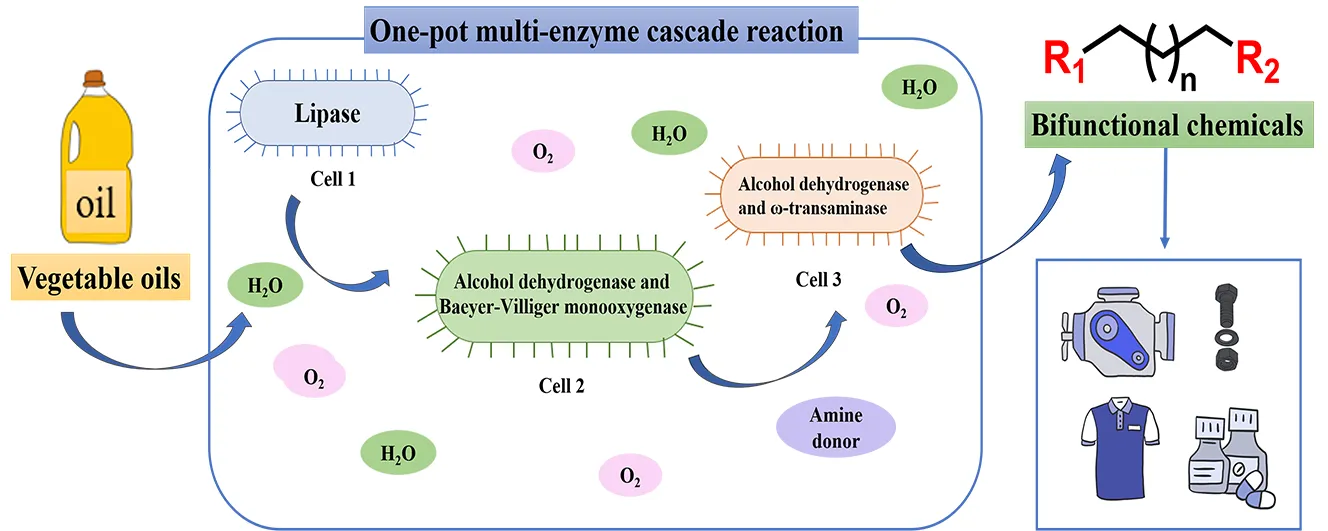
1. Introduction
Bifunctional chemicals are compounds with two functional groups, commonly at the α- and ω- positions of the carbon chain [
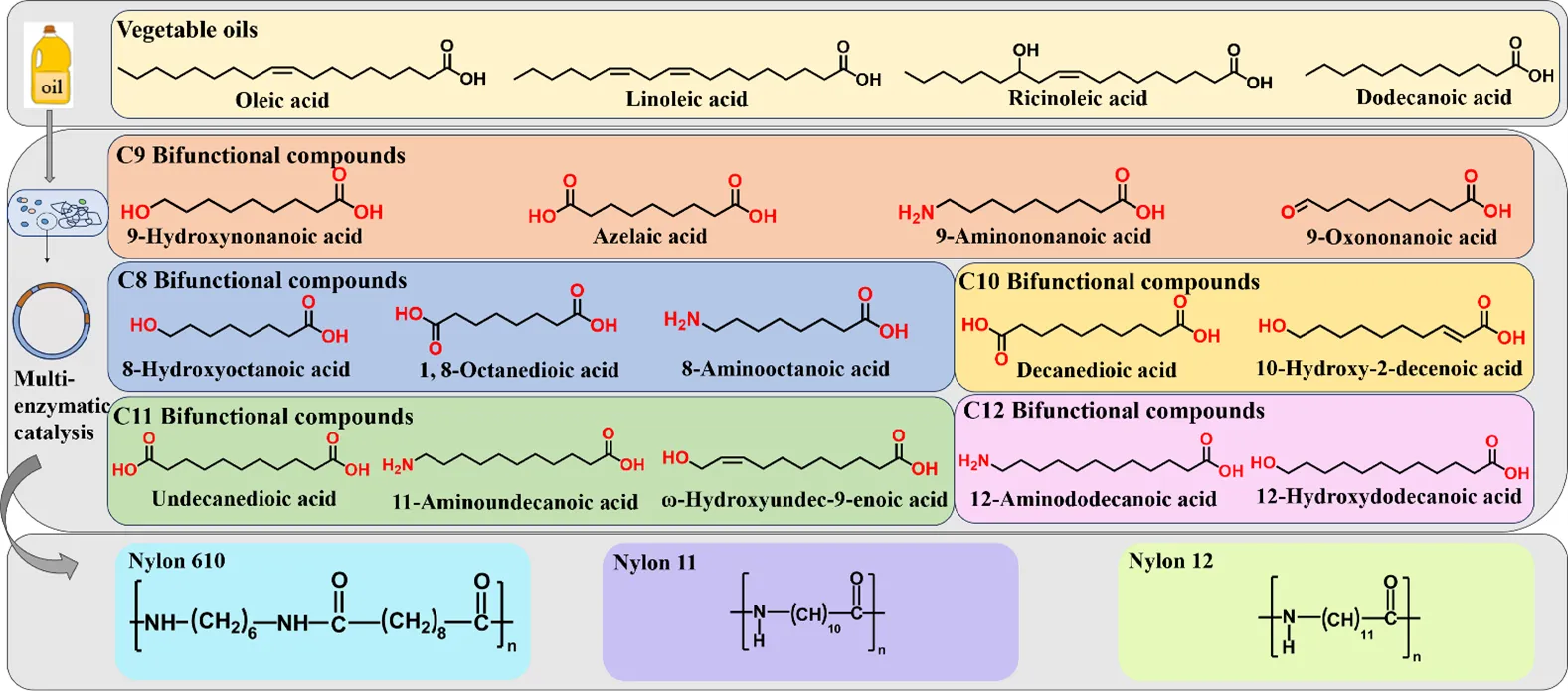
1]. As shown in , long-chain unsaturated fatty acids such as oleic, linoleic and ricinoleic acid, as well as medium-chain saturated fatty acids such as lauric acid, which are abundant in vegetable oils, are suitable substrates for the fermentation of different bifunctional compounds. Although such processes require a variety of enzymes, such as hydratase, alcohol dehydrogenase, monooxygenase, lipase and transaminase, the resulting bifunctional chemicals are valuable precursors for a wide range of plastics, lubricants, fragrances, coatings, fuels and pharmaceuticals [
2,
3]. Nylon is a general term for thermoplastic resins containing repeating amide groups on the main chain of the molecule, and is also the common name for polyamide, which can be biosynthesized or chemically synthesized. According to their molecular structure, the most commonly used nylons are either copolymers of two different kinds of monomers, or homopolymers of a single bifunctional monomer [
4]. Nylons are commonly used plastics with a current global production of more than 8 million tons per annum, which is expected to grow at the rate of 2.2%, reaching up to 10.4 million tons by 2027 [
5]. Nylon is widely used because of its excellent resistance to heat, abrasion and corrosion, while also offering excellent electrical insulation [
6]. In its early days, nylon was mainly used as a textile material, but later it was applied as an engineering plastic to replace steel, iron, copper and other materials. Nylon-based engineering plastics are often used in wear-resistant parts, transmission structures, and household electrical appliances [
7]. Fatty acid-derived diacids, diamines and ω-amino acids are important bio-based nylon monomers. For example, decanedioic acid can be used to synthesize nylon 610, ω-aminoundecanoic acid can be used to synthesize nylon 11, and ω-aminododecanoic acid can be used to synthesize nylon 12. Nylon 11 was the earliest bio-based polyamide product with excellent performance, offering better flexibility, dimensional stability, and impact resistance than comparable plastics with shorter monomer.
. The structural formulae of bifunctional chemicals and their application in the synthesis of nylon.
Previously, bifunctional monomers were mostly produced through chemical methods, but the synthesis process mainly relies on highly polluting and energy-intensive chemical oxidation processes. These chemical processes require the use of large quantities of corrosive chemicals such as concentrated sulfuric and nitric acid, which cause many environmental problems such as greenhouse gas emissions and ozone depletion [
8]. Catalysts based on precious and transition metals were previously commonly used to produce adipic acid [
9], but this method has limitations such as poor catalyst stability, high cost and difficulties in recovery of precious metals. To address these issues, scientists have been exploring new green methods and process technologies for the efficient synthesis of bifunctional compounds. With the development of synthetic biotechnology, in vitro biosynthesis has received increasing attention. It is an area of synthetic biology that focuses on the construction of synthetic multi-enzyme pathways outside the cell to convert cheap substrates into desired products. The in vitro synthetic biology platform has some distinctive advantages, such as high titer, production rate and yield (TRY), as well as easier product separation and simpler process control compared to whole-cell biocatalysis [
10]. Multi-enzyme cascades are becoming a revolutionary tool for the selective synthesis of functional molecules [
11]. One-pot biocatalytic processes offer multiple advantages, such as no need to isolate/purify reaction intermediates, reduced environmental impact, and improved atomic economy [
12]. Bifunctional compounds derived from renewable resources have attracted increasing attention due to their potential to reduce harmful greenhouse gas emissions and circumvent the rising price of fossil raw materials, while also completely avoiding the use of increasingly expensive precious metal catalysts and the need for metal recovery [
13]. Vegetable oils are one of the most important renewable resources and have received widespread attention due to their accessibility, sustainability, non-toxicity, low cost, and potential for chemical modification [
14]. The global annual production of vegetable oils surpassed 200 million tons in 2015, with palm and soybean oil accounting for more than 50% of the total [
15]. Vegetable oils are widely available in nature and are produced in huge quantities. However, the majority of vegetable oils are still used simply for the production of edible oils and feed additives, with only a small proportion used for the synthesis of biodiesel and bio-based chemicals [
16]. Among them, soybean, palm, castor, linseed and sunflower oil are the most commonly used raw materials for the biosynthesis of bifunctional compounds [
17]. Vegetable oils are mainly composed of triglycerides and a small number of diglycerides and monoglycerides. All of them are composed of glycerol and saturated or unsaturated fatty acids [
18]. With the development of biotechnology, the use of biocatalytic processes with good selectivity and mild reaction conditions to achieve highly selective functionalization and refined value addition of fatty acids is receiving increasing attention. Compared to carbohydrates and lignin, fatty acids have a highly reduced aliphatic structure and therefore offer unique advantages for the production of bifunctional chemicals [
19]. With the increasing demand, there is also a need for complete biosynthetic routes capable of producing fine chemicals from inexpensive feedstocks. With the development of synthetic biotechnology, artificially designed multi-enzyme cascades can replace many conventional methods for the production of bifunctional compounds. Low-cost and easily available raw materials can be converted into high value-added products that humans need through a one-pot multi-enzyme cascade under mild and environmentally friendly reaction conditions. This review mainly summarizes the research progress in the production of bifunctional chemicals from vegetable oils using one-pot multi-enzyme cascades in recent years. It also briefly describes the classification of multi-enzyme cascade reactions, the characteristics of different C8–12 bifunctional chemicals, as well as current strategies for enhancing the production of bifunctional compounds from vegetable oils.
2. One-pot Multi-enzyme Cascade Reaction Systems
Enzyme-catalyzed reactions originate from the various metabolic pathways that have evolved in nature for organisms to sustain life processes [
20]. One-pot cascade reactions are combinations of different chemical steps in a single pot without isolating intermediates [
21]. This approach avoids some of the operational steps, thus improving the efficiency of synthesis. One-pot reactions often involve two or more catalysts and have two basic modes of operation [
22]. The first one is to add all catalysts and reagents to the reaction system at the beginning of the reaction, so that the multi-enzyme reaction is carried out simultaneously. The second approach is to add catalysts or substrates for the next step of the reaction system only after the preceding step is completed. The one-pot multi-enzyme cascade reactions have been successfully applied to the synthesis of many compounds, which can be broadly categorized into two enzyme, multi-enzyme, and chemical-enzyme catalyzed one-pot reactions.
2.1. Two-enzyme Catalyzed One-pot Methods
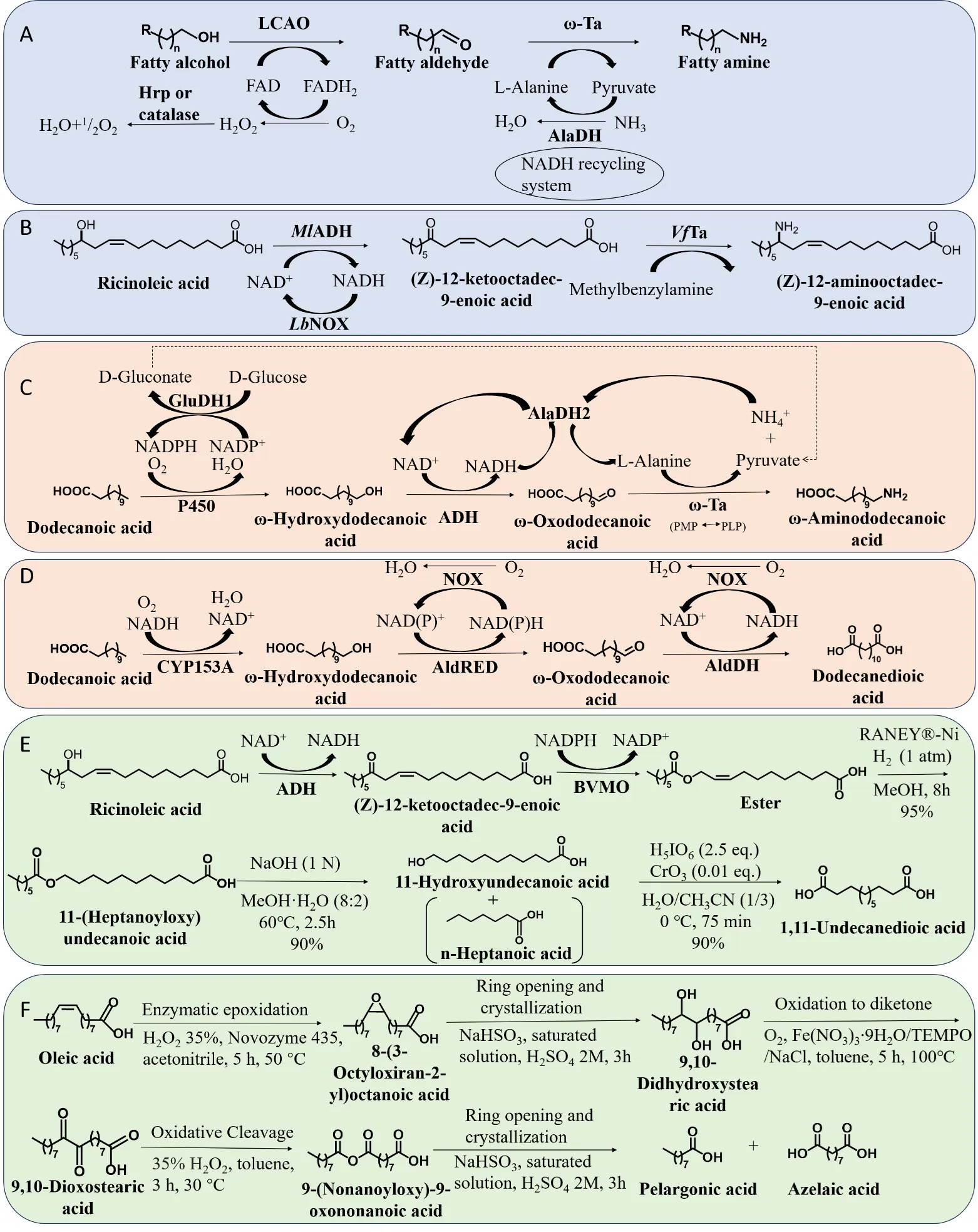
The two-enzyme one-pot method applies two core catalytic reactions catalyzed by different enzymes, but the reaction does not necessarily involve only two enzymes. An example is the construction of a two-enzyme catalytic cascade for fatty amine production from fatty alcohols catalyzed by long-chain alcohol oxidase (LCAO) and ω-transaminase (ω-Ta) [
23]. A total of five enzymes were involved in the reaction. The core components are LCAO responsible for catalyzing the conversion of long-chain fatty alcohols to intermediate aldehydes and ω-Ta responsible for their further conversion to long-chain fatty amines. The other three enzymes were used for the regeneration of cofactors and the amine donor. Horseradish peroxidase (HRP) or catalase was used to scavenge hydrogen peroxide from the system to maintain the enzyme activity. Alanine dehydrogenase (AlaDH) was used to construct the regeneration system for the amino donor D-alanine, while glucose dehydrogenase (GluDH) was used to construct the cofactor regeneration system for AlaDH. Thus, this is still considered a two-enzyme reaction system (A). As shown in B, the two-enzyme one-pot method can also be used for the synthesis of long-chain fatty amines from renewable hydroxy fatty acids. For example, two enzymes were involved in the conversion of ricinoleic acid to (
Z)-12-aminooctadec-9-enoic acid [
24]. The two enzymes were long-chain alcohol dehydrogenase from
Micrococcus luteus (
MlADH) and transaminase variant from
Vibrio fluvialis (
VfTa).
MlADH was used to catalyze the oxidation of hydroxy fatty acids to the corresponding keto fatty acids. Transaminases are generally promiscuous enzymes that can synthesize diverse amines, but most of them do not accept two large substituents next to the carbonyl functionality. To overcome this, 13 variants Ta were screened, and the
VfTa variant was finally selected. Although the NADH oxidase from
Lactobacillus brevis (
LbNOX) was also part of the system, it was only used for the regeneration of NAD
+. Thus, this is still considered a two-enzyme cascade reaction. Under the catalysis of these two key enzymes, the reaction reached a 71% conversion rate. This study contributes to the biosynthesis of long-chain fatty amines with two large substituents next to the amino group.
. Enzymatic cascade reaction systems. (A) Preparation of fatty amines from fatty alcohols by two-enzyme catalyzed one-pot method. (B) Preparation of (Z)-12-aminooctadec-9-enoic acid from ricinoleic acid by two-enzyme catalyzed one-pot method. (C) Preparation of ω-aminododecanoic acid from dodecanoic acid by multi-enzyme catalyzed one-pot method. (D) Preparation of dodecanedioic acid from dodecanoic acid by multi-enzyme catalyzed one-pot method. (E) Preparation of 1,11-undecanedioic acid from ricinoleic acid by chemical-enzyme catalyzed one-pot method. (F) Preparation of pelargonic acid and azelaic acid from oleic acid by chemical-enzyme catalyzed one-pot method.
The multi-enzyme one-pot method was developed from the two-enzyme one-pot method. It can link existing catalytic modules in tandem to form a more complete system and thereby directly generate complex compounds from simple substrates. Self-sufficiency of different modules can also be achieved by constructing cofactors or donor cycling pathways, which is an important part of designing a multi-enzyme one-pot method. ω-aminododecanoic acid (ω-AmDDA) was synthesized by a multi-enzyme catalyzed one-pot reaction using dodecanoic acid (DDA) as the main raw material (C). The reaction was continuously catalyzed by P450 monooxygenase, alcohol dehydrogenase (ADH), and ω-Ta [
25]. By introducing glucose dehydrogenase 1 (GluDH1) into the enzymatic cascade, an additional “bridge” between the artificial pathway and the natural metabolism of the host can be established. By adding AlaDH2 to the reaction system, an internal cycle of L-alanine and NAD(H) was constructed, which increased the yield of ω-AmDDA to 96.5%. A study used aldehyde reductase (AldRED) and acetaldehyde dehydrogenase (AldDH) to optimize the production of dodecanedioic acid (DDDA) from the corresponding ω-hydroxy fatty acids (D) [
26]. The cascade was combined in tandem with a CYP153A-based module for ω-hydroxylation of free fatty acids. Ultimately, this cascade enables the production of DDDA from inexpensive and readily available fatty acids. This reaction uses NADH oxidase (NOX) for cofactor regeneration, which only consumes molecular oxygen and does not require additional substrates to drive product formation. Finally, the authors produced 2.3 mM (0.53 g/L) of DDDA using a one-pot multi-enzyme cascade.
2.3. Chemical-enzyme Catalyzed One-pot Methods
Chemical-enzyme catalyzed one-pot methods can avoid the cytotoxicity of intermediates and the lack of key enzymes in some pathways. From the viewpoint of industrial synthesis, it is attractive to synthesize compounds with high optical purity by one-pot combination of chemical catalysts and enzymes. As shown in E, Jang et al. established a one-pot chemical-enzyme cascade system to synthesize 11-hydroxyundecanoic acid and 1,11-undecanedioic acid [
27]. The first part is the biotransformation pathway, where ricinoleic acid converted into an ester by alcohol dehydrogenase (ADH) and Baeyer-Villiger monooxygenase (BVMO). Then, the ester is hydrolyzed to
N-heptanoic acid and (
Z)-11-hydroxyundec-9-enoic acid. However, both products are toxic to microbial host cells and the stability of BVMO is low, so the concentration of the final product and productivity of the biosynthetic route are low. To solve the problem of product toxicity, a chemical-enzymatic one-pot method can be used. In the second part the authors developed a three-step chemical reaction involving olefin hydrogenation, ester hydrolysis and alcohol oxidation to convert the intermediate ester to undecanedioic acid. The Raney-Ni catalyst was found to be more effective and economical than other metal catalysts. A mixture of ω-hydroxy acids was treated with periodate and chromium trioxide in 75% acetonitrile at 0 °C. After simple extraction with ethyl acetate, the desired product 1,11-undecanedioic acid was obtained by recrystallization from ethyl acetate-hexane. Using a chemical-enzyme catalyzed one-pot method, the authors easily prepared up to 10 g of 1,11-undecanedioic acid in the laboratory. A study described the chemical-enzyme catalyzed one-pot transformation of oleic acid into pelargonic acid and azelaic acid [
28]. As shown in F, the entire process requires four steps, and high chemical purity was eventually achieved at a 44% separation rate. In the first step, the lipase-mediated generation of peroleic acid in the presence of 35% H
2O
2 was employed for the self-epoxidation of the unsaturated acid to the corresponding oxirane derivative. In the second step, the intermediate was converted in situ by acid catalysis to obtain the corresponding 9,10-dihydroxyl fatty acid. The third step was to oxidize the 9,10-dihydroxyl fatty acid to 9,10-dioxostearic acid. In the end, the acid was cracked through the action of 35% H
2O
2 under mild conditions, and no catalyst was required to obtain
N-nonanoic acid and azelaic acid.
Although chemical-enzymatic catalysis has certain advantages, the implementation of chemical-enzyme one-pot systems is challenging compared to pure chemical catalysis or pure enzyme catalysis. Because different catalysts require different reaction conditions and may have different reaction rates or substrate ranges, when enzymes and chemical catalysts are in the same system, mutual inhibition is likely to occur [
29]. With the development of protein engineering, the acceptance of unnatural substrates and tolerance to organic solvents can be improved by modifying the structure of the enzyme [
30].
3. Production of Bifunctional Compounds from Vegetable Oils Using One-pot Multi-enzyme Cascade Reaction Systems
With the increasing awareness of environmental pollution and the need for sustainable development, the global demand for green bifunctional chemicals is gradually increasing [
31]. Oil and fat are the most important renewable raw materials in the chemical industry. Most natural vegetable oils contain unsaturated fatty acids, such as oleic, linoleic and ricinoleic acid. Considering the inherently mild reaction conditions of biocatalysts and the consequent high chemo- and regioselectivity, fatty acid functionalization through biocatalytic manufacturing processes has received much attention [
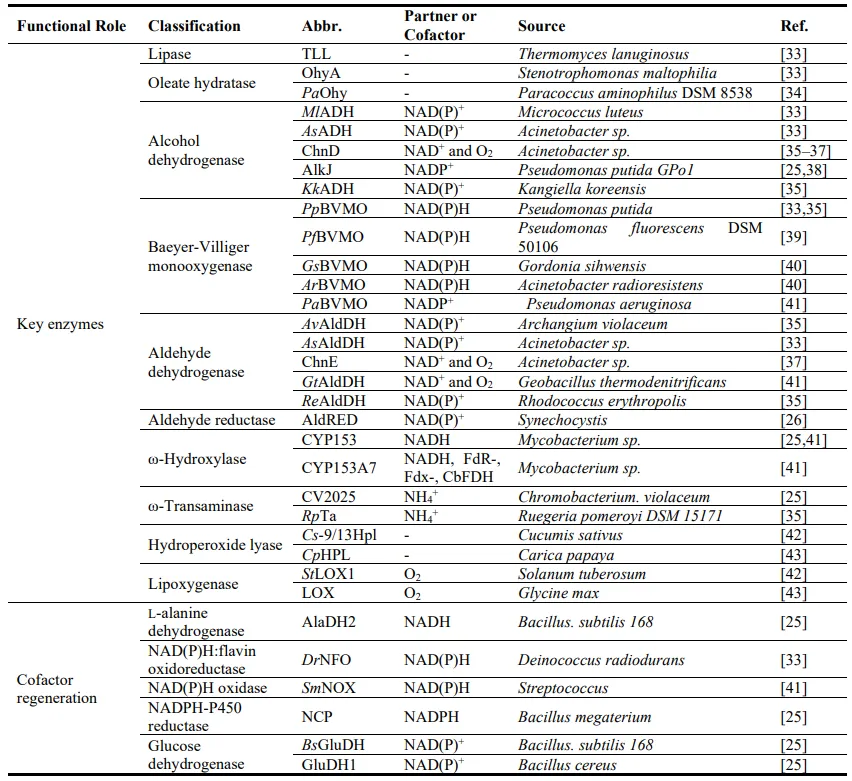
32]. Fat-derived bifunctional monomers such as ω-amino fatty acids, α,ω-dicarboxylic acids and α,ω-diamines are commonly used for synthesizing nylon. shows the key enzymes in the synthesis of C8–C12 bifunctional monomers through multi-enzyme cascade reactions.
. Biosynthetic enzymes involved in the production of bifunctional C8–C12 monomers.
Azelaic acid was previously prepared from oleic acid using hydrogen peroxide and ozone as oxidizing agents [
44]. The ozonolysis route first involves the formation of a primary ozonide via 1,3-cycloaddition. Secondly, the resulting 1,2,4-trioxolane is oxidized to a carboxylic acid under oxidative reaction conditions [
45]. Perhaps unsurprisingly, this process is not environmentally friendly. Therefore, it is important to develop a green multi-enzyme cascade route for the synthesis of C9 chemicals.
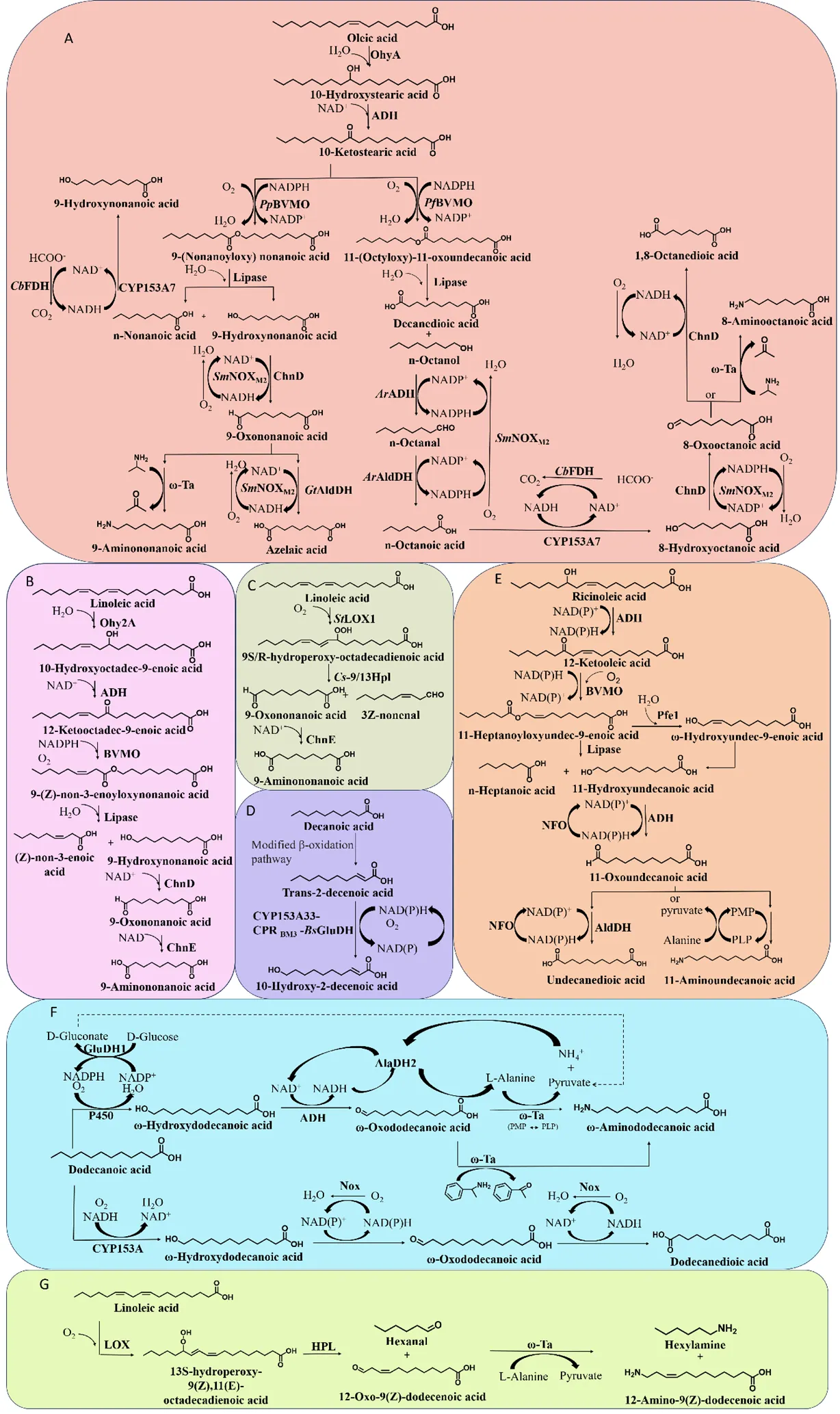
We can sustainably produce C9 bifunctional chemicals from oleic acid, which is difficult to achieve chemically, through a biocatalytic hydration-hydrogenation-oxidation-hydrolysis cascade [
46]. This reaction usually involves four enzymes, oleate hydratase (OhyA), ADH, BVMO and lipase (A). The first step in the conversion of oleic acid into C9 bifunctional compounds is the production of 10-hydroxystearic acid. An example of this is the conversion of oleic acid-rich olive oil into 10-hydroxystearic acid by a strain co-expressing
Thermomyces lanuginosus lipase (TLL) and OhyA from
Stenotrophomonas maltophilia [
37]. However, the activity of both OhyA and BVMO in this reaction system is relatively poor. To overcome the bottleneck in the OhyA step, a study was conducted to clone a number of candidate enzymes based on sequence homology with reported oleic acid hydratases [
34]. Among them, a novel OhyA from
Paracoccus aminophilus DSM 8538 (
PaOhy), was selected for its ability to efficiently catalyze the hydration of oleic acid to 10-hydroxystearic acid.
PaOhy is the most active oleic acid hydratase reported to date. After optimization of reaction conditions, 90 g·L
−1 oleic acid was successfully converted into 10-hydroxystearic acid with a volumetric productivity of 522 g·L
−1·d
−1. In a later study, random mutagenesis of
PaOhy was conducted by error-prone PCR [
47]. These mutants were next evaluated for thermal stability and combined catalytic performance for oleic acid conversion. The triple mutant
PaOhy
F233L/F122L/T15N with significantly higher activity and better thermal stability was finally obtained, showing a significant increase in the catalytic efficiency constant (
Kcat/
Km) from 33 to 119 s
−1·mM
−1.
The next step is the esterification of 10-hydroxystearic acid by ADH and BVMO. BVMO can be used to oxidize ketones to yield esters or lactones. The regioselectivity of BVMO is important in converting asymmetric ketone substrates to produce normal and abnormal esters. The
PpBVMO of
Pseudomonas putida KT2440 favors the insertion of an oxygen atom at the high substitution site of the asymmetric linear aliphatic ketone to generate the “normal” ester and the corresponding ω-hydroxy fatty acids by hydrolysis [
48]. One study utilized MlADH from Micrococcus luteus and
PpBVMO for biotransformation [
33]. Moreover, BVMOs with high regioselectivity and catalytic activity are favorable for the production of medium-chain ω-hydroxy fatty acids from long-chain keto fatty acids. Thirteen genes potentially encoding BVMOs were selected by protein BLAST in the NCBI database using the amino acid sequence of
PpBVMO as the probe [
40]. They were cloned and heterologously expressed in
E. coli BL21 (DE3) to screen enzymes with higher catalytic activity and normal regioselectivity, including 10-ketostearic acid among the substrates. Finally, BVMOs from
Gordonia sihwensis (
GsBVMO) and
Acinetobacter radioresistens (
ArBVMO) were confirmed to have higher catalytic activity than the probe
PpBVMO. Among them,
GsBVMO had the highest catalytic activity and superior regioselectivity (97:3). Determination of the substrate range confirmed that
GsBVMO is more suitable for converting long-chain aliphatic keto acids and medium-chain aliphatic ketones.
Finally, the resulting esters are cleaved by lipase to yield the desired C9 bifunctional chemicals (A). In one study, TLL was used to convert the ester to nonanedioic acid and 9-hydroxynonanoic acid. Through a cascade of hydration-hydrogenation-oxidation-hydrolysis reactions, 110 g·L
−1 of olive oil was converted into 35.2 g·L
−1 of 9-hydroxynonanoic acid [
33]. During the production of 9-hydroxynonanoic acid,
N-nonanoic acid was formed from oleic acid, and
N-non-3-enoic acid was formed from linoleic acid [
37].
N-nonanoic acid can also be transformed into 9-hydroxynonanoic by introducing a hydroxy group. One study used CYP153A7 for this transformation [
36]. The next reaction was to convert 9-hydroxynonanoic acid into azelaic acid using alcohol dehydrogenase (ChnD) and aldehyde dehydrogenase (ChnE). During these reactions, 9-oxonanoic acid and nonanedioic acid are generated. In addition, 9-hydroxynonanoic acid can also be converted into 9-aminononanoic acid using ChnD and ω-TA.
Linoleic acid can be isolated from soybean oil and microalgal oil, which is another renewable source of fatty acids for the production of bifunctional chemicals [
49]. However, most of the reported fatty acid double-bond hydratases exhibit high activity on oleic acid and rather low activity on linoleic acid. Therefore, a study investigated the cloning of novel fatty acid hydratase enzymes with activity towards linoleic and oleic acid [
50]. The putative fatty acid double-bond hydratase OhyA2 (B) was highly active on both oleic and linoleic acid, making it suitable to effectively hydrate unsaturated fatty acids released from olive and soybean oil, thus improving the conversion efficiency of fatty acids into C9 chemicals. Lipoxygenase (LOX) is a non-heme iron dioxygenase that catalyzes the insertion of oxygen into the double bonds of polyunsaturated fatty acids such as linoleic acid. Based on the regioselectivity of oxygen insertion, plant lipoxygenases can be divided into 9- and 13-lipoxygenases [
51]. An in vitro pathway combining 9-lipoxygenase and 9/13-hydroperoxyl cleavage enzymes was capable of converting linoleic acid into 9-oxononanoic acid via a hydroperoxy intermediate (C) [
52]. The resulting one-pot multi-enzyme process yielded 29 mg·L
−1 of azelaic acid in 8 h with 34% substrate conversion and 47% selectivity [
42].
. Production of bifunctional compounds from vegetable oils using the one-pot multi-enzyme cascade reaction systems. (A) Production of C8-C10 bifunctional compounds from oleic acid. (B) Production of C9 bifunctional compounds from linoleic acid. (C) Production of C9 bifunctional compounds from linoleic acid. (D) Production of C10 bifunctional compounds from decanoic acid. (E) Production of C11 bifunctional compounds from ricinoleic acid. (F) Production of C12 bifunctional compounds from dodecanoic acid. (G) Production of C12 bifunctional compounds from linoleic acid.
Traditional processes mostly used chemical cracking of castor oil to prepare decanedioic acid, which caused environmental and safety issues. The advent of new transition metal catalysts has yielded alternatives to harmful oxidants in the synthesis of dicarboxylic acids. A multi-step chemical route for the production of decanedioic acid from cellulose-derived furfural and acetylpropionic acid was successfully developed [
53]. However, high temperatures (180 °C) and pressures (3 MPa H
2) were used in this process, which limits its environmental appeal. As a result, there is great interest and demand for the development of greener and more sustainable alternatives to existing chemical processes.
The major difference in the production of C8 and C10 compounds from oleic acid compared to the production of C9 compounds is that the BVMO used in the third step must have abnormal regioselectivity. Therefore, by using OhyA and different types of BVMOs, the diversity of synthetic pathways can be broadened.
PfBVMO from
Pseudomonas fluorescens DSM 50106 favors the generation of “abnormal” esters and the corresponding α,ω-dicarboxylic acids [
54]. Thus, the regioselectivity of the BVMO is key in determining the type of ester generated and downstream processing [
55]. It was reported that the generation of 10-ketostearic acid from oleic acid produced two types of esters due to the selection of different BVMOs [
46]. Notably, PpBVMO preferentially produces 9-(nonanoyloxy) nonanoic acid, while
PfBVMO produces 11-(octyloxy)-11-oxoundecanoic acid (A). The 9-(nonanoyloxy) nonanoic acid is then converted by lipase to produce an
N-nonanoic acid and a 9-hydroxynonanoic acid. Similarly, 11-(octyloxy)-11-oxoundecanoic acid is converted into N-octanol and decanedioic acid. In one study a variant of MlADH (MlADHM4) was utilized to oxidize 10-hydroxystearic acid to 10-oxostearic acid. BVMO from Pseudomonas aeruginosa (PaBVMO) has abnormal regioselectivity for long-chain keto acids. It was able to insert an oxygen atom near the carbonyl portion of 10-oxostearic acid to give 11-(octyloxy)-11-oxoundecanoic acid [
41]. In the next step, the lipase converts 11-(octyloxy)-11-oxoundecanoic acid to decanedioic acid and
N-octanol, the latter of which is further converted to
N-octanoic acid by ADH, AldDH and NOX. The next reaction is similar to the formation of 9-hydroxynonanoic acid, azelaic acid and 9-aminononanoic acid from
N-nonanoic acid. As shown in A,
N-octanoic acid is hydroxylated by CYP153A to yield 8-hydroxynonanoic acid. Then, 1,8-octanedioic acid is generated via ChnD and AlaDH. Finally, 8-aminooctanoic acid is generated via ChnD and ω-Ta.
Decanoic acid is a by-product of the hydrolysis of coconut oil to produce lauric acid. Notably, decanoic acid can be used to synthesize C10 bifunctional compounds. A study efficiently biosynthesized 10-hydroxy-2-decenoic acid (10-HAD) from decanoic acid using the NAD(P)H-dependent P450 regeneration system in a whole-cell biocatalyst [
56]. The resulting 10-HDA is a hydroxylated α,β-unsaturated medium-chain carboxylic acid with antibacterial, antioxidant, anti-inflammatory, immunomodulatory, and antitumor effects [
57]. In the first step, decanoic acid is converted into trans-2-decenoic acid in
E. coli expressing a modified β-oxidation pathway (D). The second step is the bioconversion of trans-2-decenoic acid to 10-HDA by the P450 enzyme variant supported by NADPH regeneration via
BsGluDH. Ultimately, the yield of 10-HDA reached 486.5 mg·L
−1 by two-step sequential biosynthesis using decanoic acid as substrate.
3.3. C11 Bifunctional Compounds
The main component of castor oil, ricinoleic acid, already contains –COOH, –OH, and –C=C– functional groups, so it is often used as starting material to synthesize C11 bifunctional chemicals. Undecanedioic acid and 11-aminoundecanoic acid were previously produced from castor oil derivatives by cracking at 450 to 500 °C with the aid of a palladium catalyst, sodium hydroxide, hydrochloric acid, hydrogen bromide or ammonia [
58]. However, these chemical processes cause environmental pollution due to their need for strong acids and bases, heavy metal catalysts, organic solvents, and other toxic substances, so new catalytic methods have been explored. A study reported the generation of C11 bifunctional chemicals from ricinoleic acid via a one-pot multi-enzyme cascade reaction [
59]. The engineered biocatalyst was able to produce high concentrations of 11-hydroxyundecanoic acid and undecanedioic acid from ricinoleic acid after multilayer engineering of the
E. coli-based whole-cell biocatalyst and individual enzymes (E). The enzymes involved in the conversion process are
MlADH,
PpBVMO,
KkADH (ADH from
Kangiella koreensis),
ReAldDH (AldDH from
Rhodococcus erythropolis),
DrNFO, ChnD and
RpTa (Ta from
Ruegeria pomeroyi). Ricinoleic acid can be converted into esters by ADH and BVMO inside the cells. The key enzyme BVMO was fused with a hexa-glutamate (E6) tag, which was subsequently mutated to C302L and finally fused to maltose-binding protein (MBP). These modifications not only increased the functional expression level of BVMO in
E. coli, but also increased its structural stability and biocatalytic activity. Esters of 11-heptanoyloxyundec-9-enoic acid are then hydrolyzed into 11-hydroxyundec-9-enoic acid and
N-heptanoic acid by lipases. However, 10 mM
N-heptanoic acid inhibited the C=C reducing activity of the native enzyme of
E. coli, leading to the formation of 11-hydroxyundecylenic acid. This inhibition was mitigated by the addition of adsorbent resins for in situ extraction of
N-heptanoic acid. Next, 11-hydroxyundecanoic acid was converted into undecanedioic acid by
KkADH,
ReAldDH and
DrNFO, while 11-aminoundecanoic acid was converted to undecanedioic acid by ChnD,
DrNFO and
RpTa. Ultimately, 248 mM undecanedioic acid or 232 mM 11-aminoundecanoic acid were obtained from 300 mM ricinoleic acid using the one-pot multi-enzyme cascade system. These concentrations were 500- and 640-foid higher than those produced before engineering, respectively.
ω-hydroxyundec-9-enoic acid (ω-HUA) is another C11 bifunctional chemical with a hydroxyl group at the terminal carbon atom, and is an effective structural unit for the production of polyesters [
31]. ω-HUA has been reported to have high antifungal activity, which makes it useful as an additive in cosmetics, pharmaceuticals and fertilizers [
60]. Recently, the bioproduction of ω-HUA by whole-cell biocatalysts based on
Pseudomonas tropicalis and
E. coli has enabled the development of safe and environmentally friendly processes [
61]. The bioconversion of ricinoleic acid into ω-HUA by recombinant
E. coli requires the expression of three heterologous enzymes: ADH, BVMO, and lipase Pfe1 (E). ADH first oxidized ricinoleic acid into 12-ketooleic acid, which was then oxidized to yield (
E)-11-(heptanoyloxy)undec-9-enoic acid undecanoate (11-HOUA) by BVMO using O
2 and NADPH. Subsequently, Pfe1 hydrolyzed 11-HOUA into ω-HUA and heptanoic acid [
27]. Various strategies have been developed for the bulk production of ω-HUA from ricinoleic acid using recombinant
E. coli systems. Jang et al. [
62] obtained 4.0 g·L
−1 of ω-HUA from ricinoleic acid using whole-cell biotransformation with recombinant
E. coli, which achieved a conversion rate of more than 80%. Although the resting cell production of 11-HOUA (intermediate of ω-HUA) by recombinant
E. coli showed a high titer and productivity, the final concentration and productivity of ω-HUA remained low due to the toxicity and side effects of 11-HOUA. To overcome this obstacle, adsorbent resin can be added to alleviate the inhibition. To promote the regeneration of cofactors required for ADH and BVMO reactions as well as to maintain cell viability, Cho et al. applied continuous and intermittent addition of glucose and glycerol during the biotransformation phase of ricinoleic acid [
61]. Enhanced interaction of Pfe1 with 11-HOUA and cofactor regeneration synergistically improved the efficiency of ricinoleic acid biotransformation into ω-HUA by combining PelB attachment to Pfe1 and a supplementation batching strategy [
63]. The yield of ω-HUA was increased two-fold compared to the control strain.
3.4. C12 Bifunctional Compounds
Nylon 12 monomer was previously synthesized chemically from butadiene in the form of ω-12 lactam through a number of steps such as trimerization, catalytic hydrogenation, oxidation, ketonization, oxidization and Beckman rearrangement [
64]. However, this method has many disadvantages. Firstly, butadiene is derived from petroleum and its price is subject to large fluctuations in the petroleum market. Secondly, toxic and corrosive materials such as benzene are used in the synthesis process, and a large amount of waste is generated, which puts a great pressure on the environment. Thus, there is growing pressure to develop a green route for the production of nylon 12 monomer.
There are two ways to synthesize nylon monomers, one involves the cleavage of fatty acids, and the other involves P450-catalysed ω-oxidation. Lauric acid, also known as dodecanoic acid (DDA), is a saturated fatty acid commonly used to synthesize nylon 12 monomer. Different from the biosynthesis of C8–10 bifunctional chemicals from C18 unsaturated fatty acids, the first step of ω-AmDDA synthesis requires hydroxylation at the ω-terminus (F). A study developed a multi-enzyme cascade reaction consisting of CYP153A, alcohol dehydrogenase (AlkJ) and ω-TA for the production of ω-AmDDA from DDA [
65]. Regioselective ω-hydroxylation of 1 mM DDA was achieved using whole-cell biocatalysts co-expressing CYP153A and ferric oxide reductase with almost complete conversion (>99%). When a whole-cell biocatalyst co-expressing AlkJ and ω-TA was used for sequential biotransformation of ω-hydroxydodecanoic acid (ω-OHDDA), 1.8 mM ω-OHDDA was converted into ω-AmDDA, with a conversion rate of 87% after 3 h. Finally, when a one-pot reaction using 2 mM DDA was performed using both whole-cell systems, 0.6 mM ω-AmDDA was produced after 5 h of reaction.
To construct a biosynthetic pathway for the production of ω-aminododecanoic acid (ω-AmDDA) from lauric acid, enzymes catalyzing the terminal hydroxylation of DDA, oxidation of ω-hydroxydodecanoic acid (ω-OHDDA), and amination of ω-oxododecanoic acid (ω-ODDA) need to be screened and sequentially assembled. CYP153A
G307A from
Marinobacter aquaeolei is reported to catalyze highly regioselective ω-hydroxylation of medium- and long-chain fatty acids [
66]. The flavonoid P450 BM3 from
Bacillus megaterium is a naturally occurring fusion protein composed of a fatty acid hydroxylase domain and a mammalian-like diflavin NADPH-P450 reductase (NCP) domain [
67]. It can be used to catalyze the proximal hydroxylation of long-chain fatty acids. The hydroxylation efficiency of CYP153A
G307A increased after fusion with NCP [
68]. A study introduced the chimeric fusion protein CYP153A-NCP into
E. coli to construct a ω-AmDDA synthesis pathway [
25]. The reaction was continuously catalyzed by CYP153A-NCP, BsADH
C257L and CV2025, yielding 1.9 mM ω-OHDDA from 2.5 mM DDA after 2 h of reaction in the presence of 5.0 mM NADPH, whereby no by-product peaks were detected. The authors constructed a biosynthetic cascade system for ω-AmDDA production in
E. coli BL21(DE3), assembling the three enzymes into two catalytic modules. Overexpression of glucose dehydrogenase (GluDH1) accelerated glucose metabolism and increased NADPH regeneration. This was combined with expansion of the pyruvate pool used as a substrate for AlaDH2 to produce the co-substrate L-alanine. They also introduced AlaDH2 to construct a cycle of NAD(H) and L-alanine, thereby achieving cofactor self-sufficiency and solving the amino donor problem. The authors also blocked the β-oxidation pathway, introduced the outer membrane protein AlkL to enhance the product yield, was well as increasing the PLP supply by introducing a heterologous ribulose 5-phosphate (R5P)-dependent PLP synthesis pathway. Finally, efficient synthesis of C12 bifunctional compounds from DDA was achieved, reaching a ω-AmDDA yield of up to 96.5% via whole-cell biocatalysis.
In addition to the synthesis of nylon 12 monomer from lauric acid, a study developed the synthesis of ω-AmDDA from linoleic acid [
43] (G). Seven ω-Tas were first cloned for expression in
E. coli and verified for activity in long-chain aldehydes, with the ω-Ta from
Aquitalea denitrificans having the highest activity. Next, a multi-enzyme cascade comprising lipoxygenase (LOX), hydroperoxide lyase (HPL), ω-Ta and lactate dehydrogenase was established. Ultimately, up to 12% of linoleic acid was converted into 12-aminododecenoic acid.
4. Novel Strategies for Enhancing the Production of Bifunctional Compounds from Vegetable Oils
Compared to traditional chemical catalytic processes, the multi-enzyme cascade processes were at lower production levels. As a result, recent studies were devoted to developing multi-level strategies to improve the efficiency, economy, and sustainability of multi-enzyme cascades. Sequential cofactor-dependent oxidation-reduction reactions are involved in the production of bifunctional chemicals from fatty acids. To avoid large additions of expensive cofactors, several cofactor regeneration strategies were developed. In addition, protein engineering strategies were applied to improve enzyme stability and catalytic performance. Whole-cell catalysis has been widely applied in the biosynthesis of fatty acid-derived bifunctional chemicals due to its elimination of tedious enzyme purification processes. Inhibition of the competing β-oxidation pathway, the improvement of mass transport efficiency across membranes, and the enhancement of cellular robustness are effective optimization strategies for whole-cell catalytic systems.
4.1. Cofactor Regeneration
Oxidoreductases account for approximately 25% of all known enzymes, and are responsible for intermolecular electron transfer [
69]. However, many enzymatic redox applications are limited by the dependence on cofactors, such as nicotinamide adenine dinucleotide (NADH) and its phosphorylated form (NADPH). Bioconversions that employ oxidoreductases consume a stoichiometric amount of cofactors while synthesizing the product. However, these cofactors are often expensive and have low stability, making the application of large amounts of cofactors unfeasible from a techno-economic standpoint. To solve this problem, the construction of cofactor regeneration systems has been widely studied in recent years. Compared with the direct addition of expensive cofactors, cofactor regeneration systems that can be integrated into multi-enzyme cascades have obvious advantages. The construction of cofactor regeneration systems is not only inexpensive but also has high atom economy. A study reported that by introducing
DrNFO into
E. coli expressing
KkADH and
GtAldDH followed by optimizing the relative expression of the three genes for α,ω-dicarboxylic acid production, an engineered biocatalyst with a cofactor regeneration system was successfully constructed, which was 1.6-fold more active than
E. coli without cofactor regeneration [
70]. NAD(P)
+-dependent ADH and AldDH can form a cofactor regeneration system by coupling with NAD(P)H dehydrogenase (NOX). Catalyst II was constructed for the conversion of 11-hydroxyundecanoic acid into undecanedioic acid by tandem coupling of
KkADH,
ReAldDH and
DrNFO. Catalyst III was constructed for the conversion of 11-hydroxyundecanoic acid into ω-aminoundecanoic acid by combining ChnD,
DrNFO and
RpTa in series. These two cofactor-regeneration methods can be used to produce C11 bifunctional chemicals. The final productivity of biocatalyst II with the cofactor regeneration system was 1.4 times higher than that of
E. coli expressing
KkADH and
ReAldDH alone. The specific activity of biocatalyst III with the cofactor regeneration system was 2.0-fold higher than that of
E. coli expressing ChnD and
RpTa without the cofactor regeneration system [
35]. In the production of C12 bifunctional compounds, Ye et al. [
25] introduced AlaDH2 to construct an L-alanine—NAD(H) cycle, which bridged the cofactor requirement gap of the ω-hydroxydodecanoic acid to ω-aminododecanoic acid pathway. The inexpensive ammonium source for this pathway can also be used to convert endogenous pyruvate into L-alanine. A thermotolerant NAD
+-dependent ADH from
Bacillus thermophilus was used for the biocatalytic synthesis of ω-oxododecanoic acid methyl ester, a key intermediate for the production of C12 bifunctional compounds, from the corresponding long-chain alcohol [
71].
A study investigated the modification of cofactor specificity in NAD
+-dependent
MlADH to convert long-chain fatty acids into industrially relevant bifunctional C9 chemicals [
72]. This was first done by characterizing the biochemical properties of
MlADH to identify engineering targets that alter its cofactor specificity. The corresponding mutants were then constructed based on substrate binding models and observed structural features. The mutants of
MlADH were tested in biotransformation reactions together with
PpBVMO. The
MlADH variants were found to be able to efficiently catalyze the biotransformation using NADP+ as the sole cofactor.
MlADH
D37S/A38R/V39S/T15I was successfully coupled to PpBVMO for the redox biotransformation of C18 fatty acids into C9 chemicals, resulting in an 1800-fold increase of NADP+ utilization efficiency.
4.2. Protein Engineering
To develop efficient whole-cell biocatalysts, it is necessary to optimize the gene expression levels of the enzymes in the biotransformation pathway and thereby increase the activity of the kinetically limiting enzymes involved [
73]. An example of such a system is the engineering of a BVMO-based
E. coli biocatalyst for the oxidative cleavage of long-chain fatty acids to produce medium-chain bifunctional chemicals, especially ω-hydroxy carboxylic and α,ω-dicarboxylic acids. The instability of BVMO under oxidative and thermal stresses as well as its limited specific activity remain major barriers to the industrial application of these biocatalysts. For example, BVMO has been reported to be sensitive to oxidative stress [
74]. Cysteine and methionine are amino acids that contain sulfur groups and rare highly sensitive to reactive oxygen species (ROS). The interaction of cysteine with hydrogen peroxide and other oxidants generates sulfonic acid intermediates and sulfites [
75,
76]. Notably,
PpBVMO contains 12 Met and 8 Cys residues. Among them, Cys302 is located approximately 8.7 A inside the active site and may be expose to intrinsic ROS [
77]. Therefore, many studies used Cys302 as an engineering target for protein modification. Woo et al. [
78] replaced Cys302 with Leu, which made the engineered enzyme (i.e.,
E6BVMO
C302L) more stable to oxidation and thermal stress. The catalytic activity of the E. coli whole-cell biocatalyst based on
E6BVMO
C302L was also higher than that of the biocatalyst based on E6BVMO. Maltose-binding protein (MBP) can enhance the functional expression and structural stability of fusion proteins. When MBP was fused to
E6BVMO
C302L, its catalytic efficiency (
Kcat/
Km) was 1.8-fold and 2.2-fold higher than that of
E6BVMO
C302L and E6BVMO, respectively [
35]. These results suggest that MBP, as a solubility tag of considerable size, contributes not only to the stability of BVMO but also to its catalytic efficiency. The increased catalytic efficiency could be attributed to the increased stability of BVMO, as the enzyme has very low stability under normal reaction conditions. The whole-cell biocatalyst constructed by co-expressing MBP-
E6BVMO
C302L with MlADH was able to convert 200 mM ricinoleic acid into 152 mM 11-heptanoyloxyundec-9-enoic acid within 12 h. This was 1.3-fold higher than the
E. coli-based biocatalyst expressing
E6BVMO
C302L. H
2O
2 is involved in several enzyme-catalyzed reactions as an oxidant or toxic by-product. A study predicted the enzymatic channels of several BVMOs by using CAVER and/or Protein Energy Landscape Exploration software [
79]. Amino acid residues susceptible to oxidation by H
2O
2 (e.g., methionine and tyrosine) and those located in the vicinity of the predicted H
2O
2 migration pathway were replaced with less reactive or inert amino acids (e.g., leucine and isoleucine). Ultimately, H
2O
2-resistant enzyme variants were designed to improve the tolerance of BVMOs to H
2O
2. PpBVMO
C302L/M340L/M380L produced a 2-fold higher titer of 9-(nonanoyloxy) nonanoic acid in cascade fatty acid biotransformation. A study identified a novel BVMOs from
Gordonia sihwensis (
GsBVMO) with higher normal regioselectivity [
40]. Moreover, the specificity constant (
Kcat/
Km) of its engineered variant
GsBVMO
C308L was increased 8.9-fold compared to the parental enzyme. Using
GsBVMO
C308L as an effective biocatalyst, 10-ketostearic acid was converted into 9-(nonanoyloxy)nonanoic acid with higher yield and specificity.
4.3. Blocking the β-oxidation Pathway
Fatty acid β-oxidation is the main pathway of fatty acid degradation. Fatty acid substrates can be metabolized by the endogenous β-oxidation pathway upon entry into
E. coli cells, which may affect the final yield of bifunctional compounds. For example, DDA can be degraded via the β-oxidation pathway, leading to a decrease of ω-AmDDA acid production [
80]. To increase the accumulation of fatty acids, it is necessary to block the β-oxidation of fatty acids. Extracellular long-chain fatty acids are transported across the membrane into the cytoplasm by the action of the outer membrane proteins FadL and acyl CoA synthase (FadD). First acyl CoA dehydrogenase (FadE) catalyzes the generation of enoyl CoA from lipoyl CoA. Hydroxyacyl CoA is then converted to enoyl CoA through hydration by enoyl CoA hydratase and oxidation by β-hydroxyacyl CoA dehydrogenase. Next, β-keto-lipoyl CoA thiolase hydrolyzes the β-keto-lipoyl CoA disulfide bond to produce acetyl CoA and lipoyl CoA, resulting in the shortening of the hydrocarbon chain by two carbon atoms. FadD and FadE are the key enzymes associated with β-oxidation [
81]. They respectively catalyze the first and second steps of β-oxidation, and deletion of the
fadD or
fadE gene can block β-oxidation and significantly enhance fatty acid accumulation [
82]. Considering the Δ
fadE strains still synthesizes acyl CoA from fatty acids, the deletion of
fadD may be more effective in preventing fatty acid degradation [
80]. In a study on the production of C12 bifunctional compounds,
E. coli BL21(DE3)-Δ
fadD was constructed using a CRISPR/Cas9 based method. After 20 h of biotransformation with 2.5 mM DDA, the yield of ω-AmDDA increased to 78.8%, resulting in a product titer of 1.97 mM [
25]. In a study on the synthesis of nylon 12 monomer, Ahsan et al. also knocked out the
fadD gene to enhance the production of ω-AmDDA [
38]. The conversion of 2 mM DDA yielded 0.6 mM ω-AmDDA after 5 h of reaction.
4.4. Improving Mass Transport
Fatty acid transport across cell membranes into the cytoplasm of microbial cells limits substrate availability and compromises membrane integrity, which in turn reduces cell viability and biotransformation capacity. The outer layer of the outer membrane of
E. coli is mainly composed of lipopolysaccharide, which affects the whole-cell biotransformation of hydrophobic molecules by slowing down their transport to intracellular enzymes through the cell membrane. Moreover, hydrophobic substrates are often toxic to microorganisms because fatty acid passage disrupts cell membranes and lowers intracellular pH [
83]. Mass transport can be enhanced by engineering the native cellular machinery. FadL protein in the outer membrane of
E. coli binds extracellular long-chain fatty acids with relatively high affinity. Several charged amino acid residues within FadL participate in the interaction with charged fatty acid groups. One study showed that the whole-cell biotransformation rate of long-chain fatty acids was significantly affected by the amount of FadL in the cell membrane [
84]. With the increase of FadL expression, the specific whole-cell biotransformation rate of 10-hydroxyoctadecane acid increased more than 5-fold. However, when FadL was overexpressed, the rate of multi-step biotransformation of ricinoleic acid in an engineered whole-cell biocatalyst was significantly reduced. This indicates that overexpression of FadL seems to have a negative impact on the balanced expression of intermediate enzymes in the cytoplasm. Therefore, the balanced expression of FadL is conducive to the biotransformation of long-chain fatty acids. In addition, AlkL, an outer membrane protein of
Pseudomonas putida GPo1, was found to enhance the uptake of hydrophobic substrates by
E. coli, and effective ω-oxo-functionalization of fatty acid methyl esters was achieved after fine-tuning the expression level of AlkL [
85]. The final methyl dodecanoate oxidizing activity was increased 62-fold to 87 U·g·cd
−1. Another study developed a whole-cell biocatalyst that can form caveolae-like structures to promote substrate endocytosis [
86]. Expression of caveolin-1 in
E. coli increased the rate of fatty acid transport and the intracellular fatty acid concentration, complementing substrate endocytosis. In addition, fatty acid endocytosis attenuated substrate cytotoxicity in
E. coli. The engineered
E. coli cells that formed caveolae-like structures were able to convert 15 mM ricinoleic acid into 9.6 mM (
Z)-11-(heptanoyloxy)undec-9-enoic acid, resulting in 2-fold higher bioconversion than the control strain. Moreover, when 30 mM ricinoleic acid was added to the whole-cell biotransformation, control cells produced 7 mM (
Z)-11-(heptanoyloxy)undec-9-enoic acid, whereas the engineered cells with caveolae-like structures produced 13 to 17 mM (
Z)-11-(heptanoyloxy)undec-9-enoic acid. Thus, the endocytosing cells can more rapidly incorporate fatty acid substrates than control cells, so that fatty acid-induced cytotoxicity was mitigated and the performance of the biocatalyst was improved. Furthermore, the intracellular fatty acid concentration was kept high, which contributed to more efficient bioconversion of fatty acids and vegetable oils into value-added products. A team developed a novel synthetic multi-enzyme complex for in vitro cascade transformation of renewable triacylglycerols (TAGs) into fatty alcohols based on the
E. coli-INP-cohesin-dockerin surface assembly system [
87]. Three customized enzymes, lipase (Lip), carboxylate reductase (CR), and aldehyde reductase (AldRED), were co-assembled on the surface of
E. coli cells. The cell-Lip-CR-AldRED complex and its non-complexed counterpart in the free enzyme mixture exhibited
Kcat/
Km values of 1.52 × 10
5 and 3.63 × 10
3 M
−1·s
−1, respectively. The biocatalytic performance of the cell-based multi-enzyme complexes was greatly improved compared to the mixture of three free enzymes, as evidenced by the higher
Kcat/
KM values. The multi-enzyme complex formed a substrate channel, achieving 73% conversion. Moreover, the complex showed high stability when exposed to increased temperatures, extreme pH, polar organic solvents and high substrate concentrations. Finally, the complex also exhibited good recoverability in continuous biotransformation.
4.5. Enhancing Strain Tolerance
E. coli BL21 (DE3) is widely used for the expression of various recombinant proteins. However, the overexpression of recombinant proteins usually imposes a metabolic burden on the host, especially in the case of toxic proteins. To overcome this, a dynamic equilibrium-based production system was constructed to precisely reduce the host burden and increase recombinant protein production. This system can effectively reduce optimization costs and is suitable for rapid development of various types of expressing hosts [
88]. The system consists of three modules, including basic growth-related genes (rRNA, RNAP core enzyme, sigma factor), prediction and overexpression of key proteins using the enzyme-constrained model ec_iECBD_1354, and dynamic regulation of the intensity of expression of growth-related and key proteins based on load-driven promoters. Using this system, a number of high-load proteins, including autolysins and membrane proteins of
E. coli, can be produced with increased yields. Among them, cytosine transporter protein showed the most dramatic improvement, with a 4.02-fold increase in yield compared to the parental strain. The glutamic acid-dependent acid resistance (GDAR) system is responsible for the removal of intracellular protons by catalytic glutamate decarboxylation and is inactive in
E. coli BL21(DE3). The
rcsB and
dsrA genes encode regulatory factors. RcsB acts as an important component of GDAR regulation by forming a heterodimer with the GDAR transcription factor. DsrA, on the other hand, is a small RNA involved in stress-induced metabolism in
E. coli. Activation of the GDAR system in this strain by overexpression of the
rcsB and
dsrA genes resulted in increased resistance not only to HCl but also to
N-heptanoic acid [
89]. Furthermore, activation of the GDAR system allowed recombinant
E. coli BL21 (DE3) expressing MlADH and PpBVMO to withstand product concentrations above 60% during the bioconversion of ricinoleic acid to
N-heptanoic and 11-hydroxyundecan-9-dioic acid.
The low solubility of the oil in water leads to inefficient enzyme catalysis and low product titers, limiting the biosynthesis of fatty acid-derived chemicals. Advanced strategies such as artificial spore were implied to enhance the whole-cell catalysis of hydrophobic organic substrate. Compared to traditional whole-cell catalysis, artificial spores are encapsulated living cells that are not only robust but also metabolically active and thus have great biosynthetic potential. A study presented a simple and versatile method to prepare artificial spores based on live
E. coli cells by biocompatible polydopamine coating and demonstrated their challenging interfacial catalytic ability [
90]. The biocompatibility of the spore formation process was tested in the study. Compared to uncoated
E. coli cells, artificial spores had a similar growth capacity, although they showed a delayed growth curve. This suggests that artificial spore formation is a gentle process that allows cell division even after coating. Moreover, the artificial spores maintained almost twice the enzyme activity of the natural cells following 2 h of UV irradiation. The enzyme activity of the artificial spores was also somewhat higher when the cells were treated with aqueous-toluene medium. In addition, the spore shell protected the cells from heat and acetonitrile solvent. These results demonstrate the ability of artificial spore coats to protect cells against a wide range of environmental stresses. Emulsions have been prepared by mixing artificial spores with organic solvents, and it was found that the artificial spores were completely aggregated in the outer layer of the emulsion surface, resulting in micrometer-sized pores. This tightly packed porous structure indicates that there was a sufficient number of cells in the emulsion and that these pores facilitate substrate entry into the structure, which should favor rapid mass transfer during catalysis. In addition, the cells recovered from the emulsion remain metabolically active. The protective coating enabled the living cells to stabilize aqueous organic emulsions, leading to effective interfacial biocatalysis of simple reactions and multi-enzyme cascades. Thus, this artificial spore-based platform technology is expected to lay the foundation for the next generation of whole-cell catalysis involving oil substrates.
5. Conclusions
Value-added bifunctional compounds can be produced from vegetable oils or related fatty acids by microbial enzymatic conversion reactions. These bifunctional compounds can be used in the production of cosmetics, plastics, industrial materials, etc., and have bright prospects for applied research and development. In the production of C8–C10 bifunctional compounds, the main catalytic components used are oleate hydratase, alcohol dehydrogenase, Baeyer-Villiger monooxygenase, esterase, P450 hydroxylase, and transaminase, etc. However, most biocatalytic reactions are still in the laboratory research stage and cannot achieve industrial scale production presently. This is mainly due to insufficient enzyme stability and low catalytic activity. In addition, the low tolerance of E. coli, the strong hydrophobicity of the substrate, and the high toxicity of the product to the cells are also the main reasons for the current low substrate loading and low production intensity. In order to solve the above problems, a discussion is developed in this review. The construction of a cofactor regeneration system helps to promote a positive shift in the reaction equilibrium. This improves the efficiency of the overall cofactor cycle and avoids large additions of expensive cofactors thereby reducing production costs. Protein engineering is the targeted modification of proteins based on the understanding of known structure and function of proteins with the help of computer-aided design. Adding protein tags enhance the soluble expression of enzymes, and targeted mutagenesis can enhance the stability of enzymes to improve catalytic performance. Blocking the β-oxidation pathway effectively reduced the degradation of fatty acid substrates in whole-cell catalysis. Overexpression of the fatty acid transporter protein FadL and introduction of the outer membrane protein AlkL are both able to increase the efficiency of transmembrane transport of substrates, which improve the overall biocatalytic performance. It has been reported that development of the GDAR system improved the tolerance of the strain to N-heptanoic acid. Advanced strategies such as artificial spore have been implied to alleviate inefficient enzyme catalysis and low product titers caused by the low solubility of the oil substrate, which is expected to improve the whole-cell biosynthesis of fatty acid-derived bifunctional chemicals. Although these issues have been improved, multi-enzyme cascade reactions involve multiple reactions and enzymes, making them more complex than single-step reactions. Multiple factors need to be considered in practical applications of multi-enzyme cascade reactions. The coordination between multiple heterologous enzymes and the compatibility of reaction conditions are key issues in multi-enzyme cascades and a challenging challenge. Therefore, systematic engineering optimization and applied research depending on the type of reaction as well as the product are still needed to increase the potential of bifunctional compounds as industrial products. We can combine the design of biotransformation pathways with computers to obtain cascade enzyme catalytic elements by means such as directed evolution. In the future, we can construct efficient engineered multi-enzyme cascade processes for achieving large-scale industrial applications based on advanced strategies.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This work was financially supported by the National Key Research and Development Program of China (No. 2021YFC2101100), the National Natural Science Foundation of China (No. 22178173), the Jiangsu Agricultural Science and Technology Independent Innovation Fund Project (No. CX(22)3015), the Jiangsu Synergetic Innovation Center for Advanced Bio-Manufacture (No. XTC2204).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Oh H-J, Kim S-U, Song J-W, Lee J-H, Kang W-R, Jo Y-S, et al. Biotransformation of Linoleic Acid into Hydroxy Fatty Acids and Carboxylic Acids Using a Linoleate Double Bond Hydratase as Key Enzyme.
Adv. Synth. Catal. 2015,
357, 408–416.
[Google Scholar]
-
2.
Chung H, Yang JE, Ha JY, Chae TU, Shin JH, Gustavsson M, et al. Bio-based production of monomers and polymers by metabolically engineered microorganisms.
Curr. Opin. Biotechnol. 2015,
36, 73–84.
[Google Scholar]
-
3.
Richter M, Boldescu V, Graf D, Streicher F, Dimoglo A, Bartenschlager R, et al. Synthesis, Biological Evaluation, and Molecular Docking of Combretastatin and Colchicine Derivatives and their hCE1-Activated Prodrugs as Antiviral Agents.
ChemMedChem 2019,
14, 469–483.
[Google Scholar]
-
4.
Lin L, Ledesma-Amaro R, Ji XJ, Huang H. Multienzymatic synthesis of nylon monomers from vegetable oils.
Trend. Biotech. 2023,
41, 150–153.
[Google Scholar]
-
5.
Hirschberg V, Rodrigue D. Recycling of polyamides: Processes and conditions.
J. Polym. Sci. 2023,
61, 1937.
[Google Scholar]
-
6.
Winnacker M, Rieger B. Biobased Polyamides: Recent Advances in Basic and Applied Research.
Macromol. Rapid Commun. 2016,
37, 1391–1413.
[Google Scholar]
-
7.
Shakiba M, Rezvani Ghomi E, Khosravi F, Jouybar S, Bigham A, Zare M, et al. Nylon—A material introduction and overview for biomedical applications.
Polym. Adv. Tech. 2021,
32, 3368–3383.
[Google Scholar]
-
8.
Uyama H. Functional polymers from renewable plant oils.
Polym. J. 2018,
50, 1003–1011.
[Google Scholar]
-
9.
Tembe GL, Bandyopadhyay AR, Ganeshpure PA, Satish S. Catalytic Dimerization of Alkyl Acrylates.
Catal. Rev. 1996,
38, 299–327.
[Google Scholar]
-
10.
Zhang Y-HP, Sun J, Ma Y. Biomanufacturing: history and perspective.
J. Ind. Microbiol. Biotechnol. 2017,
44, 773–784.
[Google Scholar]
-
11.
Finnigan W, Hepworth LJ, Flitsch SL, Turner NJ. RetroBioCat as a computer-aided synthesis planning tool for biocatalytic reactions and cascades.
Nat. Catal. 2021,
4, 98–104.
[Google Scholar]
-
12.
Speranza G, Morelli C, Orlandi M, Scarpellini M, Manitto, P. Fate of the oxygen atoms in the diol-dehydratase-catalyzed dehydration of meso-butane-2,3-diol.
Helvetica Chim. Acta 2001,
84, 335–344.
[Google Scholar]
-
13.
Kunduru KR, Basu A, Haim Zada M, Domb AJ. Castor Oil-Based Biodegradable Polyesters.
Biomacromolecules 2015,
16, 2572–2587.
[Google Scholar]
-
14.
Su Y, Ma S, Wang B, Xu X, Feng H, Hu K, et al. High-performance castor oil-based polyurethane thermosets: Facile synthesis and properties.
React. Funct. Polym. 2023,
183, 105496.
[Google Scholar]
-
15.
Danov SM, Kazantsev OA, Esipovich AL, Belousov AS, Rogozhin AE, Kanakov EA. Recent advances in the field of selective epoxidation of vegetable oils and their derivatives: A review and perspective.
Catal. Sci. Tech. 2017,
7, 3659–3675.
[Google Scholar]
-
16.
Seo JH, Lee SM, Lee J, Park JB. Adding value to plant oils and fatty acids: Biological transformation of fatty acids into omega-hydroxycarboxylic, alpha,omega-dicarboxylic, and omega-aminocarboxylic acids.
J. Biotechnol. 2015,
216, 158–166.
[Google Scholar]
-
17.
Ribeiro AR, Silva SS, Reis RL. Challenges and opportunities on vegetable oils derived systems for biomedical applications.
Biomater. Adv. 2022,
134, 112720.
[Google Scholar]
-
18.
Tran MH, Lee EY. Production of polyols and polyurethane from biomass: A review.
Environ. Chem. Lett. 2023,
21, 2199–2223.
[Google Scholar]
-
19.
Zhou Y, Wu S, Bornscheuer UT. Recent advances in (chemo)enzymatic cascades for upgrading bio-based resources.
Chem. Commun. 2021,
57, 10661–10674.
[Google Scholar]
-
20.
Lopez-Gallego F, Schmidt-Dannert C. Multi-enzymatic synthesis.
Curr. Opin. Chem. Biol. 2010,
14, 174–183.
[Google Scholar]
-
21.
Barber DM, Duriš A, Thompson AL, Sanganee HJ, Dixon DJ. One-Pot Asymmetric Nitro-Mannich/Hydroamination Cascades for the Synthesis of Pyrrolidine Derivatives: Combining Organocatalysis and Gold Catalysis.
ACS Catal. 2014,
4, 634–638.
[Google Scholar]
-
22.
Jiang C, Cheng G, Xu F, Wu Q. One Pot Enzyme-Catalyzed Cascade Benefit Systems.
Mini-Rev. Org. Chem. 2021,
18, 282–295.
[Google Scholar]
-
23.
Pickl M, Fuchs M, Glueck SM, Faber K. Amination of omega-Functionalized Aliphatic Primary Alcohols by a Biocatalytic Oxidation-Transamination Cascade.
ChemCatChem 2015,
7, 3121–3124.
[Google Scholar]
-
24.
Lee DS, Song J-W, Voß M, Schuiten E, Akula RK, Kwon Y-U, et al. Enzyme Cascade Reactions for the Biosynthesis of Long Chain Aliphatic Amines from Renewable Fatty Acids.
Adv. Synth. Catal. 2019,
361, 1359–1367.
[Google Scholar]
-
25.
Ge J, Yang X, Yu H, Ye L. High-yield whole cell biosynthesis of Nylon 12 monomer with self-sufficient supply of multiple cofactors.
Metab. Eng. 2020,
62, 172–185.
[Google Scholar]
-
26.
Lim S, Yoo H, Sarak S, Kim B, Yun H. A multi-enzyme cascade reaction for the production of α,ω-dicarboxylic acids from free fatty acids.
J. Ind. Eng. Chem. 2021,
98, 358–365.
[Google Scholar]
-
27.
Jang H-Y, Singha K, Kim H-H, Kwon Y-U, Park J-B, et al. Chemo-enzymatic synthesis of 11-hydroxyundecanoic acid and 1,11-undecanedioic acid from ricinoleic acid.
Green Chem. 2016,
18, 1089–1095.
[Google Scholar]
-
28.
Brenna E, Colombo D, Di Lecce G, Gatti FG, Ghezzi MC, Tentori F, et al. Conversion of Oleic Acid into Azelaic and Pelargonic Acid by a Chemo-Enzymatic Route.
Molecules 2020,
25, 1882.
[Google Scholar]
-
29.
Denard CA, Hartwig JF, Zhao H. Multistep One-Pot Reactions Combining Biocatalysts and Chemical Catalysts for Asymmetric Synthesis.
ACS Catal. 2013,
3, 2856–2864.
[Google Scholar]
-
30.
Wang M, Si T, Zhao H. Biocatalyst development by directed evolution.
Bioresour. Technol. 2012,
115, 117–125.
[Google Scholar]
-
31.
Biermann U, Bornscheuer U, Meier MA, Metzger JO, Schäfer HJ. Oils and fats as renewable raw materials in chemistry.
Angew. Chem. Int. Ed. Engl. 2011,
50, 3854–3871.
[Google Scholar]
-
32.
Shoda S-I, Uyama H, Kadokawa J, Kimura S, Kobayashi S. Enzymes as Green Catalysts for Precision Macromolecular Synthesis.
Chem. Rev. 2016,
116, 2307–2413.
[Google Scholar]
-
33.
Kang SH, Kim TH, Park JB, Oh DK. Increased Production of omega-Hydroxynonanoic Acid and alpha,omega-Nonanedioic Acid from Olive Oil by a Constructed Biocatalytic System.
J. Agric. Food Chem. 2020,
68, 9488–9495.
[Google Scholar]
-
34.
Wu YX, Pan J, Yu HL, Xu JH. Enzymatic synthesis of 10-oxostearic acid in high space-time yield via cascade reaction of a new oleate hydratase and an alcohol dehydrogenase.
J. Biotechnol. 2019,
306, 100008.
[Google Scholar]
-
35.
Kim T-H, Kang S-H, Han J-E, Seo E-J, Jeon E-Y, Choi G-E, et al. Multilayer Engineering of Enzyme Cascade Catalysis for One-Pot Preparation of Nylon Monomers from Renewable Fatty Acids.
ACS Catal. 2020,
10, 4871–4878.
[Google Scholar]
-
36.
Chong G-G, Ding L-Y, Qiu Y-Y, Qian X-L, Li C-X, Pan J, et al. All-Carbon-Atom Refinery of Oleic Acid into Bifunctional Chemicals Using Artificial Consortia of Escherichia coli Strains.
ACS Sustain. Chem. Eng. 2022,
10, 13125–13132.
[Google Scholar]
-
37.
Seo EJ, Yeon YJ, Seo JH, Lee JH, Boñgol JP, Oh Y, et al. Enzyme/whole-cell biotransformation of plant oils, yeast derived oils, and microalgae fatty acid methyl esters into n-nonanoic acid, 9-hydroxynonanoic acid, and 1,9-nonanedioic acid.
Bioresour. Technol. 2018,
251, 288–294.
[Google Scholar]
-
38.
Ahsan M, Patil MD, Jeon H, Sung S, Chung T, Yun H. Biosynthesis of Nylon 12 Monomer, ω-Aminododecanoic Acid Using Artificial Self-Sufficient P450, AlkJ and ω-TA.
Catalysts 2018,
8, 400.
[Google Scholar]
-
39.
Kirschner A, Altenbuchner J, Bornscheuer UT. Baeyer-Villiger-Monooxygenase fromPseudomonas fluorescens DSm50106.
Chem. Ing. Tech. 2006,
78, 1408–1409.
[Google Scholar]
-
40.
Zhang GX, You ZN, Yu JM, Liu YY, Pan J, Xu JH, et al. Discovery and Engineering of a Novel Baeyer-Villiger Monooxygenase with High Normal Regioselectivity.
ChemBioChem 2021,
22, 1190–1195.
[Google Scholar]
-
41.
Chong GG, Ding LY, Qiu YY, Qian XL, Dong YL, Li CX, et al. Building Flexible Escherichia coli Modules for Bifunctionalizing N-Octanol: The Byproduct of Oleic Acid Biorefinery.
J. Agric. Food Chem. 2022,
70, 10543–10551.
[Google Scholar]
-
42.
Otte KB, Kittelberger J, Kirtz M, Nestl BM, Hauer B. Whole-Cell One-Pot Biosynthesis of Azelaic Acid.
ChemCatChem 2014,
6, 1003–1009.
[Google Scholar]
-
43.
Coenen A, Ferrer M, Jaeger KE, Schörken U. Synthesis of 12-aminododecenoic acid by coupling transaminase to oxylipin pathway enzymes.
Appl. Microbiol. Biotechnol. 2023,
107, 2209–2221.
[Google Scholar]
-
44.
Benessere V, Cucciolito ME, De Santis A, Di Serio M, Esposito R, Ruffo F, et al. Sustainable Process for Production of Azelaic Acid Through Oxidative Cleavage of Oleic Acid.
J. Am. Oil Chem. Soc. 2015,
92, 1701–1707.
[Google Scholar]
-
45.
Todea A, Deganutti C, Spennato M, Asaro F, Zingone G, Milizia T, et al. Azelaic Acid: A Bio-Based Building Block for Biodegradable Polymers.
Polymers 2021,
13, 4091.
[Google Scholar]
-
46.
Song JW, Jeon EY, Song DH, Jang HY, Bornscheuer UT, Oh DK, et al. Multistep enzymatic synthesis of long-chain alpha,omega-dicarboxylic and omega-hydroxycarboxylic acids from renewable fatty acids and plant oils.
Angew. Chem. Int. Ed. Engl. 2013,
52, 2534–2537.
[Google Scholar]
-
47.
Sun QF, Zheng YC, Chen Q, Xu JH, Pan J. Engineering of an oleate hydratase for efficient C10-Functionalization of oleic acid.
Biochem. Biophys. Res. Commun. 2021,
537, 64–70.
[Google Scholar]
-
48.
Rehdorf J, Kirschner A, Bornscheuer UT. Cloning, expression and characterization of a Baeyer-Villiger monooxygenase from Pseudomonas putida KT2440.
Biotechnol. Lett. 2007,
29, 1393–1398.
[Google Scholar]
-
49.
Custódio L, Soares F, Pereira H, Barreira L, Vizetto-Duarte C, João M, et al. Fatty acid composition and biological activities of Isochrysis galbana T-ISO,
Tetraselmis sp. and
Scenedesmus sp.: possible application in the pharmaceutical and functional food industries.
J. Appl. Phycol. 2013,
26, 151–161.
[Google Scholar]
-
50.
Jeon E-Y, Seo J-H, Kang W-R, Kim M-J, Lee J-H, Oh D-K, et al. Simultaneous Enzyme/Whole-Cell Biotransformation of Plant Oils into C9 Carboxylic Acids.
ACS Catal. 2016,
6, 7547–7553.
[Google Scholar]
-
51.
Liavonchanka A, Feussner I. Lipoxygenases: occurrence, functions and catalysis.
J. Plant Physiol. 2006,
163, 348–357.
[Google Scholar]
-
52.
Otte KB, Kirtz M, Nestl BM, Hauer B. Synthesis of 9-oxononanoic acid, a precursor for biopolymers.
ChemSusChem 2013,
6, 2149–2156.
[Google Scholar]
-
53.
Li X-L, Zhang K, Jiang J-L, Zhu R, Wu W-P, Deng J, et al. Synthesis of medium-chain carboxylic acids or α,ω-dicarboxylic acids from cellulose-derived platform chemicals.
Green Chem. 2018,
20, 362–368.
[Google Scholar]
-
54.
Kirschner A, Altenbuchner J, Bornscheuer UT. Cloning, expression, and characterization of a Baeyer-Villiger monooxygenase from Pseudomonas fluorescens DSM 50106 in
E. coli.
Appl. Microbiol. Biotechnol. 2007,
73, 1065–1072.
[Google Scholar]
-
55.
Yu JM, Liu YY, Zheng YC, Li H, Zhang XY, Zheng GW, et al. Direct Access to Medium-Chain alpha,omega-Dicarboxylic Acids by Using a Baeyer-Villiger Monooxygenase of Abnormal Regioselectivity.
ChemBioChem 2018,
19, 2049–2054.
[Google Scholar]
-
56.
Wang L, Wang L, Wang R, Wang Z, Wang J, Yuan H, et al. Efficient Biosynthesis of 10-Hydroxy-2-decenoic Acid Using a NAD(P)H Regeneration P450 System and Whole-Cell Catalytic Biosynthesis.
ACS Omega 2022,
7, 17774–17783.
[Google Scholar]
-
57.
Collazo N, Carpena M, Nuñez-Estevez B, Otero P, Simal-Gandara J, Prieto MA. Health Promoting Properties of Bee Royal Jelly: Food of the Queens.
Nutrients 2021,
13, 543.
[Google Scholar]
-
58.
Koh M-H, Kim H, Shin N, Kim H-S, Yoo D, Kim Y-G. Divergent Process for C10, C11and C12ω-Amino Acid and α,ω-Dicarboxylic Acid Monomers of Polyamides from Castor Oil as a Renewable Resource.
B Korean Chem. Soc. 2012,
33, 1873–1878.
[Google Scholar]
-
59.
Kim TH, ang SH, Park JB, Oh DK. Construction of an engineered biocatalyst system for the production of medium-chain alpha,omega-dicarboxylic acids from medium-chain omega-hydroxycarboxylic acids.
Biotechnol. Bioeng. 2020,
117, 2648–2657.
[Google Scholar]
-
60.
Yang KM, Kim BM, Park JB. Omega-Hydroxyundec-9-enoic acid induces apoptosis through ROS-mediated endoplasmic reticulum stress in non-small cell lung cancer cells.
Biochem. Biophys. Res. Commun. 2014,
448, 267–273.
[Google Scholar]
-
61.
Cho YH, Kim SJ, Kim HW, Kim JY, Gwak JS, Chung D, et al. Continuous supply of glucose and glycerol enhances biotransformation of ricinoleic acid to (E)-11-(heptanoyloxy) undec-9-enoic acid in recombinant
Escherichia coli.
J. Biotechnol. 2017,
253, 34–39.
[Google Scholar]
-
62.
Jang H-Y, Jeon E-Y, Baek A-Y, Lee S-M, Park J-B. Production of ω-hydroxyundec-9-enoic acid and N-heptanoic acid from ricinoleic acid by recombinant Escherichia coli-based biocatalyst.
Process Biochem. 2014,
49, 617–622.
[Google Scholar]
-
63.
Cho YH, Kim SJ, Kim JY, Lee DH, Park K, Park YC. Effect of PelB signal sequences on Pfe1 expression and omega-hydroxyundec-9-enoic acid biotransformation in recombinant
Escherichia coli.
Appl. Microbiol. Biotechnol. 2018,
102, 7407–7416.
[Google Scholar]
-
64.
Yapa Mudiyanselage A, Viamajala S, Varanasi S, Yamamoto K. Simple Ring-Closing Metathesis Approach for Synthesis of PA11, 12, and 13 Precursors from Oleic Acid.
ACS Sustain. Chem. Eng. 2014,
2, 2831–2836.
[Google Scholar]
-
65.
Ahsan MM, Jeon H, P Nadarajan S, Chung T, Yoo HW, Kim BG, et al. Biosynthesis of the Nylon 12 Monomer, omega-Aminododecanoic Acid with Novel CYP153A, AlkJ, and omega-TA Enzymes.
Biotechnol. J. 2018,
13, e1700562.
[Google Scholar]
-
66.
Honda Malca S, Scheps D, Kühnel L, Venegas-Venegas E, Seifert A, Nestl BM, et al. Bacterial CYP153A monooxygenases for the synthesis of omega-hydroxylated fatty acids.
Chem. Commun. 2012,
48, 5115–5117.
[Google Scholar]
-
67.
Munro AW, Leys DG, McLean KJ, Marshall KR, Ost TW, Daff S, et al. P450 BM3: the very model of a modern flavocytochrome.
Trend. Biochem. Sci. 2002,
27, 250–257.
[Google Scholar]
-
68.
Scheps D, Honda Malca S, Richter SM, Marisch K, Nestl BM, Hauer B. Synthesis of omega-hydroxy dodecanoic acid based on an engineered CYP153A fusion construct.
Microb. Biotechnol. 2013,
6, 694–707.
[Google Scholar]
-
69.
Hollmann F, Arends IWCE, Holtmann D. Enzymatic reductions for the chemist.
Green Chem. 2011,
13, 2285–2314.
[Google Scholar]
-
70.
Shin KC, Kang S-H, Lee T-E, Kim T-H, Oh D-K. Pentadecanedioic acid production from 15-hydroxypentadecanoic acid using an engineered biocatalyst with a co-factor regeneration system.
J. Am. Oil Chem. Soc. 2022,
99, 675–683.
[Google Scholar]
-
71.
Kirmair L, Seiler DL, Skerra A. Stability engineering of the Geobacillus stearothermophilus alcohol dehydrogenase and application for the synthesis of a polyamide 12 precursor.
Appl. Microbiol. Biotechnol. 2015,
99, 10501–10513.
[Google Scholar]
-
72.
Seo EJ, Kim HJ, Kim MJ, Kim JS, Park JB. Cofactor specificity engineering of a long-chain secondary alcohol dehydrogenase from Micrococcus luteus for redox-neutral biotransformation of fatty acids.
Chem. Commun. 2019,
55, 14462–14465.
[Google Scholar]
-
73.
Park YJ, Lee KH, Baek MS, Kim D-M. High-throughput engineering of initial coding regions for maximized production of recombinant proteins.
Biotechnol. Bioprocess Eng. 2017,
22, 497–503.
[Google Scholar]
-
74.
Opperman DJ, Reetz MT. Towards practical Baeyer-Villiger-monooxygenases: design of cyclohexanone monooxygenase mutants with enhanced oxidative stability.
ChemBioChem 2010,
11, 2589–2596.
[Google Scholar]
-
75.
Schöneich C. Mechanisms of protein damage induced by cysteine thiyl radical formation.
Chem. Res. Toxicol. 2008,
21, 1175–1179.
[Google Scholar]
-
76.
Schöneich C. Thiyl radicals and induction of protein degradation.
Free Radic. Res. 2015,
50, 143–149.
[Google Scholar]
-
77.
Dijkman WP, de Gonzalo G, Mattevi A, Fraaije MW. Flavoprotein oxidases: classification and applications.
Appl. Microbiol. Biotechnol. 2013,
97, 5177–5188.
[Google Scholar]
-
78.
Woo JM, Jeon EY, Seo EJ, Seo JH, Lee DY, Yeon YJ, et al. Improving catalytic activity of the Baeyer-Villiger monooxygenase-based Escherichia coli biocatalysts for the overproduction of (Z)-11-(heptanoyloxy)undec-9-enoic acid from ricinoleic acid.
Sci. Rep. 2018,
8, 10280.
[Google Scholar]
-
79.
Seo EJ, Kim M-J, Park S-Y, Park S, Oh D-K, Bornscheuer U, et al. Enzyme Access Tunnel Engineering in Baeyer-Villiger Monooxygenases to Improve Oxidative Stability and Biocatalyst Performance.
Adv. Synth. Catal. 2021,
364, 555–564.
[Google Scholar]
-
80.
Bae JH, Park BG, Jung E, Lee PG, Kim BG. fadD deletion and fadL overexpression in
Escherichia coli increase hydroxy long-chain fatty acid productivity.
Appl. Microbiol. Biotechnol. 2014,
98, 8917–8925.
[Google Scholar]
-
81.
Wang D, Wu H, Thakker C, Beyersdorf J, Bennett GN, San KY. Efficient free fatty acid production in engineered
Escherichia coli strains using soybean oligosaccharides as feedstock.
Biotechnol. Prog. 2015,
31, 686–694.
[Google Scholar]
-
82.
Jawed K, Mattam AJ, Fatma Z, Wajid S, Abdin MZ, Yazdani SS. Engineered Production of Short Chain Fatty Acid in Escherichia coli Using Fatty Acid Synthesis Pathway.
PLoS ONE 2016,
11, e0160035.
[Google Scholar]
-
83.
Royce LA, Yoon JM, Chen Y, Rickenbach E, Shanks JV, Jarboe LR. Evolution for exogenous octanoic acid tolerance improves carboxylic acid production and membrane integrity.
Metab. Eng. 2015,
29, 180–188.
[Google Scholar]
-
84.
Jeon EY, Song JW, Cha HJ, Lee SM, Lee J, Park JB. Intracellular transformation rates of fatty acids are influenced by expression of the fatty acid transporter FadL in
Escherichia coli cell membrane.
J. Biotechnol. 2018,
281, 161–167.
[Google Scholar]
-
85.
Julsing MK, Schrewe M, Cornelissen S, Hermann I, Schmid A, Bühler B. Outer membrane protein AlkL boosts biocatalytic oxyfunctionalization of hydrophobic substrates in
Escherichia coli.
Appl. Environ. Microbiol. 2012,
78, 5724–5733.
[Google Scholar]
-
86.
Shin J, Yu J, Park M, Kim C, Kim H, Park Y, et al. Endocytosing
Escherichia coli as a Whole-Cell Biocatalyst of Fatty Acids.
ACS Synth. Biol. 2019,
8, 1055–1066.
[Google Scholar]
-
87.
Xu Y, Li F, Yang K, Qiao Y, Yan Y, Yan J. A facile and robust non-natural three enzyme biocatalytic cascade based on
Escherichia coli surface assembly for fatty alcohol production.
Energy Convers. Manag. 2019,
181, 501–506.
[Google Scholar]
-
88.
Zhang ZX, Wang YZ, Nong FT, Xu Y, Ye C, Gu Y, et al. Developing a dynamic equilibrium system in
Escherichia coli to improve the production of recombinant proteins.
Appl. Microbiol. Biotechnol. 2022,
106, 6125–6137.
[Google Scholar]
-
89.
Woo JM, Kim JW, Song JW, Blank LM, Park JB. Activation of the Glutamic Acid-Dependent Acid Resistance System in Escherichia coli BL21(DE3) Leads to Increase of the Fatty Acid Biotransformation Activity.
PLoS ONE 2016,
11, e0163265.
[Google Scholar]
-
90.
Sun Z, Hübner R, Li J, Wu C. Artificially sporulated
Escherichia coli cells as a robust cell factory for interfacial biocatalysis.
Nat. Commun. 2022,
13, 3142.
[Google Scholar]