Beyond Oil: Molecular Breeding for Multifunctional Innovation in Oilseed Brassica

Beyond Oil: Molecular Breeding for Multifunctional Innovation in Oilseed Brassica
Yingying Zhou 1 Iram Batool 1 Kangni Zhang 1 Ling Xu 2 Qian Huang 3 Fakhir Hannan 1 Yongqi Sun 1 Tongjun Qin 1 Wan Xu 4 Wu Qian 5 Ahsan Ayyaz 1,2,* Weijun Zhou 1,*
Received: 10 March 2026 Revised: 07 April 2026 Accepted: 08 June 2026 Published: 12 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
Graphical Abstract
1. Introduction
Brassica napus L. (rapeseed, canola) is the world’s second most significant oilseed crop, with an annual production exceeding 75 million metric tons. It basically yields vegetable oil for human consumption and a high-protein meal as an essential component of animal feed. Traditional breeding has been in a yield paradigm whereby emphasis is placed on the production of more seed and better yield in oil content and fatty acid composition [1]. Low-erucic acid and low-glucosinolate (double-low) rapeseed varieties marked a breakthrough in the nutritional and feed quality, substantially increasing the crop’s value [2]. These breeding efforts have already been fruitful; the oil content of contemporary hybrid varieties is now more than 50%, and yield has increased by about 1.5% each year over the last several decades [3]. Moreover, the utility of rapeseed extends far beyond its seeds; the oil is not only a major source of edible oils but also a key feedstock for biodiesel and industrial lubricants [4]. Majorly, in livestock and aquaculture feed, defatted seed meal (35–45% protein) is used, which lowers the need to import soybean meal [5]. B. napus also offers significant ecological advantages. It is a prolific producer of nectar and promotes the health of the pollinators and the apiculture industry [6].
The fact that it grows very fast and also because of its high biomass makes it an effective cover crop as well as green manure, and this helps to organize the soil and recycle of nutrients as well as the process of carbon sequestration. Additionally, in the genus Brassica, a few of them have been domesticated about the vegetative structures, such as B. rapa, as a leafy vegetable or ruminant fodder crop [7]. Brassica is also a good source of bioactive compounds with high nutraceutical and pharmaceutical value. Glucosinolates and hydrolysates (e.g., sulforaphane), phenols, and tocopherols are anti-inflammatory, antioxidant, and anti-carcinogenic compounds that could be useful as the ingredients in functional foods and cosmetics [8]. The species is pre-adapted to abiotic stressors (e.g., cold, drought) and is suited to various cropping systems (e.g., winter cropping, rotations), which gives it a place as a backbone to climate-resilient intensive sustainable agriculture [9]. Although rapeseed has multiple potential applications, its full exploitation of the crop is yet to be achieved, partly because breeding has been emphasized on oil maximization. Therefore, a paradigm shift is required, which would not be the oil-centric model of breeding but would create this multifaceted crop in all aspects. This will involve decoding the complicated genetic systems that dictate not only seed characteristics but also vegetative biomass, metabolic capacity, and stress offenses [10]. Modern molecular tools enable this transformation. With the availability of high-quality reference genomes, advanced multi-omics approaches such as genomics, phenomics, metabolomics (Figure 1), and precision breeding technologies, we now possess an unprecedented capacity to dissect and re-engineer these complex traits [11]. Hence, this review focuses on the emerging concept of oilseed Brassica as a multipurpose biological system for sustainable production. Leveraging its genetic and genomic plasticity, we discuss innovative molecular breeding technologies for the simultaneous optimization of yield, quality, and resilience, as well as for adding value to non-seed tissues. We also provide a vision of how this designer crop could be integrated into systems that address global needs for nutrition, renewable biomass, and ecological health.
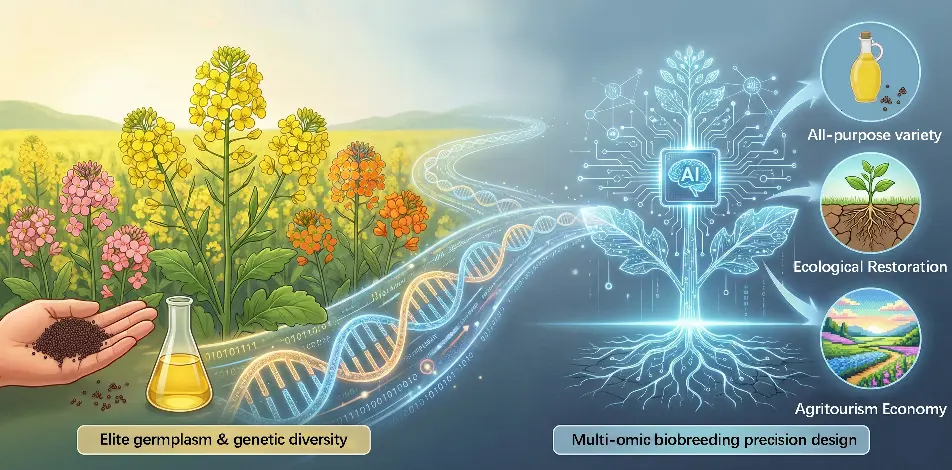
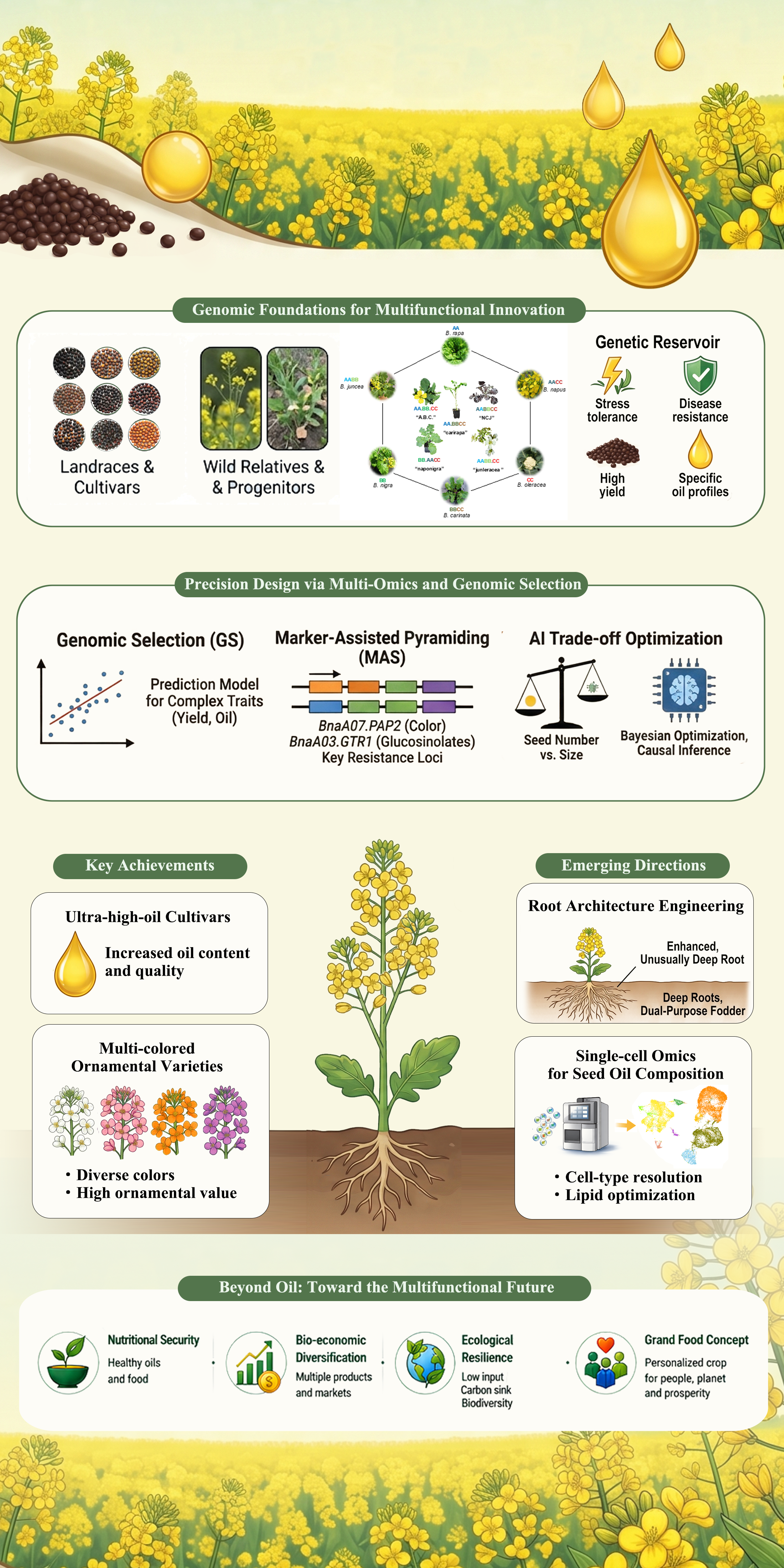
Figure 1. Multifunctional landscape and integrative applications of modern Brassica napus. This figure shows the path from elite genetic resources to AI-driven design. It highlights how modern rapeseed serves multiple goals, including high-quality oil production, ecological restoration, and the agri-tourism economy to support the “Grand Food Concept”.
2. Genetic Diversity and Genomic Foundations for Multifunctional Innovation
The remarkable adaptability of B. napus is rooted in its complex genetic architecture. As an allotetraploid, it has evolved within the Triangle of U, creating a genomic foundation that supports high phenotypic plasticity. This section explores this evolutionary history and how it has endowed rapeseed with functional traits, natural genetic variation, and phenotypic diversity, as well as the modern techniques that can be applied to generate new germplasm resources. Together, these constitute the genetic toolbox needed to engineer rapeseed beyond its traditional role in oil production.
2.1. Evolutionary Origin and Genomic Architecture of Brassica napus
The allotetraploid species in U’s triangle are among the youngest known allopolyploids. Brassica napus in particular is thought to have formed less than 7500 years ago through hybridization between B. rapa and B. oleracea, followed by chromosome doubling [12].
The genus Brassica is an outstanding agricultural genus of the Brassicaceae family, traditionally consisting of three diploid species B. rapa (AA genome, n = 10), B. nigra (BB genome, n = 8), and B. oleracea (CC genome, n = 9), and three allotetraploid species B. juncea (AABB, n = 18), B. napus (AACC, n = 19), and B. carinata (BBCC, n = 17). They have genetic relationships defined by the Triangle of U (Figure 2).
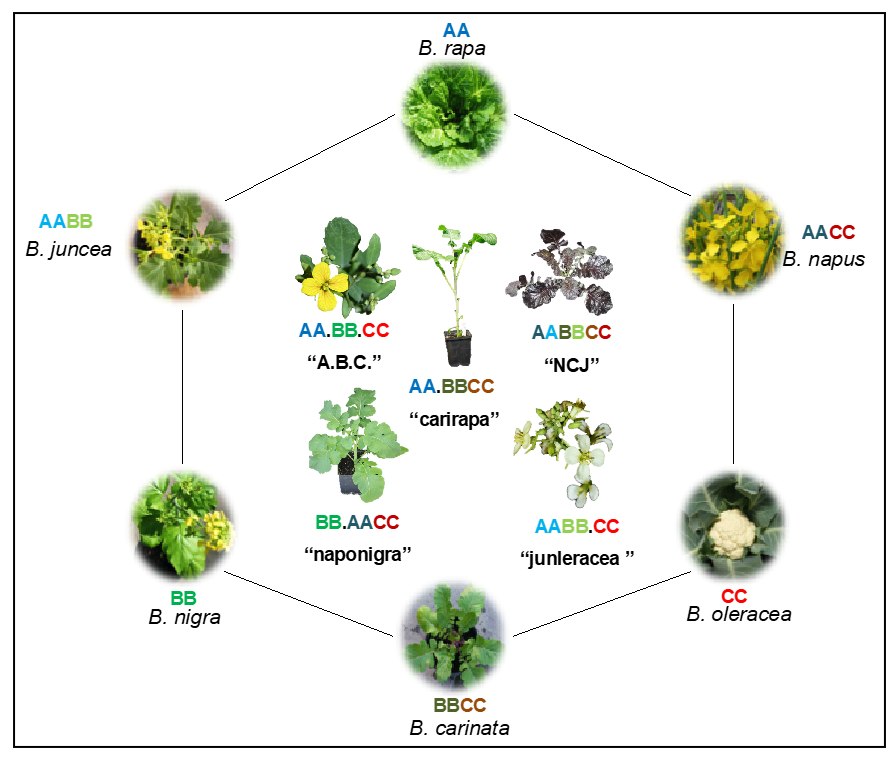
Figure 2. Beyond legacy of “U’s Triangle”, showing the genetic relationships among the six classical species of the genus Brassica, plus the five novel allohexaploids. Reproduced with permission from our previous Ref. [13]. The traditional “Triangle of U” as a hexagon, with established diploid species Brassica rapa, B. nigra, B. oleracea with AA, BB and CC genome complements, the established allotetraploid species B. juncea, B. napus and B. carinata with AABB, AACC and BBCC genome complements, and possible combinations of allohexaploids which can be derived from crosses between them: “naponigra” (B. napus × B. nigra), “junleracea” (B. juncea × B. oleracea), “carirapa” (B. carinata × B. rapa), “NCJ” (B. napus × B. carinata × B. juncea) and “A.B.C.” (B. rapa × B. oleracea × B. nigra).
The allotetraploid B. napus (AnAnCnCn) was formed due to either natural hybridization and whole-genome duplication of its parents, B. rapa (ArAr) and B. oleracea (CoCo) [13]. Despite the traditional Triangle of U model, which explains the basic genomic arrangement of rapeseed (B. napus), recent evidence shows that allopolyploid gene introgression, especially that of the B. rapa ancestor, has contributed much more to the diversification of the latter [14], which has contributed to its genetic diversity and adaptation. Most allopolyploid species have asymmetric sub-genomes on structural and functional levels, which results in genome shock that supports evolutionary innovation [15]. The Cn sub-genome has undergone more loss of genes, increased sequence diversification and accumulation of transposable elements (TEs) than the An sub-genome in B. napus [16]. B. napus represents a young and still evolving allopolyploid system. Its genomic flexibility provides a basis for phenotypic variation and creates opportunities for further genetic improvement.
2.2. Genetic Resources for Multifunctional Traits
B. napus is not only a global source of oilseeds but also provides vegetables and fodder. Major repositories worldwide store tens of thousands of germplasm accessions [17]. This global germplasm comprising landraces, elite cultivars, wild relatives, and synthetic polyploids harbors extensive natural variation due to its allotetraploid origin (AACC, 2n = 38), providing the genetic foundation for phenotypic plasticity and broad adaptability. To exploit these resources, researchers employ pan-genome sequencing, genome-wide association studies (GWAS), and large-scale transcriptomic platforms (e.g., BnIR) for in-depth characterization [18]. For multifunctional innovation, B. napus germplasm can be classified based on target phenotypes and relevant genes (Table 1).
Table 1. Functional classification of Brassica napus genetic resources and key genes for multifunctional traits.
|
Functional Category |
Trait |
Representative Gene(s) |
|---|---|---|
|
Flower Color & Ornamental Value |
Anthocyanin biosynthesis |
|
|
Petal color variation |
||
|
Yield and Seed Traits |
Seed color |
|
|
Flowering time & floral transition |
||
|
Seed size/seed number |
BnEOD3 (CYP78A6) [32], BnKLUH (CYP78A5) [32], BnARF18 [33], BnDA1 [34] |
|
|
Silique number and shape |
BnCLV3, BnCLV1, BnCLV2 [35] |
|
|
Pod shattering resistance |
BnSHP1 [36], BnSHP2 [37], BnIND [38], BnALC [39], BnJAG.A08 [40] |
|
|
Oil content and fatty acid composition |
BnLPAT2 [41], BnLPAT5 [41], BnFAD2 [42], BnDGAT1 [43], BnGPAT9 [44] |
|
|
Root Architecture & Nutrient Efficiency |
Root system architecture |
|
|
Nitrogen-use efficiency |
||
|
Stress and Resilience Traits |
Drought tolerance |
|
|
Disease resistance |
||
|
Cold/Heat tolerance |
||
|
Quality and Value-added Traits |
Glucosinolate content |
|
|
Protein quality and seed storage |
||
|
Pigment & antioxidant metabolism |
This practical categorization, incorporating recent genomic and gene-editing discoveries, highlights how existing genetic diversity can be harnessed to diversify B. napus for ornamental, productive, and resilient applications. By combining genomic variants with high-throughput phenotypic data, it is now possible to precisely identify quantitative trait loci (QTLs) and major genes influencing traits such as flower color, root architecture, quality components, and stress resistance [67]. Research on asymmetrical sub-genome evolution and epigenetic control (e.g., chromatin accessibility differences) has revealed the molecular nature of variation of functional traits across accessions, and is increasingly turning germplasm banks into a genetic toolbox to breed with precision.
2.3. Innovation of New Genetic Resources
To overcome the genetic constraints imposed by decades of intense selection for oil-related traits, it is essential to develop new germplasm resources that facilitate the versatile expansion of B. napus. One of the most effective approaches is interspecific hybridization and the production of synthetic polyploids to broaden the gene pool [68]. Allotriploid (ArAnCn) strategies enhance diversity, but the creation of allohexaploid Brassica (AABBCC, 2n = 6x = 54), also known as Super Brassica, is the new horizon of innovation of genomes [69]. This novel allohexaploid has a potential to bring together genetic variation and favorable features in all six Brassica species, with the help of other allelic heterosis. The goal of merging the A, B, C genomes is to increase the high yield of B. napus (AC) with the improved disease resistance (e.g., blackleg, club root) and stronger abiotic stress tolerance (e.g., salt and drought) of the B genome (B. nigra).
Such resources offer an avenue of multi-trait breeding, which combines the applications of crops to high-protein feed, bioenergy, and industrial oils. In addition to structural genomic modifications, the contemporary germplasm innovation is starting to revolve around the area of epigenetics. Hybridization causes a massive expression change of genes and epigenetic reprogramming, known as transcriptome shock [70]. Another new opportunity to alter sub-genome-specific DNA methylation patterns is focused on the manipulation of the RdDM pathway, such as the mutation of such key genes as BnaDCL3 and BnaRDR2, which are expected to alter the sub-genome-specific patterns of DNA methylation [71]. This epigenetic editing, coupled with the de novo domestication of allohexaploids, could promise to stabilize nascent genomes and produce new phenotypic variation that could not be achieved in other ways. Despite persistent challenges with genomic instability and reduced fertility in early generations, significant progress in chromosome engineering and selection systems has established synthetic resources as the foundation for next-generation, multi-purpose Brassica crops so-called “chassis”.
3. Precision Design of Multifunctional Oilseed Brassica via Multi-Omics and Genomic Selection (GS)
Technologies for improving canola (B. napus) are now based on precision breeding, integrating genomics, multi-omics networks, and intelligent prediction (Figure 3). The aim of these approaches is to broaden the functional scope of rapeseed beyond a traditional oil crop, toward a more versatile crop with applications in ornamental, edible, feed, and ecological systems. The following sections describe three key components that enable precision breeding: genomic prediction, marker-assisted trait stacking, and systems-level integration of omics.
3.1. Integrated Genomic Prediction and Marker-Assisted Trait Pyramiding
Molecular prediction has revolutionized plant breeding by enabling the direct application of whole-genome markers in estimating the potential genetic capabilities instead of depending on phenotypic selection. Genomic prediction (GP) became a reality in the early 21st century using statistical models, including the genomic best linear unbiased prediction (GBLUP) and Bayesian models [72]. These methods predict the breeding value of individuals since the sum total of the minuscule impact of thousands of single nucleotide polymorphisms (SNPs) get the polygenic nature of intricate phenomena [73]. In B. napus, high-density genotyping and sequencing technologies have supported the application of GS across breeding programs, allowing in silico prediction of trait performance under different environments.
GS considers the entire picture of genome rather than just a few large major quantitative trait loci (QTLs) like seed yield, oil quality, and stress resistance, which is particularly effective in the case of complex polygenic phenotypes such as seed yield [67]. This has been caused by the emergence of pan-genomic information and multi-environment scale-level experiments, which have resulted in genomic selection algorithms that integrate machine learning, environmental covariates, and gene-by-gene epistatic interactions [74]. These advancements have made it possible to select in the early-stages of breeding based on genomic information and shortened the breeding period of rapeseed by a significant margin [75]. Complementary marker-assisted selection (MAS) allows stacking major-effect alleles precisely to provide a hybrid strategy of combining extensive genomic prediction with highly accurate gene localization [76]. It has also transformed Brassica by integration of GS and MAS, such that Brassica ceased to be dependent on phenotypes during breeding, but prediction-based breeding and data breeding. Big and heterogeneous collections of germplasm have played a significant role in calibration of genomic predictions. For instance, the genome-wide association studies and whole-genome resequencing, which find SNP genotype loci that have an effect on the trait, and used in models like GBLUP to improve prediction accuracy in early selection [77].
In the meantime, marker-assisted gene stacking is more specific in that it increases the effects of the specifically targeted gene loci. Natural allelic variants to genes like BnFAD2 (oleic acid content), BnWRKY70 (disease resistance), and BnTTG1 (flavonoid biosynthesis) have been utilized in B. napus to provide more than one trait into elite germplasm without being transgenically altered [27,49,78]. Therefore, GS to select the background and MAS to select the foreground, making it a two-level breeding approach. One of the challenges of breeding plants is how to trade of between divergent characteristics (e.g., yield and seed size, defense and growth rate). Recent progress in multi-trait genomic prediction and causal inference modeling now allows breeders to measure and optimize these interactions, helping produce the best varieties [79].
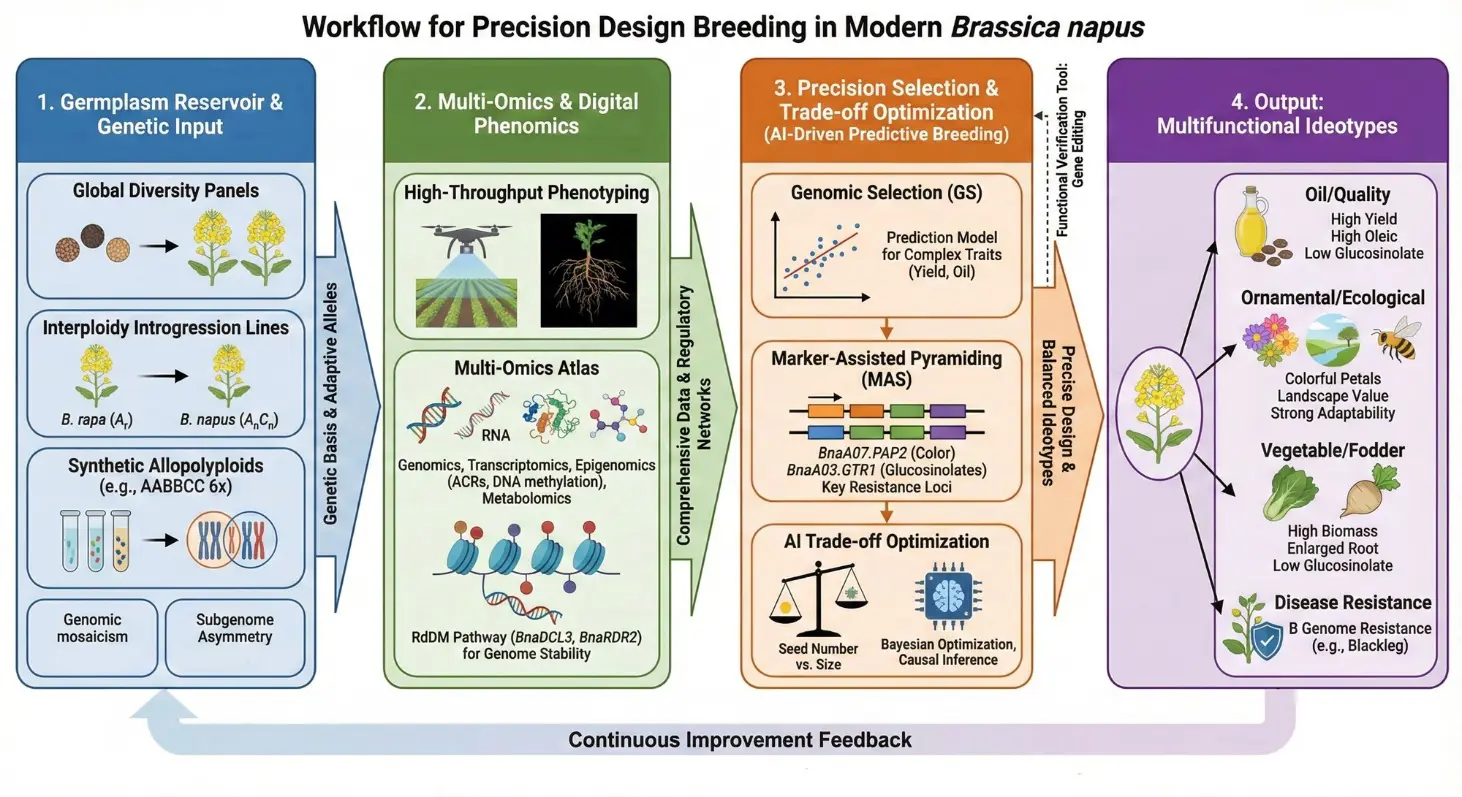
Figure 3. It is the breeding of ideotypes of multifunctional Brassica napus. Four integrated modules, (1) Input: expansion of genetic diversity by using diversity panels and synthetic polyploids; (2) Data: bridging high-throughput phenomics and multi-omics to decode regulatory networks; (3) Core: AI-innovative genomic selection and marker-assisted pyramiding; (4) Output: non-transgenic ideotypes by using genetic diversity adapted to oil quality, ornamental value, and ecological resilience convert genetic diversity to elite varieties. It has a feedback mechanism whereby it continuously contributes to the germ plasm store.
3.2. Multi-Omics Networks Driving Optimization of Seed Oil Composition
Multi-omics technologies have transformed plant biology by enabling the simultaneous interrogation of genetic, transcriptional, proteomic, and metabolic layers of regulation. In B. napus, this integrated framework is particularly valuable because key agronomic and industrial traits, especially seed oil content and composition, are governed by highly interconnected regulatory networks rather than single loci [80,81]. Genomic variation derived from pan-genome resources provides the structural basis of diversity, while transcriptomic profiling captures dynamic gene regulation during development and stress responses [67,82]. Proteomics and metabolomics further complement these datasets by linking gene activity to enzymatic function and metabolic flux, particularly in lipid biosynthesis pathways [83].
A representative example of multi-omics integration in B. napus is triacylglycerol (TAG) biosynthesis and fatty acid (FA) metabolism, which directly determine oil yield and quality. Key enzyme-coding genes such as acetyl-CoA carboxylase (ACCase), stearoyl-ACP desaturase (SAD), diacylglycerol acyltransferase (DGAT), and phospholipid:diacylglycerol acyltransferase (PDAT) play central roles in regulating carbon flux toward lipid accumulation [84]. At the regulatory level, transcription factors including WRI1, LEC1, and ABI3 integrate carbon metabolism with storage lipid biosynthesis during seed development [85]. Multi-omics studies combining transcriptomic and proteomic data have revealed coordinated developmental modules associated with early fatty acid synthesis, TAG assembly, and oil body formation, suggesting that oil accumulation is governed by tightly synchronized gene–protein networks rather than isolated pathways.
In addition to transcriptional regulation, epigenetic mechanisms such as chromatin accessibility and histone modifications further contribute to the control of lipid metabolism, indicating that three-dimensional genome organization may influence the activity of lipid-related genes. Metabolomic profiling provides complementary evidence by identifying key intermediate and cofactor limitations in fatty acid elongation and desaturation pathways, thereby revealing metabolic bottlenecks that shape both oil yield and quality. Together, these findings highlight that seed oil traits in B. napus emerge from multi-layered regulatory interactions spanning the genome to the metabolome.
Recent advances in single-cell and spatial transcriptomics further refine this framework by revealing cell-type-specific metabolic specialization within developing seeds [86,87]. When integrated with spatial metabolomics and chromatin architecture mapping, these approaches enable the reconstruction of spatially resolved metabolic microenvironments during seed development. Coupled with machine learning and network modeling approaches such as weighted gene co-expression network analysis (WGCNA), these datasets allow the identification of regulatory modules underlying lipid accumulation and stress adaptation [39]. Process-based and AI-assisted models further support predictive simulation of carbon allocation between competing pathways, providing a basis for rational design of oilseed ideotypes with tailored fatty acid profiles.
3.3. Intelligent Breeding Pipelines for Multifunctional Ideotypes
The introduction of intelligent breeding systems that incorporate data, artificial intelligence (AI), and systems modeling has led to a new generation of crop advancement. Traditional breeding practices are founded on a large-scale application of empirical selection in addition to breeder knowledge; however, the rising levels of multi traits target in crops such as rapeseed make prediction as well as simulation-based decision-making systems mandatory. Donald introduced the ideotype concept, which is a vision of a model or an idea of a plant optimized to the selected environmental and agronomic goals [88]. The use of modern computational tools, mathematical modeling, optimization algorithms, and data synthesis with the help of artificial intelligence makes the quantitative ideal type design possible.
It is now possible to predict phenotypic performance using machine-learning models, especially the convolutional neural network (CNN) and gradient boosting models, using the large-scale multi-omics and environmental measurements, and in-silico screen before field identification [89]. AI-assisted optimization methods enable plant breeders to simultaneously combine multiple traits, thereby overcoming traditional trade-offs such as growth versus disease resistance and yield versus quality. Together with digital phenotyping solutions (e.g., drone imaging and real-time stress sensors), breeders would be able to simulate the performance of plants at various time points and create optimal plant architecture, which is not only adapted to the environment but also achieves production objectives. This approach brings about a shift towards involving plant breeding as a process of trial and error to an engineered, data-driven procedure.
For B. napus, this technology can be used to design multi-purpose ideotypes suited to various contexts: ornamental varieties for tourism, dual-purpose varieties for vegetable and fodder production, and stress-tolerant varieties for marginal lands. The essence of molecular breeding is to translate genetic potential into ideotypes that are effective across various environments. The ideal ideotype is one that is characterized by high yield of oil, appealing floral color, desirable edible or fodder quality, and ecological resilience, in the case of B. napus. The problem of achieving this balance is that it needs smart optimization systems that are capable of a trade-off of traits. The models of Bayesian optimization and causal inference enable the weighting of traits based on the environment and stage of development, where the optimal combinations of traits can be determined [90]. AI also combines multi-omics data and time-series of phenomics to project genotype performance, and neural networks demonstrate the ability of individual alleles to enhance yield without decreasing stress tolerance [91]. Process-based models are models that simulate the flow of both carbon and lipids in order to enhance the efficiency of resource allocation [68]. These tools combined allow testing virtually, breeding faster with the result of multifunctional ideal type varieties as the future of sustainable agriculture.
4. Expanding the Diversified Application of B. napus
4.1. Ornamental and Ecological Innovations in Multi-Colored Rapeseed
The applications of rapeseed (B. napus) have expanded far beyond its traditional uses in edible oil, biofuels, and animal feed to include high-value ornamental and ecological roles. Originally bearing yellow flowers, modern rapeseed has evolved through interspecific hybridization and selective breeding to display a diverse palette of flower colors, including orange, red, pink, and purple (Figure 4). This shift toward ornamental and ecological applications addresses the need to diversify agricultural products and promote agro-tourism, while also enhancing the crop’s nutritional and medicinal value [92]. These diverse colors are scientifically rooted in the flavonoid biosynthetic pathway, specifically in the accumulation of anthocyanins. Anthocyanins are water-soluble pigments that produce the red, pink, blue, and purple color on different organs of plants. Metabolically expressed, the color of the petals depends on concentrations and composition of individual anthocyanidins; pelargonidin type generates orange-to-red color, whereas cyanidin and peonidin type generate pink-to-fuchsia color, and delphinidin, petunidin, and malvidin type generate purple to blue color. A recent metabolomic profiling has discovered 79 anthocyanins and associated metabolites in the colorful genotypes, and the presence of high levels of cyanidins specifically suggested their involvement in the red pigmentation process [93]. A conserved set of structural genes and transcription factors governs the molecular control of this transition. The major structural enzymes are Phenylalanine Ammonia-Lyase (PAL), Chalcone Synthase (CHS), Flavanone 3-Hydroxylase (F3H), Dihydroflavonol 4-Reductase (DFR) and Anthocyanin Synthase (ANS). The MYB-bHLH-WD40 (MBW) regulatory complex coordinates these. The major regulators of B. napus are BnaA07.PAP2 (induces anthocyanin genes to make apricot turn to red) and BnaA03.ANS (key to red pigmentation). Integrated transcriptomics has continued to perfect this network [93,94]. Multi-colored rapeseed can offer very important non-aesthetic services ecologically. Colorful flowers are also noticed, which attract the pollinators, leading to an increase in the success of pollination and seed production. In addition, flavonoids and anthocyanins assist the resistance of plants against stress, as they are antioxidants that eliminate destructive free radicals. They prevent exposure of plants to UV rays, heavy metals, drought, salt stress, and frost. How ornamental properties can also enhance ecological resilience is demonstrated by purple-stem plants that are more efficient photosynthesis systems and stronger antioxidants during periods of drought due to their richness in anthocyanins. Together, these findings show that variation in flower color in B. napus is not simply a visible trait. It reflects regulated changes in flavonoid metabolism and can be directed through molecular breeding and network-level regulation.

Figure 4. Representative examples of phenotypic diversity in multi-colored Brassica napus. Field demonstration of the multifunctional “Zheda Colorful Rapeseed”. This series exhibits phenotypic diversity, boasting over 15 different petal colors. The color innovation is not limited to flowers but extends to multiple vegetative and reproductive organs, including leaves, stems, branches, and pods.
4.2. Unlocking Fodder and Vegetable Potential via Root Architecture Plasticity
This high plasticity of B. napus root architecture gives it a biological basis to grow its application as either fodder or a vegetable. Even though it has primarily been bred as a seed and oil crop, its hybrid nature of B. rapa and B. oleracea enables it to inherit the gene of thickening of the root, storage of carbohydrates as well as vegetative development. Recent genomic and transcriptomic findings demonstrate that such concealed qualities can be unleashed through molecular breeding, thus establishing new possibilities for multi-purpose rapeseed varieties [67]. Root thickening has already been well characterized in B. rapa, which would be valuable in this enhancement [95]. In a recent study, it was demonstrated through histological and transcriptomic studies that impactful cellular processes include quickening of the growth and expansion of the parenchyma cells and the cambium during the formation of the hypocotyl-tuber [96]. Hormone interactions (particularly auxin and cytokinin balance), sugar build up, and cell wall remodeling control this process. A number of the hub genes were found, such as Bra-EXP3, Bra-STP1.1, Bra-IAA1, Bra-ARF17, Bra-CYP735A2, and Bra-FLOR1 [97]. These genes have orthologs in B. napus and are differentially expressed in root and hypocotyl tissues indicating that marker-assisted selection or transcriptome-based breeding [67] may reinstate the same developmental toolkit. The SUS pathway and HXK pathway regulate the breakdown and transportation of sucrose and the root strength of Brassica plants that produce storage roots [98]. The balance of hormones controls the division of the cells in the cambium. In B. napus, increased BnARF7, BnLBD29, and BnEXPA10 are associated with increased root biomass, which shows that specific regulation of the mentioned genes might alter the growth of plants to edible or fodder phenotypes [99]. This type of multifunctional potential is exemplified by a hypocotyl-tuber-forming germplasm of B. napus that grows and fruit in field conditions and retains normal flowering and seed-setting ability (Figure 5). These results show that B. napus has untapped potential. These findings suggest the potential to develop multi-purpose B. napus ideotypes combining edible, fodder, and oilseed functions. Root architecture plasticity in B. napus also represents a useful breeding target. It offers a way to develop dual-purpose ideotypes that combine vegetative biomass production with stable reproductive performance.

Figure 5. Multifunctional Brassica napus exhibiting hypocotyl-tuber development with normal flowering and seed-setting capabilities. This ideotype supports a dual-purpose utilization model, combining the harvest of nutrient-rich hypocotyl-tubers with stable seed productivity within a single life cycle, though it requires high soil fertility for optimal growth. The germplasm displays distinct skin color variations, including purplish (top) and greenish (bottom) phenotypes. Both variants feature a dense, solid, whitish interior of good quality, with no pithiness (hollow centers).
4.3. Ecosystem Services and Roles in Sustainable Cropping Systems
B. napus is increasingly recognized not only as an oilseed crop but also as a functional component of agroecosystems, especially when used as a cover crop or included in diversified rotations. Its contributions include improvements in soil structure, nutrient cycling, reduced nutrient losses, and suppression of soilborne diseases.
As a cover crop, B. napus improves soil physical properties. Cover crops can alleviate soil compaction and reduce its susceptibility to further compaction, although the extent of these effects depends on species, growth period, and root traits [100]. Brassica species with deep taproots, such as rapeseed, are able to penetrate compacted layers and thereby loosen the soil. In no-till systems, root penetration through compacted layers (0–50 cm) follows the order forage radish > rapeseed > rye [100]. After decomposition, these roots leave macropores that improve water infiltration, soil aeration, and root growth of subsequent crops [100,101]. Cover crops also reduce soil penetration resistance, with decreases of up to 65% reported in surface soils [102], and can reduce plow pan compaction over the long term [103].
Beyond structural effects, B. napus contributes to nutrient retention and recycling. Cover crops effectively take up residual soil nitrogen and reduce nitrate leaching. A synthesis of 16 studies reported reductions in NO3 leaching from 6% to 94%, depending on species and conditions [104], while global analyses estimate an average reduction of about 50% in irrigated systems [105,106]. Brassica species are particularly efficient in nitrogen uptake [107,108]. For example, rye can retain 46–56 kg·ha−1 of excess nitrogen in maize systems [109]. These nutrients are gradually released after termination, thereby improving nutrient use efficiency and limiting losses. Cover crops also take up phosphorus and convert it into organic forms, lowering soil P levels and reducing runoff risk [110]. Root traits further affect these processes. Rapeseed has an extensive lateral root system and a favorable C:N ratio, which supports efficient nutrient capture and recycling.
Including B. napus in crop rotations also provides clear agronomic and ecological benefits. Diverse rotations improve nutrient cycling, increase soil organic matter, and enhance the availability of nutrients such as nitrogen and potassium compared with monoculture systems [111]. They also support more diverse soil microbial communities, which are important for nutrient turnover and plant health. Canola hosts a complex rhizosphere microbiome that contributes to nutrient uptake, stress tolerance, and disease resistance. Root exudates are key drivers of these communities, shaping microbial composition and activity.
Rotation frequency is an important factor. Continuous or short rotations of canola are linked to higher abundance of soilborne pathogens such as Alternaria, Leptosphaeria, and Phaeomycocentrospora, and to increased disease incidence, including blackleg [111]. In contrast, more diverse rotations, such as canola–wheat or canola–pea–barley systems, favor beneficial microorganisms, including Pseudomonas, Serratia, and Penicillium, which contribute to nutrient solubilization and pathogen suppression. This highlights the importance of rotation design.
Brassica species can also suppress diseases when used in rotations or as green manure. Field studies show that Brassica crops can reduce diseases such as black scurf, common scab, and powdery scab in potato systems, with reductions ranging from 16% to 78% depending on conditions [112]. These effects are often linked not only to glucosinolate-derived biofumigation but also to broader changes in soil microbial communities. Similar effects have been reported in non-Brassica rotations, suggesting that microbial shifts and organic matter inputs play a central role.
Finally, ecosystem services can be further improved by using mixed-species cover crops. Because no single species provides all functions, combining Brassica species with legumes or grasses can take advantage of complementary traits such as nitrogen fixation, carbon input, root diversity, and weed suppression [113,114]. These mixtures enhance system resilience and productivity, reinforcing the role of B. napus in diversified cropping systems.
5. Challenges and Opportunities
5.1. Advancing Agronomic Technologies via UAV Remote Sensing
The practical use of multi-functional rapeseed is not only based on genetic innovation but also the concomitant creation of exact agronomic management. Unmanned aerial vehicles (UAVs), defined as remotely piloted or autonomous airborne platforms equipped with sensors for data acquisition, have emerged as a powerful tool in agricultural remote sensing. Classical approaches to field phenotyping can be time consuming and labor intensive, and thus must be slowly replaced by high-throughput phenotyping (HTP) [115]. The remote sensing technology of the UAV has turned out to be a revolution. Multi-source sensors and deep learning algorithms used in the UAV platforms present a chance to provide a database to predict complex yield variables and determine the non-destructive physiological conditions of crops, which could assist in the large-scale production of rapeseed (Figure 6).
One of the most important features of field nutrient management is the real-time monitoring of chlorophyll content because chlorophyll content is a good indicator of the nitrogen status and photosynthetic efficiency of plants [116]. The latest studies have demonstrated that with the help of background removal and feature fusion, it is possible to significantly increase the quality of the prediction of the chlorophyll content in rapeseed leaves. The sensitivity of the color and spectral indices to the canopy biochemical parameters was greatly improved using the ExG-ExR threshold segmentation technique to eliminate the soil and weed interference. Moreover, the gradual integration of RGB and multispectral characteristics enabled thorough canopy characterization, resulting in the development of powerful machine learning models (including support vector regression) that retained high predictive stability across growth phases under varying nitrogen conditions [117]. In addition to physiological monitoring, structural information based seed yield estimation is another future field in precision agriculture. In rapeseed, where the deep canopy cover and the dense pod cover are present, the common vertical overhead imaging device would tend to cause systematic underestimation of the yield. The 3D reconstruction of the organs and the segmentation of the biomass and volume can be highly correlated (R2 = 0.98), using Multi-view oblique imaging technology, 3D Gaussian scattering (3DGS), and Segment Anyone Model (SAM) to achieve the results. The algorithm is better than the structure-from- motion-based algorithms in the recovery of the canopy structure and lost pods [118]. This has replaced the black box spectral correlations with the biological basis organ counting and given genotype selection and harvesting strategies a resource that is understandable.
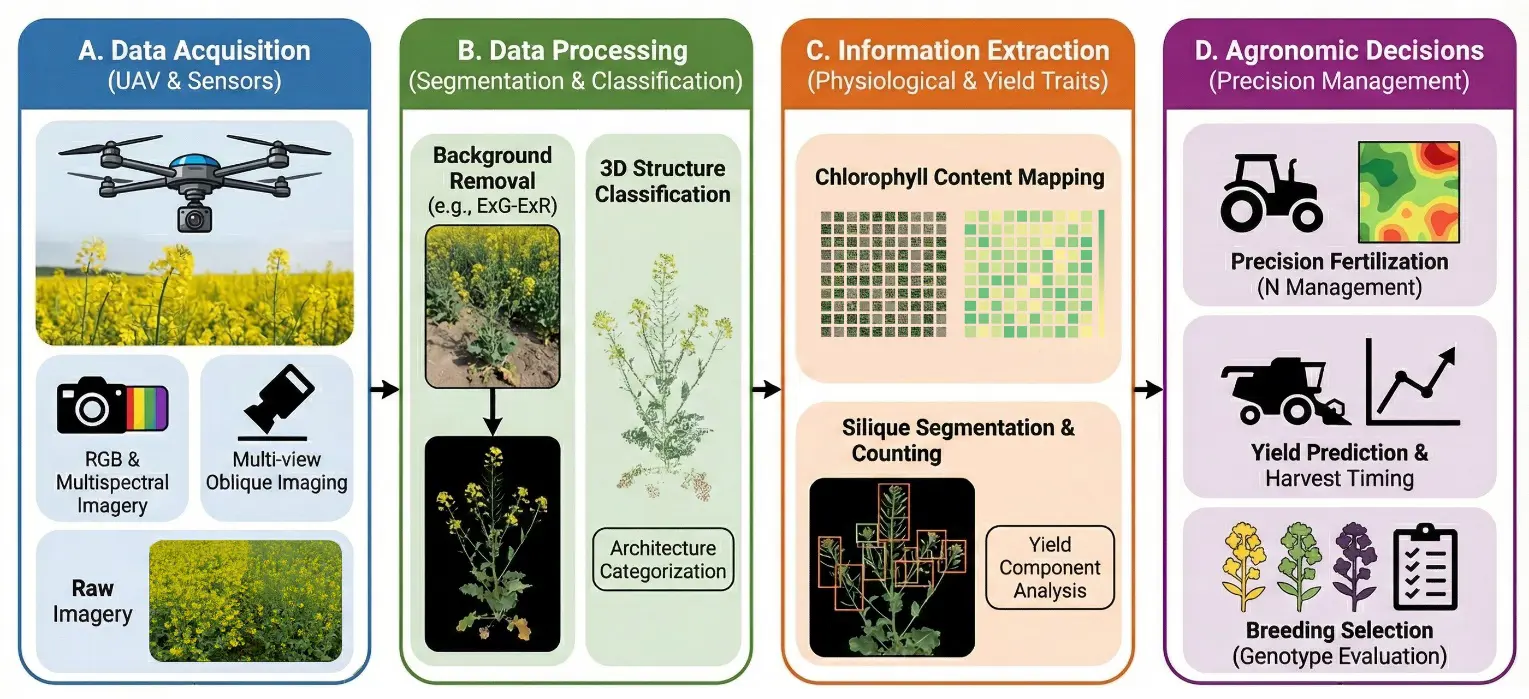
Figure 6. The precision agronomic management and the high throughput phenotyping of B. napus with UAVs. The pictorial workflow shows the transformation of the high-throughput data into action agronomic decisions. (A). Data Acquisition: The data is collected by the UAVs fitted with field sensors on high-resolution images. (B). Data Processing: Data processing of Soil background: Data is processed using tools to identify the shape of the plant. (C). Information Extraction: Software is applied to map yield (chlorophyll and siliques or pods). (D). Agronomic Decisions: Data can be used to assist in vindicating and harvesting time, plus the preferable varieties.
5.2. Challenges in Scaling the Multifunctional Rapeseed System
The transition of B. napus from a single-purpose oil crop to a multifunctional system that integrates food, energy, and ecosystem services is constrained by several factors, including biological trade-offs, gaps in the value chain, and the difficulty of evaluating ecological functions.
One of the main issues is the trade-off among yield, oil, and protein. Nitrogen input is a clear example. More nitrogen generally increases biomass and protein content, but oil accumulation often declines because carbon resources are shared between lipid and protein synthesis [119]. This pattern is well documented: higher nitrogen levels lead to lower oil content and higher protein content in rapeseed [120,121,122]. Nitrogen also affects key metabolic enzymes, such as ACCase, DGAT2, and PEPC, which, in turn, alter carbon allocation [123,124,125]. This makes it difficult to improve all target traits at the same time. Higher nitrogen inputs that increase yield can also increase lodging risk due to stem elongation and reduced mechanical strength [126]. Field evidence shows that yield gains under high-input conditions are often accompanied by reduced lodging resistance. In addition, yield components interact in compensatory ways. In B. napus, increases in silique number do not always translate into higher yield because of reductions in harvest index, and grain number and grain weight are often negatively associated [127,128]. At the ecological level, growth and defense are also in tension. Enhanced resistance to pathogens such as Sclerotinia sclerotiorum, for example, through pectin-related pathways, can come at the cost of growth and productivity [129,130]. Together, these patterns show that multifunctional optimization is constrained across metabolic, structural, and ecological levels.
Even if these biological constraints are addressed, practical challenges remain along the value chain. Rapeseed has potential uses in several sectors, but these are not well connected. In the protein sector, processing remains a bottleneck. Standard methods reduce protein quality, while improved technologies are still expensive and not widely used [131]. In the bioenergy sector, although rapeseed biomass could support renewable energy production, high costs and limited use of by-products remain major issues [132]. At the farm level, inefficient nitrogen use, often below 40%, further reduces economic returns and increases environmental risks [133].
Another challenge is that many ecosystem services are difficult to quantify. Rapeseed depends strongly on insect pollination, which contributes substantially to both yield and quality, with estimates suggesting that pollinators support 15–30% of agricultural production [134,135]. However, pollination is highly variable and sensitive to management practices and landscape context, making it difficult to standardize. In addition, services such as soil improvement, carbon cycling, and biodiversity support are difficult to quantify and incorporate into economic decision-making. This lack of measurable indicators limits the integration of ecological benefits into scalable production systems.
In response to these systemic constraints, our previous work on the Zheda rapeseed series provides a representative pathway to resolve the bottlenecks in multifunctional system scaling. By integrating genetic improvement with agronomic optimization and value-chain extension, these cultivars simultaneously enhance seed yield, oil content, and multi-purpose utilization. Furthermore, the development of ornamental rapeseed landscapes has enabled the expansion of rapeseed into the tertiary sector, creating a “flowering economy” that links agricultural production with rural tourism. This tri-sectoral integration illustrates a feasible model in which productivity, ecological sustainability, and diversified economic value are synergistically aligned, offering a practical solution to overcome the fragmentation and instability of conventional rapeseed value chains.
5.3. From Conventional Breeding to Intelligent Multi-Omics Design
Breeding in B. napus is shifting from phenotype-based selection toward data-informed design, driven by the complexity of traits such as yield, oil content, and stress tolerance. Traditional breeding, which relies on phenotypic selection and empirical crossing, has delivered substantial gains but remains limited in resolving complex trait interactions. Advances in genomics and high-throughput sequencing have enabled the identification of quantitative trait loci and candidate genes through GWAS and linkage mapping. Through GWAS and linkage mapping, many loci linked to yield, oil traits, and stress responses have been identified [136,137]. Even so, genomic data on its own does not fully account for phenotypic variation, which has led to increasing interest in integrating different types of omics data.
This need has driven the development of multi-omics approaches that combine genomics with transcriptomic, proteomic, and metabolomic data. Integrative analyses, such as those linking GWAS with TWAS and eQTL mapping, have improved the resolution of regulatory networks controlling oil biosynthesis and fatty acid composition [138]. These studies show that lipid metabolism is coordinated across pathways involved in carbon allocation, fatty acid synthesis, and triacylglycerol assembly, rather than being controlled by single enzymes [139]. Similar approaches have identified genes and pathways associated with yield components, including silique number, seed weight, and biomass accumulation [140]. These studies have shifted the focus from single genes to interacting networks.
At the same time, predictive breeding methods are becoming more widely used. Genomic selection allows breeders to estimate breeding values directly from genotype data, reducing reliance on long field trials. When combined with high-throughput phenotyping and environmental information, it has shown good performance in predicting yield stability and oil content in B. napus [141,142]. Machine learning approaches further improve prediction by identifying complex relationships among traits that are difficult to capture with conventional models [75].
Rather than focusing on individual traits, breeding efforts now aim to balance productivity, quality, and ecological performance. Multi-omics models can be used to examine carbon allocation among growth, storage lipids, and defense pathways, helping to address trade-offs such as yield versus oil content or growth versus stress resistance [143]. They also allow the design of varieties suited to different applications, including high-oleic oil, bioenergy use, and ornamental or ecological functions (Figure 7). Resources such as BnIR further support this process by integrating multi-omics datasets for trait prediction [144].
Even with these advances, several issues remain unresolved. Data integration is still challenging, especially when combining different types of omics data. Gene-environmental interactions are difficult to interpret, and predictive models do not always perform well under field conditions. More work is needed to standardize data, improve model transparency, and link molecular data with field performance. Continued advances in artificial intelligence and systems biology are expected to make it likely to design rapeseed varieties with coordinated improvements across multiple traits.
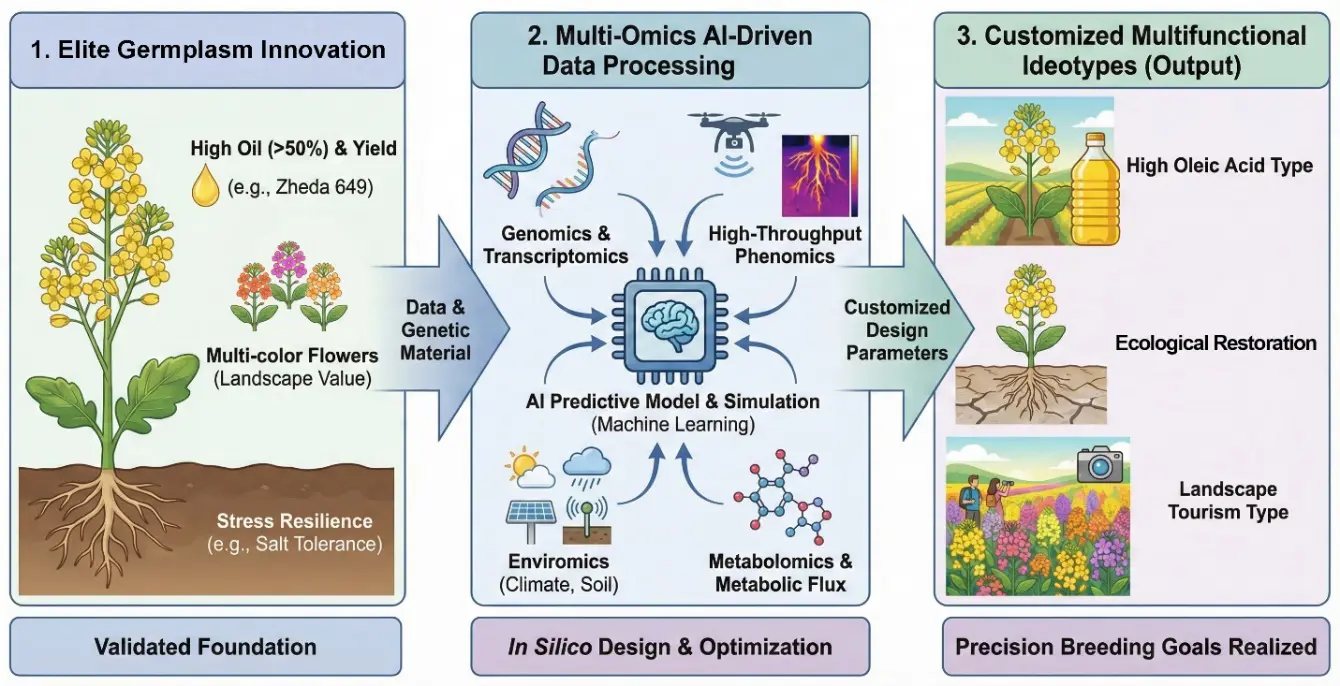
Figure 7. Future-oriented multi-omics intelligent breeding for Brassica napus. (Left): Utilizing high-quality basic materials, such as the “Zheda” series, these materials serve as a high-quality genetic foundation, possessing characteristics such as high oil content and diverse petal colors. (Middle): Multidimensional datasets, including genomics, digital phenomics, and environmental variables, will be integrated into advanced AI-driven predictive models. In this process, existing successful varieties will serve as crucial “ground truth” data and validation sets to improve machine-learning algorithms. (Right): The ultimate goal is to develop versatile, ideal varieties tailored to specific industrial and ecological scenarios, including specialty varieties with high oleic acid content, salt-tolerant varieties for land reclamation, and varieties with high ornamental value for landscape tourism.
6. Conclusions
B. napus cultivation is undergoing a paradigm shift, evolving from a single-purpose oil crop into a multipurpose and multifunctional ideotype. This review demonstrates the interactions between genomic knowledge, precision breeding techniques, and enhanced agronomic techniques in enhancing the value of this valuable crop. This is because the success of new varieties produced has shown that interspecific hybridization, marker-assisted selection, and gene pyramiding could beat the classical trade-off between high yield and high quality. With the implementation of landscape value, high-quality nutrition, and ecological and social benefits as potential applications of B. napus, the diverse demands within the scope of the Grand Food Concept will be met. However, additional development is still needed. The future of breeding must evolve from traditional experience-based strategies to intelligent design and data-driven approaches. It will demand the integration of multi-omics with AI-driven models of prediction to design cultivars in-silico to match the particular environment. To conclude, the utilization of the genetic, functional potential of B. napus can fully transform the crop into a major crop in green, sustainable, and diversified agriculture in the world.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the author(s) used Google Gemini AI tools to produce conceptual illustrative elements and icons only. After using this tool/service, the author(s) reviewed and edited the content as needed and take(s) full responsibility for the content of the article.
Acknowledgments
We thank Rui Sun and Ping Yang from Agricultural Experiment Station of Zhejiang University for their assistance.
Author Contributions
Y.Z.: Writing—Original draft preparation, Formal analysis, Visualization. I.B. and A.A.: Methodology, Validation, Formal analysis, Investigation, Data Curation, Writing—original draft; K.Z., F.H., Y.S. and T.Q.: Validation, Investigation, Data Curation; Q.H., L.X., W.X. and W.Q.: Investigation, Validation; W.Z.: Conceptualization, Methodology, Resources, Writing—review & editing, Supervision, Project administration, funding acquisition. All authors agreed with the publication of the manuscript.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data presented in this study is available in the article and supplementary materials.
Funding
This study was funded by Science and Technology Department of Zhejiang Province (grant number 2023C02002-3), Agriculture and Rural Affairs Department of Zhejiang Province (grant number 2023ZDXT01), and Collaborative Innovation Center for Modern Crop Production co-sponsored by Province and Ministry (CIC-MCP).
Declaration of Competing Interest
Author Wu Qian is employed by Wenzhou Jiayou Seed Industry Co., Ltd., Wenzhou 325014, China. The authors declare that this company was not involved in the literature search strategy, data interpretation, manuscript writing, or the decision to publish. The commercial affiliation did not influence the review’s conclusions or the presentation of information.
References
- Attia Z, Pogoda CS, Reinert S, Kane NC, Hulke BS. Breeding for sustainable oilseed crop yield and quality in a changing climate. Theor. Appl. Genet. 2021, 134, 1817–1827. DOI:10.1007/s00122-021-03770-w [Google Scholar]
- Grygier A. Mustard seeds as a bioactive component of food. Food Rev. Int. 2023, 39, 4088–4101. DOI:10.1080/87559129.2021.2015774 [Google Scholar]
- Hajinajaf N, Bakhsh AF, Shahsavar SK, Sanjarian F, Rahnama H. Boosting plant oil yields: The role of genetic engineering in industrial applications. Biofuel Res. J. 2024, 11, 2105–2145. DOI:10.18331/BRJ2024.11.2.5 [Google Scholar]
- Kumar V, Kaushik D, Kaur J, Rasane P, Sayyad A, Oz F, et al. Significance of Brassica crops in oilseed industry, crop improvement strategies in Brassica species. Appl. Sci. 2026, 1, 37–57. DOI:10.1007/978-981-95-3861-4_2 [Google Scholar]
- Gołębiewska K, Fraś A, Gołębiewski D. Rapeseed meal as a feed component in monogastric animal nutrition—A review. Ann. Anim. Sci. 2022, 22, 1163–1183. DOI:10.2478/aoas-2022-0020 [Google Scholar]
- Bansal A, Abrol DP. Diversity and abundance of insect pollinators affecting seed production in mustard (Brassica napus L.). J. Apic. 2023, 38, 315–323. DOI:10.17519/apiculture.2023.11.38.4.315 [Google Scholar]
- Rijal R, Sharm AK, Kumar A. Economic importance of Brassica crops. In Crop Improvement Strategies in Brassica Species: Applied Science; Springer: Berlin/Heidelberg, Germany, 2026; Volume 1, pp. 1–36. [Google Scholar]
- Sharma S, Bala M, Kaur G, Tayyab S, Feroz SR. Chemical composition of oil and cake of Brassica juncea: Implications on human and animal health. In The Brassica juncea Genome; Springer: Berlin/Heidelberg, Germany, 2022; pp. 29–55. [Google Scholar]
- Batool M, El-Badri AM, Hassan MU, Yang H, Wang C, Yan Z, et al. Drought stress in Brassica napus: Effects, tolerance mechanisms, and management strategies. J. Plant Growth Regul. 2023, 42, 21–45. DOI:10.1007/s00344-021-10542-9 [Google Scholar]
- Zhang D, Lu Y, Ma W, Zhao J. The epigenetic regulation of agronomic traits and environmental adaptability in Brassicas. Plant Cell Environ. 2025, 48, 8915–8927. DOI:10.1111/pce.70177 [Google Scholar]
- Tan Z, Han X, Dai C, Lu S, He H, Yao X, et al. Functional genomics of Brassica napus: Progress, challenges, and perspectives. J. Integr. Plant Biol. 2024, 66, 484–509. DOI:10.1111/jipb.13635 [Google Scholar]
- Chalhoub B, Denoeud F, Liu S, Parkin IAP, Tang H, Wang X, et al. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science 2014, 345, 950–953. DOI:10.1126/science.1253435 [Google Scholar]
- Zhang K, Mason AS, Farooq MA, Islam F, Quezada-Martinez D, Hu D, et al. Challenges and prospects for a potential allohexaploid Brassica crop. Theor. Appl. Genet. 2021, 134, 2711–2726. DOI:10.1007/s00122-021-03845-8 [Google Scholar]
- Wang T, Dijk AD, Bucher J, Liang J, Wu J, Bonnema G, et al. Interploidy introgression shaped adaptation during the origin and domestication history of Brassica napus. Mol. Biol. Evol. 2023, 40, 199. DOI:10.1093/molbev/msad199 [Google Scholar]
- Zhou J, Ma M, Zhang Q, Ni S, Zhao H, Wen J, et al. Genomic and epigenomic coordination maintains subgenome transcriptional balance in allotetraploid Brassica napus. Hortic. Res. 2025, 13, 266. DOI:10.1093/hr/uhaf266 [Google Scholar]
- Tong C, Kole C, Liu L, Cheng X, Huang J, Liu S. The asymmetrical evolution of the mesopolyploid Brassica oleracea genome. In The Brassica oleracea Genome; Springer: Berlin/Heidelberg, Germany, 2021. [Google Scholar]
- El-Esawi MA. Genetic diversity and evolution of Brassica genetic resources: From morphology to novel genomic technologies—A review. Plant Genet. Resour. 2017, 15, 388–399. DOI:10.1017/S1479262116000058 [Google Scholar]
- Admas T, Jiao S, Pan R, Zhang W. Pan-omics insights into abiotic stress responses: Bridging functional genomics and precision crop breeding. Funct. Integr. Genomics 2025, 25, 128. DOI:10.1007/s10142-025-01633-x [Google Scholar]
- Qian X, Zheng W, Hu J, Ma J, Sun M, Li Y, et al. Identification and expression analysis of DFR gene family in Brassica napus L. Plants 2023, 12, 2583. DOI:10.3390/plants12132583 [Google Scholar]
- Liang K. Genetic regulation of pigmentation in Brassica napus flowers. Comput. Mol. Biol. 2025, 15, 65–74. DOI:10.5376/cmb.2025.15.0006 [Google Scholar]
- Vu TT, Jeong CY, Nguyen HN, Lee D, Lee SA, Kim JH, et al. Characterization of Brassica napus flavonol synthase involved in flavonol biosynthesis in Brassica napus L. J. Agric. Food Chem. 2015, 63, 7819–7829. DOI:10.1021/acs.jafc.5b02994 [Google Scholar]
- Zhang B, Liu C, Wang Y, Yao X, Wang F, Wu J, et al. Disruption of a CAROTENOID CLEAVAGE DIOXYGENASE 4 gene converts flower colour from white to yellow in Brassica species. New Phytol. 2015, 206, 1513–1526. DOI:10.1111/nph.13335 [Google Scholar]
- Ye S, Hua S, Ma T, Ma X, Chen Y, Wu L, et al. Genetic and multi-omics analyses reveal BnaA07.PAP2 as the key gene conferring anthocyanin-based color in Brassica napus flowers. J. Exp. Bot. 2022, 73, 6630–6645. DOI:10.1093/jxb/erac312 [Google Scholar]
- Zhao C, Safdar LB, Xie M, Shi M, Dong Z, Yang L, et al. Mutation of the PHYTOENE DESATURASE 3 gene causes yellowish-white petals in Brassica napus. Crop J. 2021, 9, 1124–1134. DOI:10.1016/j.cj.2020.10.012 [Google Scholar]
- Liu Y, Ye S, Yuan G, Ma X, Heng S, Yi B, et al. Gene silencing of BnaA09.ZEP and BnaC09.ZEP confers orange color in Brassica napus flowers. Plant J. 2020, 104, 932–949. DOI:10.1111/tpj.14970 [Google Scholar]
- Lian J, Lu X, Yin N, Ma L, Lu J, Liu X, et al. Silencing of BnTT1 family genes affects seed flavonoid biosynthesis and alters seed fatty acid composition in Brassica napus. Plant Sci. 2017, 254, 32–47. DOI:10.1016/j.plantsci.2016.10.012 [Google Scholar]
- Xie T, Chen X, Guo T, Rong H, Chen Z, Sun Q, et al. Targeted knockout of BnTT2 homologues for yellow-seeded Brassica napus with reduced flavonoids and improved fatty acid composition. J. Agric. Food Chem. 2020, 68, 5676–5690. DOI:10.1021/acs.jafc.0c01126 [Google Scholar]
- Zhai Y, Yu K, Cai S, Hu L, Amoo O, Xu L, et al. Targeted mutagenesis of BnTT8 homologs controls yellow seed coat development for effective oil production in Brassica napus L. Plant Biotechnol. J. 2020, 18, 1153–1168. DOI:10.1111/pbi.13281 [Google Scholar]
- Calderwood A, Lloyd A, Hepworth J, Tudor EH, Jones DM, Woodhouse S, et al. Total FLC transcript dynamics from divergent paralogue expression explains flowering diversity in Brassica napus. New Phytol. 2021, 229, 3534–3548. DOI:10.1111/nph.17131 [Google Scholar]
- Jiang L, Li D, Jin L, Ruan Y, Shen WH, Liu C. Histone lysine methyltransferases BnaSDG8.A and BnaSDG8.C are involved in the floral transition in Brassica napus. Plant J. 2018, 95, 672–685. DOI:10.1111/tpj.13978 [Google Scholar]
- Yi L, Chen C, Yin S, Li H, Li Z, Wang B, et al. Sequence variation and functional analysis of a FRIGIDA orthologue (BnaA3.FRI) in Brassica napus. BMC Plant Biol. 2018, 18, 32. DOI:10.1186/s12870-018-1253-1 [Google Scholar]
- Khan MH, Hu L, Zhu M, Zhai Y, Khan SU, Ahmar S, et al. Targeted mutagenesis of EOD3 gene in Brassica napus L. regulates seed production. J. Cell Physiol. 2021, 236, 1996–2007. DOI:10.1002/jcp.29986 [Google Scholar]
- Liu J, Hua W, Hu Z, Yang H, Zhang L, Li R, et al. Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed. Proc. Natl. Acad. Sci. USA 2015, 112, 5123–5132. DOI:10.1073/pnas.1502160112 [Google Scholar]
- Wang JL, Tang MQ, Chen S, Zheng XF, Mo HX, Li SJ, et al. Down-regulation of BnDA1, whose gene locus is associated with the seeds weight, improves the seeds weight and organ size in Brassica napus. Plant Biotechnol. J. 2017, 15, 1024–1033. DOI:10.1111/pbi.12696 [Google Scholar]
- Yang Y, Zhu K, Li H, Han S, Meng Q, Khan SU, et al. Precise editing of CLAVATA genes in Brassica napus L. regulates multilocular silique development. Plant Biotechnol. J. 2018, 16, 1322–1335. DOI:10.1111/pbi.12872 [Google Scholar]
- Liu J, Zhou R, Wang W, Wang H, Qiu Y, Raman R, et al. A Copia-like retrotransposon insertion in the upstream region of the SHATTERPROOF1 gene, BnSHP1.A9, is associated with quantitative variation in pod shattering resistance in oilseed rape. J. Exp. Bot. 2020, 71, 5402–5413. DOI:10.1093/jxb/eraa281 [Google Scholar]
- Zaman QU, Chu W, Shi Y, Hao M, Mei D, Jacqueline B, et al. Characterization of SHATTERPROOF homoeologs and CRISPR-Cas9-mediated genome editing enhances pod-shattering resistance in Brassica napus L. CRISPR J. 2021, 4, 360–370. DOI:10.1089/crispr.2020.0129 [Google Scholar]
- Zhai Y, Cai S, Hu L, Yang Y, Amoo O, Fan C, et al. CRISPR/Cas9-mediated genome editing reveals differences in the contribution of INDEHISCENT homologues to pod shatter resistance in Brassica napus L. Theor. Appl. Genet. 2019, 132, 2111–2123. DOI:10.1007/s00122-019-03341-0 [Google Scholar]
- Braatz J, Harloff HJ, Mascher M, Stein N, Himmelbach A, Jung C. CRISPR-Cas9 targeted mutagenesis leads to simultaneous modification of different homoeologous gene copies in polyploid oilseed rape (Brassica napus). Plant Physiol. 2017, 174, 935–942. DOI:10.1104/pp.17.00426 [Google Scholar]
- Zaman QU, Chu W, Hao M, Shi Y, Sun M, Sang SF, et al. CRISPR/Cas9-mediated multiplex genome editing of JAGGED gene in Brassica napus L. Biomolecules 2019, 9, 725. DOI:10.3390/biom9110725 [Google Scholar]
- Zhang K, He J, Yin Y, Chen K, Deng X, Yu P, et al. Lysophosphatidic acid acyltransferase 2 and 5 commonly, but differently, promote seed oil accumulation in Brassica napus. Biotechnol. Biofuels Bioprod. 2022, 15, 83. DOI:10.1186/s13068-022-02182-2 [Google Scholar]
- Shi J, Lang C, Wang F, Wu X, Liu R, Zheng T, et al. Depressed expression of FAE1 and FAD2 genes modifies fatty acid profiles and storage compounds accumulation in Brassica napus seeds. Plant Sci. 2017, 263, 177–182. DOI:10.1016/j.plantsci.2017.07.014 [Google Scholar]
- Taylor DC, Zhang Y, Kumar A, Francis T, Giblin EM, Barton DL, et al. Molecular modification of triacylglycerol accumulation by over-expression of DGAT1 to produce canola with increased seed oil content under field conditions. Botany 2009, 87, 533–543. DOI:10.1139/B08-101 [Google Scholar]
- Chen X, Truksa M, Snyder CL, El-Mezawy A, Shah S, Weselake RJ. Three homologous genes encoding sn-glycerol-3-phosphate acyltransferase 4 exhibit different expression patterns and functional divergence in Brassica napus. Plant Physiol. 2011, 155, 851–865. DOI:10.1104/pp.110.169482 [Google Scholar]
- Haelterman L, Louvieaux J, Chiodi C, Bouchet AS, Kupcsik L, Stahl A, et al. Genetic control of root morphology in response to nitrogen across rapeseed diversity. Physiol. Plant 2024, 176, 14315. DOI:10.1111/ppl.14315 [Google Scholar]
- Ahmad N, Ibrahim S, Tian Z, Kuang L, Wang X, Wang H, et al. Quantitative trait loci mapping reveals important genomic regions controlling root architecture and shoot biomass under nitrogen, phosphorus, and potassium stress in rapeseed (Brassica napus L.). Front. Plant Sci. 2022, 13, 994666. DOI:10.3389/fpls.2022.994666 [Google Scholar]
- Feng Y, Cui R, Wang S, He M, Hua Y, Shi L, et al. Transcription factor BnaA9.WRKY47 contributes to the adaptation of Brassica napus to low boron stress by up-regulating the boric acid channel gene BnaA3.NIP5;1. Plant Biotechnol. J. 2020, 18, 1241–1254. DOI:10.1111/pbi.13288 [Google Scholar]
- Zhang Q, Chen H, He M, Zhao Z, Cai H, Ding G, et al. The boron transporter BnaC4.BOR1;1c is critical for inflorescence development and fertility under boron limitation in Brassica napus. Plant Cell Environ. 2017, 40, 1819–1833. DOI:10.1111/pce.12987 [Google Scholar]
- Sun Q, Lin L, Liu D, Wu D, Fang Y, Wu J, et al. CRISPR/Cas9-mediated multiplex genome editing of the BnWRKY11 and BnWRKY70 genes in Brassica napus L. Int. J. Mol. Sci. 2018, 19, 2716. DOI:10.3390/ijms19092716 [Google Scholar]
- Kagale S, Divi UK, Krochko JE, Keller WA, Krishna P. Brassinosteroid confers tolerance in Arabidopsis thaliana and Brassica napus to a range of abiotic stresses. Planta 2007, 225, 353–364. DOI:10.1007/s00425-006-0361-6 [Google Scholar]
- Xu P, Cai W. Functional characterization of the BnNCED3 gene in Brassica napus. Plant Sci. 2017, 256, 16–24. DOI:10.1016/j.plantsci.2016.11.012 [Google Scholar]
- Wang Z, Mao H, Dong C, Ji R, Cai L, Fu H, et al. Overexpression of Brassica napus MPK4 enhances resistance to Sclerotinia sclerotiorum in oilseed rape. Mol. Plant Microbe Interact. 2009, 22, 235–244. DOI:10.1094/MPMI-22-3-0235 [Google Scholar]
- Wang Z, Fang H, Chen Y, Chen K, Li G, Gu S, et al. Overexpression of BnWRKY33 in oilseed rape enhances resistance to Sclerotinia sclerotiorum. Mol. Plant Pathol. 2014, 15, 677–689. DOI:10.1111/mpp.12123 [Google Scholar]
- Potlakayala SD, DeLong C, Sharpe A, Fobert PR. Conservation of NON-EXPRESSOR OF PATHOGENESIS-RELATED GENES1 function between Arabidopsis thaliana and Brassica napus. Physiol. Mol. Plant Pathol. 2007, 71, 174–183. DOI:10.1016/j.pmpp.2008.01.003 [Google Scholar]
- Yan L, Zeng L, Raza A, Lv Y, Ding X, Cheng Y, et al. Inositol improves cold tolerance through inhibiting CBL1 and increasing Ca2+ influx in rapeseed (Brassica napus L.). Front. Plant Sci. 2022, 13, 775692. DOI:10.3389/fpls.2022.775692 [Google Scholar]
- Liang Z, Li M, Liu Z, Wang J. Genome-wide identification and characterization of the Hsp70 gene family in allopolyploid rapeseed (Brassica napus L.) compared with its diploid progenitors. PeerJ 2019, 7, e7511. DOI:10.7717/peerj.7511 [Google Scholar]
- Zhang T, Chang Y, Wang J, Wang N, Wang Y, Chen Q, et al. Cloning and expression analysis of a BnICE1 from Brassica napus L. Sci. Agric. Sin. 2013, 1, 205–214. 10.3864/j.issn.0578-1752.2013.01.024 [Google Scholar]
- Jhingan S, Harloff HJ, Abbadi A, Welsch C, Blümel M, Tasdemir D, et al. Reduced glucosinolate content in oilseed rape (Brassica napus L.) by random mutagenesis of BnMYB28 and BnCYP79F1 genes. Sci. Rep. 2023, 13, 2344. DOI:10.1038/s41598-023-28661-6 [Google Scholar]
- Nour-Eldin HH, Madsen SR, Engelen S, Jørgensen ME, Olsen CE, Andersen JS, et al. Reduction of antinutritional glucosinolates in Brassica oilseeds by mutation of genes encoding transporters. Nat. Biotechnol. 2017, 35, 377–382. DOI:10.1038/nbt.3823 [Google Scholar]
- Neal CS, Fredericks DP, Griffiths CA, Neale AD. The characterisation of AOP2: A gene associated with the biosynthesis of aliphatic alkenyl glucosinolates in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 170. DOI:10.1186/1471-2229-10-170 [Google Scholar]
- Miquel M, Trigui G, d’Andréa S, Kelemen Z, Baud S, Berger A, et al. Specialization of oleosins in oil body dynamics during seed development in Arabidopsis seeds. Plant Physiol. 2014, 164, 1866–1878. DOI:10.1104/pp.113.233262 [Google Scholar]
- Peng D, Zhou B, Jiang Y, Tan X, Yuan D, Zhang L. Enhancing freezing tolerance of Brassica napus L. by overexpression of a stearoyl-acyl carrier protein desaturase gene (SAD) from Sapium sebiferum (L.) Roxb. Plant Sci. 2018, 272, 32–41. DOI:10.1016/j.plantsci.2018.03.028 [Google Scholar]
- Huang KL, Li Y, Wang H, Tian J, Fu YF, Zheng Y, et al. Phosphorylation of BnLEC1 by BnSnRK2;2 is crucial for modulating lipid synthesis in seeds of Brassica napus. Seed Biol. 2024, 3, e009. DOI:10.48130/seedbio-0024-0009 [Google Scholar]
- Zeng DZ, Tian LS, Guo SX, Cai YF, Yang JP, Deng W, et al. Cloning and analysis of the phytoene synthase (BnPSY) gene in Brassica napus L. In Proceedings of the 13th International Rapeseed Congress, Prague, Czech Republic, 5–9 June 2011; pp. 928–932. [Google Scholar]
- Jiang J, Tian L, Guo S, Yu Q, Zeng D, Niu Y. Molecular cloning of BnZEP and its expression in petals of different colors in Brassica napus L. Turk. J. Agric. For. 2015, 39, 377–386. DOI:10.3906/tar-1403-17 [Google Scholar]
- Endrigkeit J. Identifikation und Charakterisierung von Genen der Tocopherol-Biosynthese aus Raps (Brassica napus L.). Ph.D. Dissertation, Christian-Albrechts-Universität zu Kiel, Kiel, Germany, 2008. [Google Scholar]
- Chikkaputtaiah C, Debbarma J, Baruah I, Havlickova L, Boruah HPD, Curn V. Molecular genetics and functional genomics of abiotic stress-responsive genes in oilseed rape (Brassica napus L.): A review of recent advances and future. Plant Biotechnol. Rep. 2017, 11, 365–384. DOI:10.1007/s11816-017-0458-3 [Google Scholar]
- Hu D, Jing J, Snowdon RJ, Mason AS, Shen J, Meng J, et al. Exploring the gene pool of Brassica napus by genomics-based approaches. Plant Biotechnol. J. 2021, 19, 1693–1712. DOI:10.1111/pbi.13636 [Google Scholar]
- Yu J, Hu F, Dossa K, Wang Z, Ke T. Genome-wide analysis of UDP-glycosyltransferase super family in Brassica rapa and Brassica oleracea reveals its evolutionary history and functional characterization. BMC Genom. 2017, 18, 474. DOI:10.1186/s12864-017-3844-x [Google Scholar]
- Kesidis CE. The effect of formation pathway on allopolyploids between Brassica carinata, Brassica napus, Brassica juncea and Sinapis arvensis. Ph.D. Dissertation, Carleton University, Ottawa, Canada, 2019. [Google Scholar]
- Quan C, Dou S, Dai C. The dynamics of ACR and DNA methylation impact asymmetric subgenome dominance in allotriploid Brassica species. bioRxiv 2025, 2025-02. DOI:10.1101/2025.02.16.638486 [Google Scholar]
- Fikere M, Barbulescu DM, Malmberg MM, Shi F, Koh JC, Slater AT, et al. Genomic prediction using prior quantitative trait loci information reveals a large reservoir of underutilised blackleg resistance in diverse canola (Brassica napus L.) lines. Plant Genome 2018, 11, 170100. DOI:10.3835/plantgenome2017.11.0100 [Google Scholar]
- Afzal M, Alghamdi SS, Rahman MH, Ahmad A, Farooq T, Alam M, et al. Current status and future possibilities of molecular genetics techniques in Brassica napus. Biotechnol. Lett. 2018, 40, 479–492. DOI:10.1007/s10529-018-2510-y [Google Scholar]
- Edukondalu B, Aswini N, Amaresh, Krishnappa G, Soundharya B, Nikhitha G, et al. Accelerating genetic gain through integrated genomic selection in crop plants. J. Appl. Genet. 2026, 67, 249–269. DOI:10.1007/s13353-025-01034-7 [Google Scholar]
- Gu J, Guan Z, Jiao Y, Liu K, Hong D. The story of a decade: Genomics, functional genomics, and molecular breeding in Brassica napus. Plant Commun. 2024, 5, 100884. DOI:10.1016/j.xplc.2024.100884 [Google Scholar]
- Cobb JN, Biswas PS, Platten JD. Back to the future: Revisiting MAS as a tool for modern plant breeding. Theor. Appl. Genet. 2019, 132, 647–667. DOI:10.1007/s00122-018-3266-4 [Google Scholar]
- Shi J, Ni X, Huang J, Fu Y, Wang T, Yu H, et al. CRISPR/Cas9-mediated gene editing of BnFAD2 and BnFAE1 modifies fatty acid profiles in Brassica napus. Genes 2022, 13, 1681. DOI:10.3390/genes13101681 [Google Scholar]
- Calderwood A, Siles L, Eastmond PJ, Kurup S, Morris RJ. A causal inference and Bayesian optimisation framework for modelling multi-trait relationships—Proof-of-concept using Brassica napus seed yield under controlled conditions. PLoS ONE 2023, 18, e0290429. DOI:10.3390/genes13101681 [Google Scholar]
- Anas M, Basri S, Chaurasiya N, Ali M. Artificial intelligence-assisted omics techniques in plant defense: Recent advancements and future prospects. In AI-Enhanced Plant Omics; CABI: Wallingford, UK, 2026; p. 143. [Google Scholar]
- Ijaz S, Iqbal J, Abbasi BA, Yaseen T, Rehman S, Kazi M, et al. Role of OMICS-based technologies in plant sciences. In OMICs-Based Techniques for Global Food Security; Wiley: Hoboken, NJ, USA, 2024; pp. 45–66. [Google Scholar]
- Liu H, Zhao W, Hua W, Liu J. A large-scale population based organelle pan-genomes construction and phylogeny analysis reveal the genetic diversity and the evolutionary origins of chloroplast and mitochondrion in Brassica napus L. BMC Genom. 2022, 23, 339. DOI:10.1186/s12864-022-08573-x [Google Scholar]
- Farooq MA, Hong Z, Islam F, Noor Y, Hannan F, Zhang Y, et al. Comprehensive proteomic analysis of arsenic induced toxicity reveals the mechanism of multilevel coordination of efficient defense and energy metabolism in two Brassica napus cultivars. Ecotoxicol. Environ. Saf. 2021, 208, 111744. DOI:10.1016/j.ecoenv.2020.111744 [Google Scholar]
- Li H, Che R, Zhu J, Yang X, Li J, Fernie AR, et al. Multi-omics-driven advances in the understanding of triacylglycerol biosynthesis in oil seeds. Plant J. 2024, 117, 999–1017. DOI:10.1111/tpj.16545 [Google Scholar]
- Zhang WJ, Tang LP, Peng J, Zhai LM, Ma QL, Zhang XS, et al. A WRI1-dependent module is essential for the accumulation of auxin and lipid in somatic embryogenesis of Arabidopsis thaliana. New Phytol. 2024, 242, 1098–1112. DOI:10.1111/nph.19689 [Google Scholar]
- Liew LC, You Y, Auroux L, Oliva M, Peirats-Llobet M, Ng S, et al. Establishment of single-cell transcriptional states during seed germination. Nat. Plants 2024, 10, 1418–1434. DOI:10.1038/s41477-024-01771-3 [Google Scholar]
- Lee TA, Illouz-Eliaz N, Nobori T, Xu J, Jow B, Nery JR, et al. A single-cell, spatial transcriptomic atlas of the Arabidopsis life cycle. Nat. Plants 2025, 11, 1960–1975. DOI:10.1038/s41477-025-02072-z [Google Scholar]
- Ma N, Yuan J, Li M, Li J, Zhang L, Liu L, et al. Ideotype population exploration: Growth, photosynthesis, and yield components at different planting densities in winter oilseed rape (Brassica napus L.). PLoS ONE 2014, 9, e114232. DOI:10.1371/journal.pone.0114232 [Google Scholar]
- Wu Y, Xie L. AI-driven multi-omics integration for multi-scale predictive modeling of genotype-environment-phenotype relationships. Comput. Struct. Biotechnol. J. 2025, 27, 265–277. DOI:10.1016/j.csbj.2024.12.030 [Google Scholar]
- Norouzi MA, Ahangar L, Payghamzadeh K, Sabouri H, Sajadi SJ. Investigation of genetic diversity of different spring rapeseed (Brassica napus L.) genotypes and yield prediction using machine learning models. Genet. Resour. Crop Evol. 2024, 71, 4519–4532. DOI:10.1007/s10722-024-01915-6 [Google Scholar]
- Sharma SS, Pandey A, Kashyap A, Goyal L, Garg P, Kushwaha R, et al. CRISPR/Cas9: Efficient and emerging scope for Brassica crop improvement. Planta 2025, 262, 14. DOI:10.1007/s00425-025-04727-9 [Google Scholar]
- Chen YY, Lu HQ, Jiang KX, Wang YR, Wang YP, Jiang JJ. The flavonoid biosynthesis and regulation in Brassica napus: A review. Int. J. Mol. Sci. 2022, 24, 357. DOI:10.3390/ijms24010357 [Google Scholar]
- Li F, Gong Y, Mason AS, Liu Q, Huang J, Ma M, et al. Research progress and applications of colorful Brassica crops. Planta 2023, 258, 45. DOI:10.1007/s00425-023-04205-0 [Google Scholar]
- Zhang K, Yang S, Zou J, Huang Q, Chen S, Yan G, et al. Genome-wide association study reveals genomic regions impacting yield-related traits in allohexaploid Brassica with AABBCC genomes. Ann. Bot. 2025, mcaf309. DOI:10.1093/aob/mcaf309 [Google Scholar]
- Hearn DJ, O’Brien P, Poulsen SM. Comparative transcriptomics reveals shared gene expression changes during independent evolutionary origins of stem and hypocotyl/root tubers in Brassica (Brassicaceae). PLoS ONE 2018, 13, e0197166. DOI:10.1371/journal.pone.0197166 [Google Scholar]
- Nuruzzaman M, Sato M, Okamoto S, Hoque M, Shea DJ, Fujimoto R, et al. Comparative transcriptome analysis during tuberous stem formation in kohlrabi (B. oleracea var. gongylodes) at early growth periods (seedling stages). Physiol. Plant 2022, 174, e13770. DOI:10.1111/ppl.13770 [Google Scholar]
- Liu M, Bassetti N, Petrasch S, Zhang N, Bucher J, Shen S, et al. What makes turnips: Anatomy, physiology and transcriptome during early stages of its hypocotyl-tuber development. Hortic. Res. 2019, 6, 38. DOI:10.1038/s41438-019-0119-5 [Google Scholar]
- Yuan R, Zeng X, Zhao S, Wu G, Yan X. Identification of candidate genes related to stem development in Brassica napus using RNA-Seq. Plant Mol. Biol. Rep. 2019, 37, 347–364. DOI:10.1007/s11105-019-01158-1 [Google Scholar]
- Yang R, Wang Q, Wang J, Wang X, Zhao J, Li N, et al. JmjC protein-mediated histone demethylation: Regulating growth, development, and stress adaptation in Brassica rapa. Horticulturae 2025, 11, 1424. DOI:10.3390/horticulturae11121424 [Google Scholar]
- Chen G, Weil RR. Penetration of cover crop roots through compacted soils. Plant Soil. 2010, 331, 31–43. DOI:10.1007/s11104-009-0223-7 [Google Scholar]
- Cresswell HP, Kirkegaard JA. Subsoil amelioration by plant-roots: The process and the evidence. Aust. J. Soil. Res. 1995, 33, 221–239. DOI:10.1071/SR9950221 [Google Scholar]
- Folorunso OA, Rolston DE, Prichard T, Loui DT. Soil surface strength and infiltration rate as affected by winter cover crops. Soil. Technol. 1992, 5, 189–197. DOI:10.1016/0933-3630(92)90021-R [Google Scholar]
- Abdollahi L, Munkholm LJ. Tillage system and cover crop effects on soil quality: I. Chemical, mechanical, and biological properties. Soil. Sci. Soc. Am. J. 2014, 78, 262–270. DOI:10.2136/sssaj2013.07.0301 [Google Scholar]
- Blanco-Canqui H, Shaver TM, Lindquist JL, Shapiro CA, Elmore RW, Francis CA, et al. Cover crops and ecosystem services: Insights from studies in temperate soils. Agron. J. 2015, 107, 2449–2474. DOI:10.2134/agronj15.0086 [Google Scholar]
- Quemada M, Baranski M, Nobel-de Lange MNJ, Vallejo A, Cooper JM. Meta-analysis of strategies to control nitrate leaching in irrigated agricultural systems and their effects on crop yield. Agric. Ecosyst. Environ. 2013, 174, 1–10. DOI:10.1016/j.agee.2013.04.018 [Google Scholar]
- Tonitto C, David MÁ, Drinkwater LE. Replacing bare fallows with cover crops in fertilizer-intensive cropping systems: A meta-analysis of crop yield and N dynamics. Agric. Ecosyst. Environ. 2006, 112, 58–72. DOI:10.1016/j.agee.2005.07.003 [Google Scholar]
- Kristensen HL, Thorup-Kristensen K. Root growth and nitrate uptake of three different catch crops in deep soil layers. Soil. Sci. Soc. Am. J. 2004, 68, 529–537. DOI:10.2136/sssaj2004.5290 [Google Scholar]
- Dabney SM, Delgado JA, Reeves DW. Using winter cover crops to improve soil and water quality. Commun. Soil. Sci. Plant Anal. 2001, 32, 1221–1250. DOI:10.1081/CSS-100104110 [Google Scholar]
- Rasse DP, Ritchie JT, Peterson WR, Wei J, Smucker AJ. Rye cover crop and nitrogen fertilization effects on nitrate leaching in inbred maize fields. Agron. J. 2000, 29, 298–304. DOI:10.2134/jeq2000.00472425002900010037x [Google Scholar]
- Villamil MB, Bollero GA, Darmody RG, Simmons FW, Bullock DG. No-till corn/soybean systems including winter cover crops: Effects on soil properties. Soil. Sci. Soc. Am. J. 2006, 70, 1936–1944. DOI:10.2136/sssaj2005.0350 [Google Scholar]
- Town JR, Dumonceaux T, Tidemann B, Helgason BL. Crop rotation significantly influences the composition of soil, rhizosphere, and root microbiota in canola (Brassica napus L.). Environ. Microbiome 2023, 18, 40. DOI:10.1186/s40793-023-00495-9 [Google Scholar]
- Larkin RP, Lynch RP. Use and effects of different Brassica and other rotation crops on soilborne diseases and yield of potato. Horticulturae 2018, 4, 37. DOI:10.3390/horticulturae4040037 [Google Scholar]
- Kramberger B, Gselman A, Kristl J, Lešnik M, Šuštar V, Muršec M, et al. Winter cover crop: The effects of grass–clover mixture proportion and biomass management on maize and the apparent residual N in the soil. Eur. J. Agron. 2014, 55, 63–71. DOI:10.1016/j.eja.2014.01.001 [Google Scholar]
- Tosti G, Benincasa P, Farneselli M, Tei F, Guiducci M. Barley–hairy vetch mixture as cover crop for green manuring and the mitigation of N leaching risk. Eur. J. Agron. 2014, 54, 34–39. DOI:10.1016/j.eja.2013.11.012 [Google Scholar]
- Angidi S, Madankar K, Tehseen MM, Bhatla A. Advanced high-throughput phenotyping techniques for managing abiotic stress in agricultural crops: A comprehensive review. Crops 2025, 5, 8. DOI:10.3390/crops5020008 [Google Scholar]
- Meena V, Dotaniya ML, Meena MD, Jat RS, Meena MK, Choudhary RL, et al. Sensor based precise nitrogen application augmented productivity and profitability of mustard (Brassica juncea L.). PLoS ONE 2024, 19, e0304206. DOI:10.1371/journal.pone.0304206 [Google Scholar]
- Li J, Xie T, Chen Y, Zhang Y, Wang C, Jiang Z, et al. High-throughput unmanned aerial vehicle-based phenotyping provides insights into the dynamic process and genetic basis of rapeseed waterlogging response in the field. J. Exp. Bot. 2022, 73, 5264–5278. DOI:10.1093/jxb/erac242 [Google Scholar]
- Shen Y, Zhou H, Yang X, Lu X, Guo Z, Jiang L, et al. Biomass phenotyping of oilseed rape through UAV multi-view oblique imaging with 3DGS and SAM model. Comput. Electron. Agric. 2025, 235, 110320. DOI:10.1016/j.compag.2025.110320 [Google Scholar]
- Yu S, Du S, Yuan J, Hu Y. Fatty acid profile in the seeds and seed tissues of Paeonia L. species as new oil plant resources. Sci. Rep. 2016, 6, 26944. DOI:10.1038/srep26944 [Google Scholar]
- Zhao Y, Ning P, Feng X, Ren H, Cui M, Yang L. Characterization of stem nodes associated with carbon partitioning in maize in response to nitrogen availability. Int. J. Mol. Sci. 2022, 23, 4389. DOI:10.3390/ijms23084389 [Google Scholar]
- Nunes-Nesi A, Fernie AR, Stitt M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. DOI:10.1093/mp/ssq049 [Google Scholar]
- Tang M, Guschina IA, O’Hara P, Slabas AR, Quant PA, Fawcett T, et al. Metabolic control analysis of developing oilseed rape (Brassica napus cv. Westar) embryos shows that lipid assembly exerts significant control over oil accumulation. New Phytol. 2012, 196, 414–426. DOI:10.1111/j.1469-8137.2012.04262.x [Google Scholar]
- Zhai J, Xue J, Zhang Y, Zhang G, Shen D, Wang Q, et al. Effect of nitrogen application rate on lodging resistance of spring maize stalks under integrated irrigation with water and fertilizer. J. Maize Sci. 2021, 29, 137–144. 10.13597/j.cnki.maize.science.20210518 [Google Scholar]
- Liu Q, Wu K, Harberd NP, Fu X. Green revolution DELLAs: From translational reinitiation to future sustainable agriculture. Mol. Plant 2021, 14, 547–549. DOI:10.1016/j.molp.2021.03.015 [Google Scholar]
- Zhu J, Dai W, Chen B, Cai G, Wu X, Yan G. Research progress on the effect of nitrogen on rapeseed between seed yield and oil content and its regulation mechanism. Int. J. Mol. Sci. 2023, 24, 14504. DOI:10.3390/ijms241914504 [Google Scholar]
- Wu W, Shah F, Duncan RW, Ma BL. Grain yield, root growth habit and lodging of eight oilseed rape genotypes in response to a short period of heat stress during flowering. Agric. For. Meteorol. 2020, 287, 107954. DOI:10.1016/j.agrformet.2020.107954 [Google Scholar]
- Diepenbrock W. Yield analysis of winter oilseed rape (Brassica napus L.): A review. Field Crops Res. 2000, 67, 35–49. DOI:10.1016/S0378-4290(00)00082-4 [Google Scholar]
- Slafer GA, Savin R, Sadras VO. Coarse and fine regulation of wheat yield components in response to genotype and environment. Field Crops Res. 2014, 157, 71–83. DOI:10.1016/j.fcr.2013.12.004 [Google Scholar]
- Wang D, Jin S, Chen Z, Shan Y, Li L. Genome-wide identification of the pectin methylesterase inhibitor genes in Brassica napus and expression analysis of selected members. Front. Plant Sci. 2022, 13, 940284. DOI:10.3389/fpls.2022.940284 [Google Scholar]
- Wang D, Lu Q, Jin S, Fan X, Ling H. Pectin, lignin and disease resistance in Brassica napus L.: An update. Horticulturae 2023, 9, 112. DOI:10.3390/horticulturae9010112 [Google Scholar]
- Mupondwa E, Li X, Wanasundara JP. Technoeconomic prospects for commercialization of Brassica (cruciferous) plant proteins. J. Am. Oil Chem. Soc. 2018, 95, 903–922. DOI:10.1002/aocs.12057 [Google Scholar]
- Suchocki T. Energy utilization of rapeseed biomass in Europe: A review of current and innovative applications. Energies 2024, 17, 6177. DOI:10.3390/en17236177 [Google Scholar]
- Herath A, Ma BL, Shang J, Liu J, Dong T, Jiao X, et al. On-farm spatial characterization of soil mineral nitrogen, crop growth, and yield of canola as affected by different rates of nitrogen application. Can. J. Soil. Sci. 2017, 98, 1–14. DOI:10.1139/CJSS-2017-0024 [Google Scholar]
- Klein AM, Vaissière BE, Cane JH, Steffan-Dewenter I, Cunningham SA, Kremen C, et al. Importance of pollinators in changing landscapes for world crops. Proc. R. Soc. B 2007, 274, 303–313. DOI:10.1098/rspb.2006.3721 [Google Scholar]
- Halder S, Ghosh S, Khan R, Khan AA, Perween T, Hasan MA. Role of pollination in fruit crops: A review. Pharma Innov. J. 2019, 8, 695–702. Available online: https://www.researchgate.net/profile/Shuvadeep-Halder/publication/333532191_Role_of_pollination_in_fruit_crops_A_review/links/5cf23e8e92851c4dd01f855b/Role-of-pollination-in-fruit-crops-A-review.pdf (accessed on 28 January 2026).
- Zandberg JD, Fernandez CT, Danilewicz MF, Thomas WJ, Edwards D, Batley J. The global assessment of oilseed Brassica crop species yield, yield stability and the underlying genetics. Plants 2022, 11, 2740. DOI:10.3390/plants11202740 [Google Scholar]
- Zhang Y, Zhang H, Zhao H, Xia Y, Zheng X, Fan R, et al. Multi-omics analysis dissects the genetic architecture of seed coat content in Brassica napus. Genome Biol. 2022, 23, 86. DOI:10.1186/s13059-022-02647-5 [Google Scholar]
- Zhao Q, Wang F, Xiong A, Li S, Wang Y, Lei X, et al. Regulation of fatty acid synthesis in oilseed crops: Multidimensional insights and strategies for enhancing oil quality. Crit. Rev. Biotechnol. 2025, 45, 1737–1753. DOI:10.1080/07388551.2025.2529591 [Google Scholar]
- Bu M, Fan W, Li R, He B, Cui P. Lipid metabolism and improvement in oilseed crops: Recent advances in multi-omics studies. Metabolites 2023, 13, 1170. DOI:10.3390/metabo13121170 [Google Scholar]
- Zhang C, Gong R, Zhong H, Dai C, Zhang R, Dong J, et al. Integrated multi-locus genome-wide association studies and transcriptome analysis for seed yield and yield-related traits in Brassica napus. Front. Plant Sci. 2023, 14, 1153000. DOI:10.3389/fpls.2023.1153000 [Google Scholar]
- Raza A, Razzaq A, Mehmood SS, Hussain MA, Wei S, He H, et al. Omics: The way forward to enhance abiotic stress tolerance in Brassica napus L. GM Crops Food 2021, 12, 251–281. DOI:10.1080/21645698.2020.1859898 [Google Scholar]
- Mahmood U, Li X, Fan Y, Chang W, Niu Y, Li J, et al. Multi-omics revolution to promote plant breeding efficiency. Front. Plant Sci. 2022, 13, 1062952. DOI:10.3389/fpls.2022.1062952 [Google Scholar]
- Gao G, Zhang L, Tong P, Yan G, Wu X. Enhancing oil content in oilseed crops: Genetic insights, molecular mechanisms, and breeding approaches. Int. J. Mol. Sci. 2025, 26, 7390. DOI:10.3390/ijms26157390 [Google Scholar]
- Yu W, Wang X, Wang H, Wang W, Cheng H, Mei D, et al. Optimization and application of genome prediction model in rapeseed: Flowering time, yield components, and oil content as examples. Hortic. Res. 2025, 12, uhaf115. DOI:10.1093/hr/uhaf115 [Google Scholar]
- Yang Z, Wang S, Wei L, Huang Y, Liu D, Jia Y, et al. BnIR: A multi-omics database with various tools for Brassica napus research and breeding. Mol. Plant 2023, 16, 775–789. DOI:10.1016/j.molp.2023.03.007 [Google Scholar]