Comprehensive Effects of Flashing/Pulsed Light on Microalgae: Molecular Mechanisms and Biotechnological Applications

Comprehensive Effects of Flashing/Pulsed Light on Microalgae: Molecular Mechanisms and Biotechnological Applications
Received: 07 March 2026 Revised: 30 March 2026 Accepted: 08 June 2026 Published: 25 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Concept, Historical Development and Research Evolution
The historical evolution of research surrounding the use of flashing/pulsed light in the cultivation of microalgae is based on the essential observation that illumination or intermittent light, which refers to short high-intensity light pulses followed by dark periods such that the time-averaged light intensity equals that of continuous illumination, can enhance biomass yields when compared to continuous light as a result of temporal decoupling in photosynthesis [1,2].
Research on the mechanism of light quality and photoinhibition began in the early foundation period (1993–2007). It was reported that blue light causes acidification on the surface of the cell in macroalgae [3], while similar light-sensing mechanisms were subsequently explored in microalgae, and mechanistic models of PSII photodamage were put forward [4], as well as the concepts of ultrahigh algal productivity under optimized light delivery [5]. These early studies identified kinetic mismatches between photochemical reactions and carbon fixation in photosynthesis, which flashing light later helped to overcome. During the experimental validation period from 2011 to 2015, empirical evidence emerged for the flashing light effect. In several studies, it was demonstrated that Chlamydomonas reinhardtii exhibited higher photosynthetic efficiency and growth rate under flashing regimes [6], gained performance benefits from continuous background light supplemented with flashing light [7], and showed increased performance under attenuated flashing regimes [8]. During a systematic synthesis period (2016), the first major review consolidated the available information in this field and acknowledged that flashing light can improve biomass and metabolites in microalgal biotechnology [1]. During the engineering realization period (2017–2020), practically oriented reactor designs multiplied and encompassed LED flashing light for production [9], 10 kHz pulsing in biofilms yielding a 7-fold net photosynthesis enhancement [10], and hydrodynamic baffles or fiber-optic systems inducing natural flashing [11].
During the phase of parameter refinement and metabolic targeting (2021–2023), the emphasis of research switched to refined parameters and selected outcomes: flashing-induced proteins, polyunsaturated fatty acids (PUFAs), and pigments [12,13]; theoretical models of the predicted cell growth at variable flashing conditions [14]. During the modeling and scale-up period (2024), integrated predictive mathematical modeling of high-intensity/high-frequency regimes and photosynthetic unit (PSU) dynamics provided frameworks for minimizing photoinhibition and supporting industrialized scale-up [15].
To avoid ambiguity in the subsequent sections, we explicitly define and distinguish the various illumination regimes discussed in this review, as they operate on fundamentally different timescales and elicit distinct physiological responses [1,2]. Flashing light and pulsed light are used interchangeably to describe regimes with microsecond-to-millisecond (µs–ms) light pulses followed by dark periods of comparable duration [1,6], typically generated by electronic control of LEDs [9]. Intermittent light is a broader term encompassing any discontinuous illumination from milliseconds to seconds [1]. Light-dark (L/D) cycles refer to second-to-hour scale alternations (e.g., 12 h:12 h) that mimic natural diel rhythms, primarily influencing circadian regulation rather than photochemical efficiency [16]. We also distinguish externally imposed pulsing (programmed LED switching) from hydrodynamically induced light-dark exposure (passive cycling via mixing, baffles, or bubbles) [11,17,18]. Key parameters governing efficacy include frequency (f, Hz), duty cycle (DC = light period/total period), peak intensity (I0, µmol·m−2·s−1), and average intensity (Iavg = I0 × DC) [6,19]. Figure 1 provides a conceptual schematic graph comparing continuous light, high-frequency pulsed light (µs–ms), low-frequency flashing light (ms–s), and diel L/D cycles (h) with their typical effects on PSII dynamics.
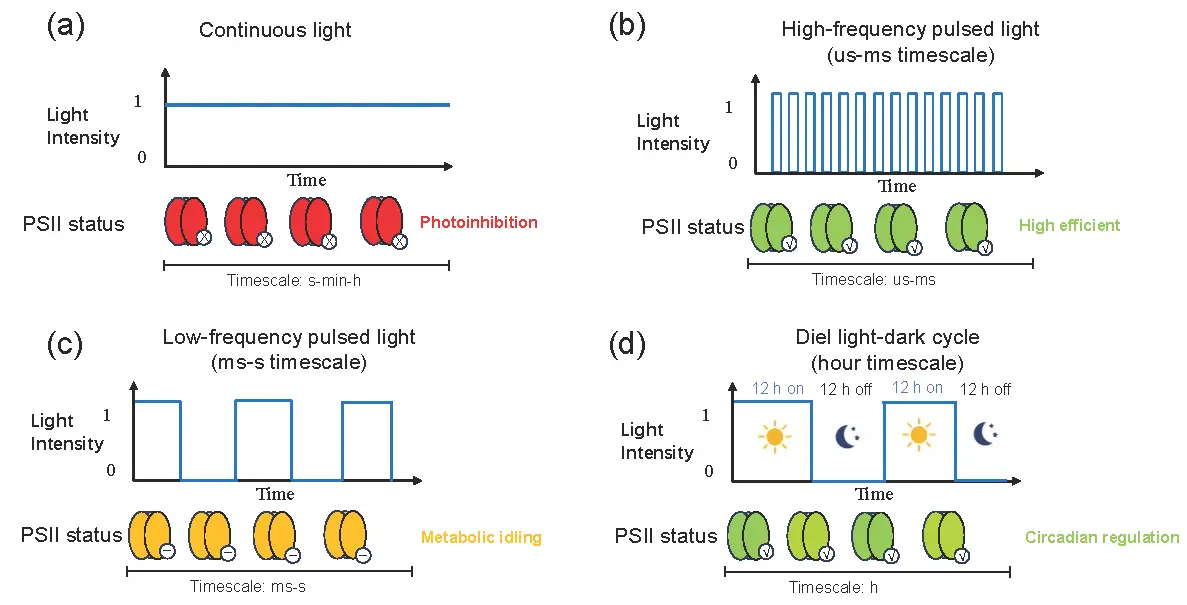
Figure 1. Schematic illustration of four illumination regimes and their effects on PSII reaction centers. (a) Continuous light: constant intensity → PQ pool over-reduction → PSII centers closed (red), photoinhibition. (b) High-frequency pulsed light (µs–ms): short pulses + dark intervals → PQ re-oxidation → PSII open (green), high quantum yield. (c) Low-frequency flashing light (ms–s): longer dark periods → partial recovery but risk of metabolic idling (yellow). (d) Diel light-dark cycle (h): extended dark → circadian regulation, no rapid photochemical gain. Key parameters: frequency, duty cycle, peak and average intensity.
2. The Flashing/Pulsed Light Effect: Phenomenon, Principle and Advantages
The flashing/pulsed light effect is a well-documented phenomenon in which intermittent illumination at an equivalent time-averaged intensity yields superior photosynthetic efficiency, biomass productivity, and target metabolite accumulation compared to continuous illumination [1]. This effect fundamentally arises from the kinetic decoupling of the ultrafast photochemical reactions within the photosystems (picosecond to millisecond scale) from the significantly slower downstream biochemical processes, notably the Calvin-Benson cycle (millisecond to second scale) [2]. By providing brief dark intervals, the flashing/pulsed light allows the enzymatic machinery to process the products of the light reactions, thus reducing photoinhibition and improving quantum yield [6,14]. This principle has been validated across a wide range of microalgal species, demonstrating its considerable promise for biotechnological applications. To provide a clear conceptual framework, we distinguish four interconnected levels of analysis throughout this review. At the molecular/cellular level, we focus on intrinsic mechanisms such as PSII reaction center dynamics, plastoquinone (PQ) pool redox state, and the temporal decoupling of electron transport from carbon fixation. At the physiological level, we discuss cellular responses, including cell growth rate, biomass accumulation, and the synthesis of high-value metabolites. At the reactor scale, we consider extrinsic effects induced by engineering design, such as hydrodynamically generated light-dark cycling, light gradients, self-shading, and bubble-induced scattering. At the engineering outcome level, we evaluate overall performance metrics including biomass productivity, energy consumption, and cost-effectiveness. This layered framework is intended to help readers distinguish between cell-autonomous phenomena and process-induced effects.
2.1. Enhanced Biomass Productivity
A primary advantage of flashing/pulsed light effect is the significant enhancement of biomass production. For instance, by a flashing light-inducing effect via hydrodynamic baffles in a flat-plate photobioreactor, the dry cell weight of Chlorella sp. was increased by 39.23% over conventional designs [17]. It is important to note that while some reports show dramatic increases, such as a 61-fold higher biomass for Nannochloropsis sp. under low-frequency blue light (45 min light:15 min dark), this comparison was made against variable natural light, highlighting the immense potential of optimized light delivery rather than a direct comparison under controlled, identical average intensities [11]. Recent studies further confirm that ultra-short flashes of 50–100 μs can significantly accelerate the specific growth rates of microalgal cells compared to continuous illumination at an integrated identical intensity [20]. Furthermore, this effect is highly effective in immobilized cultures; for instance, high-frequency flashing light at 10 kHz achieved 7-fold net photosynthetic activity higher than continuous light in biofilm photobioreactors, effectively overcoming the severe light gradients typically found in dense cell layers [10].
Further illustrating the potential for innovative light delivery, a synergistic strategy was demonstrated in which supplementing high-intensity flashing light with a low-intensity continuous background light boosted Dunaliella salina biomass by up to 64% compared to continuous illumination at the same integrated intensity [7]. This principle of synergistic illumination extends from continuous background light to macro-scale temporal patterns. For instance, one study systematically compared the effects of a 1 Hz flash under either continuous light or a 12 h:12 h light-dark cycle across multiple taxa. The results revealed a potent synergistic effect, particularly under the diurnal cycle, where the dry cell weight of Acutodesmus obliquus increased by 5.16-fold compared to the cycle without flashes [21]. Similar enhancements were observed in Chlorella vulgaris (1.34-fold increase in dry cell weight under a light-dark cycle with flash), Uronema sp. (2.77-fold increase under continuous light with flash), and Micractinium reisseri (M. reisseri) (1.52-fold increase under continuous light with flash), underscoring that the efficacy of a flashing light strategy is highly contingent on the foundational lighting conditions.
2.2. Improved Photosynthetic Efficiency and Cell Growth Rate
At the physiological level, flashing light directly improves the efficiency of photon conversion into biomass, thus pulsed light with high photon flux densities can achieve a higher biomass yield per photon than continuous light of the same integrated flux [2], indicating an enhanced quantum yield. This is robustly demonstrated in Chlamydomonas reinhardtii, where high-frequency flashing light (e.g., 100 Hz) matched or exceeded photosynthetic efficiency and specific growth rates of microalgal cells under continuous light with equivalent time-averaged intensity [6].
2.3. Directed Metabolic Flux for High-Value Products
Beyond promoting cell growth, intermittent illumination serves as a powerful tool for metabolic engineering, steering carbon flux toward the synthesis of specific high-value compounds [22]. This strategy can effectively overcome the common trade-off between cell growth and secondary metabolite production. For example, a combination of diurnal cycles and superimposed flashing light induced a 3.31-fold increase of total fatty acid content in M. reisseri [21]. Similarly, the flashing light effect is recognized as a potent strategy to induce the simultaneous accumulation of both biomass and fucoxanthin [23]. Furthermore, longer-period intermittent light, such as a 12 h:12 h diurnal cycle, while mechanistically distinct from high-frequency flashes, has also been shown to improve light utilization efficiency and increase the production of proteins and essential amino acids in Dunaliella salina [16].
The strategic modulation of flash parameters can further serve as a “metabolic switch” to partition carbon into specific pools [24]. Specifically, while pulse durations around 10 μs favor the synthesis of high-value pigments such as astaxanthin and lutein, ultra-short flashes with a 10 μs light pulse followed by a 90 μs dark interval have been found to dramatically enhance the accumulation of PUFAs, including the essential omega-3 and omega-6 fatty acids [20].
The strategic application of flashing light as a “finishing step” in two-stage cultivation has proven highly effective for triggering the biosynthesis of high-value metabolites. Recent studies demonstrate that low-frequency regimes can dramatically reshape the biochemical profile of microalgae without the prolonged durations required by nutrient starvation [12,13]. For instance, applying low-frequency flashing light (5–50 Hz) at a 0.05 duty cycle with a high instantaneous intensity (6000 μmol photons·m−2·s−1) for four days significantly boosted the accumulation of proteins, pigments, and omega-3 fatty acids across multiple taxa. In Nannochloropsis gaditana, this regime increased the productivities of eicosapentaenoic acid (EPA) and specific carotenoids, such as violaxanthin, by up to 3-fold compared to continuous illumination [12]. The potency of this frequency-modulated induction was further highlighted in Diacronema lutheri and Tetraselmis striata. A three-day exposure to 5 Hz pulsing light (0.05 duty cycle, 300 μmol photons·m−2·s−1 as average intensity) resulted in a 4.6-fold increase in the productivities of fucoxanthin, diatoxanthin, EPA, and docosahexaenoic acid (DHA) in D. lutheri. Under identical 5 Hz parameters, T. striata exhibited a 1.3-fold enhancement in lutein, zeaxanthin, and EPA production [13]. These findings underscore that short-term (3–4 days) low-frequency light stimulation can serve as a robust metabolic trigger, effectively steering carbon flux toward complex lipids and xanthophylls in a strain-dependent manner. This demonstrates that flashing regimes can be tailored to induce specific biochemical pathways without the standard growth-productivity trade-off.
3. Core Molecular Mechanisms Underpinning the Biological Effects
Flashing/pulsed light optimizes photosynthesis across scales, and the superiority of the flashing effect is rooted in its profound remodeling of the core physiological processes in microalgal photosynthesis. The mechanisms involve a comprehensive optimization spanning from photosystem dynamics and the redox state of the electron transport chain to the spatiotemporal regulation of energy carriers (Figure 2).
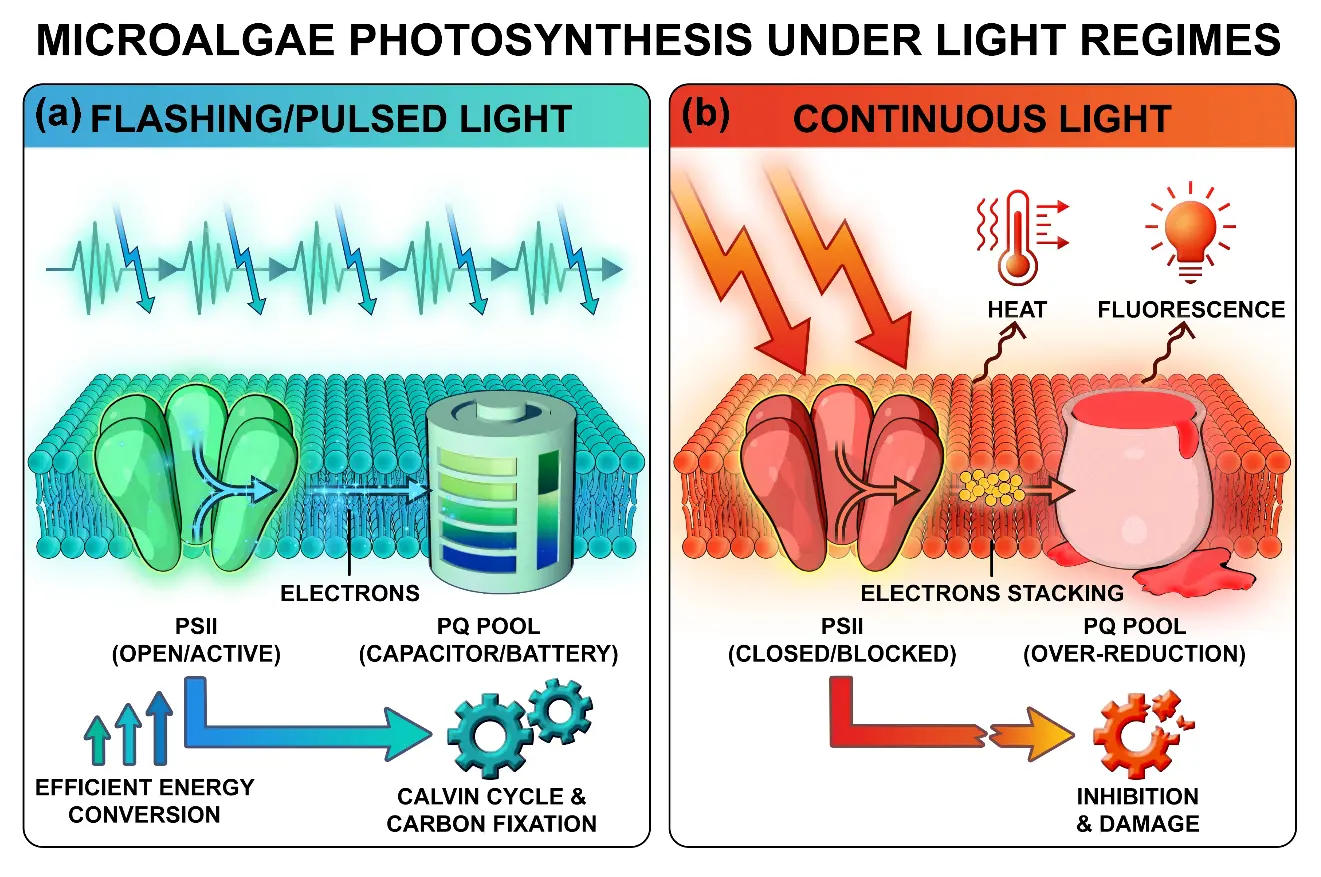
Figure 2. Conceptual illustration of the core molecular mechanisms differentiating flashing/pulsed light and continuous light regimes in microalgal photosynthesis. (a) Flashing/pulsed light: intermittent light pulses allow for brief dark intervals, enabling the re-oxidation of the photosynthetic electron transport chain. This keeps PSII reaction centers “open” and active. The plastoquinone (PQ) pool functions as an “electron capacitor”, storing electrons during light flashes and releasing them steadily during dark periods. This temporal decoupling leads to efficient energy conversion and drives the Calvin Cycle for carbon fixation. (b) Continuous high light: constant high-intensity illumination leads to the over-reduction of the PQ pool and causes PSII reaction centers to become “closed” or blocked. The resulting “electron traffic jam” forces excess energy to be dissipated as heat and fluorescence, leading to photoinhibition and cellular damage.
3.1. Dynamic Regulation of PSII
At the molecular level, under continuous high light, the primary electron acceptor of PSII, plastoquinone (QA), is rapidly reduced, causing a large fraction of reaction centers (RCs) to enter a “closed” state where they can no longer perform efficient charge separation [4]. This forces excess excitation energy to be dissipated as heat or fluorescence, leading to photoinhibition [4]. The microsecond-to-millisecond dark intervals provided by pulsed light offer a critical recovery window. During this time, QA is re-oxidized, allowing the closed RCs to “reopen” and regain their capacity for photochemical energy conversion [2,7]. This is thought to provide the necessary time for the rate-limiting step of the photosynthetic electron transport chain—the oxidation of the plastoquinone (PQ) pool—thereby ensuring that the entire electron flow remains fluent [14]. This is further supported by evidence that flashes as short as 50 μs combined with adequate dark intervals allow the photosynthetic machinery to bypass photo-inhibitory stress and the formation of reactive oxygen species typically induced by high-intensity continuous illumination [20]. Such microsecond-scale pulsing light may enable a more efficient reset of the PSII reaction centers, maintaining a higher fraction of “open” centers even under extreme peak intensities.
3.2. Energy Buffering by Plastoquinone (PQ) Pool
The PQ pool, acting as a mobile electron carrier connecting PSII to the cytochrome b6f complex, has been proposed as a conceptual electron buffer or “capacitor” due to its capacity to accept and store electrons, thereby buffering the electron transport chain against transient overload [25]. In microalgae, the PQ pool dynamics play an essential role in balancing electron flow under varying light conditions [26]. This analogy helps to illustrate how the PQ pool can transiently store electrons during light flashes and release them during dark periods. Under continuous highlight, the PQ pool remains in a deeply reduced state, exerting feedback inhibition on PSII and impeding linear electron flow. In a pulsed light regime, however, the large bolus of electrons generated during a light flash can be transiently stored in the oxidized PQ pool [27]. Subsequently, during the dark period, these stored electrons are steadily released to the downstream cytochrome b6f complex and ultimately utilized for carbon fixation [28]. This “peak shaving” effect on the electron flow not only prevents energy waste and photodamage during the intense light pulses but also achieves a more efficient and uniform distribution of light energy across the entire light-dark cycle. Thus, PQ pool stores/releases electrons in a pulsed light regime for efficient electron flow, under optimally tuned conditions, the fraction of open PSII reaction centers can approach unity, suggesting near-maximal photochemical efficiency [2].
3.3. Temporal Decoupling and Integration
High-frequency flashing light facilitates a partial temporal decoupling between the electron generation during light reactions and the carbon fixation processes of dark reactions. During these ultrashort, high-intensity light flashes, the instantaneous rate of linear electron transport can significantly surpass the maximum steady-state threshold observed under continuous illumination [9,29]. The resulting NADPH and ATP serve as transiently sequestered pools of reducing power and chemical energy. In the ensuing dark intervals, enzymatic processes such as carbon fixation continue by utilizing these accumulated energy carriers, is thought to achieve a temporal integration of light energy and enhancing the overall energy conversion efficiency [8]. Experimental data at high frequencies showed that net photosynthetic quantum yields can reach 0.161 at 10 kHz, 51 μmol photons·m−2·s−1 [10]. This value exceeds the traditionally accepted theoretical maximum of the Z-scheme under continuous illumination, although it should be noted that this comparison involves different illumination regimes. As noted by the original authors, this unusually high value was an observation requiring further investigation rather than definitive proof of a mechanism [10]. One interpretation is that high-frequency pulsing light may optimize the coordination between rapid electron generation and slower enzymatic CO2 fixation, but this remains a hypothesis that requires additional experimental validation. It is important to note that the temporal decoupling discussed here refers to the intrinsic kinetic mismatch between photochemical reactions (picosecond-to-millisecond) and enzymatic carbon fixation (millisecond-to-second) within the cell. This is conceptually distinct from hydrodynamically induced light-dark cycling at the reactor scale, which passively alternates cell exposure between illuminated and dark zones without altering the fundamental kinetics of the photosynthetic apparatus. The latter will be discussed in Section 5.3.
3.4. System-Level Photoprotection and Global Metabolic Regulation
In a pulsed light regime, appropriate dark intervals facilitate the activation and relaxation of essential photoprotective mechanisms, most notably non-photochemical quenching (NPQ), which effectively mitigates the risk of photo-oxidative damage. Beyond its role as an energy source, flashing light has been suggested to act as an environmental signal that may influence the cellular metabolic network. By modulating the levels of intracellular phytohormones, specifically gibberellic acid (GA3), the signaling effect has been proposed as a potential pathway to simultaneously enhance both microalgal growth and the production of high-value compounds [21]. However, this remains an emerging concept, and the precise molecular circuitry underlying phytohormone-mediated responses to pulsed light is not yet fully elucidated. Ultimately, the systematic adjustment of duty cycles and frequencies may contribute to a reorganization of carbon flux between primary biomass synthesis and secondary metabolism, offering a theoretical framework for photobioreactor design [24]. Further studies are needed to distinguish direct photosynthetic effects from indirect signaling-mediated regulation. This intermittent signaling dictates the distribution of energy between primary biomass synthesis and photoprotective secondary metabolism, providing a theoretical foundation for the rational design of advanced photobioreactors that synchronize artificial lighting with the intrinsic kinetic bottlenecks of specific microalgal species [24]. Thus, NPQ and phytohormone signaling may help balance cell growth and secondary metabolism under flashing light.
Despite these photoprotective and regulatory benefits, flashing/pulsed light regimes that deviate from species-specific optimal windows can trigger adverse effects, which must be acknowledged and mitigated. Improperly designed flashing/pulsed light can induce photoinhibition, and ROS burst when peak intensity overwhelms PSII electron transport or dark periods are insufficient for PQ pool re-oxidation [4,19]. Low-frequency regimes with long dark intervals may cause metabolic idling, while excessive frequencies (>10 kHz) offer diminishing returns [6,30]. Mitigation strategies include: (i) maintaining duty cycles below 0.5 and selecting species-specific optimal frequencies (100–1000 Hz for cell growth, 5–50 Hz for metabolites) [6,31,32]; (ii) employing two-stage cultivation to separate cell growth from stress-induced production [31,33]; and (iii) real-time chlorophyll fluorescence monitoring as an early-warning indicator of photoinhibition [32]. Recognizing these constraints is essential for robust industrial applications.
4. Critical Parameters and Optimization
The efficacy of the flashing light effect is not universal; rather, it is strictly governed by a suite of interdependent physical and engineering parameters. In fact, efficacy depends on frequency, duty cycle, and spectrum. The optimization of these factors is essential to synchronize artificial illumination with the intrinsic biological rhythms of the photosynthetic apparatus in microalgal cells.
4.1. Frequency
Current research typically categorizes pulsed light into low-frequency (≤50 Hz) and mid-to-high frequency (>50 Hz) regimes. Evidence suggests the existence of an “optimal frequency window” wherein photosynthetic performance and cell growth rates of microalgae are maximized; frequencies outside this threshold often prove suboptimal or may even exert inhibitory effects [2]. Crucially, the selection of pulse frequency can be strategically tailored to meet specific metabolic objectives or target product profiles. Low-frequency pulsing light (e.g., 5 Hz or 50 Hz) has emerged as a potent tool for inducing the accumulation of high-value secondary metabolites (see Section 2.3 for detailed examples). In brief, studies have reported significant increases in EPA, carotenoids, fucoxanthin, and DHA under low-frequency (5–50 Hz) regimes [12,13].
4.2. Duty Cycle
The duty cycle, defined as the fraction of the light period relative to the total cycle duration [19], functions alongside pulse frequency to dictate the dynamic illumination landscape experienced by microalgal cells [6]. While the duty cycle directly modulates the time-averaged photon flux density (PFD), the resulting biological response is characteristically non-linear. Under high-intensity flashing regimes, excessive pulse durations (i.e., high duty cycles) can lead to photosystem overload and subsequent photoinhibition, where surplus excitation energy is dissipated as heat, thereby diminishing net oxygen evolution and cell growth efficiency [19]. Conversely, an optimized low duty cycle (e.g., 0.05–0.2) paired with periodic dark intervals facilitates the recovery of the photosynthetic apparatus. This allows the NADPH and ATP generated during the light pulse to be fully utilized by downstream enzymatic reactions such as carbon fixation during the dark phase, significantly enhancing overall energy conversion efficiency [8,14]. In practical applications, the efficacy of the duty cycle is inextricably coupled with frequency. In low-frequency regimes (e.g., 1–10 Hz), productivity may decline even at identical duty cycles if the dark intervals are excessively long, leading to metabolic idling. In contrast, high-frequency pulsing (e.g., 100 Hz) combined with an appropriate duty cycle enables microalgae to effectively integrate light energy, as the organism perceives the fluctuating environment as quasi-continuous [8]. This synergy allows for photosynthetic efficiencies that match or even exceed those observed under continuous light of the same average intensity. For instance, at a duty cycle of 0.5, flashing light at 1 Hz and 10 Hz has been shown to decrease biomass productivity by an average of 10%, whereas 100 Hz pulsing increased biomass productivity by 35% compared to continuous illumination [8].
4.3. Spectral Specificity
Light quality is defined by the spectral distribution of photons, typically categorized by wavelength into blue (400–500 nm), green (500–600 nm), and red (600–700 nm) light [25]. Each spectral band interacts uniquely with the microalgal photosynthetic apparatus, influencing both the rate of electron transport and the activation of specific signaling pathways [25,34]. Light quality plays a pivotal role in modulating the physiological state of microalgae through the activation of specific photoreceptors and signaling pathways [35]. Among the visible spectrum, blue light is particularly influential. Comparative studies on diatoms with varying cell sizes have demonstrated that 100 Hz pulsed blue light can achieve biovolume yields identical to those of continuous blue light at the same peak intensities. Specifically, this pulsed regime yielded a specific growth rate (0.97 d−1), statistically indistinguishable from that under continuous illumination (1.01 d−1). This highlights its potential for more energy-efficient biomass production, as the identical yield was achieved with only 50% of the total photon dose at a 50% duty cycle, effectively doubling the energy conversion efficiency [35].
More sophisticated cultivation strategies involve the dynamic adjustment of pulse wavelengths to align with the varying metabolic requirements of different cell growth phases [36]. In beta-carotene production by Dunaliella salina, a novel strategy transitions from flashing regimes that combine red and blue light to exclusive blue light pulsing at the onset of the stationary phase. This wavelength-switching approach increased beta-carotene content to 10 mg·g−1, a productivity gain exceeding 300% compared to continuous flashing controls using mixed red and blue light [36].
The practical implementation of these high-efficiency pulsed illumination schemes increasingly relies on the integration of advanced optoelectronic technologies. For instance, the combination of concentrator photovoltaics (CPV) and high-performance LEDs allows for the conversion of solar energy into pulsed red light, which can then be delivered to indoor photobioreactors, a strategy that offers global feasibility for renewable-energy-based microalgal production [5]. Consequently, future research prioritizes the bio-optical design of photobioreactors, where the synergistic relationship between cell size and light quality selection serves as a core optimization parameter [35]. The optimization of flashing/pulsed light parameters is inherently species-specific, as different microalgae possess distinct photosynthetic apparatus and metabolic regulatory networks. To provide a practical reference for strain selection and reactor operation, we summarizes representative species-specific optimized regimes in Table 1 and their reported enhancements in biomass or high-value compound productivity, compiled from the studies cited above.
Table 1. Species-specific responses and optimized flashing/pulsed light parameters for enhanced biomass and high-value metabolite production in microalgae.
|
Species |
Target Product/Outcome |
Light Regime (Frequency, Duty Cycle, Intensity, Wavelength) |
Enhancement vs. Control |
Refs. |
|---|---|---|---|---|
|
Chlamydomonas reinhardtii |
Photosynthetic efficiency & growth |
100 Hz, 0.5 duty cycle, ~200 μmol·m−2·s−1, white |
Matched or exceeded continuous light |
[6] |
|
Dunaliella salina |
Biomass |
Continuous background + flashing (1 Hz, 0.5 duty cycle) |
+64% vs. continuous light at same avg. intensity |
[7] |
|
Nannochloropsis sp. |
Biomass |
Low-frequency blue LED (45 min light:15 min dark) |
61-fold vs. variable natural light |
[11] |
|
Chlorella sp. |
Biomass |
Hydrodynamic baffle-induced flashing (flat-plate PBR) |
+39.23% dry cell weight |
[11] |
|
Nannochloropsis gaditana |
EPA, violaxanthin |
Low-frequency (5–50 Hz), 0.05 duty cycle, 6000 μmol·m−2·s−1 (peak), 4 days |
2–3 fold increase in productivity |
[12] |
|
Diacronema lutheri |
Fucoxanthin, EPA, DHA |
5 Hz, 0.05 duty cycle, 300 μmol·m−2·s−1 (avg.), 3 days |
Up to 4.6-fold increase |
[13] |
|
Tetraselmis striata |
Lutein, zeaxanthin, EPA |
5 Hz, 0.05 duty cycle, 300 μmol·m−2·s−1 (avg.), 3 days |
1.3-fold enhancement |
[13] |
|
Acutodesmus obliquus |
Dry cell weight |
1 Hz flash superimposed on 12 h:12 h light-dark cycle |
5.16-fold vs. cycle without flash |
[19] |
|
M. reisseri |
Total fatty acids |
1 Hz flash + diurnal cycle |
3.31-fold increase |
[19] |
|
Dunaliella salina |
β-carotene |
Red-to-blue wavelength-switching flashing (stationary phase) |
>300% increase (to 10 mg·g−1) |
[32] |
|
Synechocystis sp. PCC 6803 |
Light utilization |
Four light regions (FLR) optimized by ANN |
76.36% accuracy improvement |
[37] |
Note: This table summarizes representative examples. Actual optimal parameters may vary with reactor design, culture density, and nutrient conditions. EPA: eicosapentaenoic acid; DHA: docosahexaenoic acid; PBR: photobioreactor; FLR: four light regions; ANN: artificial neural network.
4.4. Limitations, Negative Findings, and Context-Dependent Efficacy
Not all flashing regimes outperform continuous light. In Chlamydomonas reinhardtii, 1 Hz and 10 Hz (0.5 duty cycle) decreased biomass productivity by 10% versus continuous light, while 100 Hz increased it by 35% [8]. Some species show weaker responses due to differences in antenna size or PQ pool capacity [12,14]. Under high cell densities, self-shading can attenuate the flashing effect [36].
Poorly designed regimes can be detrimental. Peak intensities >6000 µmol·m−2·s−1 cause ROS burst and photoinhibition [4,12]; duty cycles >0.7 reduce dark periods below the time needed for PQ re-oxidation [19]; and ultra-low frequencies (≤0.1 Hz) lead to metabolic idling [8].
Comparing results across studies is challenging. Average intensity definitions differ [2,6]; reactor geometry and mixing introduce passive light-dark cycles [11,37]; culture density affects light penetration [36]; and species-specific apparatus leads to divergent responses [12,36]. Many studies lack key parameters such as mixing speed, light path length, cell density, or waveform [9,19]. Future studies should report pulse parameters (frequency, duty cycle, peak, and average intensity), waveform, reactor geometry, mixing, culture density, species, and growth stage to improve comparability.
5. Biotechnological Applications
5.1. Autotrophic Cultivation
In autotrophic systems of microalgae, pulsed illumination primarily aims to maximize biomass productivity and CO2 fixation rates. By optimizing light-dark cycles through hydrodynamic mixing or controlled LED pulsing, studies report up to a 50% increase in biomass productivity with CO2 fixation rates rising from 0.62 to 0.87 g CO2·L−1·d−1 [38]. The efficacy depends heavily on the interplay between frequency, duty cycle, and light intensity [8]. Furthermore, the optimized CO2 aeration with 0.8 mm bubbles enhances cell growth rates by 32.4%, using bubbles as scattering agents to modulate flashing frequency and light penetration [39].
5.2. Mixotrophic Cultivation
In mixotrophic cultivation, microalgae simultaneously assimilate inorganic carbon (CO2) via photosynthesis and organic carbon (e.g., glucose, acetate) via respiration, enabling higher biomass productivity than that of autotrophic growth while retaining light-driven biosynthesis [40]. However, the coexistence of photoautotrophic and heterotrophic metabolisms introduces inherent complexity that is not present in autotrophic systems, making the application of pulsed light particularly challenging yet potentially rewarding.
Interaction between photosynthesis and respiration. Under mixotrophic conditions, electrons derived from both photosystem II (PSII) and mitochondrial respiration converge on the plastoquinone (PQ) pool and the cytochrome b6f complex, creating competition for electron transport chain capacity [30]. Organic carbon assimilation supplies additional ATP and NADPH, potentially alleviating energy demand during dark intervals of pulsed light regimes. However, this dual supply can also trigger feedback inhibition on photosynthetic electron transport when the cellular energy charge remains high, a phenomenon known as photosynthetic down-regulation [41]. Pulsed light may mitigate this by providing dark intervals that allow PQ re-oxidation and dissipation of excess reducing equivalents, thereby restoring redox balance [31].
Influence on redox balance and carbon partitioning. The intermittent nature of pulsed light can steer carbon flux toward different endpoints. During light pulses, rapid electron generation produces NADPH and ATP; during dark intervals, these carriers are consumed by downstream processes. If dark intervals are too short, over-reduction occurs; if too long, metabolic idling reduces efficiency [8]. In mixotrophic systems, the presence of organic carbon complicates this balance: glucose tends to repress photosynthetic gene expression (akin to a “Crabtree effect”), whereas acetate integrates more readily with phototrophic metabolism [30,40]. Thus, the optimal pulsed light regime depends on the organic carbon source—glucose-based mixotrophy may require lower light intensities or lower duty cycles, while acetate requires sufficient illumination to maximize synergy [40].
Associated challenges. Beyond regulatory crosstalk, rapid biomass accumulation in mixotrophic cultures intensifies self-shading, thereby attenuating the impact of specific light regimes on individual cells [41]. Utilization of organic carbon also leads to higher oxygen demand and potential ROS production, which pulsed light must manage. Despite these challenges, pulsed light has shown promise in wastewater treatment applications, where flashing light at 1000 Hz (60% duty cycle) achieved 97% nitrate and 70% phosphate removal [33], and 100 Hz, 0.3 duty cycle increased biomass by 35.8% and NH4+-N removal by 44.2% in dark biogas slurry with 25.1% energy savings [32]. These examples demonstrate that, despite the complexity, optimized pulsed regimes can enhance both biomass and pollutant removal. Future studies should systematically map the interactions among organic substrate type, pulsed light parameters, and redox state to enable the rational design of energy-efficient mixotrophic processes.
5.3. Implications for Innovative Reactor Design
At the reactor scale, innovative photobioreactor (PBR) designs aim to induce high-frequency light-dark cycling through hydrodynamic control and integrated lighting. These extrinsic light-dark alternations are mechanistically distinct from the intrinsic temporal decoupling described in Section 3; they passively modulate the light exposure of cells as they circulate between illuminated and dark zones, thereby leveraging the molecular mechanisms of photosynthesis without altering the cellular machinery itself. Innovative photobioreactor (PBR) designs focus on inducing high-frequency light-dark cycling through hydrodynamic control and integrated lighting. Structural modifications, such as internal baffles or lantern-shaped draft tubes, accelerate cell movement between light and dark zones, boosting biomass dry weight by nearly 40% [17]. Other designs utilize internal fiber optics or wavy-bottom geometries to provide robust, high-frequency (3–10 Hz) stimulation [18,37]. Furthermore, advanced LED systems enable precise control over pulse parameters and wavelength-switching strategies (e.g., red-to-blue), serving as a “metabolic switch” to trigger secondary metabolite synthesis [6,8]. These innovations bridge fundamental photosynthetic mechanisms with scalable, energy-efficient industrial applications.
5.4. Emerging AI-Driven Adaptive Lighting Systems
Recent advances incorporate artificial intelligence (AI) and machine learning (ML) to optimize light regimes in real time. However, most current AI applications focus on general light optimization (e.g., continuous or diurnal regimes), while direct application to flashing/pulsed light control remains an emerging area. Nevertheless, the methodologies developed provide a foundation for future adaptive pulsed-light systems.
Established applications. AI models have been successfully applied to predict optimal light intensities, durations, and spectra under static conditions. For example, an ANN-GA model for Parachlorella kessleri optimized polyphenol production under white light intensity of 29 µE·m−2·s−1 and 12:12 light-dark cycles (R2 = 0.97, <5% error) [42]; an XGBoost model predicted cell growth in a tubular photobioreactor (R2 = 0.9997, +7.19% biomass) with real-time sensor control [43]; an ANN model optimized four light regions in Synechocystis sp. PCC 6803 (R2 = 0.97, 76.36% accuracy improvement) [44]. And LSTM/SVR models captured light history effects in the outdoor cultivation of Phaeodactylum tricornutum [45]. Reported advantages include biomass enhancement of 15–57% and a 20–43% increase in high-value compounds.
Future potential for pulsed-light control. Direct AI-driven optimization of flashing/pulsed light parameters (frequency, duty cycle, peak intensity) is still nascent. Reinforcement learning has been successfully applied for autonomous pH control in photobioreactors [46], and hybrid modeling with transfer learning has been validated for adapting predictive models to new microalgal strains [47]. These approaches could be extended to real-time regulation of flashing regimes, but several additional requirements must be met: (i) high-frequency sensors capable of capturing µs–ms scale photosynthetic responses; (ii) training datasets encompassing multiple frequencies, duty cycles, and peak intensities; (iii) low-latency control systems to adjust pulsing parameters at sub-second timescales; and (iv) integration of CFD models to account for hydrodynamically induced light-dark cycling. A pilot-level step toward such integration is the Algal Autobioreactor (AAB), which employs a real-time adaptive AI system (RIPAAP) for autonomous algal density and optical performance monitoring [48].
Technical bottlenecks and solutions. Several challenges remain as follows. Data scarcity and species variability limit model generalization across different strains and reactors [42,45]; sensor reliability and cost hinder real-time closed-loop control [43]; and light attenuation in high-density cultures persists even with adaptive lighting. Feasible solutions include reinforcement learning for autonomous control and hybrid modeling combined with transfer learning to overcome data limitations.
Although direct AI-adaptive flashing is still emerging, predictive models show strong promise for dynamic pulsing in high-density cultures [49]. Addressing the data and system requirements outlined above will be critical for transforming flashing light effects from laboratory demonstrations to intelligent, energy-efficient industrial processes.
5.5. Energy Consumption, Cost Bottlenecks, and Solutions
Although flashing/pulsed light offers biological benefits, its practical viability depends on net energy savings and production costs. Quantitative comparisons among light regimes show that 100 Hz square flashing blue light achieves identical biomass yield as continuous blue light at the same peak intensity, effectively halving energy input (50% saving) [35]. For Mychonastes homosphaera, cool-white LED consumed the least energy (228.6 W·h·day−1) to produce maximum biomass (512.0 mg·L−1·day−1) [50].
Regarding unit biomass energy metrics, LED-assisted systems increased biomass yields by 24–34% over natural light controls, but the minimum biomass selling price was higher (80.76–91.37 USD·kg−1 vs. 68.85 USD·kg−1 for control) [51]. The electrical-to-chemical energy conversion efficiency of LEDs is approximately 6–8% [52]. For flashing light specifically, a 100 Hz, 0.3 duty cycle regime reduced light energy consumption by 25.1% compared to continuous light at the same biomass output [32].
Several solutions can address cost/energy bottlenecks. Photovoltaic integration reduces LED operational costs by up to 16.9% [51]. CFD-guided reactor optimization enhances light/dark cycling frequency and reduces energy use [31]. Species selection based on cell optical properties—large diatoms have 1.75× lower absorbance than small cells—improves light penetration and reduces the need for artificial lighting [35]. Model-based optimization balances productivity against energy input [53], and low-cost Arduino-based automation lowers economic barriers for smaller applications [52]. Collectively, these strategies indicate that optimized flashing light systems can narrow the economic gap toward commercial viability.
6. Summary and Future Perspectives
Flashing/pulsed light provides a transformative means of manipulating the spatiotemporal distribution of photons to optimize photosynthesis at the molecular level. Through mitigating photoinhibition, exploiting the PQ pool as an electronic buffer, and facilitating the temporal decoupling and synergy of light-dark reactions, these strategies have enabled breakthrough improvements in light energy utilization efficiency. Consequently, the potential to enhance biomass productivity in cultivation systems and to steer the targeted accumulation of high-value metabolites is now well established, offering a robust foundation for the sustainable production of microalgal biomass. The current development in AI-adaptive systems signal a shift toward real-time, sensor-driven control for optimized flashing regimes.
Looking forward, we anticipate that metabolic reprogramming and artificial intelligence will drive the next wave of innovation in flashing/pulsed light systems. Preliminary evidence suggests that increased light intensity under optimized pulsed regimes can enhance cell growth and metabolite production [20], and wavelength switching has begun to function as a metabolic switch [24]. AI models are increasingly being applied to optimize light regimes for microalgal growth [49], and pilot projects have demonstrated closed-loop systems incorporating AI-coupled adaptive flashing with real-time sensor control and hybrid CFD-AI predictive models, pointing toward intelligent, autonomous illumination systems. These emerging trends, while still in early stages, are expected to mature in the coming years and are discussed in more detail in Section 5.4.
Future priorities in research and industrial applications mainly focus on three strategic frontiers: (1) elucidating the specific molecular mechanisms and regulatory networks underlying cell responses to pulsed light across diverse taxa and cultivation modes, with a particular emphasis on the complex metabolic crosstalk in mixotrophic systems; (2) developing intelligent, adaptive lighting systems that integrate advanced sensing and real-time control for scaling-up; and (3) achieving rational reactor design and hybrid computational fluid dynamics (CFD)-AI integration to enable intelligent, sustainable commercialization. Ultimately, the successful applications of these lab-scale “flashing effects” into industrial reality will depend on the interdisciplinary convergence of photonics, physiology, bioprocess engineering, and auto-control technology.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the authors used DeepSeek-V4 to assist with preliminary language polishing. The authors have thoroughly reviewed and revised all content generated or suggested by the tool and assume full responsibility for the accuracy, originality, and scientific integrity of the published work.
Author Contributions
B.W.: Writing—Original Draft, Literature Search/Data Curation; D.W.: Writing—Review & Editing, Supervision and Funding Acquisition.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during the current study are available from the corresponding author on reasonable request.
Funding
This work was supported by National Key Research and Development Program for Food Nutrition and Safety (2023YFF1104100), and the Key Project of Guangzhou Research & Development Program (2025B03J0093).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Abu-Ghosh S, Fixler D, Dubinsky Z, Iluz D. Flashing light in microalgae biotechnology. Bioresour. Technol. 2016, 203, 357–363. DOI:10.1016/j.biortech.2015.12.057 [Google Scholar]
- Zarmi Y. High-intensity pulsed-light cultivation of unicellular algae: Photosynthesis continues in the dark. Heliyon 2024, 10, e27224. DOI:10.1016/j.heliyon.2024.e27224 [Google Scholar]
- Schmid R, Dring MJ. Rapid, Blue-Light-Induced Acidifications at the Surface of Ectocarpus and Other Marine Macroalgae. Plant Physiol. 1993, 101, 907–913. DOI:10.1104/pp.101.3.907 [Google Scholar]
- Han BP. A Mechanistic Model of Algal Photoinhibition Induced by Photodamage to Photosystem-II. J. Theor. Biol. 2002, 214, 519–527. DOI:10.1006/jtbi.2001.2468 [Google Scholar]
- Gordon JM, Polle JEW. Ultrahigh bioproductivity from algae. Appl. Microbiol. Biotechnol. 2007, 76, 969–975. DOI:10.1007/s00253-007-1102-x [Google Scholar]
- Vejrazka C, Janssen M, Streefland M, Wijffels RH. Photosynthetic efficiency of Chlamydomonas reinhardtii in flashing light. Biotech Bioeng. 2011, 108, 2905–2913. DOI:10.1002/bit.23270 [Google Scholar]
- Abu-Ghosh S, Fixler D, Dubinsky Z, Iluz D. Continuous background light significantly increases flashing-light enhancement of photosynthesis and growth of microalgae. Bioresour. Technol. 2015, 187, 144–148. DOI:10.1016/j.biortech.2015.03.119 [Google Scholar]
- Vejrazka C, Janssen M, Streefland M, Wijffels RH. Photosynthetic efficiency of Chlamydomonas reinhardtii in attenuated, flashing light. Biotechnol. Bioeng. 2012, 109, 2567–2574. DOI:10.1002/bit.24525 [Google Scholar]
- Schulze PSC, Guerra R, Pereira H, Schüler LM, Varela JCS. Flashing LEDs for Microalgal Production. Trends Biotechnol. 2017, 35, 1088–1101. DOI:10.1016/j.tibtech.2017.07.011 [Google Scholar]
- Martín-Girela I, Curt MD, Fernández J. Flashing light effects on CO2 absorption by microalgae grown on a biofilm photobioreactor. Algal Res. 2017, 25, 421–430. DOI:10.1016/j.algal.2017.06.008 [Google Scholar]
- Yustinadiar N, Manurung R, Suantika G. Enhanced biomass productivity of microalgae Nannochloropsis sp. in an airlift photobioreactor using low-frequency flashing light with blue LED. Bioresour. Bioprocess. 2020, 7, 43. DOI:10.1186/s40643-020-00331-9 [Google Scholar]
- Lima S, Schulze PSC, Schüler LM, Rautenberger R, Morales-Sánchez D, Santos TF, et al. Flashing light emitting diodes (LEDs) induce proteins, polyunsaturated fatty acids and pigments in three microalgae. J. Biotechnol. 2021, 325, 15–24. DOI:10.1016/j.jbiotec.2020.11.019 [Google Scholar]
- Schüler LM, Walter JM, Kato H, Suzuki H, Hulatt CJ, Rautenberger R, et al. High-value compound induction by flashing light in Diacronema lutheri and Tetraselmis striata CTP4. Bioresour. Technol. Rep. 2022, 19, 101158. DOI:10.1016/j.biteb.2022.101158 [Google Scholar]
- Fierro Ulloa JI, Lu LD, Bernard O. Theoretical growth rate of microalgae under high/low-flashing light. J. Math. Biol. 2023, 86, 48. DOI:10.1007/s00285-023-01871-2 [Google Scholar]
- Saccardo A, Wolf J, Bezzo F, Hankamer B. Modelling dynamics of photosynthetic units and microalgae growth based on high throughput pulsed light screens. Chem. Eng. J. 2024, 490, 151684. DOI:10.1016/j.cej.2024.151684 [Google Scholar]
- Sui Y, Muys M, Vermeir P, D’Adamo S, Vlaeminck SE. Light regime and growth phase affect the microalgal production of protein quantity and quality with Dunaliella salina. Bioresour. Technol. 2019, 275, 145–152. DOI:10.1016/j.biortech.2018.12.046 [Google Scholar]
- Zhang Q, Yan C, Xue S, Wu X, Wang Z, Cong W. A novel flat plate photobioreactor for microalgae cultivation. Chin. J. Biotechnol. 2015, 31, 251–257. Available online: https://europepmc.org/article/med/26062346 (accessed on 1 March 2026).
- Moroni M, Lorino S, Cicci A, Bravi M. Design and Bench-Scale Hydrodynamic Testing of Thin-Layer Wavy Photobioreactors. Water 2019, 11, 1521. DOI:10.3390/w11071521 [Google Scholar]
- Vejrazka C, Janssen M, Benvenuti G, Streefland M, Wijffels RH. Photosynthetic efficiency and oxygen evolution of Chlamydomonas reinhardtii under continuous and flashing light. Appl. Microbiol. Biotechnol. 2013, 97, 1523–1532. DOI:10.1007/s00253-012-4390-8 [Google Scholar]
- Shankar U, Lenka SK, Ackland ML, Callahan DL. Flashing white light stimulates enhanced growth, pigment and fatty acid accumulation in microalgae under high-intensity illumination cycles. Algal Res. 2025, 91, 104224. DOI:10.1016/j.algal.2025.104224 [Google Scholar]
- Choi Y, Kim H, Kumaran RS, Song H, Song K, Kim KJ, et al. Enhanced growth and total fatty acid production of microalgae under various lighting conditions induced by flashing light. Eng. Life Sci. 2017, 17, 976–980. DOI:10.1002/elsc.201700001 [Google Scholar]
- Morelli L, Patwari P, Pruckner F, Bastide M, Fabris M. Specific light-regime adaptations, terpenoid profiles and engineering potential in ecologically diverse Phaeodactylum tricornutum strains. Algal Res. 2025, 86, 103920. DOI:10.1016/j.algal.2025.103920 [Google Scholar]
- Wang S, Wu S, Yang G, Pan K, Wang L, Hu Z. A review on the progress, challenges and prospects in commercializing microalgal fucoxanthin. Biotechnol. Adv. 2021, 53, 107865. DOI:10.1016/j.biotechadv.2021.107865 [Google Scholar]
- Marchese A, Lima S, Cosenza A, Giambalvo F, Scargiali F. Effects of Light Quality Adjustment in Microalgal Cultivation: Flashing Light and Wavelength Shifts in Photobioreactor Design. Processes 2025, 13, 1159. DOI:10.3390/pr13041159 [Google Scholar]
- Shankar U, Lenka SK, Ackland ML, Callahan DL. Review of the structures and functions of algal photoreceptors to optimize bioproduct production with novel bioreactor designs for strain improvement. Biotechnol. Bioeng. 2022, 119, 2031–2045. DOI:10.1002/bit.28116 [Google Scholar]
- Nikkanen L, Solymosi D, Jokel M, Allahverdiyeva Y. Regulatory electron transport pathways of photosynthesis in cyanobacteria and microalgae: Recent advances and biotechnological prospects. Physiol. Plant 2021, 173, 514–525. DOI:10.1111/ppl.13404 [Google Scholar]
- Deák Z, Sass L, Kiss É, Vass I. Characterization of wave phenomena in the relaxation of flash-induced chlorophyll fluorescence yield in cyanobacteria. Biochim. et Biophys. Acta (BBA)—Bioenerg. 2014, 1837, 1522–1532. DOI:10.1016/j.bbabio.2014.01.003 [Google Scholar]
- Weng JH, Shieh YJ. Salicylhydroxamic acid (SHAM) inhibits O2 photoreduction which protects nitrogenase activity in the cyanobacterium Synechococcus sp. RF-1. Photosynth. Res. 2004, 82, 151–164. DOI:10.1007/s11120-004-9369-6 [Google Scholar]
- Abu-Ghosh S, Fixler D, Dubinsky Z, Solovchenko A, Zigman M, Yehoshua Y, et al. Flashing light enhancement of photosynthesis and growth occurs when photochemistry and photoprotection are balanced in Dunaliella salina. Eur. J. Phycol. 2015, 50, 469–480. DOI:10.1080/09670262.2015.1069404 [Google Scholar]
- Alcántara C, Fernández C, García-Encina PA, Muñoz R. Mixotrophic metabolism of Chlorella sorokiniana and algal-bacterial consortia under extended dark-light periods and nutrient starvation. Appl. Microbiol. Biotechnol. 2015, 99, 2393–2404. DOI:10.1007/s00253-014-6125-5 [Google Scholar]
- Li T, Kirchhoff H, Gargouri M, Feng J, Cousins AB, Pienkos PT, et al. Assessment of photosynthesis regulation in mixotrophically cultured microalga Chlorella sorokiniana. Algal Res. 2016, 19, 30–38. DOI:10.1016/j.algal.2016.07.012 [Google Scholar]
- Chen C, Tao F, Han T, Gao F, Dong T, Jiang W, et al. Photoacclimation caused by high frequency flashing light assists Chlorella sp. M-12 wastewater treatment and biomass accumulation in dark color biogas slurry. J. Appl. Phycol. 2022, 34, 2929–2940. DOI:10.1007/s10811-022-02840-w [Google Scholar]
- Keramati A, Azizi S, Hashemi A, Pajoum Shariati F. Effects of flashing light–emitting diodes (LEDs) on membrane fouling in a reciprocal membrane photobioreactor (RMPBR) to assess nitrate and phosphate removal from whey wastewater. J. Appl. Phycol. 2021, 33, 1513–1524. DOI:10.1007/s10811-021-02388-1 [Google Scholar]
- Hallmann A. Sensing a rainbow of colors: Algal photoreceptors. Front. Plant Sci. 2025, 16, 1684559. DOI:10.3389/fpls.2025.1684559 [Google Scholar]
- Eilertsen HC, Strømholt J, Bergum JS, Eriksen GK, Ingebrigtsen R. Mass Cultivation of Microalgae: II. A Large Species Pulsing Blue Light Concept. BioTech 2023, 12, 40. DOI:10.3390/biotech12020040 [Google Scholar]
- Mirzaie MAM, Lak ZJ, Meybodi FH. A New Shifting Strategy in Flashing Light Based on the Cell Growth Curve to Enhance β-carotene Biosynthesis in Dunaliella salina. Curr. Microbiol. 2021, 78, 2733–2740. DOI:10.1007/s00284-021-02542-5 [Google Scholar]
- Olivieri G, Moroni M, Janssen M, Piersanti L, Mezza D, Bravi M. Model-Based Prediction of Perceived Light Flashing in Recirculated Inclined Wavy-Bottomed Photobioreactors. Processes 2021, 9, 1158. DOI:10.3390/pr9071158 [Google Scholar]
- Ye Q, Cheng J, Guo W, Xu J, Li K, Zhou J. Serial lantern-shaped draft tube enhanced flashing light effect for improving CO2 fixation with microalgae in a gas-lift circumflux column photobioreactor. Bioresour. Technol. 2018, 255, 156–162. DOI:10.1016/j.biortech.2018.01.127 [Google Scholar]
- Song Y, Cheng J, Yang Y, Gao Z. Irradiance penetration distribution and flashing light frequency simultaneously affected with microalgal cell absorption and CO2 bubble scattering in a raceway pond. Sci. Total Environ. 2023, 864, 160988. DOI:10.1016/j.scitotenv.2022.160988 [Google Scholar]
- Patel AK, Joun JM, Hong ME, Sim SJ. Effect of light conditions on mixotrophic cultivation of green microalgae. Bioresour. Technol. 2019, 282, 245–253. DOI:10.1016/j.biortech.2019.03.024 [Google Scholar]
- Arora N, Philippidis GP. Unraveling metabolic alterations in Chlorella vulgaris cultivated on renewable sugars using time resolved multi-omics. Sci. Total Environ. 2021, 800, 149504. DOI:10.1016/j.scitotenv.2021.149504 [Google Scholar]
- Fernández Izquierdo P, Patiño Coral M, Ortiz Benavides F. Application of an artificial neural network coupled to a genetic algorithm for the production of polyphenols in Parachlorella kessleri grown under mixotrophic conditions. Algal Res. 2024, 77, 103331. DOI:10.1016/j.algal.2023.103331 [Google Scholar]
- Tummawai T, Rohitatisha Srinophakun T, Padungthon S, Sukpancharoen S. Application of Artificial Intelligence and Image Processing for the Cultivation of Chlorella sp. Using Tubular Photobioreactors. ACS Omega 2024, 9, 46017–46029. DOI:10.1021/acsomega.4c05971 [Google Scholar]
- Yu T, Fan F, Huang L, Wang W, Wan M, Li Y. Artificial neural networks prediction and optimization based on four light regions for light utilization from Synechocystis sp. PCC 6803. Bioresour. Technol. 2024, 394, 130166. DOI:10.1016/j.biortech.2023.130166 [Google Scholar]
- Yeh YC, Syed T, Brinitzer G, Frick K, Schmid-Staiger U, Haasdonk B, et al. Improving microalgae growth modeling of outdoor cultivation with light history data using machine learning models: A comparative study. Bioresour. Technol. 2023, 390, 129882. DOI:10.1016/j.biortech.2023.129882 [Google Scholar]
- Caparroz M, Guzmán JL, Berenguel M, Acién FG. A novel data-driven model for prediction and adaptive control of pH in raceway reactor for microalgae cultivation. New Biotechnol. 2024, 82, 1–13. DOI:10.1016/j.nbt.2024.04.001 [Google Scholar]
- Riezzo L, Kay H, Feng Y, Jing K, Zhang D. Accelerating bioprocess digital twin development by integrating hybrid modelling with transfer learning. Chem. Eng. J. 2025, 511, 162018. DOI:10.1016/j.cej.2025.162018 [Google Scholar]
- Azimi M, Mahdavinejad M, Yeganeh M, Zamani Z. Real-time adaptive algae bioreactor facades: A novel method for dynamic daylight and thermal efficiency optimization in buildings. Energy Convers. Manag. 2025, 346, 120409. DOI:10.1016/j.enconman.2025.120409 [Google Scholar]
- Wu Y, Shan L, Zhao W, Lu X. Harnessing Artificial Intelligence to Revolutionize Microalgae Biotechnology: Unlocking Sustainable Solutions for High-Value Compounds and Carbon Neutrality. Mar. Drugs 2025, 23, 184. DOI:10.3390/md23050184 [Google Scholar]
- Borah D, Eldiehy KSH, AL-Hothaly KA, Deka D. A Novel Approach Towards RSM-Based Optimization of LED-Illuminated Mychonastes homosphaera Culture, Emphasizing Input Energy: An Industrial Perspective of Microalgae Cultivation. Phycology 2025, 5, 62. DOI:10.3390/phycology5040062 [Google Scholar]
- Magalhães IB, Jesus Júnior MMD, França BT, Silva TA, Pereira ASAP, Souza LCDA, et al. Shining a light on outdoor algal systems for wastewater treatment: How artificial light enhancement impacts biomass costs and life cycle. J. Environ. Manag. 2025, 391, 126510. DOI:10.1016/j.jenvman.2025.126510 [Google Scholar]
- Singh V, Mishra V. A review on the current application of light-emitting diodes for microalgae cultivation and its fiscal analysis. Crit. Rev. Biotechnol. 2023, 43, 665–679. DOI:10.1080/07388551.2022.2057274 [Google Scholar]
- Saccardo A, Porcelli A, Borella L, Sforza E, Bezzo F. Model-based optimisation of microalgae growth under high-intensity and high-frequency pulsed light conditions. J. Clean. Prod. 2024, 469, 143238. DOI:10.1016/j.jclepro.2024.143238 [Google Scholar]