Progress and Prospects in Breeding Research on Key Aromatic Species of the Lamiaceae Family

Progress and Prospects in Breeding Research on Key Aromatic Species of the Lamiaceae Family
Shixiao Chen 1,2 Yuanyuan Feng 1 Rui Fan 1 Xuejun Li 3 Chaoyun Hao 1,4,5,* Yanli Huang 2,*
Received: 19 March 2026 Revised: 28 April 2026 Accepted: 18 May 2026 Published: 29 May 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Introduction
The Lamiaceae family represents one of the most species-rich lineages of angiosperms [1]. Within this family, the subfamily Nepetoideae comprises several economically important aromatic genera, including Mentha, Ocimum, Origanum, Thymus, Rosmarinus, Lavandula, and Perilla [1]. Quadrangular stems and opposite leaves typically characterize these plants, and they possess specialized glandular structures that synthesize essential oils through diverse secondary metabolic pathways [2]. These essential oils, composed mainly of terpenoids, phenolic compounds, and other volatile constituents, not only confer characteristic aromas but also exhibit a wide range of bioactivities, including anti-inflammatory, antibacterial, antioxidant, and digestive-promoting effects [3]. These properties constitute the biochemical basis for their utilization in food, medicine, health care, and related industries [4].
Phylogenetic evidence suggests that the high species diversity of aromatic Lamiaceae plants is closely associated with multiple centers of origin, and that diversification shows clear geographical patterns. Mentha, Lavandula, Origanum, Rosmarinus, and Thymus originated predominantly in the Mediterranean region, whereas Ocimum and Perilla originated mainly in the mountainous regions of tropical Asia and East Asia [5,6,7,8,9,10,11]. These native distribution patterns shaped the chronological order in which different aromatic plants were discovered, utilized, and disseminated by humans. The use of aromatic herbs from the Lamiaceae family dates back thousands of years. In classical civilizations, the ancient Egyptians, Greeks, and Romans used rosemary, thyme, oregano, and lavender for ritual incense, mummification, aromatic bathing, purification, and healing practices [9,12,13,14]. Mint and basil were also recorded for their use in relieving gastrointestinal discomfort and promoting physical and mental well-being [15]. Basil was regarded as a sacred herb in ancient India [6], whereas perilla has long been used for both medicinal [16] and culinary purposes in China and later became established in Japan, Korea, and Southeast Asia through cultural exchange within the East Asian cultural sphere [16,17].
The subsequent dissemination of these aromatic plants was facilitated by military expansion, long-distance trade, religious and cultural exchange, and colonial migration. Mediterranean herbs gradually spread across Europe, the Americas, and Oceania through the expansion of Roman legions, trade along the Silk Road, monastic exchange, and European colonial activities. In parallel, Asian-origin basil and perilla expanded outward along ancient spice routes and regional cultural networks, becoming integrated into local culinary traditions, folk medicine, garden cultivation, and symbolic systems. With the development of plant chemistry and pharmacology in modern times, the practical value of these species has been increasingly supported by scientific evidence. In the food sector, mint, basil, and oregano have become widely used natural herbs in teas, desserts, Western cuisine, and meat processing [18]. Rosemary essential oil, owing to its strong antioxidant activity, has been developed as a natural food preservative capable of extending shelf life [19]. In medicine and health-related applications, menthol is widely used in throat lozenges and topical anti-pruritic formulations; rosemary extracts are incorporated into functional dietary supplements; and lavender essential oil is widely applied in aromatherapy for emotional regulation and sleep improvement [20]. The traditional medicinal value of thyme and mint has also been supported by modern pharmacological studies, particularly with respect to their antibacterial, antiviral, and immunomodulatory activities [14].
Driven by advances in breeding technologies and the deep processing of natural products, cultivars with enhanced aroma profiles and improved stress tolerance have continued to emerge. The value of aromatic plants in the Lamiaceae family is therefore no longer limited to traditional uses in ritual practices, medicine, seasoning, and cultural symbolism. Instead, their applications are expanding into natural medicines, functional foods, eco-friendly personal care products, and the aromatherapy industry. Meanwhile, because of their strong adaptability, some species have become naturalized and spread in introduced regions [21], forming a plant group with combined economic value, ecological relevance, and cultural significance.
This review systematically summarizes the global breeding history of these representative aromatic Lamiaceae genera, integrates their utilization and dissemination trajectories, and divides their development into four major phases. According to Figure 1, the first phase is the pre-breeding stage (before 1000 BCE) [6,13,15], driven primarily by basic human survival needs, during which wild resources were utilized directly without the development of artificial cultivation or directed selection; The second stage is the early introduction and preliminary domestication stage (1000–500 BCE), during which the expansion of ancient trade facilitated the cross-regional dissemination of species, and the domestication of germplasm began through simple phenotypic selection under artificial cultivation; The third phase is the conventional breeding stage, from 500 BCE to the late 20th century, which was driven by increasing commercial demand. During this period, clonal selection [22,23], phenotypic selection, and hybridization were gradually developed and widely applied [24], enabling the stable retention of desirable traits and the formation of diverse regionally distinctive local germplasm. The fourth phase is the modern molecular breeding stage, from the 21st century to the present, which has developed alongside scientific and technological advances. This stage includes molecular breeding strategies based on genome sequencing [25], identification of genes associated with essential oil biosynthesis and stress tolerance [2,26], and marker-assisted selection [27].
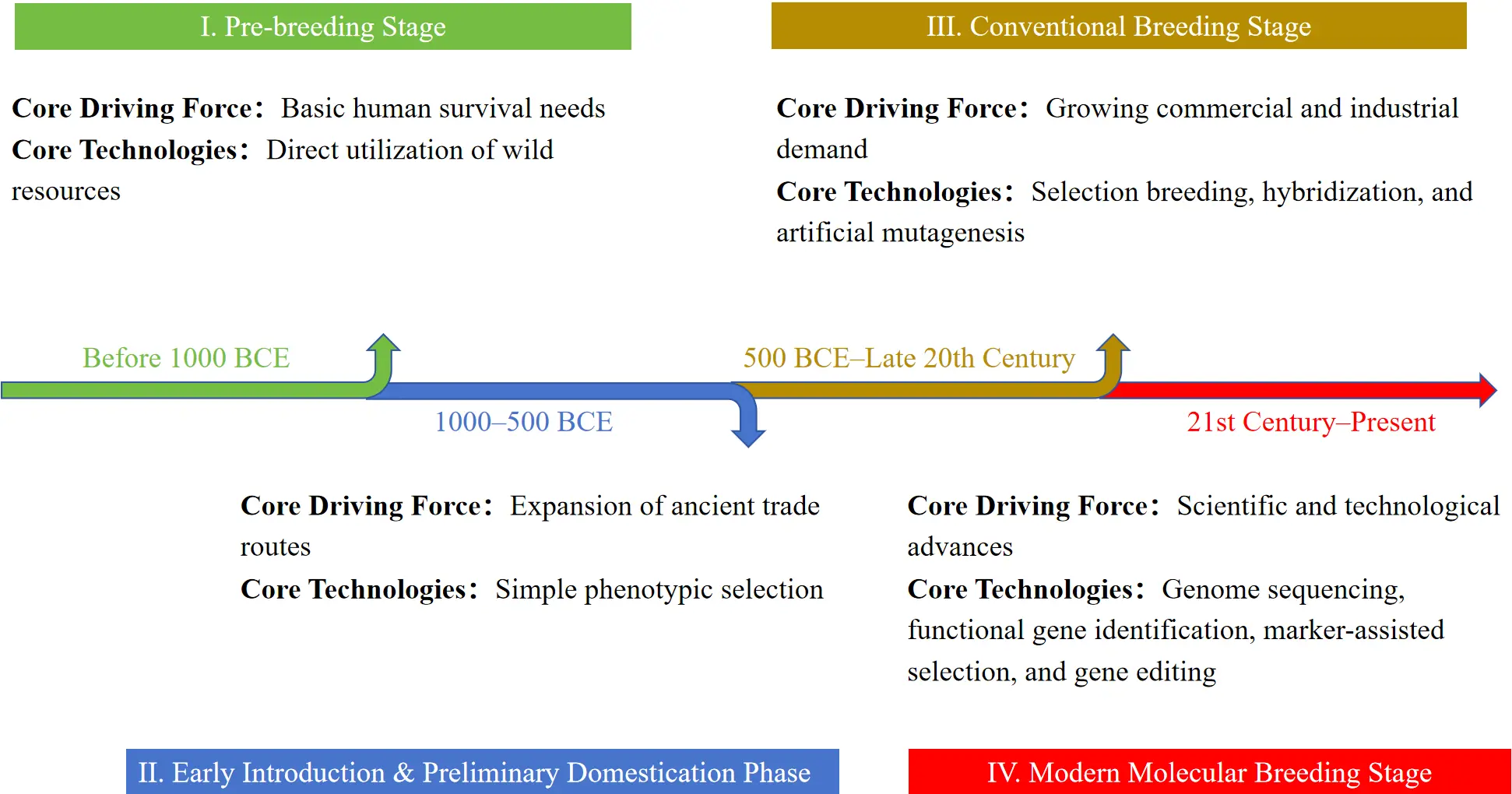
Despite substantial progress in the breeding of these key aromatic plant genera, their commercial improvement still faces several major constraints [28]. These include low genetic transformation efficiency in most species [29], limited genomic resources for underutilized taxa [25], long breeding cycles associated with traditional selection [30], the lack of comprehensive germplasm evaluation systems [31], and fragmented phenotype–genotype association databases [27]. Future research should prioritize: (1) the establishment of globally standardized databases for aromatic Lamiaceae germplasm resources [32]; (2) the integration of transcriptomics, metabolomics, proteomics, and other multi-omics approaches to elucidate the genetic regulatory networks underlying essential oil biosynthesis and stress resistance [33]; and (3) the optimization of gene-editing and genetic transformation systems for both major and underutilized aromatic Lamiaceae species [34,35].
2. Global Distribution, Collection, and Conservation of Lamiaceae Aromatic Plant Germplasm Resources
2.1. Origin and Spread of Crops
The origin and dissemination of key aromatic genera in Lamiaceae exhibit characteristics of “polycentric origin and gradient diffusion”. Their cross-regional spread was not an isolated botanical phenomenon, but rather closely associated with major geopolitical and economic transitions in human history. From the integration of Europe under the Roman Empire to the global expansion of European colonialism, the dissemination trajectories of these plants reflect the processes of imperial expansion, the development of trade networks, and intercultural exchange (Figure 2). Based on their historical and geographical patterns of spread, this study proposes two major “dissemination complexes”. The first is the Mediterranean dissemination complex, represented by mint, rosemary, lavender, thyme, and oregano, whose early expansion was promoted by the Roman Empire and later globalized through European colonial powers. The second is the Asian dissemination complex, represented by basil and perilla, whose dissemination pathways differed markedly, with westward spread along ancient spice trade routes toward the Mediterranean world [36,37].
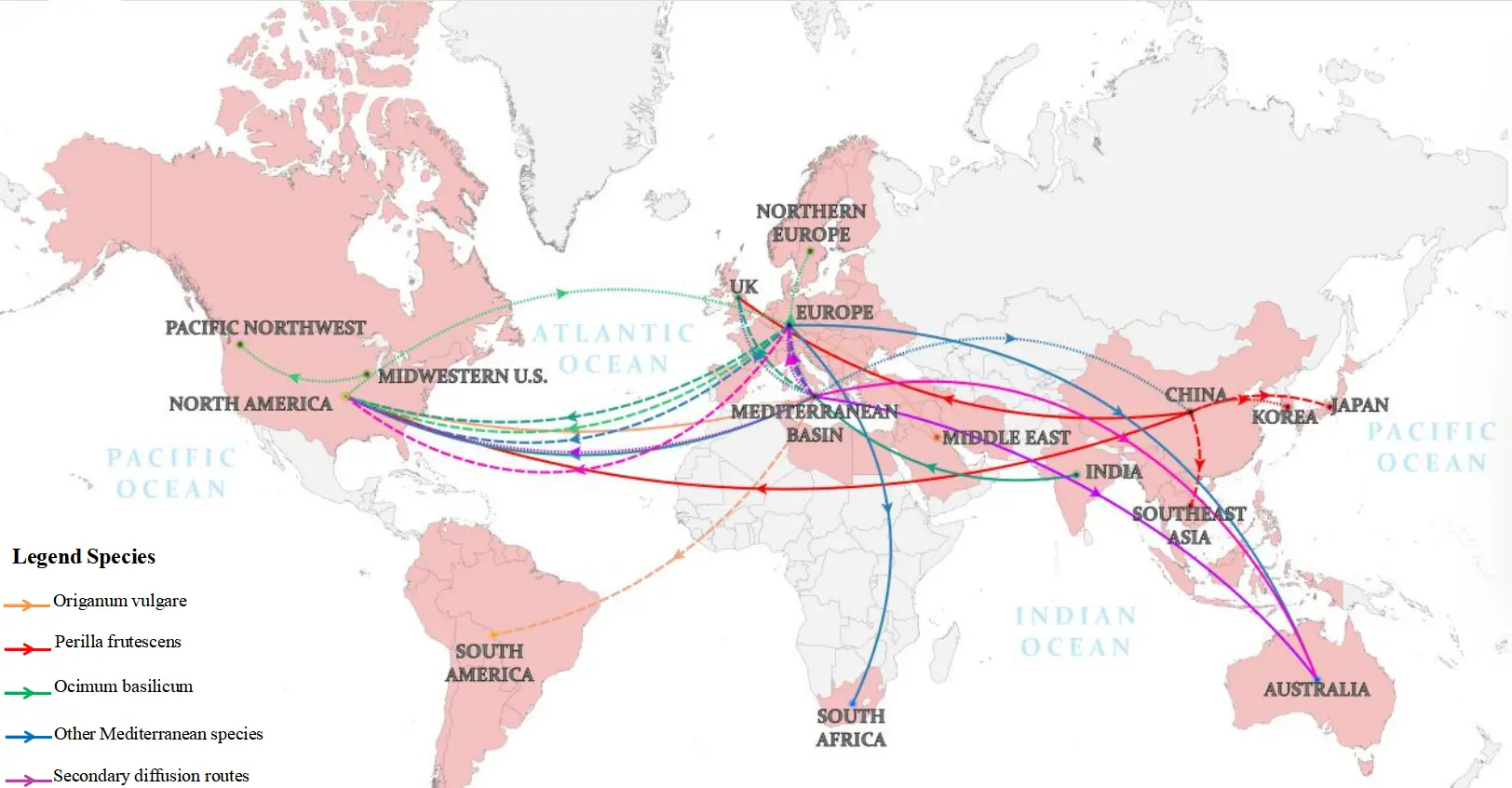
The global dissemination of key aromatic genera in Lamiaceae can be divided into three major periods. The first period, from 1000 BCE to 500 CE, was driven primarily by ancient land and maritime trade. The core routes originated in the Mediterranean region and Southwest Asia and extended eastward to East Asia via the ancient Silk Road. Roman legions and administrative officials facilitated the spread of mint, rosemary, and thyme from the Mediterranean region to northern Europe and Britain, whereas Persian and Arab merchants introduced basil from India to the Mediterranean region [14,18,38,39,40,41,42]. In the 2nd century CE, mint and basil were introduced into China through the Silk Road, leading to the establishment of early localized cultivation areas in Hainan, Yunnan, and other regions. During the same period, rosemary also entered China through Silk Road exchanges [40].
The second period, from the 15th to the 17th century, corresponded to the Age of Discovery and was driven mainly by European colonial expansion and cross-cultural exchange. European colonizers introduced mint, lavender, thyme, and rosemary to North America; Spanish colonizers brought oregano to Latin America; and Portuguese traders introduced basil to Indonesia [23,25,40,43]. Within the East Asian cultural sphere, perilla was introduced from China to Japan during the 8th–9th centuries through Buddhist monks and broader cultural exchanges [15]. It subsequently spread to Southeast Asian regions, such as Vietnam, through Chinese migration, while its introduction to the Korean Peninsula occurred earlier [16,17].
The third period, from the 17th century to the present, represents the era of globalized trade and commercialized breeding. Modern breeding technologies have substantially improved the climatic adaptability of key aromatic genera in Lamiaceae. For example, the “Mitcham” mint developed in the United States has been successfully cultivated in Canada and Australia, while the “large-leaf basil” bred in China has been widely promoted in Southeast Asia. The center of commercial mint cultivation has shifted from the U.S. Midwest to the Pacific Northwest, and the United States has become an important global exporter of mint oil [24]. Perilla was introduced to Europe and North America as an ornamental plant in the mid-19th century, whereas rosemary, lavender, and other herbs spread to South Africa and Oceania through colonial expansion [16,44,45].
In the modern context of crop dissemination and industrial development, climate adaptability has become a key factor shaping the geographical distribution of key aromatic genera in Lamiaceae. In Mediterranean-climate regions such as Italy, Greece, and Morocco, oregano and mint are among the dominant crops. In tropical and subtropical regions, including India, Hainan, China, and Brazil, basil holds a prominent position. In temperate regions such as the United States and Germany, mint is cultivated on a large scale. Meanwhile, the clustering effect of the essential oil processing industry has further accelerated the global expansion of these crops. Benefiting from its well-developed essential oil extraction industry, India has become one of the world’s largest producers and exporters of basil and mint, with annual essential oil exports exceeding 20,000 metric tons (FAO, 2023). Some species, such as perilla and thyme, have naturalized in introduced regions, including New Zealand and Australia, and, in certain cases, have become invasive [7,46,47]. These patterns indicate the transition of key aromatic genera in Lamiaceae from traditional culinary and medicinal plants to globally commercialized crops with significant economic and ecological relevance.
2.2. Germplasm Resource Collection and Preservation
The systematic collection of global key aromatic genera in Lamiaceae germplasm resources began in the 1960s, driven by the international initiative for plant genetic resource conservation and rising demand for aromatic crop diversity in the food and pharmaceutical industries [48] (FAO, 1967). Currently, a global conservation network has been established, led by international institutions and supported by national research organizations. This network integrates in situ conservation (wild population protection) and ex situ conservation (seed banks and living collections) to safeguard the long-term maintenance of genetic diversity (IPGRI, 2001). The International Plant Genetic Resources Institute (IPGRI) has established a core germplasm repository for Lamiaceae species in Rome, Italy, preserving 1300 accessions covering 15 species of Mentha, 8 species of Ocimum, and 5 species of Origanum. Wild resources account for 40% of the collection, including 12 wild accessions of the endangered species Salvia pratensis L. Such wild germplasm is particularly valuable for breeding stress-tolerant cultivars due to its abundant genetic variation [49] (IPGRI).
Major germplasm repositories in key producing countries exhibit distinct characteristics tailored to regional agricultural demands: India’s National Bureau of Plant Genetic Resources (NBGPR) preserves 850 accessions of Lamiaceae aromatic plant resources, with a focus on cultivars, indigenous basil varieties (e.g., “Tula” basil) of high medicinal value, and wild mint resources adapted to the tropical climate of the Indian subcontinent. The institute has established a preliminary database associating essential oil composition with genotypes, supporting marker-assisted selection for high-essential-oil-yielding lines [27]. The United States Department of Agriculture (USDA) germplasm repository in Maryland, USA, preserves 620 accessions, mainly from the genus Mentha genus (including Mentha piperita L.), such as the highly disease-resistant “USDA-N2” mint. This repository uses next-generation sequencing to characterize the genetic background of mint accessions, accelerating the discovery of genes associated with essential oil synthesis [49]. The National Lamiaceae aromatic plant Germplasm Resource Garden in Yunnan Province, established in 2005, functions as a key repository for both aromatic and medicinal Lamiaceae in China. It maintains 580 accessions representing 23 distinct species with diverse functional traits. Indigenous resources—including Mentha haplocalyx Briq. (wild mint) from Yunnan and Ocimum americanum L. (large-leaf basil) from Hainan—accounting for more than 60% of the collection, reflecting their strong adaptability to China’s diverse climatic conditions from the subtropical coastal regions to the arid northwest zones. To strengthen its ex-situ conservation, the garden has also established a cryopreservation system for Lamiaceae seeds, effectively extending the preservation duration of rare germplasm to over 30 years [50].
2.3. Research on Genetic Diversity of Germplasm Resources
Genetic diversity in the Lamiaceae family is manifested at morphological, molecular, and ecological levels [31]. Morphologically, remarkable intraspecific variation exists. Within the genus Mentha, peppermint (M. piperita L.) displays long-oval leaves and a high essential oil content (1.5–2.5%), while spearmint (M. spicata L.) has lanceolate leaves rich in carvone [31]. In the genus Ocimum, ‘Sweet Basil’ presents features green leaves with a mild aroma; ‘Purple Basil’ is characterized by anthocyanin accumulation and purple foliage [51]; and ‘Lemon Basil’ exudes a lemon-like fragrance due to limonene [52]. Studies on Indian holy basil (Ocimum tenuiflorum L.) further support the presence of substantial morphological and germplasm diversity [53]. Physiologically, different varieties exhibit obvious divergence in stress tolerance. Mediterranean oregano shows strong drought and heat resistance, Chinese mint varieties from Yunnan can tolerate low temperatures ranging from −5 °C to 0 °C, and Indian basil varieties adapt well to high-humidity environments (80–90% RH) [53]. Drought stress significantly affects growth and essential oil yield in basil, with differential responses detected among sweet basil (O. basilicum L.), African basil (O. gratissimum L.), and American basil (O. americanum L.) [54]. At the molecular level, studies using markers like SSR and other markers have clarified the genetic structure of Lamiaceae species. For Mentha, genetic differentiation is strongly with correlation between genetic differentiation and geographical distribution: the genetic distance between Mediterranean and East Asian populations is the largest (similarity coefficient 0.32), while genetic similarity between European populations exceeds 0.75 [55]. Molecular phylogenetic analysis of Ocimum suggests genetic divergence between cultivated basil and its wild relative Ocimum americanum L. occurred approximately 5000 years ago, consistent with the timeframe of human domestication timelines [56]. In addition, chloroplast genome studies have identified genetic diversity hotspots in cpDNA genes like matK and rbcL, and other cpDNA genes, which are valuable for species identification and phylogenetic analysis [28]. The rbcL sequence has been verified as an effective DNA barcoding for Mentha, enabling the discrimination of morphologically cryptic species. Combined use of matK and rbcL has successfully supported identification and classification across multiple Lamiaceae taxa [57].
In summary, studies on Lamiaceae molecular genetic diversity support three major conclusions: (1) Lamiaceae species generally maintain high genetic diversity, with wild germplasms and geographically isolated populations exhibiting significantly greater variation than cultivated varieties. (2) Such diversity patterns are mainly driven by geographical isolation (promoting allopatric speciation and genetic divergence) and human domestication (reducing genetic variation via artificial selection). (3) Chloroplast genes such as matK and rbcL serve as reliable markers for assessing genetic diversity and phylogenetic relationships, providing technical support for germplasm identification and resource classification. Meanwhile, single-copy nuclear genes with higher variability than cpDNA show good potential for species-level phylogenetic analysis [58].
The application of genetic diversity has become a core driver of modern breeding programs, with an obvious trend of multi-regional cooperation. In India, the cross between medicinal ‘Tulsi’ basil and high-yielding ‘Sweet Basil’ produced the variety ‘Basil No. 1’, with 2.1% essential oil content and strong mildew resistance, effectively balancing medicinal and economic values [53]. In the United States, researchers introduced disease-resistant genes from wild mint (Mentha longifolia) to improve the main cultivar ‘Mitchem’, increasing root rot resistance by 40% and providing key genetic support for stable production [49]. In China, the Institute of Botany, Chinese Academy of Sciences, successfully bred the new lavender variety ‘Luoshen’, which exhibits superior heat and humidity (81.9% survival rate) and high content of linalool and linalyl acetate, with considerable market potential [59]. Alongside these applications, progress in basic research—including genome sequencing of common lavender, population diversity analysis and molecular breeding of thyme, and innovation of new oregano germplasms—has deepened understanding of genetic mechanisms in Lamiaceae. Exploring wild resources remains a key strategy to broadening the genetic foundation for breeding, with antioxidant genes from Salvia africana L. and cold-tolerance genes from South American wild basil representing promising genetic resources for future breakthrough cultivar improvement [60,61].
3. Breeding History of Key Aromatic Genera in Lamiaceae
The development of global key aromatic genera in Lamiaceae breeding technology can be categorized into four distinct phases based on breeding methodologies: the pre-breeding stage, early introduction and preliminary domestication, conventional breeding, and modern molecular breeding [30]. The latter two stages have generated a wide range of important cultivars (Table 1 and Table 2), which are widely cultivated in major production regions across Southeast Asia, the Americas, and Africa. Historically, breeding efforts were dominated by traditional producing countries such as India, Italy, and the United States. Since the mid-20th century, emerging producers including China, Brazil, and Morocco have gradually established systematic breeding programs.
Table 1. Global Dissemination Matrix of Key Aromatic Genera in Lamiaceae.
|
Species |
Exact Origin |
Ancient Spread (500 BCE) |
Medieval & Early Modern Spread (500–1700 CE) |
Modern Spread and Commercialization (Post–1700 CE) |
|---|---|---|---|---|
|
Mint (Mentha spp.) |
Mediterranean Basin [5] |
Origin: Mediterranean → Destination: Northern Europe; Agent: Roman legions and administrators [38] |
Origin: Europe → Destination: North America; Agent: British and other European colonists [38] |
Origin: U.S. Midwest → Destination: U.S. Pacific Northwest (shift of commercial cultivation center); U.S. exports mint oil globally [38] |
|
Basil (Ocimum basilicum) |
India (Punjab and Himachal Pradesh foothills) [6] |
Origin: India → Destination: Mediterranean; Agent: Persian/Arab merchants via overland and maritime spice routes [41] |
Origin: Italy → Destination: France, England; Agent: Culinary and cultural exchange [42]; Origin: Europe → Destination: Americas; Agent: European colonists [21] |
Global culinary popularization, especially promoted in the U.S. by Italian immigrant communities [43] |
|
Perilla (Perilla frutescens) |
China/Indian mountains [7] |
Origin: China → Destination: Korea; Agent: Introduced before the Unified Silla period [17] |
Origin: China → Destination: Japan; Agent: Transmitted during the 8th–9th centuries through cultural/monastic exchanges [16]; Origin: China → Destination: Southeast Asia (e.g., Vietnam); Agent: Chinese migration and cultural influence [16] |
Introduced to the West (England, U.S.) as an ornamental plant in the mid-19th century; later became an invasive species in some regions [16] |
|
Rosemary (Salvia rosmarinus) |
Arid, rocky regions of the Mediterranean (including Portugal and northwestern Spain) [8] |
Origin: Mediterranean → Destination: Britain; Agent: Roman legions (43 CE) [40]; Origin: Mediterranean → Destination: China; Agent: Silk Road trade (c. 220 CE) [40] |
Origin: Europe → Destination: Americas; Agent: Early 17th-century European colonists [40] |
Spread to South Africa and Australia through European colonial expansion [44] |
|
Lavender (Lavandula spp.) |
Western Mediterranean mountains, Middle East, India [13] |
Origin: Mediterranean → Destination: Europe (including Britain); Agent: Romans, used for bathing and preservation [20] |
Origin: Europe → Destination: North America; Agent: English Puritan colonists (1620) |
Spread to Oceania (Australia, New Zealand) through European colonial expansion [45] |
|
Thyme (Thymus vulgaris) |
Southwestern Europe and southeastern Italy [10] |
Origin: Mediterranean → Destination: Europe (including Britain); Agent: Roman legions, used for purification and as a symbol of courage [9] |
Origin: Europe → Destination: North America; Agent: Early European colonists [23] |
Naturalized successfully in New Zealand, Australia, and other regions; becomes an invasive species in some areas [46] |
|
Oregano (Origanum vulgare) |
Mediterranean and Western Asian mountains [11] |
Origin: Mediterranean → Destination: Europe; Agent: Roman expansion [43]; Origin: Europe → Destination: Asia; Agent: Trade routes [43] |
Origin: Spain → Destination: Latin America; Agent: Spanish colonists [43] |
Brought to North America by Italian/Greek immigrants in the early 20th century; became widespread after World War II [43] |
Table 2. Table of diverse variety characteristics of key aromatic genera in Lamiaceae at different stages.
|
Breeding Method |
Variety Type |
Representative Variety |
Pedigree |
Breeding Institution |
Breeding Year |
Characteristics |
Molecular Marker Technology |
Application Scenarios |
Promotion Area |
|---|---|---|---|---|---|---|---|---|---|
|
Domestication Breeding |
Oregano, Basil, Mint, Sage |
Greek Oregano |
Natural domestication of wild oregano |
Greek local farmers/agricultural cooperatives |
Early 1900s |
Carvacrol content 60–70%, citronellal content 10–15%, drought and barren-tolerant, growth cycle 120 days |
/ |
Western food seasoning, essential oil extraction |
Greece, Italy, Mexico |
|
Italian Basil |
Artificial domestication of wild basil |
Lombardy Agricultural Research Center, Italy |
Mid-1900s |
Carvacrol content 60–70%, linalool content 8–10%, drought-tolerant, fresh yield up to 30 t/hm2 |
/ |
Fresh side dish, spice processing |
Italy, France, Spain |
||
|
Indian Tulsi Basil |
Traditional domestication of wild tulsi basil |
Karnataka Agricultural Research Institute, India |
Late 1900s |
Oleanolic acid (uric acid class) content 1.2%, rosmarinic acid content 0.8%, significant medicinal value, heat, and humidity-tolerant |
/ |
Traditional medicine, religious sacrificial spice |
India, Sri Lanka, Nepal |
||
|
Clonal Selection |
Mint, Basil |
Michigan Improved Mint |
Clonal screening of natural mutants of spearmint |
Horticultural Research Center, USDA |
1935 |
Essential oil content 2.3%, menthol content 78%, menthone content 5%, mint rust-resistant, fresh grass yield 60 t/hm2 |
/ |
Food additives, daily chemical flavor preparation |
USA, India, China |
|
Basil No. 3 |
Breeding Clonal propagation of superior individual sweet basil |
Indian Institute of Spices Research (IISR) |
1968 |
Powdery mildew-resistant (incidence < 5%), yield increased by 35% compared with ordinary sweet basil, essential oil content 1.8% |
/ |
Large-scale spice cultivation, deep processing |
India, Pakistan, Bangladesh |
||
|
Shanghai Mint No. 1 |
Directional clonal selection of Jiangsu purple-stem mint |
Horticultural Research Institute, Shanghai Academy of Agricultural Sciences |
1972 |
Strong cold resistance, can tolerate winter low temperature of −8 °C, root overwintering survival rate 90%, essential oil content 2.1% |
/ |
Open-field cultivation in northern China, mint tea processing |
East China, North China (China) |
||
|
Selection Breeding |
Oregano, Mint, Basil |
Italian Oregano No. 1 |
Continuous individual plant selection of wild oregano natural mutants |
Council for Agricultural Research and Economics (CREA), Italy |
1956 |
Root rot (fusarium oxysporum)-resistant, incidence reduced from 60% to 15%, carvacrol content 65% |
/ |
Mediterranean agricultural planting, spice export |
Italy, Greece, Turkey |
|
USDA-M5 Mint |
γ-ray (60Co) mutagenesis of mint cultivar ‘Spearmint’ |
Crop Genetics Research Institute, USDA |
1981 |
Essential oil content increased by 20% compared with the parent, citral content 12%, continuous cropping-tolerant, suitable for mechanized harvesting |
/ |
Industrial essential oil extraction, large-scale cultivation |
USA, Canada, Australia |
||
|
Basil No. 8 |
EMS chemical mutagenesis of basil cultivar ‘Genovese’ |
Indian Institute of Spices Research (IISR) |
1993 |
Early flowering (15 days earlier than the parent), growth period 65 days, double cropping per year possible, fresh grass yield 45 t/hm2 |
/ |
Protected agriculture rapid cultivation, fresh-cut vegetables |
India, Brazil, Yunnan (China) |
||
|
Molecular Marker-Assisted Breeding |
Mint, Oregano |
Marker-Assisted Mint No. 1 |
SSR marker (primer RM128)-assisted selection of mint germplasm |
Institute of Vegetables and Flowers, Chinese Academy of Agricultural Sciences (CAAS) |
2010 |
Menthol content increased to 82%, fusarium wilt (fusarium oxysporum)-resistant, saline-alkali tolerant (pH 7.5–8.5) |
SSR marker |
High-value essential oil production, saline-alkali land cultivation |
North China, Northwest China (China), Europe |
|
Basil, Oregano |
SSR-Basil No. 5 |
SSR marker (primer Bs11)-assisted pyramiding breeding of basil germplasm |
Spice Research Center, Brazilian Agricultural Research Corporation (EMBRAPA) |
2015 |
Essential oil yield increased by 40%, vanillin content 0.5%, heat-tolerant (normal growth above 35 °C) |
SSR marker |
Large-scale cultivation in tropical regions, flavor production |
Brazil, Southeast Asia, South America |
|
|
Oregano |
SNP-Oregano No. 2 |
SNP marker (locus OreSNP23)-assisted selection of oregano genome |
Agricultural Biotechnology Laboratory, University of Florence (Italy) |
2018 |
Carvacrol content up to 75%, salt-tolerant (NaCl concentration 0.6%), drought tolerance index increased to 0.85 |
SNP marker |
Cultivation in arid and semi-arid regions, medicinal extraction |
Italy, Greece, North Africa |
Note: Abbreviations of breeding institutions in this table are defined as follows: USDA: United States Department of Agriculture; IISR: Indian Institute of Spices Research; CREA: Council for Agricultural Research and Economics (Italy); CAAS: Chinese Academy of Sciences; EMBRAPA: Brazilian Agricultural Research Corporation.
3.1. Pre-Breeding Stage
The pre-breeding stage, before 1000 BCE, represents the earliest phase of human interaction with aromatic herbaceous plants. Human use during this period was mainly driven by basic survival needs, including food acquisition, disease relief, pain alleviation, and improvements in living environments [4]. At this stage, plant utilization was characterized primarily by passive adaptation and direct harvesting. Conscious artificial cultivation and targeted selection had not yet been established [62]. The distribution of these herbs was determined entirely by natural dispersal, and human intervention was limited to the collection and simple use of wild resources [62].
Among the representative Lamiaceae aromatic herbs, Mentha, Rosmarinus, Lavandula, Thymus, and Origanum originated mainly in the Mediterranean region, where they grew naturally in coastal mountains, coastal zones, and arid rocky habitats under the influence of distinctive regional climatic and edaphic conditions. Ocimum originated in the foothill regions of India and Southeast Asia [41], whereas Perilla was native to mountainous areas of China and India. During this period, these taxa remained confined to their respective native ecosystems, with no substantial interregional plant exchange. Human use of these seven aromatic herbs was therefore local, small-scale, and closely associated with immediate survival needs. Their early utilization can be summarized in three major aspects.
Medicinal use and disease prevention constituted the most fundamental applications of these herbs. In the absence of effective medical systems, early human communities relied on wild plants to relieve discomfort, treat illness, and prevent disease. The ancient Egyptian Ebers Papyrus, dating to approximately 1550 BCE, records the use of mint to alleviate indigestion, reflecting an early medicinal application derived from the collection of wild mint [15]. Thyme was used by the ancient Egyptians in mummification, taking advantage of its natural preservative and antibacterial properties to prolong the preservation of bodies [63]. Basil was also recorded in Indian Vedic texts approximately 5000 years ago [6]. In its early use, basil was applied mainly for simple medicinal purposes and had not yet acquired the religious significance it later acquired in later cultural contexts. During this period, medicinal use generally involved the direct collection of wild stems, leaves, or seeds, followed by simple processing methods such as crushing, soaking, or steeping to obtain active components. No artificial cultivation or systematic selection of these plants had yet been practiced.
Culinary seasoning represented another important early use. During hunting and gathering activities, early humans discovered that the stems, leaves, and seeds of certain aromatic herbs could mask undesirable odors in food and improve flavor. These plants were therefore gradually used as natural seasonings. Oregano, native to the Mediterranean mountains, was initially collected by the ancient Greeks from wild populations to season simple foods; its name, meaning “joy of the mountains” [43], reflects the sensory value associated with its aroma. Thyme stems and leaves were also used for flavoring, and wild thyme was often added to cooked game to enhance taste. At this stage, culinary use depended entirely on wild resources. The harvested amount was generally limited to immediate dietary needs, and practices such as storage, large-scale collection, or cultivation had not yet developed.
In addition to medicinal and culinary uses, aromatic herbs were also used to improve living environments. The distinctive scents of some herbs helped mask damp or musty odors in dwellings. For example, early humans collected lavender and placed it in living spaces for deodorization, while mint was used around dwellings because its scent could help repel mosquitoes and reduce insect disturbance. These practices did not require cultivation but relied solely on the direct collection of wild plants, representing an early and rudimentary exploitation of the functional traits of aromatic herbs.
Overall, the pre-breeding stage was characterized by the direct utilization of naturally distributed wild germplasm, without domestication, artificial cultivation, or directed selection [64]. With the subsequent development of human civilization, particularly the expansion of the Roman Empire, the rise of ancient spice trade networks, and later European colonial activities, these aromatic herbs gradually spread beyond their native ranges and entered a new phase of cross-regional dissemination and utilization.
3.2. Early Introduction and Initial Domestication and Cultivation Phase
During the early introduction, initial domestication, and cultivation phase, from 1000 to 500 BCE, ancient regional trade networks gradually developed. Trade along the Mediterranean coast and across the Asian continent became increasingly active, serving as a major driving force in the cross-regional dissemination of aromatic herbaceous plants. In contrast to the predominantly natural dispersal that characterized the pre-breeding stage, plant spread during this period was largely mediated by human transport. The main objective was to reproduce the uses of native herbs in new habitats to meet basic needs such as food seasoning and medicinal treatment [65], thereby breaking the earlier distribution pattern in which these seven herbs had remained largely confined to their native regions.
Building on the introduction of plant species across regions, the utilization of aromatic herbs shifted from simple collection to preliminary domestication efforts. The core mechanism of this process involved basic phenotypic selection: based on immediate needs, humans selected plants that were robust, had a stronger aroma, and exhibited more pronounced medicinal effects, preserving their seeds or rhizomes for subsequent cultivation, thereby gradually achieving the initial domestication of plant germplasm. For mint, this involved selecting plants with a more intense aroma, aligning with the medicinal requirement described in the “Shennong Bencao Jing” that mint “treats foul odors and abdominal distension”, driving humans to prioritize individuals with superior medicinal efficacy and stronger fragrance. The line “passing the mint to dance” in the “Chu Ci”, later verified by scholars to refer to mint, corroborates the widespread recognition and tendency toward its use during this stage [66]. For oregano and thyme, humans selected individuals with more vigorous growth and better flavor-enhancing properties. Theophastus’s records of oregano’s vermifuge and flavor-enhancing effects, along with Hippocrates’s discussions of over forty uses for oregano in his works, both indirectly reflect humanity’s tendency to screen for superior phenotypes. As for thyme, its status as a “symbol of courage” as recorded in “The Odyssey” drove humans to prioritize individuals with a more intense aroma and robust growth. Domestication at this stage remained in its early stages; the core traits of these herbaceous plants had not undergone fundamental changes, retaining many wild characteristics, and stable cultivated varieties had not yet been established. Human intervention was limited to selection and simple cultivation.
3.3. Conventional Breeding Phase
The conventional breeding stage, from 500 BCE to the late 20th century, was characterized by increasing commercial demand for aromatic herbs. During this period, vegetative propagation and empirical selection were widely adopted [67], enabling the stable preservation of desirable traits. This process ultimately promoted the formation of diverse locally adapted germplasm resources and represented a pivotal stage in the transition of aromatic herb domestication from early utilization to more mature cultivation systems.
3.3.1. Clonal Selection Breeding
Clonal selection breeding is one of the primary breeding methods for aromatic crops in the Lamiaceae family and has been most widely applied in the genus Mentha, particularly in peppermint and spearmint [68]. This method is generally based on local varieties developed during the traditional empirical selection stage and represents a core component of conventional breeding systems. It is characterized by technical maturity, operational simplicity, and strong practical applicability. Its main advantage lies in the stable preservation of varietal genetic characteristics, effectively avoiding trait segregation caused by sexual reproduction and ensuring the inheritance of superior traits across generations.
In 1954, the U.S. Department of Agriculture screened natural variation populations of “Mitcham” mint and identified an elite individual with an essential oil content of 2.3% and a menthol content of 78%. Through vegetative propagation methods such as rhizome propagation and cuttings, the clonal variety “Mitcham Improved” was developed. Owing to its stable quality and high productivity, this variety became one of the major cultivated types for mint essential oil production worldwide and was subsequently introduced into major production areas in several countries [25]. In 1975, to address the limitations of traditional spearmint, the UK Council for Agricultural and Food Research screened local clonal materials and identified an individual plant with an essential oil content of 1.8% and a carvone content exceeding 90%. Through cutting propagation, “British Spearmint No. 1” was developed. This variety is well adapted to the climate of the United Kingdom and has become a major cultivar for regional spearmint essential oil production. In 1986, the Indian Institute of Spices Research screened powdery mildew-resistant individuals from sweet basil clones and developed “Basil No. 3”. In areas with a high incidence of powdery mildew, this variety increased yield by 35% compared with the control and effectively reduced the adverse effects of the disease. In 1988, the Japanese Institute of Aromatic Plants screened imported “Mitcham Improved” mint clones and selected an individual with a mild aroma and a menthol content of 65–70%, leading to the development of “Japanese Mint No. 1”. This variety is well suited to local climatic conditions and is mainly used in high-end cosmetics and food flavoring. In 1990, Chinese researchers selected cold-tolerant individuals from traditional local mint varieties and developed “Shanghai Mint No. 1”. This variety can safely overwinter at −8 °C, is adapted to the climatic conditions of eastern China, and has become a major cultivar in the region [69].
Despite its practical value, clonal selection breeding has inherent limitations. These include limited genetic variation, the accumulation of pathogens during long-term vegetative propagation, potential clonal degeneration, difficulty in generating continuous innovation through genetic recombination [70], and relatively narrow ecological adaptability of selected varieties. innovation through genetic recombination, and the limited ecological adaptability of the varieties.
3.3.2. Selection Breeding
Selection breeding has been widely used [71] for varietal improvement in aromatic Lamiaceae plants and can be broadly divided into two categories: selection from natural variation and selection following induced mutation [72,73].
Selection from natural variation relies on the identification and utilization of superior individuals within existing populations. In 1962, the Italian Agricultural Research Institute screened wild oregano populations and selected individuals resistant to root rot, leading to the development of “Italian Oregano No. 1”. This variety reduced disease incidence in continuous cropping fields from 50% to 15%. In 1978, Morocco developed “Moroccan Mint” through selective breeding. This material has an essential oil menthol content of 45% and is suitable for perfume production.
Induced mutation breeding mainly involves the creation of novel variation through radiation or chemical mutagenesis, followed by the selection of desirable mutants. In 1985, mint seeds were irradiated with gamma rays in the United States, resulting in a mutant with a 20% increase in essential oil content, which was subsequently developed into “USDA-M5”. In 2000, India treated basil seeds with ethyl methanesulfonate (EMS) and developed the early-flowering variety “Basil No. 8”. This variety shortened the growth period from 90 days to 65 days, making it suitable for multi-season cultivation.
However, selection breeding also has clear limitations. Its efficiency is restricted by the low frequency of favorable mutations and the relatively long breeding cycle. For example, the selection breeding cycle for oregano generally requires 5–8 years. In addition, this approach is often constrained by the genetic background of the original material, making it difficult to achieve breakthroughs in traits.
3.3.3. Hybrid Breeding
Hybrid breeding is one of the core approaches for improving key aromatic genera in Lamiaceae [24]. By recombining parental traits, this method enables the coordinated improvement of yield, quality, and stress tolerance [56,74]. Hybrid breeding in the genus Mentha is among the most representative examples. In 1790, British breeders crossed peppermint (M. piperita) with spearmint (M. spicata) to develop “lemon mint”, which combines the cooling sensation of mint with a lemon-like aroma. In 1992, researchers in the United States crossed “Mitcham” mint with wild mint (M. longifolia) and developed “USDA-M8”, which showed a 50% improvement in rust resistance and achieved an essential oil yield of 120 kg·ha−1 [71].
In basil, hybrid breeding has focused mainly on combining superior quality with stress tolerance. In 2005, India crossed “Sweet Basil”, a high-yielding type, with “Tulsi” basil, which has high medicinal value, and developed “Basil No. 10”. The essential oil of this variety contains 1.2% eugenol, and its yield is 25% higher than that of “Sweet Basil”. In 2010, the Chinese Academy of Tropical Agricultural Sciences developed “Qiong Basil No. 1” by crossing “large-leaf basil” with “lemon basil”. Its essential oil contains 30% limonene, and the variety is suitable for cultivation in tropical regions [74].
Interspecific hybridization is an important strategy for broadening the genetic base of aromatic Lamiaceae crops. In 1998, Italy developed the aphid-resistant variety “Italian Basil No. 6” by crossing basil (O. basilicum) with wild basil (O. americanum). Nevertheless, interspecific hybridization remains constrained by reproductive isolation. For example, the hybridization success rate between Mentha and Ocimum is less than 5%, and embryo rescue techniques are often required to improve hybrid recovery and hybridization efficiency [75].
3.4. Modern Molecular Breeding Stage
3.4.1. Genome Assembly and Analysis of Cultivars
To date, chromosome-level genome assemblies have been accomplished for multiple key aromatic genera in Lamiaceae worldwide. In 2017, the Kunming Institute of Botany, Chinese Academy of Sciences, completed the genome assembly of peppermint (Mentha piperita L. ‘Mitcham’), with a size of 620 Mb and a contig N50 of 25.3 Mb. The genome was successfully anchored to 12 pseudo-chromosomes and annotated with 58,234 protein-coding genes. Among these annotated genes, 128 were associated with terpenoid biosynthesis (e.g., the TPS gene family), providing a solid molecular foundation for investigating the mechanisms of essential oil synthesis [25]. In 2020, Cornell University (USA) completed the genome assembly of covering Ocimum basilicum L. ‘Genovese’, covering 385 Mb with a contig N50 of 18.7 Mb. The study identified 45 members of the MYB gene family involved in anthocyanin synthesis, which explains the color difference between “purple basil” and “green basil” [51].
In 2022, a research team from the University of Florence (Italy) completed the draft genome assembly of oregano (Origanum vulgare L.) with a genome size of approximately 410 Mb. A key gene regulating carvacrol biosynthesis, CYP71D1, was identified in the genome. This finding is consistent with the conclusion regarding the terpenoid metabolic diversity in four Lamiaceae aromatic plants revealed by Bornowski et al. [76], further clarifying the molecular synthesis mechanism of the core active components in oregano essential oil. In 2023, a research team from the University of Thessaly Greece, conducted a high-throughput pre-breeding evaluation of Greek oregano (Origanum vulgare L. subsp. hirta). By constructing a genotype fingerprint using SSR molecular markers and integrating essential oil component analysis, they identified three specialized genotypes, establishing precise linkage between molecular markers and phenotypic traits [77].
In recent years, genomic research on aromatic Lamiaceae plants has achieved a series of breakthroughs, providing critical insights into their evolutionary history and the mechanisms underlying the biosynthesis of active compounds. In 2021, a team from the Institute of Botany, Chinese Academy of Sciences, completed the chromosome-level genome assembly of Lavandula angustifolia Mill. Through comparative genomic analysis, they revealed two whole-genome duplication events in Lamiaceae plants and identified key gene clusters regulating monoterpene essential oil biosynthesis, laying a molecular basis for the directional improvement of lavender essential oil quality [78]. In 2023, the research team led by Professor Chengwei Yang at South China Normal University constructed the first chromosome-level reference genome of Salvia rosmarinus L. spreng, elucidating the evolutionary mechanism of the rosmarinic acid biosynthetic pathway. They found that the tandem duplication and functional divergence of key enzyme genes are the core drivers of high antioxidant accumulation in rosemary [79]. In 2024, the team led by Professor Dangquan Zhang at Henan Agricultural University further completed a high-quality genome assembly of Salvia rosmarinus L. spreng using the multi-resistant new line ‘Albus-2’. This work not only improved the continuity and completeness of the genome but also deeply dissected the molecular regulatory network linking antioxidant biosynthesis and environmental adaptation, providing key targets for its stress-resistant breeding and active ingredient metabolic engineering [80].
3.4.2. Identification and Utilization of Genes Associated with Key Breeding Traits
Disease Resistance Genes
Major diseases affecting aromatic Lamiaceae plants include downy mildew, root rot, and rust, making the mining of disease resistance genes a core focus of molecular breeding efforts. Sweet basil (Ocimum basilicum L.) is highly susceptible to downy mildew caused by Peronospora belbahrii Thines, and to date, no commercial resistant varieties have been developed. In 2018, studies confirmed that the highly resistant wild basil (Ocimum americanum L. var. americanum PI 500945) harbors the dominant downy mildew resistance gene Pb1—specifically a pair of dominant genes Pb1A and Pb1A’. By combining interspecific hybridization with embryo rescue technology, this resistance trait was successfully transferred to susceptible sweet basil lines, and the disease resistance of the F1 and backcross progenies was stably inherited [81]. In 2020, an American research team identified a quantitative trait locus (QTL) associated with rust resistance on chromosome 5 of mint through a genome-wide association study (GWAS). The Tm-1 gene located within this locus can inhibit the proliferation of Puccinia menthae, (the causal agent of mint rust). The introduction of Tm-1 into ‘Mitcham’ mint significantly enhanced its rust resistance by 45% [82].
Quality-Related Genes
Their essential oil composition primarily determines the commercial quality of Lamiaceae aromatic plants. Research into the mining and functional verification of quality regulatory genes has mostly focused on the terpenoid and phenolic biosynthetic pathways [83], leading to the characterization of several core functional genes and regulatory mechanisms. In Mentha, MmPMR (menthol reductase gene) is a key determinant of menthol biosynthesis. Specifically expressed in leaves, it and directly catalyzes the rate-limiting step of menthol formation. Functional validation experiments have demonstrated that the overexpression of MmPMR significantly increases menthol accumulation by 30%, thereby providing a pivotal molecular target for the directional breeding of Mentha varieties with superior essential oil quality [84]. In Ocimum tenuiflorum L., ObUGT (UDP-glycosyltransferase gene) acts as a core regulatory gene in the ursolic acid biosynthetic pathway, with its overexpression leading to a 25% increase in ursolic acid content. Concurrently, the terpene synthase (TPS) gene family in Ocimum tenuiflorum L. exhibits significant differences in expression abundance and patterns, which directly drives the chemical diversity of essential oil and serves as a critical molecular basis for quality formation [33]. Studies on Rosmarinus officinalis L. have revealed that the interaction between arbuscular mycorrhizal fungi and the plants can significantly up-regulate the transcription level of terpene synthase genes via signal transduction pathways. This process accelerates the conversion of terpenoid precursors into essential oil components and ultimately enhances essential oil yield, and thereby providing a novel regulatory strategy for the efficient and green production of Rosmarinus officinalis L. essential oil [85].
Stress-Resistant Genes
In recent years, research on the stress resistance of key aromatic genera in Lamiaceae has focused on the molecular response mechanisms to drought, low-temperature, and salt stress, with remarkable progress achieved. Regarding drought tolerance, Bai et al. (2023) identified the McWRKY57-like gene (a group IIc WRKY transcription factor) in Mentha canadensis L. Under drought stress, the transcriptional level of this gene in leaves was up-regulated by 33 folds; its heterologous expression in Arabidopsis thaliana enhanced leaf chlorophyll content, soluble sugar, soluble protein, and proline levels, while increasing the activities of antioxidant enzymes including catalase, superoxide dismutase, and peroxidase, thereby alleviating yield loss under drought conditions [86]. For cold tolerance, Rezaie et al. (2020) analyzed the effects of low-temperature stress on Ocimum basilicum L. They found that cold stress altered the antioxidant defense system and phenylpropanoid metabolism, and up-regulated the expression of genes involved in phenylpropanoid biosynthesis, thereby enhancing the cold tolerance of plants [87]. In addition, Wang et al. (2024) performed comparative transcriptome analysis of Ocimum basilicum L. under salt stress, identifying ObWRKY16 and ObPAL2 as key regulators of salt stress response. Overexpression of these two genes in transgenic Arabidopsis thaliana significantly improved salt tolerance, thereby expanding our understanding of the genetic regulatory network of salt stress in Lamiaceae plants [88].
3.4.3. Advances in Genetic Transformation and Gene Editing Technologies
Agrobacterium-mediated transformation is the core technique for genetic transformation of key aromatic genera in Lamiaceae, with mature systems established for several species. Yu et al. (2022) established a stable Agrobacterium-mediated genetic transformation protocol for Mentha piperita L. and Mentha canadensis L. using internodal explants, achieving 100% regeneration rates and producing 3 to 4 transgenic lines per 500–600 explants [29]. Deschamps et al. (2002) established the first Agrobacterium-mediated transformation system for Ocimum basilicum L. and O. citriodorum L., achieving a 96% infection rate using leaf explants and successfully obtaining stable transgenic plants [89]. The gene editing technology for key aromatic genera in Lamiaceae is still in the initial stage, with most studies focusing on the regulation of essential oil components and the improvement of agronomic traits. Yun et al. (2023) utilized CRISPR/Cas9 to knockout the PfHd3a gene in perilla (Perilla frutescens (L.) Britt.), resulting in significantly delayed flowering and a ~50% increase in leaf biomass [90]. Currently, some Lamiaceae aromatic plant species suffer from low regeneration efficiency, and the proportion of stably edited plants is less than 5%, indicating that the relevant technical systems require further optimization.
4. Advances in Seedling Propagation Techniques
The propagation techniques of key aromatic genera in Lamiaceae are selected based on their biological characteristics and industrial production requirements, with seed propagation, cutting propagation, grafting propagation, and tissue culture propagation being the four core technical methods currently in use. Each technique has unique advantages and scopes of application.
4.1. Seed Propagation
Seed propagation is the core method for commercial seedling production of annual and biennial key aromatic genera in Lamiaceae, and serves as the fundamental basis for their germplasm innovation and hybrid breeding. In relevant studies, technical measures, including seed priming, substrate optimization, and environmental regulation, have effectively addressed the industrial problems of low natural germination rates and uneven seedling emergence in some species, providing solid technical support for large-scale seedling cultivation. Customized technical schemes have been developed for different species: When sweet basil seeds were primed with salicylic acid and potassium nitrate, their germination rate and seedling growth potential were significantly improved under salinity stress [91]. Under conditions of 25 ± 2 °C and 70% relative humidity, oregano seeds soaked in 200 mg/L GA3 for 8 h under showed an increase in germination rate from 65% to 89.5%, which significantly reduced seedling loss caused by fungal contamination [92]. Rosemary seeds subjected to 4 °C stratification for 30 d, combined with 100 mg/L GA3 soaking, had their germination rate increased from less than 20% to 68% [92]. Lavender seeds treated with 50 mg/L GA3 for 12 h had exhibited 35% better germination uniformity and a 7–10 day shorter seedling cycle [93]. Seed propagation is characterized by a high propagation coefficient, low cost, and simple operation, which can meet the demand for commercial large-scale production and provide genetic variation for elite variety breeding. However, it has certain limitations: perennial species with heterozygous genomes exhibit certain severe trait segregation in their progeny. Species such as lavender and rosemary have small seeds with long dormancy periods, and their field germination rate is less than 30% without proper treatment. Additionally, their germination is sensitive to environmental conditions, leading to fluctuations in seedling quality across batches and increased management difficulty [94].
4.2. Cutting Propagation
Cutting propagation is the mainstream asexual propagation technology for perennial key aromatic genera in Lamiaceae, including mint, lavender, and rosemary. It serves as a means to stably preserve the excellent essential oil quality and agronomic traits of mother plants, while avoiding trait segregation in seed propagation, thereby ensuring the commercial quality uniformity of spices. Relevant research on cutting selection, exogenous regulator application, substrate optimization, and precise environmental control has effectively addressed the problems of low rooting rate and poor root development in some species, improving the efficiency and stability of seedling cultivation. Customized propagation protocols have been formulated for different species. For mint upper semi-lignified stem segments, treated with 100 mg/L NAA treatment, exhibit a 98.5% rooting rate when planted in peat-perlite-vermiculite (2:1:1) substrate [95]. For Lavandula angustifolia Mill. annual stems with 30–50% lignification, when soaked in a mixture of 200 mg/L IBA + 50 mg/L NAA under suitable conditions, show a significant increase in rooting rate, increasing from 55% to 92.1% [96]. For rosemary cuttings pretreated at 4 °C and treated with ABT1 rooting powder, treated with rooting elevated rise from 38% to 79.3% [97]. This technology features simple operation and a short seedling cycle, making it suitable for large-scale production; however, it has obvious limitations. The supply of high-quality cuttings is limited by the quantity of the mother plant, resulting in a much lower propagation coefficient than with seed or tissue culture propagation. Cuttings are prone to carrying pathogens that can cause basal rot; their rooting is sensitive to environmental conditions, leading to batch-to-batch variation. Long-term continuous cutting propagation leads to germplasm degradation of mother plants, requiring regular germplasm renewal [67].
4.3. Grafting Propagation
Grafting propagation is a specialized asexual propagation technique for key aromatic genera in Lamiaceae. It involves fusing scions of elite varieties with stress-tolerant rootstocks to form symbiotic plants through wound healing. This approach not only fully preserves the superior traits of scions, such as essential oil quality and agronomic performance, but also enhances the plants’ resistance to diseases, cold, and salt-alkali stress through the well-developed root systems of rootstocks, while effectively mitigating the obstacles posed by continuous cropping. Currently, the application of this technique is far less widespread than that of cutting propagation, and research and practical application of grafting for oregano and mint remain relatively limited. Mature protocols have only been established for specific species; for instance, approach-grafting sweet basil scions onto Ocimum gratissimum L. rootstocks reduces the incidence of fusarium wilt by 45% in continuously cropped soil [98]. Grafting facilitates the complementary advantages of scions and rootstocks, thereby stabilizing spice quality and improving plant growth performance [99]. However, this technique imposes high requirements on operational expertise and labor costs. Additionally, there are significant variations in graft compatibility among different species, and tailored grafting schemes for oregano, mint, and related species have not yet been developed. Furthermore, its propagation coefficient is considerably lower than that of cutting and seed propagation, and the prolonged seedling cycle further increases production inputs [99].
4.4. Tissue Culture Propagation
Tissue culture is a core technology for efficient asexual propagation and germplasm preservation of key aromatic genera in Lamiaceae. By inoculating explants onto artificial media under aseptic conditions, complete plants are regenerated through dedifferentiation and redifferentiation, enabling the rapid acquisition of large quantities of high-quality seedlings with consistent genetic traits [100]. This technology overcomes limitations of traditional asexual propagation, such as low propagation coefficients and seasonal restrictions, and can produce virus-free seedlings to address germplasm degradation in species like mint and lavender caused by long-term asexual propagation, thereby providing a technical platform for germplasm innovation and genetic engineering improvement [101]. Adapted technical systems have been established for different species: Mint shoot tips achieve efficient proliferation (proliferation coefficient > 15) on MS medium supplemented with 6-BA and NAA, and their virus-free seedlings exhibit a 20% higher essential oil yield [101]; Sweet basil cotyledons form regenerated plants via callus induction, with a 99% rooting rate and >95% transplant survival rate [102]; Rosemary stem segments resolve vitrification issues by optimizing the hormone ratios of the medium, shortening the proliferation cycle to 25 days [103]; Lavender realizes somatic embryogenesis through embryogenic callus culture, which improves propagation efficiency and enables long-term germplasm preservation [104]. Tissue culture is characterized by high propagation coefficients and short seedling cycles, making it suitable for annual factory-scale seedling production; it also preserves maternal plant traits and enhances plant stress resistance [100]. However, this technology imposes strict requirements on laboratory conditions and operational skills. The costs for media and aseptic environments are relatively high, rendering it unsuitable for small-scale growers. Immature regeneration systems persist in some species (e.g., oregano), with unresolved issues including explant browning and transplant difficulties. Tissue culture seedlings have weak root systems, requiring refined management; long-term culture is prone to somatic variation, which necessitates regular trait screening [105].
5. Outlook
At present, traditional selection breeding and crossbreeding remain the core approaches for breeding key aromatic genera in Lamiaceae. Although improved lines with increased essential oil content and optimized agronomic traits have been developed for major varieties such as Mentha haplocalyx Briq., Lavandula angustifolia L., Ocimum basilicum L., and Rosmarinus officinalis L., the overall breeding efficiency is relatively low with a long cycle, and the breeding progress of varieties with disease resistance, high quality, and high yield is sluggish. In recent years, modern biotechnologies, including molecular markers, multi-omics analysis, and tissue culture, have been gradually integrated into breeding research, achieving phased breakthroughs in the regulatory mechanisms of key traits, precise identification of germplasm resources, and high-efficiency seedling raising. However, the high-efficiency genetic transformation system for most species remains imperfect, and biotechnology-based breeding is still in transition from laboratory exploration to industrial application, with no large-scale, commercially available biotech-bred varieties available so far. To break through the bottlenecks in industrial development and provide sustainable variety support for the Lamiaceae aromatic plant industry, future breeding efforts should focus on the following core directions.
5.1. Breeding Objectives: Focus on Disease Resistance, Quality Improvement, High Yield, and Medicinal Breeding
Disease resistance is the primary core objective of Lamiaceae aromatic plant breeding. In actual production, the frequent occurrences of fusarium root rot, powdery mildew, downy mildew, continuous cropping obstacles, as well as abiotic stresses such as drought and salinization, severely restrict the stability of yield and quality. Therefore, it is urgent to directionally cultivate varieties with broad-spectrum disease resistance and high stress tolerance (Figure 3), reduce agricultural chemical input, and consolidate the foundation for stable industrial production [106]. High quality is the core commercial attribute of varieties. It is necessary to systematically analyze the genetic mechanisms underlying the synthesis of characteristic flavor substances (e.g., linalool, menthol, and thymol), directionally improve the components and content of essential oils, and cultivate high-quality lines that meet the demands of the food and daily chemical industries. Meanwhile, it is imperative to strengthen the medicinal breeding direction based on food and medicinal homology, and targetedly increase the content of bioactive components with both edible and medicinal values in varieties so as to expand the dimensions of their medicinal development and application [107]. High yield is the core demand for the large-scale development of the key aromatic genera in Lamiaceae industry. It is required to optimize plant type, improve photosynthetic efficiency and seed setting rate through agronomic trait improvement, and combine the precise pyramiding of disease resistance and high quality traits to breed new high-yield varieties with excellent comprehensive agronomic traits, thereby realizing the coordinated improvement of disease resistance, high quality, and high yield [108].
5.2. Intensifying the Collection, Identification, and Innovative Utilization of Germplasm Resources
Germplasm resources serve as the material foundation for key aromatic genera in Lamiaceae breeding. Lamiaceae plants exhibit abundant germplasm diversity, and their wild relatives harbor excellent genetic resources related to disease resistance, stress tolerance, high essential oil content, and high bioactive component content [109]. For example, the introduction, cultivation, and comprehensive evaluation of basil germplasm resources have confirmed the diversity of these traits in wild materials [109]. However, most currently cultivated varieties have a narrow genetic background and a single base of breeding materials. It is thereby necessary to conduct systematic field collection, ex-situ conservation, and precise identification of key aromatic genera in Lamiaceae germplasm resources, construct a standardized and information-based germplasm resource bank, and clarify the agronomic traits, resistance performance, and genetic background of the resources. Such systematic collection and conservation work has been effectively applied in the process of lavender resource introduction and new variety breeding [110]. Meanwhile, DNA barcoding technology can act as a core method for precise identification this technology has been used to identify 13 types of medicinal and edible spice plants in the Lamiaceae family [111], and has also realized species differentiation of important Lamiaceae medicinal plants in India [112]. At the same time, it is essential to break the barriers to germplasm resources circulation, strengthen cross-regional and international germplasm exchange and cooperation, broaden the genetic background through intraspecific crossing and Interspecific distant hybridization, and explore and pyramid excellent alleles associated with disease resistance, high quality, high yield, and high bioactive component content. For instance, wild relatives have become core germplasm resources for mint variety improvement [68], providing a reference for other key aromatic genera in Lamiaceae. In this way, abundant material support can be provided for breeding innovation and solving the current problems of insufficient genetic diversity and difficult pyramiding of excellent traits in current breeding.
5.3. Accelerating the Construction of an Efficient and High-Throughput Seedling Breeding System
Seed propagation and cutting propagation are the main propagation methods for key aromatic genera in Lamiaceae in production, both of which have obvious shortcomings: seed propagation is prone to genetic segregation, and perennial varieties have low germination rate and long dormancy period; cutting propagation has a low propagation coefficient and limited supply of high-quality cuttings, and long-term asexual propagation is likely to cause germplasm degradation, which affects the stable expression of disease resistance, high quality and high yield traits of varieties [113]. As an efficient asexual propagation technology, tissue culture can rapidly produce large numbers of high-quality seedlings with consistent genetic traits and cultivate virus-free seedlings, thereby solving the problem of germplasm degradation at the source, which is an important development direction for large-scale seedling production of key aromatic genera in Lamiaceae. At present, preliminary tissue culture regeneration systems have been established for some varieties, but there are still technical bottlenecks, such as explant browning, vitrification, low rooting rates, and low survival rates during transplanting and acclimatization [114]. It is urgent to optimize the medium formula, hormone ratio and environmental control conditions according to the characteristics of different species, establish an efficient, stable and standardized tissue culture regeneration system, and construct a high-throughput seedling breeding system combined with soilless culture and factory seedling raising technology to realize the annual supply of high-quality seedlings, thus providing solid technical support for the rapid popularization and industrial application of new disease-resistant, high-quality and high-yield varieties [115].
5.4. Integrating and Innovating Modern Biotechnology to Break Through the Bottlenecks of Traditional Breeding
Most key aromatic genera in Lamiaceae are perennial plants, and some varieties have high genomic heterozygosity and complex ploidy. Traditional breeding has such bottlenecks as a long cycle, low efficiency of trait selection, and incompatibility of distant hybridization, making it difficult to rapidly realize the precise pyramiding of disease resistance, high quality, and high yield traits. Modern biotechnology provides precise and efficient tools to break through these problems. Interspecific distant hybridization combined with embryo rescue technology can overcome reproductive isolation, introduce the excellent traits of disease resistance, high yield, and high bioactive component content from wild relatives into cultivated varieties, and create breakthrough germplasm materials [75]. Molecular marker-assisted selection technology can realize the early and precise identification of target traits such as disease resistance, high quality, and high bioactive component content, eliminate inferior individual plants, greatly shorten the breeding cycle, and improve breeding efficiency [116]. Whole-genome sequencing and pan-genome analysis can elucidate the molecular regulatory networks of key traits related to disease resistance, essential oil synthesis, and yield, explore functional genes and molecular markers, and provide a theoretical basis for precise breeding [117]. Meanwhile, it is necessary to accelerate the construction of an efficient genetic transformation system, and combine with gene editing technology to realize the precise regulation of key genes related to essential oil synthesis, disease and stress resistance, yield, and bioactive component synthesis, so as to breed breakthrough new varieties with disease resistance, high quality, and high yield. Haploid breeding technology can rapidly obtain homozygous lines and solve the problem of the long breeding cycle of heterozygous varieties, while genome-wide association study can locate gene loci of key traits and provide precise targets for Molecular marker-assisted selection [118,119].
5.5. Cross-Disciplinary Collaboration and Technology Transformation for Sustainable Industrial Development
Key aromatic genera in Lamiaceae are characteristic economic crops with edible, medicinal, and industrial values. The cultivation and popularization of new disease-resistant, high-quality, and high-yield varieties is not a single-discipline work, and the industrial chain covers breeding, planting, processing, application, and other links. It is necessary to strengthen cross-disciplinary cooperation among breeding science, botany, molecular biology, processing engineering, agricultural mechanization, and other disciplines to realize the whole-chain connection from variety cultivation to industrial application. At the same time, it is essential to establish an in-depth cooperation mechanism between breeding institutions and planting bases, processing enterprises, pharmaceutical and food enterprises, promote the integrated development of breeding, propagation, popularization and application, and accelerate the demonstration, popularization and technology transformation of excellent new varieties with disease resistance, high quality, high yield and food-medicinal homology, so as to enable the rapid industrial application of breeding achievements [120].
In addition, it is required to strengthen international exchange and joint research on breeding technology, share research achievements and technical experience, jointly address global industrial challenges such as diseases, pests and climate change, popularize innovative technologies such as molecular breeding, green seedling raising and high-efficiency cultivation, improve the breeding level and production efficiency of the global key aromatic genera in Lamiaceae industry, and promote variety upgrading and the high-quality and sustainable development of the industry [121].
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the authors used GPT in order to correct grammar errors. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the published article.
Acknowledgments
The authors thank Chaoyun Hao (Spice and Beverage Research Institute, Chinese Academy of Tropical Agricultural Sciences) for the academic guidance and support throughout the preparation of this review.
Author Contributions
S.C. was responsible for conceptualization, literature collection, and original manuscript writing. Y.F. and R.F. contributed equally to literature screening, data integration, and manuscript revision. Y.H. and C.H. supervised the study, provided academic guidance, and revised the manuscript as corresponding authors. X.L. provided technical support and material supplementation for the review.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Funding
This research received no external funding.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Li B, Cantino PD, Olmstead RG, Bramley GLC, Xiang C-L, Ma Z-H, et al. A large-scale chloroplast phylogeny of the Lamiaceae sheds new light on its subfamilial classification. Sci. Rep. 2016, 6, 34343. DOI:10.1038/srep34343 [Google Scholar]
-
Krause ST, Liao P, Crocoll C, Boachon B, Förster C, Leidecker F, et al. The biosynthesis of thymol, carvacrol, and thymohydroquinone in Lamiaceae proceeds via cytochrome P450s and a short-chain dehydrogenase. Proc. Natl. Acad. Sci. USA 2021, 118, e2110092118. DOI:10.1073/pnas.2110092118 [Google Scholar]
-
Luo W, Du Z, Zheng Y, Liang X, Huang G, Zhang Q, et al. Phytochemical composition and bioactivities of essential oils from six Lamiaceae species. Ind. Crops Prod. 2019, 133, 357–364. DOI:10.1016/j.indcrop.2019.03.025 [Google Scholar]
-
Gülçin İ, Karageçili H. The Lamiaceae Family Plants Ethnobotanical Properties, Ethnopharmacological Uses, Phytochemical Studies and Their Utilization in Public or Current Clinical Practices: A Review. Rec. Nat. Prod. 2025, 466–487. DOI:10.25135/rnp.529.2505.3521 [Google Scholar]
-
Mint. Available online: https://academics.hamilton.edu/foodforthought/Our_Research_files/mint.pdf?q=the-mint-plant-and-its-uses#:~:text=Mint%20originated%20in%20the%20Mediterranean,to%20Britain%20and%20eventually%20America (accessed on 20 October 2025).
-
The History & Magic of Basil. Available online: https://www.plantspecialists.com/blog/history-of-basil (accessed on 20 October 2025).
-
Perilla frutescens—Perilla-Mint. Available online: https://gobotany.nativeplanttrust.org/species/perilla/frutescens/ (accessed on 20 October 2025).
-
Rosemary. Available online: https://www.mccormickscienceinstitute.com/resources/culinary-spices/herbs-spices/rosemary#:~:text=It%20is%20native%20to%20the,prefers%20well%20drained%2C%20sandy%20soil (accessed on 20 October 2025).
-
A History of Lavender. Available online: https://www.highcountrygardens.com/content/gardening/lavender-history (accessed on 20 October 2025).
-
Thymus vulgaris. Available online: https://plants.ces.ncsu.edu/plants/thymus-vulgaris/ (accessed on 20 October 2025).
-
Oregano. Available online: https://www.britannica.com/plant/oregano (accessed on 20 October 2025).
-
Herbs in History: Mint. Available online: https://www.ahpa.org/herbs_in_history_mint (accessed on 20 October 2025).
-
Lavender History. Available online: https://hitchinlavender.com/lavender-history/#:~:text=It%20is%20widely%20believed%20that,scent%20could%20still%20be%20detected (accessed on 20 October 2025).
-
Thyme. Available online: https://en.wikipedia.org/wiki/Thyme (accessed on 20 October 2025).
-
Civil War Medicinal Herb Garden—How Mint Was Used as a Remedy. Available online: https://www.civilwarmed.org/mint/ (accessed on 20 October 2025).
-
Shiso. Available online: https://en.wikipedia.org/wiki/Shiso (accessed on 20 October 2025).
-
Perilla frutescens. Available online: https://en.wikipedia.org/wiki/Perilla_frutescens (accessed on 20 October 2025).
-
Romans: Food and Health. Available online: https://www.english-heritage.org.uk/learn/story-of-england/romans/food-and-health/ (accessed on 20 October 2025).
-
Rosemary, Rosemarinus officinalis. Available online: https://hort.extension.wisc.edu/articles/rosemary-rosemarinus-officinalis/ (accessed on 20 October 2025).
-
Wash Up with Lavender. Available online: https://sdzwildlifeexplorers.org/animals/lavender#:~:text=In%20Roman%20times%2C%20people%20put,spreading%20the%20plant%20throughout%20Europe (accessed on 20 October 2025).
-
Sweet Basil. Available online: https://cornellbotanicgardens.org/explore/exhibits/seeds-of-survival-and-celebration/featured-plants/sweet-basil (accessed on 20 October 2025).
-
Early Settler Gardens. Available online: https://teara.govt.nz/en/gardens/page-2 (accessed on 20 October 2025).
-
Thyme. Thymus vulgaris. Available online: https://www.monticello.org/house-gardens/in-bloom-at-monticello/thyme/ (accessed on 20 October 2025).
-
Yaghini H, Sabzalian MR, Rahimmalek M, Garavand T, Maleki A, Mirlohi A. Seed set in inter specific crosses of male sterile Mentha spicata with Mentha longifolia. Euphytica 2020, 216, 46. DOI:10.1007/s10681-020-2578-z [Google Scholar]
-
Talbot SC, Pandelova I, Lange BM, Vining KJ. A first look at the genome structure of hexaploid “Mitcham” peppermint (Mentha × piperita L.). G3 Genes Genomes Genet. 2024, 14, jkae195. DOI:10.1093/g3journal/jkae195 [Google Scholar]
-
Pyne RM, Honig JA, Vaiciunas J, Wyenandt CA, Simon JE. Population structure, genetic diversity and downy mildew resistance among Ocimum species germplasm. BMC Plant Biol. 2018, 18, 69. DOI:10.1186/s12870-018-1284-7 [Google Scholar]
-
Chowdhury T, Mandal A, Roy SC, De Sarker D. Diversity of the genus Ocimum (Lamiaceae) through morpho-molecular (RAPD) and chemical (GC–MS) analysis. J. Genet. Eng. Biotechnol. 2017, 15, 275–286. DOI:10.1016/j.jgeb.2016.12.004 [Google Scholar]
-
Nazar N, Howard C, Slater A, Sgamma T. Challenges in Medicinal and Aromatic Plants DNA Barcoding—Lessons from the Lamiaceae. Plants 2022, 11, 137. DOI:10.3390/plants11010137 [Google Scholar]
-
Yu X, Chen Z, Li S, Qi X, Fang H, Bai Y, et al. A stable method for Agrobacterium-mediated transformation of Mentha piperita and Mentha canadensis using internodal explants. In Vitro Cell. Dev. Biol.-Plant. 2022, 58, 1038–1047. DOI:10.1007/s11627-022-10294-5 [Google Scholar]
-
Novak J. Molecular support for genetic improvement of Lamiaceae. In Proceedings of the I International Symposium on the Labiatae: Advances in Production, Biotechnology and Utilisation, Sanremo, Italy, 22 February 2006. DOI:10.17660/ActaHortic.2006.723.4 [Google Scholar]
-
Gupta AK, Mishra R, Singh AK, Srivastava A, Lal RK. Genetic variability and correlations of essential oil yield with agro-economic traits in Mentha species and identification of promising cultivars. Ind. Crops Prod. 2017, 95, 726–732. DOI:10.1016/j.indcrop.2016.11.041 [Google Scholar]
-
IPGRI. Crop Genetic Resources for Climate Resilience. 2025. Available online: https://www.frontiersin.org/research-topics/78498/smart-breeding-and-crop-genetic-resources-for-climate-resilience (accessed on 3 March 2026).
-
Chiang T-Y, Kumar Y, Khan F, Rastogi S, Shasany AK. Genome-wide detection of terpene synthase genes in holy basil (Ocimum sanctum L.). PLoS ONE 2018, 13, e0207097. DOI:10.1371/journal.pone.0207097 [Google Scholar]
-
Xu S, Li F, Zhou F, Li J, Cai S, Yang S, et al. Efficient targeted mutagenesis in tetraploid Pogostemon cablin by the CRISPR/Cas9-mediated genomic editing system. Hortic. Res. 2024, 11, uhae021. DOI:10.1093/hr/uhae021 [Google Scholar]
-
Varshney G, Hasley JAR, Navet N, Tian M. CRISPR/Cas9-mediated mutagenesis of sweet basil candidate susceptibility gene ObDMR6 enhances downy mildew resistance. PLoS ONE 2021, 16, e0253245. DOI:10.1371/journal.pone.0253245 [Google Scholar]
-
Tripathi T. Spices in Indian history: A multifaceted exploration of trade, medicine and religious practices. Int. J. Appl. Res. 2024, 10, 4–11. DOI:10.22271/allresearch.2024.v10.i8a.11909 [Google Scholar]
-
Modiri M, Semnani AA. Maritime Silk Road or spice road (cultural and civilizational opportunities). In Proceedings of the Eleventh United Nations Conference on the Standardization of Geographical Names, New York, NY, USA, 8–17 August 2017. [Google Scholar]
-
Mint Farming in Washington. Available online: https://www.historylink.org/File/20562 (accessed on 20 October 2025).
-
The Story of Mint Through the Ages in 2 Minutes. Available online: https://medium.com/@jacklohan20/the-story-of-mint-through-the-ages-in-2-minutes-33a553241866 (accessed on 20 October 2025).
-
Rosemary. Available online: https://en.wikipedia.org/wiki/Rosemary (accessed on 20 October 2025).
-
Herbs in History: Basil. Available online: https://www.ahpa.org/herbs_in_history_basil (accessed on 20 October 2025).
-
Peter KV. Handbook of Herbs and Spices, 3rd ed.; Woodhead publishing: Cambridge, UK, 2006; pp. 285–365. [Google Scholar]
-
Oregano. Available online: https://floridaheritagefoods.com/french-collection/oregano/ (accessed on 20 October 2025).
-
Rosemary. Available online: https://www.britannica.com/plant/rosemary (accessed on 20 October 2025).
-
Davidson A. The Oxford Companion to Food, 3rd ed.; Oxford University Press: Oxford, UK, 2014; pp. 73–107. [Google Scholar]
-
Our Unique Flora and Fauna. Available online: https://www.pitches-store.co.nz/about-us/blog/post/our-unique-flora-and-fauna (accessed on 20 October 2025).
-
Thymus vulgaris. Available online: https://www.nzpcn.org.nz/flora/species/thymus-vulgaris/ (accessed on 20 October 2025).
-
FAO. The State of Food and Agriculture. 1967. Available online: https://www.fao.org/3/a0075e00/ (accessed on 3 March 2026).
-
López-Hernández F, Cortés AJ. Whole Transcriptome Sequencing Unveils the Genomic Determinants of Putative Somaclonal Variation in Mint (Mentha L.). Int. J. Mol. Sci. 2022, 23, 5291. DOI:10.3390/ijms23105291 [Google Scholar]
-
Plitta-Michalak BP, Naskręt-Barciszewska MZ, Barciszewski J, Chmielarz P, Michalak M. Epigenetic Integrity of Orthodox Seeds Stored under Conventional and Cryogenic Conditions. Forests 2021, 12, 288. DOI:10.3390/f12030288 [Google Scholar]
-
Phippen W, Simon J. Anthocyanin inheritance and instability in purple basil (Ocimum basilicum L.). J. Hered. 2000, 91, 289–296. DOI:10.1093/jhered/91.4.289 [Google Scholar]
-
Gurav TP, Dholakia BB, Giri AP. A glance at the chemodiversity of Ocimum species: Trends, implications, and strategies for the quality and yield improvement of essential oil. Phytochem. Rev. 2022, 21, 879–913. DOI:10.1007/s11101-021-09767-z [Google Scholar]
-
Malav P, Pandey A, Bhatt KC, Gopala Krishnan S, Bisht IS. Morphological variability in holy basil (Ocimum tenuiflorum L.) from India. Genet. Resour. Crop Evol. 2015, 62, 1245–1256. DOI:10.1007/s10722-015-0227-5 [Google Scholar]
-
Cáceres F, Vallès J, Garnatje T, Gras A. Ethnobotanical insights into the medicinal and food uses of Lamiaceae in the Mediterranean region: A systematic review and meta-analysis. Plants People Planet 2025, 1–17. DOI:10.1002/ppp3.70146 [Google Scholar]
-
Chen X, Zhang F, Yao L. Chloroplast DNA molecular characterization and leaf volatiles analysis of mint (Mentha; Lamiaceae) populations in China. Ind. Crops Prod. 2012, 37, 270–274. DOI:10.1016/j.indcrop.2011.11.011 [Google Scholar]
-
Matthew JO, Oziegbe M, Azeez SO, Ajose TE, Okoyo ME. Polyploidization and speciation: Patterns of natural hybridization and gene flow in basil (Ocimum spp.). Not. Sci. Biol. 2022, 14, 11289. DOI:10.55779/nsb14311289 [Google Scholar]
-
Thakur VV, Tiwari S, Tripathi N, Tiwari G, Sapre S. DNA barcoding and phylogenetic analyses of Mentha species using rbcL sequences. Ann. Phytomed. 2016, 5, 59–62. Available online: https://www.researchgate.net/profile/Vishwa-Vijay-Thakur/publication/306400737_DNA_barcoding_and_phylogenetic_analyses_of_mentha_species_using_rbcL_sequences/links/57bd4de408ae6c703bc5dafe/DNA-barcoding-and-phylogenetic-analyses-of-mentha-species-using-rbcL-sequences.pdf (accessed on 20 October 2025).
-
Curto MA, Puppo P, Ferreira D, Nogueira M, Meimberg H. Development of phylogenetic markers from single-copy nuclear genes for multi locus, species level analyses in the mint family (Lamiaceae). Mol. Phylogenet. Evol. 2012, 63, 758–767. DOI:10.1016/j.ympev.2012.02.010 [Google Scholar]
-
Guo XY. Lavender “Luoshen” Obtains National Plant Variety Rights with Exceptional Waterlogging Resistance, Successfully Survives Summer in Beijing. China Flower News 2023, W04, 1–3. DOI:10.38297/n.cnki.nzghh.2023.000104 [Google Scholar]
-
Kamatou GPP, Makunga NP, Ramogola WPN, Viljoen AM. South African Salvia species: A review of biological activities and phytochemistry. J. Ethnopharmacol. 2008, 119, 664–672. DOI:10.1016/j.jep.2008.06.030 [Google Scholar]
-
Siqueira IR, Vanzella C, Lovatel GA, Bertoldi K, Spindler C, Moysés FDS, et al. American Basil, Ocimum americanum, Has Neuroprotective Properties in the Aging Process. Nutrients 2025, 17, 2368. DOI:10.3390/nu17142368 [Google Scholar]
-
Li B, Kim M. Cultivation of Perilla frutescens (Lamiaceae) in prehistoric Korea. J. Archaeol. Sci. Rep. 2021, 40, 103224. DOI:10.1016/j.jasrep.2021.103224 [Google Scholar]
-
Ancient Rome’s Sacred Plants: History and Symbolism. Available online: https://weirditaly.com/2023/01/30/ancient-romes-sacred-plants-history-and-symbolism/#:~:text=Romans%20are%20credited%20with%20spreading,liqueurs%20with%20its%20aromatic%20scent (accessed on 20 October 2025).
-
D’Agostino A, Di Marco G, Marvelli S, Marchesini M, Rizzoli E, Rolfo MF, et al. Neolithic dental calculi provide evidence for environmental proxies and consumption of wild edible fruits and herbs in central Apennines. Commun. Biol. 2022, 5, 1384. DOI:10.1038/s42003-022-04354-0 [Google Scholar]
-
Naithani S. History and Science of Cultivated Plants, 1st ed.; Oregon State University: Corvallis, OR, USA, 2021; pp. 33–34. [Google Scholar]
-
[Chinese Herbal Medicine at the Door] Mint Cool and Fragrant. Available online: https://pujian.com.my/chinese-herbal-medicine-at-the-door-mint-cool-and-fragrant/ (accessed on 20 October 2025).
-
Waman AA, Smitha GR, Bohra P. A Review on Clonal Propagation of Medicinal and Aromatic Plants through Stem Cuttings for Promoting their Cultivation and Conservation. Curr. Agri. Res. 2019, 7, 122–138. DOI:10.12944/carj.7.2.01 [Google Scholar]
-
Vining KJ, Hummer KE, Bassil NV, Lange BM, Khoury CK, Carver D. Crop Wild Relatives as Germplasm Resource for Cultivar Improvement in Mint (Mentha L.). Front. Plant Sci. 2020, 11, 1217. DOI:10.3389/fpls.2020.01217 [Google Scholar]
-
Wang YP, Fu LG. Preliminary Identification and Analysis of Mentha in Suburban Areas of Shanghai. Chung Ts’ao Yao 2001, 74–75. Available online: https://kns.cnki.net/kcms2/article/abstract?v=niGKtOOGHVft8Sl_QwtlBFzE_5g-1FXcmwOVS_fBgeYiOHE2FFxmF4hYrn138C8q_i4goJ5GaN67MlRN83o1DqJBabsDQ08HK9cj-UGbIplTfUIcjrvcUY1tesh_E6TfO_zo_BWhoz1vXUexTO38yOYZFlEILbxJRucCtbrEMxBCVdv2qcMYMQ==&uniplatform=NZKPT&language=CHS (accessed on 20 October 2025).
-
White O, Biswas M, Wendawek A, Dussert Y, Kebede F, Nichols R, et al. Maintenance and expansion of genetic and trait variation following domestication in a clonal crop. Mol. Ecol. 2023, 32, 4165–4180. DOI:10.1111/mec.17033 [Google Scholar]
-
Vining KJ, Zhang Q, Tucker AO, Smith C, Davis TM. Mentha longifolia (L.) L.: A Model Species for Mint Genetic Research. HortScience 2005, 40, 1225–1229. DOI:10.21273/hortsci.40.5.1225 [Google Scholar]
-
Prasad P, Singh V, Aftab N, Gupta A, Kishor R, Kushwaha HK, et al. Gamma irradiation-induced variability in morpho-agronomic and oil quality traits of Mentha piperita L. Int. J. Radiat. Biol. 2021, 97, 737–745. DOI:10.1080/09553002.2021.1893855 [Google Scholar]
-
Kumari N, Singh M. Effect of Chemical Mutagen on Seed Germination, Morphological and Essential Oil Content in Ocimum basilicum L. Asian J. Biol. Life Sci. 2024, 12, 460–465. DOI:10.5530/ajbls.2023.12.61 [Google Scholar]
-
Gurav TP, Jayaramaiah RH, Punekar SA, Dholakia BB, Giri AP. Generation of novelties in the genus Ocimum as a result of natural hybridization: A morphological, genetical and chemical appraisal. Ind. Crops Prod. 2020, 156, 112859. DOI:10.1016/j.indcrop.2020.112859 [Google Scholar]
-
Rogo U, Fambrini M, Pugliesi C. Embryo Rescue in Plant Breeding. Plants 2023, 12, 3106. DOI:10.3390/plants12173106 [Google Scholar]
-
Bornowski N, Hamilton JP, Liao P, Wood JC, Dudareva N, Buell CR. Genome sequencing of four culinary herbs reveals terpenoid genes underlying chemodiversity in the Nepetoideae. DNA Res. 2020, 27, dsaa016. DOI:10.1093/dnares/dsaa016 [Google Scholar]
-
Sarrou E, Martinidou E, Palmieri L, Poulopoulou I, Trikka F, Masuero D, et al. High throughput pre-breeding evaluation of Greek oregano (Origanum vulgare L. subsp. hirtum) reveals multi-purpose genotypes for different industrial uses. J. Appl. Res. Med. Aromat. Plants. 2023, 37, 100516. DOI:10.1016/j.jarmap.2023.100516 [Google Scholar]
-
Li J, Wang Y, Dong Y, Zhang W, Wang D, Bai H, et al. The chromosome-based lavender genome provides new insights into Lamiaceae evolution and terpenoid biosynthesis. Hortic. Res. 2021, 8, 53. DOI:10.1038/s41438-021-00490-6 [Google Scholar]
-
Han D, Li W, Hou Z, Lin C, Xie Y, Zhou X, et al. The chromosome-scale assembly of the Salvia rosmarinus genome provides insight into carnosic acid biosynthesis. Plant J. 2023, 113, 819–832. DOI:10.1111/tpj.16087 [Google Scholar]
-
Lai Y, Ma J, Zhang X, Xuan X, Zhu F, Ding S, et al. High-quality chromosome-level genome assembly and multi-omics analysis of rosemary (Salvia rosmarinus) reveals new insights into the environmental and genome adaptation. Plant Biotechnol. J. 2024, 22, 1833–1847. DOI:10.1111/pbi.14305 [Google Scholar]
-
Ben-Naim Y, Falach L, Cohen Y. Transfer of Downy Mildew Resistance from Wild Basil (Ocimum americanum) to Sweet Basil (O. basilicum). Phytopathology 2018, 108, 114–123. DOI:10.1094/phyto-06-17-0207-r [Google Scholar]
-
Edwards J, Parbery DG, Taylor PA, Halloran GM. Effects of Puccinia menthae on growth and yield of Todd’s Mitcham peppermint (Mentha × piperita). Aust. J. Agric. Res. 1999, 50, 1273–1278. DOI:10.1071/AR99013 [Google Scholar]
-
Ali M, Li P, She G, Chen D, Wan X, Zhao J. Transcriptome and metabolite analyses reveal the complex metabolic genes involved in volatile terpenoid biosynthesis in garden sage (Salvia officinalis). Sci. Rep. 2017, 7, 16074. DOI:10.1038/s41598-017-15478-3 [Google Scholar]
-
Davis EM, Ringer KL, McConkey ME, Croteau R. Monoterpene Metabolism. Cloning, Expression, and Characterization of Menthone Reductases from Peppermint. Plant Physiol. 2005, 137, 873–881. DOI:10.1104/pp.104.053306 [Google Scholar]
-
Leggatt E, Griffiths A, Budge S, Stead AD, Gange AC, Devlin PF. Addition of Arbuscular Mycorrhizal Fungi Enhances Terpene Synthase Expression in Salvia rosmarinus Cultivars. Life 2023, 13, 315. DOI:10.3390/life13020315 [Google Scholar]
-
Bai Y, Zhang T, Zheng X, Li B, Qi X, Xu Y, et al. Overexpression of a WRKY transcription factor McWRKY57-like from Mentha canadensis L. enhances drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2023, 23, 216. DOI:10.1186/s12870-023-04213-y [Google Scholar]
-
Rezaie R, Abdollahi Mandoulakani B, Fattahi M. Cold stress changes antioxidant defense system, phenylpropanoid contents and expression of genes involved in their biosynthesis in Ocimum basilicum L. Sci. Rep. 2020, 10, 5290. DOI:10.1038/s41598-020-62090-z [Google Scholar]
-
Wang Y, Ye H, Ren F, Ren X, Zhu Y, Xiao Y, et al. Comparative Transcriptome Analysis Revealed Candidate Gene Modules Involved in Salt Stress Response in Sweet Basil and Overexpression of ObWRKY16 and ObPAL2 Enhanced Salt Tolerance of Transgenic Arabidopsis. Plants 2024, 13, 1487. DOI:10.3390/plants13111487 [Google Scholar]
-
Simon J, Deschamps C. Agrobacterium tumefaciens -mediated transformation of Ocimum basilicum and O. citriodorum. Plant Cell Rep. 2002, 21, 359–364. DOI:10.1007/s00299-002-0526-0 [Google Scholar]
-
Yun HR, Chen C, Kim JH, Kim HE, Karthik S, Kim HJ, et al. Genome-edited HEADING DATE 3a knockout enhances leaf production in Perilla frutescens. Front. Plant Sci. 2023, 14, 1133518. DOI:10.3389/fpls.2023.1133518 [Google Scholar]
-
Younes al-Aboud MF, Abedi B, Aroiee H, Bayanati M, Sayyad-Amin P. Improving the Germination and Growth of Basil Seed (Ocimum basilicum L.) under Salinity Stress Conditions with the Use of Priming Salicylic Acid and Potassium Nitrate. Russ. J. Plant Physiol. 2025, 72, 9–13. DOI:10.1134/s1021443724608966 [Google Scholar]
-
Madeiras AM, Boyle TH, Autio WR. Stratification, gibberellic acid, scarification, and seed lot influence on rosemary seed germination. Seed Technol. 2009, 31, 55–65. Available online: https://www.jstor.org/stable/23433506 (accessed on 20 October 2025).
-
Szekely-Varga Z, Kentelky E, Cantor M. Effect of gibberellic acid on the seed germination of Lavandula angustifolia mill. Rom. J. Hortic. 2021, 2, 169–176. DOI:10.51258/RJH.2021.22 [Google Scholar]
-
Ekren S, Gökçöl A, Yıldırım Keskinoğlu A, Paylan İC. Optimizing germination performance of Lamiaceae family seeds: Insights from research. J. Plant Dis. Prot.2024, 131, 999–1008. DOI:10.1007/s41348-023-00857-y [Google Scholar]
-
Triharyanto E, Pujiasmanto B, Pardono P, Fa’izah AT. Response of Growth and Yield of Mint (Mentha spicata L.) Cuttings to Auxin and Composition of Planting Media. Agrotechnol. Res. J. 2023, 7, 1–7. DOI:10.20961/agrotechresj.v7i1.76700 [Google Scholar]
-
Karakaş İ, İzci B. Effects of Rooting Mediums and Growth Regulating Agents on Rooting Parameters of Lavender and Lavandin Cuttings (Lavandula sp.). J. Agri. Pro. 2024, 5, 138–152. DOI:10.56430/japro.1485102 [Google Scholar]
-
Lykokanellos G, Lagogiannis I, Liopa-Tsakalidi A, Barla SA, Salachas G. Optimizing Rooting and Growth of Salvia rosmarinus Cuttings in Soilless Systems Affected by Growth Regulators. Plants 2025, 14, 2210. DOI:10.3390/plants14142210 [Google Scholar]
-
Hollick JR, Kubota C. Effect of Self- and Inter-Cultivar Grafting on Growth and Nutrient Content in Sweet Basil (Ocimum basilicum L.). Front. Plant Sci. 2022, 13, 921440. DOI:10.3389/fpls.2022.921440 [Google Scholar]
-
Loupit G, Brocard L, Ollat N, Cookson SJ. Grafting in plants: recent discoveries and new applications. J. Exp. Bot. 2023, 74, 2433–2447. DOI:10.1093/jxb/erad061 [Google Scholar]
-
Abdalla N, El-Ramady H, Seliem MK, El-Mahrouk ME, Taha N, Bayoumi Y, et al. An Academic and Technical Overview on Plant Micropropagation Challenges. Horticulturae 2022, 8, 677. DOI:10.3390/horticulturae8080677 [Google Scholar]
-
Vaidya BN, Asanakunov B, Shahin L, Jernigan HL, Joshee N, Dhekney SA. Improving micropropagation of Mentha × piperita L. using a liquid culture system. In Vitro Cell. Dev. Biol.-Plant. 2019, 55, 71–80. DOI:10.1007/s11627-018-09952-4 [Google Scholar]
-
Tsuro M, Yasue S, Otagaki S. Varietal Differences of Regenerative Ability in Sweet Basil (Ocimum basilicum L.). Environ. Control. Biol. 2025, 63, 91–95. DOI:10.2525/ecb.63.91 [Google Scholar]
-
Misra P, Chaturvedi HC. Micropropagation of Rosmarinus officinalis L. Plant Cell Tiss. Org. Cult. 1984, 3, 163–168. DOI:10.1007/BF00033737 [Google Scholar]
-
Kruglova NN, Seldimirova OA, Zinatullina AE, Yegorova NA. Histological Aspects of a Long-Term In Vitro Culture of Morphogenic Calluses of Lavandula angustifolia Mill. Biol. Bull. Russ. Acad. Sci. 2025, 52, 6. DOI:10.1134/S1062359024608760 [Google Scholar]
-
Goleniowski ME, Flamarique C, Bima P. Micropropagation of oregano (Origanum vulgare × applii) from meristem tips. In Vitro Cell. Dev. Biol.-Plant. 2003, 39, 125–128. DOI:10.1079/IVP2002361 [Google Scholar]
-
Avasiloaiei DI, Calara M, Brezeanu PM, Murariu OC, Brezeanu C. On the Future Perspectives of Some Medicinal Plants within Lamiaceae Botanic Family Regarding Their Comprehensive Properties and Resistance against Biotic and Abiotic Stresses. Genes 2023, 14, 955. DOI:10.3390/genes14050955 [Google Scholar]
-
Chakrabartty I, Mohanta YK, Nongbet A, Mohanta TK, Mahanta S, Das N, et al. Exploration of Lamiaceae in Cardio Vascular Diseases and Functional Foods: Medicine as Food and Food as Medicine. Front. Pharmacol. 2022, 13, 894814. DOI:10.3389/fphar.2022.894814 [Google Scholar]
-
Krishnamurthy KS, Sharon A, Meena RS, Janakiram T, Nirmal Babu K. Improved Varieties of Spice Crops. In Handbook of Spices in India: 75 Years of Research and Development; Ravindran PN, Sivaraman K, Devasahayam S, Babu KN, Eds.; Springer Nature: Singapore, 2024; pp. 283–396. [Google Scholar]
-
Zhang XB. Introduction, Cultivation, Identification and Comprehensive Evaluation of Basil Germplasm Resources. Ph.D. Dissertation, Hainan University, Haikou, China, 2013. [Google Scholar]
-
Li H, Bai HT. A Romantic Legend from the Mediterranean to the Foot of the Tianshan Mountains: Introduction of Lavender Resources and New Variety Breeding. Life World 2019, 16–21. Available online: https://kns.cnki.net/kcms2/article/abstract?v=niGKtOOGHVf8Oyika0XixaKbMo3HXtzWro6L2XNMR6oKbuZuBQCbuyG9ADsalRXjAJOTy5cIF1khWOfr0_Zx2Ix-oc9u6zPt4h6xulyVfcM6C61aLhLHIjibzBl6crlfKzlkHrCeM7srfkAMSPQT8ue4bhmq7fYZe-ACcCJsLfXuuAT705n0Og==&uniplatform=NZKPT&language=CHS (accessed on 20 October 2025).
-
Zheng MD, Wang ML, Yu JQ. Identification of 13 medicinal and edible aromatic plants from labiatae based on dna barcodes. Mod. Food Sci. Technol. 2025, 41, 341–348. DOI:10.13982/j.mfst.1673-9078.2025.4.0225 [Google Scholar]
-
Thakur VV, Tripathi N, Tiwari S. DNA barcoding of some medicinally important plant species of Lamiaceae family in India. Mol. Biol. Rep. 2021, 48, 3097–3106. DOI:10.1007/s11033-021-06356-3 [Google Scholar]
-
Gowda MRS, Arpitha K, Gamyashree K, Prabhu KN, Kumar AN, Srinivas KVNS, et al. Chemical and molecular diversity of rosemary (Salvia rosmarinus L.) clones. Genet. Resour. Crop Evol. 2024, 71, 2003–2018. DOI:10.1007/s10722-023-01758-7 [Google Scholar]
-
Permadi N, Akbari SI, Prismantoro D, Indriyani NN, Nurzaman M, Alhasnawi AN, et al. Traditional and next-generation methods for browning control in plant tissue culture: current insights and future directions. Curr. Plant Biol. 2024, 38, 100339. DOI:10.1016/j.cpb.2024.100339 [Google Scholar]
-
Davidović Gidas J, Zeljković S, Đekić N, Đurić G. LED lights and plant growth regulators enhance the in vitro mass propagation of rosemary (Rosmarinus officinalis L.). Eur. J. Hortic. Sci. 2025, 90, 1. DOI:10.1079/ejhs.2025.0008 [Google Scholar]
-
Hasan N, Choudhary S, Naaz N, Sharma N, Laskar RA. Recent advancements in molecular marker-assisted selection and applications in plant breeding programmes. J. Genet. Eng. Biotechnol. 2021, 19, 128. DOI:10.1186/s43141-021-00231-1 [Google Scholar]
-
Vining KJ, Johnson SR, Ahkami A, Lange I, Parrish AN, Trapp SC, et al. Draft Genome Sequence of Mentha longifolia and Development of Resources for Mint Cultivar Improvement. Mol. Plant 2017, 10, 323–339. DOI:10.1016/j.molp.2016.10.018 [Google Scholar]
-
Giuseppe A, Raffaella EM. The First Genome-Wide Mildew Locus O Genes Characterization in the Lamiaceae Plant Family. Int. J. Mol. Sci. 2023, 24, 13627. DOI:10.3390/ijms241713627 [Google Scholar]
-
Gao Q, Yue G, Li W, Wang J, Xu J, Yin Y. Recent progress using high-throughput sequencing technologies in plant molecular breeding. J. Integr. Plant Biol. 2012, 54, 215–227. DOI:10.1111/j.1744-7909.2012.01115.x [Google Scholar]
-
Yi D, Wang Z, Peng M. Comprehensive Review of Perilla frutescens: Chemical Composition, Pharmacological Mechanisms, and Industrial Applications in Food and Health Products. Foods 2025, 14, 1252. DOI:10.3390/foods14071252 [Google Scholar]
-
Kaur S, Seem K, Ali A, Jaiswal S, Gumachanamardi P, Kaur G, et al. A comprehensive review on nutritional, nutraceutical, and industrial perspectives of perilla (Perilla frutscens L.) seeds—An orphan oilseed crop. Heliyon 2024, 10, e33281. DOI:10.1016/j.heliyon.2024.e33281 [Google Scholar]