Emerging Technologies Empowering the Biosynthesis of Paclitaxel

Emerging Technologies Empowering the Biosynthesis of Paclitaxel
Xiaonan Liu 1,*,† Yuanyuan Zhang 1,† Jing Li 2,† Huifeng Jiang 3,4,*
Received: 26 January 2026 Revised: 24 February 2026 Accepted: 30 April 2026 Published: 18 May 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
Graphical Abstract
1. Introduction
Paclitaxel (Taxol) is a tetracyclic diterpene compound isolated from the rare Taxus species, standing out as one of the most potent natural anticancer agents ever discovered. Its remarkable therapeutic efficacy stems from a unique mechanism of action: it stabilizes microtubules by preventing their depolymerization, thereby arresting mitosis and inhibiting cancer cell proliferation [1]. This compound has attracted widespread scientific interest not only for its clinical importance but also for its limited natural availability, complex molecular structure, and the intricate biosynthetic pathway required for its production. Recent research on paclitaxel biosynthesis has achieved great progress, and almost all the enzymes involved in its pathway have been identified. However, substrate and functional promiscuity among key enzymes transform paclitaxel biosynthesis into a complex metabolic network, making it highly challenging to fully decipher the reaction process [2]. Moreover, although the synthesis of taxa-4(5),11(12)-diene in microorganisms can reach gram-scale yields, subsequent oxidation steps remain difficult to advance. Several recent review articles have summarized progress in elucidating the biosynthetic mechanism of paclitaxel, as well as the key challenges hindering the sustainable production of taxane compounds, and also proposed the state-of-the-art synthetic biology and artificial intelligence strategies to overcome these challenges [3,4,5]. This work focuses mainly on advances in the evolutionary origin, pathway identification, and artificial biosynthesis of paclitaxel formation. The emerging technologies, such as complex genomic sequencing, the mpXsn method, MALDI-IMS, dropout experiments, and the oxidases toolbox, will also be discussed, as well as the catalytic mechanism studies of key catalytic components.
The yew tree is a rare species listed as a national key protected wild plant, often praised as “Green Gold” for its unique medicinal, timber, and ornamental value. The sequencing of the Taxus genome, especially the releases of high-quality genomes for Taxus chinensis [6], Taxus wallichiana [7], and Taxus yunnanensis [8], has substantially accelerated research on paclitaxel biosynthesis [9]. These genomic resources, integrated with comprehensive transcriptomic datasets and advanced spatial metabolomics techniques like MALDI-IMS, have enabled unprecedented insights into the evolutionary origins and metabolic intricacies of taxane biosynthesis. For instance, MALDI-IMS analysis of Pseudotaxus chienii, combined with its genome sequencing, has provided crucial clues about the origin and evolution of this pathway [10]. Furthermore, innovative methodologies such as multiplexed perturbation × single nuclei sequencing (mpXsn), which couples single-nucleus RNA-seq with high-throughput phenotypic perturbation, offer a powerful platform for deconstructing complex plant metabolic networks and identifying regulatory factors, such as the nuclear transport factor 2 (NTF2)-like protein FoTO1 [11].
Significant progress has been made in elucidating the paclitaxel biosynthetic pathway, with nearly all involved enzymes now identified. Recent research has yielded pivotal discoveries that overcome long-standing barriers. Key breakthroughs include the elucidation of the oxetane ring-forming mechanism at C4, C5, and C20 positions [12,13,14,15], the identification of the nuclear transport factor 2 (NTF2)-like protein FoTO1 (which complexes with TXS and T5αOH to enhance early pathway yield by 10 to 17-fold) [11], T9αOHs for different pathways, T7AT and different T1βOHs, deacetylases and taxane C-9-oxidase (T9ox), β-phenylalanine-CoA ligase (PCL), and the clarification of final steps such as C2′α hydroxylation and 3′-N benzoylation [16]. Concurrently, enzyme engineering has produced innovative tools, such as artificial oxidase toolboxes derived from TteUPO mutants, enabling site-directed oxidation of taxanes at multiple positions and offering unconventional strategies for creating paclitaxel analogs [17]. In addition, versatile 2-oxoglutarate-dependent dioxygenases, such as Tm576, TcOGD1, and TcOGD2, could catalyze skeleton reconstructions and oxidation modifications of toxoids, which expanded the chemical space of taxane diterpenoids [2,18].
While advances in synthetic biology have made green and sustainable paclitaxel intermediates (particularly taxadiene), production via engineered cell factories is increasingly viable. Reconstruction of the downstream pathway after taxa-4(5),11(12)-diene still faces critical bottlenecks, including intrinsically low P450 catalytic efficiency, poor compatibility with heterologous hosts, and metabolic flux diversion due to enzyme promiscuity [4]. Consequently, a central research focus is the reconstruction and enhancement of the complete pathway in amenable heterologous hosts such as Escherichia coli, yeast, Nicotiana benthamiana, and cyanobacteria. Current strategies to overcome these hurdles include: (1) developing novel chassis systems (such as non-conventional yeasts, fungi, algae) for better compatibility with plant P450s; (2) employing hybrid biosynthetic-chemical approaches; (3) deploying regulatory scaffolds like FoTO1 to minimize off-target oxidation; (4) utilizing machine learning and synthetic microbial consortia for pathway optimization [5]. This review summarizes recent progress across evolutionary origin studies, pathway identification, and artificial biosynthesis strategies (Figure 1). By offering a comprehensive analysis of key enzymatic discoveries and cutting-edge synthetic biology approaches, it aims to enhance the understanding of paclitaxel biosynthesis and catalysis, thereby laying a foundation for the systematic development of sustainable production platforms.
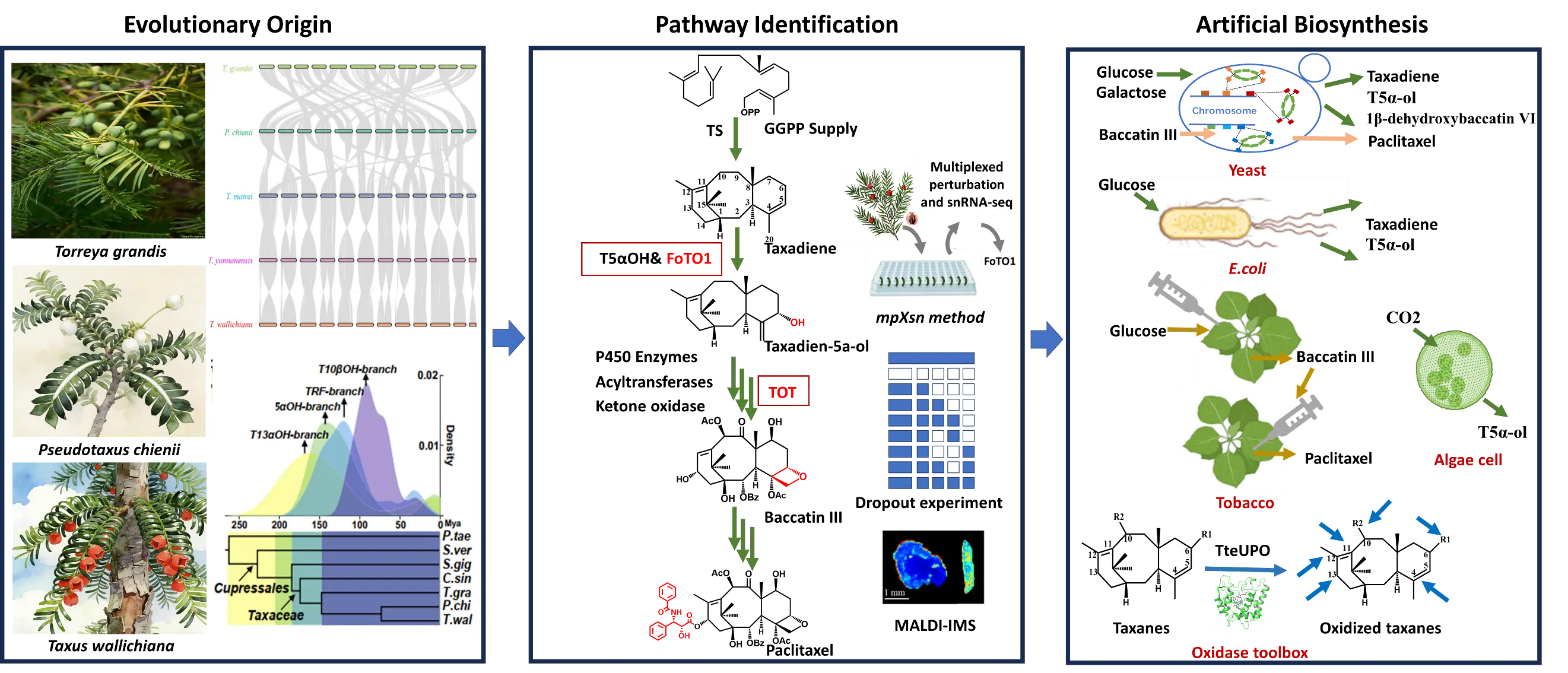
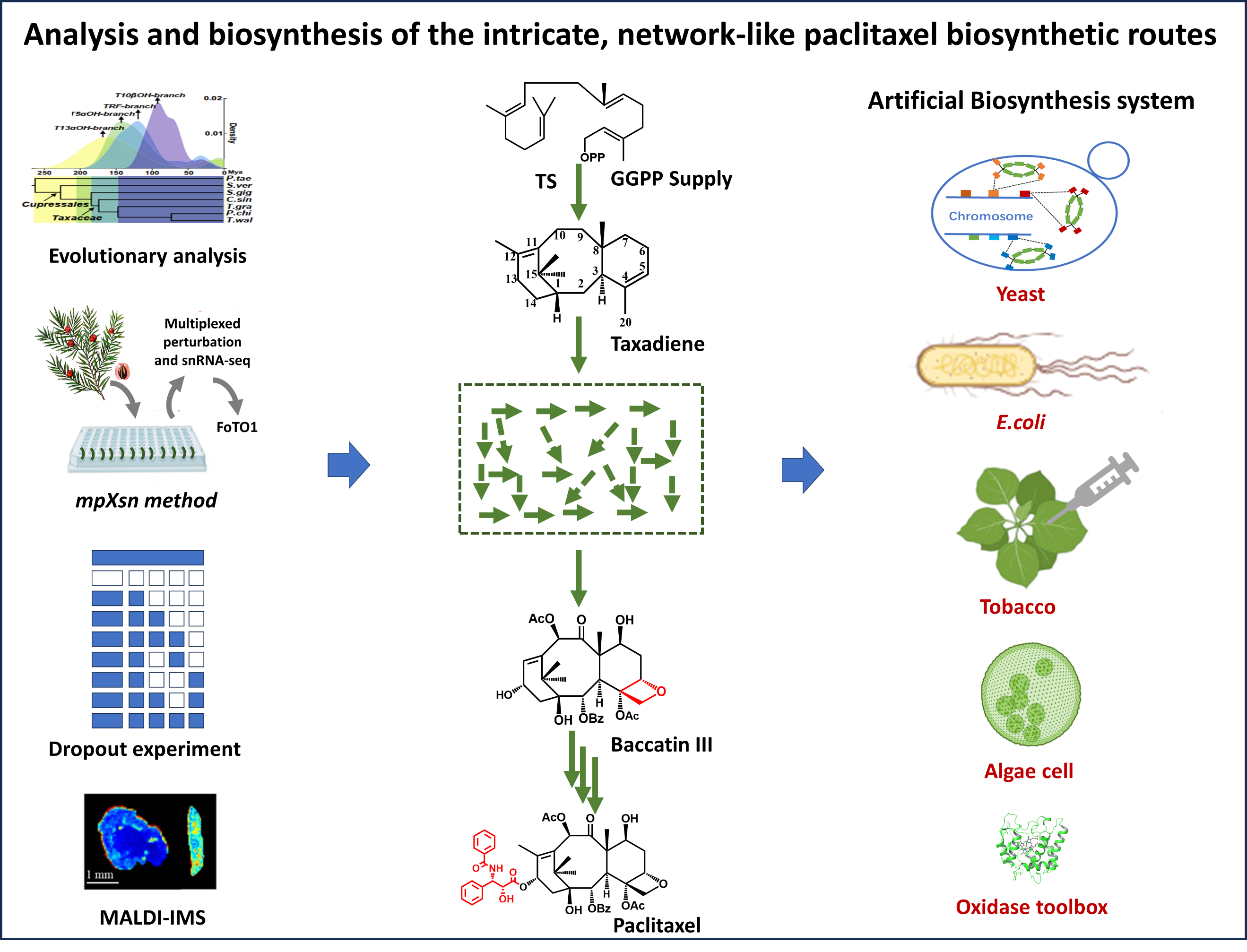
Figure 1. The evolution and synthetic biology research on paclitaxel biosynthesis. Single arrow represents a one-step reaction, three arrows represent multi-step reactions. Green arrows indicate reactions catalyzed by enzymes naturally involved in the taxol biosynthesis pathway, blue arrows indicate catalysis by oxidases not derived from the Taxus speicies. Red boxes highlight key catalytic steps.
2. Advances in the Paclitaxel Biosynthetic Pathway Identification
As a starting point for understanding the paclitaxel synthesis, a systematic review of its biosynthetic pathway is provided below. The biosynthetic pathway of paclitaxel is generally divided into three stages. The first stage involves the synthesis of the key taxane core skeleton, which leads to the formation of taxa-4(5),11(12)-diene. The second stage involves the formation of baccatin III from taxadiene via several hydroxylation and acetylation modifications. The third stage comprises the synthesis of the β-phenylisoserinoyl-CoA side chain and attaching this side chain to baccatin III followed by a series of additional modifications that ultimately result in the formation of paclitaxel (Figure 2).
2.1. Biosynthesis of the Taxa-4(5),11(12)-Diene
The biosynthesis of the taxane core begins with geranylgeranyl pyrophosphate (GGPP), which is a universal precursor to diterpenes. GGPP is formed through the condensation of one molecule of dimethylallyl pyrophosphate (DMAPP) with three molecules of isopentenyl pyrophosphate (IPP), catalyzed by geranylgeranyl pyrophosphate synthase (GGPPS). IPP and DMAPP are primarily synthesized through the 2-C-methyl-D-erythritol-4-phosphate (MEP) pathway and the mevalonate (MVA) pathway [19]. Additionally, an isomerization reaction between DMAPP and IPP can be catalyzed by isopentenyl diphosphate isomerase (IDI). Subsequently, taxa-4(5),11(12)-diene synthase (TS) catalyzes the cyclization of GGPP to generate the taxane diterpene backbone [19,20], which comprises approximately 95% taxa-4(5),11(12)-diene, and taxa-4(20),11(12)-dienetaxadiene with other minor diterpene scaffolds. In plant cells, although the cytosolic MVA pathway can also generate IPP, both GGPPS and TS harbor intrinsic plastid-targeting peptides and thus preferentially utilize IPP synthesized via the MEP pathway in plastids [20]. Following the taxa-4(5),11(12)-diene backbone, a cascade of tailoring modifications ultimately leads to the formation of baccatin III.
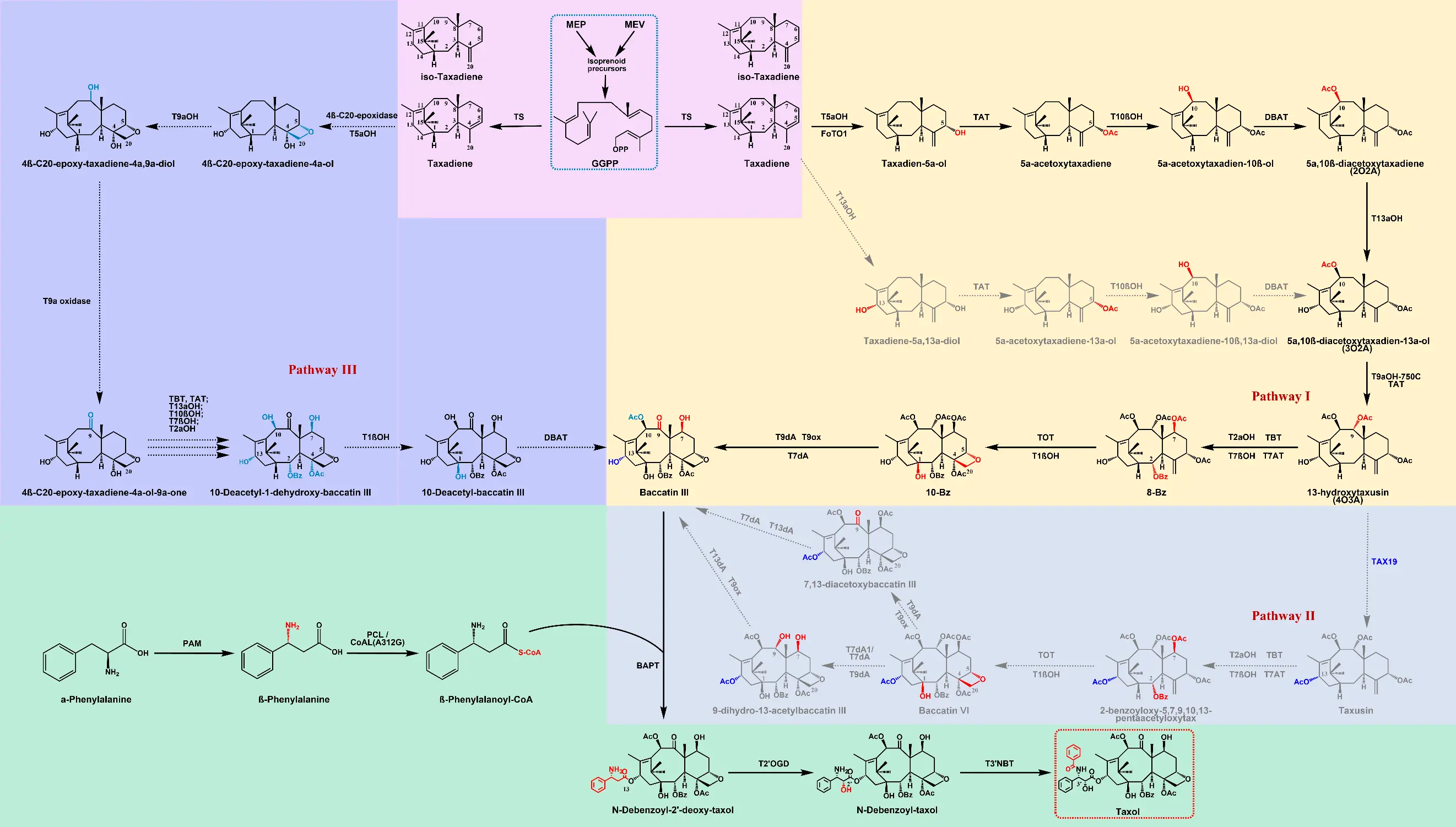
Figure 2. The recently discovered biosynthetic pathway of paclitaxel. TS, taxa-4(5),11(12)-diene synthase; T5αOH, taxane 5α-hydroxylase; TAT, taxa-4(5),11(12)-diene-5α-ol-O-acetyl transferase; T10βOH, taxane 10β-hydroxylase; T13αOH, taxane 13α-hydroxylase; T2αOH, taxane 2α-hydroxylase; T9αOH, taxane 9α-hydroxylase; T7βOH, taxane 7β-hydroxylase; T1βOH, taxane 1β-hydroxylase; TBT, taxane 2α-O-benzoyltransferase; DBAT, 10-deacetyl baccatin III-10-O-acetyltransferase; BAPT, baccatin III-3-amino, 13-phenylpropanoyltransferase; TOT, taxane oxetanase; TAX19, taxane C13-O-acetyltransferase; PAM, phenylalanine aminomutase; PCL, β-phenylalanine coenzyme A ligase; CoA(A312G), a variant of a CoA ligase from Penicillium chrysogenum; T7AT, taxane C-7β-O-acyltransferase; T7dA/T7dA1, taxane 7β-O-deacetylase; T9dA, taxane 9α-O-deacetylase; T9ox, taxane C-9-oxidase; T13dA, taxane C13-O-deacetylase; T2/OGD, taxane 2′α-hydroxylase; T3′NBT, 3’-N-debenzoyl-′-deoxytaxol-N-benzoyltransferase. Single arrow represents a one-step reaction, while multiple arrows represent multi-step reactions. Dashed arrow: putative pathway; Blue dashed box: key precursors; Red dashed box: the final product taxol; Red color: newly added structure in each step; Blue color: putative structure.
2.2. Biosynthesis of the Baccatin III
During the tailoring steps in paclitaxel biosynthesis, cytochrome P450 enzymes mediate most hydroxylation reactions. Several P450 enzymes have been identified that catalyze hydroxylation at the C2, C5, C7, C10, and C13 positions. Recent studies have reported progress in elucidating the functions of the C4β-C20 epoxidase, C1 hydroxylase (T1βOH), different C9 hydroxylases (T9αOHs), C9 oxygenase, taxane-2′α-hydroxylase (T2′αOH), 3′-N-debenzoyl-2′-deoxytaxol N-benzoyltransferase (DBTNBT), and so on [12,15]. After years of effort, especially the explosive innovative discoveries in the past two years, the catalytic components involved in the pathway from taxa-4(5),11(12)-diene to paclitaxel have been almost completely identified (Table 1). It has long been assumed that among all hydroxylation reactions, the first to occur is C5 hydroxylation, catalyzed by taxa-4(5),11(12)-diene-5α-hydroxylase T5αOH [21]. Taxa-4(20),11(12)-diene is directly converted by T5αOH to yield taxa-4(20),11(12)-dien-5α-ol (T5α-ol) [22]. When taxa-4(5),11(12)-diene serves as the substrate, T5αOH converts it into T5α-ol via C5 hydroxylation coupled with double-bond rearrangement. Additionally, T5αOH can catalyze the formation of multiple mono- and di-oxidized byproducts, including OCT (5(12)-oxa-3(11)-cyclotaxane) and iso-OCT (5(11)-oxa-3(11)-cyclotaxane) [21]. Recently, the FoTO1 was found to be crucial for promoting the formation of the desired product T5α-ol during the first oxidation, resolving a long-standing bottleneck in the paclitaxel pathway reconstitution [11]. Subsequently, owing to the high catalytic promiscuity of P450 enzymes and acyltransferases, particularly in oxetane ring formation, multiple catalytic mechanisms have been proposed. The turning point occurred when an α-ketoglutarate/Fe (II)-dependent dioxygenase (ODD), named as TB328, was identified as the catalyst for C4β-C20 epoxidation. This discovery led to the hypothesis that an ODD enzyme mediated reaction accelerates the taxa-4(5),11(12)-diene oxidation cascade [15]. Subsequently, the formation mechanism of oxetane was elucidated by different enzymes. Specifically, recent reports demonstrate that the P450 enzyme T5αOH can also orchestrate oxetane ring formation through a two-step epoxidation mechanism, fundamentally revising earlier assumptions regarding its catalytic limits [23]. In 2024, enzymatic activity assays performed on 58 key candidate genes in a tobacco host system identified a bifunctional P450 enzyme, named TOT1, that participates in the oxetane ring formation of paclitaxel [13]. While recent systematic reviews have chronicled these conceptual shifts [24]. CYP725A55 [13] and TmCYP1 [14] are also the same P450 enzyme that catalyzes the oxetane ester formation via a cascade oxidation-concerted acyl rearrangement mechanism. These divergent results of oxetane ring formation in paclitaxel biosynthesis also suggest that the pathway to paclitaxel may involve a complex network instead of a linear network.
In previous studies, two alternative pathways have been proposed for the downstream reactions following the C5 hydroxylation of taxa-4(5),11(12)-diene: In the first proposed route, T5α-ol undergoes acetylation at the 5α-hydroxyl group, catalyzed by taxa-4(5),11(12)-diene-5α-ol-O-acetyltransferase (TAT), forming Taxa-4(20),11(12)-dien-5α-yl-acetate. This intermediate is then hydroxylated at the C10 position by taxane-10β-hydroxylase (T10βOH), producing Taxa-4(20),11(12)-dien-5α-acetoxy-10β-ol. Alternatively, the CYP725A family taxane 13α-hydroxylase (T13αOH) catalyzes the hydroxylation of T5α-ol at the C13 position, yielding Taxa-4(20),11(12)-diene-5α-13α-diol. Further modifications occur through hydroxylation at C1, C2, C7, and C9, acylation at C2, C5, and C10, and ketonization at C9, although the precise sequence of these steps remains unclear. It was proposed that TB328 and P450 enzyme T5αOH act synergistically to generate 4β-C20-epoxy-taxa-4(5),11(12)-diene-4α-ol, which is subsequently modified through C9 hydroxylation and oxidation. Using the same transient expression system and dropout experiments, taxane-1β-hydroxylase (T1βOH), taxane-9α-hydroxylase (T9αOH), and taxane-9α-oxidase were further identified [15]. However, the precise sequences of the downstream reactions remain unresolved. Subsequently, the intermediate 2-debenzoyltaxane undergoes regioselective benzoylation catalyzed by taxane-2α-O-benzoyltransferase (TBT) to yield 10-deacetylbaccatin III (10-DAB). This is followed by the final modification of the taxane core, in which 10-deacetylbaccatin III-10-O-acetyltransferase (DBAT) catalyzes acetylation of the C10 hydroxyl group of 10-DAB, resulting in the formation of baccatin III [25]. The early steps of the paclitaxel biosynthetic pathway were reconstructed in a tobacco system, demonstrating that taxa-4(20),11(12)-dien-5α-ol can be sequentially converted by early paclitaxel pathway enzymes either in the order TAT-T10βOH-DBAT-T13αOH or T13αOH-TAT-T10βOH-DBAT, producing 5α,10β-diacetoxy-13α-ol as an intermediate for subsequent reactions, ultimately leading to the formation of the 5α,10β-diacetoxy-13-one intermediate [26]. Based on the newly discovered oxetane ring-forming enzyme, the research groups of Jianbin Yan and Zhihua Zhou constructed biosynthetic pathways for baccatin III and 5α,10β-diacetoxy-13-one in tobacco and Saccharomyces cerevisiae, respectively. In addition, both teams identified the enzyme T9αOH1, which is responsible for hydroxylation at the C9 position of the taxane core [12,13]. Using single-nucleus transcriptomic sequencing and a multi-gene perturbation strategy, the group identified key catalysts for paclitaxel, including T9αOH-750C, the scaffold protein FoTO1, two T1βOH variants (T1βOH-184 and T1βOH-686), and T9ox. In contrast to previous findings, they also discovered T7AT, which catalyzes acetylation or benzoylation at the C-7β position, as well as T9dA and T7dA, which remove acetyl groups at the C-9 and C-7 positions, respectively. T7AT is particularly important as it is required for the function of TOT and facilitates the formation of the oxirane ring, a crucial step in the pathway [11]. In 2026, the baccatin III biosynthetic pathway was further elucidated. Through the transcriptomic analysis of T. media cell cultures, four previously unreported deacetylases (T7dA1, T79dA, T13dA1, and T13dA2) were identified belonging to the carboxylesterase family, which plays a pivotal role in the deacetylation of plant-derived natural products. Substrate-feeding experiments utilizing a N. benthamiana transient expression system demonstrated that T7dA1 exhibits higher catalytic activity than the previously reported T7dA. Furthermore, these assays revealed that T79dA functions as a bifunctional deacetylase capable of removing acetyl groups at the C7 and C9 positions [27]. Most importantly, the discovery of the C13-deacetylases T13dA1 and T13dA2 uncovered a cryptic “C13α-O-acetylation-deacetylation module” within the paclitaxel biosynthetic pathway. This critical finding repositioned classic taxoids, such as taxusin and baccatin VI, as bona fide on-pathway intermediates rather than metabolic byproducts. Although TAX19 has been characterized as a native taxane C13α-O-acetyltransferase in Taxus , the baccatin III biosynthetic route necessitates a hydroxyl group at the C13α position [28]. This structural requirement further substantiates the existence of a dynamic “C13α-O-acetylation-deacetylation” process during biosynthesis. Within this framework, 13-hydroxytaxusin is converted to taxusin via the action of TAX19. A subsequent catalytic cascade involving T2αOH, TBT, T7βOH, T7AT, TOT, and T1βOH then transforms taxusin into baccatin VI. From this point, baccatin VI can proceed through two distinct routes to generate baccatin III. In Pathway I, the sequence initiates with deacetylation at the C7 and C9 positions. In vitro steady-state kinetics confirmed the specific catalytic order of T13dA and T9ox, demonstrating that baccatin VI yields baccatin III through the sequential action of T7dA1, T9dA, T13dA, and T9ox. Conversely, Pathway II begins with C9 ketonization orchestrated by T9dA and T9ox. Following this step, the intermediate undergoes deacetylation at the C7 and C13 positions, catalyzed by T7dA1 and T13dA1 respectively, to ultimately form baccatin III [27]. In summary, although different research groups have drawn divergent conclusions regarding the catalytic sequence in the baccatin III biosynthetic pathway, recent studies on oxetane ring formation, the nuclear transport factor 2 (NTF2)-like protein (FoTO1), the discovery of new functional enzymes, and the construction of heterologous taxane biosynthetic pathways have collectively expanded our understanding of the functional diversity of paclitaxel P450 enzymes. Additionally, such observations offer robust proof that paclitaxel biosynthesis is rooted in a coordinated metabolic network rather than a traditional, strictly linear catalytic process.
Furthermore, within Taxus plants, once the cyclized taxa-4(5),11(12)-diene core is formed, it can be modified along multiple branching pathways through hydroxylation or acylation at other positions to generate diverse taxane compounds. A P450 monooxygenase, taxane-14β-hydroxylase (T14βOH), which functions in an alternative branch of the taxane biosynthetic pathway, was subsequently identified [29]. Studies have shown that the expression of T14βOH is negatively correlated with paclitaxel content, suggesting that multiple biosynthetic routes for taxanes may exist in Taxus, forming a complex synthetic network. After the formation of the tricyclic diterpene core, various enzymatic modifications lead to the production of diverse taxane compounds. Subsequent studies have shown that, in addition to the 19 reactions required to convert GGPP to paclitaxel, several branching pathways involving alternative hydroxylation and acylation reactions lead to the formation of nonproductive taxanes, which is a major cause of the accumulation of numerous intermediate byproducts [25].
2.3. Biosynthesis, Attachment, and Modification of the β-Phenylalanyl Side Chain
Initially, phenylalanine aminomutase (PAM) catalyzes the isomerization of α-phenylalanine to β-phenylalanine [30]. Subsequently, β-phenylalanyl-CoA ligase (PCL) activates this side chain to form β-phenylalanyl-CoA [31]. In 2023, efforts to characterize side-chain attachment successfully identified the native β-phenylalanyl-CoA ligase sequence utilizing a tobacco transient expression system [15]. To further optimize this catalytic step, subsequent comparative assays in 2025 evaluated three distinct CoA ligases: the native TBPCCL from Taxus baccata, AAE16 from Taxus chinensis, and an engineered fungal variant, CoAL(A312G), derived from Penicillium chrysogenum [16]. These functional comparisons demonstrated that CoAL(A312G) possesses significantly superior catalytic activity relative to both plant-derived enzymes. Previous studies have demonstrated that baccatin III:3-amino-3-phenylpropanoyltransferase (BAPT) catalyzes the acylation of baccatin III at the C13 position using β-phenylalanyl-CoA as the acyl donor, generating β-phenylalanyl baccatin III [32]. This intermediate is subsequently hydroxylated at the C2′position of the side chain by taxane-2′α-hydroxylase (T2′αOH) to yield 3′N-debenzoyltaxol. Finally, 3′-N-debenzoyl-2′-deoxytaxol N-benzoyltransferase (DBTNBT) catalyzes the benzoylation of the side-chain C3′ nitrogen atom, producing paclitaxel [32]. In 2025, Sotirios C. Kampranis’ team revised the final two key steps of side-chain biosynthesis in the paclitaxel pathway, specifically C2′α-hydroxylation and 3′-N-benzoylation. They identified taxoid-2′-oxoglutarate-dependent dioxygenase (T2′OGD) as the enzyme responsible for C2′ hydroxylation and taxoid-3′-N-benzoyltransferase (T3′NBT) as the enzyme responsible for 3′-N-benzoylation, which was different from the previously proposed roles of T2′αOH (TB506) and DBTNBT. They also investigated the sequential order from baccatin III to paclitaxel. First, the C13 hydroxyl group of baccatin III is conjugated with β-phenylalanyl-CoA by BAPT to form the intermediate N-debenzoyl-2′-deoxytaxol. Subsequently, T2′OGD hydroxylates the C2′ position of this intermediate, generating N-debenzoyltaxol. Finally, T3′NBT transfers a benzoyl group to the N3′ position of the side chain, yielding the final product paclitaxel. Moreover, the study revealed that the last two steps of paclitaxel biosynthesis do not necessarily require baccatin III as the substrate. Instead, 10-deacetylbaccatin III can also serve as a substrate for T2′OGD, undergoing C2′ hydroxylation and subsequent N3′ benzoylation, followed by DBAT-mediated acetylation to produce paclitaxel. These findings demonstrate that while the action of BAPT is not strictly fixed in order, the sequence of T2′OGD and T3′NBT is invariant, as N3′ benzoylation can only occur after C2′ hydroxylation has been completed [16].
Table 1. The identified paclitaxel biosynthetic enzymes and references.
|
Name |
Abbreviation |
Accession Number |
References |
|
|---|---|---|---|---|
|
1 |
Geranylgeranyl diphosphate synthase |
GGPPS |
AF081514 |
[33] |
|
2 |
Taxa-4(5),11(12)-diene synthase |
TS |
AY364469 |
[34] |
|
3 |
Taxa-4(5),11(12)-diene-5α-hydroxylase |
T5αOH |
AY289209 |
[35] |
|
4 |
Taxa-4(5),11(12)-diene-13α-hydroxylase |
T13αOH |
AY056019 |
[36] |
|
5 |
Taxa-4(5),11(12)-diene-5α-ol-O-acetyl transferase |
TAT |
AF190130 |
[37] |
|
6 |
Taxa-4(5),11(12)-diene-10β-hydroxylase |
T10βOH |
AF318211/ AY563635 |
|
|
7 |
Taxa-4(5),11(12)-diene-7β-hydroxylase |
T7βOH |
AY307951 |
[38] |
|
8 |
Taxa-4(5),11(12)-diene-2α-hydroxylase |
T2αOH |
AY518383 |
[39] |
|
9 |
Taxane-2α-O-benzoyl transferase |
TBT |
AF297618 |
[40] |
|
10 |
10-deacetylbaccatin III-10-O-acetyl transferase |
DBAT |
AF193765 |
[41] |
|
11 |
Phenylalanine aminomutase |
PAM |
AY582743 |
[30] |
|
12 |
β-phenylalanoyl-CoA ligase |
PCL |
KM593667 |
[31] |
|
13 |
Baccatin III:3-amino, 13-phenylpropanoyltransferase |
BAPT |
AY082804 |
[31] |
|
14 |
Taxane-2′α-hydroxylase |
T2′αOH |
KP178208 |
[32] |
|
15 |
N-benzoyl transferase |
DBTNBT |
AF466397 |
[42] |
|
16 |
Taxa-4(5),11(12)-diene-14β-hydroxylase |
T14βOH |
AY188177 |
[29] |
|
17 |
Cytochrome P450 reductase |
TcCPR |
AY571340 |
[29] |
|
18 |
C4β-C20 epoxidase |
TB328 |
[15] |
|
|
19 |
Taxane 1β-hydroxylase |
T1βOH |
[15] |
|
|
20 |
Taxane 9α-hydroxylase |
T9αOH |
[15] |
|
|
21 |
Taxane 9α-dioxygenase |
T9OX |
[15] |
|
|
22 |
Phenylalanine-CoA ligase |
PCL |
[15] |
|
|
23 |
Taxane oxetanase |
TOT |
[12] |
|
|
24 |
Taxane 9α-hydroxylase |
T9αOH1 |
[12] |
|
|
25 |
Taxane 9α-hydroxylase |
CYP725A37 |
PP197199/PP197200 |
[13] |
|
26 |
Taxane oxetanase |
CYP725A55 |
PP197201 |
[13] |
|
27 |
Acyltransferase |
AT5 |
PP197202 |
[13] |
|
28 |
CoA ligase |
CoAL(A312G) |
[16] |
|
|
29 |
Taxoid-2′-oxoglutarate-dependent dioxygenase |
T2′OGD |
PQ015324 |
[16] |
|
30 |
Taxoid-3′-N-benzoyltransferase |
T3′NBT |
PQ015327 |
[16] |
|
31 |
Taxane 9α-hydroxylase |
T9αOH-750C |
[11] |
|
|
32 |
Taxane 1β-hydroxylase |
T1βOH-184 |
[11] |
|
|
33 |
Taxane 1β-hydroxylase |
T1βOH-686 |
[11] |
|
|
34 |
Taxane 7β-O-acyltransferase |
T7AT |
[11] |
|
|
35 |
DeAc898 |
T9dA |
[11] |
|
|
36 |
DeAc1023 |
T7dA |
[11] |
|
|
37 |
Taxane C7β-O-deacetylase |
T7dA1 |
PX848829 |
[27] |
|
38 |
Taxane bifunctional deacetylase |
T79dA |
PX848830 |
[27] |
|
39 |
Taxane C13-O-deacetylase |
T13dA1 |
PX848827 |
[27] |
|
40 |
Taxane C13-O-deacetylase |
T13dA2 |
PX848828 |
[27] |
|
41 |
Taxoid acetyltransferase |
TAX19 |
AY628434 |
[28] |
3. Multi-Omics Approaches Facilitating the Evolutionary Origins Illustration of Paclitaxel Biosynthesis
In recent years, omics technologies, including genomics, transcriptomics, proteomics, and metabolomics, have been extensively applied to paclitaxel biosynthesis, functional gene discovery, transcription factor analysis, and the detection of associated metabolites. These approaches provide robust technical support for molecular biology studies and the metabolic pathway analysis of paclitaxel. Moreover, the generation of large-scale omics data and integrative multi-omics analyses has laid a solid foundation for investigating the biosynthesis and regulatory mechanisms of secondary metabolites in Taxus.
3.1. Applications of Genomic Analysis in Paclitaxel Biosynthesis
In recent years, three Chinese research teams have successively released the genomic data of Taxus wallichiana var. mairei [6], Himalayan yew (Taxus wallichiana) [7], and Taxus yunnanensis [8]. These studies have systematically investigated the evolutionary origins of the paclitaxel biosynthetic pathway and its key gene clusters, providing valuable insights into the formation and regulatory mechanisms of paclitaxel biosynthesis. Currently, the cloned hydroxylases involved in paclitaxel biosynthesis from Taxus species are classified into the CYP725A subfamily [7]. Through genome analysis of Taxus mairei, researchers revealed the genomic distribution and regulatory patterns of the cytochrome P450 family and discovered that paclitaxel biosynthetic genes are clustered on chromosome 9 of Taxus. Furthermore, by constructing a co-regulatory gene network, they identified 17 CYP725A genes, three transferases, and 10 transcription factors [6]. In addition, chromosome-level whole-genome sequencing of Taxus wallichiana (Himalayan yew) enabled the resolution of key gene clusters responsible for paclitaxel biosynthesis, leading to the identification of 31 candidate P450 genes. Within this cluster, the isoenzymes TS-2 and T5αOH-2, homologous to TS and T5αOH, were identified. Furthermore, the study demonstrated that tandem gene duplication is the primary driving force behind the evolution of gene families involved in paclitaxel biosynthesis in Taxus, providing essential genetic resources for the comprehensive elucidation of the paclitaxel biosynthetic pathway [7]. Genomic analysis of Taxus yunnanensis (Yunnan yew) revealed that the gene clusters involved in paclitaxel biosynthesis are predominantly located on chromosome 12 and that the hydroxylase gene families associated with paclitaxel biosynthesis have undergone significant expansion, thereby laying a solid foundation for understanding the formation of the paclitaxel biosynthetic pathway [8]. Relevant genomic studies will enhance our understanding of the paclitaxel biosynthetic pathway and its genetic basis, contributing to the resolution of key scientific questions in gymnosperm genome evolution. The candidate genes and transcription factors identified through these efforts provide valuable genetic resources and molecular targets for paclitaxel biosynthesis research, facilitating the design and application of heterologous paclitaxel production.
To investigate the conservation of the paclitaxel biosynthesis pathway within closely related Taxaceae species, Shen’s team generated a chromosome-level genome assembly of Pseudotaxus chienii and performed matrix-assisted laser desorption/ionization imaging mass spectrometry (MALDI-IMS) analysis. This approach yielded a series of spatially resolved metabolite maps comprising 9999 data points, revealing tissue-specific accumulation patterns of bioactive taxanes. Comparative genomic analysis across P. chienii, Taxus wallichiana, and Torreya grandis further indicated that disruption of TBT and TOT1 (CYP1) enzymatic steps may redirect metabolic flux toward taxusin or related analogs rather than paclitaxel. Notably, no taxa-4(5),11(12)-diene synthase (TS) homolog was identified in T. grandis, suggesting the absence of a functional taxane biosynthesis pathway in this species. Collectively, this genomic study of P. chienii provides important insights into the evolutionary origins and diversification of taxane biosynthesis [10].
3.2. Application of Transcriptome Analysis in Paclitaxel Biosynthesis
With the advancement of high-throughput sequencing technologies, a significant number of Taxus transcriptome datasets have been released. During 2024, a total of 1254 transcriptome datasets could be available in GenBank. Transcriptome-based analyses have identified several candidate genes and transcription factors involved in the paclitaxel biosynthesis pathway [43]. Moreover, comparative transcriptome analyses have been employed to investigate the tissue-specific distribution and differential accumulation of taxanes across various Taxus species [44]. Recent mining of these publicly available resources has uncovered several previously uncharacterized catalytic components, including C4β-C20 epoxidase, T9αOH, PCL [31], and T2′αOH [15]. In addition, using RNA-Seq technology, researchers have investigated the regulatory mechanisms of paclitaxel and other taxanes, identifying several transcription factors that modulate paclitaxel biosynthesis. Analyses and functional studies of transcription factor families, including ERF, bHLH, WRKY, MYC, and MYB, have identified regulators of paclitaxel biosynthetic gene expression, thereby contributing to increased paclitaxel accumulation in transgenic Taxus cell lines [45]. The discovery of these functional genes and transcription factors provides critical insights for advancing the genetic engineering of paclitaxel biosynthesis and elucidating the regulatory mechanisms attributing its production.
3.3. Application of Multi-Omics Approaches in Paclitaxel Biosynthesis
Integrative multi-omics analyses allow the capture of dynamic regulations in Taxus plants and the paclitaxel biosynthetic pathway from multiple perspectives, thus facilitating the exploration of the complex mechanisms underlying paclitaxel biosynthesis. Combined transcriptome and metabolome analyses help elucidate the regulatory networks of gene expression and metabolic pathways responsible for metabolite synthesis in Taxus, laying a foundation for delineation of taxane biosynthesis routes. Similarly, integrating proteomic and metabolomic data provides deeper insights into the mechanisms underlying gene expression regulation. Yu et al. systematically analyzed Taxus stem tissues using proteomic and metabolomic approaches. Metabolomic profiling revealed tissue-specific accumulation of metabolites in the stems, while proteomic analysis identified 10 key enzymes involved in paclitaxel biosynthesis. Additionally, they found that TmMYB3, specifically expressed in the phloem, regulates paclitaxel biosynthesis by activating the expression of TBT and TS within the pathway, highlighting its critical role in transcriptional regulation of paclitaxel production. The authors conducted a quantitative analysis of paclitaxel, 10-deacetylpaclitaxel, baccatin III, and 10-DAB in four major stem tissues using ultra-performance liquid chromatography-tandem mass spectrometry (UPLC-MS/MS). The results revealed that paclitaxel and 10-DAB predominantly accumulated in the phloem, baccatin III mainly in the pith, and 3′-N-debenzoyltaxol (DAP) primarily in the cortex and phloem, while all four metabolites exhibited the lowest levels in the xylem [46]. Recently, the same authors employed advanced mass spectrometry imaging techniques to generate spatial distribution maps of taxane compounds in young Taxus stems. Furthermore, the study revealed that T14βOH, T5αOH, and TS are predominantly expressed in endodermal cell populations, T10βOH and DBTNBT are primarily expressed in xylem parenchyma cells, and DBAT is expressed in epidermal cell populations. Through single-cell expression profiling, numerous cell type-specific transcription factors, including members of the MYB, TEM, RAV, and NAC families, were identified, and their downstream target genes were predicted. This work established a cell type-resolved expression atlas of young Taxus stems at single-cell resolution, providing a foundation for further studies on the cell-specific regulation of taxane biosynthesis [47]. Liang and colleagues, based on genomic and transcriptomic data from Taxus yunnanensis, Taxus wallichiana var. mairei, and related species, conducted gene co-expression analyses across multiple tissues (leaves, roots, stems, and arils) under various treatments, including methyl jasmonate induction. Through this approach, they successfully identified the two key enzymes catalyzing the final steps of paclitaxel biosynthesis: taxoid-2′-oxoglutarate-dependent dioxygenase (T2′OGD) and taxoid-3′-N-benzoyltransferase (T3′NBT) [16]. Recently, a strategy called “multiplexed perturbation × single nuclei” (mpXsn) was developed, in which 136 different treatments, including hormones, pathogens, and metabolic intermediates, were applied to Taxus tissues. Following the treatments, the tissues underwent single-nucleus RNA sequencing, enabling the construction of a highly accurate gene co-expression network. This network provided a clearer understanding of gene interactions and their roles in paclitaxel biosynthesis. A key strength of the mpXsn strategy is its ability to partition known paclitaxel biosynthesis genes into three distinct expression modules corresponding to the early, middle, and late stages of the biosynthetic pathway. From this, eight important new genes, including T9αOH-750C, T7AT, and T1βOH, were identified. In this study, mpXsn served as a powerful discovery tool, with metabolomics used to assess phenotypic outcomes and validate findings, while proteomics provided insights into the underlying mechanisms [11]. All results were interpreted within the larger genomic context. This multi-omics approach successfully addressed the challenges posed by the large and complex genome of Taxus, providing valuable insights into paclitaxel biosynthesis.
4. Advances in Microbial and Plant Chassis Engineering for the Biosynthesis of Paclitaxel
As a disruptive innovation in biotechnology, synthetic biology offers novel solutions to critical challenges in resource sustainability, environmental protection, and related fields. The biosynthesis of plant natural products has emerged as a research hotspot of synthetic biology. To date, intermediate pathways of paclitaxel and the heterologous production of several precursors have been successfully reconstructed in diverse chassis organisms, including Escherichia coli, Saccharomyces cerevisiae, Yarrowia lipolytica, tobacco cells, and endophytic fungi (Table 2).
4.1. Advances in Paclitaxel Biosynthesis Using Prokaryotic Chassis
Early studies directly overexpressed IDI, GGPPS, and TS in E. coli, achieving a taxa-4(5),11(12)-diene yield of approximately 0.5 mg/L [48]. Subsequent overexpression of the endogenous MEP pathway in E. coli further increased taxa-4(5),11(12)-diene production to 1.3 mg/L. A multivariate modular metabolic engineering strategy partitioned the paclitaxel biosynthetic pathway into two functional modules for systematic precursor enhancement: an upstream module containing four rate-limiting genes (dxs, idi, ispD, and ispF) for IPP production, and a downstream module containing two heterologous genes (GGPPS and TS) for taxa-4(5),11(12)-diene generation. By optimizing both the native MEP pathway and the taxa-4(5),11(12)-diene biosynthetic pathway, the researchers successfully overexpressed TS and T5αOH in E. coli, achieving an approximately 15,000-fold increase in taxa-4(5),11(12)-diene production, reaching 1 g/L-the highest reported yield to date. Simultaneously, taxa-4(5),11(12)-dien-5α-ol (T5α-ol) production increased 2400-fold, reaching 58 ± 3 mg/L [49], thereby laying the foundation for subsequent P450 mediated oxidation to downstream metabolites. The study also demonstrated that when exogenous indole concentrations exceeded approximately 100 mg/L, taxa-4(5),11(12)-diene synthesis was severely inhibited, and further increases in indole concentration suppressed cell growth, with the inhibitory effect being highly strain-dependent. The biochemical mechanism underlying the interaction between indole and the isoprenoid pathway remains unclear; however, the results suggest that indole may act synergistically with isoprenoid-derived terpenoids to inhibit cell growth. Subsequently, the same research group [50] combined the GGPPS and TS genes with E. coli native MEP pathway to investigate the effects of different promoters and cellular backgrounds on taxa-4(5),11(12)-diene production. They found that JM109 (DE3) produced approximately 2.5 times more taxa-4(5),11(12)-diene than the BL21 (DE3) strain. Metabolic engineering strategies incorporating the mevalonate (MVA) pathway alongside the TS gene into E. coli yielded a modest taxa-4(5),11(12)-diene output of only 11.3 ± 0.5 mg/L [8]. Prompted by these limitations in bacterial hosts, parallel studies shifted the production platform from bacteria to filamentous fungi. Initial optimizations in Alternaria alternata TPF6 evaluated the relative strengths of six heterologous promoters. The integration of these characterized promoters for the targeted overexpression of heterologous IDI, tHMGR (truncated 3-hydroxy-3-methylglutaryl-CoA reductase), and TS ultimately established a stable fungal biosynthesis system, achieving a taxa-4(5),11(12)-diene titer of 61.9 ± 6.3 μg/L [19].
Recent engineering efforts directed toward the de novo synthesis of paclitaxel precursors established a heterologous MVA pathway in E. coli. The researchers first screened key enzymes in the pathway, including TS and cytochrome P450 reductase (CPR), and optimized both the linker between T5αOH and CPR fusion proteins and the fermentation conditions. Ultimately, the oxidized taxane yield reached 27 mg/L, with T5α-ol production reaching 7 mg/L, representing a 12-fold and 23-fold increase, respectively. In a parallel approach, co-expression of GGPPS and TS in E. coli achieved a taxa-4(5),11(12)-diene yield of 93.5 mg/L in shake-flask cultures. Expansion of the microbial production platform to Bacillus subtilis involved overexpressing a plant-derived TS gene alongside a customized synthetic operon. This operon carried genes from B. subtilis’s native MEP pathway, as well as IspA (encoding geranyl-geranyl and farnesyl diphosphate synthase), which enabled the production of taxa-4(5),11(12)-diene in B. subtilis 168 [51]. Furthermore, by overexpressing the crtE gene (encoding GGPPS) to enhance GGPP supply, the strain achieved a substantial increase in taxa-4(5),11(12)-diene production. Compared with a strain expressing only TS and relying on B. subtilis’s native pathway, the overexpression of MEP pathway enzymes, IspA and GGPPS resulted in an 83-fold increase in taxa-4(5),11(12)-diene yield. The engineered strain produced a total taxa-4(5),11(12)-diene yield of 17.8 mg/L, demonstrating that B. subtilis can serve as a promising platform for paclitaxel production. Subsequently, taxa-4(5),11(12)-diene undergoes the first oxidation step catalyzed by T5αOH. However, in E. coli, the absence of an internal membrane system and the lipophilic N-terminal region of P450 reduce its solubility, making the successful expression of P450 enzymes in E. coli particularly challenging. N-terminal engineering of the P450 enzyme, coupled with the systematic optimization of its coupling with CPR, successfully facilitated high-level T5αOH expression in E. coli and achieved a hydroxylated taxane yield of 570 ± 45 mg/L [52]. Heterologous introduction of the DBAT gene into E. coli, supplemented by the feeding of exogenous 10-DAB, further enabled the successful semi-synthesis of the key intermediate baccatin III.
Due to the absence of efficient isoprenoid biosynthetic pathways, the use of E. coli and other prokaryotic hosts for the heterologous production of plant metabolites presents inherent limitations. Additionally, prokaryotic hosts often produce target proteins in insoluble and nonfunctional forms, and the supply of NADPH, a crucial cofactor for plant cytochrome P450 enzymes, is severely limited in prokaryotic systems. Furthermore, prokaryotes lack the compartmentalization found in eukaryotic cells, which creates distinct intracellular environments in eukaryotes, enabling the spatial and temporal partitioning of intermediates during final product formation. Consequently, unicellular eukaryotes, such as yeast, with their membrane-bound organelles and cellular compartmentalization, have attracted considerable attention for heterologous production [53].
4.2. Advances in Paclitaxel Biosynthesis Using Yeast Chassis
Microbial co-culture platforms integrating E. coli and S. cerevisiae successfully enabled the biosynthesis of oxygenated taxanes. In this system, E. coli was utilized for its rapid growth capacity and efficient taxa-4(5),11(12)-diene production, while S. cerevisiae was chosen for its abundant intracellular membrane structures that enable the functional expression of cytochrome P450 enzymes. However, ethanol produced by S. cerevisiae acts as a growth inhibitor for E. coli. To overcome this limitation, the researchers employed xylose as the carbon source, which can only be metabolized by E. coli. During xylose consumption, E. coli generates acetate, which can be utilized by S. cerevisiae without leading to ethanol production. Ultimately, this strategy resulted in the production of 33 mg/L of oxygenated taxanes. DeJong et al. successfully introduced eight genes involved in the paclitaxel biosynthetic pathway (GGPPS, TS, T5αOH, T10βOH, T13αOH, TAT, TBT, and DBAT) into S. cerevisiae using two plasmids. The expression of these genes was verified through both in vivo and in vitro experiments, and the production of GGPP and taxa-4(5),11(12)-diene was detected in the system. This work demonstrated the functional activity of fusion proteins in yeast and confirmed that the native isoprenoid precursors IPP and DMAPP are sufficient to initiate the terpenoid biosynthetic pathway. However, the yield of T5α-ol was very low. Engels et al. successfully introduced heterologous genes encoding isoprenoid biosynthetic enzymes along with regulatory factors that suppress competing pathways into S. cerevisiae. When yeast co-expressed HMGR, the mutated regulatory protein UPC2-1, GGPPS, and TS, the production of taxa-4(5),11(12)-diene reached 8.7 mg/L. Subsequently, Ding et al. identified the most catalytically efficient GGPPS variant by protein modeling and molecular docking, and co-expressed it with ERG20, tHMGR, and TS in S. cerevisiae, resulting in a taxa-4(5),11(12)-diene titer of 72.8 mg/L [54].
Integration of a tagged and multi-copy TS gene driven by the GAL1 promoter resulted in a 22-fold enhancement in taxa-4(5),11(12)-diene titer. Subsequent fermentation of this engineered platform produced 57 ± 3 mg/L at 30 °C, and the yield was further elevated to 129 mg/L by reducing the cultivation temperature to 20 °C [55]. Further genetic modifications to strain LRS5 involved the introduction of genes encoding T5αOH, a homologous CPR, and the TAT gene from Taxus cuspidata, yielding the advanced strain LRS6. Through optimization of fermentation conditions and pH control, the researchers achieved a 2.7-fold increase in oxygenated taxane production, reaching 78 mg/L [56]. Subsequent efforts to maximize T5αOH activity at both enzymatic and whole-cell levels in S. cerevisiae revealed that the uncoupling of T5αOH and CPR interactions potentially facilitates the formation of oxygenated taxanes. Through the construction of fusion proteins to enhance T5α-ol production, they achieved a maximum oxygenated taxane yield of approximately 361.4 ± 52.4 mg/L using 10 mL resting cell cultures, with T5α-ol reaching 38.1 ± 8.4 mg/L [57]. Integration of a push-pull strategy with the endogenous MVA pathway in yeast successfully maximized precursor supply while minimizing competitive by-pathway metabolism. Enhancing the soluble expression of TS via SUMO fusion, coupled with the targeted overexpression of tHMG1, GGS1, and TS, ultimately yielded a taxa-4(5),11(12)-diene titer of 101.4 mg/L during fed-batch fermentation. Walls and colleagues addressed nutrient limitation by enhancing culture nutrition, and under optimized conditions in a 1 L bioreactor, they achieved a maximum taxa-4(5),11(12)-diene titer of 71 ± 8 mg/L after 95 h of cultivation. The main products of T5αOH, iso-OCT, OCT, and T5α-ol-accumulated to 16 ± 3, 44 ± 3, and 42 ± 4 mg/L, respectively, while the TAT product T5α-Ac reached a maximum of 21 ± 0.3 mg/L, nearly sixfold higher than previously reported levels [56]. However, due to the poor expression and low catalytic activity of T5αOH in heterologous hosts, its primary products remained OCT and its isomer iso-OCT, with less than 10% of the taxa-4(5),11(12)-diene precursor ultimately converted to the target T5α-ol. As observed in E. coli, the high-level catalytic activity of P450 enzymes in yeast is kinetically dependent on efficient intracellular NADPH supply. Additionally, the formation of by-products and other endogenous metabolites in yeast interferes with the heterologous production of paclitaxel.
In 2024, the biosynthetic pathway for highly oxidized paclitaxel intermediates was reconstructed in S. cerevisiae by feeding T5α-ol. This work systematically elucidated the complete biosynthetic route from the C5 mono-oxygenated product of taxa-4(5),11(12)-diene to the highly oxidized 1β-dehydroxybaccatin VI, advancing a comprehensive understanding of paclitaxel biosynthesis. The researchers identified the paclitaxel 9α-hydroxylase responsible for the fourth hydroxylation step, as well as CYP725A55, which catalyzes oxetane ring formation via a cascade oxidation coupled with acyl rearrangement mechanism, and the acetyltransferase responsible for C7-OAc formation. Furthermore, the study highlighted the need to further clarify the complete biosynthetic pathway from 1β-dehydroxybaccatin VI to baccatin III, as well as the parallel biosynthetic pathways leading to paclitaxel, and emphasized the importance of confirming the necessity of taxane-1β-hydroxylase and taxane-9α-oxidase in pathway construction. This systematic work provides a solid foundation for fully elucidating paclitaxel biosynthesis and its heterologous production in yeast cells.
In 2025, the research group of Sotirios C. Kampranis employed a tobacco-based system to successfully identify and validate the catalytic activities of CoAL(A312G), T2′OGD, and T3′NBT [16]. Subsequently, they reconstructed the paclitaxel side-chain biosynthetic pathway in Saccharomyces cerevisiae. However, due to poor protein stability and solubility of BAPT in yeast cells, the intermediate N-debenzoyl-2′-deoxytaxol could not be detected. To address this limitation, the team applied Protein-Sol prediction, site-directed mutagenesis, and computational approaches to generate a novel BAPT variant (BAPTm). Introduction of BAPTm increased the yield of N-debenzoyl-2′-deoxytaxol by 27%. Furthermore, by fusing maltose-binding protein (MBP) at the N-terminus via a tetra-amino acid linker (IGGG), the solubility of BAPTm (designated MBPig3BAPTm) was further improved. Ultimately, the integration of CoAL(A312G), MBPig3BAPTm, T2′OGD, and T3′NBT into S. cerevisiae, together with supplementation of baccatin III and β-phenylalanine, enabled the successful detection of paclitaxel, achieving a final titer of 0.59 ± 0.03 μg/L.
4.3. Progress in Paclitaxel Biosynthesis Using Plant-Based Chassis Systems
Plants are capable of harnessing sunlight and atmospheric CO2 to produce secondary metabolites [58]. Leveraging the abundant endogenous IPP and DMAPP in plant cells, parts of the paclitaxel biosynthetic pathway have been successfully introduced into Arabidopsis thaliana [59]. Following overexpression of the TS gene under the constitutive CaMV 35S promoter, taxa-4(5),11(12)-diene was successfully detected in homozygous plants, demonstrating functional expression of the recombinant protein in A. thaliana. However, overexpression of the relevant genes adversely affected plant growth, including reduced hypocotyl length, leaf chlorosis, slowed growth, and delayed flowering. These effects were hypothesized to result either from the cytotoxicity of taxa-4(5),11(12)-diene or from TS expression interfering with the biosynthesis of other essential terpenoids, such as gibberellins or carotenoids, during development. To mitigate these effects, the TS promoter was replaced with a glucocorticoid-inducible promoter, which prevented GGPP flux from being directed into the taxa-4(5),11(12)-diene pathway during plant growth and development, thereby reducing the detrimental phenotypic impacts of TS expression. Nevertheless, taxa-4(5),11(12)-diene production still remained low.
Studies have shown that tobacco is also an excellent host for heterologous paclitaxel production. The TS and T5αOH genes were introduced into tobacco and achieved trichome-specific expression. However, the engineered plants did not produce the desired T5α-ol but instead accumulated the byproduct OCT [60]. Implementation of a chloroplast compartmentalization strategy in N. benthamiana spatially segregated TS, T5αOH, and CPR. This was combined with an optimized supply of isoprenoid precursors, resulting in the production of taxa-4(5),11(12)-diene and T5α-ol at 56.5 µg/g and 1.3 µg/g of fresh leaf tissue, respectively [61]. Parallel development of transplastomic tobacco lines initially resulted in minimal taxa-4(5),11(12)-diene accumulation when TS was co-expressed in both chloroplasts and the cytosol. Remarkably, when TS was expressed with an added chloroplast transit peptide, taxa-4(5),11(12)-diene levels in chloroplasts increased significantly, reaching 87.8 µg/g of cell dry weight. These results highlight the importance of transporting intermediates into chloroplasts and their subsequent post-translational modifications for achieving high-level taxa-4(5),11(12)-diene production [62].
Integration of transcriptomics, cell biology, metabolomics, and single-enzyme knockout experiments identified the minimal gene set required for heterologous paclitaxel biosynthesis. By transiently expressing a truncated plastid-targeted diterpene synthase (nsTXS) and geranylgeranyl diphosphate synthase (nsGGPS) in N. benthamiana, along with the cytosolic mevalonate (MVA) pathway’s rate-limiting enzyme 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), they established a functional biosynthetic pathway for taxa-4(5),11(12)-diene. This led to the production of approximately 100 µg of taxa-4(5),11(12)-diene per gram of fresh leaf tissue. By incorporating previously characterized enzymes along with newly identified catalysts—including a C4β-C20 epoxidase, taxane 1β-hydroxylase (T1βOH), taxane 9α-hydroxylase (T9αOH), taxane 9α-dioxygenase, and phenylalanyl-CoA ligase (PCL)-the team successfully synthesized the key intermediate baccatin III and reconstituted its conversion to paclitaxel in N. benthamiana [15]. In April 2025, Sotirios C. Kampranis’s team identified a novel acyl-CoA ligase, CoAL(A312G), which functions in coupling β-phenylalanoyl-CoA, an inherently unstable intermediate, with baccatin III via BAPT to yield N-debenzoyl-2′-deoxytaxol. Co-expression of CoAL(A312G) with BAPT and PAM significantly enhanced the biosynthetic efficiency, achieving an exogenous baccatin III conversion rate exceeding 60%. The study also pinpointed T2′OGD and T3′NBT as key enzymes responsible for the final steps of paclitaxel biosynthesis. When PAM, CoAL(A312G), BAPT, T2′OGD, and T3′NBT were co-expressed in tobacco leaves, paclitaxel accumulation reached 1.99 ± 0.19 μg per gram of fresh leaf weight. Notably, the C4β-C20 epoxidase, an α-ketoglutarate-dependent dioxygenase, overcame the first biosynthetic bottleneck in paclitaxel metabolic engineering. While functional expression was not achieved in E. coli or S. cerevisiae, the tobacco chassis facilitated its activity, enabling baccatin III production. These results define a minimal pathway for biosynthesis, suggesting a metabolic network rather than a strictly linear sequence.
Notably, the newly identified C4β-C20 epoxidase, an α-ketoglutarate-dependent dioxygenase, overcomes the first biosynthetic bottleneck in paclitaxel metabolic engineering. Cofactors or chaperone proteins within the tobacco chassis likely facilitated the functional expression of the C4β-C20 epoxidase, enabling the production of baccatin III in plant cells, whereas functional expression was not achieved in E. coli or yeast. Although the yields of 10-deacetylbaccatin III and baccatin III in this heterologous system are lower than those in the native paclitaxel-producing environment, these results define a complete, minimal pathway for their biosynthesis. Furthermore, the researchers observed that this pathway likely represents a metabolic network rather than a strictly linear sequence.
In January 2024, a streamlined baccatin III biosynthetic route was identified. By transiently expressing TOT and T9αOH-1 alongside seven other known biosynthetic genes (TXS, T5αOH, T13αOH, T2αOH, T7βOH, TAT, and TBT) in N. benthamiana, the researchers successfully confirmed baccatin III production. Their biochemical analyses showed that these core genes are co-regulated by the plant hormone jasmonate, with highly correlated expression patterns, indicative of tight synergistic control within the cell. Subcellular localization experiments demonstrated that GGPP is first converted to taxa-4(5),11(12)-diene in chloroplasts by TXS. Taxa-4(5),11(12)-diene is then transferred to the cytosol via plastid-endoplasmic reticulum (ER) contact sites, where six ER-anchored oxidases (T2αOH, T5αOH, T7βOH, T9αOH, T13αOH, and TOT) collaborate with two cytosolic acyltransferases (TAT and TBT) to produce baccatin III [12]. Systematic reconstruction of the early paclitaxel biosynthetic network recently enabled the structural elucidation of four novel T5αOH products. Chemical profiling reveals that a significant portion of these newly isolated compounds represent over-oxidized derivatives originating from the primary mono-oxygenated metabolites [26]. Their engineered system successfully reconstructed a six-step biosynthetic pathway for the production of 5α,10β-diacetoxy-taxa-4(5),11(12)-diene-13α-ol, which can be isolated. Additionally, the team updated the overall biosynthetic pathway for baccatin III, identifying the nuclear transport factor 2 (NTF2) domain-containing protein, FoTO1, as a cofactor that selectively increases taxa-4(5),11(12)-diene-5α-ol production to over 90%. By heterologously expressing seven newly identified genes (FoTO1, T9αOH-750C, T7AT, T1βOH-184, T1βOH-686, T9dA, T7dA) and nine previously known genes (TS, T5αOH, TAT, T10βOH, DBAT, T13αOH, T2αOH, TBT, TOT) in N. benthamiana, the team achieved the complete biosynthesis of baccatin III from GGPP, with yields ranging from 10 to 30 μg/g dry weight (DW), comparable to the natural content found in Taxus needles. This achievement fills several critical gaps in the paclitaxel biosynthesis pathway and lays the groundwork for the sustainable production of paclitaxel. Recent studies on the baccatin III biosynthetic pathway have identified taxusin and baccatin VI as key intermediates. The discovery of the enzyme T13dA, alongside other deacetylases, has clarified the metabolic network leading to baccatin III. The integration of a C13α-O-acetylation-deacetylation module has facilitated the de novo production of taxanes in plant hosts. By introducing the genes T13dA1, T79dA, and T7dA1, researchers reconstructed 18-gene and 19-gene baccatin III biosynthetic pathways in Nicotiana benthamiana. With the utilization of the C9-hydroxylase T9αH-750C, the engineered 19-gene pathway produced a baccatin III yield of 23 µg/g dry weight (DW). This yield is comparable to that of the previously established 17-gene pathway. However, unlike the 17-gene route that lacks the C13-deacetylation module, the 18-gene and 19-gene pathways emulate the endogenous metabolic network of Taxus. The inclusion of the acetylation-deacetylation mechanism directs the metabolic flux through the intermediates taxusin and baccatin VI. This pathway configuration circumvents potential catalytic bottlenecks associated with simplified linear routes. Consequently, these reconstructed pathways validate the biological function of the acetylation-deacetylation cycle and provide a functional framework for the future metabolic engineering of taxanes in plant chassis [27].
Meanwhile, significant advancements have been made in the development of transformation systems for Taxus species. Recently, the team led by Mercedes Bonfill reported the establishment of distinct transformed lines of Taxus baccata, mediated by Rhizobium rhizogenes A4. Although transgenic Taxus roots exhibit relatively slow growth, a dedifferentiation strategy was employed to obtain callus cultures, followed by the establishment of suspension cell systems. To enhance paclitaxel production, two key limiting genes, BAPT and DBTNBT, were overexpressed in a two-stage culture system under dual elicitation treatments. Compared to wild-type lines, paclitaxel content in DBTNBT-overexpressing lines increased more than fourfold, while BAPT-overexpressing lines exhibited a twofold increase. This marks the first successful transformation of Taxus seedlings via direct inoculation with two engineered R. rhizogenes A4 strains carrying paclitaxel biosynthetic genes. Earlier protocols utilizing the wild-type R. rhizogenes A4 strain successfully induced hairy root formation in Taxus through a strictly in vitro infection process [63]. Transcriptional analysis of the paclitaxel biosynthetic genes further revealed that GGPPS, TS, and DBAT were the most responsive to DBTNBT overexpression and dual elicitation, with their expression exhibiting a gradual and sustained increase.
Table 2. Heterologous expression systems for paclitaxel.
|
Synthetic Systems |
Products |
Yield |
Research Methods |
Publication Date |
References |
|---|---|---|---|---|---|
|
Escherichia coli |
Taxa-4(5),11(12)-diene |
1.3 mg/L |
Overexpression of IDI, GGPPS, TS, and DXP |
2001 |
[48] |
|
Escherichia coli |
Taxa-4(5),11(12)-diene |
1 g/L |
Multivariate modular metabolic engineering with overexpression of TS and T5αOH |
2010 |
[64] |
|
Escherichia coli |
Taxa-4(5),11(12)-diene-5α-ol |
58 ± 3 mg/L |
|||
|
Escherichia coli |
Taxa-4(5),11(12)-diene |
4.5 mg/g DW |
JM109(DE3) strain, 22 °C |
2012 |
[65] |
|
CladosporiumTPF6 |
Taxa-4(5),11(12)-diene |
61.9 ± 6.3 μg/L |
alcA promoter, overexpression of IDI and tHMGR |
2017 |
[19] |
|
Escherichia coli |
Oxidized taxanes |
27 mg/L |
TbrTS, Taxus CPR, T5αOH-GSTGS-CPR, introduction of a heterologous MVA pathway |
2022 |
[66] |
|
Escherichia coli |
Taxa-4(5),11(12)-diene-5α-ol |
7 mg/L |
|||
|
Escherichia coli |
Taxa-4(5),11(12)-diene |
93.5 mg/L |
Fusion expression of GGPP and TS |
2022 |
[67] |
|
Escherichia coli |
Oxidized taxanes |
570 ± 45 mg/L |
N-terminal modification of P450 enzyme |
2016 |
[52] |
|
Bacillus subtilis |
Taxa-4(5),11(12)-diene |
17.8 mg/L |
Overexpression of the MEP pathway, GGPPS, and TS |
2019 |
[51] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene |
8.7 mg/L |
Co-expression of tHMGR, the mutant regulatory protein UPC2-1, GGPPS, and TS |
2008 |
[68] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene |
72.8 mg/L |
YSG50 strain, GGPPSbc |
2014 |
[54] |
|
Saccharomyces cerevisiae |
Oxidized taxanes |
33 mg/L |
Co-culture of Escherichia coli and Saccharomyces cerevisiae |
2015 |
[49] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene |
129 mg/L |
Multi-copy TS with an MBP tag, 20 °C |
2020 |
[55] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-ol |
20 mg/L |
Fermentation process optimization, pH adjustment |
2020 |
[56] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-yl acetate |
3.7 mg/L |
|||
|
Saccharomyces cerevisiae |
Oxidized taxanes |
78 mg/L |
|||
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-ol |
42 mg/L |
2×YP medium, statistically determined screening design |
2022 |
[69] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-yl acetate |
22 mg/L |
|||
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-ol |
38.1 ± 8.4 mg/L |
Promoter pHXT7, fusion expression of T5OH and CPR, assayed in resting cells under neutral pH conditions |
2022 |
[57] |
|
Saccharomyces cerevisiae |
Oxidized taxanes |
361.4 ± 52.4 mg/L |
|||
|
Yarrowia lipolytica |
Taxa-4(5),11(12)-diene |
101.4 mg/L |
Fusion expression of SUMO with TS, with overexpression of tHMG1, GGS1, and TS |
2023 |
[70] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene |
215 mg/L |
Computational metabolic engineering |
2023 |
[71] |
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-ol |
43.65 mg/L |
|||
|
Saccharomyces cerevisiae |
Taxa-4(5),11(12)-diene-5α-yl acetate |
26.2 mg/L |
|||
|
Saccharomyces cerevisiae |
1β-dehydroxybaccatin VI |
-- |
Co-expression of 12 identified genes in cells with feeding of taxadien-5α-ol |
2024 |
[13] |
|
Arabidopsis thaliana |
Taxa-4(5),11(12)-diene |
600 ng/g DW |
Glucocorticoid-inducible expression of TS |
2004 |
[59] |
|
Nicotiana benthamiana |
Taxa-4(5),11(12)-diene |
50 μg/g DW |
Expression of TS with MeJA induction and silencing of PSY and PDS |
2014 |
[72] |
|
Nicotiana benthamiana |
Taxa-4(5),11(12)-diene |
56.6 ± 3.2 µg/g fresh weight |
Compartmentalization strategy with co-expression of DXS and GGPPS |
2019 |
[61] |
|
Nicotiana benthamiana |
Taxadien-5α-ol |
1.3 ± 0.5 µg/g fresh weight |
|||
|
Nicotiana tabacum cv. Xanthi |
Taxa-4(5),11(12)-diene |
87.8 μg/g DW |
N-terminal chloroplast transit peptide fusion of TS |
2021 |
[62] |
|
Nicotiana benthamiana |
Baccatin III |
154.87 ng/g fresh weight |
Transient co-expression of C4β-C20 epoxidase, T9αOH, T1βOH, and T9OX along with nine other known genes |
2023 |
[15] |
|
Nicotiana benthamiana |
Paclitaxel |
64.29 ng/g fresh weight |
Transient co-expression of PCL with BAPT, PAM, DBTNBT, and T2′OH |
2023 |
[15] |
|
Nicotiana benthamiana |
Taxa-4(5),11(12)-diene-5α-ol |
-- |
Attenuation of T5αOH expression using the weak constitutive NOS promoter |
2024 |
[26] |
|
Nicotiana benthamiana |
5α,10β-Diacetoxy-taxa-4(20),11-diene-13α-ol |
42 µg/g DW |
(NOS)T10βOH, DBAT and (NOS)T13αOH |
2024 |
[26] |
|
Nicotiana benthamiana |
Baccatin III |
50 ng/g DW |
Transient co-expression of TOT and T9αOH-1 along with seven other known biosynthetic genes |
2024 |
[12] |
|
Nicotiana benthamiana |
Baccatin III |
10–30 μg/g DW |
Transient expression of 7 newly discovered genes and 9 additional previously known genes. |
2025 |
[73] |
|
Nicotiana benthamiana |
Baccatin III |
23 μg/g DW |
Transient co-expression of 18-gene/19-gene pathway including C13-acetylation-deacetylation module |
2026 |
[27] |
4.4. Research Progress on Paclitaxel Biosynthesis by Taxus-Associated Endophytic Fungi
Additionally, it has been discovered that certain endophytic fungi associated with Taxus species are capable of producing paclitaxel. Initial screening and mutagenic optimization of one such isolate successfully elevated its native paclitaxel titer from 125.7 μg/L to 314.07 μg/L. Combinatorial treatments utilizing physical and chemical mutagenesis generated alternative strains producing 448.52 μg/L paclitaxel, with subsequent protoplast fusion events further improving the yield to 468.62 μg/L. Wang et al. [74] were the first to apply the REMI method to obtain a paclitaxel-producing fungal strain (BT2) with hygromycin B resistance. Similarly, genomic integration of a hygromycin B phosphotransferase gene, strictly driven by the trpC promoter, achieved protoplast regeneration frequencies exceeding 6% in targeted endophytes. These approaches have successfully facilitated the transfer of paclitaxel biosynthetic genes into engineered fungi to enhance paclitaxel production. Collectively, these genetic manipulation strategies facilitated the preliminary transfer of selective markers and vector elements into fungal platforms. Agrobacterium-mediated transformation of Cladosporium cladosporioides subsequently yielded the MD2 strain, a process strictly reliant on optimized co-culture conditions for stable integration. Liu et al. developed a highly efficient Agrobacterium-mediated transformation method for the paclitaxel-producing endophytic fungus Ozonium sp. EFY21 and optimized several factors affecting transformation efficiency and transformant stability. However, none of these studies have integrated the biosynthetic genes for paclitaxel into the genomes of the transformed fungi. Targeted genetic editing, such as the CRISPR/Cas9 system, was used to disrupt sterol metabolism in filamentous fungi by knocking out both squalene synthase and cycloartenol synthase. This concurrent knockout successfully redirected metabolic flux, thereby enhancing paclitaxel accumulation [75].
A comparative assessment of paclitaxel yields across different endophytic fungal strains reveals substantial variability, with the highest reported yield of 468.62 μg/L still orders of magnitude lower than that required for industrial applications. Several key bottlenecks limit the commercial application of this approach, including: (1) strain instability, with paclitaxel production often declining after repeated subculturing; (2) intrinsically low yields that remain insufficient for cost-effective production; and (3) challenges in fermentation scalability, as endophytic fungi typically exhibit slow growth and complex nutritional requirements; (4) random mutagenesis and protoplast fusion offer moderate yield improvements but lack predictability and sustainability. Agrobacterium-mediated transformation has not yet achieved stable integration of complete paclitaxel pathways. CRISPR/Cas9-based metabolic reprogramming is promising but remains at an early stage. Overall, current research on Taxus endophytic fungi remains relatively limited. Compared to E. coli, S. cerevisiae, and tobacco chassis, the genetic manipulation and cultivation systems for Taxus-derived endophytic fungi remain underdeveloped. With the continued advancement of omics-driven data mining, synthetic biology, and analytical technologies, future studies on endophytic fungi are expected to provide further insights into the biosynthesis of paclitaxel. However, given the current yield and stability constraints, endophytic fungal platforms are more likely to serve as valuable models for understanding paclitaxel biosynthesis rather than as immediate industrial production hosts.
4.5. Engineering Cyanobacteria to Produce Paclitaxel Precursors from CO2
Algae have emerged as advanced cellular factories, demonstrating substantial progress in the production of high-value compounds such as biofuels, pharmaceuticals, and nutraceuticals. Their rapid growth, adaptability to diverse environments, and inherent capacity for photosynthetic carbon fixation position them as promising platforms for sustainable biotechnological applications. Advances in genetic engineering and metabolic optimization have further enabled the tailored design of algal systems for efficient synthesis of complex molecules, providing environmentally sustainable alternatives to conventional manufacturing processes.
Although synthetic biology has facilitated the de novo synthesis of paclitaxel precursors in various microbial hosts, the complete biosynthesis of paclitaxel remains challenging due to limited oxidation efficiency in heterotrophic microorganisms. Ma’s team engineered Synechocystis sp. PCC 6803 with modular metabolic pathways comprising enzymes from the MEP pathway and the paclitaxel biosynthetic cascade to produce T5α-ol. The optimized strain DIGT-P560 achieved production titers of up to 17.43 mg/L of oxygenated taxanes and 4.32 mg/L of T5α-ol. Transcriptomic analysis of DIGT-P560 further indicated that establishing a flux toward oxygenated taxanes may enhance photosynthetic electron transfer efficiency and central metabolism in the engineered strain, thereby alleviating metabolic disturbances caused by the introduction of exogenous genes. This work represents the first demonstration of photosynthetic production of taxa-4(5),11(12)-diene-5α-ol from CO2 in cyanobacteria, underscoring the potential of engineered cyanobacteria as biosolar cell factories for high-value terpenoid production and offering new avenues for rational strain engineering and optimization [76].
5. Advances in the Engineering and Catalytic Mechanisms of Key Enzymes Involved in Paclitaxel Biosynthesis
In the early stages of paclitaxel biosynthesis, although taxa-4(5),11(12)-diene synthase (TS) exhibits low catalytic efficiency and produces multiple byproducts [77], it is not considered the primary rate-limiting step in the pathway. Instead, the hydroxylation of taxa-4(5),11(12)-diene at the C5 position, catalyzed by taxa-4(5),11(12)-diene-5α-hydroxylase (T5αOH), has been identified as the critical bottleneck and principal rate-limiting step in paclitaxel biosynthesis. Directed evolution, a widely applied enzyme engineering strategy, emulates the principles of genetic mutation and natural selection, introducing iterative mutations and screening cycles within comparatively short timeframes. Despite notable progress, the engineering of T5αOH and other cytochrome P450 enzymes in the paclitaxel pathway through directed evolution remains highly challenging, primarily owing to limited knowledge of their catalytic mechanisms and the absence of efficient high-throughput screening strategies.
Rational and semi-rational enzyme engineering strategies, which depend on sequence, structural, and functional information, enhance the likelihood of beneficial mutations and significantly reduce experimental workload. Amino acid substitutions within the catalytic active site can enhance enzymatic activity; however, achieving such improvements requires precise conformational control to sustain a delicate balance between thermodynamic flexibility and structural rigidity [78]. The integration of computational approaches with experimental validation has proven highly valuable for elucidating catalytic mechanisms and guiding rational or semi-rational protein engineering, particularly in improving substrate recognition and catalytic efficiency of P450 enzymes. Recently, substantial breakthroughs in pathway design within plant hosts have provided promising solutions to the persistent challenges associated with the chemical catalysis by P450 enzymes in paclitaxel biosynthesis [53].
5.1. Engineering of the Key Enzyme TS
TS consists of 862 amino acid residues, including an N-terminal plastid-targeting signal of approximately 80 residues and a catalytic active site structured by three helical regions [79]. The C-terminal domain (S553–V862) corresponds to a class I terpene cyclase, whereas the N-terminal domain (M107–I135) and the “insertion” domain (S349–Q552) constitute a degenerate class II terpene cyclase [80]. Class I terpene cyclase employs a trinuclear metal cluster that coordinates the conserved motifs DDXXD and (N,D)DXX(S,T)XXXE, thereby facilitating diphosphate ionization of isoprenoid substrates to generate carbocation intermediates and initiate catalysis.
In contrast, class II terpene cyclase utilizes aspartate residues within the DXDD motif to catalyze protonation of isoprenoid double bonds or oxirane moieties, thereby initiating carbocation formation. Despite differences in protein folding and substrate activation mechanisms between the two cyclase classes, the structural architecture of TS suggests an evolutionary relationship in terpene biosynthesis [79]. The catalytic activity of TS is mediated primarily by its C-terminal domain. Initially, a cluster of three Mg2+ ions binds GGPP and promotes diphosphate ionization. The subsequent diphosphate hydrolysis generates a reactive carbocation intermediate, while the enzyme’s active-site pocket shifts conformationally from an open to a closed state. This conformational transition enables the carbocation to undergo a cascade of rearrangements and cyclization reactions, ultimately yielding multiple taxa-4(5),11(12)-diene isomers [81].
The reaction pathway involving transient intermediates has increasingly emerged as a focal point of TS research. Kinetic and thermodynamic computational analyses have revealed that carbocation rearrangements catalyzed by TS are strongly influenced by the enzyme’s microenvironment and intrinsic conformational dynamics. Factors such as strong electrostatic interactions dependent on cation positioning, orientation, and conformational shifts, the spatial architecture of the active site, and the presence of water molecules in the catalytic pocket collectively modulate the reaction energy barrier, resulting in distinct product configurations [81]. A notable feature of TS is its active-site cavity, which is considerably larger than the final product, taxa-4(5),11(12)-diene. This spatial redundancy is considered a primary factor contributing to the enzyme’s product promiscuity. In parallel, extensive mutagenesis studies have identified additional structural determinants critical to catalysis in TS. Beyond the previously identified Y688 residue, tryptophan at position 753 plays a pivotal role in stabilizing carbocation intermediates via π–cation interactions. Remarkably, substitution of this residue with histidine results in a complete shift of the product profile, converting taxa-4(5),11(12)-diene into the alternative diterpene cembrene A [82,83].
TS is a key enzyme in paclitaxel biosynthesis, directing metabolic flux into the taxane pathway through cyclization activity. During fermentation, TS cyclizes geranylgeranyl pyrophosphate (GGPP) to yield 93.2% taxa-4(5),11(12)-diene, with minor byproducts of 4.7% taxa-4(20),11(12)-diene and additional diterpene scaffolds. Subsequently, only 10% of taxa-4(5),11(12)-diene is converted by taxa-4(5),11(12)-diene-5α-hydroxylase (T5αOH) into the target product, taxa-4(5),11(12)-diene-4(20),11(12)-5α-ol [22]. Evidence suggests that T5αOH has a higher substrate affinity for taxa-4(20),11(12)-diene than for taxa-4(5),11(12)-diene. Targeted mutagenesis near the TS active site revealed that substitution of tyrosine 688 with leucine TS (Y688L) significantly enhanced production of taxa 4(20),11(12) diene. When the TS (Y688L) variant was introduced into a T5αOH-expressing strain, taxa-4(5),11(12)-diene-5α-ol yield increased 2.4-fold. This study indicates that engineering TS to favor taxa-4(20),11(12)-diene formation can channel substrates directly into T5αOH-mediated P450 hydroxylation, bypassing the epoxide cleavage pathway and markedly increasing the proportion of the target product [22]. With the elucidation of the TS crystal structure [79], systematic mutagenesis of key residues has been performed. Based on structural modeling, several TS variants were engineered, including S713T, W753H, W753H/C830A, V584K/L/M, S587A, V714A/I, C803A/S, and Y841F. All variants retained catalytic activity, with S713T preserving 97.4% of wild-type enzymatic activity [84].
5.2. Advances in the Study of the Catalytic Mechanism of Taxa-4(5),11(12)-Diene-5α-Hydroxylase (T5αOH)
Before the foundation of FoTO1, investigations into the catalytic mechanism of T5αOH have mainly focused on two proposed pathways: the radical-rebound mechanism and the epoxidation mechanism [82,83]. Yadav et al. [85] suggested that competitive proton abstraction by the ferryl-oxo species at the C3, C13, and C20 positions contributes to the diversity of T5αOH oxidation products. Edgar and colleagues [22,23] hypothesized that taxa-4(5),11(12)-diene is initially converted by T5αOH into an unstable epoxide intermediate, which subsequently undergoes non-specific cleavage to yield various oxidized products, including taxa-4(5),11(12)-diene-5α-ol, in accordance with the epoxidation mechanism. Meanwhile, the isomeric form taxa-4(20),11(12)-diene bypasses the epoxide cleavage step and produces the target product taxa-4(5),11(12)-diene-5α-ol directly via the canonical P450 radical-rebound mechanism. Barton et al. further demonstrated that taxa-4(5),11(12)-diene-4(5)-epoxide is acid-sensitive and undergoes multiple rearrangements under varying chemical conditions, particularly in the presence of iron-porphyrin, thereby highlighting challenges to its use as a viable intermediate [83]. Since the initial discovery of T5αOH in Taxus in 1996, numerous strategies have been implemented, including N-terminal engineering, optimization of the electron transfer chain, ribosomal switch regulation, resting-cell cultivation, endoplasmic reticulum remodeling, and subcellular compartmentalization [53]. In parallel, extensive optimization of catalytic conditions, including temperature, O2 concentration, pH, and cofactors, has also been performed [56,60]. However, significant breakthroughs in the molecular-level engineering of T5αOH are still lacking. Yadav et al. developed a homology model of T5αOH based on six P450 structures and designed 53 variants. Among these, six mutants showed enhanced performance in either conversion efficiency or T5α-ol selectivity. Notably, the K131R mutant increased conversion at the expense of selectivity, whereas the V374L mutant enhanced selectivity but reduced conversion. Building on in-depth analyses of the paclitaxel biosynthetic gene cluster, our research group conducted large-scale functional screening of candidate sequences from the CYP725A subfamily of Himalayan yew. Two homologs, T5αOH-1 and T5αOH-2, were identified and experimentally confirmed to catalyze C5 hydroxylation of taxa-4(5),11(12)-diene. Nevertheless, their catalytic performance did not exceed that of previously reported T5αOH [7].
In 2024, our research group, in collaboration with Binju Wang’s group and Zhenzhan Chang’s group, employed X-ray crystallography, molecular dynamics (MD) simulations, quantum mechanics/molecular mechanics (QM/MM) calculations, and quantum mechanics (QM) computations to elucidate the crystal structure of the CYP725A4-taxa-4(5),11(12)-diene complex. Through comprehensive computational analyses, the catalytic mechanism of CYP725A4 in the native paclitaxel biosynthesis pathway was elucidated [86]. The results indicate that oxidation of taxa-4(5),11(12)-diene generates a zwitterionic intermediate, which can proceed along two competing transformation routes. In one pathway, an epoxide is formed and subsequently undergoes a water-mediated rearrangement to yield the target product, taxa-4(5),11(12)-diene-5α-ol. In the alternative pathway, protonation of the oxygen atom in the zwitterionic intermediate facilitates hydrogen shifts and carbon–oxygen coupling, leading to the formation of the side products OCT/iso-OCT. In contrast, taxa-4(20),11(12)-diene undergoes direct C5 hydroxylation, yielding taxa-4(5),11(12)-diene-5α-ol without formation of the epoxide intermediate. These crystallographic and computational studies provide valuable insights into the catalytic mechanism of CYP725A4 and lay the groundwork for its further engineering to enhance performance in paclitaxel biosynthesis.
5.3. Enzyme Engineering of T10βOH
In a recent study, the structural model of the N-terminally truncated enzyme t20T10βOH was predicted using AlphaFold2. The model features an N-terminal transmembrane domain connected to a catalytic core comprising 12 α-helices and antiparallel β-sheets, with the heme cofactor positioned between helices I and L. Building on this structure, the PROSS tool was employed to predict 45 single-point mutations with the potential to improve protein folding, each of which was subsequently evaluated energetically using FoldX. Experimental validation identified three beneficial mutations (I75F, L226K, and S345V) which enhanced substrate conversion by 2.1-, 1.4-, and 1.4-fold, respectively, compared to the wild-type enzyme. Mechanistic analysis suggested that I75F increases hydrophobicity at the substrate channel entrance to facilitate substrate access, L226K modulates enzyme conformation through altered surface charge distribution, and S345V stabilizes the protein core via enhanced hydrophobic interactions. Further combination of these mutations yielded double and triple mutants, among which the triple mutant I75F/L226K/S345V achieved the highest substrate conversion (2.8-fold increase) and exhibited a 9.5-fold elevation in cytochrome P450 T10βOH heterologous expression. This study provides a valuable engineering framework for enhancing the de novo biosynthesis of T10βOH and related taxanes [87].
5.4. Designing an Oxidase Toolbox for Site-Directed Oxidation of Taxanes
Although the biosynthetic pathway of paclitaxel has been progressively elucidated, its overall efficiency remains constrained by the limited activity and product selectivity of the native hydroxylases involved. To address this limitation, the researchers focused on identifying non-native oxidases capable of functionalizing taxa-4(5),11(12)-diene as alternatives to cytochrome P450 enzymes. Among 29 screened fungal oxidases, TteUPO was selected due to its superior activity toward taxa-4(5),11(12)-diene and high protein expression level. Based on this candidate, an artificial enzyme toolbox was developed using engineered variants of TteUPO, enabling site-specific oxidation of taxanes at multiple positions (C-4, C-6, C-10, C-11, C-12, and C-13). A collection of TteUPO mutants with distinct regioselectivities toward the taxa-4(5),11(12)-diene skeleton was constructed and utilized for pathway engineering. Moreover, by integrating the taxa-4(5),11(12)-diene biosynthetic pathway and enabling heterologous expression of a selected TteUPO mutant in E. coli, the de novo biosynthesis of oxidized taxa-4(5),11(12)-diene derivatives was achieved in this engineered chassis. Together, these findings provide an innovative strategy to establish efficient synthetic routes toward paclitaxel and to generate structurally diverse analogs for medicinal evaluation, thereby establishing a versatile platform for developing artificial synthetic pathways to paclitaxel and related taxanes [17].
5.5. Advances in the Mechanistic Study of Oxetane Ring Formation
Recent breakthroughs have clarified the mechanism of oxetane ring formation. Unlike other candidate enzymes implicated in oxetane formation, TB328 is not a P450 enzyme but rather an α-ketoglutarate/Fe(II)-dependent dioxygenase (ODD). Through transient expression in N. benthamiana, the researchers demonstrated that TB328 catalyzes the conversion of taxusin into taxusin-4β,20-epoxide, thereby enabling the de novo biosynthesis of baccatin III in the tobacco chassis. However, attempts to express TB328 in E. coli or S. cerevisiae strains engineered for taxa-4(5),11(12)-diene production were not successful. Furthermore, the detailed reaction mechanism remains unresolved because the low yield of the oxidized product prevents reliable NMR characterization [15].
In 2023, the research team led by Sotirios C. Kampranis published a study entitled challenged prevailing hypotheses that the C5 position of taxa-4(5),11(12)-diene-5α-ol is first acetylated and then oxidized through multiple steps to form the oxetane ring. The researchers observed taxane compounds in Taxus species that contained oxetane rings without prior acetylation, which led them to propose a “cyclization-first, acetylation-later” mechanism [23]. In their study, candidate genes were screened through genomic and transcriptomic analyses, and validation was conducted using two heterologous expression systems (S. cerevisiae and N. benthamiana) in combination with structural characterization of the resulting products. They ultimately identified CYP725A4 (also known as T5αOH) as the enzyme responsible for oxetane ring formation. The authors discovered that CYP725A4 catalyzes oxidation at multiple positions on the taxa-4(5),11(12)-diene precursor, potentially forming the oxetane ring through a two-step epoxidation mechanism, thereby revising the previous understanding of its oxidative products and catalytic mechanism. Moreover, the study demonstrated that both taxa-4(5),11(12)-diene and taxa-4(20),11(12)-diene serve as starting substrates for the generation of the target epoxidized intermediate, suggesting the presence of multiple branched biosynthetic pathways leading to paclitaxel [23].
In 2024, P450 enzyme TOT1 was found to play a key role in the formation of the oxetane ring. It did not catalyze the direct conversion of a three-membered ring intermediate into a four-membered ring [12]. Instead, the enzyme simultaneously converts the double-bond structure into both three- and four-membered ring systems. Using density functional theory (DFT) calculations in combination with substrate selectivity assays, the study demonstrated that both epoxide and oxetane products arise from a carbocation intermediate within TOT1. However, oxetane formation was shown to be energetically and kinetically more favorable, providing a molecular explanation for its predominance and indicating that the epoxide is not a necessary intermediate in this process [12]. Almost concurrently, the catalytic mechanism of TOT1’s isoenzyme, CYP725A55, for the same reaction was elucidated by Zhihua Zhou’s team and Jungui Dai’s team. Isotope labeling experiments confirmed that the oxetane ring is likely formed through a direct oxidation-acyl rearrangement process, consistent with the predictions of Yan’s team [13]. These findings overturn three decades of conventional understanding of this structural motif and fill a critical gap in elucidating the molecular mechanism of ring-expansion reactions that generate oxygen-containing four-membered rings, a process previously observed only in plants.
5.6. Engineering of DBAT in the Paclitaxel Biosynthetic Pathway
In recent years, owing to the challenges in modifying the substrate specificity and enhancing the catalytic activity of T5αOH, increasing attention has been directed toward engineering DBAT, a downstream enzyme in the paclitaxel biosynthetic pathway. Alanine scanning was employed to identify the active site of DBAT, leading to the design of a double mutant (G38R/F301V) that exhibited a sixfold increase in catalytic efficiency relative to the wild type. A single-point mutant (G38R) was found to exhibit a 2.15-fold improvement in catalytic efficiency [38,88]. Molecular docking analysis further revealed the critical role of His162 in DBAT catalysis, and alanine scanning indicated that substitution of four residues (H162, R363, G361, and I164) with alanine completely abolished enzymatic activity. Application of a semi-rational design strategy generated a double mutant (I43S/D390R) that not only enhanced catalytic efficiency but also facilitated more effective utilization of vinyl acetate, a more economical acyl donor than acetyl-CoA [89].
5.7. Engineering of Key Enzymes for the Conversion of Paclitaxel Natural Analogues
At present, paclitaxel obtained from nursery-cultivated Taxus species remains one of the primary sources for clinical use. However, its natural abundance is extremely low, ranging from 0 to 0.069%, with the highest concentrations occurring in the bark. By contrast, the content of its natural analog, 7-β-xylosyl-10-deacetylpaclitaxel (XDT), is generally much higher than that of paclitaxel. Typically, removal of the xylosyl moiety from XDT yields 10-deacetylpaclitaxel (DT), which can subsequently be converted to paclitaxel through acetylation at the C10 position. Chen et al. conducted single and multiple site-directed mutagenesis at 11 residues of the fungal glycosidases LXYL-P1-1 and LXYL-P1-2 to identify amino acid positions associated with enhanced activity, ultimately obtaining variants with improved catalytic performance. Among the 17 mutants, the E12 variant (A72T/V91S) displayed the highest β-xylosidase and β-glucosidase activities, exceeding those of LXYL-P1-2 by 2.8- and 3-fold, respectively [90]. During paclitaxel extraction, these analogs are often discarded, leading to resource waste and potential environmental contamination. The utilization of XDT as a substrate for paclitaxel semi-synthesis holds substantial practical value in reducing bioconversion costs.
In summary, engineering of key enzymes within the paclitaxel biosynthetic pathway constitutes an effective strategy. Such modifications can enhance substrate utilization and selectivity and improve catalytic efficiency, particularly through the targeted design of critical enzymes. These advances provide a solid foundation for future heterologous biosynthesis and industrial-scale production of paclitaxel.
5.8. Recent Study on Catalytic Mechanistic Analysis of FoTO1
Regarding recent studies on the catalytic mechanism elucidation of FoTO1, which is an NTF2-like superfamily protein in paclitaxel biosynthesis, two preprint articles have recently been published online. Jiang and Liu’s team demonstrates that FoTO1 is a dedicated epoxide isomerase that stereospecifically redirects a reactive epoxide intermediate toward productive T5α-ol formation [91]. Through in vitro reconstitution, mutagenesis, and QM/MM calculations, the authors show that a pre-organized D68/D149 catalytic dyad electrostatically activates epoxide ring opening via an isomerization mechanism, in contrast to the water-mediated hydrolysis employed by canonical epoxide hydrolases such as LEH. Modular dissection of the C-terminal extension further reveals a functional partition between catalytic integrity and productive coupling with T5αOH, mediated by specific hydrophobic contacts that enforce precise geometric complementarity at the binary complex interface. Together, these results demonstrate how electrostatic activation and enzyme association cooperate to control the fate of a highly reactive intermediate in paclitaxel biosynthesis. Sattely and McClune’s group systematically elucidates the dual role of FoTO1 [73]. On one hand, FoTO1 functions as an epoxide hydrolase that catalyzes the selective conversion of taxadiene-4(5)-epoxide to T5α-ol. On the other hand, through protein-protein interactions with atypical cytochrome P450 enzymes, FoTO1 serves a non-catalytic scaffolding function that organizes a multi-enzyme complex to enhance pathway efficiency. The study also demonstrates that the non-catalytic function of FoTO1 is generalizable across multiple plant diterpenoid biosynthetic pathways, providing new engineering strategies for heterologous production of plant natural products. Collectively, these two complementary studies not only resolve the long-standing catalytic mechanism of FoTO1 but also reveal its unprecedented non-catalytic scaffolding role, marking a significant advance in our understanding of paclitaxel biosynthesis and offering new avenues for pathway engineering in heterologous hosts.
6. The Emerging Technologies Development
To address the bottlenecks in paclitaxel pathway elucidation, enzyme engineering, and heterologous production, a suite of operationally defined emerging technologies has been deployed, with clear input materials, output data, and direct practical applications value.
6.1. MpXsn: High-Resolution Gene Co-Expression Network for Pathway Discovery
Recent advances in multi-omics and synthetic biology have provided a suite of emerging technologies that can be operationally deployed to overcome specific bottlenecks in paclitaxel biosynthesis. The multiplexed perturbation × single-nucleus sequencing (mpXsn) method takes as input Taxus tissues treated with 136 distinct perturbations, including hormones, pathway intermediates, and pathogens, followed by single-nucleus RNA sequencing [11]. Its output is a high-resolution gene co-expression network that partitions paclitaxel-associated genes into early, middle, and late expression modules. This technology has been practically applied to discover previously missing genes, especially for the auxiliary protein FoTO1, directly guiding the complete analysis of the paclitaxel synthesis pathway. Thus, mpXsn represents a paradigm shifting discovery tool that transforms correlative transcriptomic data into causal, network guided pathway elucidation.
6.2. MALDI-IMS: Spatial Metabolomics for Tissue-Specific Taxane Mapping
MALDI-imaging mass spectrometry (MALDI-IMS) operates on cryosections of Pseudotaxus tissues (such as stems, leaves, and phloem) and generates spatial metabolomic maps comprising up to 10,000 data points that reveal tissue-specific taxane distribution [10]. Practically, these maps identify where paclitaxel and its intermediates accumulate and enabled researchers to select appropriate tissue-specific promoters and design subcellular compartmentalization strategies in engineered plants. MALDI-IMS bridges a critical gap between conventional metabolomics and spatial biology by preserving tissue context, which is essential for understanding the compartmentalized nature of paclitaxel biosynthesis. However, current limitations include relatively low throughput, the need for specialized instrumentation, challenges in absolute quantification, and an inability to reliably distinguish isomeric compounds without tandem MS imaging. Nevertheless, when integrated with some other emerging technologies such as single-nucleus RNA sequencing (snRNA-seq) for cell-type resolution of gene expression, laser microdissection for targeted sampling of specific cell layers, and machine learning algorithms for automated metabolite annotation and spatial pattern recognition, MALDI-IMS can achieve a multi-dimensional understanding of the paclitaxel biosynthetic network, from spatial metabolomics to transcriptional regulation and predictive modeling.
6.3. Oxidase Toolbox: Site-Directed Oxidation Bypassing P450 Bottlenecks
The oxidase toolbox based on TteUPO mutants takes taxa-4(5),11(12)-diene as substrate and through engineered fungal unspecific peroxygenase variants expressed in E. coli, produces site-specific oxidized taxanes at C4, C6, C10, C11, C12, and C13 positions [17]. This toolbox bypasses the low-activity plant P450s and enables biosynthesis of oxidized taxanes in prokaryotic chassis while also generating diverse taxane analogs for medicinal chemistry. This represents a paradigm shift in taxane functionalization, as it replaces plant derived membrane-bound P450s with a soluble and bacterial-expressed peroxidase, thereby overcoming long standing bottlenecks in heterologous P450 expression and electron transfer chain assembly. At this stage, the regioselectivity of TteUPO mutants remains less precise than that of native P450s, and further directed evolution is needed to achieve the stereospecific oxidations required for authentic paclitaxel biosynthesis.
6.4. Gene Dropout Experiments
Gene dropout experiments using transient expression in N. benthamiana involve the specific deletion of a gene, which achieved by infiltrating tobacco leaves with Agrobacterium strains carrying candidate genes for combinatorial infiltration.This method realized the identification of indispensable pathway genes, such as C4β-C20 epoxidase, T9αOH, T1βOH, and PCL [14,15]. This approach has been practically applied to define the minimal gene set required for baccatin III and paclitaxel biosynthesis through systematic gene removal and to validate the order of pathway reactions [12]. Gene dropout experiments have been gradually extended to elucidate the biosynthetic pathways of various complex natural products. This dropout strategy is a powerful functional tool that enables accelerated determination of gene function, transforming correlative transcriptomic or genomic data into causal genetic evidence. When combined with modular pathway design and automated infiltration technologies, gene dropout screening offers an unprecedented capability for rapidly deconvoluting complex plant metabolic networks and accelerating the discovery of minimal biosynthetic routes for pharmaceutical production.
7. Conclusions and Outlooks
Paclitaxel, recognized as one of the most potent natural anticancer agents discovered to date, has undergone a diversified developmental history in its synthesis. At present, the most widely employed approach remains the semi-synthetic route [82]. Commercially available paclitaxel marketed by Bristol Myers Squibb is synthesized via the Holton–Ojima method, employing 10-deacetylbaccatin III as the precursor, with yields of up to 90%. However, the supply of semi-synthetic precursors, such as 10-deacetylbaccatin III or baccatin III, remains dependent on extraction from the scarce yew tree (Taxus spp.), severely limiting large-scale production and necessitating optimization via heterologous biosynthesis. Endophytic fungal fermentation has been investigated as an alternative source; however, yields remain low and unstable, with no reports of industrial-scale implementation to date [25]. While the co-evolution of endophytes and Taxus species has endowed these microorganisms with a diverse array of metabolic enzyme genes, their inherent genetic instability renders them currently unsuitable for use as industrial chassis cells. Consequently, endophytes are more likely to serve as a potential genetic reservoir in the future, providing critical resources for the mining of novel enzymatic parts. Plant cell culture technology provides another sustainable method for industrial-level paclitaxel production. For example, Phyton utilizes proprietary Plant Cell Fermentation (PCF®) technology to directly produce natural active pharmaceutical ingredients from suspension cultures [92]. In recent years, large-scale genome sequencing and analysis of various Taxus species have been completed, and the rapid advancement of synthetic biology has established favorable conditions for the heterologous biosynthesis of paclitaxel. This progress has enabled a paradigm shift in paclitaxel biosynthesis research, transitioning from traditional gene mining to systematic chassis reconstruction. With the recent advances in elucidating the oxetane ring formation mechanism, the nuclear transport factor 2 (NTF2)-like protein, FoTO1, and identifying previously unknown catalytic steps, long-standing critical gaps in the synthetic pathway have been bridged, thus rapidly advancing the resolution of the paclitaxel biosynthetic pathway and its heterologous synthesis. However, current research indicates that although the paclitaxel metabolic pathway has been elucidated, this knowledge has not yet been substantially translated into efficient metabolic flux within engineered chassis. Consequently, enhancing heterologous metabolic flux remains the paramount bottleneck in the field. Currently, advances in microbial and tobacco cell platforms have further facilitated the heterologous biosynthesis of paclitaxel intermediates. However, these chassis cells are fundamentally constrained by a shared core bottleneck: the high functional promiscuity and exceedingly low catalytic efficiency of plant-derived core catalytic elements, particularly cytochrome P450 enzymes and acyltransferases. Consequently, this results in the diversion of metabolic flux and the concomitant accumulation of byproducts. Furthermore, significant discrepancies exist in the efficiency of membrane protein coupling and microenvironmental adaptation across different chassis platforms. Therefore, future research directions should focus on integrating bioinformatics with high-sensitivity multi-species screening systems to systematically investigate the distribution and functional diversity of Taxus-derived cytochrome P450 enzymes and acyltransferases. Furthermore, the functional characterization of currently unidentified catalytic parts will further elucidate the reaction steps and complete the paclitaxel biosynthetic landscape. Subsequently, employing methodologies such as multi-species enzyme screening, rational design, and machine learning-assisted enzyme engineering will facilitate the elucidation of the catalytic mechanisms and evolutionary trajectories of key P450 enzymes at the molecular level. By coupling these insights with rational engineering strategies to optimize enzyme activity, the fundamental issue of enzyme promiscuity can be resolved. To address the practical imperatives of industrialized production, the assembly of paclitaxel biosynthetic modules in tobacco cells or engineered eukaryotic microbes must be combined with the simultaneous optimization of critical factors, including cofactor (NADPH, O2) regeneration, coenzyme compatibility, and precursor supply. This comprehensive approach will alleviate the limitations restricting the heterologous biosynthesis of paclitaxel, ultimately enabling highly efficient heterologous production.
Moreover, integrating chemical synthesis and biosynthesis in a complementary manner could enable greener, more efficient production of paclitaxel [93]. The superiority of this strategy lies in its cross-disciplinary complementarity: it leverages microbial chassis for the construction of the complex taxane polycyclic core skeleton, followed by the application of chemical approaches to append specific side-chain functional groups, ultimately yielding paclitaxel. This synthetic paradigm not only effectively circumvents the inefficiencies inherent in downstream biocatalytic steps but also significantly mitigates the severe environmental impact and high energy consumption associated with traditional total synthesis. In summary, the multifaceted research currently surrounding paclitaxel biosynthesis has not only systematically reconstructed its complex and extensive metabolic network at the theoretical level and elucidated the evolutionary mechanisms of core catalytic elements, but has also laid the critical groundwork for overcoming the catalytic bottlenecks associated with plant-derived cytochrome P450 enzymes. These efforts are of significant scientific importance for deepening the mechanistic understanding of paclitaxel biosynthesis, overcoming bottlenecks associated with key catalytic elements, and advancing the synthetic biology of paclitaxel and its derivatives.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the authors used DeepSeek and ChatGPT to polish specific sentences to better align with English writing standards. Following the use of these tools, the authors carefully reviewed and edited the content as necessary and assume full responsibility for the content of the published article.
Author Contributions
X.L.: designed, wrote and revised the paper. Y.Z.: wrote the paper and collected the data. J.L.: wrote the paper and collected the data. H.J.: revised the manuscript.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable to this article as no datasets were generated or analysed during the current study.
Funding
This work was supported by the National Natural Science Foundation of China (Grant No. 32371499), Natural Science Foundation of Wuhan (2025040601020158), Science and Technology Major Project of Guangxi (Guike AA24206048, Guike AA24206050-3, Guike JF2504850012), Hubei University of Technology High-Level Talent Research Startup Fund Program (4301/00960), Hubei Provincial Key R&D Major Projects (2025BCA006), Youth Science and Technology Talent Cultivation Special Program of Hubei Province (2025DJA024).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Prota AE, Katja B, Didier Z, Field JJ, José Fernando D, Karl-Heinz A, et al. Molecular mechanism of action of microtubule-stabilizing anticancer agents. Science 2013, 339, 587–590. DOI:10.1126/science.1230582 [Google Scholar]
-
Li C, Wang Y, Yin X, Sun X, Sui S, Liu J, et al. Novel alpha-KG/Fe(II)-Dependent Dioxygenases Catalyzing C1beta-Hydroxylation and Construction of 5/7/6-Skeleton of Highly Oxygenated Taxoids. Angew. Chem. Int. Ed. 2025, 137, e202517041. DOI:10.1002/ange.202517041 [Google Scholar]
-
Winegar PH, Astolfi MC, Holm SF, Hudson GA, Keasling JD. Advances in the microbial biosynthesis of therapeutic terpenoids. Curr. Opin. Biotechnol. 2025, 96, 103374. DOI:10.1016/j.copbio.2025.103374 [Google Scholar]
-
Zhu Y, Liang F, Chai P, Kampranis SC, Zhao Y. Recent Advances in the Biosynthesis and Biotechnological Production of Taxol. Engineering 2025, 4, 65–75. DOI:10.1016/j.eng.2025.12.018 [Google Scholar]
-
Tan CL, Yu X, Feng HC, Gershenzon J, Liu Y, Li SH. A synthetic biology roadmap for sustainable production of the plant-originated anti-cancer drug paclitaxel. Trends Biotechnol. 2026. DOI:10.1016/j.tibtech.2025.11.013 [Google Scholar]
-
Xiong X, Gou J, Liao Q, Li Y, Zhou Q, Bi G, et al. The Taxus genome provides insights into paclitaxel biosynthesis. Nat. Plants 2021, 7, 1026–1036. DOI:10.1038/s41477-021-00963-5 [Google Scholar]
-
Cheng J, Wang X, Liu X, Zhu X, Li Z, Chu H, et al. Chromosome-level genome of Himalayan yew provides insights into the origin and evolution of the paclitaxel biosynthetic pathway. Mol. Plant 2021, 14, 1199–1209. DOI:10.1016/j.molp.2021.04.015 [Google Scholar]
-
Song C, Fu F, Yang L, Niu Y, Tian Z, He X, et al. Taxus yunnanensis genome offers insights into gymnosperm phylogeny and taxol production. Commun. Biol. 2021, 4, 1203. DOI:10.1038/s42003-021-02697-8 [Google Scholar]
-
Zhang Y, Scossa F, Fernie AR. The genomes of Taxus species unveil novel candidates in the biosynthesis of taxoids. Mol. Plant 2021, 14, 1773–1775. DOI:10.1016/j.molp.2021.08.017 [Google Scholar]
-
Wang M, Ma R, Fang Z, Zhang L, Zhang Y, Zheng M, et al. Analysis genome of Pseudotaxus chienii reveals insights into the origin and evolution of taxane biosynthesis. Nat. Commun. 2025, 17, 1088. DOI:10.1038/s41467-025-67849-4 [Google Scholar]
-
McClune CJ, Liu JC, Wick C, De La Pena R, Lange BM, Fordyce PM, et al. Discovery of FoTO1 and Taxol genes enables biosynthesis of baccatin III. Nature 2025, 643, 582–592. DOI:10.1038/s41586-025-09090-z [Google Scholar]
-
Jiang B, Gao L, Wang H, Sun Y, Zhang X, Ke H, et al. Characterization and heterologous reconstitution of Taxus biosynthetic enzymes leading to baccatin III. Science 2024, 383, 622–629. DOI:10.1126/science.adj3484 [Google Scholar]
-
Yang C, Wang Y, Su Z, Xiong L, Wang P, Lei W, et al. Biosynthesis of the highly oxygenated tetracyclic core skeleton of Taxol. Nat. Commun. 2024, 15, 2339. DOI:10.1038/s41467-024-46583-3 [Google Scholar]
-
Li C, Yin X, Wang S, Sui S, Liu J, Sun X, et al. A Cytochrome P450 Enzyme Catalyses Oxetane Ring Formation in Paclitaxel Biosynthesis. Angewandte Chem. Int. Ed. 2024, 63, e202407070. DOI:10.1002/anie.202407070 [Google Scholar]
-
Zhang Y, Wiese L, Fang H, Alseekh S, Perez de Souza L, Scossa F, et al. Synthetic biology identifies the minimal gene set required for Paclitaxel biosynthesis in a plant chassis. Mol. Plant 2023, 16, 1951–1961. DOI:10.1016/j.molp.2023.10.016 [Google Scholar]
-
Liang F, Xie Y, Zhang C, Zhao Y, Motawia MS, Kampranis SC. Elucidation of the final steps in Taxol biosynthesis and its biotechnological production. Nat. Synth. 2025, 4, 1212–1222. DOI:10.1038/s44160-025-00800-z [Google Scholar]
-
Lai M, Weng Y, Wei J, Lu XY, Huang Z, Li H, et al. Designing an oxidase toolbox for site-directed oxidation of taxanes. Nat. Commun. 2025, 17, 841. DOI:10.1038/s41467-025-67544-4 [Google Scholar]
-
Tang X, Zhang X, Long LP, Wu S, Fan XY, Liu J, et al. Versatile 2-Oxoglutarate-Dependent Dioxygenases Catalyze Radical-Mediated Multifunctional Skeleton Reconstructions and Oxidation Modifications of Taxoids. J. Am. Chem. Soc. 2026, 148, 11333–11343. DOI:10.1021/jacs.6c01169 [Google Scholar]
-
Bian G, Yuan Y, Tao H, Shi X, Zhong X, Han Y, et al. Production of taxadiene by engineering of mevalonate pathway in Escherichia coli and endophytic fungus Alternaria alternata TPF6. Biotechnol. J. 2017, 12, 1600697. DOI:10.1002/biot.201600697 [Google Scholar]
-
Williams DC, Wildung MR, Jin AQ, Dalal D, Oliver JS, Coates RM, et al. Heterologous expression and characterization of a “Pseudomature” form of taxadiene synthase involved in paclitaxel (Taxol) biosynthesis and evaluation of a potential intermediate and inhibitors of the multistep diterpene cyclization reaction. Arch. Biochem. Biophys. 2000, 379, 137–146. DOI:10.1006/abbi.2000.1865 [Google Scholar]
-
Edgar S, Zhou K, Qiao K, King JR, Simpson JH, Stephanopoulos G. Mechanistic Insights into Taxadiene Epoxidation by Taxadiene-5alpha-Hydroxylase. ACS Chem. Biol. 2016, 11, 460–469. DOI:10.1021/acschembio.5b00767 [Google Scholar]
-
Edgar S, Li FS, Qiao K, Weng JK, Stephanopoulos G. Engineering of Taxadiene Synthase for Improved Selectivity and Yield of a Key Taxol Biosynthetic Intermediate. ACS Synth. Biol. 2017, 6, 201–205. DOI:10.1021/acssynbio.6b00206 [Google Scholar]
-
Zhao Y, Liang F, Xie Y, Duan Y-T, Andeadelli A, Pateraki I, et al. Oxetane Ring Formation in Taxol Biosynthesis Is Catalyzed by a Bifunctional Cytochrome P450 Enzyme. J. Am. Chem. Soc. 2023, 146, 801–810. DOI:10.1021/jacs.3c10864 [Google Scholar]
-
Li C, Yin X, Dai J. The History of Studies on Oxetane Ring Formation in Paclitaxel Biosynthesis. Chembiochem 2025, 26, e202400947. DOI:10.1002/cbic.202400947 [Google Scholar]
-
Tong Y, Luo YF, Gao W. Biosynthesis of paclitaxel using synthetic biology. Phytochem. Rev. 2021, 21, 863–877. DOI:10.1007/s11101-021-09766-0 [Google Scholar]
-
Liu JCT, De La Pena R, Tocol C, Sattely ES. Reconstitution of early paclitaxel biosynthetic network. Nat. Commun. 2024, 15, 1419. DOI:10.1038/s41467-024-45574-8 [Google Scholar]
-
Li C, Sun X, Chen R, Xie K, Chen D, Liu J, et al. The discovery of missing taxane C13α-O-deacetylases redelineates the biosynthetic pathway of paclitaxel. bioRxiv 2026. DOI:10.64898/2026.04.28.721278 [Google Scholar]
-
Chau M, Walker K, Long R, Croteau R. Regioselectivity of taxoid-O-acetyltransferases: Heterologous expression and characterization of a new taxadien-5alpha-ol-O-acetyltransferase. Arch. Biochem. Biophys. 2004, 430, 237–246. DOI:10.1016/j.abb.2004.07.013 [Google Scholar]
-
Jennewein S, Rithner CD, Williams RM, Croteau R. Taxoid metabolism: Taxoid 14beta-hydroxylase is a cytochrome P450-dependent monooxygenase. Arch. Biochem. Biophys. 2003, 413, 262–270. DOI:10.1016/S0003-9861(03)00090-0 [Google Scholar]
-
Walker KD, Klettke K, Akiyama T, Croteau R. Cloning, heterologous expression, and characterization of a phenylalanine aminomutase involved in Taxol biosynthesis. J. Biol. Chem. 2004, 279, 53947–53954. DOI:10.1074/jbc.M411215200 [Google Scholar]
-
Ramirez-Estrada K, Altabella T, Onrubia M, Moyano E, Notredame C, Osuna L, et al. Transcript profiling of jasmonate-elicited Taxus cells reveals a beta-phenylalanine-CoA ligase. Plant Biotechnol. J. 2016, 14, 85–96. DOI:10.1111/pbi.12359 [Google Scholar]
-
Walker K, Long R, Croteau R. The final acylation step in taxol biosynthesis: Cloning of the taxoid C13-side-chain N-benzoyltransferase from Taxus. Proc. Natl. Acad. Sci. USA 2002, 99, 9166–9171. DOI:10.1073/pnas.082115799 [Google Scholar]
-
Hefner J, Ketchum REB, Croteau R. Cloning and functional expression of a cDNA encoding geranylgeranyl diphosphate synthase from Taxus canadensis and assessment of the role of this prenyltransferase in cells induced for taxol production. Arch. Biochem. Biophys. 1998, 360, 62–74. DOI:10.1006/abbi.1998.0926 [Google Scholar]
-
Wildung MR, Croteau R. A cDNA clone for taxadiene synthase, the diterpene cyclase that catalyzes the committed step of taxol biosynthesis. J. Biol. Chem. 1996, 271, 9201–9204. DOI:10.1074/jbc.271.16.9201 [Google Scholar]
-
Jennewein S, Long RM, Williams RM, Croteau R. Cytochrome P450 taxadiene 5 alpha-hydroxylase, a mechanistically unusual monooxygenase catalyzing the first oxygenation step of taxol biosynthesis. Chem. Biol. 2004, 11, 379–387. DOI:10.1016/j.chembiol.2004.02.022 [Google Scholar]
-
Jennewein S, Rithner CD, Williams RM, Croteau RB. Taxol biosynthesis: Taxane 13 alpha-hydroxylase is a cytochrome P450-dependent monooxygenase. Proc. Natl. Acad. Sci. USA 2001, 98, 13595–13600. DOI:10.1073/pnas.251539398 [Google Scholar]
-
Walker K, Schoendorf A, Croteau R. Molecular cloning of a taxa-4(20),11(12)-dien-5alpha-ol-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli. Arch. Biochem. Biophys. 2000, 374, 371–380. DOI:10.1006/abbi.1999.1609 [Google Scholar]
-
Chau M, Jennewein S, Walker K, Croteau R. Taxol biosynthesis: Molecular cloning and of a cytochrome p450 characterization taxoid 7 beta-hydroxylase. Chem. Biol. 2004, 11, 663–672. DOI:10.1016/S1074-5521(04)00111-5 [Google Scholar]
-
Chau M, Croteau R. Molecular cloning and characterization of a cytochrome P450 taxoid 2 alpha-hydroxylase involved in Taxol biosynthesis. Arch. Biochem. Biophys. 2004, 427, 48–57. DOI:10.1016/j.abb.2004.04.016 [Google Scholar]
-
Walker K, Croteau R. Taxol biosynthesis: Molecular cloning of a benzoyl-CoA. Taxane 2 alpha-O-benzoyltransferase cDNA from Taxus and functional expression in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 13591–13596. DOI:10.1073/pnas.250491997 [Google Scholar]
-
Walker K, Croteau R. Molecular cloning of a 10-deacetylbaccatin III-10-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli. Proc. Natl. Acad. Sci. USA 2000, 97, 583–587. DOI:10.1073/pnas.97.2.583 [Google Scholar]
-
Sanchez-Munoz R, Perez-Mata E, Almagro L, Cusido RM, Bonfill M, Palazon J, et al. A Novel Hydroxylation Step in the Taxane Biosynthetic Pathway: A New Approach to Paclitaxel Production by Synthetic Biology. Front. Bioeng. Biotechnol. 2020, 8, 410. DOI:10.3389/fbioe.2020.00410 [Google Scholar]
-
Li T, Li B, Liao C, Zhang H, Wang L, Fu T, et al. Transcriptome analysis provides insights into light condition effect on paclitaxel biosynthesis in yew saplings. BMC Plant Biol. 2022, 22, 577. DOI:10.1186/s12870-022-03958-2 [Google Scholar]
-
Mubeen S, Li ZL, Huang QM, He CT, Yang ZY. Comparative Transcriptome Analysis Revealed the Tissue-Specific Accumulations of Taxanes among Three Experimental Lines of Taxus yunnanensis. J. Agric. Food Chem. 2018, 66, 10410–10420. DOI:10.1021/acs.jafc.8b03502 [Google Scholar]
-
Wang T, Chen Y, Zhuang W, Zhang F, Shu X, Wang Z, et al. Transcriptome Sequencing Reveals Regulatory Mechanisms of Taxol Synthesis in Taxus wallichiana var. Mairei. Int. J. Genom. 2019, 2019, 1596895. DOI:10.1155/2019/1596895 [Google Scholar]
-
Yu C, Luo X, Zhang C, Xu X, Huang J, Chen Y, et al. Tissue-specific study across the stem of Taxus media identifies a phloem-specific TmMYB3 involved in the transcriptional regulation of paclitaxel biosynthesis. Plant J. 2020, 103, 95–110. DOI:10.1111/tpj.14710 [Google Scholar]
-
Yu C, Hou K, Zhang H, Liang X, Chen C, Wang Z, et al. Integrated mass spectrometry imaging and single-cell transcriptome atlas strategies provide novel insights into taxoid biosynthesis and transport in Taxus mairei stems. Plant J. 2023, 115, 1243–1260. DOI:10.1111/tpj.16315 [Google Scholar]
-
Huang Q, Roessner CA, Croteau R, Scott AI. Engineering Escherichia coli for the Synthesis of Taxadiene, a Key Intermediate in the Biosynthesis of Taxol. Bioorganic Med. Chem. 2001, 9, 2237–2242. DOI:10.1016/S0968-0896(01)00072-4 [Google Scholar]
-
Zhou K, Qiao K, Edgar S, Stephanopoulos G. Distributing a metabolic pathway among a microbial consortium enhances production of natural products. Nat. Biotechnol. 2015, 33, 377–383. DOI:10.1038/nbt.3095 [Google Scholar]
-
Boghigian BA, Armando J, Salas D, Pfeifer BA. Computational identification of gene over-expression targets for metabolic engineering of taxadiene production. Appl. Microbiol. Biotechnol. 2012, 93, 2063–2073. DOI:10.1007/s00253-011-3725-1 [Google Scholar]
-
Abdallah II, Pramastya H, van Merkerk R, Sukrasno, Quax WJ. Metabolic Engineering of Bacillus subtilis Toward Taxadiene Biosynthesis as the First Committed Step for Taxol Production. Front. Microbiol. 2019, 10, 218. DOI:10.3389/fmicb.2019.00218 [Google Scholar]
-
Biggs BW, Lim CG, Sagliani K, Shankar S, Stephanopoulos G, De Mey M, et al. Overcoming heterologous protein interdependency to optimize P450-mediated Taxol precursor synthesis in Escherichia coli. Proc. Natl. Acad. Sci. USA 2016, 113, 3209–3214. DOI:10.1073/pnas.1515826113 [Google Scholar]
-
Mutanda I, Li J, Xu F, Wang Y. Recent Advances in Metabolic Engineering, Protein Engineering, and Transcriptome-Guided Insights Toward Synthetic Production of Taxol. Front. Bioeng. Biotechnol. 2021, 9, 632269. DOI:10.3389/fbioe.2021.632269 [Google Scholar]
-
Ding MZ, Yan HF, Li LF, Zhai F, Shang LQ, Yin Z, et al. Biosynthesis of Taxadiene in Saccharomyces cerevisiae: Selection of geranylgeranyl diphosphate synthase directed by a computer-aided docking strategy. PLoS ONE 2014, 9, e109348. DOI:10.1371/journal.pone.0109348 [Google Scholar]
-
Nowrouzi B, Li RA, Walls LE, d’Espaux L, Malci K, Liang L, et al. Enhanced production of taxadiene in Saccharomyces cerevisiae. Microb. Cell Factories 2020, 19, 200. DOI:10.1186/s12934-020-01458-2 [Google Scholar]
-
Walls LE, Malci K, Nowrouzi B, Li RA, d’Espaux L, Wong J, et al. Optimizing the biosynthesis of oxygenated and acetylated Taxol precursors in Saccharomyces cerevisiae using advanced bioprocessing strategies. Biotechnol. Bioeng. 2021, 118, 279–293. DOI:10.1002/bit.27569 [Google Scholar]
-
Nowrouzi B, Lungang L, Rios-Solis L. Exploring optimal Taxol(R) CYP725A4 activity in Saccharomyces cerevisiae. Microb. Cell Factories 2022, 21, 197. DOI:10.1186/s12934-022-01922-1 [Google Scholar]
-
Jiang Z, Kempinski C, Bush CJ, Nybo SE, Chappell J. Engineering Triterpene and Methylated Triterpene Production in Plants Provides Biochemical and Physiological Insights into Terpene Metabolism. Plant Physiol. 2016, 170, 702–716. DOI:10.1104/pp.15.01548 [Google Scholar]
-
Besumbes O, Sauret-Gueto S, Phillips MA, Imperial S, Rodriguez-Concepcion M, Boronat A. Metabolic engineering of isoprenoid biosynthesis in Arabidopsis for the production of taxadiene, the first committed precursor of Taxol. Biotechnol. Bioeng. 2004, 88, 168–175. DOI:10.1002/bit.20237 [Google Scholar]
-
Rontein D, Onillon S, Herbette G, Lesot A, Werck-Reichhart D, Sallaud C, et al. CYP725A4 from yew catalyzes complex structural rearrangement of taxa-4(5),11(12)-diene into the cyclic ether 5(12)-oxa-3(11)-cyclotaxane. J. Biol. Chem. 2008, 283, 6067–6075. DOI:10.1074/jbc.M708950200 [Google Scholar]
-
Li J, Mutanda I, Wang K, Yang L, Wang J, Wang Y. Chloroplastic metabolic engineering coupled with isoprenoid pool enhancement for committed taxanes biosynthesis in Nicotiana benthamiana. Nat. Commun. 2019, 10, 4850. DOI:10.1038/s41467-019-12879-y [Google Scholar]
-
Fu J, Xu W, Huang W, Wang B, Li S, Zhang J, et al. Importation of taxadiene synthase into chloroplast improves taxadiene production in tobacco. Planta 2021, 253, 107. DOI:10.1007/s00425-021-03626-z [Google Scholar]
-
He J, Plácido JPA, Pateraki I, Kampranis S, Favero BT, Lütken H. Hairy root induction of Taxus baccata L. by natural transformation with Rhizobium rhizogenes. Horticulturae 2023, 9, 4. DOI:10.3390/horticulturae9010004 [Google Scholar]
-
Ajikumar PK, Xiao WH, Tyo KE, Wang Y, Simeon F, Leonard E, et al. Isoprenoid pathway optimization for Taxol precursor overproduction in Escherichia coli. Science 2010, 330, 70–74. DOI:10.1126/science.1191652 [Google Scholar]
-
Boghigian BA, Salas D, Ajikumar PK, Stephanopoulos G, Pfeifer BA. Analysis of heterologous taxadiene production in K- and B-derived Escherichia coli. Appl. Microbiol. Biotechnol. 2012, 93, 1651–1661. DOI:10.1007/s00253-011-3528-4 [Google Scholar]
-
Wu Q-Y, Huang Z-Y, Wang J-Y, Yu H-L, Xu J-H. Construction of an Escherichia coli cell factory to synthesize taxadien-5α-ol, the key precursor of anti-cancer drug paclitaxel. Bioresour. Bioprocess. 2022, 9, 82. DOI:10.1186/s40643-022-00569-5 [Google Scholar]
-
Wang J-Y, Huang Z-Y, Wu Q-Y, Pan J, Li C-X, Xu J-H. Facile biosynthesis of taxadiene by a newly constructed Escherichia coli strain fusing enzymes taxadiene synthase and geranylgeranyl pyrophosphate synthase. Process Biochem. 2022, 122, 129–136. DOI:10.1016/j.procbio.2022.09.005 [Google Scholar]
-
Engels B, Dahm P, Jennewein S. Metabolic engineering of taxadiene biosynthesis in yeast as a first step towards Taxol (Paclitaxel) production. Metab. Eng. 2008, 10, 201–206. DOI:10.1016/j.ymben.2008.03.001 [Google Scholar]
-
Walls LE, Martinez JL, Rios-Solis L. Enhancing Saccharomyces cerevisiae Taxane Biosynthesis and Overcoming Nutritional Stress-Induced Pseudohyphal Growth. Microorganisms 2022, 10, 163. DOI:10.3390/microorganisms10010163 [Google Scholar]
-
Xu M, Xie W, Luo Z, Li CX, Hua Q, Xu J. Improving solubility and copy number of taxadiene synthase to enhance the titer of taxadiene in Yarrowia lipolytica. Synth. Syst. Biotechnol. 2023, 8, 331–338. DOI:10.1016/j.synbio.2023.04.002 [Google Scholar]
-
Min L, Han JC, Zhang W, Gu CC, Zou YP, Li CC. Strategies and Lessons Learned from Total Synthesis of Taxol. Chem. Rev. 2023, 123, 4934–4971. DOI:10.1021/acs.chemrev.2c00763 [Google Scholar]
-
Hasan MM, Kim HS, Jeon JH, Kim SH, Moon B, Song JY, et al. Metabolic engineering of Nicotiana benthamiana for the increased production of taxadiene. Plant Cell Rep. 2014, 33, 895–904. DOI:10.1007/s00299-014-1568-9 [Google Scholar]
-
Wick C, Somani A, Liu JC, Karunadasa SS, Xu SL, Fordyce PM, et al. FoTO1 orchestrates Taxol biosynthesis through catalytic and non-catalytic mechanisms. BioRxiv 2026. DOI:10.64898/2026.03.21.713420 [Google Scholar]
-
Wang Y, Guo B, Miao Z, Tang K. Transformation of taxol-producing endophytic fungi by restriction enzyme-mediated integration (REMI). FEMS Microbiol. Lett. 2007, 273, 253–259. DOI:10.1111/j.1574-6968.2007.00801.x [Google Scholar]
-
El-Sayed ASA, Abdel-Ghany SE, Ali GS. Genome editing approaches: Manipulating of lovastatin and taxol synthesis of filamentous fungi by CRISPR/Cas9 system. Appl. Microbiol. Biotechnol. 2017, 101, 3953–3976. DOI:10.1007/s00253-017-8263-z [Google Scholar]
-
Zhong J, Wang Y, Chen Z, Yalikun Y, He L, Liu T, et al. Engineering cyanobacteria as a new platform for producing taxol precursors directly from carbon dioxide. Biotechnol. Biofuels Bioprod. 2024, 17, 99. DOI:10.1186/s13068-024-02555-9 [Google Scholar]
-
Ansbacher T, Freud Y, Major DT. Slow-Starter Enzymes: Role of Active-Site Architecture in the Catalytic Control of the Biosynthesis of Taxadiene by Taxadiene Synthase. Biochemistry 2018, 57, 3773–3779. DOI:10.1021/acs.biochem.8b00452 [Google Scholar]
-
Liu H, Chen Q. Computational protein design for given backbone: Recent progresses in general method-related aspects. Curr. Opin. Struct. Biol. 2016, 39, 89–95. DOI:10.1016/j.sbi.2016.06.013 [Google Scholar]
-
Koksal M, Jin Y, Coates RM, Croteau R, Christianson DW. Taxadiene synthase structure and evolution of modular architecture in terpene biosynthesis. Nature 2011, 469, 116–120. DOI:10.1038/nature09628 [Google Scholar]
-
Soliman S, Tang Y. Natural and engineered production of taxadiene with taxadiene synthase. Biotechnol. Bioeng. 2015, 112, 229–235. DOI:10.1002/bit.25468 [Google Scholar]
-
Van Rijn JPM, Escorcia AM, Thiel W. QM/MM study of the taxadiene synthase mechanism. J. Comput. Chem. 2019, 40, 1902–1910. DOI:10.1002/jcc.25846 [Google Scholar]
-
Kanda Y, Ishihara Y, Wilde NC, Baran PS. Two-Phase Total Synthesis of Taxanes: Tactics and Strategies. J. Org. Chem. 2020, 85, 10293–10320. DOI:10.1021/acs.joc.0c01287 [Google Scholar]
-
Barton NA, Marsh BJ, Lewis W, Narraidoo N, Seymour GB, Fray R, et al. Accessing low-oxidation state taxanes: Is taxadiene-4(5)-epoxide on the taxol biosynthetic pathway? Chem. Sci. 2016, 7, 3102–3107. DOI:10.1039/C5SC03463A [Google Scholar]
-
Schrepfer P, Buettner A, Goerner C, Hertel M, van Rijn J, Wallrapp F, et al. Identification of amino acid networks governing catalysis in the closed complex of class I terpene synthases. Proc. Natl. Acad. Sci. USA 2016, 113, 958–967. DOI:10.1073/pnas.1519680113 [Google Scholar]
-
Yadav VG. Unraveling the multispecificity and catalytic promiscuity of taxadiene monooxygenase. J. Mol. Catal. B Enzym. 2014, 110, 154–164. DOI:10.1016/j.molcatb.2014.10.004 [Google Scholar]
-
Song X, Wang Q, Zhu X, Fang W, Liu X, Shi C, et al. Unraveling the Catalytic Mechanism of Taxadiene-5α-hydroxylase from Crystallography and Computational Analyses. ACS Catal. 2024, 14, 3912–3925. DOI:10.1021/acscatal.3c05807 [Google Scholar]
-
Zhang MF, Xie WL, Chen C, Li CX, Xu JH. Computational redesign of taxane-10β-hydroxylase for de novo biosynthesis of a key paclitaxel intermediate. Appl. Microbiol. Biotechnol. 2023, 107, 7105–7117. DOI:10.1007/s00253-023-12784-x [Google Scholar]
-
Li BJ, Wang H, Gong T, Chen JJ, Chen TJ, Yang JL, et al. Improving 10-deacetylbaccatin III-10-β-O-acetyltransferase catalytic fitness for Taxol production. Nat. Commun. 2017, 8, 15544. DOI:10.1038/ncomms15544 [Google Scholar]
-
Lin SL, Wei T, Lin JF, Guo LQ, Wu GP, Wei JB, et al. Bio-production of Baccatin III, an Important Precursor of Paclitaxel by a Cost-Effective Approach. Mol. Biotechnol. 2018, 60, 492–505. DOI:10.1007/s12033-018-0090-7 [Google Scholar]
-
Chen JJ, Liang X, Wang F, Wen YH, Chen TJ, Liu WC, et al. Combinatorial mutation on the β-glycosidase specific to 7-β-xylosyltaxanes and increasing the mutated enzyme production by engineering the recombinant yeast. Acta Pharm. Sin. B 2019, 9, 626–638. DOI:10.1016/j.apsb.2018.11.003 [Google Scholar]
-
Bai J, Li J, Zhang Y, Chang H, Zhang N, Liu Y, et al. FoTO1 is an epoxide isomerase in paclitaxel biosynthesis. BioRxiv 2026. DOI:10.64898/2026.03.30.715439 [Google Scholar]
-
Zhu X, Liu X, Liu T, Wang Y, Ahmed N, Li Z, et al. Synthetic biology of plant natural products: From pathway elucidation to engineered biosynthesis in plant cells. Plant Commun. 2021, 2, 100229. DOI:10.1016/j.xplc.2021.100229 [Google Scholar]
-
Li J, Liu X, Zhu X, Liu J, Zhang L, Ahmed N, et al. Biochemical synthesis of taxanes from mevalonate. Synth. Syst. Biotechnol. 2024, 9, 694–700. DOI:10.1016/j.synbio.2024.05.002 [Google Scholar]