Utilization of Post-Mortem Oocytes for In Vitro Embryo Production for Preservation of Adapted High Genetic Value Germplasm of Jersey Cattle

Utilization of Post-Mortem Oocytes for In Vitro Embryo Production for Preservation of Adapted High Genetic Value Germplasm of Jersey Cattle
Syed Mohmad Shah
Abrar A. Malik
*
 Arif A. Pandit
Faheem A. Bhat
Syed H. Yaqoob
Sofi Imran Ul Umar
Rameez A. Dar
Anees A. Shah
Riaz A. Shah
Arif A. Pandit
Faheem A. Bhat
Syed H. Yaqoob
Sofi Imran Ul Umar
Rameez A. Dar
Anees A. Shah
Riaz A. Shah
Received: 15 April 2026 Revised: 20 May 2026 Accepted: 16 June 2026 Published: 22 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
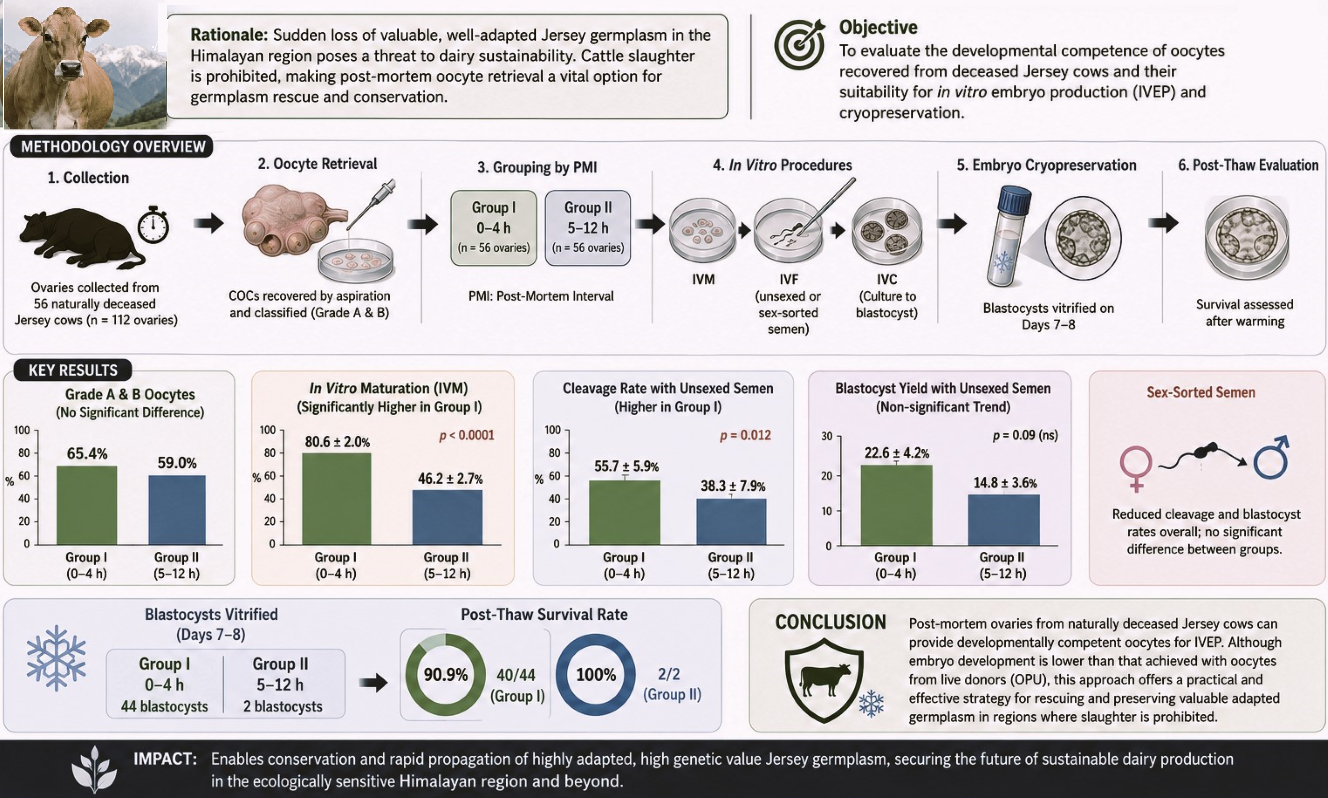
Graphical Abstract
1. Introduction
Jersey and Holstein Friesian crossbreds form the main milch herd in the Indian subcontinent, more so in the ecologically sensitive Himalayan Kashmir region. Characterized by a modified humid continental climate under the Köppen-Geiger classification system, the region experiences four distinct macro-seasons: a cold, severe winter (December to February) with mean temperatures frequently dropping below 0 °C; a wet, transitional spring (March to May); a temperate, relatively dry summer (June to August) with maximum temperatures averaging between 25 °C and 32 °C; and a dry, cooling autumn (September to November). The average annual precipitation across the valley floors ranges between 700 mm and 1200 mm, varying substantially with localized changes in elevation.
These crossbreds raised through decades of cross-breeding of local nondescript cattle using artificial insemination have shown very good adaptation to the temperate climatic conditions of the region, forming a valuable genetic resource and paving the way for the establishment of the purebred replacement stock, highly adapted to the region while harboring the exotic genome. With recent trends shifting towards procurement of the pure-bred germplasm from other states of the country, this valuable germplasm faces neglect from the emerging dairy entrepreneurs who barely acknowledge their pre-and post-adaptation advantage. These stakeholders often face challenges, including decreased productivity and high mortality, which lead to decreased motivation in dairy entrepreneurship. After an outburst of a huge number of such cases and reports, it was concluded that these purebreds imported from other countries fail to adapt to the climatic conditions of the Kashmir valley. The breeding policy of the Union Territory (UT) of Jammu & Kashmir also aims for up to 75% of the exotic inheritance in the temperate regions of the Kashmir valley and up to 50% only in the subtropical plains of the Jammu region [1] (J&K state Livestock Breeding Policy, 2019). With a clear preference for cross-bred cattle in J&K UT, the cross-bred cattle population has already cross 50% mark and is further increasing due to increasing artificial inseminations using exotic germplasm [2] (20th Livestock Census). The average national mortality rate of 4–20% in dairy cattle due to sudden disease outbreaks, pandemics, accidents, and other causes, poses a great threat to this upgraded and highly adapted germplasm [3] (Annual report 2023–2024, Dept. of Animal husbandry and dairying, Government of India), thus compelling the need for technological interventions for conservation and rapid propagation of this valuable germplasm. The application of assisted reproductive techniques (ARTs) offers a reliable solution not only for conservation and rapid multiplication of this germplasm but also for future applications in conservation of other endangered/threatened species, because 27% of the 5969 mammalian species evaluated are facing extinction [4] (IUCN Red List of Threatened Species, 2022). The current state of extinction is estimated to be 1000 times the normal rate of extinction, primarily attributed to deforestation, habitat loss, human-wildlife conflicts, hunting, and global warming [5]. The application of such ARTs, such as oocyte aspiration followed by in vitro maturation, in vitro fertilization, and in vitro embryo production, provides a valuable tool for not only germplasm conservation but also for the rapid development of a superior replacement dairy herd, which is essentially needed in the Indian subcontinent. This assumes more importance in regions like India, where there are religious prohibitions on cattle slaughter, which leads to loss of valuable germplasm following the sudden death of elite dairy cows, thereby presenting a critical bottleneck for breeding programs. Such restrictions eliminate access to abattoir-sourced ovaries, which remain the most widely used source of oocytes for in vitro embryo production (IVEP) worldwide, without involving the skill and resource-demanding ovum-pick-up technical setup from the live animals. Under these constraints, ovaries retrieved post-mortem from naturally deceased or accident-related animals provide a pragmatic emergency source of oocytes for germplasm conservation. It will also ameliorate the restrictions on female gamete resource availability for research and thus enhance reliance on female gamete rescue from post-mortem animals, exacerbating the need for further refinement of the techniques involved. Developmental potential of post-mortem oocytes is highly sensitive to handling conditions and, critically, to the post-mortem interval (PMI). Prolonged PMI exacerbates ischemic and oxidative injury, spindle disruption, and cytoplasmic deterioration, thereby reducing oocyte competence [6,7,8]. Prior studies in ruminants have confirmed that delayed recovery compromises maturation and embryo development, but systematic evaluation in cattle under slaughter-ban conditions remains limited. The use of sex-sorted semen further complicates efficiency. Although it enables female-biased calf production in dairy herds, its application is consistently associated with reduced cleavage, blastocyst development, and pregnancy rates [9,10,11] Optimization strategies may alleviate, but not fully eliminate, these disadvantages [12]. Consequently, the objective of this study was to evaluate the developmental competence and post-thaw cryosurvival of oocytes salvaged from naturally deceased Jersey cattle under slaughter-ban constraints, comparing a rapid response <6 h versus an extended (6–12 h) post-mortem interval, utilizing both conventional and sex-sorted semen.
2. Materials and Methods
All procedures complied with institutional and national guidelines for animal research. Ovaries were obtained only from cows that had died due to non-zoonotic, non-notifiable diseases within 10–12 h of death, and informed owner consent was obtained for the same. All chemicals and media were of cell culture grade and obtained from Sigma Aldrich unless otherwise specified.
2.1. Source of Ovaries and Transportation
Ovaries were recovered post-mortem from Jersey cows that had died from accidental causes or non-infectious and non-notifiable diseases. Animals accessible within 10–12 h of death were included in the study, preferably on the post-mortem table. Immediately after removal, ovaries were rinsed in sterile transport medium consisting of 0.9% NaCl supplemented with 400 IU/mL penicillin and 200 µg/mL streptomycin as described earlier [13]. Ovaries were transported at 30–35 °C in a pre-warmed sterile thermos flask containing the medium and transported to the laboratory within 1–2 h of collection. A total of 56 cows were included, yielding 112 ovaries. Ovaries from each cow were processed separately to allow evaluation of cow effect; however, data were pooled within each PMI group for statistical comparison.
2.2. Oocyte Retrieval and Grading
Visible follicles (2–10 mm) were aspirated individually using an 18-G needle attached to a 10 mL syringe (BD Falcon). Aspiration medium consisted of TCM-199 supplemented with 6 mg/mL BSA and 5000 IU/mL Heparin. Follicular contents were pooled per ovary, allowed to sediment for 10 min, and examined under a stereomicroscope. Cumulus–oocyte complexes (COCs) were washed three times in handling medium and graded according to cumulus investment and cytoplasmic appearance [14]. Grade A and B Complete cumulus cover with several compact layers and a few slightly loose cell layers; oocytes were selected for in vitro maturation (IVM), while Grades C (partial cumulus cover and loose cell layers) and D (No cumulus cover and COCs showing clear signs of atresia) were excluded.
2.3. In Vitro Maturation (IVM)
Selected Grade A&B oocytes were washed three times in serum-free BO-IVM medium (Cat. No. I2521). Groups of 15–20 COCs were cultured in 100 µL droplets of BO-IVM medium under mineral oil at 38.5 °C in a humidified incubator (5% CO2, 21% O2, balance N2) for 22 h. Cumulus expansion was used as the primary indicator of maturation, and nuclear maturation was confirmed by polar body extrusion following partial denudation [15,16].
2.4. Semen Preparation
Frozen straws of conventional (unsexed) and sex-sorted semen were procured from Genus ABS Pune, India. Frozen semen straws from the same bull were used. For performing the IVF, frozen semen straws were thawed at 37 °C for 30 s and processed by two-step centrifugation at 300× g for 5 min each in BO-Semen Prep medium (IVF Bioscience, Falmouth, UK). The final pellet was resuspended to yield a working concentration of 2.0 × 106 sperm/mL. Sperm concentration was measured using the hemocytometer method (Neubauer Chamber).
2.5. In Vitro Fertilization (IVF)
Groups of mature oocytes (25 oocytes per group) were washed in BO-IVF medium (IVF Bioscience, Falmouth, UK) and transferred into 100 µL droplets under mineral oil. Sperm suspensions were added to achieve a final sperm to oocyte ratio of approximately 2 × 104 sperm per oocyte. Coincubation was carried out at 38.5 °C in 5% CO2 for 16–18 h. A subset of 10 oocytes per group was denuded and examined for polar body extrusion to validate nuclear maturation.
2.6. Embryo Culture and Quality Assessment
Presumptive zygotes were denuded by brief exposure to 0.5 mg/mL hyaluronidase for 2–3 min and washed in BO-IVC medium (IVF Bioscience, Falmouth, UK). Zygotes were cultured in 50 µL droplets (5 µL per zygote) of BO-IVC medium under mineral oil, equilibrated in 5% O2, 5% CO2, 90% N2 at 38.5 °C [17]. Cleavage was assessed at 48 h post-insemination. Blastocyst development was evaluated on Days 7–8 and classified according to IETS criteria. A subset of blastocysts was stained with Hoechst 33342 for cell number evaluation. Fluorescent images were captured using a Leica DMi8 inverted microscope equipped with the Leica Application Suite X (LAS X) software (Leica Microsystems, Wetzlar, Germany), and cell numbers were quantified using ImageJ software (version 1.54, National Institutes of Health, Bethesda, MD, USA).
2.7. Vitrification and Warming of Blastocysts
Expanded, Day 7–8 IETS Grade 1–2 blastocysts, with intact TE were vitrified using the BO Vitri-Cool kit (IVF Bioscience, Falmouth, UK) following the manufacturer’s protocol. Embryos were equilibrated for a total of 10 min, loaded onto vitrification devices (0.25 cc French Straw, IMV Technologies), and plunged into liquid nitrogen. Warming was performed using the BO Vitri-Warm (IVF Bioscience, Falmouth, UK) kit by sequentially diluting cryoprotectants at 37 °C. Post-warming recovery was assessed after 3–4 h of incubation at 38.5 °C, with survival defined as blastocoel re-expansion and intact morphology.
2.8. Statistical Analysis
Data were analyzed using Systat software (version 27.0; SPSS Inc., Chicago, IL, USA). Percentage data were arcsine-transformed before analysis. Oocyte recovery, IVM rates, cleavage, and blastocyst development were compared between groups using one-way ANOVA. Fisher’s least significant difference (LSD) test was applied for multiple comparisons. Cow identity was included as a random effect to account for clustering. Results are presented as mean ± SEM, and p < 0.05 was considered significant.
3. Results
3.1. Oocyte Retrieval and Morphological Assessment
A total of 112 ovaries were collected post-mortem from 56 Jersey cows. Ovaries were classified into Group I: PMI up to 6 h (n = 58 ovaries) and Group II: PMI up to 6.1–12 h (n = 54 ovaries). Overall, 847 COCs were recovered: 442 from Group I (7.6 ± 0.1/ovary) and 405 from Group II (7.5 ± 0.1/ovary). The difference in recovery was not significant (p = 0.301). Morphological assessment showed that 65.4% (289/442) of oocytes in Group I and 59.0% (239/405) in Group II were Grade A/B, while 34.6% and 41.0% were Grade C/D, respectively (p = 0.238) (Table 1). In total, 528 Grade A/B oocytes (289 from Group I, 239 from Group II) were selected for IVM.
Table 1. Recovery and morphological grading of COCs from post-mortem Jersey cow ovaries collected at 0–6 h (Group I) or 6.1–12 h (Group II) after death.
|
Parameter |
PMI |
Number of Ovaries |
Number of COCs |
Mean COCs per Ovary (±SE) |
Good Quality COCs (Grade A/B) |
Poor Quality COCs (Grade C/D) |
|---|---|---|---|---|---|---|
|
Group I |
Up to 6 h |
58 |
442 |
(7.62 a ± 0.1) |
289 (65.4%) |
153 (34.6%) |
|
Group II |
6–12 h |
54 |
405 |
(7.50 a ± 0.1) |
239 (59.0%) |
166 (41.0%) |
|
Total |
112 |
847 |
(7.56 ± 0.07) |
528 (62.3%) |
319 (37.7%) |
Values are presented as mean ± standard error (SE). Percentages represent the proportion of each grade within the total COCs for that group. Data with same superscript do not differ significantly (p > 0.05).
3.2. In Vitro Maturation
Of the 528 Grade A/B oocytes, 344 achieved nuclear maturation as assessed by cumulus expansion and polar body extrusion (Figure 1). Group I showed significantly higher maturation (233/289; 80.6 ± 2.0%) compared with Group II (111/239; 46.2 ± 2.7%; p < 0.0001). Validation by denudation confirmed polar body extrusion in 9/10 oocytes from Group I and 7/10 from Group II (Table 2).
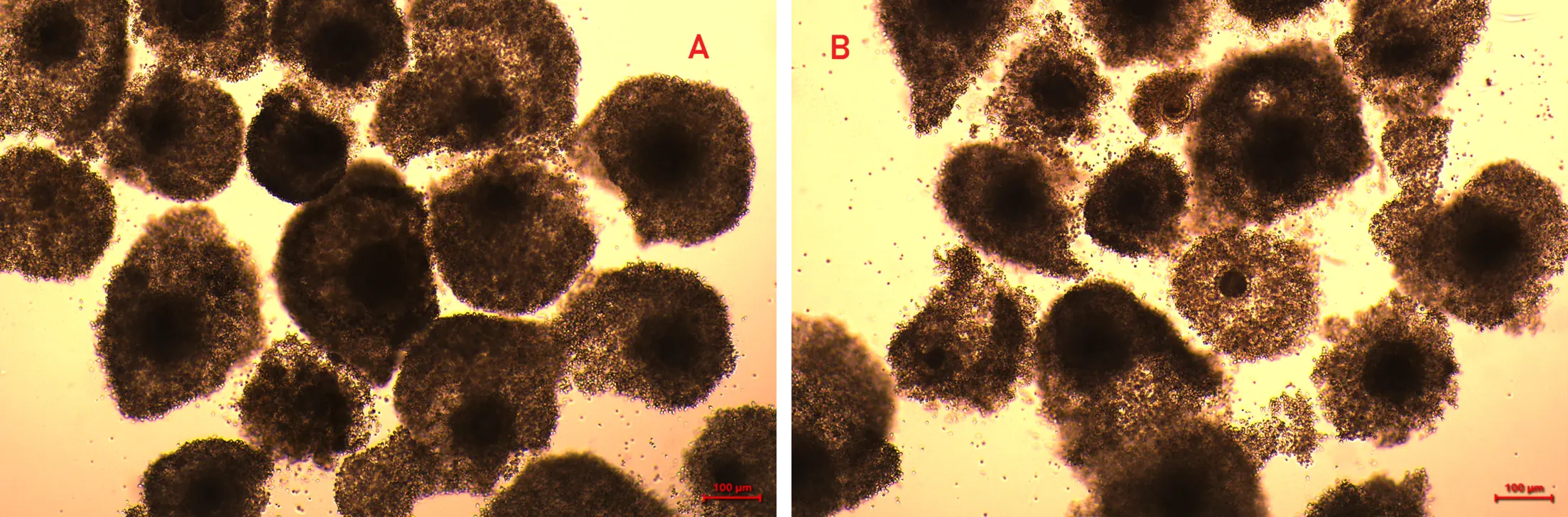
Figure 1. In vitro matured oocytes from post-mortem Jersey cow ovaries. (A) Group I oocytes (0–6 h PMI) showing complete cumulus expansion. (B) Group II oocytes (6.1–12 h PMI) showing reduced cumulus expansion.
Table 2. Nuclear maturation of Grade A/B oocytes retrieved from post-mortem Jersey cow ovaries at different post-mortem intervals.
|
Group |
COCs for IVM (n) |
No. Matured (n/N) |
Maturation Rate (% ± SE) |
Polar Body Extrusion (n/10) |
|---|---|---|---|---|
|
I |
289 |
233/289 |
80.6 ± 2.0 a |
9/10 |
|
II |
239 |
111/239 |
46.2 ± 2.7 b |
7/10 |
Values are presented as mean ± standard error (SEM). Values with different superscripts within a column differ significantly (p < 0.05).
3.3. Embryo Development After IVF with Unsexed and Sex-Sorted Semen
A total of 324 mature oocytes were subjected to IVF: 223 from Group I and 101 from Group II (Table 3), using sexed and conventional semen from the same Jersey bull. The results of cleavage and blastocyst formation (Figure 2) were achieved as:
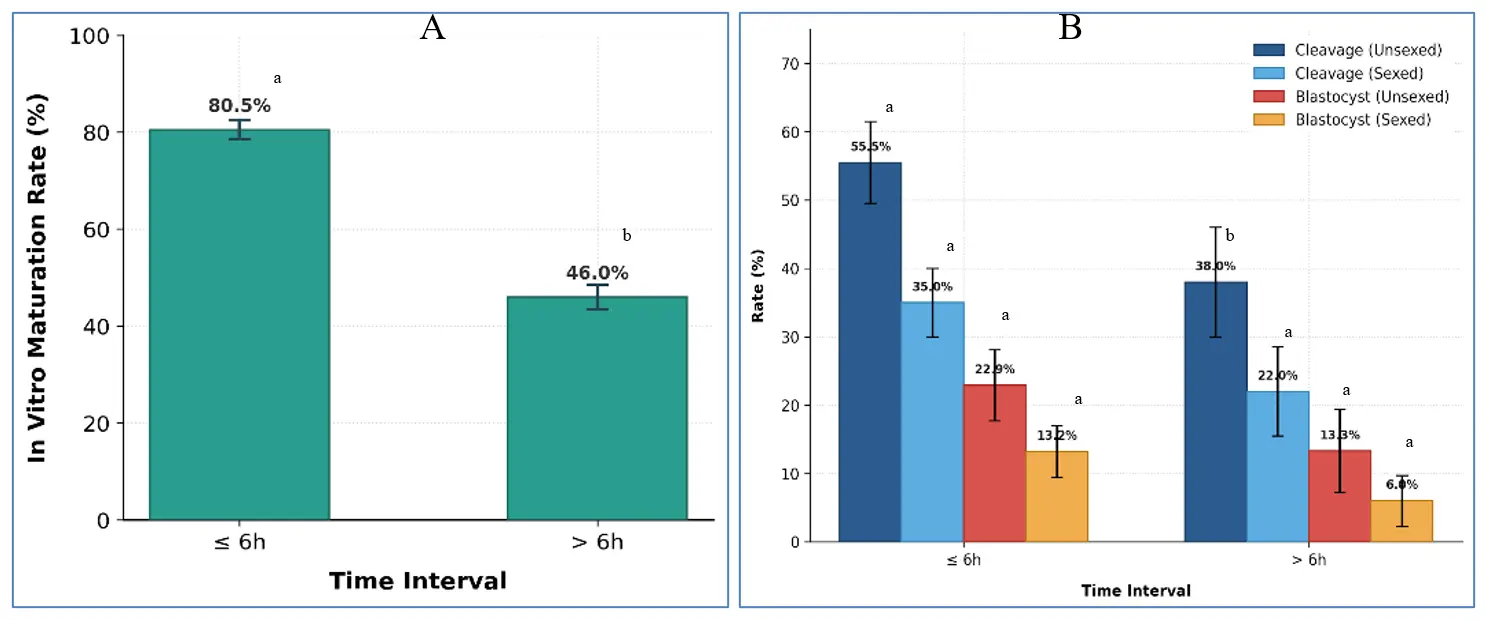
Figure 2. Developmental competence of oocytes retrieved at different post-mortem intervals. (A) IVM rates were significantly higher in Group I compared with Group II (p < 0.0001). (B) Cleavage rates and Blastocyst development rates following IVF with unsexed or sex-sorted semen. Group I showed significantly higher cleavage with unsexed semen (p = 0.012), while sex-sorted differences were not significant. Blastocyst development in Group I showed numerical but not significant advantages. (Bars represent mean ± SEM, Bars that share a common letter are not significantly different from each other. Bars that do not share a common letter are significantly different (p < 0.05)).
Unsexed semen: Cleavage was significantly higher in Group I (124/223; 55.7 ± 5.9%) than in Group II (39/101; 38.3 ± 7.9%; p = 0.012). Blastocyst formation also favoured Group I (51/223; 22.9 ± 5.2%) compared with Group II (13/101; 13.3 ± 6.1%), although the difference was not statistically significant (p = 0.089).
Sex-sorted semen: Cleavage was 39/111 (35.0 ± 5.3%) in Group I and 11/50 (22.0 ± 6.5%) in Group II (p = 0.074). Blastocyst rates were 15/111 (13.2 ± 3.8%) in Group I and 3/50 (6.0 ± 3.7%) in Group II (p = 0.162).
On Day 7–8, the majority of Group I blastocysts were of superior quality, with 63.6% classified as IETS Grade 1, compared with predominantly Grade 2 embryos in Group II. Hoechst staining (Figure 3) confirmed higher mean total cell numbers in Group I blastocysts (138.4 ± 6.7) compared with Group II (102.3 ± 5.9; p < 0.001).
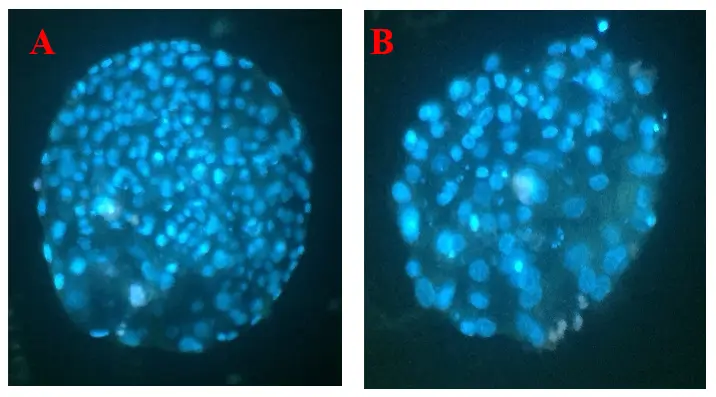
Table 3. Cleavage and blastocyst development rates of oocytes retrieved from post-mortem Jersey cow ovaries after IVF with unsexed or sex-sorted semen.
|
Treatment |
Group |
No. of COCs |
Cleavage Rate (% ± SEM) |
Blastocyst Rate (% ± SEM) |
|---|---|---|---|---|
|
IVF (Unsexed semen) |
I |
112 |
55.7 ± 5.9 a |
22.9 ± 5.2 a |
|
II |
51 |
38.3 ± 7.9 b |
13.3 ± 6.1 a |
|
|
IVF (Sexed semen) |
I |
111 |
35.0 ± 5.3 a |
13.2 ± 3.8 a |
|
II |
50 |
22.0 ± 6.5 a |
6.0 ± 3.7 a |
Data are presented as mean ± SEM. Values with different superscripts within a column differ significantly (p < 0.05).
3.4. Vitrification and Warming
Expanded Day 7–8 IETS Grade 1–2 blastocysts meeting pre-specified quality criteria (IETS Grade 1–2; fully expanded; intact TE) were considered for cryopreservation. From Group I, 44 blastocysts were vitrified; 40/44 (90.9%) survived warming and re-expanded within 3–4 h, with Grade 1 embryos showing 96.4% re-expansion (Table 4). From Group II, although 10 blastocysts were obtained in Group II, only two met vitrification criteria and were cryopreserved. Both survived warming and re-expanded, albeit with inferior morphology compared with Group I. Thus, earlier ovary retrieval increased the number of vitrifiable embryos and was associated with higher post-warming competence (Figure 4).
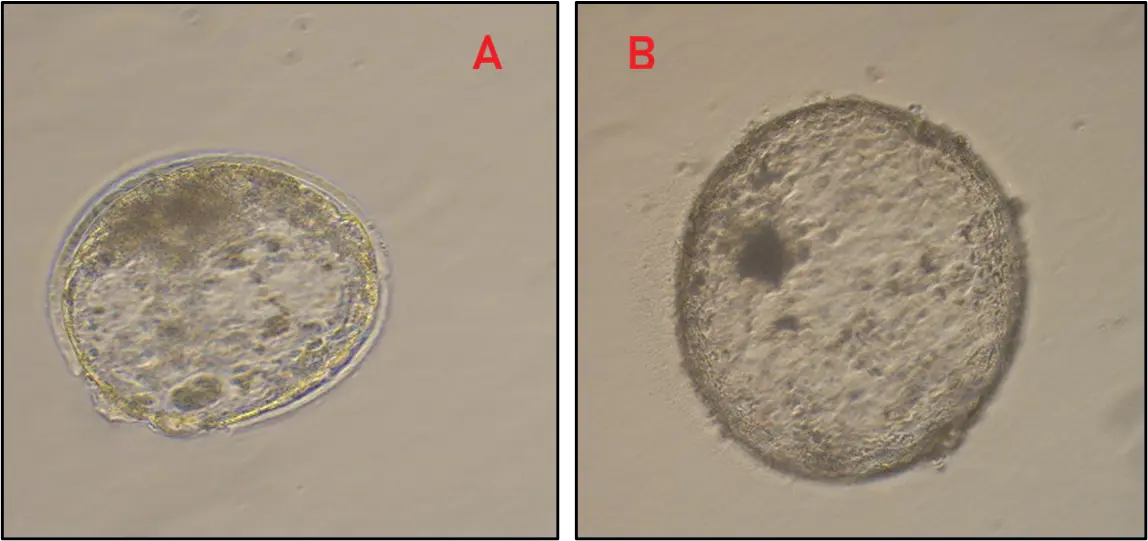
Figure 4. In vitro produced blastocysts after vitrification and warming. Blastocysts displayed (A) hatching, indicative of high cryotolerance. (B) complete blastocoel re-expansion.
Table 4. Vitrification and survival outcomes of Day 7–8 expanded IETS Grade 1–2 blastocysts derived from post-mortem Jersey cow ovaries. Embryos were vitrified if fully expanded and with intact trophectoderm. Survival was assessed by re-expansion and morphological integrity within 3–4 h of warming.
|
Group |
Vitrifiable Embryos (n) |
Survived Warming (n, %) |
|---|---|---|
|
I |
44 |
40 (90.9%) |
|
II |
2 |
2 (100%) |
4. Discussion
This study demonstrates that post-mortem ovaries retrieved from naturally deceased Jersey cows can provide a viable and reliable oocyte source for IVEP. This approach offers a practical framework for rescuing the valuable, climate-adapted germplasm in specific geo-climatic zones, particularly in regions where slaughter bans restrict access to conventional abattoir derived ovaries.
Our key finding is that PMI is a critical determinant of developmental competence. Oocytes retrieved within 6 h post-mortem had significantly higher oocyte maturation, cleavage rates, and embryo quality compared to later recovered oocytes. Although small sample sizes limited our statistical power to detect differences in final blastocyst percentages between the temporal groups, total embryonic cell counts and post-thaw cryosurvival explicitly underscore the superior developmental potential of oocytes salvaged during the early 0–6 h window.
5. Relevance in Slaughter-Ban Settings
In regions like India, where indigenous and/or highly adapted/upgraded dairy germplasm constitutes a critical socioeconomic asset, the rescue of deceased elite animals through ARTs assumes greater importance both for germplasm conservation and for sustainable, reliable livelihood provision to stakeholders. Legislation prohibiting cattle slaughter and thus the inability to access abattoir ovaries restricts the large-scale implementation of IVEP programs. Opportunistic post-mortem retrieval provides a supplementary pathway to preserve genetics that would otherwise be lost, particularly in elite dairy herds where sudden mortality events may erase decades of selective breeding progress. The integration of such approaches into national germplasm conservation and dairy strengthening programs could complement AI and MOET, especially when combined with embryo cryopreservation and sex-sorted semen strategies [18,19].
6. Post-Mortem Interval and Developmental Competence
The marked decline in IVM rates beyond 6 h PMI (80.6% vs. 46.2%) underscores the importance of retrieval timing. Prolonged warm ischemia and oxidative stress after death accelerate cytoplasmic degeneration, spindle disorganization, and maternal RNA degradation, thereby reducing the developmental competence [6,7,17]. Similar time-sensitive effects have been documented in sheep and goats, where extended PMI has been shown to reduce oocyte quality and embryo development [8]. Collectively, these findings suggest that a less than 6 h window represents a biologically critical threshold for germplasm recovery across species.
7. Impact of Sex-Sorted Semen
As anticipated, fertilization with sex-sorted semen reduced cleavage and blastocyst formation across both groups, consistent with systematic reviews and meta-analyses [9,10,20]. While numerical trends favored earlier retrieval, the differences in blastocyst rates were not statistically significant, reflecting both the reduced efficiency of sex-sorted semen and sample size constraints. This lack of divergence reflects a compounding effect: the inherent velocity and structural vulnerabilities of sexed spermatozoa, combined with our constrained sample. These findings align with reports that pregnancy rates following transfer of embryos derived from sex-sorted semen remain lower compared with conventional semen [11,21]. Nonetheless, the present results indicate that post-mortem oocytes are compatible with sex-biased embryo production, albeit at reduced efficiency.
8. Embryo Development and Cryosurvival
Cleavage rates with unsexed semen were significantly higher in Group I (55.7%) compared with Group II (38.3%), whereas blastocyst rates showed only a numerical advantage, a pattern consistent with earlier reports linking delayed ovary processing to reduced fertilization and blastocyst yield in cattle and buffalo [7,22]. Crucially, blastocysts derived from ≤6 h retrieval exhibited higher cell numbers (138.4 vs. 102.3) and superior cryotolerance, supporting the premise that embryo cell number is a reliable indicator of developmental quality and implantation potential [23,24]. Enhanced post-warming survival in this group further aligns with evidence that oxidative stress mitigation improves bovine embryo viability [25]. High re-expansion rates of vitrified embryos from Group I (90.9%) suggest that embryos derived from oocytes recovered within 6 h of death retain cryotolerance for long-term preservation; however, due to fewer embryos from later retrievals, the sample size was not enough to statistically evaluate their vitrifiable quality. These findings are consistent with studies showing that optimized vitrification protocols enhance survival and enable direct transfer in cattle [26,27,28]. Collectively, the results indicate that early retrieval not only increases the number of transferable embryos but may also ensures the generation of cryoresilient embryos, which is one of the critical prerequisites for efficient breeding programs and germplasm banking.
9. Conclusions
Oocytes retrieved from post-mortem Jersey cow ovaries retained the capacity to mature, fertilize, and form blastocysts in vitro, with outcomes strongly influenced by the timing of retrieval. Collection within 6 h of death yielded significantly higher maturation rates, superior embryo quality, and greater cryosurvival compared with retrieval after 6 h. The recovery of developmentally competent blastocysts and their successful vitrification and thawing confirm the potential utility of this approach.
Acknowledgments
The authors acknowledge financial assistance from the Holistic Agriculture Development Programme (HADP 23) of the Government of J&K UT for conducting this research.
Author Contributions
Conceptualization: S.M.S. and A.A.M.; Methodology: A.A.M., S.M.S, R.A.D. and S.H.Y.; Investigation: A.A.M., F.A.B., S.H.Y. and S.I.U.U.; Data curation: A.A.M, S.M.S and S.I.U.U.; Formal analysis: A.A.M., S.M.S. and A.A.S.; Writing—original draft preparation: S.M.S, A.A.M.; Writing—review and editing: S.M.S., A.A.P., R.A.S. and A.A.M.; Supervision: S.M.S, R.A.S; Project administration: S.M.S., A.A.P; Funding acquisition: R.A.S, S.M.S, A.A.P. All authors have read and agreed to the published version of the manuscript.
Ethics Statement
Ethical approval was obtained from the Institutional Animal Ethics Committee (IAEC) (Protocol Code: P23DAR07. Approval Dated: 12/08/2024) prior to the commencement of the study. No live animals were used in the study.
Informed Consent Statement
Not applicable.
Data Availability Statement
The data supporting the findings of this study are available from the corresponding author upon reasonable request.
Funding
This work was financially supported by the Holistic Agriculture Development Programme (HADP-23), Government of Jammu and Kashmir Union Territory.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Government of Jammu and Kashmir, Animal/Sheep Husbandry & Fisheries Department. Jammu and Kashmir State Livestock Breeding Policy, 2019; Government Order No. 74-ASH of 2019, dated 08 June 2019; Civil Secretariat: Srinagar, Jammu and Kashmir, India, 2019. Available online: https://faolex.fao.org/docs/pdf/IND227451.pdf (accessed on 18 May 2026).
- Department of Animal Husbandry and Dairying. 20th Livestock Census-2019: All India Report; Ministry of Fisheries, Animal Husbandry and Dairying, Government of India: New Delhi, India, 2019. Available online: https://dahd.gov.in/sites/default/files/2023-06/Annual2019-20.pdf (accessed on 18 May 2026).
- Department of Animal Husbandry and Dairying. Annual Report 2023–2024; Ministry of Fisheries, Animal Husbandry and Dairying, Government of India: New Delhi, India, 2024. Available online: https://dahd.gov.in/sites/default/files/2024-10/AnnualReport202324.pdf (accessed on 18 May 2026).
- International Union for Conservation of Nature (IUCN). The IUCN Red List of Threatened Species, Version 2022-2. Available online: https://www.iucnredlist.org(accessed on 18 May 2026).
- Ceballos G, Ehrlich PR, Raven PH. Vertebrates on the brink as indicators of biological annihilation and the sixth mass extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. DOI:10.1073/pnas.1922686117 [Google Scholar]
- Sidova M, Tomankova S, Abaffy P, Kubista M, Sindelka R. Effects of post-mortem and physical degradation on RNA integrity and quality. Biomol. Detect. Quantif. 2015, 5, 3–9. DOI:10.1016/j.bdq.2015.08.002 [Google Scholar]
- Blondin P, Coenen K, Guilbault LA, Sirard MA. In vitro production of bovine embryos: Developmental competence is acquired before maturation. Theriogenology 1997, 47, 1061–1075. DOI:10.1016/S0093-691X(97)00063-0 [Google Scholar]
- Van Soom A, Mateusen B, Leroy J, De Kruif A. Assessment of mammalian embryo quality: What can we learn from embryo morphology? Reprod. Biomed. Online 2003, 7, 664–670. DOI:10.1016/S1472-6483(10)62089-5 [Google Scholar]
- Álvarez-Gallardo H, Kjelland ME, Pérez-Martínez M, Villaseñor-González F, Romo-García S. Evaluation of novel SexedULTRA-4M technology for in vitro bovine embryo production. Anim. Reprod. 2022, 19, e20220018. DOI:10.1590/1984-3143-AR2022-0018 [Google Scholar]
- Reese S, Pirez MC, Steele H, Kölle S. The reproductive success of bovine sperm after sex-sorting: A meta-analysis. Sci. Rep. 2021, 11, 17366. DOI:10.1038/s41598-021-96834-2 [Google Scholar]
- Murphy EM, Crowe AD, Thompson L, Moore SG, McDonald M, Hordern E, et al. Fertility in lactating dairy cows following timed embryo transfer with fresh in vitro-produced embryos derived from conventional or sex-sorted semen. J. Dairy Sci. 2025, 108, 11635–11650. DOI:10.3168/jds.2025-27056 [Google Scholar]
- Sartori R, Balistrieri M, Silva LOE, Consentini CEC, Melo LFE, Pontes GCS, et al. Pregnancy loss in cattle with emphasis on embryo transfer programs. Anim. Reprod. 2025, 22, e20250045. DOI:10.1590/1984-3143-AR2025-0045 [Google Scholar]
- Malik AA, Shah RA, Yaqoob SH, Gayas MA, Handoo N, Magray SN, et al. Comparative developmental competence and gene expression in laparoscopically retrieved- and abattoir-derived goat oocytes. Front. Vet. Sci. 2026, 13, 1737059. DOI:10.3389/fvets.2026.1737059 [Google Scholar]
- Aguila L, Treulen F, Therrien J, Felmer R, Valdivia M, Smith LC. Oocyte selection for in vitro embryo production in bovine species: Noninvasive approaches for new challenges of oocyte competence. Animals 2020, 10, 2196. DOI:10.3390/ani10122196 [Google Scholar]
- Parrish JJ, Susko-Parrish JL, Leibfried-Rutledge ML, Critser ES, Eyestone WH, First NL. Bovine in vitro fertilization with frozen-thawed semen. Theriogenology 1986, 25, 591–600. DOI:10.1016/0093-691X(86)90143-3 [Google Scholar]
- Shah SM, Saini N, Ashraf S, Zandi M, Manik RS, Singla SK, et al. Development, characterization, and pluripotency analysis of buffalo (Bubalus bubalis) embryonic stem cell lines derived from in vitro-fertilized, hand-guided cloned, and parthenogenetic embryos. Cell. Reprogram. 2015, 17, 306–322. DOI:10.1089/cell.2014.0098 [Google Scholar]
- Soto-Heras S, Paramio MT. Impact of oxidative stress on oocyte competence for in vitro embryo production programs. Res. Vet. Sci. 2020, 132, 342–350. DOI:10.1016/j.rvsc.2020.07.013 [Google Scholar]
- Sánchez JM, Simintiras CA, Lonergan P. Aspects of embryo-maternal communication in establishment of pregnancy in cattle. Anim. Reprod. 2019, 16, 376–385. DOI:10.21451/1984-3143-AR2019-0075 [Google Scholar]
- van Rumpt M, Rabaglino MB. The role of machine learning in decoding the molecular complexity of bovine pregnancy: A review. Reprod. Fertil. Dev. 2024, 37, RD24141. DOI:10.1071/RD24141 [Google Scholar]
- Ngcobo JN, Nedambale TL, Sithole SM, Mtileni B, Mpofu TJ, Ramukhithi FV, et al. A systematic review on the prospects of X- and Y-sexed semen in ruminant livestock: Implications for conservation, a South African perspective. Front. Vet. Sci. 2024, 11, 1384768. DOI:10.3389/fvets.2024.1384768 [Google Scholar]
- Mikkola M, Andersson M, Taponen J. Transfer of cattle embryos produced with sex-sorted semen results in impaired pregnancy rate and increased male calf mortality. Theriogenology 2015, 84, 1118–1122. DOI:10.1016/j.theriogenology.2015.06.012 [Google Scholar]
- Shahzad Q, Waqas M, Pu L, Wadood AA, Xie L, Husna AU, et al. Seasonality and photoperiod influence in vitro production of buffalo embryos. Reprod. Domest. Anim. 2020, 55, 1115–1123. DOI:10.1111/rda.13749 [Google Scholar]
- Rabel RAC, Marchioretto PV, Bangert EA, Wilson K, Milner DJ, Wheeler MB. Pre-implantation bovine embryo evaluation: From optics to omics and beyond. Animals 2023, 13, 2102. DOI:10.3390/ani13132102 [Google Scholar]
- Hansen PJ, Tríbulo P. Regulation of present and future development by maternal regulatory signals acting on the embryo during the morula to blastocyst transition—insights from the cow. Biol. Reprod. 2019, 101, 526–537. DOI:10.1093/biolre/ioz030 [Google Scholar]
- da Silva Feuchard VL, Oliveira CS, Saraiva NZ, Quintão CCR, Oliveira LZ. Reactive oxygen species attenuation improves the quality of vitrified-warmed bovine embryos. Anim. Reprod. 2025, 22, e20240035. DOI:10.1590/1984-3143-AR2024-0035 [Google Scholar]
- Sanches BV, Lunardelli PA, Tannura JH, Cardoso BL, Pereira MH, Gaitkoski D, et al. A new direct transfer protocol for cryopreserved IVF embryos. Theriogenology 2016, 85, 1147–1151. DOI:10.1016/j.theriogenology.2015.11.029 [Google Scholar]
- Oliveira CS, da Silva Feuchard VL, de Freitas C, da Silva Rosa PM, dos Camargo AJR, Saraiva NZ. In-straw warming protocol improves survival of vitrified embryos and allows direct transfer in cattle. Cryobiology 2020, 97, 222–225. DOI:10.1016/j.cryobiol.2020.02.007 [Google Scholar]
- Martínez-Rodero I, García-Martínez T, Ordóñez-León EA, Vendrell-Flotats M, Olegario Hidalgo C, Esmoris J, et al. A shorter equilibration period improves post-warming outcomes after vitrification and in straw dilution of in vitro-produced bovine embryos. Biology 2021, 10, 142. DOI:10.3390/biology10020142 [Google Scholar]