1. Introduction
Terpenoids represent a large and diverse group of isoprene-based natural products, with over 80,000 distinct structural types identified. Terpenoids are typically classified based on the number of isoprene units they contain, following the isoprene rule: monoterpenoids (C10), sesquiterpenoids (C15), diterpenoids (C20), triterpenoids (C30), tetraterpenoids (C40) () [
1]. Many terpenoids exhibit significant pharmacological functions and biological activities. For example, citronellol exhibits properties such as anti-inflammatory, anti-convulsant, anti-hyperalgesia and anti-cholesterol [
2]. (+)-Nootkatone possesses activities such as anticancer, antiplatelet aggregation, antimicrobial, and anti-inflammation [
3]. Forskolin is known for its anticancer, antiasthmatic and antihypertensive activities [
4]. Protopanaxadiol exhibits a wide range of activities including anti-cancer, anti-inflammatory, anti-oxidant, hepatoprotectant, anti-lipogenic, wound-healing, and anti-obesity effects [
5]; Violaxanthin has been found to display antiproliferative, anti-inflammatory and photoprotective effects [
6]. Furthermore, the structural diversity of terpenoids also enables their extensive applications in biofuels [
7], food [
8,
9], cosmetic [
10,
11] and agriculture industries [
12]. This versatility offers immense potential and commercial value.
Currently, terpenoids are primarily obtained through plant extraction [
13] or chemical synthesis [
10,
11]. However, plant extraction is hindered by long cultivation cycles and susceptibility to environmental factors. In addition, the concentration of synthetic terpenoids in plants is low, and structural analogs are commonly found, which makes it difficult to be separated and purified. On the other hand, chemical synthesis methods for terpenoids require a large amount of organic reagents, which is not environmentally friendly. Furthermore, due to the intricate stereoscopic structures and complexity of most terpenoids, their synthesis and separation using chemical methods are challenging [
14]. In recent years, with the rapid development of metabolic engineering and synthetic biology, microorganisms are gradually being developed as efficient platforms for the green industrial production of terpenoids [
15]. Compared to traditional plant extraction and chemical synthesis methods, the utilization of fast-growing microorganisms offers several advantages. Notably, it significantly reduces the production cycle from months to just a few days, while also lowering production costs. Furthermore, the ease of separating the final products is enhanced as the target compounds are enriched within the microbial cell factory. Therefore, the microbial fermentation approach for terpenoid production not only enhances stability, controllability, and productivity but also promotes eco-friendliness [
1,
16].
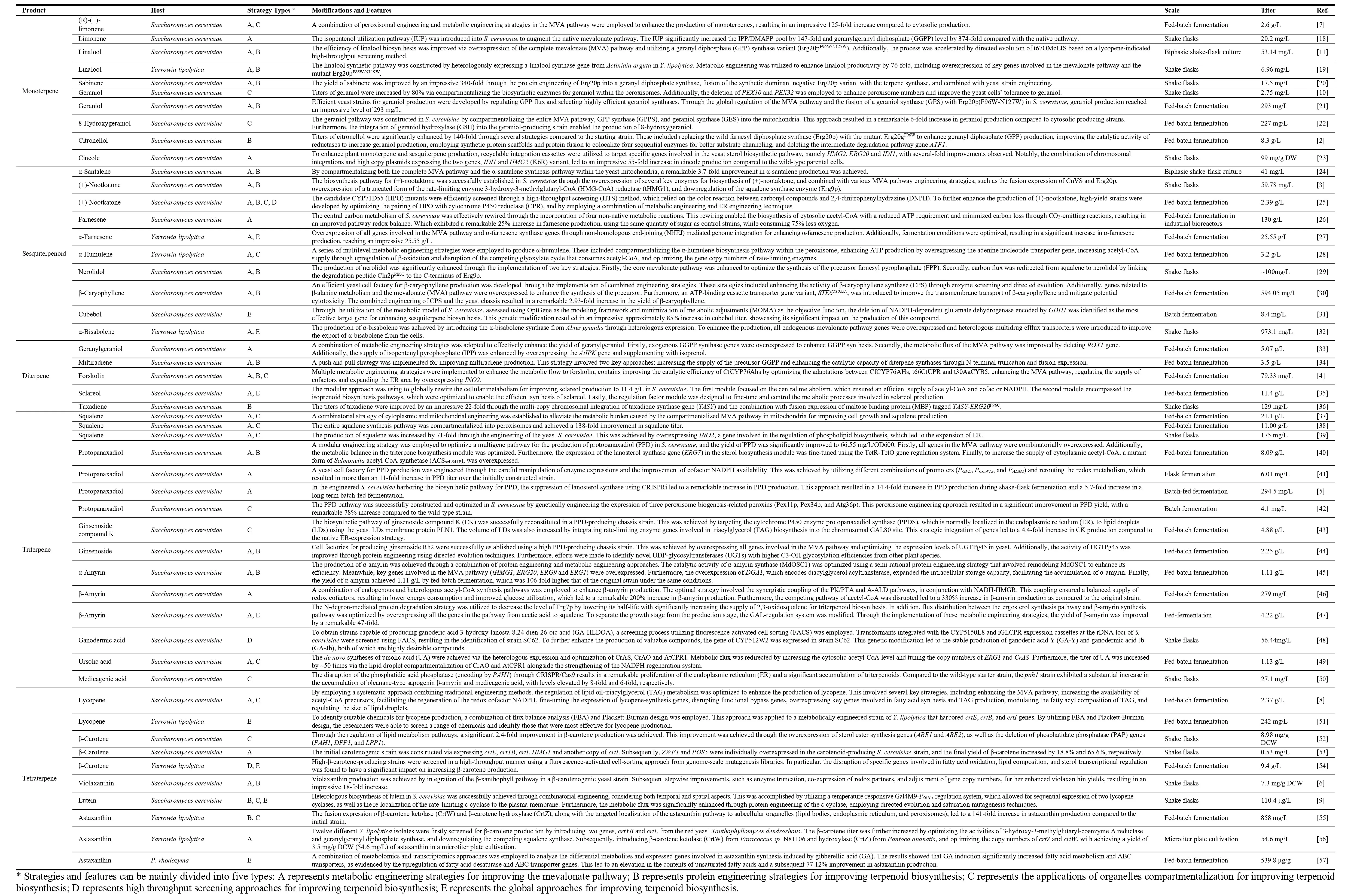
Terpenoid biosynthesis involves two main pathways: the mevalonate (MVA) pathway and the methylerythritol phosphate (MEP) pathway [
1]. Prokaryotes naturally possess the MEP pathway, while fungi predominantly use the MVA pathway. To date, various microorganisms have been used for producing terpenoids, with yeast being particularly advantageous and focused (). Yeast possesses several key attributes that make it versatile for terpenoid synthesis. Firstly, it exhibits tolerance to low pH and osmotic stress as well as resistance towards phage infections. More importantly, yeast can offer a wide range of precursors for most terpenoids through its endogenous MVA pathway, including GPP, FPP, GGPP and squalene (). In addition, yeast possesses subcellular structures such as the endoplasmic reticulum, Golgi apparatus, and peroxisome, along with post-translational modification mechanisms, which are beneficial for the efficient expression of exogenous enzymes from animals and plants. Moreover,
Saccharomyces cerevisiae, a commonly used yeast strain, is generally-recognized-as-safe (GRAS) host and has a well-characterized genetic background, making it widely used for food and alcohol production [
17]. Considering all these factors, yeast emerges as an ideal host for terpenoids synthesis [
1,
16].
This review provides a comprehensive examination of the diverse strategies employed to enhance terpenoid production in yeasts. Specifically, we introduce several engineering strategies targeting the mevalonate pathway, as well as protein engineering strategies for terpene biosynthesis. Additionally, we thoroughly analyze the applications of various organelles, assessing both their advantages and limitations in terms of compartmentalization. Furthermore, we explore the development and prospects of global approaches and high-throughput methods for the creation of efficient cell factories. Finally, we extensively discuss the future perspectives and challenges associated with constructing yeast strains capable of producing terpenoids.
. The biosynthesis of different classes of terpenoids in S. cerevisiae and Y. lipolytica using the mevalonate pathway.
. Examples of engineering yeasts for production of terpenoids.
2. Metabolic Engineering Strategies for Improving the Mevalonate Pathway
The mevalonate (MVA) pathway in yeasts serves as the source of precursors for the synthesis of terpenoids. Terpenoids are synthesized mainly from two C5 precursors [
23], namely isopentenyl diphosphate (IPP) and dimethylallyl diphosphate (DMAPP). As shown in , IPP and DMAPP results from the multi-step conversion of acetyl-CoA through the MVA pathway. Notably, the conversion of HMG-CoA to mevalonate by HMG-CoA reductase represents the rate-limiting step of the MVA pathway [
1,
58]. These two five-carbon (C5) skeletons are then ligated in diverse proportions to produce different pyrophosphate precursors, such as geranyl pyrophosphate (GPP), farnesyl pyrophosphate (FPP) and geranylgeranyl pyrophosphate (GGPP). These precursors serve as substrates for a series of rearrangement, cyclization, methylation, and other reactions, ultimately leading to the diverse array of natural terpenoids () [
15]. Considering the significance of the MVA pathway, the metabolic mechanism of the endogenous MVA pathway has been completely elucidated in yeast. Consequently, one of the primary approaches to significantly enhance the efficient synthesis of terpenoids is to focus on augmenting the flux of this pathway through the implementation of metabolic engineering techniques.
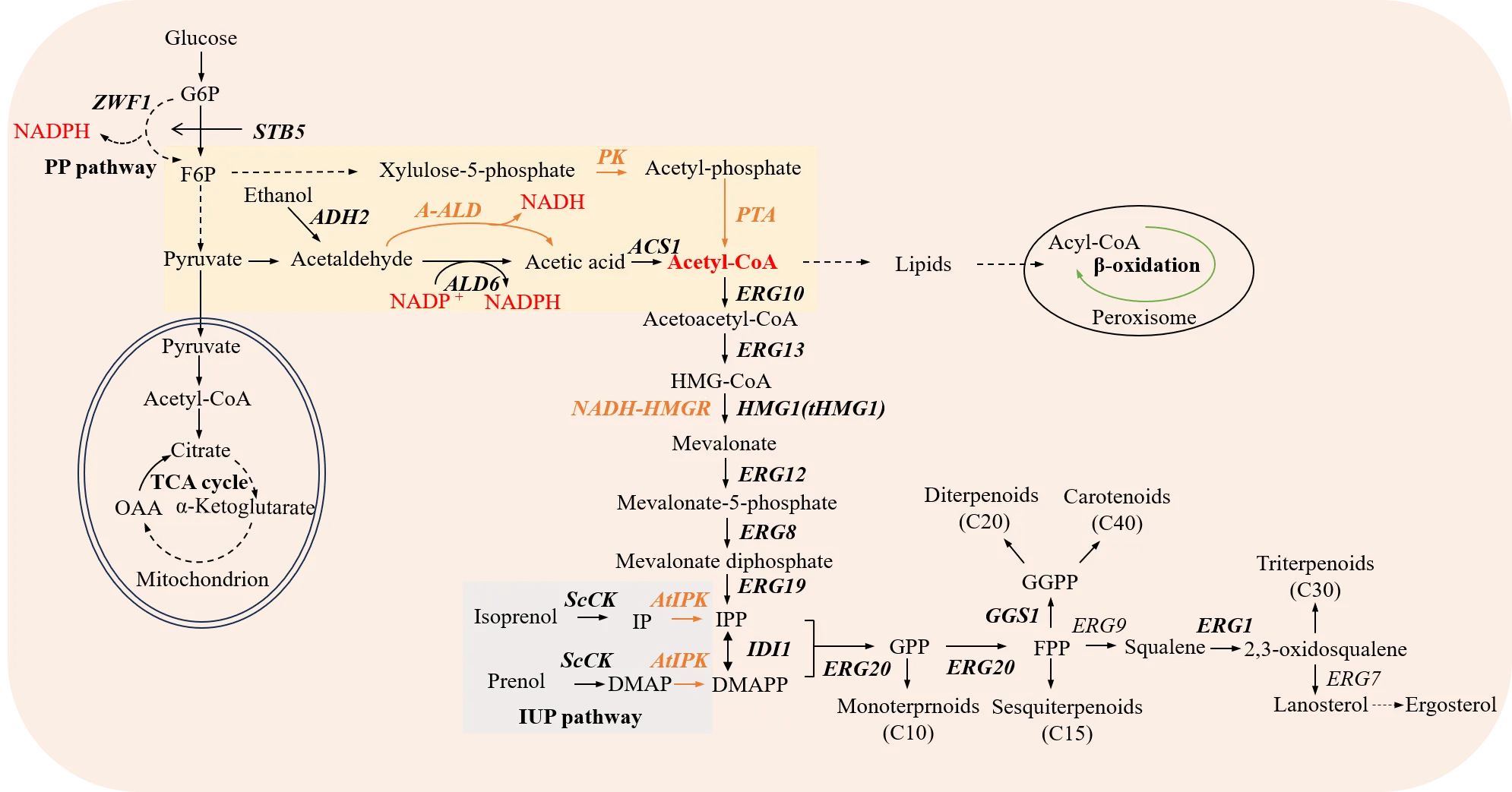
. Manipulations of the mevalonate pathways for optimizing biosynthesis of terpenoids in yeast. Overexpressed endogenous genes are shown in black bold. Additionally, the introduction of heterologous acetyl-CoA synthesis pathways and the synthetic isopentenol utilization pathway (IUP) in yeast is represented in orange. The solid and dotted arrows indicate single and multiple steps, respectively. PK, phosphoketolase gene from Leuconostoc mesenteroides; PTA, phosphotransacetylase gene from Clostridium kluyveri; A-ALD, acetylating acetaldehyde dehydrogenase gene from E. coli; NADH-HMGR, NADH-dependent HMG-CoA reductase variant gene from Silicibacter pomeroyi; AtIPK, isopentenyl phosphate kinase gene from Arabidopsis thaliana; ScCK, choline kinase gene; ZWF1, glucose-6-phosphate dehydrogenase gene; AHD2, alcohol dehydrogenase gene; ALD6, acetaldehyde dehydrogenase gene; ACS1, acetyl-CoA synthase gene; ERG10, acetyl-CoA thiolase gene; ERG13, HMG-CoA synthase gene; HMG1, 3-hydroxy-3-methyl glutaryl coenzyme A reductase gene; tHMG1, truncated HMG1; ERG12, mevalonate kinase gene; ERG8, phosphomevalonate kinase gene; ERG19, mevalonate diphosphate decarboxylase gene; IDI1, IPP isomerase gene; ERG20, geranyl/farnesyl diphosphate synthase gene; GGS1, geranylgeranyl diphosphate synthase gene; ERG9, squalene synthase gene; ERG1, squalene monooxygenase gene; ERG7, lanosterol synthase gene.
Enhancing precursor supply is one of the most common strategies to improve production of biochemicals. In the case of terpenoid production, overexpression of the key enzymes in the MVA pathway, such as HMG-CoA reductase (HMGR), is a widely used approach for improving IPP and DMAPP supply [
1,
58]. HMGR is a membrane-bound protein localized in the endoplasmic reticulum (ER), consisting of an N-terminal membrane anchor and a C-terminal catalytic domain [
58]. Overexpression of either intact or N-terminal truncated HMGR has been shown to enhance the production of diverse terpenoids [
1,
15,
56,
59,
60]. In previous studies, an N-terminal-truncated HMGR was usually suggested more effective, because the N-terminal truncation of HMGR can prevent self-degradation and make it more stable in the cytoplasm [
1,
15,
59]. However, Kildegaard et al. found that the native HMGR is superior for β-carotene production in
Y. lipolytica [
56]. IPP isomerase (IDI) is responsible for the interconversion between IPP and DMAPP and plays a crucial role in the flux distribution of GPP and FPP [
59]. Overexpression of IDI is also a common strategy to enhance the biosynthesis of terpenoids. For example, over-expression of
IDI1 enhanced cineole by 24-fold in
S. cerevisiae [
23]. Besides HMGR and IDI, most of the pathway genes in the MVA pathway (
ERG10,
ERG13,
ERG12,
ERG8 and
ERG19) have also been over-expressed to increase production of terpenoids [
32,
44]. Such as, overexpression of all genes in MVA pathway improved the bisabolene production with 251-fold, 29-fold and 72-fold enhancement in α-bisabolene, β-bisabolene and γ-bisabolene production, respectively [
32].
Furthermore, introducing heterologous synthesis pathways for improving IPP and DMAPP precursors is also a new strategy for yeasts to synthesize terpenoids. The isopentenol utilization pathway (IUP) was reported to provide universal C5 precursors for terpenoid synthesis, bypassing the lengthy and strictly regulated natural MVA pathway [
15,
61]. This pathway utilizes choline kinase (ScCK) from
S. cerevisiae and isopentenyl phosphokinase (
AtIPK) from
A. thaliana to directly phosphorylate the secondary substrate isoprenoid alcohol into IPP and DMAPP at the expense of two molecules of ATP () [
62]. Compared with the MVA pathway, the IUP pathway has the advantages of fewer steps and a higher metabolic flow [
18,
62]. For example, the introduction of a two-step isopentenol utilization pathway (IUP) into
S. cerevisiae resulted in a 147-fold increase in the IPP/DMAPP pool compared to the native MVA pathway [
18]. Cofeeding isoprenol and prenol further enhanced the geranyl diphosphate (GPP) content for monoterpene biosynthesis and elevated the GGPP level by 374-fold.
2.2. Improving the Supply of Acetyl-CoA and Cofactors
Increasing the supply of cytosolic acetyl-CoA is supposed to be an effective strategy to improve terpenoids yield [
15]. In native
S. cerevisiae, cytosolic acetyl-CoA is synthesized from the pyruvate dehydrogenase (PDH) bypass or during ethanol reoxidation, which involves pyruvate decarboxylase (PDC), alcohol dehydrogenase (ADH), aldehyde dehydrogenases (ALD) and acetyl-CoA synthetase (ACS) [
1,
46]. Common strategies for enhancing the cytosolic acetyl-CoA pool include manipulating the PDH-bypass, and removing the competitive glyoxylate shunt pathway by knockouting
MLS1 (encode malate synthase) and
CIT2 (encode peroxisomal citrate synthase) [
1,
46,
49,
63]. However, the endogenous PDH bypass for acetyl-CoA synthesis exhibits low efficiency owing to the high energy cost with the ATP-requiring reactions catalyzed by ACS (2 ATP for one acetyl-CoA formation) [
1,
46]. To address this limitation, Meadows et al. rewired the central carbon metabolism in
S. cerevisiae using four non-native metabolic reactions by acetaldehyde dehydrogenase acylating (ADA, namely A-ALD), xylulose-5-phosphate (X5P)-specific phosphoketolase (xPK), phosphotransacetylase (PTA) and NADH-consuming HMG-CoA reductase (NADH-HMGR) () [
26]. This rewiring generated cytosolic acetyl-CoA with lower ATP cost, reduced carbon loss to CO
2-emitting reactions and improved pathway redox balance, and resulted in a 25% increase in the production of β-farnesene while requiring 75% less oxygen. In another work, Liu et al. compared the effects of different acetyl-CoA synthesis pathways for β-amyrin synthesis [
46]. Among these pathways, the combination of PK/PTA and A-ALD pathways together with NADH-HMGR has been found to result in a highly efficient glucose conversion to β-amyrin, achieving a high attainable yield of 0.296 g/g glucose. This combination allows for the satisfaction of ATP consumption in the PK/PTA pathway through the reoxidation of NADH generated in the A-ALD pathway. As a result, a more balanced redox cofactors and a much lower ATP consumption is achieved than any single pathway coupled β-amyrin synthesis.
The redox cofactor NADPH plays a critical role in the metabolism of
S. cerevisiae. In addition to replacing the NADPH-dependent HMGR with the NADH-dependent NADH-HMGR to alter the cofactor dependence, replacing the NADH-generating enzyme with a NADPH-generating enzyme can also enhance the availability of NADPH (). For example, the availability of NADPH was enhanced by 1.3-fold through replacing a NADH-generating enzyme (Ald2p) with a functionally equivalent enzyme (Ald6p) that generates NADPH instead, and protopanaxadiol (PPD) production was improved 4.5-fold in
S. cerevisiae [
41]. Furthermore, the supply of NADPH can be increased by promoting the pentose phosphate (PP) pathway, where the rate-limiting step is catalyzed by glucose-6-phosphate dehydrogenase (encoded by
ZWF1 gene in yeast) [
53,
64]. In a recent reported, it has been shown that overexpression of the
ZWF1 gene increased NADPH concentrations by 1.4-fold in
S. cerevisiae [
53]. This led to a 59.9% improvement in lycopene production and an 18.8% improvement in β-carotene production. In parallel, the
STB5 gene has been reported to be a basal regulator of the pentose phosphate (PP) pathway, and its overexpression increased NADPH concentrations by 1.2-fold in
S. cerevisiae [
41]. Moreover, NADH kinase is capable of directly phosphorylating NADH to NADPH without directly impacting the metabolic pathway. Overexpression of
POS5, which encodes a mitochondrial NADH kinase that converts NADH to NADPH, significantly enhanced lycopene yield by 81.4% and β-carotene yield by 65.6% in
S. cerevisiae [
53]. When controlled by
PGAL7, the overexpression of
POS5 in
S. cerevisiae improved the ursolic acid (UA) and oleanolic acid (OA) titers to 692.3 and 253.4 mg/L, respectively [
49]. Consequently, this study offers a novel approach to construct cell factories for efficient terpenoid synthesis, particularly those that necessitate substantial quantities of NADPH.
2.3. Inhibition of Endogenous Competition Pathway
The precursor pool of GPP, FPP, and GGPP can also be promoted by down-regulating the downstream genes or genes in competitive pathways. For example,
ERG9, which encodes squalene synthase responsible for squalene synthesis, is a key node in competition with FPP for the production of sesquiterpenoid [
3], diterpenoid [
34] and tetraterpenoid [
56]. To increase the supply of FPP pool for the production of diterpenoid and tetraterpenoid, the native promoter of
ERG9 was replaced it with a weak promoter
PHXT1 or truncated, which has been shown to improve sclareol production to 310.5 mg/L [
35] and β-carotene titer by 2.5 fold [
56], respectively. In parallel, Peng et al. linked degradation peptide CLN2
PEST to the C-terminus of Erg9p for down regulating Erg9p at the translational level, which improved nerolidol titre by 86% to 100 mg L
−1 [
29].
In a similar way, down-regulating
ERG7 expression and reducing Erg7p activity are proven to be important strategies to increase triterpenoid titer by reducing 2,3-oxidosqualene flux into endogenous ergosterol pathway [
65]. Various strategies have been employed to down-regulate
ERG7 transcript expression in engineered yeasts, such as replacing the native
ERG7 promoter with a methionine sensitive
MET3 promoter [
66] or copper-repressible CTR3 promoter [
67], fine-tuning of ERG7 expression using CRISPR interference (CRISPRi) [
5] and the tetracycline repressor-tetracycline operator (TetR-TetO)-based gene regulation system [
40]. These transcript regulation strategies have proven to be effective in enhancing triterpenoid titer in yeast. Additionally, critical amino acid residues of Erg7p were engineered to decrease Erg7p activity, which resulted in 7.3- and 3-fold increases in the titers of dammarane-type and lupane-type triterpenoids, respectively [
68]. Interestingly, a degron-mediated protein degradation strategy was employed to lower the half-life of Erg7p, resulting in 3 to 8-fold increase in 2,3‑oxidosqualene-derived triterpenoid production and a 155-fold increase in 2,3:22,23-dioxidosqualene-derived triterpenoid production [
47]. These studies demonstrated the effectiveness of deregulating the native competition within the ergosterol pathway to optimize flux in triterpenoid production and offered a novel perspective on the biosynthesis of triterpenoids.
3. Protein Engineering Strategies for Improving Terpenoid Biosynthesis in Yeasts
The construction of a
de novo terpenoid synthesis pathway in yeasts typically involves the introduction of specific terpene synthases that can convert different precursors derived from the MVA pathway into target products. To improve synthesis efficiency and product yield, factors such as enzyme activity and selectivity need to be considered. Protein engineering strategies can be used to optimize and enhance enzyme properties, such as, rational design or semi-rational design, protein fusion expression and directed evolution of enzymes.
3.1. Rational Design or Semi-rational Design
For enzymes with known enzyme structure and catalytic mechanism, rational design or semi-rational design can effectively improve the specificity and activity of enzymes. In the terpenoid synthesis pathway, some enzymes are promiscuous and can catalyze the production of a wide range of natural and non-natural substrates. For example, the enzyme Erg20p functions as both a GPP synthase (GPPS) and FPP synthase (FPPS), posing challenges in separating the two synthases during monoterpenoids biosynthesis [
11,
59]. To increase the monoterpenoids titer, the mutant
ERG20F96W-N127W has been employed to improve the GPP pool by rational protein engineering design of Erg20p in
S. cerevisiae, achieved the 10-fold increase in sabinene production [
20]. A similar strategy was also employed to modify Erg20p with introducing mutant ERG20
F88W-N119W via rational protein engineering design in
Y. lipolytica, and resulted in about 0.56 mg/L of linalool production [
19]. For the majority of enzymes in the terpenoid biosynthetic pathway, their crystalline structure, active site residues and catalytic mechanism have not been resolved, posing challenges for rational design or semi-rational design approaches. Fortunately, molecular modeling tools can be employed to simulate the putative protein structure. Through homology modeling and docking, key residues were identified of CYP72A63 that is the P450 enzyme for licorice triterpenoid biosynthesis [
69]. Then the structure model aided to rationally design the promiscuous CYP72A63 for improving the
in vivo activity, regio- and chemoselectivity based on computation-guided mutations. The promiscuous CYP72A63 was successfully remodeled into an enzyme oxidizing a specific C-H bond into hydroxyl, aldehyde and carboxylic acid, respectively. In addition, the catalytic activity of MdOSC1 for α-amyrin biosynthesis was optimized via homology modeling and docking [
45]. The triple mutant MdOSC1
N11T/P250H/P373A was obtained by semi-rational protein engineering, and the yield of α-amyrin was increased to 11-fold higher than that of the control.
3.2. Protein Fusion Expression
Protein fusion is a convenient strategy for enhancing enzyme catalytic efficiency through increasing the concentration of local intermediates, and efficiently redirecting metabolic flux to the desired product [
21,
27,
59]. For example, fusion of the double mutant Erg20p(F96W-N127W) with SpSabS1 resulted in a 3.5-fold increase in sabinene yield [
20]. The design addresses a significant challenge in yeast monoterpene production by tackling the sequential nature of the yeast farnesyl diphosphate synthase reaction, without affecting the growth characteristics of the tested yeast strains. In another report, co-expression of Erg20p and a-farnesene synthase (FS) with a GGGS amino acid linker improves the catalytic efficiency of α-farnesene synthase on FPP, avoiding FPP branching metabolism pathways and improving α-farnesene titer by 4.8-fold [
27]. The same group also indicated that fusion expression of Erg20p(F96W-N127W) and tVoGES after optimizing the linker led to a 23.5-fold increase in geraniol production compared to the strain with solely overexpressing the
tVoGES gene [
21]. The authors also observed a decrease in squalene accumulation compared to the strain overexpressing a single enzyme, indicating that the fusion of tVoGES-Erg20p(F96W-N127W) improved the GPP flux during geraniol production while reducing the GPP flux towards squalene synthesis pathway. Similarly, the fusion expression of GGPP synthase (Bts1p) and Erg20pF96C with GGGS linker was adopted to increase the GGPP accumulation [
4]. Meanwhile, the fusion expression of CfCYP76AH15, t66CfCPR and t30AaCYB5 not only decreased production of the intermediate 11-oxo-manoyl oxide, but also significantly increased the production of forskolin to 583.24 μg/L. Additionally, taxadiene production was enhanced up to 22-fold through fusion expression of maltose binding protein (MBP)-tagged taxadiene synthase (TASY) and Erg20p, along with effective copy number optimization [
36]. This can be attributed to the enhancement of protein expression and solubility, along with the increased availability of FPP for the synthetic pathway. Furthermore, fusion of β-carotene ketolase (CrtW) from
Paracoccus sp. with β-carotene hydroxylase (CrtZ) from
Haematococcus pluvialis minimized the substrate-enzyme distance, increased reaction rates, and obtained 2.8-fold improvement in the production of astaxanthin [
55]. The strain expressing the CrtW-Z fusion exhibited superior performance compared to the strain with the CrtZ-W fusion protein, indicating that the order of fusion proteins also plays a crucial role.
3.3. Directed Evolution of Enzymes
Directed evolution can also be used to improve the catalytic activity of enzymes through iterative mutations and screening of libraries. Mutant libraries can be constructed using traditional techniques such as error-prone PCR (ep-PCR) [
30], DNA shuffling and the staggered extension process (StEP) to achieve random mutagenesis [
61]. However, random mutagenesis-based directed evolution will generate a very large mutant library, encompassing numerous neutral and negative mutants alongside a few positive mutants. In order to accurately obtain positive mutants with desired traits from such a vast library, it is necessary to establish a fast, simple and reliable high-throughput screening method. Consequently, directed evolution finds its primary application in modifying reaction catalytic enzymes that can be linked to growth or pigment synthesis [
11,
30,
61]. For instance, a high-throughput screening method was developed based on precursor DMAPP toxicity and used for isoprene synthase (ISPS) directed evolution towards enhanced catalytic activity for isoprene production [
70]. Two positive mutants ISPS
F340L/I478V and ISPS
A570T were obtained from the mutant library generated by ep-PCR, and isoprene-forming activity were improved by 1.6-fold and 1.8-fold than the wild type, respectively. Another instance involves the engineering of a truncated linalool synthase (t67OMcLIS) from
M. citrata through directed evolution, facilitated by the development of a competition-based color-indicated high-throughput screening method [
11]. The variant t67OMcLIS
E343D/E352H, exhibiting increased enzyme activity, was successfully screened from a library generated by ep-PCR, resulting in a 52.7% increase of the linalool titer in
S. cerevisiae.
4. Harnessing Yeast Subcellular Compartments for Improving Terpenoid Biosynthesis
Yeast harbors various subcellular compartments, which offer numerous advantages that cannot be overlooked. These compartments possess the ability to concentrate substrates and enzymes, and provide a unique cellular environment that can respond to biosynthesis pathways with different needs, such as separating target synthesis pathways from competitive pathways and serving as storage sites for intermediate products that mitigate the toxic effect of their accumulation on host cells [
71]. Therefore, the use of subcellular compartments to synthesize terpenoids in yeast has attracted more and more attention.
4.1. Harnessing Mitochondria
Mitochondria, a double-membrane organelle, is rich in acetyl-CoA, ATP, and cofactors. Notably, the level of acetyl-CoA in mitochondria is 20–30 times higher than that in cytoplasm and nucleus, which is a promising site for terpenoid synthesis [
72]. By targeting the geraniol biosynthetic pathway into mitochondria, the geraniol precursor GPP was protected from depletion by the cytoplasmic ergosterol pathway, resulting in a six-fold increase in geraniol production compared to strains confined to the cytoplasm [
22]. In addition, SquM6 strain with squalene-specific titer of 79.78 mg/g DCW was obtained by locating the squalene synthesis pathway in mitochondria [
37]. However, the strain’s growth was significantly inhibited. To overcome this challenge, they successfully resolved the issue by overexpressing
tHMG1 in the cytoplasm. Finally, the squalene titer reached 21.1 g/L and the specific titer was 437.1 mg/g dcw after the two-stage fermentation process by combining cytoplasmic and mitochondrial engineering.
4.2. Harnessing Peroxisomes
Peroxisomes, single-layer membrane organelles, play a crucial role in generating a substantial pool of acetyl-CoA through β-oxidation. These organelles can also be genetically engineered to regulate their quantity and size [
42,
73], and their presence is not necessary for yeast to grow [
73]. Consequently, peroxisomes have emerged as attractive synthetic sites for natural product synthesis [
7]. By targeting 10 genes involved in the squalene synthesis pathway to the peroxisomes, the compartmentalization of squalene synthesis was achieved in
S. cerevisiae [
38]. Through further optimizing the supply of ATP, NADPH, acetyl-CoA in the peroxisome, and combining the dual cytoplasmic-peroxisomal engineering strategy, the squalene titer was significantly increased to 1698.02 mg/L and reached 11.00 g/L after a two-stage fed-batch fermentation. In another study, an increase in the number and copy number of peroxisomes was achieved by editing three peroxisome enzymes, Pex34p, Pex11p and Atg36p [
42]. Then by reconstructing the protopanaxadiol biosynthesis pathway in the peroxisome engineered strain, the protopanaxadiol yield was about 78% higher than that of the parent strain.
4.3. Harnessing Endoplasmic Reticulum
Endoplasmic reticulum (ER) is the base of a series of important biological macromolecules, such as proteins, lipids, sugars, and nucleic acids. It also provides a natural environment for many pathway enzymes and P450 enzymes involved in terpenoid synthesis. The expansion of the ER space offers several benefits, including enhancing the synthesis and folding capacity of ER proteins, alleviating metabolic constraints caused by limited enzyme abundance, and increasing the capacity of ER-targeted P450 and CPR enzymes. Currently, ER can be expanded in
S. cerevisiae by knocking out the phosphatidic acid phosphatase-encoding
PAH1 gene or by overexpressing INO2, a key regulator of ER size [
39,
50]. As an example, the deletion of
PAH1 gene stimulated the production of recombinant triterpene biosynthesis enzymes and resulted in a substantial increase in the accumulation of various triterpene skeletons [
50]. In the ER-engineered
pah1 yeast, the production of β-amyrin, medicagenic acid, and medicagenic-28-
O-glucoside was enhanced by eight-fold, six-fold, and 16-fold, respectively, compared to the wild-type strain. Furthermore, the impact of
PAH1 knockout was also evaluated for the production of other types of terpenoids. The results showed that
pah1 cells led to an approximately two-fold increase in sesquiterpene artemisinic acid production, but had an adverse effect on β-carotene production, which does not depend on ER-localized P450s. On the other hand, strains overexpressing
INO2 demonstrated a 71-fold increase in squalene production and an eight-fold increase in cytochrome P450-mediated protopanaxadiol production compared to strains without
INO2 overexpression [
39]. Transcriptome analysis revealed that in the ER-expanded strain, there was a comprehensive reorganization of central carbon metabolism, along with significant alterations in the transcriptional levels of ER-resident chaperones. These changes were suggested to enhance glucose uptake, improve protein folding, and facilitate the assembly of functional metabolic pathways.
4.4. Harnessing Lipid Droplets
In yeast, lipid droplets (LDs), consisting of triacylglycerols (TAGs) and sterol esters (SEs), are promising storage organelles for lipophilic compounds. Due to their hydrophobic properties, lipid droplets were also acted as a reservoir of terpenoids, while anchoring specific enzymes onto their surface facilitates efficient production of terpenoid scaffolds. For example, lipid droplet is the storage organelle of the cytochrome P450 enzyme protopanaxadiol (PPD) synthase (PPDS) substrate dammarenediol-II (DD) [
43]. By targeting PPDS to lipid droplets (LDs) using the yeast PLN1 protein, the efficiency of converting DD to PPD was significantly improved by 394%, with the conversion rate of DD increasing from 17.4% to 86.0%. In another work, it was demonstrated that lipid droplets size can be regulated by overexpression of fatty acid desaturase (
OLE1) and deletion of Seipin (
FLD1), in addition to overexpressing key genes involved in fatty acid synthesis and TAG production. These modifications led to a 25% increase in lycopene production compared to the original high-yield strain. The yield of lycopene in fed-batch fermentation reached 2.37 g/L and 73.3 mg/g cdw, respectively. In addition, the regulation of lipid metabolic pathways can significantly impact the accumulation of lipophilic compounds in yeast. In the case of microbial production of β-carotene, overexpressing the sterol ester synthesis genes
ARE1 and
ARE2 resulted in a 1.5-fold increase, while deleting the phosphatidate phosphatase genes (
PAH1,
DPP1, and
LPP1) led to a two-fold increase [
52]. By combining these two strategies, β-carotene production was amplified by 2.4 times compared to the initial strain. This study also provided valuable insights into the accumulation of other lipophilic compounds in yeast.
5. The Potential of Incorporating High Throughput Approaches for Improving Terpenoid Biosynthesis
Developing high-throughput techniques is a critical part of strain engineering, which allows researchers to rapidly screen a large number of microbial strains or genetic modifications, saving time and resources compared to traditional methods [
74]. One of the key advantages of high-throughput approaches is their ability to quickly and efficiently screen and identify optimal strains or genetic modifications that exhibit desired traits, such as higher productivity, yield, or stability. This enables researchers to quickly identify and select the most promising candidates for further development.
A commonly employed method for strain screening involves leveraging the inherent characteristics of the products, such as color, fluorescence, or growth status. For example, researchers established a fluorescence-activated cell sorting method that rapidly screened millions of mutants and identified novel gene targets involved in lipid metabolism, sterol metabolism, signal transduction, and stress response [
54]. This was achieved by analyzing the β-carotene content and fluorescence intensity of cells using HPLC and FCM, respectively. Notably, disrupting genes that affect fatty acid oxidation, lipid composition, and sterol transcriptional regulation (
4CL-8,
GCS, and
YIsterTF) increased β-carotene significantly. Finally, by engineering these targets in a high-β-carotene production, a strain that produced 9.4 g/L β-carotene was constructed. In the case of (+)-nootkatone, an important and expensive natural sesquiterpene compound, its microbial production is hindered by the low catalytic efficiency of its cytochrome P450-P450 reductase (HPO-CPR) system. To address this, a high-throughput screening (HTS) method was developed based on the color reaction between carbonyl compounds and 2,4-dinitrophenylhydrazine (DNPH) [
25]. This method enabled the rapid screening of candidate HPO mutants for activity. By optimizing the pairing of HPO and CPR and employing semi-rational design, an optimal mutant HPO_M18 with a catalytic performance 2.54 times higher than the initial strain was obtained.
High-throughput approaches involve the use of automated and rapid techniques to screen a large number of microbial strains or genetic modifications for desired traits or characteristics. An automated high-throughput screening platform based on HPLC and UPLC-MS analytical techniques was established to functionally identify hundreds of
Ganoderma lucidum cytochromes P450 in
S. cerevisiae [
48]. This platform enabled the successful achievement of a series of highly efficient heterologous biosynthesis of ganoderic acid in
S. cerevisiae, along with the identification of key genes involved in the ganoderic acid biosynthetic pathway. Specifically, the catalytic reaction process of CYP512W2 and the formation of type II ganoderic acid were thoroughly investigated. Validation through shake flask fermentation of the high-yielding type II ganoderic acid strains, obtained after metabolic modification, showed that the yields of type II ganoderic acid reached more than 50 mg/L in all cases. This represents a significant improvement compared to the traditional artificial cultivation production method, with ganoderic acid yields increased by 1–4 orders of magnitude and the production efficiency enhanced by 2–5 orders of magnitude. Similarly, the screening progress of microbial colonies was expedited through the direct evaluation of microbial bioproduction colonies using supercritical fluid extraction-supercritical fluid chromatography-triple quadrupole mass spectrometry (SFE-SFC-MS/MS) [
75]. The online SFE-SFC-MS/MS system offers great potential for high-throughput analyses, due to the automated extraction of metabolites without the need for pre-processing. The SFE-SFC-MS/MS system enables faster and easier screening of highly productive strains compared to conventional analyses, as demonstrated by the high-throughput screening of (−)--limonene bioproducers.
6. The Potential of Incorporating Global Approaches for Improving Terpenoid Biosynthesis
Global approaches aim to investigate biological systems in a holistic manner, involving the elucidation of mechanisms underlying the cellular behavior of specific cell factories or the simulation of the behavior of these systems. Incorporating global approaches has immense potential for improving the performance of microbial cell factories [
76]. By adopting a holistic perspective and considering the entire biological system, these approaches can provide valuable insights and strategies for enhancing the performance of microbial cells in industrial settings. With the rapid development of several advanced techniques, such as omics technologies and genome-scale models, several global approaches are employed for improving terpenoid biosynthesis.
Through extensive studies of transcriptomics, proteomics and metabolomics, we have gained a comprehensive understanding of the metabolic networks and regulatory mechanisms in both natural producers and engineered yeast strains. This approach enables the integration of genome-scale experimental data to uncover the underlying mechanisms, identify targets for optimization, and guide the design of genetic modifications or environmental conditions to enhance strain performance. For example, in yeast, out of the 1,470 presumed N6-Methyladenosine (m6A) peaks identified, it was observed that overexpression of IME4 (yeast m6A methyltransferase) led to significantly altered transcription levels in 94 pathway genes [
77]. In particular, IME4 overexpression elevates the mRNA levels of the methylated genes in the glycolysis, acetyl-CoA synthesis and shikimate/aromatic amino acid synthesis modules, which provides essential precursors and acetyl-CoA for terpene synthesis.
Phaffia rhodozyma is a potential strain for the biosynthesis of astaxanthin. However, the unclear metabolic characteristics of
P. rhodozyma at different metabolic stages have hindered the efficient production of astaxanthin. To shed light on this, a quadrupole time-of-flight mass spectrometry metabolomics approach was employed to investigate the changes in metabolites [
78]. The results showed that the down-regulation of purine, pyrimidine, amino acid synthesis and glycolysis pathways played a crucial role in astaxanthin biosynthesis. Similarly, a comprehensive analysis of differential metabolites and expression genes was conducted to explore the effects of gibberellin-induced astaxanthin synthesis, combining transcriptomics and metabolomics [
57]. The results showed that the application of gibberellic acid (GA) significantly enhanced fatty acid metabolism and ABC transporters, and increased unsaturated fatty acid content. This favorable alteration facilitated the accumulation of astaxanthin, thereby promoting its synthesis.
Genome-scale metabolic models (GEMs) have emerged as a computational systems biology approach for effectively interpreting and integrating multi-omic data. By converting the organism’s reactions related to metabolism, transcription, and translation into a mathematical formulation, GEMs can be modeled using optimization principles [
79]. This modeling approach enables the optimization of microbial growth-associated traits by simulating biomass formation, predicting growth rates, and identifying the requirements for cell growth. Moreover, GEMs empower cells to enhance product formation by predicting gene knockout and overexpression targets using modeling tools. For instance, a GEMs was used to identify new target genes for enhancing biosynthesis of sesquiterpenes in the
S. cerevisiae [
31]. The impact of gene deletions on flux distributions in the metabolic model of
S. cerevisiae was assessed using OptGene as the modeling framework and minimization of metabolic adjustments (MOMA) as objective function. The results revealed that the deletion of GDH1 (gene encoding NADPH-dependent glutamate dehydrogenase) was the most promising target gene for improving sesquiterpene biosynthesis in yeast. This deletion increased the availability of NADPH in the cytosol for other NADPH requiring enzymes, including HMG-CoA reductase, resulting in an approximately 85% increase in the final cubebol titer. In another work, two prediction tools, namely FOCuS (a novel gene deletion tool) and OptKnock (a standard gene deletion prediction tool), were employed to identify knockout targets for the improvement of terpenoid pathway fluxes [
80]. Notably, key knockout targets were predicted, including
LYS1,
GAP1,
AAT1,
AAT2,
TH17,
KGD-m,
MET14,
PDC1 and
ACO1, which are associated with fatty acid biosynthesis, amino acid synthesis pathways or nucleotide biosynthesis pathways. Similarly, overexpression targets such as
PFK1,
FBA1,
ZWF1,
TDH1,
PYC1,
ALD6,
TPI1,
PDX1 and
ENO1, which are involved in glycolytic and pentose phosphate pathways, were identified. Each of these targets had plausible role in improving flux toward sterol pathway and appeared to be meaningful rather than artifacts. In this regard, an
in vitro study was conducted to validate the predictions by overexpressing
ALD6 and
TPI1. The results demonstrated a 2.23- and 4.24-fold increase in squalene synthesis, respectively, compared to the control. In general, the rationale behind predicting these in silico targets was attributed to either increase the acetyl-CoA precursor pool or regenerate NADPH, both of which enhance the flux in the sterol pathway.
GEM can also be used to optimize fermentation media, offering a more efficient alternative to the laborious statistical design methods such as the Plackett-Burman design. By combining GEM (FBA) results with statistical methods such as the Plackett-Burman design, it becomes possible to develop a highly effective culture medium. Using this method, chemicals for lycopene production were screened in metabolically engineered strains [
51]. In fed-batch cultivation mode, lycopene concentrations of 126 and 242 mg/L were achieved using the FBA-independent and FBA-assisted designed media, respectively. Transcriptional studies further revealed upregulations of heterologous genes in media design based on FBA, providing evidence for the predictive power of the model.
With the rapid increase in volume and complexity of data generated from omics studies, the prevalence of extensive data in the domain of multi-omics studies necessitates elaborate computational analysis. Machine learning (ML), a data-driven approach, serves as a computational tool that aims to analyze input data, identify patterns, and generate meaningful insights in the output. ML has been applied to a wide range of biological problems [
81], such as predicting pathway dynamics from omics data [
82], determining transcription regulatory dynamics [
83], and predicting genotype-phenotype associations [
84]. Thus, the application of ML holds great promise in evaluating strain design strategies. For example, to investigate the individual contributions of five non-rate-limiting enzymes in the MVA pathway, a combinatorial library of 243
S. cerevisiae strains was created, each with an additional copy of the MVA pathway integrated into the genome and expressing the non-rate-limiting enzymes through a unique combination of promoters [
85]. Through high-throughput screening combined with ML algorithm, it was revealed that Erg12p was the key enzyme in affecting the titer of the product. Optimal expression of
ERG12 was achieved using a medium-strength promoter, which represents the ideal ‘sweet spot’ for attaining high product yield. In addition, a platform strain was created by targeting the mevalonate pathway to both the cytosol and peroxisomes. By employing this dual localization approach, terpenoid production was synergistically enhanced, resulting in a remarkable 94-fold increase in monoterpene geraniol, a 60-fold increase in sesquiterpene α-humulene, and a 35-fold increase in triterpene squalene titers, respectively. This platform strain, with its enhanced capabilities, can serve as a versatile chassis for the production of various terpenoids and terpene derivatives.
7. Prospects and Conclusions
Terpenoids, which are natural secondary metabolites with various biological activities, possess immense potential and commercial value. The rapid advancements in synthetic biology, metabolic engineering, DNA sequencing and omics analysis have facilitated the sustainable and efficient production of a wide range of terpenoids through the heterologous expression of terpene synthases in various microbial chassis, such as
S. cerevisiae and
Y. lipolytica. However, there are still several challenges need to be addressed. Many high-value terpenoids and their derivatives with relatively complex structures and synthetic pathways have not been heterologous synthesized. Even for terpenoids that have been heterologously synthesized, there is still a significant gap between the production capacity of microbial cell factories and the requirements of industrial-scale production. The main obstacles to large-scale production of terpenoids mainly include the following aspects: (a) inadequate supply of precursor substrates, energy and cofactors; (b) low catalytic efficiency and poor substrate/product specificity of terpene synthases; (c) limited understanding of natural synthetic pathways and functions of key enzymes; (d) the incompatibility between heterologous pathways and chassis cells due to differences in intracellular microenvironment.
In response to these challenges, several strategies have demonstrated their effectiveness. Firstly, a combination of metabolic engineering approaches can be utilized. This includes enhancing precursor supply by introducing alternative heterologous acetyl-CoA pathways [
26] and IUP pathway [
18,
33], optimizing the endogenous MVA pathway [
23,
32,
44] and PP pathway [
41,
53], and improving and balancing the NADPH supply [
26,
41] to increase carbon fluxes toward terpenoid synthesis. Secondly, to optimize the expression of enzymes, successive enzyme colocalization strategies such as enzyme fusion [
21,
34,
36], enzyme scaffolds [
2], organelle localization [
24,
55] can be employed. These strategies aim to enhance substrate conversion efficiency and minimize the diversion of metabolic intermediates into competing pathways. Thirdly, a comprehensive understanding of the metabolic network of natural producers and the regulatory mechanism of engineering yeast strains can be achieved through the in-depth studies of omics techniques [
37,
78,
79]. Fourthly, the adaptation mechanism between heterologous pathways and yeast cells can be analyzed to achieve a balance between cell metabolism and heterologous product synthesis through protein and metabolic engineering [
9]. Additionally, the storage and transport mechanisms of synthesized terpene products in yeast can be investigated to maximize the utilization of intracellular resources [
38,
86]. Techniques such as subcellular localization [
28] and compartmentalization regulation can be employed to create more space for storing hydrophobic terpenoids by expanding the membrane structure of yeast cells [
4,
8,
42,
43]. Exploring specific transporters for the efflux of toxic products can reduce metabolic stress on cells and improve production sustainability [
86].
In the future, the development of more efficient multi-fragment and long-fragment DNA assembly, as well as genome editing techniques, can be used to realize heterologous synthesis of more complex terpenoids in yeast. Simultaneously, methods of high-throughput screening need to be developed for real-time detection of intermediate metabolites and intracellular products. Furthermore, the construction of an automated and intelligent synthetic biology platform can expedite the assembly and optimization process of complex synthetic pathways for terpenoid production in yeast cells. These platforms can shorten the design-build-test-learning cycle of cell factories, leading to the construction of efficient, stable, and controllable yeast cell factories for terpenoid biosynthesis.
Author Contributions
Y.X. and C.L. drafted and revised the manuscript; S.S. revised the manuscript; Y.X. and R.C. filled the table and drew figure; L.Q. drew figure.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This work is supported by Key-Area Research and Development Program of Guangdong Province (2022B1111080003) and Tianjin Synthetic Biotechnology Innovation Capacity Improvement Project (TSBICIP-KJGG-009). This work was funded by Beijing University of Chemical Technology, the central university of basic research funds (BH2339).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Zhang Y, Nielsen J, Liu Z. Engineering yeast metabolism for production of terpenoids for use as perfume ingredients, pharmaceuticals and biofuels.
FEMS Yeast Res. 2017,
17, fox080.
[Google Scholar]
-
2.
Jiang G, Yao M, Wang Y, Xiao W, Yuan Y. A “push-pull-restrain” strategy to improve citronellol production in
Saccharomyces cerevisiae.
Metab. Eng. 2021,
66, 51–59.
[Google Scholar]
-
3.
Meng X, Liu H, Xu W, Zhang W, Wang Z, Liu W. Metabolic engineering
Saccharomyces cerevisiae for de novo production of the sesquiterpenoid (+)-nootkatone.
Microb. Cell Factories 2020,
60, 25–36.
[Google Scholar]
-
4.
Ju H, Zhang C, He S, Nan W, Lu W. Construction and optimization of Saccharomyces cerevisiae for synthesizing forskolin.
Appl. Microbiol. Biotechnol. 2022,
106, 1933–1944.
[Google Scholar]
-
5.
Lim SH, Baek JI, Jeon BM, Seo JW, Kim MS, Byun JY, et al. CRISPRi-guided metabolic flux engineering for enhanced protopanaxadiol production in
Saccharomyces cerevisiae.
Int. J. Mol. Sci. 2021,
22, 11836.
[Google Scholar]
-
6.
Cataldo VF, Arenas N, Salgado V, Camilo C, Ibanez F, Agosin E. Heterologous production of the epoxycarotenoid violaxanthin in
Saccharomyces cerevisiae.
Metab. Eng. 2020,
59, 53–63.
[Google Scholar]
-
7.
Dusséaux S, Wajn WT, Liu Y, Ignea C, Kampranis SC. Transforming yeast peroxisomes into microfactories for the efficient production of high-value isoprenoids.
Proc. Natl. Acad. Sci. USA 2020,
117, 31789–31799.
[Google Scholar]
-
8.
Ma T, Shi B, Ye Z, Li X, Liu M, Chen Y, et al. Lipid engineering combined with systematic metabolic engineering of Saccharomyces cerevisiae for high-yield production of lycopene.
Metab. Eng. 2019,
52, 134–142.
[Google Scholar]
-
9.
Bian Q, Zhou P, Yao Z, Li M, Yu H, Ye L. Heterologous biosynthesis of lutein in
S. cerevisiae enabled by temporospatial pathway control.
Metab. Eng. 2021,
67, 19–28.
[Google Scholar]
-
10.
Gerke J, Frauendorf H, Schneider D, Wintergoller M, Hofmeister T, Poehlein A, et al. Production of the fragrance geraniol in peroxisomes of a product-tolerant baker’s yeast.
Front. Bioeng. Biotechnol. 2020,
8, 582052.
[Google Scholar]
-
11.
Zhou P, Du Y, Xu N, Yue C, Ye L. Improved linalool production in
Saccharomyces cerevisiae by combining directed evolution of linalool synthase and overexpression of the complete mevalonate pathway.
Biochem. Eng. J. 2020,
161, 107655.
[Google Scholar]
-
12.
Otto M, Teixeira PG, Vizcaino MI, David F, Siewers V. Integration of a multi-step heterologous pathway in Saccharomyces cerevisiae for the production of abscisic acid.
Microb. Cell Factories 2019,
18, 205.
[Google Scholar]
-
13.
Mitsui R, Nishikawa R, Yamada R, Matsumoto T, Ogino H. Construction of yeast producing patchoulol by global metabolic engineering strategy.
Biotechnol. Bioeng. 2020,
117, 1348–1356.
[Google Scholar]
-
14.
Chemler JA, Koffas MAG. Metabolic engineering for plant natural product biosynthesis in microbes.
Curr. Opin. Biotechnol. 2008,
19, 597–605.
[Google Scholar]
-
15.
Li ZJ, Wang YZ, Wang LR, Shi TQ, Sun XM, Huang H. Advanced strategies for the synthesis of terpenoids in
Yarrowia lipolytica.
J. Agric. Food Chem. 2021,
69, 2367–2381.
[Google Scholar]
-
16.
Sun W, Qin L, Xue H, Yu Y, Ma Y, Wang Y, et al. Novel trends for producing plant triterpenoids in yeast.
Crit. Rev. Biotechnol. 2019,
39, 618–632.
[Google Scholar]
-
17.
Mitsui R, Yamada R. Saccharomyces cerevisiae as a microbial cell factory. In Microbial Cell Factories Engineering for Production of Biomolecules; Academic Press, Cambridge, MA, USA; 2021; pp. 319–333.
-
18.
Ma Y, Zu Y, Huang S, Stephanopoulos G. Engineering a universal and efficient platform for terpenoid synthesis in yeast.
Proc. Natl. Acad. Sci. USA 2022,
120, e2207680120.
[Google Scholar]
-
19.
Cao X, Wei LJ, Lin JY, Hua Q. Enhancing linalool production by engineering oleaginous yeast
Yarrowia lipolytica. Bioresour. Technol. 2017,
245, 1641–1644.
[Google Scholar]
-
20.
Ignea C, Pontini M, Maffei ME, Makris AM, Kampranis SC. Engineering monoterpene production in yeast using a synthetic dominant negative geranyl diphosphate synthase.
ACS Synth. Biol. 2014,
3, 298–306.
[Google Scholar]
-
21.
Zhao J, Bao X, Li C, Shen Y, Hou J. Improving monoterpene geraniol production through geranyl diphosphate synthesis regulation in
Saccharomyces cerevisiae.
Appl. Microbiol. Biotechnol. 2016,
100, 4561–4571.
[Google Scholar]
-
22.
Yee DA, DeNicola AB, Billingsley JM, Creso JG, Subrahmanyam V, Tang Y. Engineered mitochondrial production of monoterpenes in
Saccharomyces cerevisiae.
Metab. Eng. 2019,
55, 76–84.
[Google Scholar]
-
23.
Ignea C, Cvetkovic I, Loupassaki S, Kefalas P, Johnson CB, Kampranis SC, et al. Improving yeast strains using recyclable integration cassettes, for the production of plant terpenoids.
Microb. Cell Fact. 2011,
10, 4.
[Google Scholar]
-
24.
Dong C, Shi Z, Huang L, Zhao H, Xu Z, Lian J. Cloning and characterization of a panel of mitochondrial targeting sequences for compartmentalization engineering in
Saccharomyces cerevisiae.
Biotechnol. Bioeng. 2021,
118, 4269–4277.
[Google Scholar]
-
25.
Liu T, Li W, Chen H, Wu T, Zhu C, Zhuo M, et al. Systematic optimization of HPO-CPR to boost (+)-nootkatone synthesis in engineered
Saccharomyces cerevisiae.
J. Agric. Food Chem. 2022,
70, 15548–15559.
[Google Scholar]
-
26.
Meadows AL, Hawkins KM, Tsegaye Y, Antipov E, Kim Y, Raetz L, et al. Rewriting yeast central carbon metabolism for industrial isoprenoid production.
Nature 2016,
537, 694–697.
[Google Scholar]
-
27.
Liu Y, Jiang X, Cui Z, Wang Z, Qi Q, Hou J. Engineering the oleaginous yeast
Yarrowia lipolytica for production of α-farnesene.
Biotechnol. Biofuels 2019,
12, 296.
[Google Scholar]
-
28.
Guo Q, Shi TQ, Peng QQ, Sun XM, Ji XJ, Huang H. Harnessing
Yarrowia lipolytica peroxisomes as a subcellular factory for alpha-humulene overproduction.
J. Agric. Food Chem. 2021,
69, 13831–13837.
[Google Scholar]
-
29.
Peng B, Plan MR, Chrysanthopoulos P, Hodson MP, Nielsen LK, Vickers CE. A squalene synthase protein degradation method for improved sesquiterpene production in
Saccharomyces cerevisiae.
Metab. Eng. 2017,
39, 209–219.
[Google Scholar]
-
30.
Lu S, Deng H, Zhou C, Du Z, Guo X, Cheng Y, et al. Enhancement of β-caryophyllene biosynthesis in
Saccharomyces cerevisiae via synergistic evolution of β-caryophyllene synthase and engineering the chassis.
ACS Synth. Biol. 2023,
12, 1696–1707.
[Google Scholar]
-
31.
Asadollahi MA, Maury J, Patil KR, Schalk M, Clark A, Nielsen J. Enhancing sesquiterpene production in
Saccharomyces cerevisiae through in silico driven metabolic engineering.
Metab. Eng. 2009,
11, 328–334.
[Google Scholar]
-
32.
Zhao Y, Zhu K, Li J, Zhao Y, Li S, Zhang C, et al. High‐efficiency production of bisabolene from waste cooking oil by metabolically engineered
Yarrowia lipolytica.
Microb. Biotechnol. 2021,
14, 2497–2513.
[Google Scholar]
-
33.
Wang J, Zhu L, Li Y, Xu S, Jiang W, Liang C, et al. Enhancing geranylgeraniol production by metabolic engineering and utilization of isoprenol as a substrate in
Saccharomyces cerevisiae.
J. Agric. Food Chem. 2021,
69, 4480–4489.
[Google Scholar]
-
34.
Hu T, Zhou J, Tong Y, Su P, Li X, Liu Y, et al. Engineering chimeric diterpene synthases and isoprenoid biosynthetic pathways enables high-level production of miltiradiene in yeast.
Metab. Eng. 2020,
60, 87–96.
[Google Scholar]
-
35.
Cao X, Yu W, Chen Y, Yang SC, Zhao ZK, Nielsen J, et al. Engineering yeast for high-level production of diterpenoid sclareol.
Metab. Eng. 2023,
75, 19–28.
[Google Scholar]
-
36.
Nowrouzi B, Li RA, Walls LE, d’Espaux L, Malci K, Liang L, et al. Enhanced production of taxadiene in
Saccharomyces cerevisiae.
Microb. Cell Factories 2020,
19, 200.
[Google Scholar]
-
37.
Zhu ZT, Du MM, Gao B, Tao XY, Zhao M, Ren YH, et al. Metabolic compartmentalization in yeast mitochondria: Burden and solution for squalene overproduction.
Metab. Eng. 2021,
68, 232–245.
[Google Scholar]
-
38.
Liu GS, Li T, Zhou W, Jiang M, Tao XY, Liu M, et al. The yeast peroxisome: A dynamic storage depot and subcellular factory for squalene overproduction.
Metab. Eng. 2020,
57, 151–161.
[Google Scholar]
-
39.
Kim JE, Jang IS, Son SH, Ko YJ, Cho BK, Kim SC, et al. Tailoring the
Saccharomyces cerevisiae endoplasmic reticulum for functional assembly of terpene synthesis pathway.
Metab. Eng. 2019,
56, 50–59.
[Google Scholar]
-
40.
Zhao F, Bai P, Nan W, Li D, Zhang C, Lu C, et al. A modular engineering strategy for high‐level production of protopanaxadiol from ethanol by
Saccharomyces cerevisiae.
AIChE J. 2018,
65, 866–874.
[Google Scholar]
-
41.
Kim JE, Jang IS, Sung BH, Kim SC, Lee JY. Rerouting of NADPH synthetic pathways for increased protopanaxadiol production in
Saccharomyces cerevisiae.
Sci. Rep. 2018,
8, 15820.
[Google Scholar]
-
42.
Choi B, Kang H, Kim S, Lee P. Organelle engineering in yeast: Enhanced production of protopanaxadiol through manipulation of peroxisome proliferation in
Saccharomyces cerevisiae.
Microorganisms 2022,
10, 650.
[Google Scholar]
-
43.
Shi Y, Wang D, Li R, Huang L, Dai Z, Zhang X. Engineering yeast subcellular compartments for increased production of the lipophilic natural products ginsenosides.
Metab. Eng. 2021,
67, 104–111.
[Google Scholar]
-
44.
Wang P, Wei W, Ye W, Li X, Zhao W, Yang C, et al. Synthesizing ginsenoside Rh2 in
Saccharomyces cerevisiae cell factory at high-efficiency.
Cell Discov. 2019,
5, 5.
[Google Scholar]
-
45.
Yu Y, Rasool A, Liu H, Lv B, Chang P, Song H, et al. Engineering
Saccharomyces cerevisiae for high yield production of α-amyrin via synergistic remodeling of α-amyrin synthase and expanding the storage pool.
Metab. Eng. 2020,
62, 72–83.
[Google Scholar]
-
46.
Liu H, Fan J, Wang C, Li C, Zhou X. Enhanced β-amyrin synthesis in
Saccharomyces cerevisiae by coupling an optimal acetyl-CoA supply pathway.
J. Agr. Food Chem. 2019,
67, 3723–3732.
[Google Scholar]
-
47.
Du MM, Zhang GG, Zhu ZT, Zhao YQ, Gao B, Tao XY, et al. Boosting the epoxidation of squalene to produce triterpenoids in
Saccharomyces cerevisiae.
Biotechnol. Biofuels 2023,
16, 76.
[Google Scholar]
-
48.
Yuan W, Jiang C, Wang Q, Fang Y, Wang J, Wang M, et al. Biosynthesis of mushroom-derived type II ganoderic acids by engineered yeast.
Nat. Commun. 2022,
13, 7740.
[Google Scholar]
-
49.
Jin K, Shi X, Liu J, Yu W, Liu Y, Li J, et al. Combinatorial metabolic engineering enables the efficient production of ursolic acid and oleanolic acid in
Saccharomyces cerevisiae.
Bioresour. Technol. 2023,
374, 128819.
[Google Scholar]
-
50.
Arendt P, Miettinen K, Pollier J, De Rycke R, Callewaert N, Goossens A. An endoplasmic reticulum-engineered yeast platform for overproduction of triterpenoids.
Metab. Eng. 2017,
40, 165–175.
[Google Scholar]
-
51.
Nambou K, Jian X, Zhang X, Wei L, Lou J, Madzak C, et al. Flux balance analysis inspired bioprocess upgrading for lycopene production by a metabolically engineered strain of
Yarrowia lipolytica.
Metabolites 2015,
5, 794–813.
[Google Scholar]
-
52.
Zhao Y, Zhang Y, Nielsen J, Liu Z. Production of β‐carotene in Saccharomyces cerevisiae through altering yeast lipid metabolism.
Biotechnol. Bioeng. 2021,
118, 2043–2052.
[Google Scholar]
-
53.
Zhao X, Shi F, Zhan W. Overexpression of ZWF1 and POS5 improves carotenoid biosynthesis in recombinant
Saccharomyces cerevisiae.
Lett. Appl. Microbiol. 2015,
61, 354–360.
[Google Scholar]
-
54.
Liu M, Zhang J, Liu X, Hou J, Qia Q. Rapid gene target tracking for enhancing β-carotene production using flow cytometry-based high-throughput screening in
Yarrowia lipolytica.
Appl. Environ. Microbiol. 2022,
88, e0114922.
[Google Scholar]
-
55.
Ma Y, Li J, Huang S, Stephanopoulos G. Targeting pathway expression to subcellular organelles improves astaxanthin synthesis in
Yarrowia lipolytica.
Metab. Eng. 2021,
68, 152–161.
[Google Scholar]
-
56.
Kildegaard KR, Adiego-Pérez B, Doménech Belda D, Khangura JK, Holkenbrink C, Borodina I. Engineering of
Yarrowia lipolytica for production of astaxanthin.
Synth. Syst. Biotechnol. 2017,
2, 287–294.
[Google Scholar]
-
57.
Liu S, Yi H, Zhan H, Wang L, Wang J, Li Y, et al. Gibberellic acid‐induced fatty acid metabolism and ABC transporters promote astaxanthin production in
Phaffia rhodozyma.
J. Appl. Microbiol. 2021,
132, 390–400.
[Google Scholar]
-
58.
Liscum L, Finer-Moore J, Stroud RM, Luskey KL, Brown MS, Goldstein JL. Domain structure of 3-hydroxy-3-methylglutaryl coenzyme A reductase, a glycoprotein of the endoplasmic reticulum.
J. Biol. Chem. 1985,
260, 522–530.
[Google Scholar]
-
59.
Zhang G, Wang H, Zhang Z, Verstrepen KJ, Wang Q, Dai Z. Metabolic engineering of Yarrowia lipolytica for terpenoids production: advances and perspectives.
Crit. Rev. Biotechnol. 2022,
42, 618–633.
[Google Scholar]
-
60.
Mantzouridou F, Tsimidou MZ. Observations on squalene accumulation in Saccharomyces cerevisiae due to the manipulation of HMG2 and ERG6.
FEMS Yeast Res. 2010,
10, 699–707.
[Google Scholar]
-
61.
Li W, Cui L, Mai J, Shi TQ, Lin L, Zhang ZG, et al. Advances in metabolic engineering paving the way for the efficient biosynthesis of terpenes in yeasts.
J. Agric. Food Chem. 2022,
70, 9246–9261.
[Google Scholar]
-
62.
Chatzivasileiou AO, Ward V, Edgar SM, Stephanopoulos G. Two-step pathway for isoprenoid synthesis.
Proc. Natl. Acad. Sci. USA 2019,
116, 506–511.
[Google Scholar]
-
63.
Zhang C, Hong K. Production of terpenoids by synthetic biology approaches.
Biotechnol. Bioeng. 2020,
8, 347.
[Google Scholar]
-
64.
Kwak S, Yun EJ, Lane S, Oh EJ, Kim KH, Jin YS. Redirection of the glycolytic flux enhances isoprenoid production in
Saccharomyces cerevisiae.
Biotechnol. J. 2019,
15, e1900173.
[Google Scholar]
-
65.
Dinday S, Ghosh S. Recent advances in triterpenoid pathway elucidation and engineering.
Biotechnol. Adv. 2023,
68, 108214.
[Google Scholar]
-
66.
Kirby J, Romanini DW, Paradise EM, Keasling JD. Engineering triterpene production in
Saccharomyces cerevisiae–β‐amyrin synthase from
Artemisia annua.
FEBS J. 2008,
275, 1852–1859.
[Google Scholar]
-
67.
Bröker JN, Müller B, van Deenen N, Prüfer D, Schulze Gronover C. Upregulating the mevalonate pathway and repressing sterol synthesis in
Saccharomyces cerevisiae enhances the production of triterpenes.
Appl. Microbiol. Biotechnol. 2018,
102, 6923–6934.
[Google Scholar]
-
68.
Guo H, Wang H, Chen T, Guo L, Blank LM, Ebert BE, et al. Engineering critical amino acid residues of lanosterol synthase to improve the production of triterpenoids in
Saccharomyces cerevisiae.
ACS Synth. Biol. 2022,
11, 2685–2696.
[Google Scholar]
-
69.
Sun W, Xue H, Liu H, Lv B, Yu Y, Wang Y, et al. Controlling chemo- and regioselectivity of a plant P450 in yeast cell toward rare licorice triterpenoid biosynthesis.
ACS Catal. 2020,
10, 4253–4260.
[Google Scholar]
-
70.
Wang F, Lv X, Xie W, Zhou P, Zhu Y, Yao Z, et al. Combining Gal4p-mediated expression enhancement and directed evolution of isoprene synthase to improve isoprene production in
Saccharomyces cerevisiae.
Metab. Eng. 2017,
39, 257–266.
[Google Scholar]
-
71.
Hammer SK, Avalos JL. Harnessing yeast organelles for metabolic engineering.
Nat. Chem. Biol. 2017,
13, 823–832.
[Google Scholar]
-
72.
Weinert BT, Iesmantavicius V, Moustafa T, Schölz C, Wagner SA, Magnes C, et al. Acetylation dynamics and stoichiometry in
Saccharomyces cerevisiae.
Mol. Syst. Biol. 2014,
10, 833.
[Google Scholar]
-
73.
Van der Klei IJ, Veenhuis M. Yeast peroxisomes: Function and biogenesis of a versatile cell organelle.
Trends Microbiol. 1997,
5, 502–509.
[Google Scholar]
-
74.
Leavell MD, Singh AH, Kaufmann-Malaga BB. High-throughput screening for improved microbial cell factories, perspective and promise.
Curr. Opin. Biotechnol. 2020,
62, 22–28.
[Google Scholar]
-
75.
Takekana M, Yoshida T, Yoshida E, Ono S, Horie S, Vavricka CJ, et al. Online SFE-SFC-MS/MS colony screening: A high-throughput approach for optimizing (-)-limonene production.
J. Chromatogr. B 2023,
1215, 123588.
[Google Scholar]
-
76.
Ko YS, Kim JW, Lee JA, Han T, Kim GB, Park JE, et al. Tools and strategies of systems metabolic engineering for the development of microbial cell factories for chemical production.
Chem. Soc. Rev. 2020,
49, 4615–4636.
[Google Scholar]
-
77.
Zhu J, An T, Zha W, Gao K, Li T, Zi J. Manipulation of IME4 expression, a global regulation strategy for metabolic engineering in
Saccharomyces cerevisiae.
Acta Pharm. Sin. B 2023,
13, 2795–2806.
[Google Scholar]
-
78.
Yang H, Yang L, Du X, He N, Jiang Z, Zhu Y, et al. Metabolomics of astaxanthin biosynthesis and corresponding regulation strategies in
Phaffia rhodozyma.
Yeast 2023,
40, 254–264.
[Google Scholar]
-
79.
Dahal S, Yurkovich JT, Xu H, Palsson BO, Yang L. Synthesizing systems biology knowledge from omics using genome-scale models.
Proteomics 2020,
20, 1900282.
[Google Scholar]
-
80.
Paramasivan K, Abdulla A, Gupta N, Mutturi S. In silico target-based strain engineering of
Saccharomyces cerevisiae for terpene precursor improvement.
Integr. Biol. 2022,
14, 25–36.
[Google Scholar]
-
81.
Kim GB, Kim WJ, Kim HU, Lee SY. Machine learning applications in systems metabolic engineering.
Curr. Opin. Biotechnol. 2020,
64, 1–9.
[Google Scholar]
-
82.
Costello Z, Martin HG. A machine learning approach to predict metabolic pathway dynamics from time-series multiomics data.
NPJ Syst. Biol. Appl. 2018,
4, 19.
[Google Scholar]
-
83.
Chen L, Cai C, Chen V, Lu X. Learning a hierarchical representation of the yeast transcriptomic machinery using an autoencoder model.
BMC Bioinformat. 2016,
17, 9.
[Google Scholar]
-
84.
Ma J, Yu MK, Fong S, Ono K, Sage E, Demchak B, et al. Using deep learning to model the hierarchical structure and function of a cell.
Nat. Methods 2018,
15, 290–298.
[Google Scholar]
-
85.
Mukherjee M, Blair RH, Wang ZQ. Machine-learning guided elucidation of contribution of individual steps in the mevalonate pathway and construction of a yeast platform strain for terpenoid production.
Metab. Eng. 2022,
74, 139–149.
[Google Scholar]
-
86.
Chen M, Li M, Ye L, Yu H. Construction of canthaxanthin-producing yeast by combining spatiotemporal regulation and pleiotropic drug resistance engineering.
ACS Synth. Biol. 2022,
11, 325–333.
[Google Scholar]