Biomanufacturing with broad applications in various industries is projected to reach a market value of ~30 trillion USD by 2030, accounting for more than one third of the global manufacturing output. Future biomanufacturing of industrial products will use novel synthetic biology tools and advanced bioprocesses to convert abundant biomass and waste resources into value-added products with comparable or superior properties to replace current petroleum-based products, thus enabling circular bioeconomy with affordable energy, economic growth, and innovation in renewable energy and chemicals production. However, biomanufacturing faces many challenges in its development that requires fundamental research in synthetic biology and novel bioprocesses involving multidisciplinary teams and academic-industry partnerships. In particular, aging and lifespan of microbial cells have been largely overlooked in industrial fermentation. Only recently have microbiologists realized that many microorganisms including yeasts (e.g., Saccharomyces cerevisiae) and bacteria (e.g., Escherichia coli) have chronological and replicative life spans which dramatically impact cell viability and longevity. In this article, we will give our perspective on how synthetic biology may contribute to overcoming some challenges facing industrial biotechnology for fuels and chemicals production from renewable sources, highlighting the importance of understanding and regulating microorganism’s lifespan and aging.
1. Challenges in Industrial Fermentation
Industrial fermentation has long been used to produce various bioproducts, including ethanol, butanol, citric acid, lactic acid, lysine, xanthan gum, and antibiotics, etc. that cannot be economically produced by chemical synthesis. Recent advances in synthetic biology and metabolic engineering have enabled the biosynthesis of almost all kinds of chemicals and materials that are used in our daily life from renewable resources including plant biomass, agricultural residues, food wastes, municipal solid wastes, and industrial wastes. However, bioproduction of industrial products, including fuels and chemicals, generally cannot compete with products derived from petrochemical routes because conventional fermentation processes are limited by relatively low product titer, yield, and productivity which result in high capital, operating, and product costs. To overcome these limitations, extensive research and development efforts have focused on developing novel microbial strains and bioprocesses.
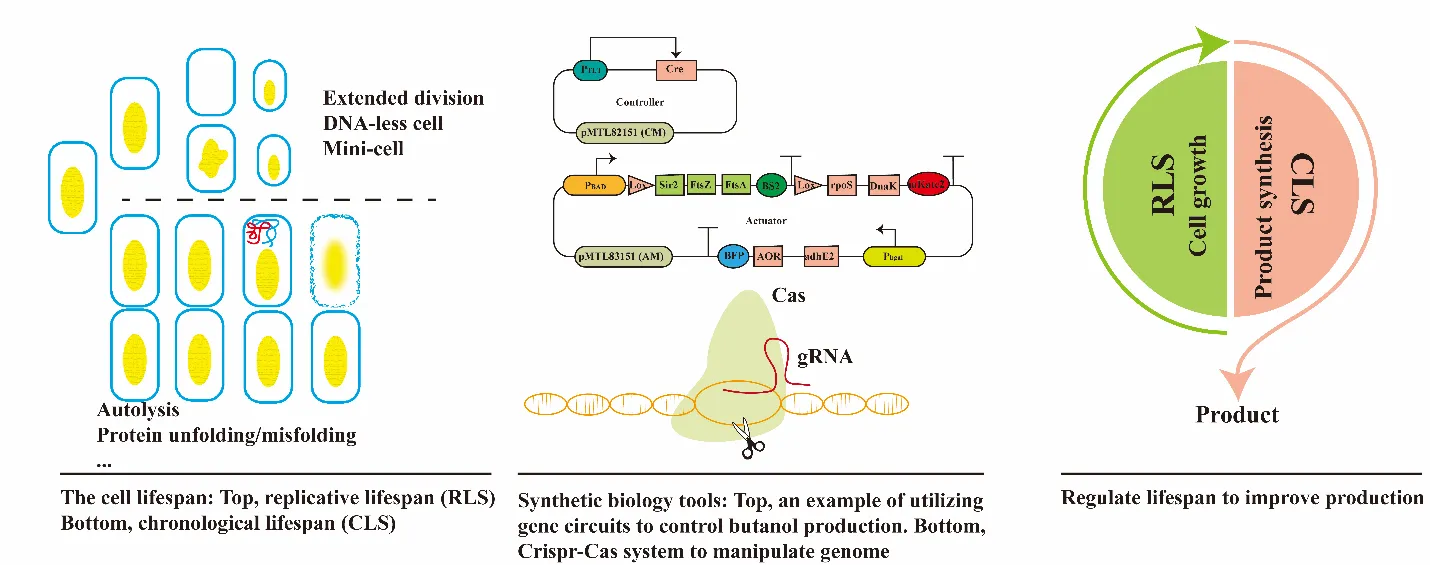
Industrial production of chemicals and fuels in microbial fermentation is often limited by low product titer, productivity or rate, and yield (TRY), poor process stability, short production duration (longevity). Understanding genes and process factors affecting microbial lifespan and aging, which have dramatic impacts on cell viability, process performance (TRY), and longevity in industrial fermentation but has been largely overlooked. Few microbial strains of industrial interest have been evaluated under different culture and stress conditions to study their effects on growth/fermentation kinetics and culture stability/longevity. Through population and transcriptomics analyses, genes/enzymes contributing to culture heterogeneity, production variability, and limited production duration or longevity can be identified. Then, synthetic biology tools including gene circuits and CRISPR genome engineering can be used to engineer strains with prolonged lifespan and mediated aging via reduced sensitivity to environmental stress. In general, microbial strains with increased lifespan or mitigated aging will be more productive for a longer duration in industrial fermentation. Such strains can be developed through the design-build-test-learn (DBTL) cycle and used in advanced continuous fermentation process with in-situ product recovery, achieving at least 50% improvements in TRY for a continuous production duration of >6 months. With lifespan engineered strains, integrated bioprocesses including feedstock pretreatment, enzymatic hydrolysis, fermentation with in-situ product separation, and product purification can be optimized with intelligent process control in future biomanufacturing.
2. Microbial Lifespan and Aging
Most bacteria typically produce two apparently (morphologically) identical cells by binary division and are considered to be nonsenescent. The aging process was believed to be specific to eukaryotes until 2004 when the signs of replicative aging in
Caulobacter crescentus were demonstrated [
1]. It was then reported that
E. coli cells divided symmetrically in morphology were functionally asymmetric [
2]. Usually, one daughter cell inherited most or all of the damaging materials such as the condensed chromosome, protein aggregates, and harmful metabolites (e.g., reactive oxygen species, ROS), which decelerated growth rate, decreased offspring production, and increased probability of death [
3]. These damages to cells could be produced under external stress as well as normal cellular physiological conditions [
3,
4], and would be kept in the mother cell in asymmetric division (
S. cerevisiae) or be inherited by only one of the progeny cells in symmetric division (
E. coli), to minimize damage impacts on descendant [
3,
5].
Microbial cell aging, one of the most fundamental but largely unexplored cellular process, involves progressive changes in morphology and function that disturb cell homeostasis, increase death probability, and shorten lifespan, which can be classified into replicative lifespan (RLS), defined by “the number of daughter cells produced before aging,” and chronological lifespan (CLS), defined as “the length of time that cells in stationary phase remain viable” [
6]. Recent studies have identified genes associated with CLS and RLS in
S. cerevisiae and
E. coli. Modulating CLS and RLS to regulate the physiological state of cells, including cell size, morphology, vitality, and stress tolerance, has recently been demonstrated as an effective strategy to increase chemicals (metabolites) production in
E. coli [
6] and wine making by yeast [
7]. Since aging is associated with oxidative stress, manipulating genes associated with stress response pathways can effectively modulate microbial lifespan and increase cell viability and metabolite production.
3. Controlling Microbial Lifespan Can Greatly Improve Fermentation Performance
The microbial lifespan or aging would have major impacts on fermentation process performance metrics—product titer, production rate and product yield (TRY), especially for continuous fermentation with recycled or immobilized cells for extended operation at high cell density and productivity. As examples, we will briefly discuss the importance of engineering the lifespan-associated genes in several non-model microorganisms—
Propionibacterium acidipropionici,
Aureobasidium pullulans, and
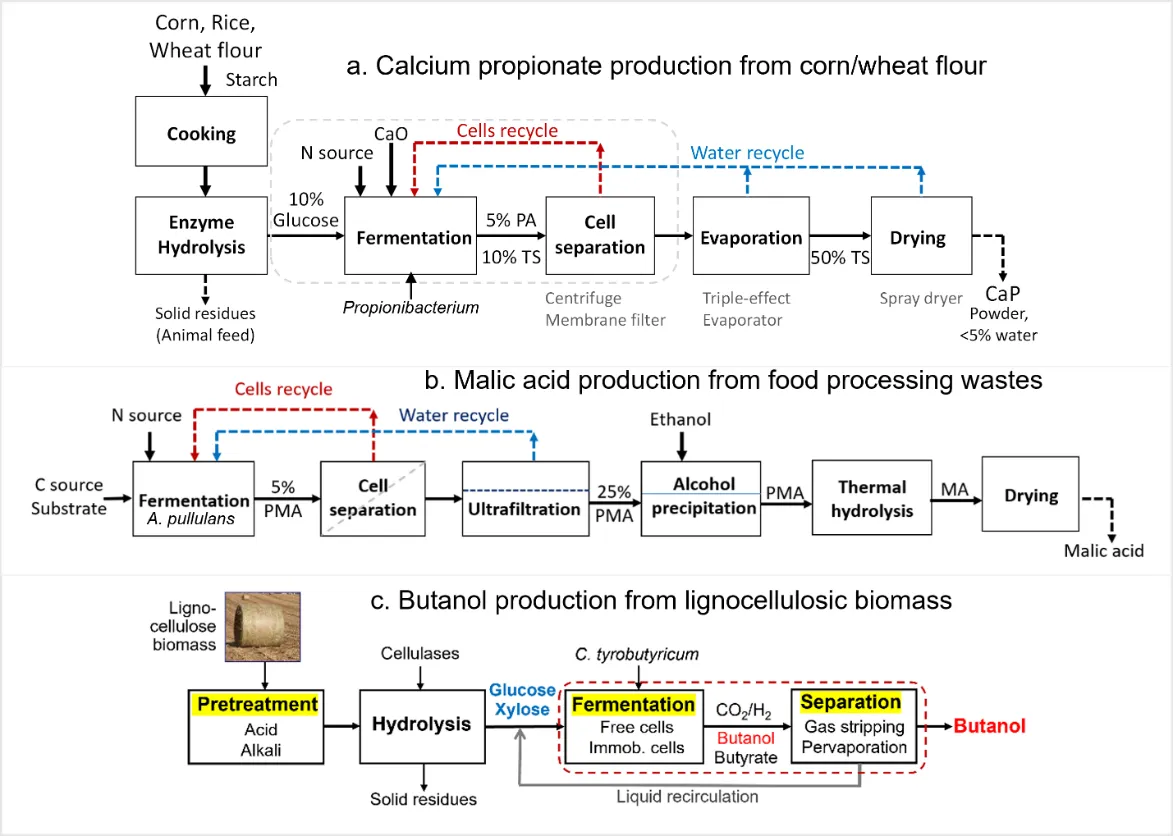
Clostridium acetobutylicum, for chemicals and fuels production from industrial wastes and lignocellulosic biomass, the most abundant renewable organic resource available on earth. shows three bioprocess flowsheets for the production of propionate [
8], malic acid [
9], and butanol [
10,
11] from corn/wheat flour, food processing wastes (e.g., soy molasses), and lignocellulosic biomass (e.g., corn stover) by
P. acidipropionici,
A. pullulans, and
Clostridium tyrobutyricum, respectively, that are currently under scale up development for commercial applications.
. Process flowsheets for calcium propionate, malic acid, and butanol production from biobased feedstocks.
One common feature in these bioprocesses is using cell recycling or immobilization to achieve high cell density in (semi-) continuous fermentation to increase volumetric productivity by 5- to 10-fold [
12,
13]. Such process intensification can greatly improve process economics by reducing both capital (CPEX) and operating expenses (OPEX) and make biomanufacturing economically attractive to compete with chemical synthesis currently used in the petroleum refinery industry. Process intensification via cell recycling or immobilization to achieve high cell density and high volumetric productivity in continuous or semi-continuous (sequential batch/fed-batch) fermentation requires deep understanding of and controlling microbial lifespan and aging effects on the bioprocess. Understanding microbial lifespan and mitigating aging effects on fermentation performance and culture longevity are thus of parament importance to future industrial fermentation for chemicals production.
Propionic acid (PA) is a specialty chemical widely used in various industries, including uses as preservatives in grains and food products and in manufacturing cellulose acetate propionate [
14]. PA fermentation usually suffers from low TRY due to strong PA inhibition and coproduction of acetate and succinate, making biobased PA difficult to compete with its counterpart produced in petrochemical processes. Increasing fermentation TRY by elevating viable cell density and removing PA
in situ to alleviate product inhibition has thus been the focus of recent research [
15]. To achieve high density of active cells in fermentation, cell recycling, retention, and/or immobilization have been proven to be efficient. For example, cell retention using an
in-situ spin filter or cell recycling via an external ultrafiltration unit have been used to increase cell density and reactor productivity in continuous fermentation processes, achieving a high productivity of 14.3 g/L∙h [
16]. However, continuous PA fermentation usually suffers from poor process stability with declining productivity due to cell aging and degeneration and low product concentrations, which greatly increases product recovery costs. The high CPEX and OPEX for continuous cell recycling or retention with membrane filtration limit their application. On the other hand, repeated or sequential batch (SB) with cell recycling by centrifugation is relatively simple to operate for high cell density (HCD) fermentation, achieving high final PA titer 25–50 g/L, yield 0.4–0.6 g/g and productivity 0.3–1.4 g/L·h [
17]. PA fermentation operated in sequential batch with cell recycling via centrifugation from our earlier lab-scale study showed greatly increased PA production in TRY due to increased viable cell density [
12]. However, when we scaled up the fed-batch PA fermentation to industrial scale (120,000 L), the fermentation time more than doubled although PA yield and titer could be maintained (unpublished data). To reduce the fermentation time and increase PA productivity in the HCD fermentation process, it is necessary to understand and control cell viability and lifespan and aging in the process.
Polymalic acid (PMA) is a hydrophilic biopolymer with potential applications as a drug carrier [
18]. Its monomer malic acid (MA) is a dicarboxylic acid widely used in food and beverage industries and can be readily converted to maleic anhydride, a platform chemical. Malic acid production via PMA fermentation by the black yeast
A. pullulans offers a novel and economically attractive solution to the disposal (utilization) of currently underutilized food processing wastes such as soy molasses [
9]. Despite extensive research and development efforts, the bioproduction of MA by MA-overproducing microbes has not been commercialized because of the high energy cost and difficulty in separating MA from citric, fumaric, and succinic acids co-produced in the fermentation [
19]. We have developed an economical and sustainable fermentation process for production of PMA and MA from low-cost lignocellulosic biomass (e.g., soybean hull) and food processing wastes (e.g., soy molasses) [
9]. PMA produced as an extracellular polyester (
Mw: 10–200 kDa) by the black yeast can be concentrated via ultrafiltration and separated with alcohol precipitation. This process can avoid contamination by other acids and thus offers an advantageous alternative to MA fermentation [
19]. Recently, we have engineered A. pullulans to overexpress heterologous pyruvate carboxylase (PYC) and malate dehydrogenase (MDH). The engineered A. pullulans produced PMA at a high titer of 96.7 g/L, high volumetric productivity of 0.90 g/L∙h, and high product yield of 0.68 g/g glucose in fed-batch fermentation [
20]. In SB fermentations with cell recycling, cell density and productivity increased with subsequent batches. Also, PMA production was decoupled from cell growth and continued in the stationary phase, which resulted in higher product yield since little carbon source would be used for cell biomass. However, further feeding glucose showed dramatically slowed PMA production probably due to aging. It is thus of interest to find out if and how CLS can be modulated in
A. aureobasidium to extend the production duration for the sequential batch fermentation process.
Butanol, a widely used industrial solvent, can be used as an advanced biofuel with superior fuel properties compared to ethanol [
21]. Butanol can also be catalytically dehydrated to butene and upgraded to C8–C12 hydrocarbon for use as sustainable aviation fuel (SAF). Industrial production of
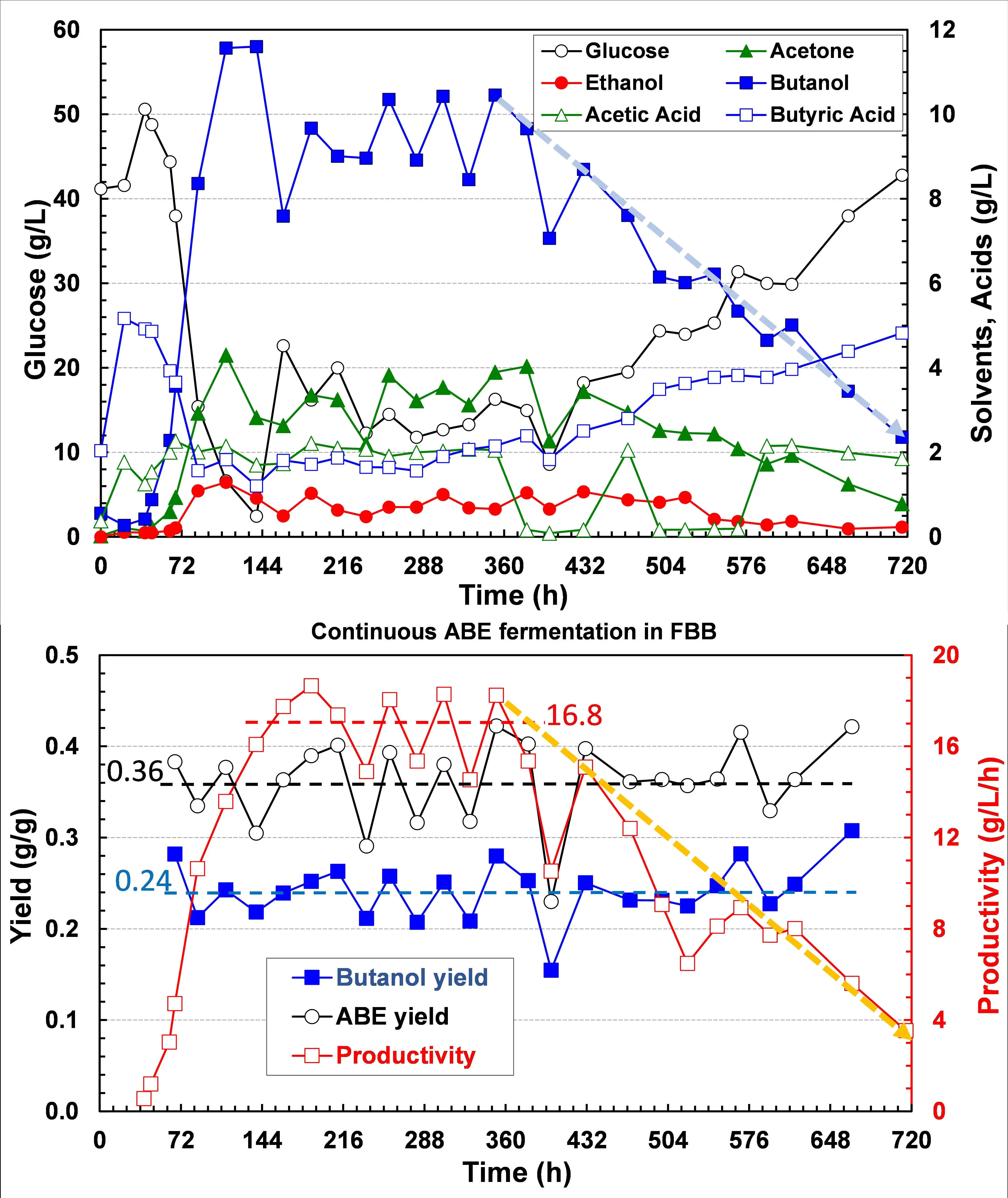
N-butanol by acetone-butanol-ethanol (ABE) fermentation and isobutanol by genetically engineered yeasts from corn starch or dextrose is limited by their low yield, productivity, and high feedstock costs. Consequently, biobutanol produced at a cost of ~$4.5/gal by existing fermentation processes is too expensive for fuel application and not competitive with petroleum-derived butanol even in the higher-value chemical market. To reduce the biobutanol production cost, we must increase butanol yield to >0.3 g/g, titer to >18 g/L, and productivity to >2 g/L·h. We developed a continuous butanol fermentation process with asporogenous C. acetobutylicum ATCC 55025 immobilized in a fibrous bed bioreactor (FBB), which gave a high cell density and productivity of ~16.8 g/L·h, which was 50-fold higher compared to conventional batchwise ABE fermentation [
13]. However, the continuous fermentation with
C. acetobutylicum showed instability and butanol production declined continuously after 400 h (see ) due to sporulation and degeneration or aging. Under stress, solventogenic clostridia sporulate and halt metabolism (thus lost solvent production). We could reprogram clostridia with histidine kinase (HK, Cac3319) knockout to eliminate sporulation and maintain solvent production [
22]. However, we also observed productivity loss over time in sequential batch fermentation with the HK knockout strain of
C. acetobutylicum and the rarely sporulating
C. tyrobutyricum engineered for butanol production [
23,
24]. It is clear that sporulation is not the only cause for productivity loss in clostridia fermentation. We hypothesize that aging is a critical cellular process controlling clostridia lifespan under both stress and normal culture conditions. It is likely that both sporulation and aging are regulated by interacting genetic networks that have never been explored before. To unlock the full potential of
Clostridium for use in industrial fermentation, it will require deep understanding of clostridia life cycle including growth, aging, and sporulation at the systems level [
25,
26,
27].
In summary, engineering genes modulating lifespan/aging in microbes is important for industrial fermentation with high productivity, stability and longevity suitable for biomanufacturing. Currently, PA, MA, and butanol are mainly produced from petroleum resources via catalytic reactions. The clean energy goal, high oil prices, and consumer demands for biobased natural chemical products have drawn a great interest towards producing these chemicals from renewable biomass feedstocks. To develop novel integrated bioprocesses with engineered microbes empowered by a deep understanding of microbial lifespan or longevity as affected by culture conditions and its impacts on fermentation performance is thus of a high priority in circular bioeconomy. To achieve the goal, synthetic biology tools can and must be used effectively to design and construct novel microorganisms, including bacteria and yeasts, with modulated lifespan for desirable outcomes [
28].
. Continuous ABE fermentation with C. acetobutylicum ATCC55025 immobilized in FBB fed with medium containing 60 g/L glucose and 5 g/L butyric acid at the dilution rate of 1.88 h<sup>−1</sup>. Butanol production declined continuously after ~400 h due to culture degeneration or cell aging with reduced vitality (productivity).
Acknowledgments
Financial supports from the National Science Foundation (Award Number 2328215), US Department of Energy ARPA-E (DE-AR0001512), US Department of Agriculture—NIFA (2020-67021-31142), and the Department of Energy—EERE (DE-EE0010300) are acknowledged. The viewpoints presented in this perspective article does not represent the opinion or position of the US government or founding agencies.
Author Contributions
Conceptualization, S.-T.Y.; Writing—Original Draft Preparation, S.-T.Y., G.W., and Z.Q.; Writing—Review & Editing, S.-T.Y.; Visualization, S.-T.Y. and G.W.; Project Administration, S.-T.Y.; Funding Acquisition, S.-T.Y.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This research was funded by the National Science Foundation (grant number 2328215), US Department of Energy ARPA-E (DE-AR0001512), US Department of Agriculture—NIFA (2020-67021-31142), and the Department of Energy—EERE (DE-EE0010300).
Declaration of Competing Interest
The authors declare no competing financial interest.
References
-
1.
Ackermann M, Stearns SC, Jenal U. Senescence in a bacterium with asymmetric division.
Science 2003,
300, 1920.
[Google Scholar]
-
2.
Stewart EJ, Madden R, Paul G, Taddei F. Aging and death in an organism that reproduces by morphologically symmetric division.
PLoS Biol. 2005,
3, e45.
[Google Scholar]
-
3.
Ksiazek K. Bacterial aging: From mechanistic basis to evolutionary perspective.
Cell. Mol. Life Sci. 2010,
67, 3131–3137.
[Google Scholar]
-
4.
Campisi J, d’Adda di Fagagna F. Cellular senescence: When bad things happen to good cells.
Nat. Rev. Mol. Cell Biol. 2007,
8, 729–740.
[Google Scholar]
-
5.
Hill SM, Hao X, Liu B, Nystrom T. Life-span extension by a metacaspase in the yeast
Saccharomyces cerevisiae.
Science 2014,
344, 1389–1392.
[Google Scholar]
-
6.
Guo L, Diao W, Gao C, Hu G, Ding Q, Ye C, et al. Engineering
Escherichia coli lifespan for enhancing chemical production.
Nat. Catal. 2020,
3, 307–318.
[Google Scholar]
-
7.
Orozco H, Matallana E, Aranda A. Genetic manipulation of longevity-related genes as a tool to regulate yeast life span and metabolite production during winemaking.
Microb. Cell Fact. 2013,
12, 1.
[Google Scholar]
-
8.
Yang H, Wang Z, Lin M, Yang ST. Propionic acid production from soy molasses by
Propionibacterium acidipropionici: Fermentation kinetics and economic analysis.
Bioresour. Technol. 2018,
250, 1–9.
[Google Scholar]
-
9.
Cheng C, Zhou Y, Lin M, Wei P, Yang ST. Polymalic acid fermentation by
Aureobasidium pullulans for malic acid production from soybean hull and soy molasses: Fermentation kinetics and economic analysis.
Bioresour. Technol. 2017,
223, 166–174.
[Google Scholar]
-
10.
Li J, Du Y, Bao T, Dong J, Tang IC, Shim H, et al.
N-Butanol production from lignocellulosic biomass hydrolysates without detoxification by
Clostridium tyrobutyricum Δack-adhE2 in a fibrous-bed bioreactor,
Bioresour. Technol. 2019,
289, 121749.
[Google Scholar]
-
11.
Lu C, Zhao J, Yang ST, Wei D. Fed-batch fermentation for
N-butanol production from cassava bagasse hydrolysate in a fibrous bed bioreactor with continuous gas stripping.
Bioresour. Technol. 2012,
104, 380–387.
[Google Scholar]
-
12.
Wang Z, Jin Y, Yang ST. High cell density propionic acid fermentation with an acid tolerant strain of
Propionibacterium acidipropionici.
Biotechnol. Bioeng. 2015,
112, 502–511.
[Google Scholar]
-
13.
Chang WL, Hou W, Xu M, Yang ST. High-rate continuous
N-butanol production by
Clostridium acetobutylicum from glucose and butyric acid in a single-pass fibrous-bed bioreactor.
Biotechnol. Bioeng. 2022,
119, 3474–3486.
[Google Scholar]
-
14.
Wang Z, Sun J, Zhang A, Yang ST. Propionic acid fermentation.
In Bioprocessing Technologies in
Biorefinery for Sustainable Production of Fuels, Chemicals, and Polymers.
John Wiley & Sons, Hoboken, NJ, USA; 2013; pp. 331–349.
-
15.
Yang ST,
Huang H, Tay A, Qin W, De Guzman L, San Nicolas EC. Extractive fermentation for
the production of carboxylic acids. In Bioprocessing
for Value-Added Products from Renewable Resources; Yang S.T. Ed.; Elsevier,
Amsterdam, Netherlands; 2006; pp. 421–446.
-
16.
Boyaval P, Corre C. Continuous fermentation of sweet whey permeate for propionic acid production in a CSTR with UF recycle.
Biotechnol. Lett. 1987,
9, 801–806.
[Google Scholar]
-
17.
Dishisha T, Ståhl A, Lundmark S, Hatti-Kaul R. An economical biorefinery process for propionic acid production from glycerol and potato juice using high cell density fermentation.
Bioresour. Technol. 2013,
135, 504–512.
[Google Scholar]
-
18.
Zou X, Cheng C, Feng J, Song X, Lin M, Yang ST. Biosynthesis of polymalic acid in fermentation: Advances and prospects for industrial application.
Crit. Rev. Biotechnol. 2019,
39, 408–421.
[Google Scholar]
-
19.
Zou X, Zhou Y, Yang ST. Production of polymalic acid and malic acid by
Aureobasidium pullulans fermentation and acid hydrolysis.
Biotechnol. Bioeng. 2013,
110, 2105–2113.
[Google Scholar]
-
20.
Qin Z, Feng J, Li Y, Zheng Y, Moore C, Yang ST. Engineering the reductive tricarboxylic acid pathway in Aureobasidium pullulans for enhanced biosynthesis of poly-L-malic acid.
Bioresour. Technol. 2024,
393, 130122.
[Google Scholar]
-
21.
Zhao J, Lu
C, Chen CC, Yang ST. Biological production of butanol and higher alcohols, in Bioprocessing Technologies in Biorefinery
for Sustainable Production of Fuels, Chemicals, and Polymers; Yang ST,
El-Enshasy HA, Thongchul N. Eds.; Wiley, Hoboken, NJ, USA; 2013; pp. 235–261.
-
22.
Du G, Zhu C, Xu M, Yang ST, Xue C. Energy-efficient butanol production by
Clostridium acetobutylicum with histidine kinase knockouts to improve strain tolerance and process robustness,
Green Chem. 2021,
23, 2155–2168.
[Google Scholar]
-
23.
Yu M, Zhang Y, Tang IC, Yang ST. Metabolic engineering of
Clostridium tyrobutyricum for
N-butanol production.
Metab. Eng. 2011,
13, 373–382.
[Google Scholar]
-
24.
Xu M, Zhao J, Yu L, Tang IC, Xue C, Yang ST. Engineering
Clostridium acetobutylicum with a histidine kinase knockout for enhanced
N-butanol tolerance and production.
Appl. Microbiol. Biotechnol. 2015,
99, 1011–1022.
[Google Scholar]
-
25.
Yoo M, Soucaille P. Trends in systems biology for the analysis and engineering of
Clostridium acetobutylicum metabolism.
Trends Microbiol. 2020,
28, 118–140.
[Google Scholar]
-
26.
Li S, Huang L, Ke C, Pang Z, Liu L. Pathway dissection, regulation, engineering and application: Lessons learned from biobutanol production by solventogenic clostridia.
Biotechnol. Biofuels 2020,
13, 39.
[Google Scholar]
-
27.
Patakova P, Branska B, Vasylkivska M, Jureckova K, Musilova J, Provaznik I, et al. Transcriptomic studies of solventogenic clostridia,
Clostridium acetobutylicum and
Clostridium beijerinckii.
Biotechnol. Adv. 2022,
58, 107889.
[Google Scholar]
-
28.
Hanczyc, M. Engineering life: A review of synthetic biology.
Artificial Life 2020,
26, 260–273.
[Google Scholar]