32.
Laursen T, Borch J, Knudsen C, Bavishi K, Torta F, Martens HJ, et al. Characterization of a dynamic metabolon producing the defense compound dhurrin in sorghum.
Science 2016,
354, 890–893.
[Google Scholar]
-
33.
Jørgensen K, Rasmussen AV, Morant M, Nielsen AH, Bjarnholt N, Zagrobelny M, et al. Metabolon formation and metabolic channeling in the biosynthesis of plant natural products.
Curr. Opin. Plant Biol. 2005,
8, 280–291.
[Google Scholar]
-
34.
Zhou F, Wang CY, Gutensohn M, Jiang L, Zhang P, Zhang D, et al. A recruiting protein of geranylgeranyl diphosphate synthase controls metabolic flux toward chlorophyll biosynthesis in rice.
Proc. Natl. Acad. Sci. USA 2017,
114, 6866–6871.
[Google Scholar]
-
35.
Ignea C, Trikka FA, Nikolaidis AK, Georgantea P, Ioannou E, Loupassaki S, et al. Efficient diterpene production in yeast by engineering Erg20p into a geranylgeranyl diphosphate synthase.
Metab. Eng. 2015,
27, 65–75.
[Google Scholar]
-
36.
Velmurugan R, Incharoensakdi A. Heterologous Expression of Ethanol Synthesis Pathway in Glycogen Deficient Synechococcus elongatus PCC 7942 Resulted in Enhanced Production of Ethanol and Exopolysaccharides.
Front. Plant Sci. 2020,
11, 74.
[Google Scholar]
-
37.
Choi SY, Sim SJ, Choi JI, Woo HM. Identification of small droplets of photosynthetic squalene in engineered Synechococcus elongatus PCC 7942 using TEM and selective fluorescent Nile red analysis.
Lett. Appl. Microbiol. 2018,
66, 523–529.
[Google Scholar]
-
38.
Choi SY, Lee HJ, Choi J, Kim J, Sim SJ, Um Y, et al. Photosynthetic conversion of CO
2 to farnesyl diphosphate-derived phytochemicals (amorpha-4,11-diene and squalene) by engineered cyanobacteria.
Biotechnol. Biofuels 2016,
9, 202.
[Google Scholar]
-
39.
Pulido P, Llamas E, Llorente B, Ventura S, Wright LP, Rodríguez-Concepción M. Specific Hsp100 Chaperones Determine the Fate of the First Enzyme of the Plastidial Isoprenoid Pathway for Either Refolding or Degradation by the Stromal Clp Protease in Arabidopsis.
PLoS Genet. 2016,
12, e1005824.
[Google Scholar]
-
40.
Lauersen KJ, Wichmann J, Baier T, Kampranis SC, Pateraki I, Møller BL, et al. Phototrophic production of heterologous diterpenoids and a hydroxy-functionalized derivative from Chlamydomonas reinhardtii.
Metab. Eng. 2018,
49, 116–127.
[Google Scholar]
-
41.
Mellor SB, Vinde MH, Nielsen AZ, Hanke GT, Abdiaziz K, Roessler MM, et al. Defining optimal electron transfer partners for light-driven cytochrome P450 reactions.
Metab. Eng. 2019,
55, 33–43.
[Google Scholar]
-
42.
Nielsen AZ, Ziersen B, Jensen K, Lassen LM, Olsen CE, Møller BL, et al. Redirecting Photosynthetic Reducing Power toward Bioactive Natural Product Synthesis.
ACS Synth. Biol. 2013,
2, 308–315.
[Google Scholar]
-
43.
Wlodarczyk A, Gnanasekaran T, Nielsen AZ, Zulu NN, Mellor SB, Luckner M, et al.
Metabolic engineering of light-driven cytochrome P450 dependent pathways into
Synechocystis sp. PCC 6803.
Metab. Eng. 2016,
33, 1–11.
[Google Scholar]
-
44.
Berepiki A, Hitchcock A, Moore CM, Bibby TS. Tapping the unused potential of photosynthesis with a heterologous electron sink.
ACS Synth. Biol. 2016,
5, 1369–1375.
[Google Scholar]
-
45.
Berepiki A, Gittins JR, Moore CM, Bibby TS. Rational engineering of photosynthetic electron flux enhances light-powered cytochrome P450 activity.
Synth. Biol. 2018,
3, ysy009.
[Google Scholar]
-
46.
Santos-Merino M, Torrado A, Davis GA, Röttig A, Bibby TS, Kramer DM, et al. Improved photosynthetic capacity and photoprotection via heterologous metabolism engineering in cyanobacteria.
Proc. Natl. Acad. Sci. USA 2020,
118, e2021523118.
[Google Scholar]
-
47.
Teicher HB, Møller BL, Scheller HV. Photoinhibition of Photosystem I in field-grown barley (Hordeum vulgare L.): Induction, recovery and acclimation.
Photosynth. Res. 2000,
64, 53–61.
[Google Scholar]