Advances in Gastrodin Production: From Native to Engineered Biosynthesis

Advances in Gastrodin Production: From Native to Engineered Biosynthesis
Xun Wang 1,2 Ziwei Jing 1,2 Fei Wang 1,2 Zhiguo Wang 1,2 Xun Li 1,2,*
Received: 03 June 2026 Revised: 18 June 2026 Accepted: 23 June 2026 Published: 29 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
Graphical Abstract
1. Introduction
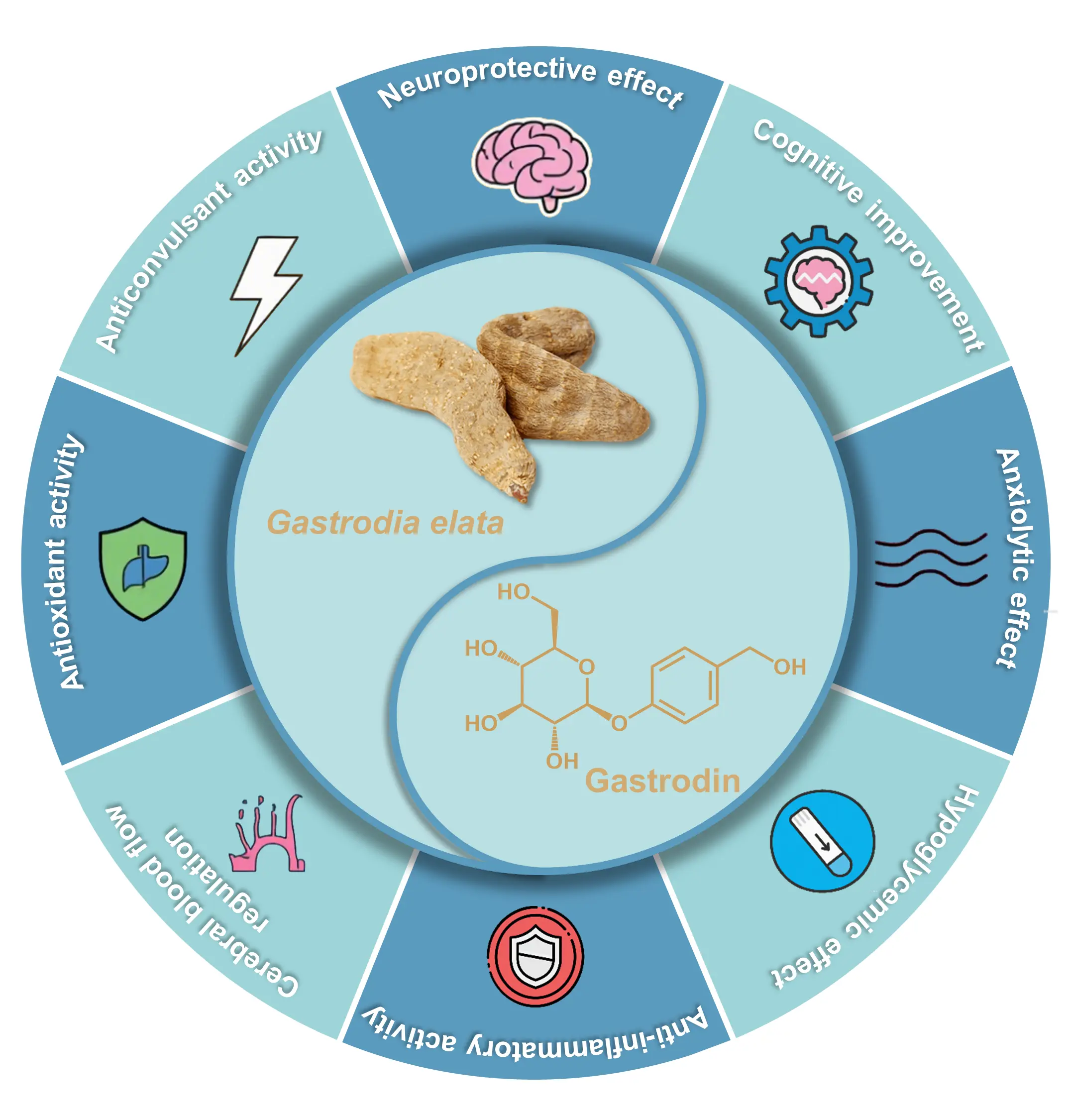
Gastrodia elata is a well-known traditional medicinal herb belonging to the orchid family [1]. It was first recorded in Classic of Materia Medica and classified among the superior herbs [2]. As a perennial plant with more than two thousand years of medicinal history [3], G. elata is mainly distributed across East and South Asia, including China, Japan, Korea, Bhutan, and India [4]. Because of its neuroprotective [5], analgesic [6], anticonvulsant [7], and anxiolytic [8] effects, it has been widely applied in the treatment of migraine [9], dizziness [10], stroke [11], epilepsy [12], and cognitive disorders [13,14]. Beyond traditional medical applications, G. elata is also consumed as a functional food ingredient, commonly added to porridge or used in herbal stews, reflecting its integration into both therapeutic and dietary practices [15].
Gastrodin, the principal bioactive compound isolated from the dried tubers of G. elata, is chemically defined as 4-(hydroxymethyl)phenyl-β-d-glucopyranoside [16]. Its structure comprises a glucose unit linked to p-hydroxybenzyl alcohol (pHBAL) through a glycosidic bond, with a molecular formula of C13H18O7 and a molecular weight of 286.27 g/mol. As the main pharmacologically active component of G. elata, gastrodin exhibits diverse biological activities and considerable potential for pharmaceutical and health-related applications. In clinical practice, gastrodin formulations such as injections [17], tablets, and capsules have been approved for treating neurasthenia, migraine, traumatic brain injury [18], and vertigo [2]. In addition, gastrodin shows pronounced neuroprotective activities [19], suggesting its potential for the management of neurodegenerative diseases, including Alzheimer’s and Parkinson’s disease [20]. In the nutraceutical and functional food industries, gastrodin has attracted increasing attention due to its natural antioxidant and neuroprotective properties [21]. These bioactivities also underpin its potential application in cosmetic formulations [18,22], where its antioxidative and anti-inflammatory characteristics contribute to skin protection and rejuvenation (Figure 1).
Despite its broad pharmacological value, the industrial development of gastrodin remains constrained by the inefficiency of traditional extraction methods. The cultivation of G. elata requires a long growth period of three to five years and highly specific ecological conditions [23,24]. The high cost of raw materials accounts for the majority of production expenses, while the extraction yield is typically below one percent [25]. These limitations restrict large-scale production and hinder the sustainable development of gastrodin-based products. The rapid advancement of metabolic engineering and synthetic biology offers promising alternatives for the sustainable production of structurally complex and high-value natural products. Successful examples include the heterologous production of artemisinin [26], paclitaxel [27], and ginsenosides [28]. Inspired by these achievements, recent studies have reconstructed the biosynthetic pathway of gastrodin in microbial hosts, establishing a foundation for scalable and cost-effective biotechnological production.
This review provides an overview of the pharmacological activities and natural biosynthetic pathways of gastrodin, summarizes current chemical and biotechnological production strategies, and discusses emerging metabolic engineering approaches for developing efficient microbial platforms for gastrodin biosynthesis.
2. Biosynthesis and Regulation of Gastrodin
Gastrodin biosynthesis is a complex metabolic process involving multiple enzyme-catalyzed reactions and interconnected pathways. The efficiency and selectivity of this process depend largely on the catalytic performance of several key enzymes. Current studies suggest that gastrodin in plants is synthesized mainly through two distinct yet convergent routes: the phenylalanine-derived phenylpropanoid pathway [29] and the toluene-derived pathway [30]. Both pathways ultimately lead to the formation of the central intermediate pHBAL, which undergoes glycosylation to yield gastrodin (Figure 2).
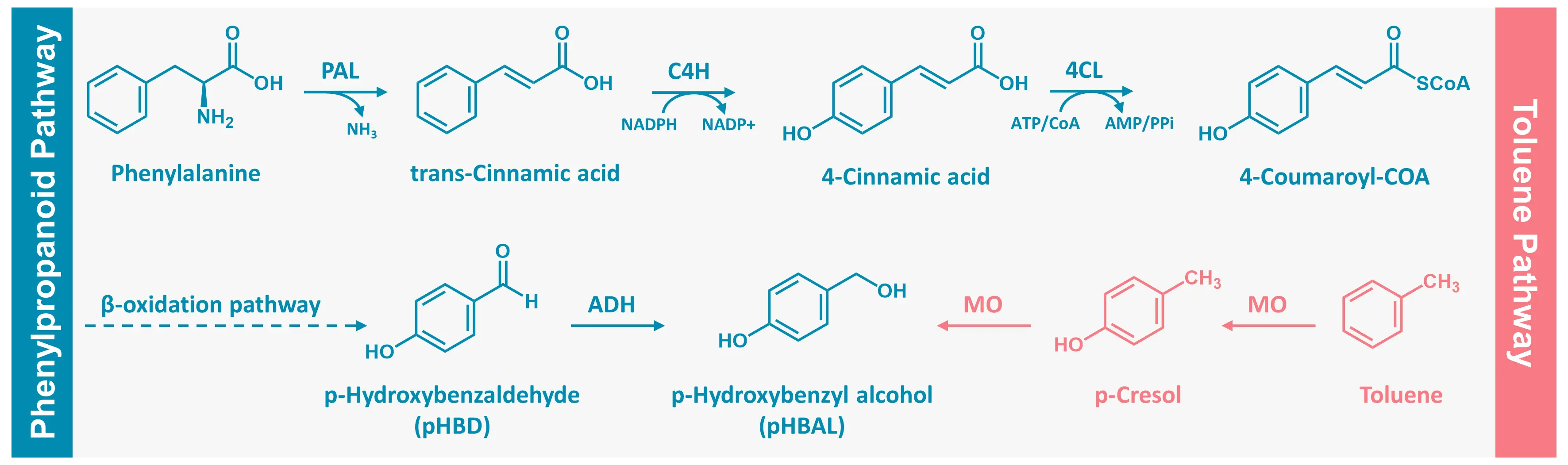
Figure 2. Two putative biosynthetic pathways of gastrodin in plants: the phenylpropanoid pathway and the toluene pathway. PAL, Phenylalanine ammonia-lyase; C4H, cinnamate 4-hydroxylase; 4CL, 4-coumarate-CoA ligase; ADH, alcohol dehydrogenase; MO, monooxygenase.
2.1. The Phenylpropanoid-Derived Pathway
This pathway originates from L-phenylalanine, which is biosynthetically derived from the shikimate pathway. In this route, phosphoenolpyruvate (PEP) and erythrose-4-phosphate (E4P) are condensed by 3-deoxy-d-arabino-heptulosonate-7-phosphate synthase (DAHP synthase) to yield 3-deoxy-d-arabino-heptulosonate-7-phosphate (DAHP). Through sequential enzymatic transformations, DAHP is converted into chorismate, the central intermediate of aromatic amino acid biosynthesis. Chorismate is subsequently rearranged by chorismate mutase to produce prephenate, which is dehydrated by prephenate dehydratase to form phenylpyruvate. The latter then undergoes transamination with glutamate as the amino donor, yielding L-phenylalanine as the final product of this upstream shikimate-derived branch. Phenylalanine ammonia-lyase (PAL) deaminates L-phenylalanine to produce trans-cinnamic acid, which is hydroxylated by cinnamate 4-hydroxylase (C4H) to form p-coumaric acid. This intermediate is subsequently activated by 4-coumarate-CoA ligase (4CL) to yield p-coumaroyl-CoA, providing the foundation for downstream reactions. Through a β-oxidation-like sequence, p-coumaroyl-CoA is converted to p-hydroxybenzaldehyde (pHBD), which is reduced by alcohol dehydrogenase (ADH) to pHBAL, the direct precursor of gastrodin [31]. The final glycosylation step, catalyzed by a UDP-dependent glycosyltransferase (UGT), transfers a glucose moiety from UDP-glucose (UDPG) to pHBAL, completing gastrodin biosynthesis.
2.2. The Toluene-Derived Pathway
An alternative biosynthetic route for gastrodin in G. elata may originate from toluene [30], an endogenous compound naturally found in many plants, which frequently serves as a general precursor for the biosynthesis of various secondary metabolites [32,33]. Transcriptomic comparisons between vegetative reproductive corms and young tubers by Tsai et al. identified two key enzyme candidates: a putative monooxygenase (TRINITY_DN54282_c0_g1) and a putative glycosyltransferase (TRINITY_DN50323_c0_g1) [30]. It was proposed that toluene is first hydroxylated by a toluene monooxygenase (TMO) to form p-cresol, which undergoes a second hydroxylation to produce pHBAL. This intermediate is then glycosylated by the identified UGT to generate gastrodin.
Although this pathway is supported by transcript-level data, subsequent genomic and transcriptomic analyses by Chen et al. revealed strong co-expression of genes involved in phenylalanine metabolism with gastrodin accumulation [31]. These findings indicate that pHBAL is more likely derived from the phenylpropanoid pathway rather than from toluene hydroxylation. Consequently, the prevailing consensus is that the phenylalanine-derived route represents the primary biosynthetic mechanism for gastrodin, while the toluene pathway may function as an auxiliary route under specific developmental or environmental conditions [34].
2.3. Key Enzymes Involved in Gastrodin Biosynthesis
Comprehensive elucidation of the gastrodin biosynthetic pathway in G. elata requires not only delineating the sequential reactions and intermediates but also identifying the rate-limiting enzymes that govern metabolic flux and product accumulation. The following sections summarize recent progress in identifying and characterizing enzymes that play essential roles in this pathway and discuss their potential biological and physiological significance.
2.3.1. Phenylalanine Ammonia-Lyase (PAL)
PAL serves as the entry enzyme of the phenylpropanoid pathway, catalyzing the deamination of L-phenylalanine to form trans-cinnamic acid. This step represents a crucial metabolic junction connecting primary metabolism to the synthesis of secondary metabolites. Transcriptomic analyses of G. elata revealed that PAL-encoding genes such as MSTRG.6339 and MSTRG.6340 are co-expressed with downstream enzymes including 4CL, both of which are implicated in gastrodin formation. These genes exhibit notably high expression levels in G. elata tubers, indicating a potential role in providing biosynthetic precursors for gastrodin [31]. In Pholidota chinensis, PAL transcript abundance shows a positive correlation with gastrodin accumulation, supporting its pivotal role in the initial stage of phenylpropanoid metabolism.
2.3.2. Cinnamate 4-Hydroxylase (C4H)
C4H, a member of the cytochrome P450 CYP73 family [35], catalyzes the para-hydroxylation of trans-cinnamic acid to produce p-coumaric acid, a rate-determining intermediate in gastrodin biosynthesis. In plant cells, C4H is localized to the endoplasmic reticulum membrane and often forms a functional complex with PAL, thereby facilitating substrate channeling and minimizing intermediate diffusion [36]. This spatial organization enhances the catalytic efficiency of the phenylpropanoid pathway. In G. elata, C4H exhibits a highly conserved sequence and function. Transcriptomic data show that C4H expression is markedly elevated in tubers relative to propagules and correlates positively with gastrodin content [30], implying that its upregulation contributes to gastrodin accumulation.
Functional studies further substantiate this role. Knosp and colleagues demonstrated that GePIF4 directly binds to the promoter region of GeC4H1 gene, activating its transcription as confirmed by dual-luciferase reporter and yeast one-hybrid assays [37]. Overexpression of GePIF4 was found to promote gastrodin accumulation, accompanied by upregulation of GeC4H1 gene expression, indicating that C4H (GeC4H1) is a key enzyme in gastrodin biosynthesis. Moreover, inhibition of C4H activity by piperonylic acid led to decreased levels of gastrodin precursors such as p-coumaric acid and pHBD, verifying that C4H functions as a rate-limiting enzyme in this biosynthetic route.
2.3.3. 4-Coumarate-CoA Ligase (4CL)
4CL ligase catalyzes the activation of p-coumaric acid with coenzyme A to form p-coumaroyl-CoA [38], a central branch-point intermediate in phenylpropanoid metabolism that supplies precursors for gastrodin synthesis. In G. elata, the 4CL gene MSTRG.8282 was identified by transcriptome analysis [31]. Its expression pattern is closely coordinated with that of PAL and C4H, showing pronounced upregulation in tubers with high gastrodin accumulation. In P. chinensis, overexpressing the 4CL gene promoted the conversion of p-coumaroyl-CoA to phenylalanine derivatives, thereby indirectly increasing gastrodin content [29].
2.3.4. Cinnamoyl-CoA Reductase (CCR) and Alcohol Dehydrogenase (ADH)
The subsequent steps involve the sequential reduction of p-coumaroyl-CoA to pHBAL [39], the immediate precursor of gastrodin. CCR catalyzes the conversion of p-coumaroyl-CoA to p-coumaraldehyde, which is subsequently reduced by ADH. In G. elata, co-expression network analysis identified CCR (MSTRG.5570) and ADH (MSTRG.8906) as strongly correlated with the accumulation of pHBAL [31]. Functional validation of ADH confirmed its catalytic activity by heterologous expression of G. elata ADH in Escherichia coli which efficiently converted p-coumaraldehyde to pHBAL, thereby providing the glycosylation substrate required for gastrodin formation [40].
2.3.5. Glucosyltransferases (GTs)
GTs catalyze the final glycosylation step of gastrodin biosynthesis by transferring a glucose moiety from UDPG to pHBAL. Transcriptomic screening identified 39 putative GT genes, which were classified into four groups [29]. Among them, transcript28360/f2p0/1592 displayed the highest expression in P. chinensis pseudobulbs and clustered phylogenetically with RsUGT from Rauvolfia serpentina, suggesting a similar catalytic function. Sequence alignment indicated strong conservation with predicted GT genes in G. elata, and co-expression analysis confirmed a positive correlation with gastrodin accumulation, establishing transcript28360/f2p0/1592 as a prime candidate for catalyzing the glycosylation step.
Further transcriptomic comparison between vegetative propagules and young tubers of G. elata revealed a glucosyltransferase gene (TRINITY_DN50323_c0_g1) that was expressed at approximately 3.2-fold higher levels in young tubers, consistent with increased gastrodin content [30]. Semi-quantitative and quantitative RT-PCR analyses supported its involvement in the terminal biosynthetic step. In another genome-wide study [31], genes annotated as hydroquinone glucosyltransferases (MSTRG.3101 and MSTRG.7014) were co-expressed with upstream phenylpropanoid genes such as PAL and 4CL, and were highly expressed in tubers. Collectively, these findings indicate that MSTRG.3101 and MSTRG.7014 are the principal candidates responsible for catalyzing the final glycosylation reaction in gastrodin biosynthesis.
2.4. Key External Factors in Gastrodin Biosynthesis
The biosynthesis of gastrodin is governed by both endogenous regulatory networks and environmental cues. Internal genetic programs determine the baseline expression of pathway enzymes, while external factors such as symbiotic interactions, nutrient availability, and abiotic stresses modulate their transcriptional and post-translational regulation. These combined influences orchestrate the dynamic control of metabolic flux through the phenylpropanoid and glycosylation pathways.
2.4.1. Symbiotic Fungal Associations
Symbiotic fungi play an indispensable role in the life cycle and metabolic activity of G. elata [23]. The species relies on a dual symbiotic relationship: Mycena species during seed germination and Armillaria mellea during vegetative growth. Mycena provides essential nutrients to the nutrient-deficient seeds, enabling the formation of protocorms, whereas A. mellea decomposes lignocellulosic material in decaying wood and transfers nutrients to the developing tubers, thereby supporting their growth and expansion.
Beyond nutritional support, fungal symbiosis exerts profound effects on secondary metabolism. Interaction with A. mellea decomposers has been shown to upregulate key enzymes in the phenylpropanoid pathway, enhancing the synthesis and glycosylation of gastrodin precursors [31]. This symbiosis also influences the metabolism of parishin (PAR), a storage form of gastrodin that is hydrolyzed into the active compound within tubers [41]. Hormonal modulation, particularly involving gibberellins and abscisic acid, is believed to mediate this process.
The functional importance of these interactions has been demonstrated through strain selection studies. En et al. developed a high-efficiency A. mellea strain (JFGL-06) that improves nutrient absorption and optimizes rhizosphere microbial composition, thereby enhancing both the yield and quality of G. elata Bl. f. glauca cultivated in the Changbai Mountain region [42]. These findings underscore that symbiotic fungi are not only critical for plant development and metabolic regulation but also serve as ecological mediators that sustain plant-microbe mutualism and ecosystem productivity.
2.4.2. Environmental Factors
The accumulation of gastrodin in G. elata is closely linked to its physiological state, which is shaped by environmental conditions. Modeling analysis using the MaxEnt framework revealed that annual precipitation, elevation, and mean temperature during the driest quarter collectively account for over 80% of the environmental influence on G. elata distribution [43]. These variables are considered key ecological determinants that indirectly regulate gastrodin biosynthesis by affecting growth and symbiotic efficiency. The optimal annual precipitation range for G. elata is estimated at 736–1256 mm. Adequate soil moisture promotes vigorous mycelial growth of A. mellea, facilitating extensive colonization of woody substrates and efficient nutrient transfer to G. elata tubers [23]. Both drought and waterlogging impede this symbiosis, thereby reducing metabolic activity. Elevation exerts an indirect yet significant influence on metabolic regulation by shaping local temperature, light intensity, and atmospheric pressure. G. elata thrives at altitudes between 767 and 2394 m, where the cool, humid climate and humus-rich soils create optimal conditions for fungal association and secondary metabolite production. Such environments favor the accumulation of phenylpropanoid-derived compounds, including gastrodin. Temperature is another critical determinant. The ideal growth temperature for G. elata is 15–25 °C [44], which coincides with maximal activity of gastrodin biosynthetic enzymes. Seasonal low-temperature dormancy is essential for tuber germination in the following year [43], and deviations from this thermal window disrupt metabolic balance. Prolonged warmth can prematurely terminate dormancy and impair gastrodin synthesis, whereas extreme cold may cause frost injury and cellular damage. Thus, appropriate temperature and moisture regimes are vital for maintaining both symbiotic functionality and secondary metabolite biosynthesis in G. elata.
3. Current Approaches for Gastrodin Production
The growing demand for natural medicines and health supplements has markedly increased the market requirement for gastrodin [45], necessitating more efficient, stable, and sustainable production strategies. Currently, gastrodin is produced mainly through four approaches: plant extraction, chemical synthesis, plant transformation, and microbial fermentation. Each method presents distinct advantages and limitations with respect to yield, scalability, environmental impact, and cost-effectiveness (Figure 3).
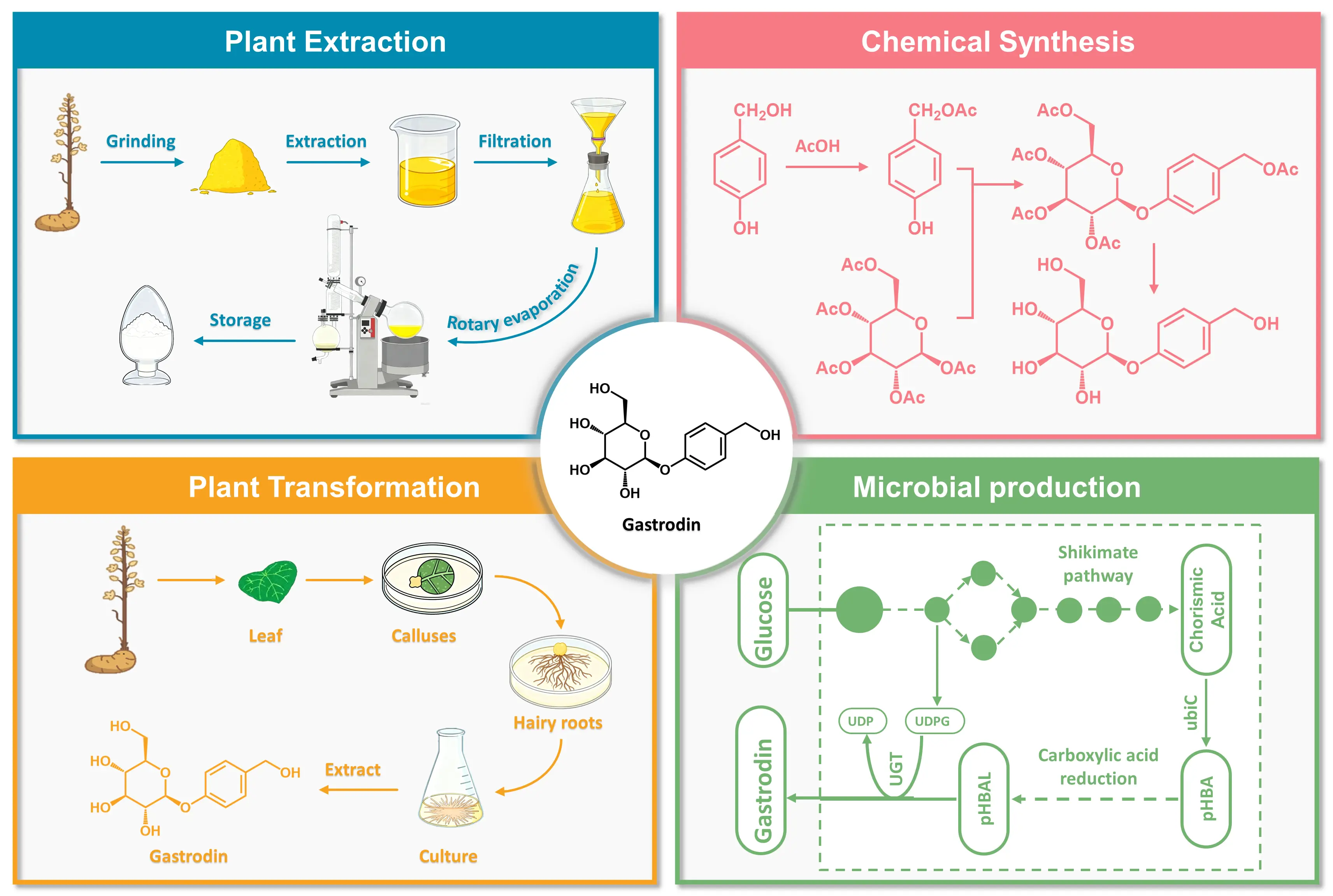
Figure 3. The production of gastrodin via plant extraction, chemical synthesis, plant transformation, and microbial fermentation.
3.1. Plant Extraction
Gastrodin is predominantly extracted from the dried tubers of G. elata [46]. Plant extraction focuses on the efficient and safe isolation of gastrodin from plant tissues and can be classified into traditional solvent-based techniques, modern physical-assisted methods, and integrated extraction-purification systems [47]. Conventional solvent reflux extraction remains the most widely used industrial method [48]. It relies on the solubility of gastrodin in solvents of varying polarity to achieve separation, typically using 60–80% ethanol or methanol as the solvent [49]. Gastrodin is released from powdered G. elata through repeated cycles of heating or maceration, yielding a crude extract. Ethanol is generally preferred due to its high extraction efficiency, low toxicity, and cost-effectiveness. For instance, Liao et al. used 70% ethanol to extract gastrodin from G. elata collected in Yunyang, achieving a yield of 0.568% as determined by HPLC analysis [50]. Modern extraction techniques such as microwave-assisted extraction (MAE) [51], ultrasound-assisted extraction (UAE) [51], and pressurized liquid extraction (PLE) [52] have improved efficiency and reduced solvent consumption. Zeng et al. optimized MAE conditions using extraction yield as the key parameter [53]. Under 55% ethanol, a liquid-to-solid ratio of 12 mL/g, microwave power of 0.4 kW, and three extraction cycles of 45 min each, the yield reached 0.632%, representing a substantial improvement over conventional reflux extraction. Similarly, the application of PLE at room temperature resulted in a gastrodin yield of 0.754%, representing a marked improvement over the 0.430% yield obtained via conventional methanol reflux [52].
Following extraction, the crude product contains impurities such as polysaccharides, amino acids, and other glycosides that necessitate purification. Macroporous resin adsorption is commonly used to remove impurities based on selective adsorption, followed by ethanol elution to obtain a purified fraction. Additional purification methods, including column chromatography and high-speed counter-current chromatography (HSCCC), are also utilized [54]. Tian et al. optimized an ultrasound-assisted enzymatic extraction process using α-amylase and established a two-phase aqueous extraction system [55]. After macroporous resin purification, the purity of gastrodin increased from 4.89% to 26.57%, representing a 5.4-fold improvement. Similarly, Li et al. used HSCCC with a biphasic solvent system of n-butanol, ethyl acetate, and water (2:3:5, v/v/v), isolating 48 mg of gastrodin with 96% purity from 500 mg of crude extract in a single run, with a recovery rate of approximately 90% [46].
3.2. Chemical Synthesis
Chemical synthesis of gastrodin has been pursued as an alternative to plant extraction due to limited natural abundance and resource constraints. Since its initial report in the 1980s, synthetic routes have evolved from the use of toxic reagents to greener and more industrially feasible processes [56]. The principal challenge remains the efficient formation of the β-glycosidic bond while balancing yield, selectivity, and environmental sustainability.
Li et al. developed a synthetic route using pentaacetyl-β-d-glucose and p-cresol as starting materials [57]. Glycosylation catalyzed by molecular sieves and Lewis acid produced 4-methylphenyl-2,3,4,6-tetra-O-acetyl-β-d-glucopyranoside. Subsequent free radical halogenation with N-halosuccinimide and reaction with acetic acid and tertiary amine yielded 4-(acetoxymethyl)phenyl-2,3,4,6-tetra-O-acetyl-β-d-glucopyranoside, which was then deacetylated to obtain gastrodin with an overall yield of 41.3% over four steps. The entire process avoided heavy metal catalysts and toxic reagents, thereby representing a greener alternative suitable for scale-up. In a simplified strategy, Wang et al. replaced p-cresol with pHBAL, thus eliminating the need for hydrogenation [58]. Esterification produced p-acetoxymethylphenol, which underwent glycosylation with pentaacetyl glucose under BF3·Et2O catalysis, followed by saponification to yield gastrodin, achieving an overall yield of 44.1% across three steps (Figure 4).
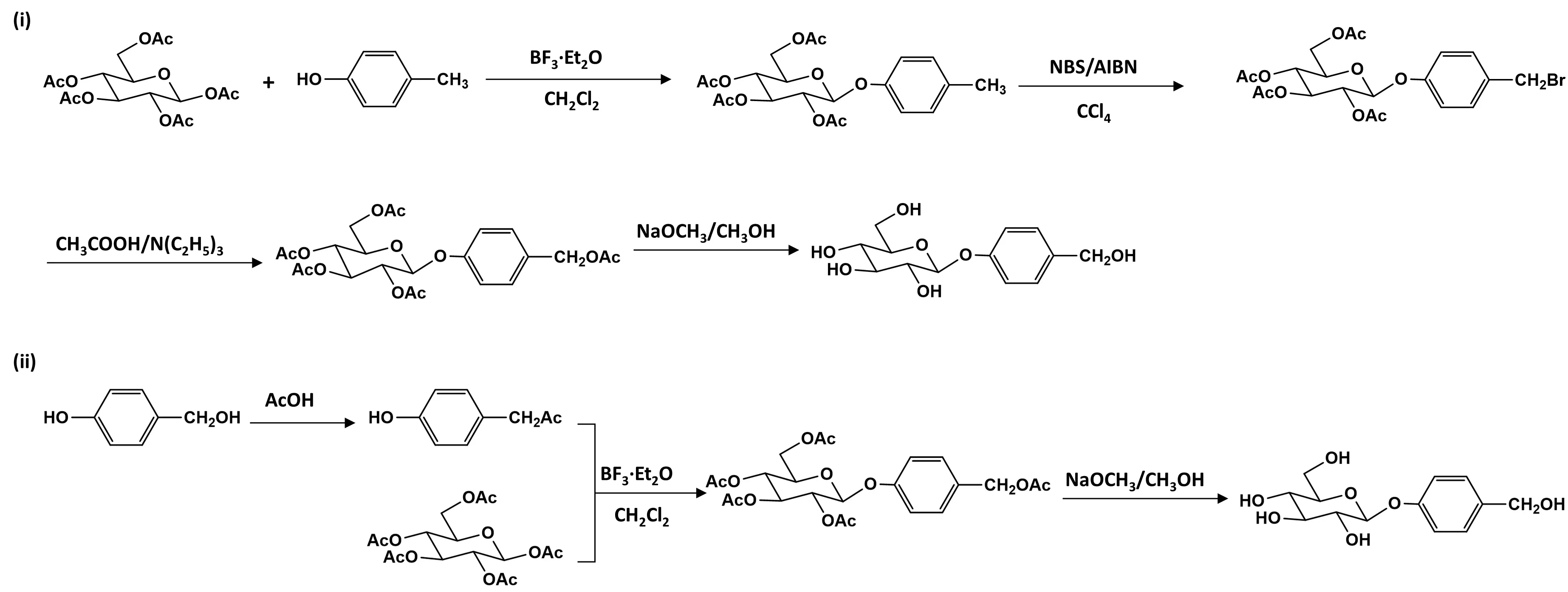
Figure 4. Proposed two chemical synthesis pathways for gastrodin. (i) Four-step synthesis of gastrodin from pentaacetyl-β-d-glucose and p-cresol; (ii) Three-step alternative synthesis of gastrodin using pHBAL as a starting material.
Despite these advances, chemical synthesis still faces limitations such as multistep reactions, moderate yields, isomer formation, and the generation of by-products that complicate purification and increase environmental burden. Although certain routes have been applied industrially, the development of more efficient, environmentally benign, and economically viable synthesis strategies remains a crucial objective for future research.
3.3. Plant-Based Biotransformation Systems
Plant transformation utilizes the intrinsic enzymatic systems of plant cells, tissues, or organs to convert exogenous precursors into gastrodin. Two main approaches have been developed: suspension-cultured cell transformation and Agrobacterium rhizogenes-mediated hairy root transformation.
Suspension-cultured cell systems, particularly those derived from the genus Datura, have been shown to bioconvert pHBD into gastrodin. In this process, reductase first reduces pHBD to pHBAL, which is subsequently glycosylated by glycosyltransferase. The addition of 0.1 mg/L salicylic acid or the maintenance of the bioreactor pressure below 0.001 MPa resulted in glycosylation rates exceeding 97% and 45%, respectively. Hairy root transformation has also demonstrated high conversion efficiency [59]. Using A. rhizogenes strain A4, Peng et al. induced hairy roots from Datura tatula leaf explants, achieving a 95% induction rate and confirming the capacity to convert pHBAL into gastrodin [60]. In a separate study, Cai et al. employed hairy roots of Panax ginseng cultured in B5 medium to transform pHBAL, yielding gastrodin that accounted for 6.65% of the dry weight within 24 h, corresponding to an 84.8% conversion rate [61].
4. Microbial Production Platforms
Unlike plant-based transformation systems that depend on the intrinsic metabolic network of G. elata, microbial biosynthesis overcomes limitations imposed by plant growth cycles and environmental constraints. By screening naturally occurring microbial strains or constructing engineered microorganisms, the catalytic potential of microbes can be harnessed to produce gastrodin efficiently. This strategy provides a controllable, sustainable, and industrially scalable alternative for gastrodin manufacture.
4.1. Whole-Cell Biotransformation of pHBAL to Gastrodin
Cui et al. co-expressed itUGT2 from Indigofera tinctoria and sucrose synthase GmSuSy from Glycine max in E. coli [62]. The itUGT2 catalyzed the conversion of pHBAL to gastrodin, while GmSuSy regenerated UDPG from sucrose, eliminating the need for external sugar donors. This whole-cell catalytic system achieved a 95% conversion rate and a yield of 220 mg/L. Zhao et al. developed an engineered E. coli strain harboring a UDPG synthesis module and a mutant itUGT2A347V [63]. By supplying pHBAL and sucrose as external substrates and enabling the intracellular self-supply of cofactor UDPG, the recombinant strain produced 4.56 g/L gastrodin in shake-flask fermentation. Xia et al. identified a AtUGT from Arabidopsis thaliana through homology analysis [64]. The enzyme showed optimal activity at pH 9.0 and 45 °C, with 2.5 mM Mg2+ enhancing its catalytic performance. Under optimized reaction conditions, pHBAL was completely converted to 2.67 g/L gastrodin within 12 h, corresponding to a 94.3% conversion rate. Utilizing a triple mutant of this AtUGTN94L/N221G/I343V with 50.1% higher activity, Wang et al. engineered an E. coli strain by deploying a push-pull metabolic strategy that combined a UDPG regeneration module with pgm deletion, ultimately achieving a gastrodin titer of 22.9 g/L and a space-time productivity of 1.9 g/L/h within 12 h [65]. Lin et al. expressed RsUGT in E. coli BL21(DE3) for the synthesis of gastrodin from pHBAL [25]. The purified enzyme (~77.7 kDa) exhibited maximal activity at 40 °C and pH 10.0, and the addition of 2% DMSO enhanced its activity by 12.1%. Under optimized enzymatic conditions, the system produced 285.3 mg/L gastrodin within 6 h, with a 99.7% molar conversion rate.
Zhu et al. screened ten fungal strains using resting-cell and batch-culture methods combined with thin-layer chromatography (TLC) and reversed-phase high-performance liquid chromatography (RP-HPLC) [66]. One of these strains, Rhizopus LN-1 exhibited the strongest catalytic ability, converting pHBD to gastrodin with a yield of 16% under optimized conditions (2.3 mg/mL substrate, 12 g cells per 120 mL medium, pH 5.5, 28 °C, 24 h). Fan et al. further identified Aspergillus foetidus ZU-G1 and Penicillium cyclopium AS 3.4513 as effective biocatalysts that converted pHBAL into 36 mg/L and 65 mg/L gastrodin, respectively [67]. Zhang et al. screened several natural filamentous fungal strains, including Armillaria luteo-virens and its mutants, and identified the variant A. luteo-virens Sacc U with superior transformation capability [68]. This strain efficiently converted pHBAL to gastrodin in a single step, producing 7.5 mg/L under shake-flask conditions. These results highlight the potential of filamentous fungi as biocatalysts for scalable gastrodin production.
These studies demonstrate that, beyond plants, certain microorganisms possess intrinsic gastrodin-synthesizing capability. Compared with plant-based systems, microbial fermentation offers shorter production cycles, simplified nutrient requirements, and more consistent process control, rendering it a more practical option for large-scale gastrodin manufacturing.
4.2. De Novo Biosynthesis of Gastrodin in Microbial Hosts
In de novo microbial biosynthesis, glucose serves as the primary carbon source for gastrodin. It is metabolized through glycolysis to PEP and through the pentose phosphate pathway to E4P. These intermediates are condensed by DAHP synthase to form DAHP, which enters the shikimate pathway and yields chorismate. Chorismate is then converted to pHBA by chorismate lyase (ubiC), marking a key branching point in precursor synthesis. Subsequently, carboxylic acid reductase (CAR) reduces pHBA to pHBD, with activation by a phosphopantetheinyl transferase such as Sfp from Bacillus subtilis or PPTcg-1 from Corynebacterium glutamicum. This adenosine triphosphate (ATP)- and nicotinamide adenine dinucleotide phosphate (NADPH)-dependent step is essential for efficient aldehyde formation. The intermediate pHBD is then reduced to pHBAL by endogenous alcohol dehydrogenases (ADHs). Finally, GTs catalyze the conjugation of pHBAL with UDP-glucose to form gastrodin.
4.2.1. Yeast Systems
Yin et al. used Saccharomyces cerevisiae BY4742 as a chassis strain to construct a de novo gastrodin biosynthetic pathway [69]. They identified a plant glucosyltransferase RsUGT with high catalytic activity toward pHBAL and introduced codon-optimized genes, including RsUGT [70], CAR from Nocardia genus [71], PPTcg-1 from C. glutamicum [71], and UbiC from E. coli [72], together with the feedback-resistant mutant ARO4K229L [73]. Multi-copy chromosomal integration and overexpression of genes (ppsA, tktA, ARO1, ARO2) enhanced precursor flux, while deletion of ARO7 activated the shikimate pathway. The resulting strain produced 2.1 g/L gastrodin in flask fermentation, a 175-fold increase relative to the parent strain. Wan et al. constructed a highly efficient Saccharomyces cerevisiae CEN.PK2-1C cell factory for de novo gastrodin biosynthesis by deploying a feedback-insensitive Aro3D154C variant and a structure-guided Aro7E198Q mutant, which redirected carbon flux toward the shikimate pathway without inducing aromatic amino acid auxotrophy. By systematically reprogramming central metabolism to reinforce E4P, PEP, NADPH, and UDPG availability, alongside introducing a semi-rationally evolved AtUGT72B1 quadruple mutant (E83T/R81E/V144M/I310L) with a 26.6-fold increase in catalytic efficiency, the integrated strain achieved a gastrodin titer of 5.44 g/L in shake flasks and a record-breaking 90.34 g/L in fed-batch fermentation [74]. Wu et al. engineered the oleaginous yeast Yarrowia lipolytica Po1fΔKu70 for high-efficiency gastrodin synthesis [75]. The introduction of UbiC, CAR, Sfp, and AS genes produced strain Gd01, yielding 1.05 g/L gastrodin. Subsequent pathway enhancement and gene knockouts (ylTRP1, ylARO10, ylPAR4, ylPHA2) increased the titer to 8.10 g/L in shake flasks and 13.22 g/L in a 5-L bioreactor. Gu et al. further optimized Y. lipolytica through systematic genome refactoring [76]. Screening and engineering of key enzymes (e.g., SylUGTS349E, ylPAR2, NcCAR, EcUbiCL220M) and multi-copy integration of biosynthetic modules improved metabolic flux. The supply of UDPG was enhanced, while competing pathways and gastrodin-degrading glucosidases were removed. In addition, a copper-inducible antisense regulation system (CATR) was implemented for dynamic control. The final strain YliGT8 achieved 9.48 g/L gastrodin in flasks and 13.4 g/L in a 5-L bioreactor, corresponding to a yield of 0.149 g/g glucose, marking the highest titer reported to date.
4.2.2. Engineered E. coli
Bai et al. successfully realized the de novo biosynthesis of gastrodin in E. coli BL21(DE3) through the rational reconstruction of a complete heterologous pathway [40]. This artificial route incorporated CAR from Nocardia, endogenous ADHs, and UGT73B6 from Rhodiola. To enhance the precursor supply, metabolic flux was redirected from central carbon metabolism toward pHBAL by overexpressing ubiC, aroGfbr, and ppsA, resulting in approximately a tenfold increase in gastrodin production compared with the initial strain. Furthermore, directed evolution of UGT73B6 generated a variant UGT73B6F389S exhibiting markedly improved catalytic efficiency toward pHBAL. Collectively, these optimizations enabled the engineered E. coli to produce 545 mg/L of gastrodin within 48 h of cultivation, establishing a foundational framework for microbial production of this high-value compound. Building upon this metabolic framework, Wang et al. further pushed the limits of de novo biosynthesis in E. coli by simultaneously addressing aromatic precursor availability and sugar donor limitations [77]. Instead of solely relying on endogenous cofactor pools, they reconstructed an independent UDPG regeneration pathway to sustainably support a highly active AtUGT triple mutant (T282S/A294E/V463L). This coordinated supply of both aglycone precursors and UDPG increased the titer to 2.18 g/L without compromising cell viability. Upon scaling up to a 1.1-L bioreactor with oxygen-controlled fed-batch fermentation, the engineered strain achieved 13.73 g/L gastrodin, representing the highest titer reported for E. coli systems to date.
5. Metabolic Engineering Strategies for Enhanced De Novo Gastrodin Biosynthesis
The microbial de novo biosynthesis of gastrodin fundamentally depends on the reconstruction of intracellular metabolic networks to convert simple carbon sources, such as glucose, into the target compound via a cascade of enzymatic reactions. The development of an efficient microbial cell factory requires carefully designed metabolic engineering strategies that direct cellular carbon flux toward the desired product while minimizing competing pathways. This section provides an overview of the major genetic and metabolic engineering approaches that have been employed to enhance gastrodin biosynthesis (Figure 5).
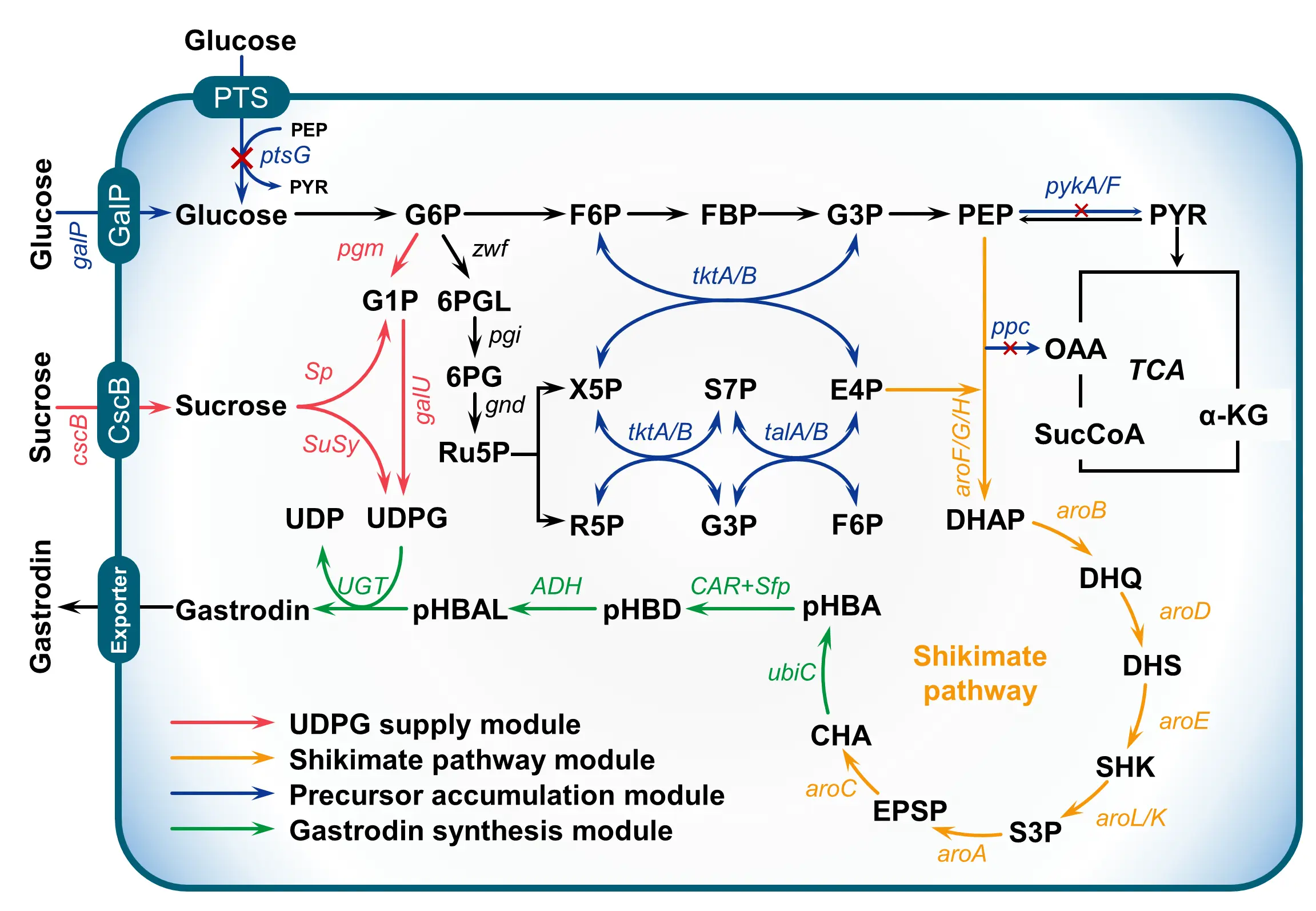
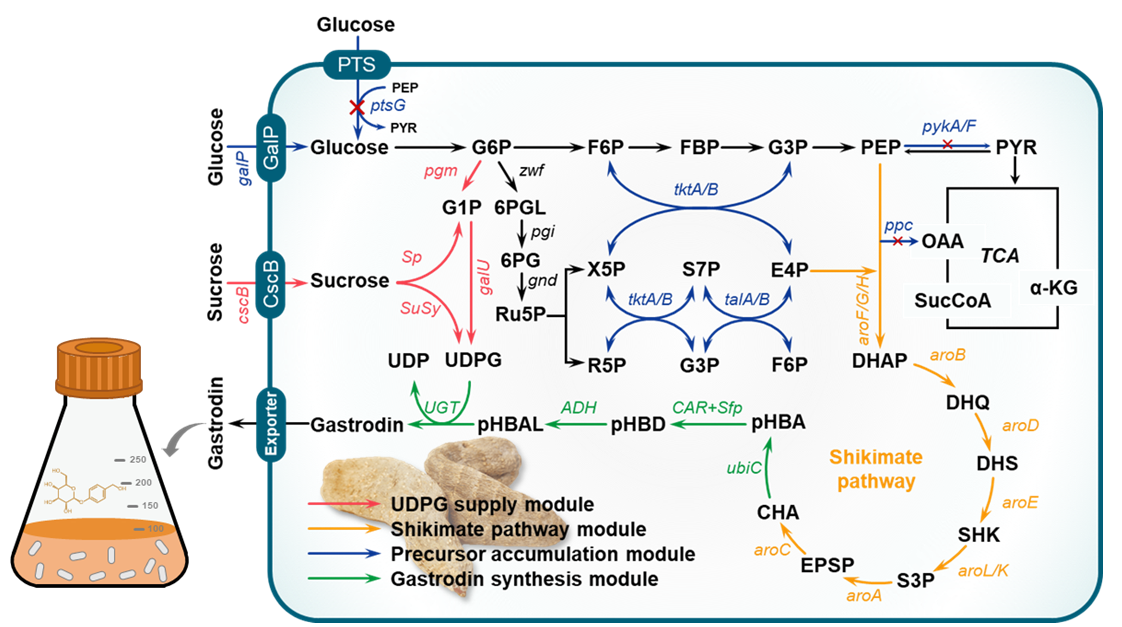
Figure 5. Common metabolic engineering strategies for microbial production of gastrodin. Metabolite abbreviations: G6P, glucose-6-phosphate; F6P, fructose-6-phosphate; FBP, fructose-1,6-biphosphate; G3P, glyceraldehyde-3-phosphate; PEP, phosphoenolpyruvate; PYR: pyruvate; G1P, glucose-1-phosphate; 6PGL, 6-phosphoglucolactone; 6PG, 6-phosphogluconate; Ru5P, ribulose-5-phosphate; X5P, xylulose-5-phosphate; R5P, ribose-5-phosphate; S7P, sedoheptulose-7-phosphate; E4P, erythrose-4-phosphate; OAA, Oxaloacetate; α-KG, α-ketoglutarate; SucCoA, succinyl-coenzyme A; DAHP, 3-deoxy-D-arabinoheptulosonate 7-phosphate; DHQ, 3-dehydroquinate; DHS, 3-dehydroshikimate; SHK, shikimate; S3P, shikimate-3-phosphate; EPSP, 5-enolpyruvyl-shikimate 3-phosphate; CHA, chorismate; pHBA, p-hydroxybenzoic acid; pHBD, p-hydroxybenzaldehyde; pHBAL, p-hydroxybenzyl alcohol; UDP, uridine diphosphate; and UDPG, uridine diphosphate glucose. The four strategies include UDPG supply module, shikimate pathway module, precursor supply module, and gastrodin synthesis module, which are shown in red, yellow, blue, and green.
5.1. Optimizing the Glycosyl Acceptor: 4-Hydroxybenzyl Alcohol (pHBAL)
Gastrodin biosynthesis critically depends on the efficient supply of its glycosyl acceptor, pHBAL. Enhancing pHBAL accumulation requires coordinated optimization of three upstream modules: precursor (PEP, E4P, and chorismate) supply, intermediate conversion efficiency, and regulatory balance within central carbon metabolism.
5.1.1. Enhancing Phosphoenolpyruvate (PEP) Supply
Precursor availability is a critical determinant of gastrodin production efficiency. Enhancing the intracellular levels of the key intermediate PEP relies on coordinated regulation of glycolysis and the pentose phosphate pathway (PPP). Two main engineering strategies are generally adopted: (i) reducing PEP consumption, and (ii) preventing carbon leakage into the tricarboxylic acid (TCA) cycle.
In E. coli, the phosphoenolpyruvate-sugar phosphotransferase system (PTS) represents the primary glucose uptake mechanism [78,79]. It couples glucose transport with phosphorylation, using PEP as a phosphate donor. This process consumes a substantial portion of intracellular PEP, thereby limiting its availability for the shikimate pathway and constraining the synthesis of downstream aromatic metabolites. To address this, researchers have disrupted the native PTS and introduced non-PTS systems, such as those involving galactose permease (encoded by galP) and glucokinase (glk), which utilize ATP rather than PEP for glucose phosphorylation. These modifications markedly reduce PEP consumption and redirect carbon flux from glycolysis to shikimate-derived pathways. Another key approach involves modulating pyruvate kinase activity. Pyruvate kinase, encoded by pykA and pykF, catalyzes the conversion of PEP to pyruvate with concomitant ATP generation. By deletion of pykA and pykF, this conversion is attenuated, thereby increasing the intracellular PEP pool available for the shikimate pathway [80,81]. Strains with both PTS disruption and pyk deletions have shown improved PEP availability, increased aromatic compound synthesis, and higher biomass yields.
Carbon leakage from glycolytic intermediates into the TCA cycle can further deplete precursors required for biosynthesis. To minimize this diversion, metabolic flux can be decoupled from the TCA cycle through targeted gene knockouts. Fujiwara et al. eliminated key enzymes such as 2-keto-3-deoxy-6-phosphogluconate aldolase (eda), phosphoenolpyruvate carboxylase (ppc), phosphoenolpyruvate carboxykinase (pck), and phosphoenolpyruvate synthase (ppsA), thereby effectively blocking carbon flux from PEP and pyruvate into the TCA cycle [82]. These modifications prevented carbon loss and increased the concentration of glycolytic intermediates, thereby strengthening precursor supply for aromatic compound biosynthesis.
5.1.2. Boosting Erythrose-4-Phosphate (E4P) Availability
E4P, an essential precursor in the shikimate pathway, is generated primarily through the PPP [83,84]. This pathway also serves as a major source of NADPH, which is required for numerous biosynthetic reactions. Enhancing E4P availability thus not only supports chorismate formation but also provides reducing power for downstream enzymatic conversions. The first rate-limiting step in the PPP is catalyzed by glucose-6-phosphate dehydrogenase (zwf), which is subject to feedback inhibition by NADPH [85]. To alleviate this regulatory constraint, overexpression of transketolase (tktA) and transaldolase (talB) has been shown to effectively increase carbon flux through the PPP [86,87]. These enzymes catalyze reversible carbon rearrangements that connect the glycolytic pathway to the pentose phosphate cycle. Their overexpression markedly elevates the intracellular E4P pool and, consequently, enhances the overall production capacity of shikimate-derived compounds.
5.1.3. Engineering the Shikimate Pathway for Chorismate Formation
Chorismate serves as a pivotal metabolic intermediate that connects central carbon metabolism to the biosynthesis of aromatic amino acids and related secondary metabolites, including gastrodin. The efficiency of chorismate accumulation largely determines the cellular capacity for pHBAL formation, the immediate precursor of gastrodin. Consequently, optimizing the shikimate pathway has become a central focus in metabolic engineering efforts aimed at boosting gastrodin biosynthesis. Current strategies to enhance chorismate production can be broadly categorized into two approaches: (i) relieving allosteric feedback inhibition to increase precursor flux through the pathway, and (ii) reinforcing the expression or activity of rate-limiting enzymes to further elevate shikimate turnover and chorismate yield. These engineering principles have been successfully applied in both prokaryotic and eukaryotic microbial hosts, providing a versatile foundation for constructing efficient gastrodin-producing cell factories.
Alleviating Feedback Inhibition
The biosynthesis of chorismate begins with the condensation of PEP and E4P to form DAHP. In E. coli, this step is catalyzed by DAHP synthases encoded by aroG, aroF, and aroH, with aroG contributing approximately 79% of total activity. The flux through this pathway is tightly controlled by feedback inhibition from aromatic amino acids, particularly tyrosine. To overcome this regulation, Li et al. engineered strain DA-16 by deleting tyrR, a global repressor of the aromatic amino acid biosynthetic operon, and by introducing feedback-resistant variants aroGD146N (designated aroGfbr) and tyrAM53I/A354V (tyrAfbr) [88]. These mutations abolished the feedback inhibition of DAHP synthase and prephenate dehydrogenase by tyrosine, thereby enabling efficient endogenous tyrosine production from glucose. Consequently, the strain no longer required exogenous tyrosine supplementation and exhibited enhanced flux toward chorismate-derived dopamine biosynthesis. A similar regulatory architecture also exists in eukaryotic systems such as S. cerevisiae. Two DAHP synthases, ARO3 and ARO4, are differentially inhibited by phenylalanine and tyrosine, respectively [89]. Site-directed mutagenesis, such as the ARO4S195F substitution, has been shown to abolish tyrosine-mediated feedback inhibition, thereby improving precursor flow toward chorismate [90]. These findings underscore that relieving feedback inhibition at the entry point of the shikimate pathway is a universal and effective strategy for enhancing aromatic precursor availability in both bacterial and yeast systems.
Reinforcing Shikimate Flux via Key Enzyme Overexpression
Beyond feedback deregulation, amplifying the expression of rate-limiting enzymes can substantially strengthen metabolic flux toward chorismate. In E. coli, overexpression of aroE (encoding shikimate dehydrogenase) and tyrB (encoding tyrosine aminotransferase) under the strong trc promoter significantly increased dopamine titers from 0.57 g/L to 0.89 g/L [88], reflecting improved conversion efficiency from chorismate to tyrosine and an overall enhancement in precursor flux. In eukaryotic hosts S. cerevisiae, pathway efficiency is limited by the low expression of the multifunctional AROM complex encoded by ARO1, which catalyzes five consecutive reactions from 3-dehydroquinate to 5-enolpyruvylshikimate-3-phosphate (EPSP) [91]. Overexpression of ARO1, or the terminal enzyme chorismate synthase (ARO2), has been demonstrated to substantially elevate chorismate accumulation [92]. Collectively, these findings demonstrate that reinforcing the activity of critical enzymes across microbial systems effectively boosts the shikimate pathway flux, leading to higher chorismate accumulation.
5.1.4. Enhancing 4-Hydroxybenzyl Alcohol (pHBAL) Production
Efficient supply of pHBAL is a prerequisite for microbial gastrodin biosynthesis. In this context, UbiC acts as a metabolic gateway, catalysing the conversion of chorismate to pHBA and thereby linking the shikimate pathway to phenolic glycoside synthesis [93] Because S. cerevisiae lacks a native ubiC homologue, functional expression of bacterial UbiC is required to establish this route [94]. Early efforts focused on overexpressing wild-type E. coli UbiC, but its limited catalytic efficiency restricted precursor accumulation. Directed evolution of UbiC has since yielded improved variants, such as EcUbiCE31Q/M34V identified by Gu et al. [76], which exhibited enhanced chorismate pyruvate-lyase activity. Heterologous expression of this mutant in strain YliUbic22 elevated pHBA titers by 1.9-fold relative to the wild-type enzyme, underscoring the importance of enzyme engineering in reinforcing the metabolic entry point for gastrodin biosynthesis.
Once pHBA is generated, its transformation into pHBAD marks the initiation of the reduction module mediated by CAR. This enzyme bridges aromatic acid metabolism with downstream alcohol formation and thus constitutes a central control node in the biosynthetic network. Functional expression of CARs from Nocardia spp. and other actinomycetes in E. coli has enabled efficient bioreduction of pHBA [40]. However, CARs are post-translationally inactive unless activated by PPTases, which convert the apoenzyme into its catalytically competent holo form through 4′-phosphopantetheine transfer. This activation, coupled with ATP- and NADPH-dependent reduction, drives the conversion of pHBA to pHBAD. A notable example is provided by Wu et al., who expressed the CAR homologue MAB4714 from Mycobacterium abscessus ATCC 19977 alongside Sfp in Y. lipolytica Po1fΔKu70 [75]. The engineered strain SC01 achieved >97% conversion of pHBA within 72 h, yielding 1.10 g/L of pHBAL. The concentration of pHBAD increased transiently to 0.21 g/L at 48 h and subsequently declined to 0.01 g/L, reflecting the efficient coupling between the CAR and downstream alcohol dehydrogenase modules.
The final reductive step toward pHBAL is catalysed by ADHs, which utilise NAD(P)H as cofactor to reduce pHBAD. Although endogenous ADHs from E. coli, Y. lipolytica, and S. cerevisiae [40,69,75] can catalyse this reaction, their native activity often limits flux through the pathway. Overexpression of efficient ADHs can both enhance reaction kinetics and mitigate aldehyde toxicity, a known bottleneck in aromatic alcohol synthesis. Wu et al. demonstrated that overexpression of ylPAR2 markedly accelerated the bioconversion of pHBAD, converting 1 g/L of substrate to 610.3 mg/L of pHBAL, 2.35 times higher than in the control strain [75]. The strong NAD(P)H affinity and synergistic interaction between ylPAR2 and CAR enabled rapid clearance of the reactive aldehyde intermediate, ultimately increasing pHBAL yield to 1.10 g/L. This tight enzymatic coupling forms the biochemical foundation for downstream glycosylation toward gastrodin production.
5.2. Optimizing UDP-Glucose (UDPG) Supply
UDPG is the essential glycosyl donor for gastrodin biosynthesis, and its intracellular availability directly determines glycosylation efficiency. Insufficient UDPG supply often constitutes a rate-limiting step in microbial systems. To address this bottleneck, recent studies have focused on reinforcing endogenous UDPG biosynthetic pathways, introducing efficient regeneration modules, and balancing precursor allocation.
5.2.1. Strengthening Endogenous UDPG Biosynthesis
In microbial hosts, the endogenous biosynthesis of UDPG proceeds via sequential reactions catalysed by phosphoglucomutase (PGM) and UTP-glucose-1-phosphate uridylyltransferase (GalU), which convert glucose-6-phosphate (G6P) into glucose-1-phosphate (G1P) and subsequently into UDPG [95]. Because UDPG serves as the universal glycosyl donor for UGT-catalysed reactions, enhancing its intracellular availability is critical for efficient gastrodin formation. Overexpression of pgm and galU markedly elevated UDPG levels, resulting in a 20-fold improvement in gastrodin production in E. coli strain G15 relative to the parental strain G1 [63]. However, G6P also represents a metabolic crossroad between biosynthetic and glycolytic fluxes. The enzyme phosphoglucose isomerase (PGI) converts G6P into fructose-6-phosphate (F6P), channelling carbon toward glycolysis and cellular energy metabolism rather than UDPG synthesis. Diverting G6P away from this competing route can therefore reinforce glycosyl donor formation. Targeted disruption of pgi effectively redirects carbon flux toward G1P accumulation. Pandey et al. demonstrated that E. coli Δpgi mutants exhibited increased G1P levels and achieved a 37.6% improvement in quercetin glycoside yield, underscoring the potential of flux reallocation strategies to boost UDPG-dependent product formation [96].
5.2.2. Establishing Heterologous UDPG Regeneration Systems
Beyond enhancing endogenous supply, the establishment of heterologous UDPG regeneration systems offers an efficient strategy for sustaining glycosylation reactions under high metabolic demand. One representative approach employs sucrose synthase (SuSy), which catalyses the reversible reaction between sucrose and UDP to generate UDPG and fructose. In a coupled glycosylation-regeneration system, Cui et al. coexpressed I. tinctoria ItUGT2 with GmSuSy, enabling in situ recycling of UDPG [62]. In this configuration, ItUGT2 mediates the transfer of glucose from UDPG to pHBAL to form gastrodin, releasing UDP, which is then reconverted to UDPG by GmSuSy. This cyclic system supported up to 37 UDPG turnovers, achieved 93% substrate conversion, and produced 535 mg/L gastrodin. Remarkably, even in the absence of exogenous UDPG, gastrodin yield increased 2.6-fold compared with the single-enzyme system, demonstrating the efficiency of SuSy-driven donor regeneration.
In addition to SuSy-mediated recycling, sucrose can also serve as a dual carbon and glycosyl donor source through phosphorylase-catalyzed cleavage. Bifidobacterium adolescentis sucrose phosphorylase (BaSP) hydrolyzes sucrose into fructose and G1P, which are subsequently utilized for cell growth and UDPG biosynthesis, respectively [97,98]. Coexpression of BaSP with uridylyltransferase (ugpA) from Bifidobacterium bifidum, together with targeted deletions to channel G1P toward glycosylation, enabled efficient coupling between primary metabolism and UDPG regeneration, thereby increasing astragalin production threefold from 570 to 1708 mg/L.
An alternative route exploits polyphosphate kinase (PPK) to catalyse the phosphorylation of UDP into UTP using polyphosphate as the phosphate donor [99]. UTP serves as a direct substrate for UDPG synthesis, thereby providing an indirect but highly effective means to sustain UDPG pools. Expression of the high-activity Pseudomonas aeruginosa PPK (PA3455) in gastrodin-producing E. coli strains increased intracellular UTP levels by 1.5-fold and enhanced gastrodin titers by 2.25-fold [63]. Together, these regeneration systems ensure the continuous provision of glycosyl donors, supporting prolonged enzymatic activity during fermentation.
5.2.3. Integrating Multi-Module Synergistic Pathways
To further optimize donor supply, Zhang et al. constructed a multi-module synergistic platform in E. coli, integrating SuSy-mediated UDPG regeneration, PPK-mediated UTP supply, and PGM/GalU-mediated endogenous UDPG synthesis [63]. This design established a coordinated network linking carbon flux redirection, precursor regeneration, and donor recycling. The resulting strain achieved balanced energy distribution, reduced accumulation of metabolic intermediates, and maximized the overall glycosylation efficiency. Implementation of this triple-module system led to a 51-fold increase in gastrodin production in strain G19 compared with the parent line, highlighting the power of modular co-optimization in UDPG metabolism engineering.
5.3. Engineering and Optimization of Glycosyltransferases (UGTs)
The UGT constitutes the rate-determining enzyme in gastrodin biosynthesis, catalyzing the transfer of a glucose moiety from UDPG to pHBAL to form the β-d-glucoside linkage characteristic of gastrodin. Rational engineering and directed evolution of UGTs have thus become central strategies to enhance catalytic turnover, substrate specificity, and cofactor utilization.
A broad range of UGTs from diverse sources has been functionally characterised for gastrodin synthesis. Gu et al. identified Solanum lycopersicum UGT (SlyUGT, NP_001307116.1) as an efficient biocatalyst and employed AlphaFold2-guided structural prediction and molecular docking to pinpoint key residues for mutagenesis [76]. Site-saturation of 16 amino acids, including Ser349 and Asn340, yielded the improved variants SlyUGTS349E and SlyUGTN340M, which exhibited enhanced activity toward pHBAL. Under shake-flask conditions, multi-copy integration and flux optimisation raised gastrodin titers to 8.3 g/L, while 5-L fed-batch fermentation achieved 13.4 g/L with a productivity of 0.149 g/g glucose, one of the highest yields reported in E. coli. Similarly, Bai et al. engineered Rhodiola UGT73B6, a natural catalyst of aromatic alcohol glycosylation [40]. Directed evolution produced the F389S variant, which displayed a twofold increase in catalytic efficiency (kcat/Km) and improved affinity for UDPG, enabling a gastrodin titer of 545 mg/L within 48 h. Yin et al. further advanced this strategy by codon-optimising R. serpentina RsUGT for S. cerevisiae expression [69]. The synthetic enzyme RsUGT exhibited 33-fold higher activity than the previously used UGT73B6F389S. After being combined with chromosomal multi-copy integration, a 2.1 g/L gastrodin titer was achieved, representing a 175-fold improvement over the parental strain. Wu et al. pursued a parallel route in Y. lipolytica, employing codon-optimized arbutin synthase (AS) from R. serpentina and integrating it with ubiC, CAR, and sfp modules to generate strain Gd07 [75]. The strain achieved 6.68 g/L gastrodin after 72 h in shake-flask fermentation, and 13.22 g/L in 5-L bioreactor culture. These findings collectively illustrate how the combination of UGT evolution, cofactor regeneration, and modular flux design has transformed gastrodin biosynthesis from a conceptual pathway into an industrially viable process.
6. Conclusions and Future Perspectives
Gastrodin is a naturally bioactive phenolic glycoside with notable neuroprotective and therapeutic potential, has drawn increasing industrial attention. Traditional extraction from G. elata is constrained by low yield and high production cost, prompting the shift toward microbial biosynthesis as a sustainable alternative. Over the past decade, significant progress has been made in elucidating the biosynthetic pathways and identifying enzymes responsible for gastrodin, laying the foundation for efficient de novo production in hosts such as E. coli, S. cerevisiae, and Y. lipolytica. Building on this framework, researchers are focusing on metabolic engineering strategies including reinforcing precursor flux, optimizing UDPG synthesis and regeneration, and improving enzymatic efficiency. These integrated strategies have markedly enhanced biosynthetic efficiency, enabling engineered strains to achieve gram-level gastrodin production and demonstrating the practical feasibility of microbial synthesis at scale (Table 1). While Y. lipolytica has demonstrated considerable potential as a microbial chassis for gastrodin biosynthesis due to its efficient glycosylation capacity and robust fermentation performance, E. coli systems still exhibit relatively low titers, mainly due to incomplete pathway integration, limited enzyme compatibility, and imbalanced cofactor utilization. Overcoming these challenges requires refined metabolic regulation, dynamic flux control, and improved transporter efficiency to enhance precursor conversion and product secretion. Moreover, optimization of gene expression parameters such as promoter strength, plasmid copy number, translation efficiency, and protein solubility can help achieve balanced enzyme activities across multi-step pathways. In parallel, advances in protein engineering and computational design, particularly structure-guided modification of UGTs, are expected to improve catalytic specificity and turnover. Future research should emphasize developing metabolically robust microbial hosts with optimized energy and redox balance, exploring novel glycosyltransferases with superior kinetics, and applying systems biology and synthetic biology tools to fine-tune metabolic flux and accelerate strain optimization. Furthermore, while significant milestones have been achieved at the laboratory scale, the transition to industrial manufacturing remains a formidable challenge. Currently, there is a critical need for comprehensive technoeconomic analyses (TEA) to evaluate the practical economic viability of these microbial platforms against traditional plant extraction. Future commercial scaling efforts must address process robustness under industrial fermentation conditions, evaluate downstream purification economics, and navigate the complex regulatory frameworks for microbially derived food and pharmaceutical ingredients. Bridging this gap will require deeper interdisciplinary collaboration among metabolic and biochemical engineers and industrial stakeholders. The ultimate aim is to create genetically stable, high-yield microbial cell factories capable of producing gastrodin efficiently and sustainably at an industrial scale.
Table 1. Microbial biosynthesis of gastrodin and strategies for optimizing associated pathways.
|
Parental Strain |
Engineering Strategy |
Substrate |
Cultivation |
Titer (g/L) |
Yield (g/g) |
Productivity (mg/L/h) |
Ref. |
|---|---|---|---|---|---|---|---|
|
E. coli BL21(DE3) |
Constructed a complete de novo biosynthetic pathway by introducing Nocardia CAR, endogenous E. coli ADHs, and Rhodiola UGT73B6; enhanced catalytic efficiency via directed evolution (F389S); simultaneously reinforced precursor supply through overexpression of aroGD146N, ppsA, and ubiC. |
Glucose |
Shake Flask |
0.55 |
0.028 |
11.35 |
[40] |
|
E. coli BL21(DE3) |
Enhanced catalytic efficiency of A. thaliana UGT (AtUGT) via triple mutant T282S/A294E/V463L, strengthened aromatic precursor supply by co-expressing key genes (aroF, aroG, tktA, aroL, ubiC), and reconstructed an independent UDPG regeneration pathway to ensure sufficient sugar donor availability. |
Glucose |
Shake flask & Fermenter |
2.18 & 13.73 |
0.11 |
143.02 |
[77] |
|
E. coli BL21(DE3) |
Established a heterologous glycosylation system by synthesizing and codon-optimizing S. lycopersicum UGT (SlyUGT) for high-level expression in E. coli, enabling efficient biotransformation of pHBAL. |
pHBAL |
Enzymatic Reaction |
0.56 |
- |
93.31 |
[100] |
|
E. coli BL21(DE3) |
Optimized enzyme expression and reaction conditions by codon-optimizing R. serpentina RsUGT, expressing it as a soluble GST-fusion protein, and enhancing in vitro activity through DMSO-assisted catalysis. |
pHBAL |
Shake Flask |
0.23 |
- |
37.67 |
[25] |
|
E. coli BL21(DE3) |
Performed rational enzyme engineering of AtUGT through sequence screening, structure-based residue analysis (His19, Asp117), and metal ion optimization (Mg2+), improving catalytic efficiency and substrate affinity. |
pHBAL |
Shake Flask |
2.67 g/L |
- |
222.5 |
[64] |
|
E. coli BL21(DE3) |
Developed a coupled glycosylation-donor regeneration system by co-expressing I. tinctoria ItUGT2 and G. max GmSuSy, establishing an in situ UDPG recycling mechanism that uses sucrose as a low-cost glucose donor. |
pHBAL |
Shake Flask |
0.22 |
- |
6.11 |
[62] |
|
E. coli BL21(DE3) |
Enhanced UDPG availability through multi-level metabolic rewiring: overexpressing endogenous pgm and galU, introducing GmSuSy and P. aeruginosa PPK for UTP regeneration, and improving glycosyltransferase performance via site-directed mutagenesis ItUGTA347V. |
pHBAL |
Shake flask & Fermenter |
4.56 & 4.25 |
- |
126.67 |
[63] |
|
E. coli BL21(DE3) |
Engineered a highly efficient glycosylation system by coupling an evolved AtUGT triple mutant M4 (N94L/N221G/I343V) with a push-pull UDPG regeneration module and pgm deletion, while optimizing pHBAL loading to mitigate substrate cytotoxicity. |
pHBAL |
Fermenter |
22.9 |
- |
1908.33 |
[65] |
|
S. cerevisiae BY4742 |
Integrated pathway and flux optimization by introducing E. coli UbiC, Nocardia CAR, and R. serpentina RsUGT; blocking competing routes (ARO7 disruption) and enhancing shikimate flux through overexpression of ppsA, tktA, and feedback-resistant ARO4K229L. |
Glucose |
Shake flask |
2.1 |
0.11 |
12.50 |
[69] |
|
S. cerevisiae CEN.PK2-1C |
Reprogrammed gastrodin biosynthesis through three modular strategies: expressing the highly active AtUGT72B1 quadruple mutant E83T/R81E/V144M/I310L, redirecting shikimate flux using Aro3D154C and Aro7E198Q, and elevating central carbon precursors (E4P, PEP) and cofactors (NADPH, UDPG). |
Glucose |
Shake flask & Fermenter |
5.44 & 90.34 |
0.23 |
470.52 |
[74] |
|
Y. lipolytica Po1fΔKu70 |
Constructed a heterologous biosynthetic module (UbiC, CAR, SFP, AS) combined with shikimate pathway enhancement (DHS1, DHS2, ylARO4K221L, aroGG146N) and deletion of competing branches; employed CRISPR/Cas9 for multiplex genome editing to streamline carbon flux. |
Glucose |
Shake flask & Fermenter |
6.68 & 13.22 |
0.10 |
91.81 |
[75] |
|
Y. lipolytica po1g |
Implemented a systems-level optimization in Y. lipolytica, integrating heterologous biosynthesis (EcUbiC, NcCAR, SyIUGT), UDPG supply enhancement via promoter engineering (yIPGM2, yIUGP1, yIAK9), glucosidase deletion, and dynamic metabolic control through the CATR regulatory system. |
Glucose |
Shake flask & Fermenter |
9.48 & 13.4 |
0.15 |
111.67 |
[76] |
Author Contributions
X.W.: Writing-original draft, Funding acquisition. Z.J.: Writing—original draft. F.W.: Supervision, Funding acquisition. Z.W.: Project administration. X.L.: Writing-review & editing, Conceptualization.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data is available from the corresponding author upon reasonable request.
Funding
This work was financially supported by the China Postdoctoral Science Foundation (Grant No. 2024M751423), the National Natural Science Foundation of Jiangsu Province (Grant No. BK20240672), the Postdoctoral Fellowship Program of CPSF (Grant No. GZC20240705), the Jiangsu Province Excellent Postdoctoral Program (Grant No. 2024ZB065).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Shukla MK, Monika, Thakur A, Verma R, Lalhlenmawia H, Bhattacharyya S, et al. Unravelling the therapeutic potential of orchid plant against cancer. South. Afr. J. Bot. 2022, 150, 69–79. DOI:10.1016/j.sajb.2022.07.005 [Google Scholar]
-
Wang YL, Bai MT, Wang X, Peng ZL, Cai CY, Xi JJ, et al. Gastrodin: a comprehensive pharmacological review. Naunyn-Schmiedebergs Arch. Pharmacol. 2024, 397, 3781–3802. DOI:10.1007/s00210-023-02920-9 [Google Scholar]
-
Li Y, Wang Z, Liu D, Wang X, Zhou H. Research advances in chemical constituents of Gastrodia elata Blume. Shandong Sci. 2016, 29, 24–29. DOI:10.3976/j.issn.1002-4026.2016.04.006 [Google Scholar]
-
Zhong Y, Li J, Liu H, Wang Y. The traditional uses, phytochemistry and pharmacology of Gastrodia elata Blume: A comprehensive review. Arab. J. Chem. 2025, 18, 106086. DOI:10.1016/j.arabjc.2024.106086 [Google Scholar]
-
Zhang Y, Zhao X, Gong L, Lai C, Liu J, Xie J. Neuroprotective effects of Gastrodia elata and its compounds in a Caenorhabditis elegans Alzheimer’s disease model. Phytomedicine 2025, 143, 156876. DOI:10.1016/j.phymed.2025.156876 [Google Scholar]
-
He P, Hu Y, Huang C, Wang X, Zhang H, Zhang X, et al. N-Butanol Extract of Gastrodia elata Suppresses Inflammatory Responses in Lipopolysaccharide-Stimulated Macrophages and Complete Freund’s Adjuvant- (CFA-) Induced Arthritis Rats via Inhibition of MAPK Signaling Pathway. Evid. -Based Complement. Altern. Med. 2020, 2020, 1658618. DOI:10.1155/2020/1658618 [Google Scholar]
-
He X, Yang Y, Yuan X, Sun Y, Li Y. Chemical composition and anticonvulsant activities of herb pair of Gastrodia elata Blume-Acorus tatarinowii Schott decoction on experimentally induced seizures in mice. Metab. Brain Dis. 2023, 38, 1877–1893. DOI:10.1007/s11011-023-01211-9 [Google Scholar]
-
Jung JW, Yoon BH, Oh HR, Ahn JH, Kim SY, Park SY, et al. Anxiolytic-Like effects of Gastrodia elata and its phenolic constituents in mice. Biol. Pharm. Bull. 2006, 29, 261–265. DOI:10.1248/bpb.29.261 [Google Scholar]
-
Zhou X, Shao J, Xie X, Xu Y, Shao T, Jin Z. Effectiveness of gastrodin for migraine: A meta-analysis. Front. Neurol. 2022, 13, 939401. DOI:10.3389/fneur.2022.939401 [Google Scholar]
-
Zhang M, Liu Y, Du H, Shi Y, Subihinur K. Research progress in the treatment of dizziness based on gastrodia elata. MEDS Chin. Med. 2023, 5, 21–24. DOI:10.23977/medcm.2023.050605 [Google Scholar]
-
Huang SC, Huang HC, Liao WL, Kao ST, Cheng CY. Neuroprotective effects of Gastrodia elata Blume on promoting M2 microglial polarization by inhibiting JNK/TLR4/T3JAM/NF-κB signaling after transient ischemic stroke in rats. Front. Pharmacol. 2024, 15, 1469602. DOI:10.3389/fphar.2024.1469602 [Google Scholar]
-
Yang L, Tsai TH. Neuroprotective effect and herbal-drug pharmacokinetic interaction of Gastrodia elata extract on valproic acid. Biomed. Pharmacother. 2022, 156, 113938. DOI:10.1016/j.biopha.2022.113938 [Google Scholar]
-
Wang Y, Zhao M, Xie C, Li L, Lin L, Li Q, et al. Fermented Gastrodia elata Bl. Alleviates Cognitive Deficits by Regulating Neurotransmitters and Gut Microbiota in D-Gal/AlCl3-Induced Alzheimer’s Disease-like Mice. Foods 2024, 13, 2154. DOI:10.3390/foods13132154 [Google Scholar]
-
Kim HJ, Moon KD, Oh SY, Kim SP, Lee SR. Ether fraction of methanol extracts of Gastrodia elata, a traditional medicinal herb, protects against kainic acid-induced neuronal damage in the mouse hippocampus. Neurosci. Lett. 2001, 314, 65–68. DOI:10.1016/S0304-3940(01)02296-0 [Google Scholar]
-
Ma JF, Xu FJ, Zhou X, Yin H, Wang H, Wang CY, et al. Quality assessment of Gastrodia elata Blume (Tianma) based on Vis-NIR spectroscopy: Discrimination of harvest times and prediction of quality indicator contents. J. Food Compos. Anal. 2024, 134, 106486. DOI:10.1016/j.jfca.2024.106486 [Google Scholar]
-
Alshalalfeh M, Sun N, Moraes AH, Utani APA, Xu Y. Conformational Distributions of Phenyl β-D-Glucopyranoside and Gastrodin in Solution by Vibrational Optical Activity and Theoretical Calculations. Molecules 2023, 28, 4013. DOI:10.3390/molecules28104013 [Google Scholar]
-
Lai Y, Wang R, Li W, Zhu H, Fei S, Shi H, et al. Clinical and economic analysis of Gastrodin injection for dizziness or vertigo: a retrospective cohort study based on electronic health records in China. Chin. Med. 2022, 17, 6. DOI:10.1186/s13020-021-00561-9 [Google Scholar]
-
Shi Z, Zhang Y, Xiao Y, Shi Z, Wei X, Wang B, et al. The protective effects of gastrodin on neurological disorders: an update and future perspectives. Front. Pharmacol. 2024, 15, 1494277. DOI:10.3389/fphar.2024.1494277 [Google Scholar]
-
Dai JN, Zong Y, Zhong LM, Li YM, Zhang W, Bian LG, et al. Gastrodin Inhibits Expression of Inducible NO Synthase, Cyclooxygenase-2 and Proinflammatory Cytokines in Cultured LPS-Stimulated Microglia via MAPK Pathways. PLoS ONE 2011, 6, e21891. DOI:10.1371/journal.pone.0021891 [Google Scholar]
-
El Menyiy N, Elouafy Y, Moubachir R, Abdnim R, Benali T, Taha D, et al. Chemistry, Biological Activities, and Pharmacological Properties of Gastrodin: Mechanism Insights. Chem. Biodivers. 2024, 21, e202400402. DOI:10.1002/cbdv.202400402 [Google Scholar]
-
He J, Li X, Yang S, Li Y, Lin X, Xiu M, et al. Gastrodin extends the lifespan and protects against neurodegeneration in the Drosophila PINK1 model of Parkinson’s disease. Food Funct. 2021, 12, 7816–7824. DOI:10.1039/D1FO00847A [Google Scholar]
-
Pei J. Inhibitory Effects of Gartrodin on Tyrosinase and Melanogenesis. Master’s Thesis, Soochow University, Suzhou, China, 2013. [Google Scholar]
-
Liu JJ, Yang XQ, Li ZY, Miao JY, Li SB, Zhang WP, et al. The role of symbiotic fungi in the life cycle of Gastrodia elata Blume (Orchidaceae): A comprehensive review. Front. Plant Sci. 2024, 14, 1309038. DOI:10.3389/fpls.2023.1309038 [Google Scholar]
-
Zhang Z, Li X, Zhang Y, Yin N, Wu G, Wei G, et al. Ecological factors impacting genetic characteristics and metabolite accumulations of Gastrodia elata. Chin. Herb. Med. 2025, 17, 562–574. DOI:10.1016/j.chmed.2024.09.002 [Google Scholar]
-
Ge L, Xia Y, Xu W, Jia R, Zhang T. Efficient Biosynthesis of Gastrodin by UDP-Glycosyltransferase from Rauvolfia serpentina. J. Microbiol. Biotechnol. 2025, 35, e2501002. DOI:10.4014/jmb.2501.01002 [Google Scholar]
-
Zhao L, Zhu Y, Jia H, Han Y, Zheng X, Wang M, et al. From Plant to Yeast-Advances in Biosynthesis of Artemisinin. Molecules 2022, 27, 6888. DOI:10.3390/molecules27206888 [Google Scholar]
-
Zhang J, Hansen LG, Gudich O, Viehrig K, Lassen LMM, Schrübbers L, et al. A microbial supply chain for production of the anti-cancer drug vinblastine. Nature 2022, 609, 341–347. DOI:10.1038/s41586-022-05157-3 [Google Scholar]
-
Hu ZF, Gu AD, Liang L, Li Y, Gong T, Chen JJ, et al. Construction and optimization of microbial cell factories for sustainable production of bioactive dammarenediol-II glucosides. Green. Chem. 2019, 21, 3286–3299. DOI:10.1039/C8GC04066D [Google Scholar]
-
Liu B, Chen J, Zhang W, Huang Y, Zhao Y, Juneidi S, Dekebo A, Wang M, Shi L, Hu X. The gastrodin biosynthetic pathway in Pholidota chinensis Lindl. revealed by transcriptome and metabolome profiling. Front. Plant Sci. 2022, 13, 1024239. DOI:10.3389/fpls.2022.1024239 [Google Scholar]
-
Tsai CC, Wu KM, Chiang TY, Huang CY, Chou CH, Li SJ, et al. Comparative transcriptome analysis of Gastrodia elata (Orchidaceae) in response to fungus symbiosis to identify gastrodin biosynthesis-related genes. BMC Genom. 2016, 17, 212. DOI:10.1186/s12864-016-2508-6 [Google Scholar]
-
Chen C, Mi H. Analysis of genes related to biosynthesis of gastrodin. Plant Physiol. J. 2021, 57, 1819–1828. DOI:10.13592/j.cnki.ppj.2021.0264 [Google Scholar]
-
Heiden A, Kobel K, Komenda M, Koppmann R, Shao M, Wildt J. Toluene emissions from plants. Geophys. Res. Lett. 1999, 26, 1283–1286. DOI:10.1029/1999GL900220 [Google Scholar]
-
Singer A, Crowley D, Thompson I. Secondary plant metabolites in phytoremediation and biotransformation. Trends Biotechnol. 2003, 21, 123–130. DOI:10.1016/S0167-7799(02)00041-0 [Google Scholar]
-
Xu Y, Wu Z, Gao Y, Zang P, Yang X, Zhao Y, et al. GePIF4 Increases the Multi-Flower/Capsule-Bearing Traits and Gastrodin Biosynthesis in Gastrodia elata. Plants 2025, 14, 1684. DOI:10.3390/plants14111684 [Google Scholar]
-
Khatri P, Chen L, Rajcan I, Dhaubhadel S. Functional characterization of Cinnamate 4-hydroxylase gene family in soybean (Glycine max). PLoS ONE 2023, 18, e0285698. DOI:10.1371/journal.pone.0285698 [Google Scholar]
-
Bassard JE, Richert L, Geerinck J, Renault H, Duval F, Ullmann P, et al. Protein-Protein and Protein-Membrane Associations in the Lignin Pathway. Plant Cell 2012, 24, 4465–4482. DOI:10.1105/tpc.112.102566 [Google Scholar]
-
Knosp S, Kriegshauser L, Tatsumi K, Malherbe L, Erhardt M, Wiedemann G, et al. An ancient role for CYP73 monooxygenases in phenylpropanoid biosynthesis and embryophyte development. Embo J. 2024, 43, 4092–4109. DOI:10.1038/s44318-024-00181-7 [Google Scholar]
-
Liu T, Yao R, Zhao Y, Xu S, Huang C, Luo J, et al. Cloning, Functional Characterization and Site-Directed Mutagenesis of 4-Coumarate: Coenzyme A Ligase (4CL) Involved in Coumarin Biosynthesis in Peucedanum praeruptorum Dunn. Front. Plant Sci. 2017, 8, 4. DOI:10.3389/fpls.2017.00004 [Google Scholar]
-
Rinaldi R, Jastrzebski R, Clough MT, Ralph J, Kennema M, Bruijnincx PCA, et al. Paving the Way for Lignin Valorisation: Recent Advances in Bioengineering, Biorefining and Catalysis. Angew. Chem. -Int. Ed. 2016, 55, 8164–8215. DOI:10.1002/anie.201510351 [Google Scholar]
-
Bai YF, Yin H, Bi HP, Zhuang YB, Liu T, Ma YH. De novo biosynthesis of Gastrodin in Escherichia coli. Metab. Eng. 2016, 35, 138–147. DOI:10.1016/j.ymben.2016.01.002 [Google Scholar]
-
Duan MZ, Bao LY, Dong L, He HY, Cheng LJ, Shi ZL, et al. Accumulation of Gastrodin, p-Hydroxybenzyl Alcohol, and Parishin in Gastrodia elata During Growth. J. Food Qual. 2025, 2025, 2555655. DOI:10.1155/jfq/2555655 [Google Scholar]
-
Yu E, Liu Q, Gao YG, Li YQ, Zang P, Zhao Y, et al. An exploration of mechanism of high quality and yield of Gastrodia elata Bl. f. glauca by the isolation, identification, and evaluation of Mycena. Front. Microbiol. 2023, 14, 1220670. DOI:10.3389/fmicb.2023.1220670 [Google Scholar]
-
Hu J, Feng Y, Zhong HT, Liu W, Tian XF, Wang YH, et al. Impact of climate change on the geographical distribution and niche dynamics of Gastrodia elata. Peerj 2023, 11, e15741. DOI:10.7717/peerj.15741 [Google Scholar]
-
Yuan QS, Wang H, Jiang WK, Ou XH, Xu J, Wang XA, et al. Dissection of seed development of Gastrodia elata at different temperatures. China J. Chin. Mater. Medica 2020, 45, 485–490. DOI:10.19540/j.cnki.cjcmm.20191204.109 [Google Scholar]
-
Dai Y, Ban W, Yang Z. Gastrodin, a Promising Natural Small Molecule for the Treatment of Central Nervous System Disorders, and Its Recent Progress in Synthesis, Pharmacology and Pharmacokinetics. Int. J. Mol. Sci. 2024, 25, 9540. DOI:10.3390/ijms25179540 [Google Scholar]
-
Li HB, Chen F. Preparative isolation and purification of gastrodin from the Chinese medicinal plant Gastrodia elata by high-speed counter-current chromatography. J. Chromatogr. A 2004, 1052, 229–232. DOI:10.1016/j.chroma.2004.09.005 [Google Scholar]
-
He Y, Wang Y, Peng X, Yang Z, Wang R. Research progress on gastrodin extraction methods. Yunnan Chem. Technol. 2025, 52, 17–19. DOI:10.3969/j.issn.1004-275X.2025.06.04 [Google Scholar]
-
Zhang M, Zhao J, Dai X, Li X. Extraction and Analysis of Chemical Compositions of Natural Products and Plants. Separations 2023, 10, 598. DOI:10.3390/separations10120598 [Google Scholar]
-
Ma Y, Meng A, Liu P, Chen Y, Yuan A, Dai Y, et al. Reflux Extraction Optimization and Antioxidant Activity of Phenolic Compounds from Pleioblastus amarus (Keng) Shell. Molecules 2022, 27, 362. DOI:10.3390/molecules27020362 [Google Scholar]
-
Liao X, Wu Z, Yang Y, Chen G, Zhan Y. The Differentiation of Total Polyphenols and Total Flavonoids Contents in Gastrodia elata Bl. f. glauca S. Chow Extracts Using Different Solvents and Their Antioxidant Capacities. Food Ferment. Ind. 2022, 48, 157–164. DOI:10.13995/j.cnki.11-1802/ts.027943 [Google Scholar]
-
Abubakar AR, Haque M. Preparation of medicinal plants: Basic extraction and fractionation procedures for experimental purposes. J. Pharm. Bioallied Sci. 2020, 12, 1–10. DOI:10.4103/jpbs.JPBS_175_19 [Google Scholar]
-
Ong ES, Heng MY, Tan SN, Yong JWH, Koh H, Teo CC, et al. Determination of gastrodin and vanillyl alcohol in Gastrodia elata Blume by pressurized liquid extraction at room temperature. J. Sep. Sci. 2007, 30, 2130–2137. DOI:10.1002/jssc.200700002 [Google Scholar]
-
Zeng S, Luo Q, Tian Y, Chen S, Zheng X, Zhao Z, et al. Microwave-assisted Extraction of Gastrodin from Gastrodiaelata and Its Content Determination. Guangdong Chem. Ind. 2019, 11. [Google Scholar]
-
Li L, Zhao J, Yang T, Sun B. High-speed countercurrent chromatography as an efficient technique for large separation of plant polyphenols: A review. Food Res. Int. 2022, 153, 110956. DOI:10.1016/j.foodres.2022.110956 [Google Scholar]
-
Tian M, Gao Q, Yi Y, Guo D, Liu Y. Study on Ultrasonic Assisted Enzyme Extraction, Aqueous Two-Phase Extraction and Macroporous Resin Purification of Gastrodin. Guid. J. Tradit. Chin. Med. Pharm. 2024, 30. DOI:10.13862/j.cn43-1446/r.2024.02.010 [Google Scholar]
-
Zhou J, Yang Y, Yang C. Chemical study on Gastrodia elata BL. Acta Chim. Sin. 1980, 38, 162–166. Available online: https://sioc-journal.cn/Jwk_hxxb/EN/Y1980/V38/I2/162 (accessed on 22 June 2026).
-
Li Y, Ma C. Method for Chemically Synthesising Gastrodin. CN102977161B, 17 December 2012. [Google Scholar]
-
Wang D. The Production Technique of Gastrodine. CN104072549B, 11 July 2014. [Google Scholar]
-
Cheng YY, Wang XL, Li C, Ji J, Liu TF, Duan KX. Highly efficient Agrobacterium rhizogenes-mediated hairy root transformation for gene functional and gene editing analysis in soybean. Plant Methods 2021, 17, 73. DOI:10.1186/s13007-021-00778-7 [Google Scholar]
-
Peng C, Zhang M, Liu Q, Zheng L, Zhang X. Study on the Hairy Root Induced from Datura tatula L. and Production Study on the Hairy Root Induced from Datura tatula L. and Production. J. Yunnan Agric. Univ. Nat. Sci. 2008, 23, 492–497.DOI:10.3969/j.issn.1004-390X.2008.04.013 [Google Scholar]
-
Cai J, Ding J, Hua Y, Li N. Establishment of biotransformation system of the gastrodin biosynthesis by hairy root of Panax ginseng. J. Plant Resour. Environ. 2005, 14, 29–31.DOI:10.3969/j.issn.1674-7895.2005.02.007 [Google Scholar]
-
Cui CX, Yan JY, Liu YT, Zhang Z, Su QY, Kong MY, et al. One-pot biosynthesis of gastrodin using UDP-glycosyltransferase itUGT2 with an in situ UDP-glucose recycling system. Enzym. Microb. Technol. 2023, 166, 110226. DOI:10.1016/j.enzmictec.2023.110226 [Google Scholar]
-
Zhang Z, Wang XY, Sha X, Qiao GH, Niu FY, Ming H, et al. Biosynthesis of gastrodin via multi-module UDPG supply and site-directed mutagensis of glycosyltransferase in Escherichia coli. Mol. Catal. 2025, 578, 114953. DOI:10.1016/j.mcat.2025.114953 [Google Scholar]
-
Xia H, Zhang ZH, Ding J, Jiang KH, Xue F. A Glycosyltransferase from Arabidopsis thaliana Enables the Efficient Enzymatic Synthesis of Gastrodin. Catal. Lett. 2024, 154, 1558–1566. DOI:10.1007/s10562-023-04406-y [Google Scholar]
-
Wang X, Zhang JL, Li T, Wang F, Wang ZG, Li X. Efficient whole-cell biotransformation for gastrodin production via glycosyltransferase engineering and process optimization. Synth. Syst. Biotechnol. 2026, 14, 10–21. DOI:10.1016/j.synbio.2026.03.014 [Google Scholar]
-
Zhu H. Studies on Biosynthesis of Gastrodin and Other Bioactive Natural Products by Microbial Transformation. Ph.D. Dissertation, Northwest University, Xian, China, 2007. [Google Scholar]
-
Fan LL, Dong YC, Xu TY, Zhang HF, Chen QH. Gastrodin Production from p-2-Hydroxybenzyl Alcohol Through Biotransformation by Cultured Cells of Aspergillus foetidus and Penicillium cyclopium. Appl. Biochem. Biotechnol. 2013, 170, 138–148. DOI:10.1007/s12010-013-0166-6 [Google Scholar]
-
Zhang HF, He GQ, Liu J, Ruan H, Chen QH, Zhang Q, et al. Production of gastrodin through biotransformation of p-2-hydroxybenzyl alcohol by cultured cells of Armillaria luteo-virens Sacc. Enzym. Microb. Technol. 2008, 43, 25–30. DOI:10.1016/j.enzmictec.2008.03.007 [Google Scholar]
-
Yin H, Hu TD, Zhuang YB, Liu T. Metabolic engineering of Saccharomyces cerevisiae for high-level production of gastrodin from glucose. Microb. Cell Factories 2020, 19, 218. DOI:10.1186/s12934-020-01476-0 [Google Scholar]
-
Hefner T, Arend J, Warzecha H, Siems K, Stöckigt J. Arbutin synthase, a novel member of the NRD1β glycosyltransferase family, is a unique multifunctional enzyme converting various natural products and xenobiotics. Bioorganic Med. Chem. 2002, 10, 1731–1741. DOI:10.1016/S0968-0896(02)00029-9 [Google Scholar]
-
Hansen EH, Moller BL, Kock GR, Buenner CM, Kristensen C, Jensen OR, et al. De. Novo Biosynthesis of Vanillin in Fission Yeast (Schizosaccharomyces pombe) and Baker’s Yeast (Saccharomyces cerevisiae). Appl. Environ. Microbiol. 2009, 75, 2765–2774. DOI:10.1128/AEM.02681-08 [Google Scholar]
-
Siebert M, Severin K, Heide L. Formation of 4-hydroxybenzoate in Escherichia coli: characterization of the ubiC gene and its encoded enzyme chorismate pyruvate-lyase. Microbiology 1994, 140 Pt 4, 897–904. DOI:10.1099/00221287-140-4-897 [Google Scholar]
-
Averesch NJH, Krömer JO. Metabolic Engineering of the Shikimate Pathway for Production of Aromatics and Derived Compounds-Present and Future Strain Construction Strategies. Front. Bioeng. Biotechnol. 2018, 6, 32. DOI:10.3389/fbioe.2018.00032 [Google Scholar]
-
Wan XW, Li DH, Tang MH, Liu ZW, Pan JY, Yu HB, et al. System-level metabolic and enzyme engineering enables high-titer gastrodin biosynthesis in yeast. Bioresour. Technol. 2026, 454, 134802. DOI:10.1016/j.biortech.2026.134802 [Google Scholar]
-
Wu YQ, Li SC, Sun BJ, Guo JY, Zheng MY, Li AT. Enhancing Gastrodin Production in Yarrowia lipolytica by Metabolic Engineering. Acs Synth. Biol. 2024, 13, 1332–1342. DOI:10.1021/acssynbio.4c00050 [Google Scholar]
-
Gu Y, Jiang YR, Li CF, Zhu J, Lu XY, Ge JY, et al. High titer production of gastrodin enabled by systematic refactoring of yeast genome and an antisense-transcriptional regulation toolkit. Metab. Eng. 2024, 82, 250–261. DOI:10.1016/j.ymben.2024.02.016 [Google Scholar]
-
Wang X, Jing ZW, Chen JM, Kong XL, Wang F, Wang ZG, et al. High-level de novo biosynthesis of gastrodin in Escherichia coli by protein and metabolic engineering. ACS Sustain. Chem. Eng. 2026, 14, 6537–6547. DOI:10.1021/acssuschemeng.6c00919 [Google Scholar]
-
Escalante A, Cervantes AS, Gosset G, Bolívar F. Current knowledge of the Escherichia coli phosphoenolpyruvate-carbohydrate phosphotransferase system: peculiarities of regulation and impact on growth and product formation. Appl. Microbiol. Biotechnol. 2012, 94, 1483–1494. DOI:10.1007/s00253-012-4101-5 [Google Scholar]
-
Meza E, Becker J, Bolivar F, Gosset G, Wittmann C. Consequences of phosphoenolpyruvate: Sugar phosphotranferase system and pyruvate kinase isozymes inactivation in central carbon metabolism flux distribution in Escherichia coli. Microb. Cell Factories 2012, 11, 127. DOI:10.1186/1475-2859-11-127 [Google Scholar]
-
Ponce E, Flores N, Martinez A, Valle F, Bolivar F. Cloning of the two pyruvate kinase isoenzyme structural genes from Escherichia coli: the relative roles of these enzymes in pyruvate biosynthesis. J. Bacteriol. 1995, 177, 5719–5722. DOI:10.1128/jb.177.19.5719-5722.1995 [Google Scholar]
-
Zhao CH, Lin Z, Dong HJ, Zhang YP, Li Y. Reexamination of the Physiological Role of PykA in Escherichia coli Revealed that It Negatively Regulates the Intracellular ATP Levels under Anaerobic Conditions. Appl. Environ. Microbiol. 2017, 83, e00316-17. DOI:10.1128/AEM.00316-17 [Google Scholar]
-
Fujiwara R, Noda S, Tanaka T, Kondo A. Metabolic engineering of Escherichia coli for shikimate pathway derivative production from glucose-xylose co-substrate. Nat. Commun. 2020, 11, 279. DOI:10.1038/s41467-019-14024-1 [Google Scholar]
-
Jiang M, Zhang H. Engineering the shikimate pathway for biosynthesis of molecules with pharmaceutical activities in E. coil. Curr. Opin. Biotechnol. 2016, 42, 1–6. DOI:10.1016/j.copbio.2016.01.016 [Google Scholar]
-
Herrmann KM, Weaver LM. THE SHIKIMATE PATHWAY. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 473–503. DOI:10.1146/annurev.arplant.50.1.473 [Google Scholar]
-
Gupte RS, Ata H, Rawat D, Abe M, Taylor MS, Ochi R, et al. Glucose-6-Phosphate Dehydrogenase Is a Regulator of Vascular Smooth Muscle Contraction. Antioxid. Redox Signal. 2011, 14, 543–558. DOI:10.1089/ars.2010.3207 [Google Scholar]
-
Chandran SS, Yi J, Draths KM, von Daeniken R, Weber W, Frost JW. Phosphoenolpyruvate availability and the biosynthesis of shikimic acid. Biotechnol. Progress. 2003, 19, 808–814. DOI:10.1021/bp025769p [Google Scholar]
-
Gu PF, Fan XY, Liang QF, Qi QS, Li Q. Novel technologies combined with traditional metabolic engineering strategies facilitate the construction of shikimate-producing Escherichia coli. Microb. Cell Factories 2017, 16, 167. DOI:10.1186/s12934-017-0773-y [Google Scholar]
-
Li X, Liu YH, Ma L, Jiang WJ, Shi TG, Li LX, et al. Metabolic engineering of Escherichia coli for high-yield dopamine production via optimized fermentation strategies. Appl. Environ. Microbiol. 2025, 91, e00159-25. DOI:10.1128/aem.00159-25 [Google Scholar]
-
Pereira SA, Livi GP. Cloning and expression of the ARO3 gene encoding DAHP synthase from Candida albicans. Gene 1993, 132, 159–165. DOI:10.1016/0378-1119(93)90191-5 [Google Scholar]
-
Helmstaedt K, Strittmatter A, Lipscomb WN, Braus GH. Evolution of 3-deoxy-D-arabino-heptulosonate-7-phosphate synthase-encoding genes in the yeast Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 2005, 102, 9784–9789. DOI:10.1073/pnas.0504238102 [Google Scholar]
-
Stogios PJ, Liston SD, Semper C, Quade B, Michalska K, Evdokimova E, et al. Molecular analysis and essentiality of Aro1 shikimate biosynthesis multi-enzyme in Candida albicans. Life Sci. Alliance 2022, 5, e202101358. DOI:10.26508/lsa.202101358 [Google Scholar]
-
Tu S, Wang JJ, Yang PM, He Y, Gong ZX, Zhong WH. Enhanced chlorogenic acid production from glucose via systematic metabolic engineering of Saccharomyces cerevisiae. Synth. Syst. Biotechnol. 2025, 10, 707–718. DOI:10.1016/j.synbio.2025.03.004 [Google Scholar]
-
Wang M, Zeng J, Zhu Y, Chen X, Guo Q, Tan H, Cui B, Song S, Deng Y. A 4-Hydroxybenzoic Acid-Mediated Signaling System Controls the Physiology and Virulence of Shigella sonnei. Microbiol. Spectr. 2023, 11, e0483522. DOI:10.1128/spectrum.04835-22 [Google Scholar]
-
Lin CY, Eudes A. Strategies for the production of biochemicals in bioenergy crops. Biotechnol. Biofuels 2020, 13, 71. DOI:10.1186/s13068-020-01707-x [Google Scholar]
-
Feng YY, Yao MD, Wang Y, Ding MZ, Zha J, Xiao WH, et al. Advances in engineering UDP-sugar supply for recombinant biosynthesis of glycosides in microbes. Biotechnol. Adv. 2020, 41, 107538. DOI:10.1016/j.biotechadv.2020.107538 [Google Scholar]
-
Pandey RP, Malla S, Simkhada D, Kim BG, Sohng JK. Production of 3-O-xylosyl quercetin in Escherichia coli. Appl. Microbiol. Biotechnol. 2013, 97, 1889–1901. DOI:10.1007/s00253-012-4438-9 [Google Scholar]
-
Pei JJ, Dong P, Wu T, Zhao LG, Fang XY, Cao FL, et al. Metabolic Engineering of Escherichia coli for Astragalin Biosynthesis. J. Agric. Food Chem. 2016, 64, 7966–7972. DOI:10.1021/acs.jafc.6b03447 [Google Scholar]
-
Pei JJ, Sun Q, Zhao LG, Shi H, Tang F, Cao FL. Efficient Biotransformation of Luteolin to Isoorientin through Adjusting Induction Strategy, Controlling Acetic Acid, and Increasing UDP-Glucose Supply in Escherichia coli. J. Agric. Food Chem. 2019, 67, 331–340. DOI:10.1021/acs.jafc.8b05958 [Google Scholar]
-
Sun C, Lou M, Li Z, Cheng F, Li Z. Combining an Enhanced Polyphosphate Kinase-Driven UDP-Glucose Regeneration System with the Screening of Key Glycosyltransferases for Efficient In Vitro Synthesis of Nucleoside Disaccharides. J. Agric. Food Chem. 2024, 72, 20557–20567. DOI:10.1021/acs.jafc.4c05329 [Google Scholar]
-
Ge L, Xu WX, Jia RB, Xia Y. Recombinant Expression and Characterization of an Alkali-tolerant UDP-Glycosyltransferase from Solanum lycopersicum and Its of Gastrodin. J. Microbiol. Biotechnol. 2025, 35, e2410029. DOI:10.4014/jmb.2410.10029 [Google Scholar]