Central Metabolism-Responsive Biosensors for Monitoring, Screening, and Engineering in Microbial Production

Central Metabolism-Responsive Biosensors for Monitoring, Screening, and Engineering in Microbial Production
Jianli Zhang 1 Xinyu Gong 1 Qi Gan 1 Yingyue Yu 1 Yuan Dou 1 Renjie Shang 2 Nicolas Lopez 1 Yajun Yan 1,*
Received: 13 April 2026 Revised: 28 April 2026 Accepted: 26 May 2026 Published: 10 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
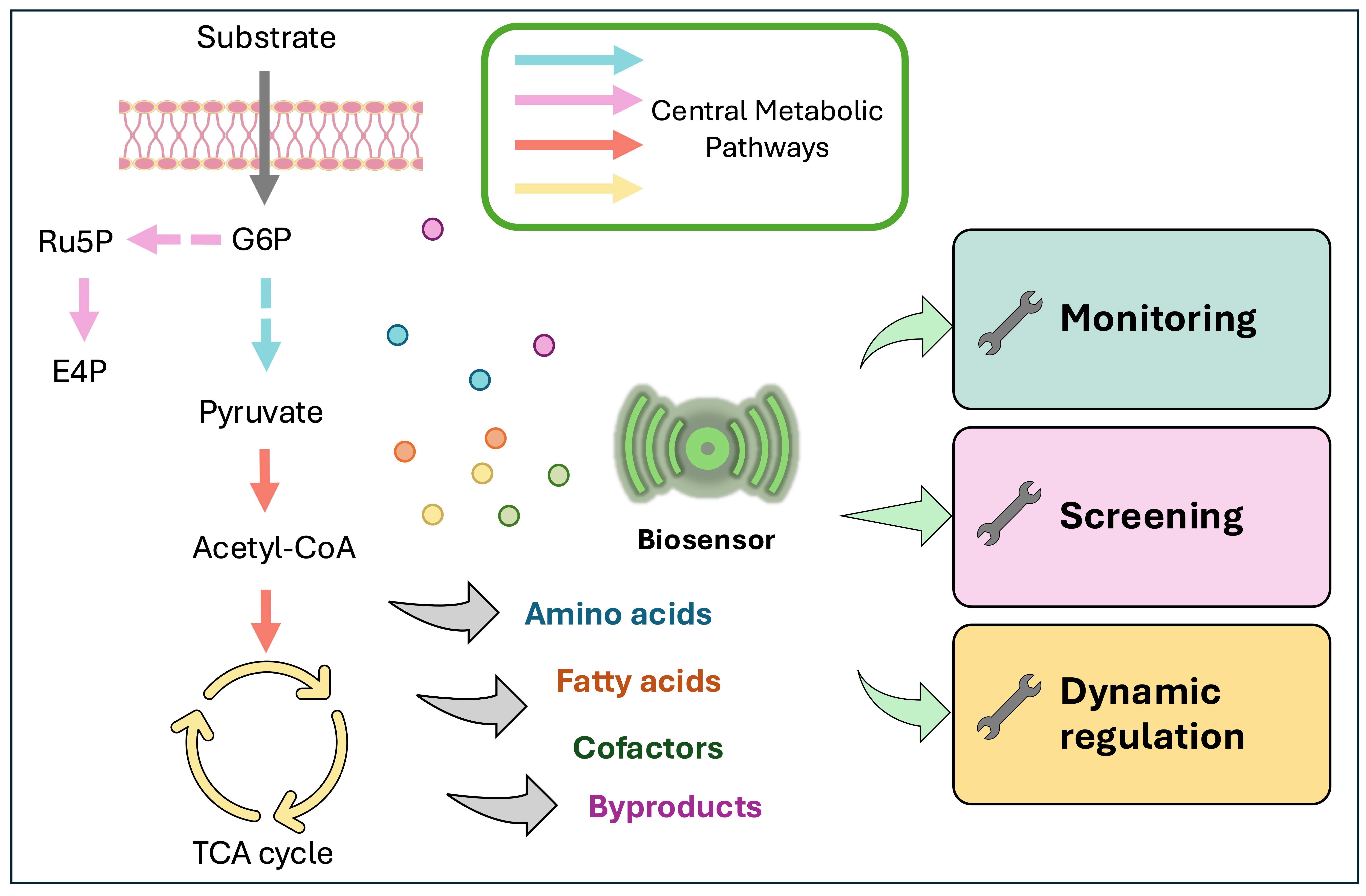
Graphical Abstract
1. Introduction
Central metabolic pathways such as glycolysis, tricarboxylic acid (TCA) cycle, and pentose phosphate (PP) pathway play pivotal roles and are highly conserved across diverse organisms. Those pathways coordinate the breakdown of carbon sources, generate energy, reduce power, and key intermediates for the synthesis of amino acids, nucleotides, fatty acids, and other cellular components to support basic cell growth, maintenance, and biosynthesis. As the central metabolic pathways are the primary routes of metabolizing the nutrients, and shunt the carbon flux to hundreds of branches, a complicated regulation network is necessary to harmonize the operating of central metabolism by responding to the nutrient availability, cellular redox status, and other environmental factors to adjust the gene expression and carbon flux in time. Transcription factor (TF) constitutes one of the most important regulation network to control the cellular pathways and metabolism. And numerous global and local TFs related to central metabolism regulation have been identified and revealed [1]. The global TFs usually detect key metabolites that are intricately linked to multiple pathways. For instance, cyclic AMP (cAMP) receptor protein (CRP) serves as a global TF in Escherichia coli, responding to cellular cAMP levels and controlling more than 180 genes [2]. ArcAB is a two-component signal transduction system that responds to respiratory conditions and regulates numerous genes related to respiratory and fermentative metabolism [3]. There are also local TF which regulate specific genes or pathways. The well-studied TFs LacI, PurR, and trpR control lactose metabolism, purine biosynthesis genes, and tryptophan biosynthesis by responding to lactose, purine nucleotides, and tryptophan respectively [4,5,6].
In the field of microbial production, the accumulation of high-value products often relies on the efficient conversion of carbon sources and other substrates. Therefore, understanding the central pathways and their regulation networks is a critical topic. With the advancement of metabolic engineering and molecular biology technologies, substantial progress has been made in modulating central metabolism to overcome key bottlenecks, such as enhancing carbon efficiency, alleviating metabolic burden, relieving cofactor shortages, and balancing competing pathways [7]. The widely employed metabolic engineering strategies for improving the biosynthetic performance include targeted gene knockouts and knock-ins, expression of high-activity heterologous enzymes, and improvement of fermentation parameters. Nevertheless, the metabolism state of microbial cell factories undergoes continuous fluctuations driven by environmental perturbations, life cycle transitions, and shifts in production phases. Consequently, static regulatory strategies are usually inadequate for real-time metabolic monitoring and timely response, resulting in suboptimal bioproduction titer.
To enable real-time metabolic sensing and achieve gene regulation in response to intracellular signals, diverse TFs have been reconstructed into biosensors and effectively employed to boost numerous synthetic pathways and support a variety of synthetic biology applications [8]. Depending on the regulatory functions of the TFs, biosensor can be tailored to respond to a wide range of targets, including metabolic intermediates, end products, byproducts, cell density and even environmental factors such as temperature, light, and pH. By responding to these fluctuations, biosensors can mediate the activation or repression of the transcription of downstream genes. When biosensors are designed to respond to pathway-specific compounds, they can be used to address pathway targeted challenges. For example, the p-coumaric acid biosensor PadR and naringenin biosensor FdeR have been adopted to activate naringenin biosynthetic enzymes and enhance its production [9]. The resveratrol biosensor engineered from TtgR was used to screen the p-coumarate: CoA ligase with improved activity for enhancing production of resveratrol and naringenin [10]. And the isoprene biosensor engineered from TbuT has been applied to screen the isoprene high producer [11]. When biosensors are engineered to detect products from central metabolic pathways, such as pyruvate, or key metabolites like amino acids, adenosine-5′-triphosphate (ATP), and NAD(P), they will possess broader applicability and can be leveraged to address metabolic issues across diverse biosynthetic situations.
In this review, we introduce central metabolism and its key metabolites from the perspective of microbial production, followed by a concise overview of the concept of biosensors. We also discuss recent advances in biosensor development and application, with a particular focus on studies of biosensors responsive to central metabolites (Figure 1). In addition, we explore the integration of biosensors with other rapidly advancing technologies, such as omics, metabolic modeling, and machine learning. We will also discuss the challenges and bottlenecks in biosensor development, along with potential avenues for future advancement.
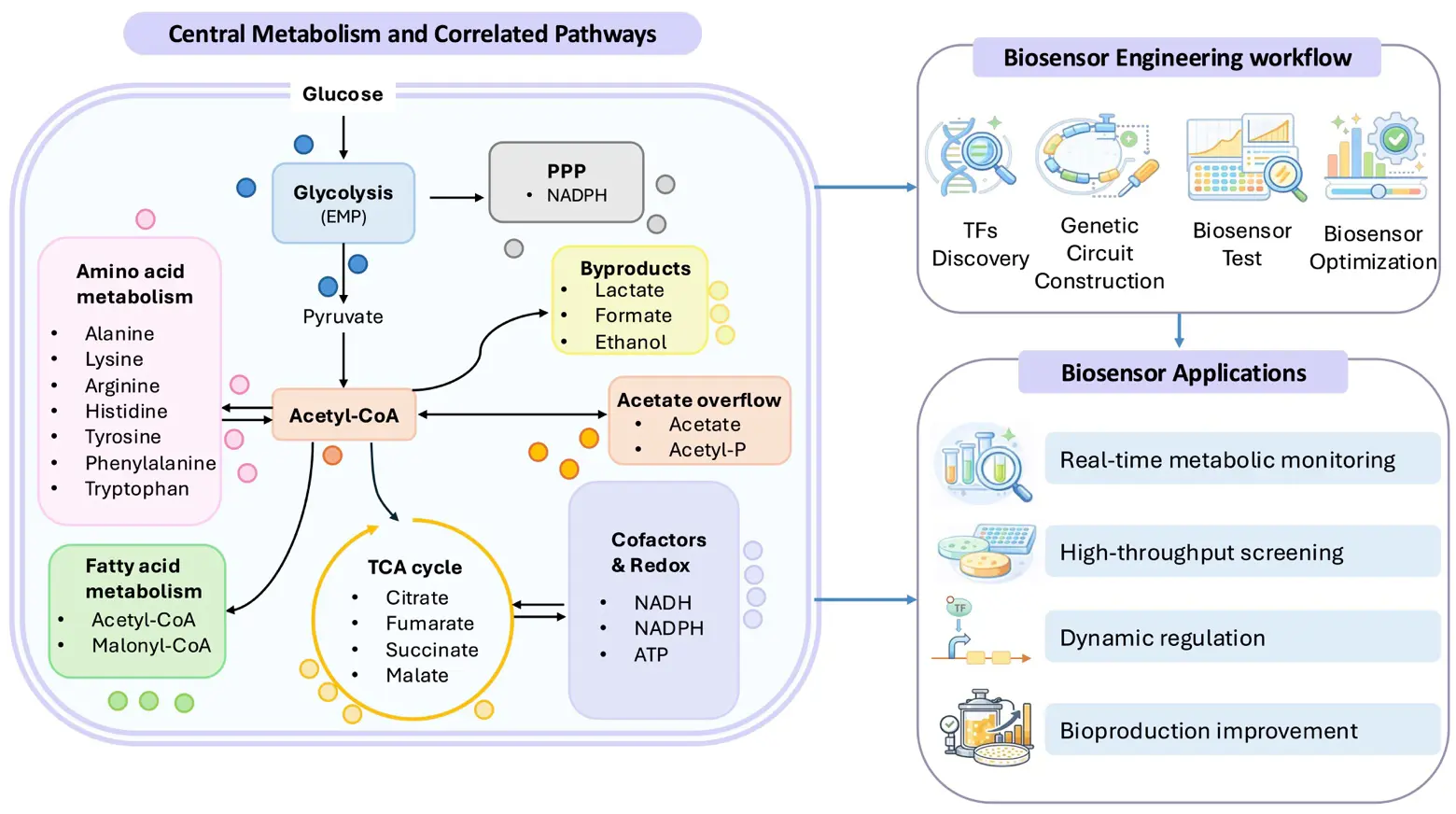
Figure 1. The schematic of central metabolism, the biosensor engineering workflow, and the applications of central metabolite-responsive biosensors. The colored dots in the “Central Metabolism and Correlated Pathway” refer to metabolite molecules that could be sensed by the biosensors. TF is the abbreviation of transcription factor.
2. Overview of Central Metabolism and Associated Pathways
The central metabolic pathway serves as the backbone of the cellular reactions, integrating nutrient uptake with energy generation and precursor supply. In the scenario of microbial production, it plays a pivotal role in determining both cell growth and biosynthetic efficiency. The major central pathways include glycolysis, the TCA cycle, and the PP pathway. They coordinate the transformation of a carbon source into ATP, reducing equivalents, and key intermediates such as pyruvate and acetyl-CoA.
Glycolysis channels sugar into pyruvate through a series of enzymatic reactions, acting as a primary entry point for carbon flux. The substrate-level phosphorylation steps mediated by phosphoglycerate kinase and pyruvate kinase are companied with a net gain of ATP. Glycolysis also produces NADH and several branch intermediates such as 3-phosphoglycerate and phosphoenolpyruvate (PEP). Pyruvate, the final product of glycolysis, serves as a central node linking glycolysis to downstream pathways. The TCA cycle is initiated by condensing oxaloacetate (OAA) and the pyruvate-derived acetyl-CoA. Through several steps, TCA cycle generates NADH, FADH2, GTP and key intermediates such as 2-ketoglutarate (2-KG). The PP pathway contributes a bunch of reducing power in the form of NADPH and provides ribose-5-phosphate (Ru5P) for nucleotide synthesis. The oxidative branch of the PP pathway, driving by glucose-6-phosphate dehydrogenase, is especially important for maintaining redox balance. The electron transport chain utilizes reducing equivalents from upstream metabolism to drive oxidative phosphorylation and sustain cellular energy demand. Together, these pathways regulate the cellular balance between catabolism and anabolism.
Beyond the core pathways, various organic acid reactions also act as critical indicators that reflect the cellular metabolic states [12]. Although not considered part of the central pathway per se, the biosynthesis of organic acids and amino acids is closely tied to central metabolic flux through shared precursors and cofactors. Organic acids such as acetate, lactate, and succinate are typically derived from glycolysis or TCA cycle intermediates and are key products or byproducts in bioproduction. Similarly, amino acids such as glutamate and aspartate are derived from 2-KG and OAA in the TCA cycle; alanine and serine are derived from pyruvate and 3-phosphoglycerate in glycolysis; histidine is derived from the PP pathway, and the three aromatic amino acids, tryptophan, phenylalanine and tyrosine are derived from both PEP and Erythrose-4-phosphate (E4P). These biosynthetic pathways are functionally integrated with central pathways and provide key signals, byproducts, and building blocks for basic cell growth.
Given the significant role of central metabolism in microbial production, many classical metabolic engineering strategies have been applied to modify central metabolism. These include knocking out competing pathways to conserve carbon flux, overexpressing rate-limiting enzymes to enhance key reactions, engineering enzymes to improve catalytic efficiency, and optimizing fermentation conditions and processes to increase overall yield. Although promising, these approaches can present significant challenges under certain conditions. For instance, deleting genes within central metabolism may lead to metabolic imbalances and disrupted carbon flux, ultimately impairing cell growth and reducing product yields [13]. Similarly, overexpressing rate-limiting enzymes can impose an excessive metabolic burden, hinder cell growth, and waste cellular resources [9]. As cellular metabolism is inherently dynamic, such static control strategies often fail to adjust regulatory strength in response to real-time changes in cellular state.
Overall, understanding and engineering central metabolic pathways and the connected routes are essential for optimizing the synthesis of target products. However, the trade-off between biomass and product yield is always challenging, and flexible real-time control strategies are in demand to divert carbon flux toward planned routes while blocking unplanned routes.
3. Overview of Transcription Factors-Based Biosensors
To address the dynamic and context-dependent bioproduction issues of microbial metabolism, TF-based biosensors have emerged as powerful tools for several purpose including real-time metabolic monitoring, high-throughput screening, and dynamic control. By coupling intracellular metabolic states to programmable gene expression, biosensors enable dynamic control strategies that are not achievable through static genetic modifications alone.
A typical TF-based biosensor comprises three functional modules: a regulator, a promoter containing the regulator-binding sequence, and an output module, such as a reporter gene. These components can be assembled to detect a wide range of intracellular signals, including pathway intermediates, end products, byproducts, and stress indicators such as ATP level, redox ratios, and temperature. The transcriptional regulator usually contains a DNA-binding domain and a ligand-binding domain. Upon binding of the effector molecules to the ligand-binding domain, the regulator undergoes a conformational change that alters its interaction with the corresponding promoter [8]. Depending on the regulatory outcome, biosensors can be classified as activators or repressors, serving different regulatory purposes (Figure 2). Owing to their versatility, biosensors have been widely applied in metabolic monitoring, high-throughput screening, and dynamic regulation.
When biosensors are used to monitor specific metabolic processes, the reporter outputs can track fluctuations of the target indicator in real-time, offering a sensitive and flexible approach that the conventional analytical or statical means often cannot achieve. Basing on the monitoring results, researchers can plan metabolic designs and refine engineering strategies. Biosensors have also been widely applied in the screening of high-producing strains and high-efficient enzymes. By converting production performance into easily detectable readouts like fluorescence, improved candidates can be rapidly identified in a high-throughput workflow. Dynamic regulation is typically designed to address complex metabolic challenges that are difficult to solve with static strategies alone [14,15]. These challenges include final product toxicity, the trade-off between cell growth and biosynthesis, the accumulation of imbalanced intermediates, and cofactor limitation. Whereas static interventions may lead to compromised results, dynamic regulation strategies can divide the fermentation process into multiple phases by responding to cellular metabolic states, thereby allowing robust biomass before turning on the biosynthetic reactions [13].
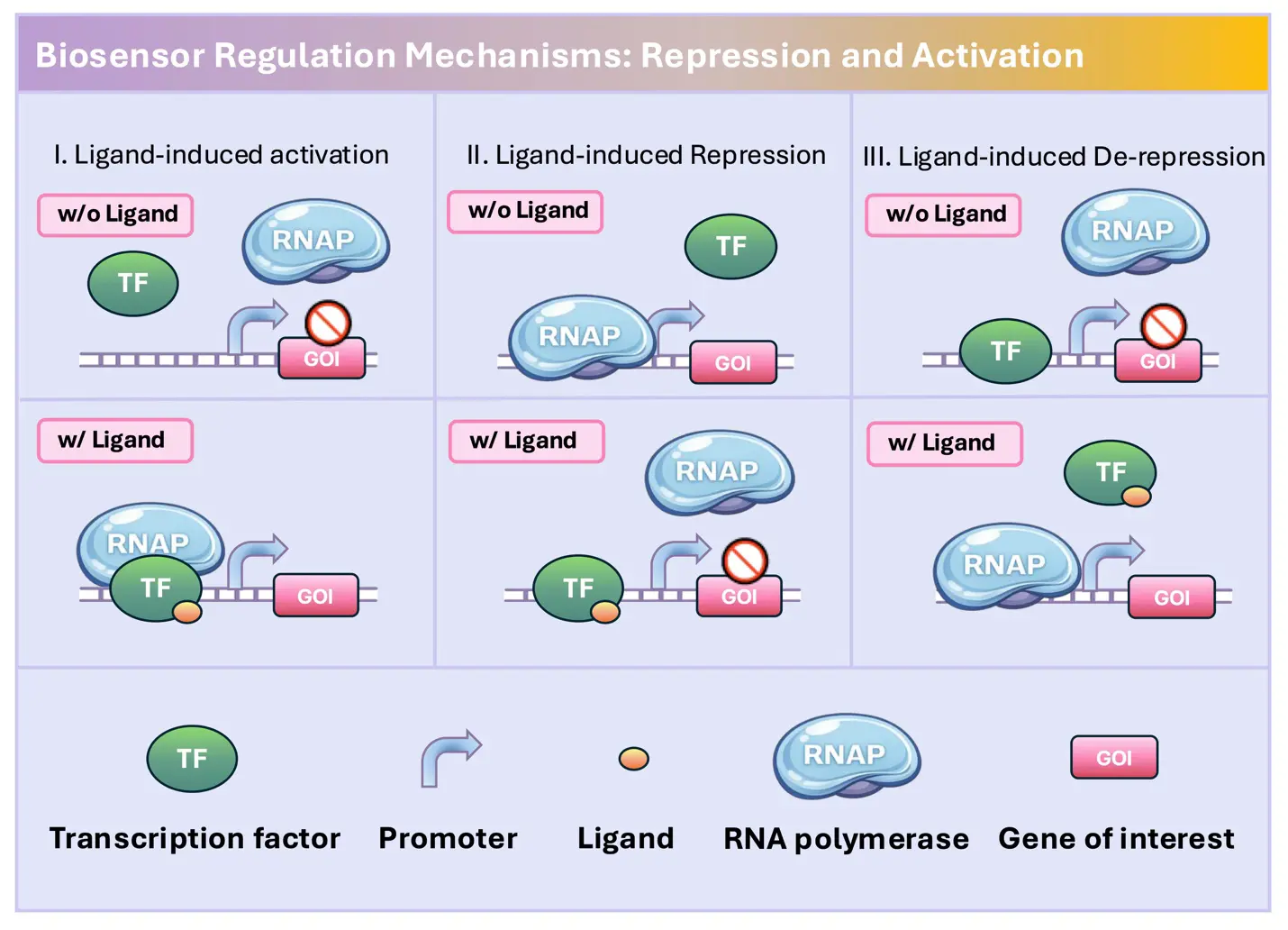
Figure 2. The mechanisms of three TFs-based biosensor types. I. Ligand-induced activation. This type of biosensor activates the transcription when the ligands present. The regulator is able to recruit RNA polymerase (RNAP) to enhance the transcription. When binding with the ligands, the regulator will undergo a conformational change which makes it binding to the promoter region or adjusting the promoter structure, turning on the downstream transcription. II. Ligand-induced Repression. This type of biosensor represses the transcription when the ligands present. The regulator is able to create a steric hindrance to block the transcription. When binding with the ligands, the regulator will undergo a conformational change, leading to the binding of regulator to the operator, inhibiting the downstream transcription. III. Ligand-induced De-repression. This type of biosensor represses the transcription when the ligands absent. When binding with the ligands, the regulator will undergo a conformational change, releasing from the promoter region and de-repressing the transcription.
Prior to implementation, wild-type TFs typically require reconstruction, characterization, and optimization. The operational range refers to the range of effector concentrations that the biosensor can detect, whereas the dynamic range refers to the difference between its minimum and maximum output signals. Depending on the intended application, biosensors must be engineered with an appropriate operational range and dynamic range to ensure adequate sensitivity and performance. In addition, tuning substrate specificity is another routine engineering procedure [16]. In some cases, the substrate specificity of broad-spectrum biosensors is narrowed to achieve selective recognition of a specific compound [17]. In others, the biosensors are engineered to respond to new substrates, thereby expanding the application scope of the original biosensors [10]. Biosensor optimization generally follows two main strategies: engineering the promoter region and modifying the regulator protein. Promoter tuning can be achieved by altering the position or sequence of regulator binding sites [18], selecting appropriate ribosome binding sites (RBS) with suitable translation strength [19], or constructing hybrid promoters to modulate transcriptional activity [20]. In parallel, the regulator protein can be optimized through either site-directed or random mutagenesis. For example, altering residues in the ligand-binding domain can modify substrate specificity or binding affinity [17], while mutations in the DNA-binding domain may influence the strength of interaction with the target promoters [21]. Collectively, these strategies enable biosensors to be tailored to specific performance requirements across diverse synthetic biology applications. In recent years, predictive tools such as machine learning have further improved the accuracy and efficiency of identifying candidate mutation sites and key residues, providing strong support for the rational design of biosensors.
4. Biosensors for Glycolysis and Fatty Acid Related Metabolites
The phosphotransferase (PTS) system comprises a series of proteins that transfer the phosphate group from phosphoenolpyruvate (PEP) to glucose, allowing the initial assimilation of glucose into glucose-6-phosphate (G6P), the first step of glycolysis. The glucose uptake rate significantly influences the maximum carbon flux and the titer of desired products. To realize monitoring of glucose uptake rate and rational carbon flux allocation based on this, Ding et al. developed a biosensor system using the transcriptional dual regulator, Mlc, which controls several genes in PTS and PEP system. Mlc is able to block the transcription of ptsG. Glucose uptake is accompanied by the dephosphorylation of EIIB~P, and EIIB can recruit Mlc to the cell membrane region, derepressing the transcription of ptsG. Basing on this mechanism, the researchers proposed a glucose uptake rate-responsive biosensor by engineering hybrid promoters containing Mlc binding sites. When the glucose uptake rate increases, the repressor Mlc is released from the hybrid promoter region, and the transcription level of the hybrid promoters elevates. Subsequently, the biosensor system was used to monitor glucose uptake of E. coli strains in different culture conditions. To explore the applicability of this biosensor on synthetic pathways, the glucose uptake rate biosensors were applied to construct feedback loops to improve the production of L-Trp, riboflavin, and D-lactic acid, respectively. On the one hand, the glucose-uptake-rate-activated genetic circuits were used to upregulate the production pathway. On the other hand, the glucose-uptake-rate-inhibited genetic circuits were used to maintain an optimal glucose uptake rate or inhibit the carbon flux competition pathways. Overall, the production titer of L-Trp, riboflavin, and D-lactic acid was increased by 91%, 108.2%, and 40.6%, respecitively [22].
Cra is a well-studied LacI/GalR family transcriptional dual regulator that responds to glycolytic flux and regulates a large number of operons involved in glycolysis, the TCA cycle, the glyoxylate shunt, and the Entner-Doudoroff pathway. While fructose-1-phosohate was proven to be the effector that can directly interact with Cra protein, this global TF has been engineered as a biosensor with either activation or repression functions by coupling with different promoters [23]. Lehning et al. employed promoters of ppsA, under the control of Cra, as a glycolytic flux biosensor. By coupling fluorescence protein with this promoter, the researchers successfully used the biosensor system to assess the glycolytic flux alteration effect caused by genetic perturbations, and identified knockout strains with altered glycolytic flux phenotypes [24]. Zhu et al. analyzed all 47 binding sites of Cra in E. coli and studied the regulatory functions and reconstructed both positive-response and negative-response biosensors by hybridizing the Cra binding sequences. After characterizing the biosensors’ dynamic range and other parameters, the negative-response biosensor was used to inhibit F6P consumption during FBP synthesis, allowing more carbon flux to Mevalonate (MVA). And both positive- and negative-response biosensors were used to improve the production of GlcNac [25]. In another study, the Cra-based biosensor promoter was used to replace the ATP synthase promoter, leading to a repression of ATP generation depend on cellular glycolysis flux. Thus, the feedback repression on pyruvate kinase was relieved, and pyruvate production was enhanced. The Cra biosensor was also used to regulate the cell membrane synthesis genes plsC, expanding the storage capacity for lycopene, and increase lycopene titer by 50-folds [26].
Pyruvate is the end product of glycolysis and also an important node in the cellular metabolic network, connecting glycolysis with multiple branches. PdhR is an Gnt family TF which the regulator activity is controlled by cellular pyruvate pool. PdhR participants in the regulation of genes involved in pyruvate dehydrogenase complex and genes on the respiratory electron transport system. When PdhR combines with its effector, pyruvate, it will be released from the corresponding promoter region, and the repression or activation effect will be relieved [27]. With the pyruvate-responsive mechanism, PdhR has been engineered to biosensors to dynamically control the metabolism flux around the pyruvate node, especially for dealing with the competition of pyruvate-derived carbon flow. Xu et al. established the pyruvate-responsive biosensors in Bacillus subtilis (B. subtilis) by employing E. coli PdhR binding box. To improve biosensor properties, they performed a series of mutations in the nonconservative region of the PdhR binding box, and an enhanced candidate with a 6.1-fold dynamic range was obtained. They further designed an antisense promoter to develop biosensors with inverted function. Finally, they applied these biosensors to build a feedback loop to control the glucaric acid synthetic pathway. When pyruvate accumulates, pgi and zwf are inhibited to restrain the carbon flow toward glycolysis and PP pathway, meanwhile, ino1 is dynamically activated, allowing the synthesis of glucaric acid precursors [28].
As an important intermediate, pyruvate can be converted to acetyl-CoA to support the running of the TCA cycle by coupling with OAA or support the synthesis of fatty acids by converting into malonyl-CoA, the building block for various fatty acids. Since fatty acids are an essential component of biomass, the development of a malonyl-CoA biosensor is beneficial to the microbial production of related compounds. Liu et al. exploited the malonyl-CoA responsive TF, FapR, from B. subtilis to develop the malonyl-CoA biosensor system. The natural FapR is involved in the regulation on fatty acid synthesis and phospholipid metabolism, the presence of malonyl-CoA antagonizes the DNA binding of FapR and releases it to initiate transcription. The researchers constructed hybrid promoters to build the biosensor in E. coli and designed an inverter by combining it with the LacI repression system. The inverter was then employed to inhibit the expression of acc genes, which catalyze the synthesis of malonyl-CoA, to achieve feedback regulation, thus relieving the toxicity caused by acc overexpression, improving cell growth conditions, and finally improving fatty acid production [29]. Li et al. also developed the malonyl-CoA biosensor in S. cerevisiae, enabling metabolite monitoring and high producer screening. The resulting recombinant yeast strain achieved enhanced titer of the malonyl-CoA derived product, 3-hydroxypropionic acid (3-HP) [30]. Another malonyl-CoA biosensor was established based on AraC-TAL, which responds to the type III polyketide triacetic acid lactone (TAL). Although this biosensor was not directly in contact with malonyl-CoA, it could use TAL to reflect the malonyl-CoA level with the help of 2-pyrone synthase (2-PS), which can produce TAL from malonyl-CoA. And the strains with enhanced malonyl-CoA production were identified by using the biosensor [31]. FadR is a bacterial repressor that responds to fatty acids/fatty acyl-CoAs. The binding of ligands with FadR antagonizes the binding of FadR with DNA. For aiding the downstream fatty acid detection, Zhang et al. developed the fatty acid/acyl-CoA responsive biosensor based on FadR TF and increased the production of biodiesel by 3-fold [32]. In another study, a FadR-based biosensor system was constructed in yeast S. cerevisiae, and was tuned by engineering the FadR expression strength, FadR species and operator numbers [33].
5. Biosensors for the TCA Cycle and Related Metabolites
Acetyl-CoA is catalyzed to enter into the tricarboxylic acid (TCA) cycle, one of the most critical pathways in central metabolism, which supplies the cell with energy, abundant reducing power, and key precursor metabolites, thereby supporting cellular respiration and sustaining healthy growth. In the early phase of microbial production, a robust TCA cycle is essential for biomass accumulation, establishing a solid foundation for fermentation. However, once the cell density reaches an optimal level, sustained high TCA cycle activity can divert a significant portion of carbon flux toward excessive cell growth. This results in inefficient carbon utilization and reduced product yield [34]. Traditional metabolic engineering approaches, such as gene knockouts, can limit TCA flux, but maintaining a low TCA cycle activity from the beginning can impair cell growth, compromise culture stability, prolong the cell growth phase, and ultimately reduce overall production efficiency. Therefore, dynamic regulation using biosensor-based tools offers a promising solution by enabling real-time monitoring of carbon flux distribution based on intracellular concentrations of TCA cycle intermediates. This approach allows the system to balance cellular growth and product formation at different fermentation stages, ensuring that valuable carbon sources are efficiently distributed to support both biomass accumulation and biosynthesis. As a result, it facilitates a dynamic equilibrium that enhances carbon utilization and overall production efficiency. To enable TCA flux-dependent monitoring and application, several biosensors responsive to TCA intermediates have been developed.
DcuS-DcuR is a two-component regulator system that responsive to various C4-dicarboxylates and citrate. The available substrates comprise several TCA intermediates such as citrate, fumarate, succinate, and malate [35,36]. The two-component regulator contains a histidine kinase (HK) and a response regulator (RR), HK is able to receive the environmental stimulus on the membrane and deliver the signal to RR by transferring phosphate groups. The phosphorylated RR binds to corresponding promoters to active or repress the downstream transcription. Thus, the two-component regulator could achieve signal transmembrane transmission [37]. DcuS-DcuR has been engineered to sense specific compounds in previous studies. For instance, Dietrich et al. utilized it as a succinate-responsive biosensor [38]. In another study, Yang et al. developed a fumaric acid-responsive two-component biosensor basing on the same two-component regulator in E. coli. By engineering promoter 5′-UTR complexes (PUTRs), the expression levels of both components and the mutagenesis of the ATP-binding site of DcuS greatly enhanced the dynamic range from 1.7-fold to 6.6-fold [39].
PcaR is an IclR family TF that reflects the ligand binding preference to multiple dicarboxylic acids. Pham et al. characterized the structure and function using the thermal shift/differential scanning fluorimetry method, demonstrating interactions between PcaR and fumaric acid, glutaric acid, malic acid, and succinic acid. Subsequently, the biosensor was constructed and tested in S. cerevisiae chassis [40]. These findings laid the foundation for further engineering and application of TCA cycle-responsive biosensors.
Due to the structural similarity among TCA cycle intermediates, the available TF often exhibit substrate specificity promiscuity. While this may allow for general monitoring of overall TCA flux, achieving a more selective response requires further fine-tunning through biosensor engineering, such as site-directed mutagenesis of ligand-binding domain.
6. Biosensors for Cofactors and Redox Homeostasis
Cofactor, especially organic coenzyme is one of the most important components of cellular metabolism. The coenzymes assist enzymatic reactions, participate in signal transduction, and can be recycled and reused as the cellular currency of energy, reducing power and specific chemical groups. For example, ATP is the main energy carrier that is involved in a large number of enzymatic reactions, supporting the basic cell growth and physiological functions. NAD(P)H serves as the electron carrier that enables most redox reactions. And CoA, the acyl-group carrier, can drive many essential cellular reactions. Since the cofactors play an indispensable role in cellular metabolism, and the shortage of cofactor supply is usually a key issue in microbial production, various engineering strategy for addressing the cofactor supply have been developed [41]. However, defining real-time cofactor levels is difficult because cofactors often exist in multiple states and undergo rapid turnover within cells due to the nature of cofactor molecules. In addition, chemical measurement methods are often laborious and time-consuming, making them unsuitable for high-throughput analysis and screening. To address these challenges, a variety of TFs-based cofactor biosensors have been identified and engineered to enable real-time monitoring of these molecules which are hard to track. These tools have enhanced our understanding of cofactor dynamics during bioproduction and provide valuable platforms for cofactor-based screening and regulatory strategies.
During aerobic respiration, ATP is mainly generated by oxidative phosphorylation, supporting cell growth, biosynthesis, and other activities. ATP fluctuates depending on multiple factors such as the carbon source, oxygen, and cell activities [42]. The absence of ATP results in hampered cell growth and decreased bioproduction. To achieve continuous monitoring of cellular ATP and the ATP level-dependent biosynthesis regulation, Mu et al. developed an ATP biosensor based on the F0-F1 ATP synthase epsilon subunit. By integrating a well-designed GFP protein, the biosensor is able to induce conformational changes of the GFP when binding with ATP and largely enhance the fluorescence level. The ATP biosensor was successfully adopted to track the ATP fluctuation in different substrates, and the superior substrates were supplemented to boost the production of fatty acid in E. coli and polyhydroxyalkanoate (PHA) in Pseudomonas putida. Furthermore, the ATP biosensor was utilized as a diagnostic tool to identify ATP limitation during limonene production, highlighting its great potential for pathway analysis and optimization [43].
NAD and NADP are essential redox cofactors for all living cells. In addition to taking part in the redox reactions, the redox cofactors are also important to the oxidative response and cell survival. The cellular redox state is determined by the ratios of NADH/NAD+ and NADPH/NADP+, with these cofactors undergoing consumption and regeneration to maintain redox homeostasis. The global redox-sensing protein (Rex) was reported to be a repression TF that controls genes related to respiration. Rex binds to specific promoter regions and functions as a transcriptional repressor. NADH and NAD+ compete for binding to Rex; when NADH binds, it induces a conformational change that causes Rex to dissociate from the promoter, relieving the repression. In contrast, NAD+ binding stabilizes Rex in its DNA-bound form, maintaining the repression. With this mechanism, Rex responds to the intracellular NADH/NAD+ ratio [44,45]. Due to the important role of Rex in cellular redox regulation, many Rex homologous have been identified, resolved, and engineered [46,47]. Liu et al. developed and characterized an B. subtilis Rex-based biosensor in E. coli. This biosensor was utilized to monitor the redox status in different respiratory chain mutants and in different carbon sources [48]. In addition to Rex, the redox-responsive promoters were also adopted to construct biosensors. Knudsen et al. exploited the yeast GPD2 promoter, which is induced under excess NADH conditions, to construct biosensors in S. cerevisiae. The GPD2-based biosensor was proved to be able to detect the perturbations of cellular redox caused by environment change and enzyme expression [49].
For the sensing of another pair of redox cofactors, NADPH/NADP+, Siedler et al. engineered the biosensors based on the [2Fe-2S] cluster-containing TF SoxRS in E. coli. The natural SoxRS regulon mediates the response to superoxide. In a high-level NADPH condition, SoxR was inactivated with the reduced [2Fe-2S] cluster, while in a low NADPH level, the oxidized [2Fe-2S] cluster causes the conformational change of SoxR, activating the expression of genes controlled by the promoter of SoxS [50]. By employing the SoxR protein and the corresponding SoxR binding region, the NADPH biosensor was engineered and demonstrated to be able to detect the varying LbAdh activities and support the high-throughput screen of NADPH-dependent enzymes [51]. Basing on SoxR and PsoxS system, Khandelwal et al. also engineered the biosensor for detecting NADP+/NADPH. In this study, lacZ was expressed under the regulation of PsoxS, when the redox ratio elevates, LacZ is generated and hydrolyzes the substrate o-nitrophenyl-β-galactoside (ONPG) into a yellow color compound o-nitrophenol (ONP) which could be measured. This study achieved real-time NADP+/NADPH monitoring and the detection of an organic pollutant triclosan [52]. In another study, Zhang et al. developed an NADPH/NADP+ biosensor in S. cerevisiae with a TF in the oxidative response pathway, Yap1p. Oxidized Yap1p undergoes a conformational change that masks its nuclear export sequence, preventing recognition by exportins and resulting in its accumulation within the nucleus. In this state, Yap1p activates the transcription of genes involved in oxidative stress defense. Conversely, in its reduced form, Yap1p is exported to the cytoplasm, thereby limiting the transcription of relative genes. By employing Yap1p and the corresponding TRX2 promoter, the researchers constructed the NADPH/NADP+ biosensors. The Yap1p-based promoter was demonstrated to detect the NADPH/NADP+ change caused by the genetic modifications. The biosensor was further applied with model-guided knockout identification and coupled with dosage-sensitive genes to connect the cell growth phenotype with the redox reporter signal, showing a promising applicability [53].
7. Biosensors for Amino Acids Metabolism
Amino acids are basic and essential molecules in all kinds of organisms, serving as the building blocks of proteins to support multiple cellular processes, such as enzymatic catalysis, structural support, and signal transduction. Additionally, amino acids are important intermediates that are involved in the TCA cycle, nucleotide synthesis pathway, and the generation of various important compounds such as neurotransmitters, porphyrins, polyamines, and phenylpropanoids. There are 20 standard amino acids that are directly encoded by codons and participant in protein generation, and these amino acids are playing crucial roles in microbial bioproduction as both the synthetic pathway final products and precursors. The amino acids products can be processed to value-added merchandises such as health supplements and animal feed additives. Meanwhile, amino acids take part in many microbial synthetic pathways of high value compounds, including L-DOPA, phenylpropanoids, and Indole derivatives. Recently, research on amino acid-responsive biosensors has grown rapidly, providing a powerful toolbox and diverse optimization strategies for engineering amino acid biosynthetic pathways.
Corynebacterium glutamicum LysG was identified as a transcriptional activator that responds to intracellular L-lysine, L-arginine, L-histidine, and L-citrulline [54]. And this TF was utilized as an L-lysine biosensor [55]. However, the wide-range substrate specificity of LysG limited its applicability in metabolic engineering cases. To address this challenge, Corte et al. semi-rationally engineered LysG into biosensors that specifically respond to L-histidine and L-arginine but eliminate L-lysine-binding capability, and the L-histidine-producing C. glutamicum variants were screened using a FACS-based screening method [56]. ArgP is a LysR-type TF that has been shown to control the transcription of the L-arginine outward transporter gene argO, and the co-effectors, L-arginine and L-lysine, show opposite effects. Specifically, L-arginine induce the activation effect on argO expression, while L-lysine causes the repression effect [57,58]. Thus, ArgP and the corresponding argO promoter have been constructed as L-arginine biosensors, allowing the relevant amino acids-inducible gene expression [59,60]. The TF Lrp from C. glutamicum is able to detect intracellular L-methionine and the branched-amino acids L-valine, L-leucine, and L-isoleucine and activates the transcription of the brnFE operon [61]. Basing on the mechanisms, Mustafi et al. developed the Lrp biosensor to respond to L-methionine and the branched-amino acids and characterized the biosensor performance when inducing with different effectors. Furthermore, the researchers successfully sorted C. glutamicum mutants with elevated excreting of L-valine, L-leucine, or L-isoleucine [62].
Aromatic amino acids comprise L-tyrosine, L-phenylalanine and L-tryptophan which are derived from shikimate pathway and serve for the synthesis of a variety of high-value microbial production compounds such as vanillin, indole derivatives, flavonoids and alkaloids. Since the shikimate pathway originates from key central metabolites PEP and E4P, maintaining a balanced supply of these precursors is critical for efficient production of aromatic compounds. Biosensors responsive to corresponding amino acids concentrations enable real-time regulation, allowing fine-tuned control and redistribution of carbon flux to support pathway balance. The L-tyrosine responsive regulator HpdA and L-phenylalanine responsive regulator PhhR of Cupriavidus necator (C. necator) were identified and characterized by Hanko et al. [63]. TyrR is a regulator which control the expression of multiple genes related to the synthesis of L-phenylalanine and L-tyrosine when L-Phenylalanine or L-tyrosine presents [64]. The feature of detecting these amino acids makes TyrR a promising biosensor tool for monitoring phenylalanine and tyrosine in urine samples [65]. TyrR was also widely developed as biosensor tool for metabolic engineering purposes, aiding the screening of hyperproducers [66,67]. TrpR is a L-tryptophan-responsive repressor that regulates the genes related to aromatic amino acids synthesis. Numerous studies have explored the engineering flexibility of the TrpR regulatory system, employing various engineering strategies to reprogram it into biosensors with distinct response performance [17,68,69]. To expand the biosensor functionality, Gong et al. designed an inverter based on TrpR and TetR, achieving the elevated production titer of tryptamine and violacein by 2.0- and 7.4-folds, respectively [70].
β-alanine is a non-proteogenic amino acid, but it serves as an important synthetic pathway intermediate. In 2020, Hanko et al. reported a β-alanine-responsive TF OapR, which was exploited from C. necator by using a genome wide identification approach. By adjusting the expression elements, OapR was successfully expressed in E. coli, worked as a β-alanine biosensor, and was further utilized for the screening of L-aspartate 1-decarboxylase [63].
8. Biosensors for Central Metabolic Byproducts
Byproducts of central metabolism are undesired compounds produced alongside central metabolic pathways. In microbial production, these byproducts often lead to substrate waste, carbon flux diversion, and inhibitory effects on biosynthesis, ultimately hindering the accumulation of target products. The accumulation of byproducts results from imbalances in carbon flux distribution and represents a non-negligible challenge for metabolic engineering. However, byproduct formations are also considered to serve as a mechanism to relieve metabolic stresses [71]. In E. coli, for example, acetate overflow metabolism is considered a route dealing with excess reducing power. When glucose is rapidly degraded, diverting carbon flux into the acetate pathway helps alleviate the accumulation of NADH generated by the TCA cycle [72]. Other compounds, such as formate, ethanol, lactate, and succinate, are also common byproducts in bioproduction scenarios. Eliminating byproduct pathways and redirecting byproduct utilization are frequently used strategies to address this issue. Due to the complexity of byproduct pathways, traditional strategies may lead to compromised results. Biosensors targeting these metabolites offer expanded strategies for addressing byproduct-related challenges. They can serve as diagnostic tools for monitoring metabolic flux imbalances and also enable the design of dynamic regulatory networks that respond to byproduct accumulation in real time.
Pseudomonas putida HpdR was identified and characterized as a 3-HP-inducible regulator with wide substrate promiscuity [73]. Recently, Zhang et al. reported the acetate responsive potential of PpHpdR. Considering the role of acetate overflow metabolism in regulating intracellular NADH levels, an acetate-responsive biosensor based on the HpdR system was constructed. This biosensor was designed to repress NADH-generating pathways and induce the expression of additional NADH dehydrogenases upon acetate accumulation, thereby enabling dynamic redox regulation. As a result, overflow metabolism in the phloroglucinol biosynthetic pathway was alleviated, leading to a 2.04-fold increase in product yield [74].
Lactate is an important overflow metabolite in cellular metabolism and a useful indicator of metabolic status. Therefore, lactate biosensors have attracted considerable attention for analytical applications in diverse biological samples. LldR is a transcriptional repressor of L-lactate and fructose/sucrose utilization operons [75]. LldR from different hosts have been identified and engineered as whole cell biosensors for detecting lactate in various samples [76,77] and as genetic elements for screening lactate high-producing targets by coupling with CRISPRi technology [78].
In addition to acetate and lactate, there are regulatory elements responsive to other byproducts that have been identified and characterized. For example, FdsR was proven to accept formate as the inducing effector [63].
9. Current Challenges, Emerging Technologies, and Future Perspectives
Although TFs-based biosensors are highly promising tools and have been widely applied in metabolic engineering to address a range of challenges, their inherent biological properties still impose several limitations. For example, the interval between transcriptional sensing and reporter protein translation can introduce response delays, and the expression of biosensor components may impose a metabolic burden on the host, potentially affecting overall cellular performance. In addition, native TFs might exhibit promiscuous substrate specificity, limited dynamic range, and narrow operational range, all of which can restrict their direct application. These challenges can be addressed by selecting efficient reporters, optimizing expression configuration, and fine-tunning biosensor components to improve performance. Another constraint is the limited repertoire of available biosensors; despite ongoing advances, current biosensors remain insufficient to comprehensively cover all key compounds within metabolic network. Fortunately, the emergence and advancement of interdisciplinary technologies have accelerated biosensor development and created new opportunities to expand their functional capabilities.
Omics technologies provide comprehensive metabolic profiling, thereby enhancing the accuracy and expanding the scope of mining novel biosensor elements [79]. Transcriptome analysis has proven to be a high-efficiency strategy for identifying promoters responsive to specific compounds. Dahl et al. evaluated the transcriptional response to toxic intermediates using whole-genome mRNA microarrays and screened promoters responding to farnesyl pyrophosphate (FPP) and 3-hydroxy-3-methylglutaryl-CoA (HMG-CoA) [80]. To overcome the limited ligand specificity of natural TFs, Nishikawa et al. developed a platform, Sensor-seq, to create biosensors with high dynamic range and novel ligand specificity for non-native pharmaceutical molecules. This method identified TF variants through measuring reporter transcripts with RNA-seq, and a TF with a large ligand binding pocket, TtgR, was selected as the engineering target. By conducting unsupervised clustering of TtgR variants generated by FuncLib [81] with Sensor-seq, TtgR mutants showing strong activity on six non-native ligands were screened out [82].
Machine learning, with its strong capabilities in data integration and predictive analysis, has offered a valuable tool in biosensor development. Owing to the structural and functional flexibility of biosensor elements, machine learning provides deeper insights into the characteristics of these elements, enabling more efficient decision-making during biosensor construction. To elucidate the connection between RBS and biosensor performance, Ding et al. studied large cross-RBSs datasets using a convolutional neural network and established a classification model, enabling predictable translation engineering of biosensors [19]. Liu et al. designed a series of de novo biosensor promoters based on a minimal core promoter framework. Using machine learning approach, they systematically modeled and analyzed the relationship between nucleotide sequences and their inducibility, providing insights into sequence-function correlations [83]. Harmer et al. developed the platform Lustro to generate high-throughput data containing the optogenetic system parameters and the corresponding induction fold. With the design of a Bayesian optimization framework that combines data-driven learning, uncertainty quantification, and experimental design, this workflow achieved high prediction accuracy and provided guidance for building multiplex light-induction programs [84].
Accurate prediction through machine learning typically requires large volumes of high-quality data. Biosensors, with their capacity for efficient and high-throughput screening, serve as an excellent source of such data, thereby offering strong support for machine learning–assisted metabolic engineering. To improve tryptophan production and obtain a hyperproducing strain, Zhang et al. combined machine learning models with mechanistic models to achieve genotype-to-phenotype predictions. Using a rational designed TrpR-based biosensor, they conducted high-throughput screening of a combinatorial library containing promoters and target genes, with fluorescence output as the phenotype readout. The resulting dataset was then used to train machine learning models, which successfully predicted combinations with elevated tryptophan production, and subsequently validated through experimental testing [85]. Overall, machine learning can facilitate the construction of optimized biosensors by predicting the performance of biological components. In turn, biosensors can generate high-quality datasets through high-throughput screening to support the development of machine learning models (Figure 3). The synergistic integration of machine learning and biosensor technologies is therefore poised to accelerate the development of more intelligent, efficient, and scalable platforms for metabolic engineering and bioproduction.
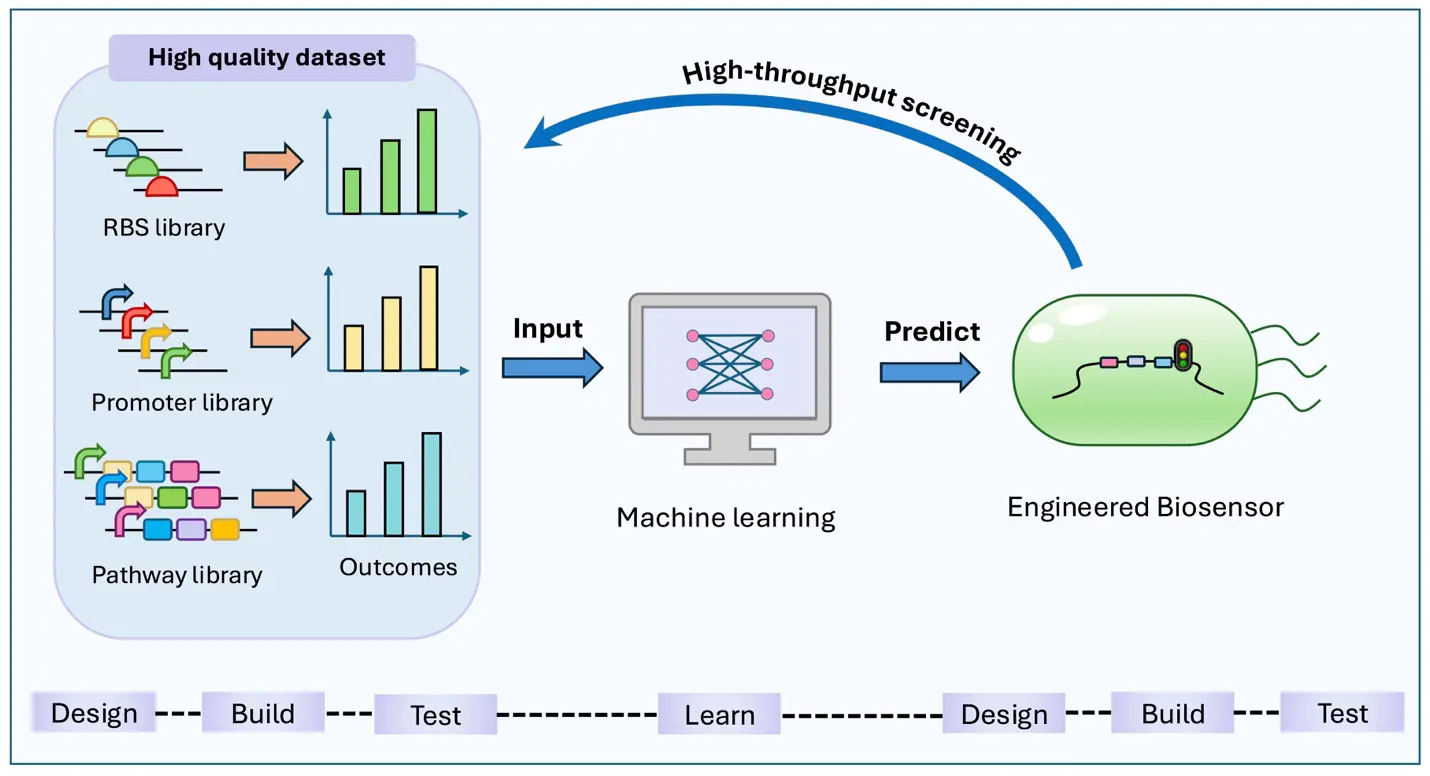
Figure 3. Machine Learning–Biosensor Design Loop. Machine learning can predict and guide the design of improved biosensors and genetic elements. In turn, high-throughput biosensor-based screening generates high-quality datasets to train machine learning models.
10. Conclusions
In this review, we systematically summarize the development and applications of biosensors responsive to central metabolism (Table 1). TFs play crucial roles in helping cells adapt to environmental fluctuations and maintain metabolic homeostasis. These regulatory proteins can be reconstructed and repurposed into biosensors that respond to a wide range of metabolic indicators. When implemented in living cells, such biosensors enable real-time monitoring of specific effectors and have been widely applied in metabolic monitoring, high-throughput screening, and the construction of dynamic regulatory circuits.
Table 1. Biosensors that are responsive to central metabolites.
|
Central Metabolite Types |
Biosensors |
Mechanisms |
Effectors |
Hosts |
References |
|---|---|---|---|---|---|
|
Glycolysis and fatty acid-derived metabolites |
Mlc |
TF |
Glucose uptake rate |
E. coli |
[22] |
|
Cra |
TF |
Fructose-1-phosphate |
E. coli |
||
|
PdhR |
TF |
Pyruvate |
Bacillus subtilis |
[28] |
|
|
FapR |
TF |
Malonyl-CoA |
E. coli, S. cerevisiae |
||
|
FadR |
TF |
Fatty acid/Fatty acyl-CoA |
E. coli, S. cerevisiae |
||
|
TCA cycle-derived metabolites |
DcuS-DcuR |
TF |
Citrate, fumarate, succinate, malate |
E. coli |
|
|
PcaR |
TF |
Fumarate, glutarate, malate, succinate |
S. cerevisiae |
[40] |
|
|
Cofactors |
F0-F1 ATP synthase epsilon subunit |
Enzyme subunit |
ATP |
E. coli, Pseudomonas putida |
[43] |
|
Rex |
TF |
NADH/NAD+ |
E. coli |
[48] |
|
|
GPD2 promoter |
Responsive promoter |
NADPH/NADP+ |
S. cerevisiae |
[49] |
|
|
SoxR |
TF |
NADPH/NADP+ |
E. coli |
||
|
Yap1p |
TF |
NADPH/NADP+ |
S. cerevisiae |
[53] |
|
|
Amino acids |
LysG |
TF |
L-lysine, L-arginine, L-histidine, L-citrulline |
Corynebacterium glutamicum, E. coli |
|
|
ArgP |
TF |
L-arginine, L-lysine |
S. cerevisiae |
||
|
Lrp |
TF |
L-methionine, L-valine, L-leucine, L-isoleucine |
Corynebacterium glutamicum |
[62] |
|
|
HpdA |
TF |
L-tyrosine |
Cupriavidus necator |
[63] |
|
|
PhhR |
TF |
L-phenylalanine |
Cupriavidus necator |
[63] |
|
|
TyrR |
TF |
L-phenylalanine, L-tyrosine |
E. coli |
||
|
TrpR |
TF |
L-tryptophan |
E. coli |
||
|
OapR |
TF |
β-alanine |
Cupriavidus necator |
[63] |
|
|
Central metabolic byproducts |
HpdR |
TF |
Acetate |
E. coli |
[74] |
|
LldR |
TF |
Lactate |
E. coli, Zymomonas mobilis |
||
|
FdsR |
TF |
Formate |
Cupriavidus necator |
[63] |
Given their central roles in biosynthesis, central metabolic pathways are frequently targets for analysis and manipulation in metabolic engineering. When metabolic imbalances arise—such as competition between growth and production, improper carbon flux distribution, or the accumulation of unwanted byproducts—overall production efficiency can be significantly compromised. Encouragingly, a growing number of biosensors capable of sensing central metabolites have been developed, providing effective tools to address these widespread challenges in metabolic engineering. In the current review, we summarize biosensors targeting glycolysis, the TCA cycle, cofactors, amino acids, and metabolic byproducts, and discusse their design strategies and application scenarios. We also examine the current limitations of biosensors and explore future directions for their development, with particular highlighting on interdisciplinary advances such as the integration of machine learning. This emerging synergy not only expands the biosensor repertoire but also harnesses the high-throughput experimental data generated by biosensors to support predictive modeling and rational design. Collectively, these developments underscore the growing importance of biosensors as versatile tools for understanding and exploiting central metabolism in advanced bioproduction.
Acknowledgments
The authors acknowledge the support of the National Institute of General Medical Sciences of the National Institutes of Health. This study was supported by the College of Engineering, The University of Georgia, Athens, Georgia, United States.
Author Contributions
Conceptualization, J.Z. and Y.Y. (Yajun Yan); Writing—Original draft preparation, J.Z.; Writing—Review & Editing, J.Z., X.G., Q.G., Y.Y. (Yingyue Yu), Y.D., R.S., N.L. and Y.Y. (Yajun Yan); Project Administration, Y.Y. (Yajun Yan); Funding Acquisition, Y.Y. (Yajun Yan).
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No new data were created or analysed in this study. Data sharing is not applicable to this article.
Funding
This work was supported by the National Institute of General Medical Sciences of the National Institutes of Health under award number R35GM128620.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Browning DF, Busby SJ. Local and global regulation of transcription initiation in bacteria. Nat. Rev. Microbiol. 2016, 14, 638–650. DOI:10.1038/nrmicro.2016.103 [Google Scholar]
-
Tsai MJ, Wang JR, Yang CD, Kao KC, Huang WL, Huang HY, et al. PredCRP: Predicting and analysing the regulatory roles of CRP from its binding sites in Escherichia coli. Sci. Rep. 2018, 8, 951. DOI:10.1038/s41598-017-18648-5 [Google Scholar]
-
Brown AN, Anderson MT, Bachman MA, Mobley HLT. The ArcAB Two-Component System: Function in Metabolism, Redox Control, and Infection. Microbiol. Mol. Biol. Rev. 2022, 86, e0011021. DOI:10.1128/mmbr.00110-21 [Google Scholar]
-
Chakerian AE, Matthews KS. Characterization of Mutations in Oligomerization Domain of Lac Repressor Protein. J. Biol. Chem. 1991, 266, 22206–22214. DOI:10.1016/S0021-9258(18)54555-5 [Google Scholar]
-
Saxild HH, Brunstedt K, Nielsen KI, Jarmer H, Nygaard P. Definition of the Bacillus subtilis PurR operator using genetic and bioinformatic tools and expansion of the PurR regulon with glyA, guaC, pbuG, xpt-pbuX, yqhZ-folD, and pbuO. J. Bacteriol. 2001, 183, 6175–6183. DOI:10.1128/JB.183.21.6175-6183.2001 [Google Scholar]
-
Gunsalus RP, Yanofsky C. Nucleotide sequence and expression of Escherichia coli trpR, the structural gene for the trp aporepressor. Proc. Natl. Acad. Sci. USA 1980, 77, 7117–7121. DOI:10.1073/pnas.77.12.7117 [Google Scholar]
-
Chae TU, Choi SY, Kim JW, Ko YS, Lee SY. Recent advances in systems metabolic engineering tools and strategies. Curr. Opin. Biotechnol. 2017, 47, 67–82. DOI:10.1016/j.copbio.2017.06.007 [Google Scholar]
-
Teng Y, Zhang J, Jiang T, Zou Y, Gong X, Yan Y. Biosensor-enabled pathway optimization in metabolic engineering. Curr. Opin. Biotechnol. 2022, 75, 102696. DOI:10.1016/j.copbio.2022.102696 [Google Scholar]
-
Jiang T, Li C, Zou Y, Zhang J, Gan Q, Yan Y. Establishing an Autonomous Cascaded Artificial Dynamic (AutoCAD) regulation system for improved pathway performance. Metab. Eng. 2022, 74, 1–10. DOI:10.1016/j.ymben.2022.08.009 [Google Scholar]
-
Xiong D, Lu S, Wu J, Liang C, Wang W, Wang W, et al. Improving key enzyme activity in phenylpropanoid pathway with a designed biosensor. Metab. Eng. 2017, 40, 115–123. DOI:10.1016/j.ymben.2017.01.006 [Google Scholar]
-
Kim SK, Kim SH, Subhadra B, Woo SG, Rha E, Kim SW, et al. A Genetically Encoded Biosensor for Monitoring Isoprene Production in Engineered Escherichia coli. ACS Synth. Biol. 2018, 7, 2379–2390. DOI:10.1021/acssynbio.8b00164 [Google Scholar]
-
Clark DP. The fermentation pathways of Escherichia coli. FEMS Microbiol. Rev. 1989, 5, 223–234. DOI:10.1016/0168-6445(89)90033-8 [Google Scholar]
-
Zou Y, Zhang J, Wang J, Gong X, Jiang T, Yan Y. A self-regulated network for dynamically balancing multiple precursors in complex biosynthetic pathways. Metab. Eng. 2024, 82, 69–78. DOI:10.1016/j.ymben.2024.02.001 [Google Scholar]
-
Zhang J, Gong X, Gan Q, Yan Y. Application of Metabolite-Responsive Biosensors for Plant Natural Products Biosynthesis. Biosensors 2023, 13, 633. DOI:10.3390/bios13060633 [Google Scholar]
-
Jiang T, Li C, Teng Y, Zhang J, Alexis Logan D, Yan Y. Dynamic Metabolic Control: From the Perspective of Regulation Logic. Synth. Biol. Eng. 2023, 1, 10012. DOI:10.35534/sbe.2023.10012 [Google Scholar]
-
Zhou GJ, Zhang F. Applications and Tuning Strategies for Transcription Factor-Based Metabolite Biosensors. Biosensors 2023, 13, 428. DOI:10.3390/bios13040428 [Google Scholar]
-
Gong X, Zhang R, Wang J, Yan Y. Engineering of a TrpR-Based Biosensor for Altered Dynamic Range and Ligand Preference. ACS Synth. Biol. 2022, 11, 2175–2183. DOI:10.1021/acssynbio.2c00134 [Google Scholar]
-
Teng Y, Gong X, Zhang J, Obideen Z, Yan Y. Investigating and Engineering an 1,2-Propanediol-Responsive Transcription Factor-Based Biosensor. ACS Synth. Biol. 2024, 13, 2177–2187. DOI:10.1021/acssynbio.4c00237 [Google Scholar]
-
Ding N, Yuan Z, Zhang X, Chen J, Zhou S, Deng Y. Programmable cross-ribosome-binding sites to fine-tune the dynamic range of transcription factor-based biosensor. Nucleic Acids Res. 2020, 48, 10602–10613. DOI:10.1093/nar/gkaa786 [Google Scholar]
-
Jiang T, Li C, Yan Y. Optimization of a p-Coumaric Acid Biosensor System for Versatile Dynamic Performance. ACS Synth. Biol. 2021, 10, 132–144. DOI:10.1021/acssynbio.0c00500 [Google Scholar]
-
Taylor ND, Garruss AS, Moretti R, Chan S, Arbing MA, Cascio D, et al. Engineering an allosteric transcription factor to respond to new ligands. Nat. Methods 2016, 13, 177–183. DOI:10.1038/nmeth.3696 [Google Scholar]
-
Ding D, Zhu Y, Bai D, Wan T, Lee SY, Zhang D. Monitoring and dynamically controlling glucose uptake rate and central metabolism. Nat. Chem. Eng. 2025, 2, 50–62. DOI:10.1038/s44286-024-00163-w [Google Scholar]
-
Huang Y, Jia KZ, Zhao W, Zhu LW. Insights into the regulatory mechanisms and application prospects of the transcription factor Cra. Appl. Environ. Microbiol. 2024, 90, e0122824. DOI:10.1128/aem.01228-24 [Google Scholar]
-
Lehning CE, Siedler S, Ellabaan MMH, Sommer MOA. Assessing glycolytic flux alterations resulting from genetic perturbations in E. coli using a biosensor. Metab. Eng. 2017, 42, 194–202. DOI:10.1016/j.ymben.2017.07.002 [Google Scholar]
-
Zhu Y, Li Y, Xu Y, Zhang J, Ma L, Qi Q, et al. Development of bifunctional biosensors for sensing and dynamic control of glycolysis flux in metabolic engineering. Metab. Eng. 2021, 68, 142–151. DOI:10.1016/j.ymben.2021.09.011 [Google Scholar]
-
Zhu Y, Gao H, Zhang J, Zhao J, Qi Q, Wang Q. De novo design of the global transcriptional factor Cra-regulated promoters enables highly sensitive glycolysis flux biosensor for dynamic metabolic control. Microb. Biotechnol. 2023, 16, 605–617. DOI:10.1111/1751-7915.14166 [Google Scholar]
-
Ogasawara H, Ishida Y, Yamada K, Yamamoto K, Ishihama A. PdhR (pyruvate dehydrogenase complex regulator) controls the respiratory electron transport system in Escherichia coli. J. Bacteriol. 2007, 189, 5534–5541. DOI:10.1128/JB.00229-07 [Google Scholar]
-
Xu X, Li X, Liu Y, Zhu Y, Li J, Du G, et al. Pyruvate-responsive genetic circuits for dynamic control of central metabolism. Nat. Chem. Biol. 2020, 16, 1261–1268. DOI:10.1038/s41589-020-0637-3 [Google Scholar]
-
Liu D, Xiao Y, Evans BS, Zhang F. Negative feedback regulation of fatty acid production based on a malonyl-CoA sensor-actuator. ACS Synth. Biol. 2015, 4, 132–140. DOI:10.1021/sb400158w [Google Scholar]
-
Li S, Si T, Wang M, Zhao H. Development of a Synthetic Malonyl-CoA Sensor in Saccharomyces cerevisiae for Intracellular Metabolite Monitoring and Genetic Screening. ACS Synth. Biol. 2015, 4, 1308–1315. DOI:10.1021/acssynbio.5b00069 [Google Scholar]
-
Li H, Chen W, Jin R, Jin JM, Tang SY. Biosensor-aided high-throughput screening of hyper-producing cells for malonyl-CoA-derived products. Microb. Cell Fact. 2017, 16, 187. DOI:10.1186/s12934-017-0794-6 [Google Scholar]
-
Zhang F, Carothers JM, Keasling JD. Design of a dynamic sensor-regulator system for production of chemicals and fuels derived from fatty acids. Nat. Biotechnol. 2012, 30, 354–359. DOI:10.1038/nbt.2149 [Google Scholar]
-
Teo WS, Hee KS, Chang MW. Bacterial FadR and synthetic promoters function as modular fatty acid sensor-regulators in Saccharomyces cerevisiae. Eng. Life Sci. 2013, 13, 456–463. DOI:10.1002/elsc.201200113 [Google Scholar]
-
Zhao M, Li Y, Wang F, Ren Y, Wei D. A CRISPRi mediated self-inducible system for dynamic regulation of TCA cycle and improvement of itaconic acid production in Escherichia coli. Synth. Syst. Biotechnol. 2022, 7, 982–988. DOI:10.1016/j.synbio.2022.05.008 [Google Scholar]
-
Davies SJ, Golby P, Omrani D, Broad SA, Harrington VL, Guest JR, et al. Inactivation and Regulation of the Aerobic C4-Dicarboxylate Transport (dctA) Gene of Escherichia coli. J. Bacteriol. 1999, 181, 5624–5635. DOI:10.1128/JB.181.18.5624-5635.1999 [Google Scholar]
-
Golby P, Davies S, Kelly DJ, Guest JR, Andrews SC. Identification and characterization of a two-component sensor-kinase and response-regulator system (DcuS-DcuR) controlling gene expression in response to C4-dicarboxylates in Escherichia coli. J. Bacteriol. 1999, 181, 1238–1248. DOI:10.1128/JB.181.4.1238-1248.1999 [Google Scholar]
-
Mascher T. Bacterial (intramembrane-sensing) histidine kinases: Signal transfer rather than stimulus perception. Trends Microbiol. 2014, 22, 559–565. DOI:10.1016/j.tim.2014.05.006 [Google Scholar]
-
Dietrich JA, Shis DL, Alikhani A, Keasling JD. Transcription factor-based screens and synthetic selections for microbial small-molecule biosynthesis. ACS Synth. Biol. 2013, 2, 47–58. DOI:10.1021/sb300091d [Google Scholar]
-
Yang H, Yang X, Lu Y, Shu Q, Zhou S, Deng Y. Engineering a fumaric acid-responsive two-component biosensor for dynamic range improvement in Escherichia coli. Syst. Microbiol. Biom. 2022, 2, 533–541. DOI:10.1007/s43393-022-00082-y [Google Scholar]
-
Pham C, Nasr MA, Skarina T, Di Leo R, Kwan DH, Martin VJJ, et al. Functional and structural characterization of an IclR family transcription factor for the development of dicarboxylic acid biosensors. FEBS J. 2024, 291, 3481–3498. DOI:10.1111/febs.17149 [Google Scholar]
-
Wang M, Chen B, Fang Y, Tan T. Cofactor engineering for more efficient production of chemicals and biofuels. Biotechnol. Adv. 2017, 35, 1032–1039. DOI:10.1016/j.biotechadv.2017.09.008 [Google Scholar]
-
Lin WH, Jacobs-Wagner C. Connecting single-cell ATP dynamics to overflow metabolism, cell growth, and the cell cycle in Escherichia coli. Curr. Biol. 2022, 32, 3911–3924.e4. DOI:10.1016/j.cub.2022.07.035 [Google Scholar]
-
Mu X, Evans TD, Zhang F. ATP biosensor reveals microbial energetic dynamics and facilitates bioproduction. Nat. Commun. 2024, 15, 5299. DOI:10.1038/s41467-024-49579-1 [Google Scholar]
-
Ravcheev DA, Li X, Latif H, Zengler K, Leyn SA, Korostelev YD, et al. Transcriptional regulation of central carbon and energy metabolism in bacteria by redox-responsive repressor Rex. J. Bacteriol. 2012, 194, 1145–1157. DOI:10.1128/JB.06412-11 [Google Scholar]
-
Brekasis D, Paget MS. A novel sensor of NADH/NAD+ redox poise in Streptomyces coelicolor A3(2). EMBO J. 2003, 22, 4856–4865. DOI:10.1093/emboj/cdg453 [Google Scholar]
-
Gyan S, Shiohira Y, Sato I, Takeuchi M, Sato T. Regulatory loop between redox sensing of the NADH/NAD(+) ratio by Rex (YdiH) and oxidation of NADH by NADH dehydrogenase Ndh in Bacillus subtilis. J. Bacteriol. 2006, 188, 7062–7071. DOI:10.1128/JB.00601-06 [Google Scholar]
-
Pagels M, Fuchs S, Pané‐Farré J, Kohler C, Menschner L, Hecker M, et al. Redox sensing by a Rex-family repressor is involved in the regulation of anaerobic gene expression in Staphylococcus aureus. Mol. Microbiol. 2010, 76, 1142–1161. DOI:10.1111/j.1365-2958.2010.07105.x [Google Scholar]
-
Liu Y, Landick R, Raman S. A Regulatory NADH/NAD+ Redox Biosensor for Bacteria. ACS Synth. Biol. 2019, 8, 264–273. DOI:10.1021/acssynbio.8b00485 [Google Scholar]
-
Knudsen JD, Carlquist M, Gorwa-Grauslund M. NADH-dependent biosensor in Saccharomyces cerevisiae: Principle and validation at the single cell level. AMB Express 2014, 4, 81. DOI:10.1186/s13568-014-0081-4 [Google Scholar]
-
Liochev SI, Fridovich I. Fumarase C, the Stable Fumarase of Escherichia coli, Is Controlled by the Soxrs Regulon. Proc. Natl. Acad. Sci. USA 1992, 89, 5892–5896. DOI:10.1073/pnas.89.13.5892 [Google Scholar]
-
Siedler S, Schendzielorz G, Binder S, Eggeling L, Bringer S, Bott M. SoxR as a Single-Cell Biosensor for NADPH-Consuming Enzymes in Escherichia coli. ACS Synth. Biol. 2014, 3, 41–47. DOI:10.1021/sb400110j [Google Scholar]
-
Khandelwal H, Mutyala S, Kim M, Kong DS, Kim C, Kim JR. In situ monitoring of intracellular NADP(+)/NADPH state: Engineered E. coli redox biosensor for carbon metabolism and toxicant detection. Biosens. Bioelectron. 2026, 294, 118213. DOI:10.1016/j.bios.2025.118213 [Google Scholar]
-
Zhang J, Sonnenschein N, Pihl TPB, Pedersen KR, Jensen MK, Keasling JD. Engineering an NADPH/NADP(+) Redox Biosensor in Yeast. ACS Synth. Biol. 2016, 5, 1546–1556. DOI:10.1021/acssynbio.6b00135 [Google Scholar]
-
Bellmann A, Vrljić M, Pátek M, Sahm H, Krämer R, Eggeling L. Expression control and specificity of the basic amino acid exporter LysE of Corynebacterium glutamicum. Microbiology 2001, 147, 1765–1774. DOI:10.1099/00221287-147-7-1765 [Google Scholar]
-
Binder S, Schendzielorz G, Stäbler N, Krumbach K, Hoffmann K, Bott M, et al. A high-throughput approach to identify genomic variants of bacterial metabolite producers at the single-cell level. Genome Biol. 2012, 13, R40. DOI:10.1186/gb-2012-13-5-r40 [Google Scholar]
-
Della Corte D, van Beek HL, Syberg F, Schallmey M, Tobola F, Cormann KU, et al. Engineering and application of a biosensor with focused ligand specificity. Nat. Commun. 2020, 11, 4851. DOI:10.1038/s41467-020-18400-0 [Google Scholar]
-
Nandineni MR, Gowrishankar J. Evidence for an arginine exporter encoded by yggA (argO) that is regulated by the LysR-Type transcriptional regulator ArgP in Escherichia coli. J. Bacteriol. 2004, 186, 3539–3546. DOI:10.1128/Jb.186.11.3539-3546.2004 [Google Scholar]
-
Marbaniang CN, Gowrishankar J. Role of ArgP (IciA) in lysine-mediated repression in Escherichia coli. J. Bacteriol. 2011, 193, 5985–5996. DOI:10.1128/JB.05869-11 [Google Scholar]
-
Skjoedt ML, Snoek T, Kildegaard KR, Arsovska D, Eichenberger M, Goedecke TJ, et al. Engineering prokaryotic transcriptional activators as metabolite biosensors in yeast. Nat. Chem. Biol. 2016, 12, 951–958. DOI:10.1038/Nchembio.2177 [Google Scholar]
-
Ambri F, D’Ambrosio V, Di Blasi R, Maury J, Jacobsen SAB, McCloskey D, et al. High-Resolution Scanning of Optimal Biosensor Reporter Promoters in Yeast. ACS Synth. Biol. 2020, 9, 218–226. DOI:10.1021/acssynbio.9b00333 [Google Scholar]
-
Lange C, Mustafi N, Frunzke J, Kennerknecht N, Wessel M, Bott M, et al. Lrp of Corynebacterium glutamicum controls expression of the brnFE operon encoding the export system for L-methionine and branched-chain amino acids. J. Biotechnol. 2012, 158, 231–241. DOI:10.1016/j.jbiotec.2011.06.003 [Google Scholar]
-
Mustafi N, Grunberger A, Kohlheyer D, Bott M, Frunzke J. The development and application of a single-cell biosensor for the detection of l-methionine and branched-chain amino acids. Metab. Eng. 2012, 14, 449–457. DOI:10.1016/j.ymben.2012.02.002 [Google Scholar]
-
Hanko EKR, Paiva AC, Jonczyk M, Abbott M, Minton NP, Malys N. A genome-wide approach for identification and characterisation of metabolite-inducible systems. Nat. Commun. 2020, 11, 1213. DOI:10.1038/s41467-020-14941-6 [Google Scholar]
-
Pittard J, Camakaris H, Yang J. The TyrR regulon. Mol. Microbiol. 2005, 55, 16–26. DOI:10.1111/j.1365-2958.2004.04385.x [Google Scholar]
-
Lin C, Jair YC, Chou YC, Chen PS, Yeh YC. Transcription factor-based biosensor for detection of phenylalanine and tyrosine in urine for diagnosis of phenylketonuria. Anal. Chim. Acta 2018, 1041, 108–113. DOI:10.1016/j.aca.2018.08.053 [Google Scholar]
-
Mahr R, von Boeselager RF, Wiechert J, Frunzke J. Screening of an Escherichia coli promoter library for a phenylalanine biosensor. Appl. Microbiol. Biotechnol. 2016, 100, 6739–6753. DOI:10.1007/s00253-016-7575-8 [Google Scholar]
-
Liu Y, Zhuang Y, Ding D, Xu Y, Sun J, Zhang D. Biosensor-Based Evolution and Elucidation of a Biosynthetic Pathway in Escherichia coli. ACS Synth. Biol. 2017, 6, 837–848. DOI:10.1021/acssynbio.6b00328 [Google Scholar]
-
Ellefson JW, Ledbetter MP, Ellington AD. Directed evolution of a synthetic phylogeny of programmable Trp repressors. Nat. Chem. Biol. 2018, 14, 361–367. DOI:10.1038/s41589-018-0006-7 [Google Scholar]
-
Li H, Zhang W, Han Y, Tang G, Lu F, Qin HM. Programming a bacterial biosensor for directed evolution of tryptophan hydroxylase via high-throughput droplet sorting. Biosens. Bioelectron. 2025, 271, 117072. DOI:10.1016/j.bios.2024.117072 [Google Scholar]
-
Gong X, Teng Y, Zhang J, Gan Q, Song M, Alaraj A, et al. Architecting a transcriptional repressor-based genetic inverter for tryptophan derived pathway regulation in Escherichia coli. Metab. Eng. 2024, 86, 66–77. DOI:10.1016/j.ymben.2024.09.008 [Google Scholar]
-
Basan M, Hui S, Okano H, Zhang Z, Shen Y, Williamson JR, et al. Overflow metabolism in Escherichia coli results from efficient proteome allocation. Nature 2015, 528, 99–104. DOI:10.1038/nature15765 [Google Scholar]
-
Vemuri GN, Altman E, Sangurdekar DP, Khodursky AB, Eiteman MA. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl. Environ. Microbiol. 2006, 72, 3653–3661. DOI:10.1128/AEM.72.5.3653-3661.2006 [Google Scholar]
-
Hanko EKR, Minton NP, Malys N. Characterisation of a 3-hydroxypropionic acid-inducible system from Pseudomonas putida for orthogonal gene expression control in Escherichia coli and Cupriavidus necator. Sci. Rep. 2017, 7, 1724. DOI:10.1038/s41598-017-01850-w [Google Scholar]
-
Zhang J, Wang J, Jiang T, Gong X, Gan Q, Teng Y, et al. Engineering an Overflow-Responsive Regulation System for Balancing Cellular Redox and Optimizing Microbial Production. Biotechnol. Bioeng. 2025, 122, 1561–1573. DOI:10.1002/bit.28976 [Google Scholar]
-
Gao YG, Suzuki H, Itou H, Zhou Y, Tanaka Y, Wachi M, et al. Structural and functional characterization of the LldR from Corynebacterium glutamicum: A transcriptional repressor involved in L-lactate and sugar utilization. Nucleic Acids Res. 2008, 36, 7110–7123. DOI:10.1093/nar/gkn827 [Google Scholar]
-
Goers L, Ainsworth C, Goey CH, Kontoravdi C, Freemont PS, Polizzi KM. Whole-cell Escherichia coli lactate biosensor for monitoring mammalian cell cultures during biopharmaceutical production. Biotechnol. Bioeng. 2017, 114, 1290–1300. DOI:10.1002/bit.26254 [Google Scholar]
-
Zúñiga A, Camacho M, Chang HJ, Fristot E, Mayonove P, Hani EH, et al. Engineered l-Lactate Responding Promoter System Operating in Glucose-Rich and Anoxic Environments. ACS Synth. Biol. 2021, 10, 3527–3536. DOI:10.1021/acssynbio.1c00456 [Google Scholar]
-
Peng Q, Bao W, Geng B, Yang S. Biosensor-assisted CRISPRi high-throughput screening to identify genetic targets in Zymomonas mobilis for high d-lactate production. Synth. Syst. Biotechnol. 2024, 9, 242–249. DOI:10.1016/j.synbio.2024.02.002 [Google Scholar]
-
Lim HG, Rychel K, Sastry AV, Bentley GJ, Mueller J, Schindel HS, et al. Machine-learning from Pseudomonas putida KT2440 transcriptomes reveals its transcriptional regulatory network. Metab. Eng. 2022, 72, 297–310. DOI:10.1016/j.ymben.2022.04.004 [Google Scholar]
-
Dahl RH, Zhang F, Alonso-Gutierrez J, Baidoo E, Batth TS, Redding-Johanson AM, et al. Engineering dynamic pathway regulation using stress-response promoters. Nat. Biotechnol. 2013, 31, 1039–1046. DOI:10.1038/nbt.2689 [Google Scholar]
-
Khersonsky O, Lipsh R, Avizemer Z, Ashani Y, Goldsmith M, Leader H, et al. Automated Design of Efficient and Functionally Diverse Enzyme Repertoires. Mol. Cell 2018, 72, 178–186.e5. DOI:10.1016/j.molcel.2018.08.033 [Google Scholar]
-
Nishikawa KK, Chen J, Acheson JF, Harbaugh SV, Huss P, Frenkel M, et al. Highly multiplexed design of an allosteric transcription factor to sense new ligands. Nat. Commun. 2024, 15, 10001. DOI:10.1038/s41467-024-54260-8 [Google Scholar]
-
Liu X, Gupta STP, Bhimsaria D, Reed JL, Rodríguez-Martínez JA, Ansari AZ, et al. De novo design of programmable inducible promoters. Nucleic Acids Res. 2019, 47, 10452–10463. DOI:10.1093/nar/gkz772 [Google Scholar]
-
Harmer ZP, Thompson JC, Cole DL, Venturelli OS, Zavala VM, McClean MN. Dynamic Multiplexed Control and Modeling of Optogenetic Systems Using the High-Throughput Optogenetic Platform, Lustro. ACS Synth. Biol. 2024, 13, 1424–1433. DOI:10.1021/acssynbio.3c00761 [Google Scholar]
-
Zhang J, Petersen SD, Radivojevic T, Ramirez A, Pérez-Manríquez A, Abeliuk E, et al. Combining mechanistic and machine learning models for predictive engineering and optimization of tryptophan metabolism. Nat. Commun. 2020, 11, 4880. DOI:10.1038/s41467-020-17910-1 [Google Scholar]