The Impacts of Ozone Inhibition on the Growth of Bacteria on Biomass Production of Haematococcus lacustris in Mixotrophic Cultivation

The Impacts of Ozone Inhibition on the Growth of Bacteria on Biomass Production of Haematococcus lacustris in Mixotrophic Cultivation
Ahmad Abd-El-Aziz
1,2,*
 Yang Su
3
Yan Shang
3
Spiros N. Agathos
2,4
Jian Li
3,*
Yang Su
3
Yan Shang
3
Spiros N. Agathos
2,4
Jian Li
3,*
Received: 29 April 2026 Revised: 29 May 2026 Accepted: 08 June 2026 Published: 24 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Introduction
The freshwater alga Haematococcus lacustris (formerly classified as Haematococcus pluvialis) has been used in the production of the carotenoid astaxanthin. Astaxanthin is a carotenoid pigment produced in cells for its antioxidant activity and has been termed the “king of carotenoids” [1,2,3]. Under stress caused by nutrient deficiency and intense light exposure, H. lacustris begins to produce high quantities of carotenoids, particularly astaxanthin. Unlike synthetic astaxanthin, algae-derived astaxanthin contains a higher proportion of the more active 3S,3’S stereoisomer, which results in a more bioactive product [4]. Additionally, algae grown in photobioreactors produce less harmful waste than those produced by chemical synthesis [5]. As a result, algae such as H. lacustris have been investigated as a sustainable method for industrial production of astaxanthin [6].
H. lacustris can use both autotrophy, if exposed to light and CO2, and heterotrophy, if given a source of organic carbon [7,8]. While the presence of intense light is necessary for triggering carotenoid production, the presence of organic carbon can improve the growth of the algae in a nitrogen-limited environment [9,10]. However, these compounds also allow the growth of heterotrophic contaminants such as bacteria and fungi, which can reduce the growth and productivity of algal culture [11,12]. Therefore, one of the major limitations of mixotrophic growth for algal biomanufacturing has been maintaining an axenic environment for the algae [13,14].
A frequently used method for establishing and maintaining an axenic environment is the application of antibiotics such as chloramphenicol, ampicillin, and penicillin [15,16]. For H. lacustris specifically, treatment of the cultures with amoxicillin has been found to be an effective approach [17]. Sodium hypochlorite (bleach) and calcium hypochlorite have been used to kill a variety of contaminating microorganisms by generating reactive oxygen species (ROS), which inhibit growth through oxidative damage [18,19]. Another approach for the production of ROS is the use of triiodide anions to inhibit bacteria in cultures of the microalgae Nannochloropsis gaditana [20]. Triiodide anions can stabilize iodine, which is a well-established antimicrobial molecule. Algae could also be co-cultured with mutualistic bacteria to increase algal growth, while the symbiotic bacteria could compete with other contaminating species [21]. Advantages of this technique include simplicity of the approach, with improved growth and output. However, this strategy would need a better understanding and control of these benign organisms, as well as additional research into the molecular interactions between mutualistic bacteria and algae. Further, because symbiotic bacteria and their biochemical interactions are specific to each algal species, this approach cannot be generalized across algal species.
One approach that has not been well explored for establishing axenic algal cultures is the application of ozone, a light blue oxidizing gas that penetrates the cell membrane of bacteria and produces ROS in cells [22,23]. Ozone has been studied for inhibiting microbial growth in food storage [24] and to prevent the growth of antibiotic-resistant bacteria in healthcare settings, including Gram-positive Staphylococcus aureus and Gram-negative Pseudomonas aeruginosa [25]. Another application of ozone as a disinfectant is in wastewater treatment [26]. An advantage of ozone disinfection is that the only byproduct released by cells is oxygen gas, making it more environmentally friendly than other methods, such as antibiotics, which can contaminate wastewater [27]. However, because ozone has not been adequately studied for establishing axenic cultures of microalgae, the optimal ozone concentration to inhibit bacterial growth without significantly affecting algal growth has not been determined.
Another factor affecting the growth of algae is light exposure—specifically, various regions of the visible spectrum which have been shown to have effects on photosynthetic efficiency [28,29]. For example, studies have shown that red light promotes the growth of H. lacustris, while blue light induces astaxanthin accumulation [30]. The light quality is a key factor affecting the photosynthetic efficiency and composition of microalgae.
The aim of this research was to optimize the concentration and flow rate of ozone used to inhibit bacterial growth in xenic cultures of H. lacustris. Additionally, different colors of light exposure were used to enhance algal growth and biomass production. The experimental design is illustrated in Figure 1.
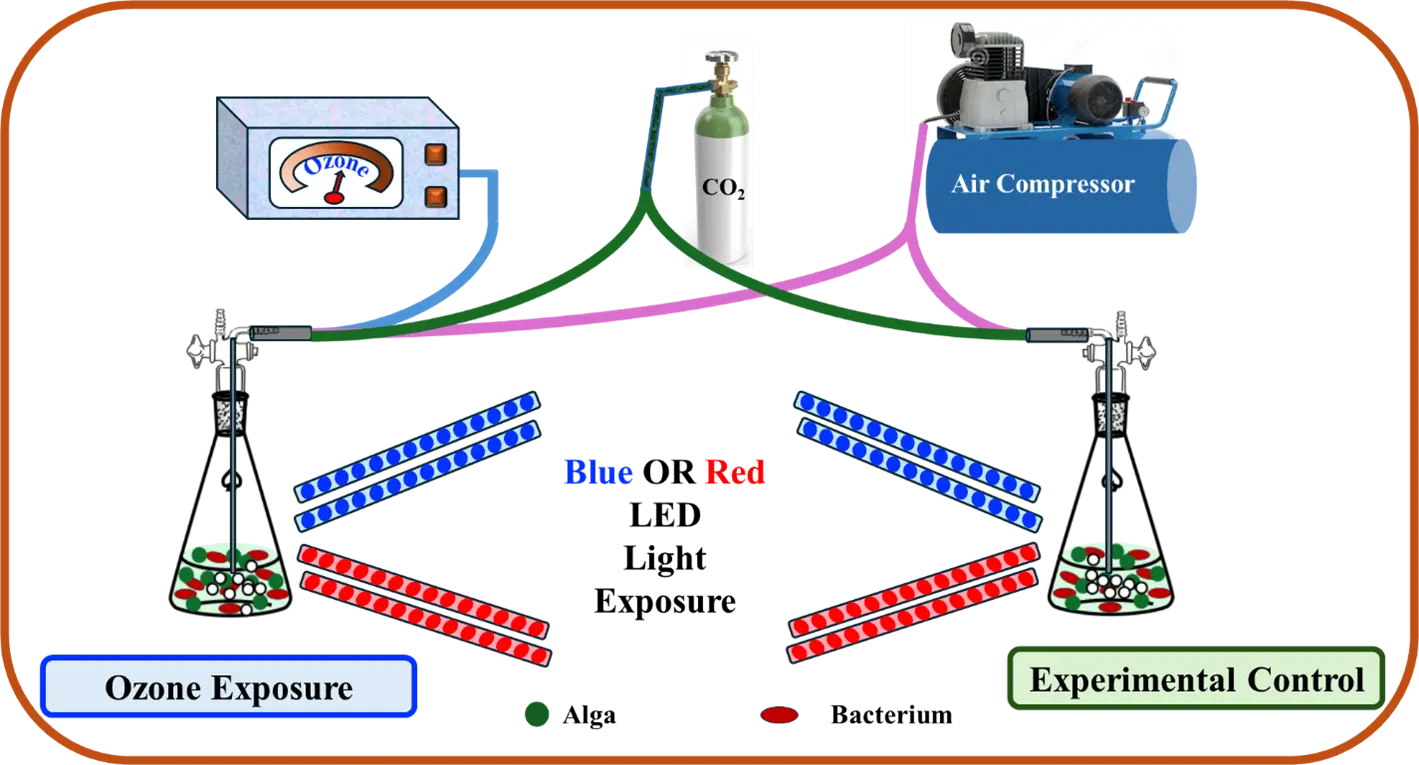
2. Materials and Methods
2.1. Ozone Concentration and Flow Rate
The H. lacustris algae solution was inoculated into six 2L flasks containing 100 mL of Bold’s Basal Medium (BBM) for three sets of parallel experiments and cultivated under white light. Continuous illumination was applied with an intensity between 50–60 lux, and the temperature was set to 25 ± 1 °C. Ozone was added to the culture using a CH-ZTW3G benchtop ozonator (Guangzhou Chuanghuan Ozone Electric Appliance Co., Ltd., Guangzhou, China) connected to an air pump. For each experimental group, the culture conditions were kept consistent, except for the ozone concentration, which was set to 0.40 mg/L, 4.00 mg/L, or 40.00 mg/L, and the ozone flow rate, which was set to 15 mL/h, 30 mL/h, or 45 mL/h. The low dose of 0.4 mg/L is equivalent to the maximum allowable concentration set by the United States Food and Drug Administration (FDA) for residual levels in bottled water (21 CFR 184.1563(c) [31]), while the higher doses were intended to study the concentration-dependent effects of ozone on the algal growth. The optical density value at 680 nm of the algae solution was measured daily using a spectrophotometer. After seven days, the number of bacteria was measured in colony forming units (CFU) by the plate count method, and the data were analyzed to find out the appropriate ozone concentration. The statistical analysis of data was performed with one-way ANOVAs using the IBM SPSS 27.0 software package.
2.2. Ozone and Different Light Sources
To investigate the effect of different light sources on the growth of algae and bacteria with and without ozone, the algae were grown in BBM in the same conditions and separated into four groups: with red light and 0.08 mg/L ozone exposure, with red light alone, with blue light and 0.08 mg/L ozone exposure, and with blue light alone. Illumination with red or blue light was provided by red or blue LED light strips with a power of 20 W per strip.
2.3. Mixotrophic Cultivation
Sodium acetate was employed as an organic carbon source to supplement the BBM medium for mixotrophic cultivation [32]. Each group consisted of two flasks of H. lacustris culture, with sodium acetate added to the medium, and grown for 16 days. One group was treated with a mixture of oxygen, carbon dioxide, and ozone, while the other group was directly injected with a mixture of oxygen and carbon dioxide. The biomass, OD value, and number of bacteria were measured every day. The biomass concentration was measured by filtering the algal culture through 5 µm glass fiber filter paper and drying it for over 24 h. The resulting mass was then weighed and divided by the volume of culture to yield the biomass concentration.
3. Results
3.1. Ozone Concentration and Flow Rate
Table 1 shows that the growth extent of H. lacustris exposed to ozone was affected compared to that of algae untreated with ozone. The OD values of H. lacustris showed an increasing trend for the negative control and 0.40 mg/L ozone groups but displayed a decreasing trend at 4.00 mg/L and 40.0 mg/L concentrations of ozone. The algal growth curve in the control group without ozone showed a steady upward trend, reaching a plateau on the seventh day of culture, and its OD value increased by about 47.1%. When the ozone concentration was 0.4 mg/L, the growth curve became flat by day seven, and its OD value increased by about 104.6% compared with the day zero value. At a concentration of 4.0 mg/L, the algal growth curve showed an overall downward trend. The OD value increased slightly on the first day of culture, but the OD value continued to decrease from the second day, and when it reached the seventh day of culture, the growth curve tended to be flat, and the OD value decreased by about 39.9% by the end of the growth period. When the introduced ozone concentration was 40.0 mg/L, the growth curve of H. lacustris decreased most rapidly compared with the other two concentrations, and its OD value continued to diminish from the first day, while the growth curve flattened on the seventh day, resulting in a decrease of 48.4% by day 7.
Table 1. Multiple comparisons of the effects of different ozone concentrations on OD values of H. lacustris.
|
Ozone Concentration (mg/L) |
OD680 |
|---|---|
|
0.00 |
0.286 ± 0.0429 c |
|
0.40 |
0.359 ± 0.0862 c |
|
4.00 |
0.191 ± 0.0348 c |
|
40.00 |
0.180 ± 0.0415 c |
Note: All OD values are mean ± standard deviation (n = 2). Different letters in superscript indicate statistically significant differences (p < 0.05) between groups.
The ozone flow rate (which was controlled by the speed of the ozone pump) was also varied to determine the effect on algal growth. For an ozone concentration of 0.40 mg/L, an upward trend in OD680 was observed by the end of the growth period at all three flow rates, which was notably higher than that of the untreated control group. The increases were ~97.8% for 15 mL/h, ~99% for 30 mL/h, and ~105% for 45 mL/h, compared to the starting values. Without the addition of ozone, the algae growth was lower, with an increase in the OD value of only ~51%, likely due to the presence of contaminating bacteria. The increase in OD value corresponded to the increase in ozone flow rate, although the overall effect is minor. At a concentration of 4.0 mg/L, when the flow rate was 15 mL/h, the OD of H. lacustris decreased by day seven by about 30%. As the rate increased, the OD values continued to decrease, by ~34% for 30 mL/h and ~39% for 45 mL/h. For the highest ozone concentration of 40.00 mg/L, when the flow rate was 15 mL/h, the OD of H. lacustris decreased by day seven by about 39%. As the flow rate increased, the OD values continued to decrease, by ~43% for 30 mL/h and ~48% for 45 mL/h. For the negative control without ozone, the final OD680 value increased by about 50% from day 0. Therefore, for 4.0 and 40.0 mg/L ozone, the growth curve decreases for all flow rates, showing that concentrations above 4.0 mg/L ozone inhibit the algal growth with no significant changes caused by the flow rate.
3.2. Antibacterial Activity
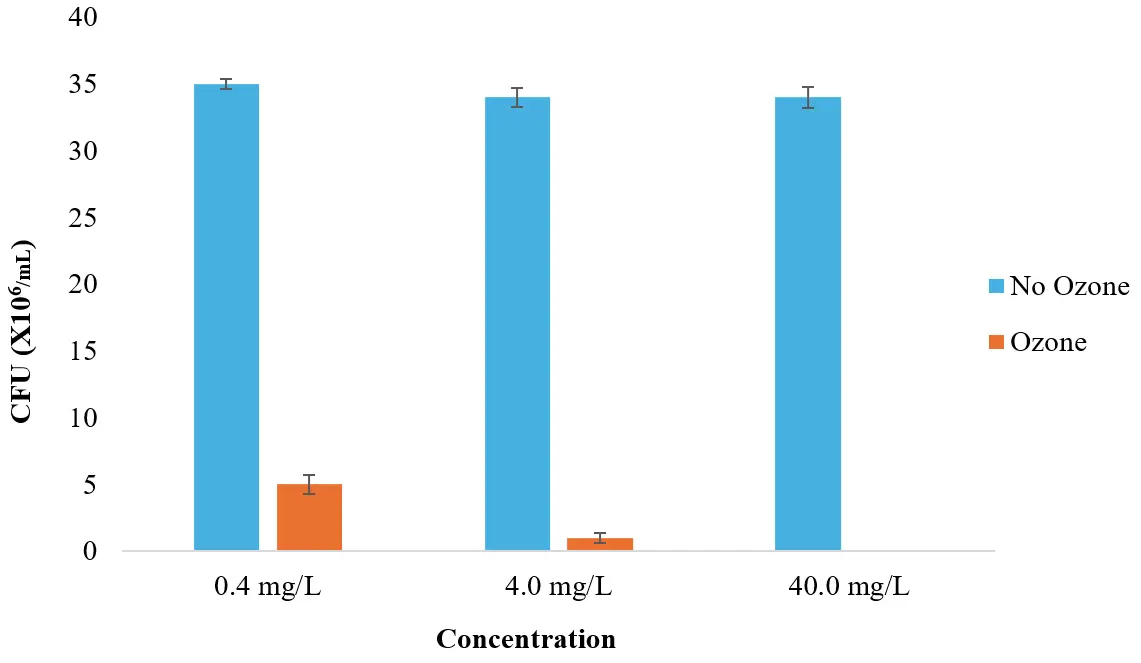
To evaluate the bacterial growth in the cultures, dilutions of 1:105, 1:106, and 1:107 were made from the 0.40 mg/L, 4.00 mg/L, and 40.00 mg/L ozone-treated groups and then cultured on agar plates. When the dilution factor was 100,000×, a larger CFU number on the bacterial plate was observed (Figure S1). As can be seen from Figure 2, the number of bacteria growing on the agar plates was much smaller after the ozone treatment than the number of bacteria growing on the bacterial plate without ozone, indicating that ozone can effectively kill bacteria. Further, as the ozone concentration increased, the number of bacteria decreased. Compared with the colony number of the control group, all bacterial strains decreased the most when the ozone concentration was 40.0 mg/L, as all miscellaneous bacteria were killed. At an ozone concentration of 0.40 mg/L, the number of bacteria decreased by ~85%, while a 4.00 mg/L concentration caused a ~97% reduction in the number of bacterial colonies.
3.3. Different Light Sources
For all following experiments, an ozone concentration of 0.08 mg/L was used to optimize the bacterial inhibition while minimizing the algal impacts. The effects of different light sources on the OD values are shown in Figure 3. Two 10 mL samples from each flask were selected and measured at a wavelength of 680 nm. The highest OD was observed for algae grown under red light, followed by blue light. The cultures all began at a comparable level of growth but exhibited observable differences after a few days (by day three for red light and by day five for blue).
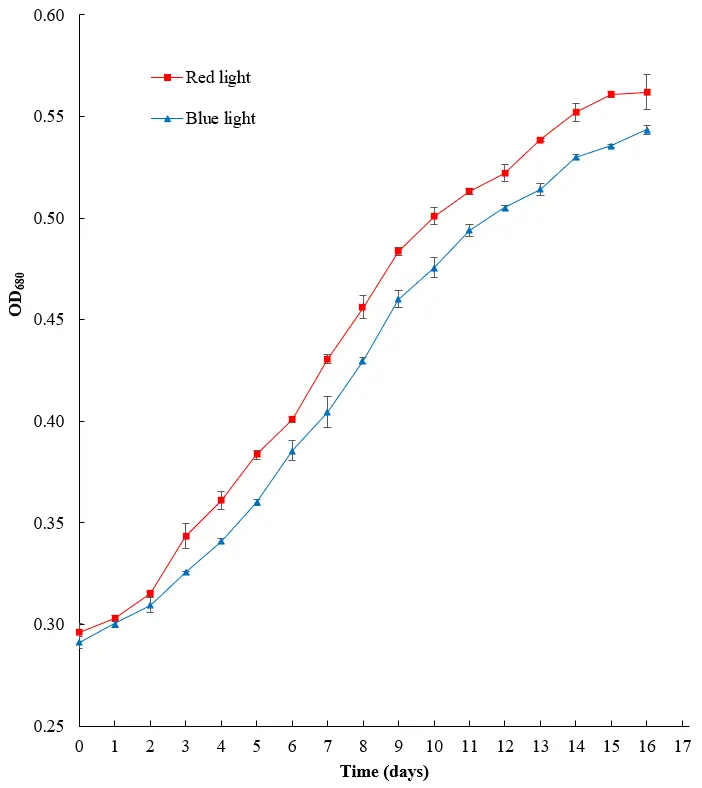
Figure 3. Effect of different sources of illumination on the OD680 value for H. lacustris culture (n = 2). Standard deviations for all points are shown as error bars.
The growth extent of the algae was also measured using the biomass concentration, as seen in Figure 4. While the biomass concentration was similar for the first few days, differences for each condition were observed after day six of cultivation. Red light produced the greatest biomass, followed by blue light. By the end of the cultivation period, the biomass of algae grown under red light was 0.02 g/L higher than that of algae grown under blue light. It can therefore be concluded that red light is the best light source for cultivating H. lacustris.
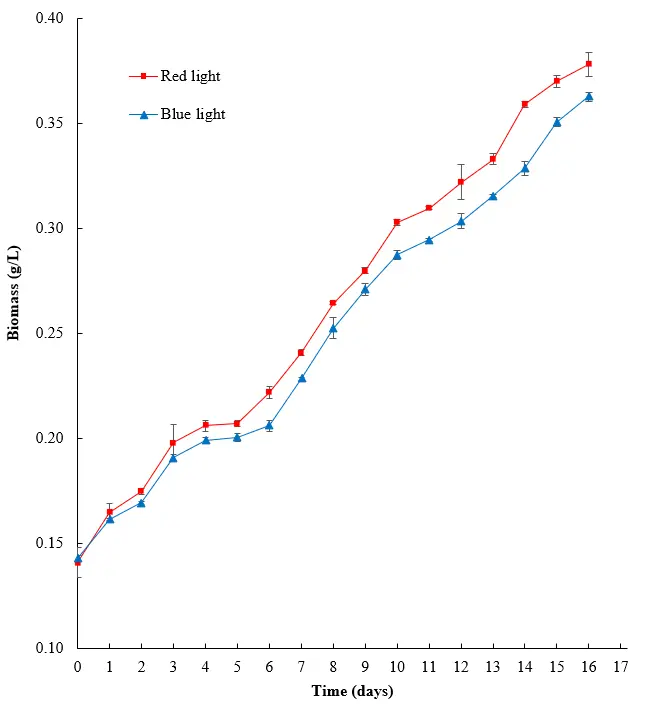
Figure 4. Effects of different light sources on the biomass of H. lacustris. Standard deviations for all points are shown as error bars.
The photosynthetic photon flux density (PPFD) between 400 and 700 nm was measured using a quantum sensor light meter. As illustrated in Figure 5, red light showed enhanced optical quantum flux for the H. lacustris cultures. The PPFD for red light was 596.112 ± 53.650 µmol·m−2·s−1, and for blue light was 476.148 ± 42.853 µmol·m−2·s−1.
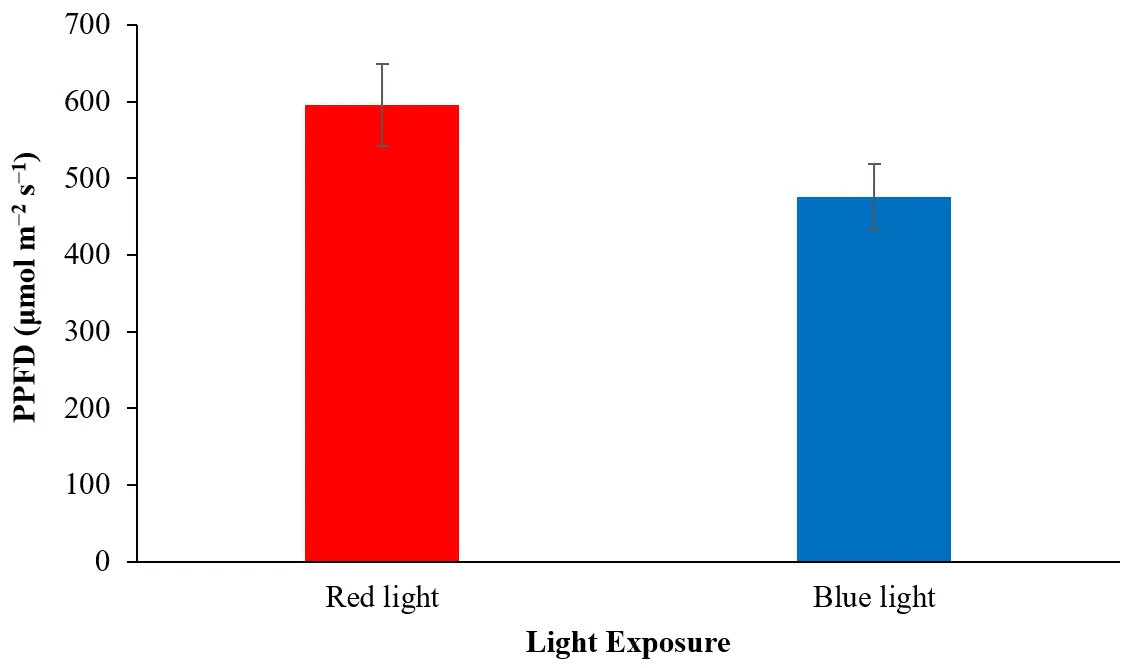
Figure 5. Optical quantum flux (PPFD) of H. lacustris cultures under red and blue light exposure (n = 2).
3.4. Ozone and Light Exposure
A combination of ozone and light exposure was applied to the algae, and the resulting OD680 values and biomass concentrations were measured, as shown in Figure 6a,b, respectively. The OD680 values measured on the first day were similar, but by day eight, algae grown under red light showed higher growth than those under blue light, with or without ozone. However, exposure to ozone gas improves algal growth under red or blue light compared to conditions with no ozone pumped into the system. For red light, the OD680 values were 0.396 ± 0.00283 with ozone and 0.370 ± 0.00141 without, while for blue light, the OD680 values were 0.349 with ozone ± 0.00141 and 0.328 ± 0.00212 without. Similar to the results for OD680, algae grown under red light show greater growth on day eight as measured by biomass concentration than blue light, with or without exposure to ozone. On the other hand, exposure to ozone improves algal biomass under red or blue light compared to conditions with no ozone pumped into the system. For algae grown under red light, the final biomass was 0.250 ± 0.00566 g/L with ozone exposure and 0.233 ± 0.00212 g/L without, while for blue light, the final biomass was 0.216 ± 0.000707 g/L with ozone exposure and 0.190 ± 0.00141 g/L without.
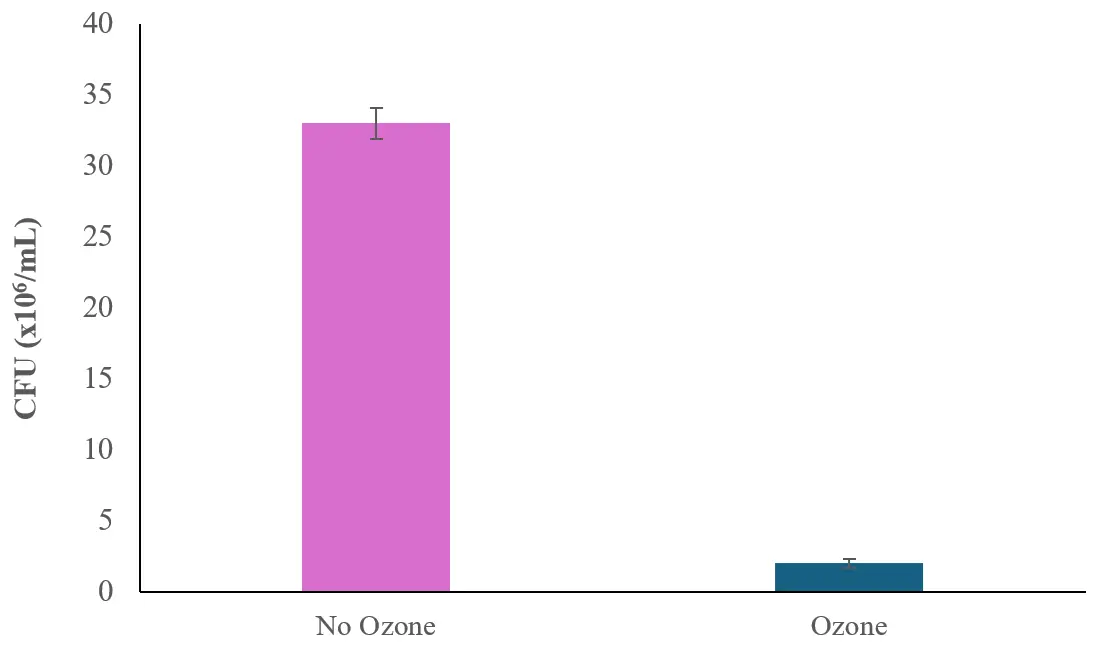
To detect bacterial growth in the cultures, 100 µL of the algal cultures grown either with or without ozone was then diluted and inoculated on Nutrient Agar plates, and the CFUs of bacteria growing on the diluted algae solution of 100,000× were observed after one day of incubation. As can be seen in Figure 7, it was found that ozone caused a large bactericidal effect in H. lacustris cultures as the bacterial number dropped from 33 × 106 ± 1.1 × 106 CFU/mL to 2 × 106 ± 0.3 × 106 CFU/mL.
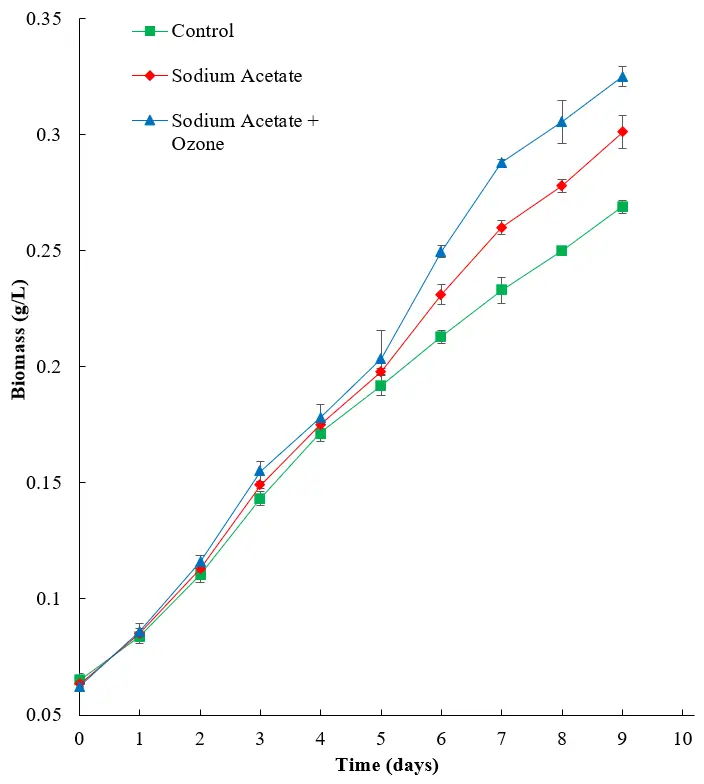
As expected, the addition of sodium acetate to the growth media allowed the algae to use the heterotrophic mode, which therefore showed improved growth compared to the negative control (Figure 8). The addition of ozone improved overall biomass production to 0.325 ± 0.00424 g/L for the sodium acetate and ozone group compared to the sodium acetate group alone (0.301 ± 0.000707 g/L), and both showed a notable improvement compared to the negative control (0.269 ± 0.00283 g/L), likely as a result of inhibited bacterial growth, allowing improved algal proliferation.
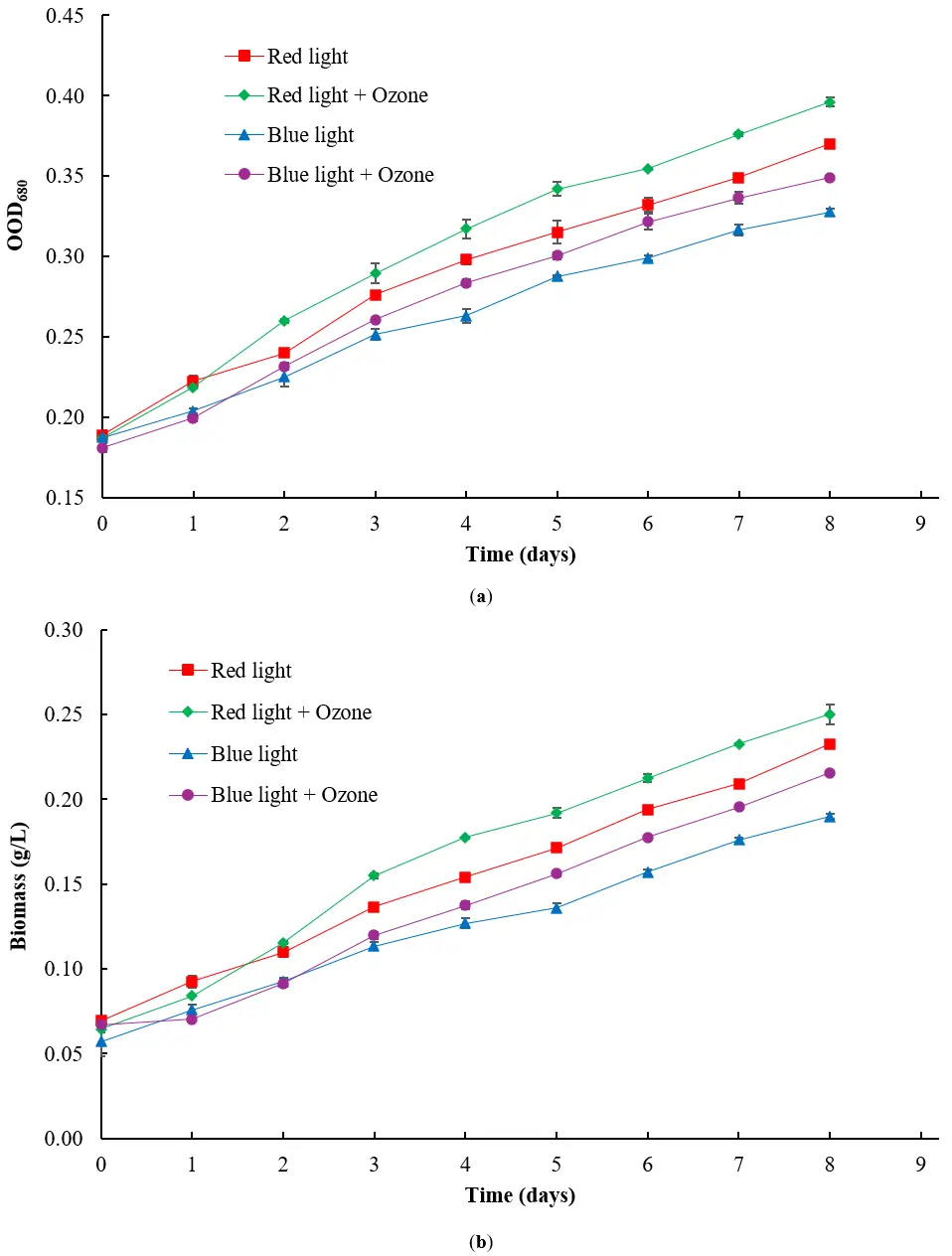
Figure 6. Effect of ozone on (a) OD680 and (b) biomass (in g/L) of H. lacustris with exposure to red or blue light (n = 2).
4. Discussion
The ozone concentration of 0.40 mg/L greatly promoted the growth of H. lacustris, while the ozone concentrations of 4.00 mg/L and 40.00 mg/L had different degrees of inhibition on its growth. The reason for this result may be that, above a certain concentration, ozone has an inhibitory effect on both bacteria and algae. The concentration of 0.40 mg/L ozone is too low to inhibit the growth of bacteria in the flask, while the 40.00 mg/L concentration of ozone can not only kill bacteria, but also inhibit the algal growth, resulting in a gradual decrease in the OD value. In addition, regardless of the presence of ozone, the growth curve of H. lacustris reaches the stationary phase flat on the seventh day of culture, indicating that the overall growth cycle is unaffected. Ozone flow rates of 15 mL/h, 30 mL/h, or 45 mL/h were also examined for their effect on algal growth. The higher the rate, the more obvious the effect of ozone, and the effect of ozone is strongest when the flow rate is 45 mL/h, but it is not significant compared to other flow rates.
Axenic cultures of H. lacustris are commonly established using antibiotic treatment, which has been examined for a range of antibiotics, and the most effective were found to be chloramphenicol, ampicillin, and penicillin [16]. For chloramphenicol, a 1.40-fold increase in cell number was observed, while for ampicillin, it was a 2.56-fold increase. For penicillin, the cell number increased by 3.59-fold. In our system, the addition of 0.40 mg/L ozone caused a ~1.38-fold increase in OD680, compared to the untreated control. Further, the addition of 0.08 mg/L ozone to cells cultivated under red or blue light resulted in a ~1.07-fold or a ~1.06-fold change, respectively, in OD680 values compared to the cells grown under the same light without ozone. A disadvantage of the antibiotic approach compared to the ozone treatment includes the higher cost of the antibiotics, especially given the relatively high concentrations required for improving algae growth, which were 500 mg/L for penicillin and 250 mg/L for ampicillin and chloramphenicol. Additionally, antibiotic usage in industrial algal cultivation can incur undesirable environmental side effects, such as the spread of antibiotic resistance genes.
Another methodology employed for the establishment of axenic microalgal cultures that involved the generation of ROS was anoxia combined with Rose Bengal, a dye that produces ROS using a photosensitive mechanism [33]. Complete decontamination (i.e., pure axenic cultures) was observed at a concentration of 3 µM Rose Bengal for four microalgal species (Tetradesmus obliquus, Desmodesmus armatus, Chlorella vulgaris, and Nannochloropsis limnetica). Notably, while more effective at decontaminating the cultures compared to antibiotics, the lack of oxygen was also equivalently harmful to the algae. By contrast, our ozone experiments showed that low doses of ozone (<4.00 mg/L) could inhibit bacterial growth without decreasing the algal growth or final biomass.
Hongxia et al. used chlorine dioxide to treat several pathogens, including the bacterium Escherichia coli, the fungus Monilinia fructicola, and the yeast Kazachstania exigua [34]. With the increase of chlorine dioxide concentration, the survival rate of bacteria gradually decreased, and chlorine dioxide had the strongest inhibitory effect on M. fructicola among the species. At a 0.1 mg/L concentration of chlorine dioxide, M. fructicola has the lowest survival rate of 1.6%. The inhibition effect on E. coli was the weakest, with a survival rate of about 2.8% at 0.1 mg/L, followed by K. exigua, with a survival rate of about 2.2%. Although the inhibitory effect of chlorine dioxide on all three organisms is stronger than the antibacterial effect of ozone in our experiment, it should also be noted that we observed the effect on algae to be minimal, while chlorine dioxide has been also used for its algicidal activity [35]. This may make chlorine dioxide less attractive as a potential treatment method for algal culture.
5. Conclusions
This study used ozone gas to inhibit bacterial growth during mixotrophic cultivation of H. lacustris. Ozone at 0.4 mg/L promoted the growth of H. lacustris by inhibiting bacterial growth, while ozone at 4.0 mg/L and 40.0 mg/L had inhibitory effects on the growth of the algae itself. The appropriate ozone concentration, therefore, could not only kill the bacteria in the culture medium but also have minimal effect on the algal growth. The flow rate of the metering pump had little impact on the effectiveness of ozone inhibition at all three speeds and concentrations examined. Red light was shown to be the best source of illumination for both strains of H. lacustris, followed by blue light, although the overall change caused by ozone is small. A combination of the changes in light exposure and addition of ozone improved the growth of the algae, with the algae grown under red light and with ozone pumped into the culture exhibiting the highest growth. To test the effect of ozone in mixotrophic culture, sodium acetate and ozone both increased the growth of H. lacustris and showed an additive effect. This research demonstrated the potential use of ozone for mixotrophic cultivation of H. lacustris in combination with cultivation under red light. These findings may therefore be useful as a potential cost-effective alternative to the addition of antibiotics to establish an axenic environment in the biosynthesis of astaxanthin or other algal bioproducts.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/1075, Figure S1: Contamination of nutrient agar plates (a) inoculated with algal culture with ozone introduced (diluted 10,000×) and (b) inoculated with control algal culture (diluted 10,000×); Table S1: Experimental data corresponding to Figure 3. Effect of different sources of illumination on the OD680 value for H. lacustris culture; Table S2: Experimental data corresponding to Figure 4. Effects of different light sources on the biomass of H. lacustris; Table S3: Experimental data corresponding to Figure 6a. Effect of ozone on OD680 of H. lacustris with exposure to red or blue light; Table S4: Experimental data corresponding to Figure 6b. Effect of ozone on biomass (in g/L) of H. lacustris with exposure to red or blue light; Table S5: Experimental data corresponding to Figure 8. Effects of sodium acetate and ozone on the biomass of H. lacustris.
Acknowledgments
The authors would like to thank Harbin Engineering University and the College of Agricultural Sciences of Panzhihua University for their support.
Author Contributions
A.A.-E.-A.: Conceptualization, Formal analysis, Methodology, Visualization, Writing—original draft, Writing—review & editing; Y.S. (Yang Su): Formal analysis, Investigation, Methodology, Validation, Writing—original draft; Y.S. (Yan Shang): Formal analysis, Investigation, Methodology, Visualization, Writing—original draft; S.N.A.: Conceptualization, Funding acquisition, Methodology, Visualization, Writing—review & editing; J.L.: Conceptualization, Investigation, Methodology, Project administration, Supervision, Visualization, Writing—review & editing.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available from the corresponding author upon reasonable request.
Funding
This research received no external funding.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper
References
-
Chang MX, Xiong F. Astaxanthin and its Effects in Inflammatory Responses and Inflammation-Associated Diseases: Recent Advances and Future Directions. Molecules 2020, 25, 5342. DOI:10.3390/molecules25225342 [Google Scholar]
-
Nair A, Ahirwar A, Singh S, Lodhi R, Lodhi A, Rai A, et al. Astaxanthin as a King of Ketocarotenoids: Structure, Synthesis, Accumulation, Bioavailability and Antioxidant Properties. Mar. Drugs 2023, 21, 176. DOI:10.3390/md21030176 [Google Scholar]
-
Medoro A, Davinelli S, Milella L, Willcox B, Allsopp R, Scapagnini G, et al. Dietary Astaxanthin: A Promising Antioxidant and Anti-Inflammatory Agent for Brain Aging and Adult Neurogenesis. Mar. Drugs 2023, 21, 643. DOI:10.3390/md21120643 [Google Scholar]
-
Snell TW, Carberry J. Astaxanthin Bioactivity Is Determined by Stereoisomer Composition and Extraction Method. Nutrients 2022, 14, 1522. DOI:10.3390/nu14071522 [Google Scholar]
-
Abdur Razzak S, Bahar K, Islam KMO, Haniffa AK, Faruque MO, Hossain SMZ, et al. Microalgae cultivation in photobioreactors: Sustainable solutions for a greener future. Green Chem. Eng. 2024, 5, 418–439. DOI:10.1016/j.gce.2023.10.004 [Google Scholar]
-
Liaqat F, Khazi MI, Bahadar A, He L, Aslam A, Liaquat R, et al. Mixotrophic cultivation of microalgae for carotenoid production. Rev. Aquac. 2023, 15, 35–61. DOI:10.1111/raq.12700 [Google Scholar]
-
Wen X, Wang Z, Ding Y, Geng Y, Li Y. Enhancing the production of astaxanthin by mixotrophic cultivation of Haematococcus pluvialis in open raceway ponds. Aquacult. Int. 2020, 28, 625–638. DOI:10.1007/s10499-019-00483-2 [Google Scholar]
-
Khazi MI, Liaqat F, Gu W, Mohamed B, Zhu D, Li J. Astaxanthin production from the microalga Haematococcus lacustris with a dual substrate mixotrophy strategy. Biotechnol. J. 2023, 18, 2300095. DOI:10.1002/biot.202300095 [Google Scholar]
-
Lutzu GA, Concas A, Damergi E, Chen L, Zhang W, Liu T. Production of Carotenoids and Astaxanthin from Haematococcus pluvialis Cultivated Under Mixotrophy Using Brewery Wastewater: Effect of Light Intensity and Cultivation Time. Appl. Sci. 2024, 14, 9704. DOI:10.3390/app14219704 [Google Scholar]
-
Tambat VS, Patel AK, Singhania RR, Vadrale AP, Tiwari A, Chen CW, et al. Sustainable mixotrophic microalgae refinery of astaxanthin and lipid from Chlorella zofingiensis. Bioresour. Technol. 2023, 387, 129635. DOI:10.1016/j.biortech.2023.129635 [Google Scholar]
-
Dawidziuk A, Popiel D, Luboinska M, Grzebyk M, Wisniewski M, Koczyk G. Assessing contamination of microalgal astaxanthin producer Haematococcus cultures with high-resolution melting curve analysis. J. Appl. Genet. 2017, 58, 277–285. DOI:10.1007/s13353-016-0378-x [Google Scholar]
-
Laezza C, Salbitani G, Carfagna S. Fungal Contamination in Microalgal Cultivation: Biological and Biotechnological Aspects of Fungi-Microalgae Interaction. J. Fungi 2022, 8, 1099. DOI:10.3390/jof8101099 [Google Scholar]
-
Vu CHT, Lee HG, Chang YK, Oh HM. Axenic Cultures for Microalgal Biotechnology: Establishment, Assessment, Maintenance, and Applications. Biotechnol. Adv. 2018, 36, 380–396. DOI:10.1016/j.biotechadv.2017.12.018 [Google Scholar]
-
Molina D, de Carvalho JC, Júnior AIM, Faulds C, Bertrand E, Soccol CR. Biological contamination and its chemical control in microalgal mass cultures. Appl. Microbiol. Biotechnol. 2019, 103, 9345–9358. DOI:10.1007/s00253-019-10193-7 [Google Scholar]
-
Wang L, Yang F, Chen H, Fan Z, Zhou Y, Lu J, et al. Antimicrobial Cocktails to Control Bacterial and Fungal Contamination in Chlamydomonas reinhardtii Cultures. BioTechniques 2016, 60, 145–149. DOI:10.2144/000114392 [Google Scholar]
-
Rayamajhi V, Byeon H, An Y, Kim T, Lee J, Lee J, et al. Screening and Selection of Antibiotics for Enhanced Production of Astaxanthin by Haematococcus lacustris. Life 2024, 14, 977. DOI:10.3390/life14080977 [Google Scholar]
-
Asatryan A, Gunasekaran M, Boussiba S, Zarka A. Establishing and validating axenic cultures of the microalga Haematococcus lacustris (Chlorophyceae). Appl. Phycol. 2022, 3, 82–97. DOI:10.1080/26388081.2022.2061871 [Google Scholar]
-
Park S, Van Ginkel SW, Pradeep P, Igou T, Yi C, Snell T, et al. The Selective Use of Hypochlorite to Prevent Pond Crashes for Algae-Biofuel Production. Water Environ. Res. 2016, 88, 70–78. DOI:10.2175/106143015X14362865227670 [Google Scholar]
-
Jeong SH, Kim W, Kwon JH. Development of a new sterilization method for microalgae media using calcium hypochlorite as the sterilant. Bioprocess. Biosyst. Eng. 2024, 47, 393–401. DOI:10.1007/s00449-024-02971-z [Google Scholar]
-
Nam K, Shin WS, Jeong BR, Park MS, Yang JW, Kwon JH. Use of a triiodide resin for isolation of axenic cultures of microalgal Nannochloropsis gaditana. Bioresour. Technol. 2015, 191, 391–394. DOI:10.1016/j.biortech.2015.03.082 [Google Scholar]
-
Abd-El-Aziz A, Elnagdy SM, Han J, Mihelič R, Wang X, Agathos SN, et al. Bacteria-Microalgae interactions from an evolutionary perspective and their biotechnological significance. Biotechnol. Adv. 2025, 82, 108591. DOI:10.1016/j.biotechadv.2025.108591 [Google Scholar]
-
Krasensky J, Carmody M, Sierla M, Kangasjärvi J. Ozone and Reactive Oxygen Species. In Encyclopedia of Life Sciences; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2017; pp. 1–9. [Google Scholar]
-
Travagli V, Iorio EL. The Biological and Molecular Action of Ozone and Its Derivatives: State-of-the-Art, Enhanced Scenarios, and Quality Insights. Int. J. Mol. Sci. 2023, 24, 8465. DOI:10.3390/ijms24108465 [Google Scholar]
-
Xue W, Macleod J, Blaxland J. The Use of Ozone Technology to Control Microorganism Growth, Enhance Food Safety and Extend Shelf Life: A Promising Food Decontamination Technology. Foods 2023, 12, 814. DOI:10.3390/foods12040814 [Google Scholar]
-
Rangel K, Cabral FO, Lechuga GC, Carvalho JPRS, Villas-Bôas MHS, Midlej V, et al. Potent Activity of a High Concentration of Chemical Ozone against Antibiotic-Resistant Bacteria. Molecules 2022, 27, 3998. DOI:10.3390/molecules27133998 [Google Scholar]
-
Yang Z, Wang Y, Lukwambe B, Nicholaus R, Yang W, Zhu J, et al. Using ozone nanobubbles, and microalgae to promote the removal of nutrients from aquaculture wastewater: Insights from the changes of microbiomes. Environ. Res. 2024, 257, 119349. DOI:10.1016/j.envres.2024.119349 [Google Scholar]
-
Langbehn RK, Michels C, Soares HM. Antibiotics in wastewater: From its occurrence to the biological removal by environmentally conscious technologies. Environ. Pollut. 2021, 275, 116603. DOI:10.1016/j.envpol.2021.116603 [Google Scholar]
-
Zhang J, Liu F, Wang Q, Gong Q, Gao X. Effect of Light Wavelength on Biomass, Growth, Photosynthesis and Pigment Content of Emiliania huxleyi (Isochrysidales, Cocco-Lithophyceae). J. Mar. Sci. Eng. 2023, 11, 456. DOI:10.3390/jmse11020456 [Google Scholar]
-
Guedes VC, Palma GM, Horta ACL. An evaluation of light wavelengths, intensity and control for the production of microalgae in photobioreactors: A review. Braz. J. Chem. Eng. 2024, 41, 1077–1090. DOI:10.1007/s43153-023-00388-x [Google Scholar]
-
Pereira S, Otero A. Haematococcus pluvialis bioprocess optimization: Effect of light quality, temperature and irradiance on growth, pigment content and photosynthetic response. Algal Res. 2020, 51, 102027. DOI:10.1016/j.algal.2020.102027 [Google Scholar]
-
United States Food and Drug Administration. Beverages: Bottled Water (21 CFR Parts 103, 129, 165, and 184); United States Food and Drug Administration: Silver Spring, MD, USA, 1995. [Google Scholar]
-
Cheng J, Fan W, Zheng L. Development of a mixotrophic cultivation strategy for simultaneous improvement of biomass and photosynthetic efficiency in freshwater microalga Scenedesmus obliquus by adding appropriate concentration of sodium acetate. Biochem. Eng. J. 2021, 176, 108177. DOI:10.1016/j.bej.2021.108177 [Google Scholar]
-
Iyer A, Monissen M, Ma Q, Osborne M, Schaedig E, Modin O, et al. Axenisation of oleaginous microalgal cultures via anoxic photosensitisation. Algal Res. 2025, 86, 103926. DOI:10.1016/j.algal.2025.103926 [Google Scholar]
-
Xin H, Yao Q, Zhang Y, Liao J, Dong J, Yu Q, et al. Antibacterial effects of chlorine dioxide on Monilinia fructicola, yeast and Escherichia coli. Chin. J. Antibiot. 2023, 48, 894–899. Available online: http://zgkss.com.cn/CN/article/downloadArticleFile.do?attachType=PDF&id=9562 (accessed on 02 June 2026).
-
He R, Zeng Y, Zhao S, Zhang J, He P, Liu J. Use of citric acid-activated chlorine dioxide to control Ulva prolifera. Mar. Pollut. Bull. 2023, 194, 115357. DOI:10.1016/j.marpolbul.2023.115357 [Google Scholar]