1. Introduction
Multiplex genome editing (MGE) enables the simultaneous modification of multiple genomic loci within a single experiment, greatly expanding the scope of genetic engineering beyond single loci modifications. This is a powerful approach for applications such as studying gene networks, disease modelling, reconstructing natural biosynthetic pathways, and engineering complex traits in diverse organisms [
1]. Among the various tools developed for MGE, the CRISPR (clustered regularly interspaced short palindromic repeats)/Cas (CRISPR-associated protein) system, first characterized as an adaptive immune mechanism in bacteria and archaea, has emerged as the most versatile genome engineering platform [
2]. The main components of the system are RNA guided endonuclease (Cas9) and small crRNA (also referred to as guide RNA (gRNA)). To achieve genome editing, Cas effector and crRNA complex binds to the target site in sequence dependent manner guided by crRNA sequence. Target recognition requires the presence of a protospacer adjacent motif (PAM) flanking the target sequence, which enables the Cas effector nuclease to generate double-stranded breaks (DSBs). Consequently, DSBs are repaired by endogenous cellular repair pathways, either non-homologous end joining (NHEJ) or homologous directed repair (HDR) pathways. The repair via error prone NHEJ pathway introduces small insertions/deletions/frame shift mutations/stop codons at the cut site. Alternatively, engagement of the HDR pathway requires exogenous or endogenous supply of homologous DNA repair template, which results in precise modifications such as large deletions and insertions, and replacement of faulty genes [
3]. The selection of a pathway for the repair of DSBs depends on many factors, such as resected DNA ends at the break point, cell cycle stage, and availability of homologous donor DNA. The NHEJ pathway operates as a relatively straightforward repair mechanism throughout the cell cycle in various cell types. NHEJ does not require extensive end resection, as the DNA ends are protected by Ku70-Ku80 protein binding. In contrast, HDR represents a more precise but complex repair pathway. HDR requires the MRN complex and CtIP proteins to facilitate extensive 5′ to 3′ end resection of duplex regions, generating stretches of single-strand DNA at the break sites. The HDR process occurs predominantly during the S/G2 phases of mitotic and meiotic cell cycles, which limits the time window for gene editing experiments and thus affects efficiency. Additionally, HDR occurs at significantly lower frequencies than NHEJ, necessitating extensive screening procedures that severely constrain MGE applications [
4]. Despite these limitations, HDR-mediated editing offers better precision compared to NHEJ. NHEJ-based DSB repair can be cytotoxic in many prokaryotic and eukaryotic systems, particularly when multiple DSBs are created simultaneously during MGE. Consequently, researchers are actively pursuing methods to enhance HDR efficiency for improved genome editing outcomes. The most common strategies employed for boosting HDR efficiency include NHEJ pathway inhibition [
5], HDR pathway activation [
6], and donor DNA template modification [
7]. Over the last decade, thousands of studies repurposed CRISPR/Cas technologies to edit or regulate individual genetic loci to achieve different outcomes. However, there is a growing interest in exploiting these tools for editing multiple genes at once to answer complex biological questions. Native CRISPR/Cas systems naturally encode one or more CRISPR arrays, making them inherently capable of MGE. When multiple crRNAs alongside Cas proteins are expressed, the system enables efficient multi locus editing, significantly broadening the range and power of genome engineering applications [
8]. Before the emergence of CRISPR-based technologies, genome editing primarily relied on methods such as homologous recombination (HR), MAGE (Multiplex Automated Genome Engineering), site-specific recombinases, and earlier nuclease-based platforms like TALENs (Transcription Activator-Like Effector Nuclease) and zinc-finger nucleases (ZFNs). These approaches enabled remarkable advances in synthetic biology and biotechnology. However, each method presented distinct limitations affecting flexibility, efficiency, and scalability. HR-based methods require a supply of exogenous donor template with large homology arms (often 500–2000 bp) to incorporate targeted modifications, insertions, or deletions, utilizing host cell native recombination machinery. The efficiency of HR is influenced by the length and the design of homologous arms, which must be tailored to the specific target site and the host system in use. These factors made this strategy inefficient, labor-intensive, and less adaptable for high-throughput or multiplex applications [
9]. Another MGE approach, MAGE, employs pools of synthetic single-stranded oligonucleotides to introduce multiple simultaneous mutations across the genome. This strategy allows rapid prototyping of genetic variants and combinatorial genome engineering, and it has been highly optimized in
Escherichia coli [
10]. To achieve higher efficiency in MAGE, host mismatch repair pathways (MMR) proteins (MutS or MutL) are often suppressed or knocked out to prevent excision of newly incorporated mutations. As a consequence, it leads to an increase in background mutations because the cell loses its ability to efficiently recognize and repair mismatches that arise naturally during DNA replication. This results in unintended genome “scarring” and elevated background mutagenesis [
11]. This further leads to compromised stability and integrity of engineered strains. Moreover, its utility is largely confined to bacterial systems, with limited applications in eukaryotes, primarily in yeast, where multiple DNA replication forks were precisely edited [
12]. Another hybrid approach, CRMAGE has combined the λ Red recombineering based MAGE technology with CRISPR/Cas9 to create a highly efficient and fast genome engineering method, demonstrated in
E. coli. While CRMAGE significantly enhances genome editing efficiency and capability, challenges such as restricted host range and system complexity restrict its widespread applications [
13]. Based on prior development of MAGE technology, a generalized broad-host vector was developed which expresses all necessary components of MAGE and the dominant negative mutant protein MutL from the
E. coli MMR system under a temperature controlled promoter (pORTMAGE). This workflow efficiently modifies multiple loci without prior modification in the host genome and any detectable off-target mutagenesis in
E. coli and Salmonella enterica [
14]. Further, site-specific recombinases (SSRs) such as Cre-loxP and FLP-FRT were engineered to edit pre-defined target sites in the genome, which offer precise integration, excision, or inversion of DNA sequences marked by recombinase specific recognition sequences. SSRs represent foundational genome editing tools widely used across multiple model organisms, including mice, mammalian systems,
Drosophila, zebrafish, and
C. elegans. They enable the generation of conditional knockouts, precise DNA modifications, and lineage tracing in neurons and other tissues. These SSR-based approaches are host-independent, allowing effective and highly specific editing of large DNA fragments, making them invaluable for developmental biology and functional genomics studies. Nonetheless, their utility is constrained by strict need of engineered target sites, difficulty in repurposing, inefficient multiplexing, possibility of genomic rearrangements, and cellular toxicity. Furthermore, the requirement for pre-existing recognition sequences (loxP, FRT sites) restricts their broader applicability beyond model organisms or engineered cell lines that have been pre-modified to contain target sites [
15,
16]. Unlike the previous examples, sequence-independent recombinases that catalyze DNA rearrangements without the presence of strict recognition sites have been reported very recently. For example, IS110 family transposases use engineered bridge RNAs to guide targeted scarless insertions, deletions, or inversions at diverse genome locations [
17]. A bridge RNA has two internal loops, enabling bispecific binding: one loop base-pairs with the target DNA site, while the other pairs with the IS element. This interaction allows the recombinase to perform scar-less genome editing by forming a synaptic complex along with direct base-pairing, bypassing complex protein–DNA recognition mechanisms [
18]. Very recently, transposases of IS110 family and its close relatives, such as IS1111 family [
19] and IS110/IS492 family [
20] have been characterized leading to expansion of their genomic target range. This method enables precise and scarless genome editing at user-selected sites, without the introduction of DSBs, offering new possibilities beyond existing CRISPR-based technologies. This technology represents a conceptual shift in genome engineering and synthetic biology strategy, especially in difficult to modify organisms. The major innovation in the field is brought by programmable nucleases, zinc finger nucleases (ZFNs), and transcription activator-like effector nucleases (TALENs), which induce DSBs at specific sites recognized by their domain sequence. These nucleases employ engineered DNA-binding protein domains fused to the catalytic domain of the FokI endonuclease to create DSBs at specific genomic sites. Designing separate protein–DNA recognition modules for each target is labor-intensive and requires extensive protein engineering, making simultaneous modification of multiple loci technically challenging and costly [
21]. These constraints limited their applications, particularly in the context of MGE, and underscored the need for more robust and versatile genome editing tools. Endonucleases belonging to CRISPR/Cas systems employ short length RNAs (crRNAs or gRNAs) to target virtually any complementary DNA sequence through Watson-Crick base pairing and enable precise and programmable targeting [
22]. The ease of retargeting by altering the gRNA sequence and its efficiency in both prokaryotic and eukaryotic systems has made CRISPR the mainstream tool for multiplexed genome engineering. Over the years, CRISPR tools have evolved and expanded to include an array of applications. These applications include deletions, knock-outs, knock-downs, knock-ins, mutations, RNA editing, epigenetic modifications, base conversions, and gene activation and repression [
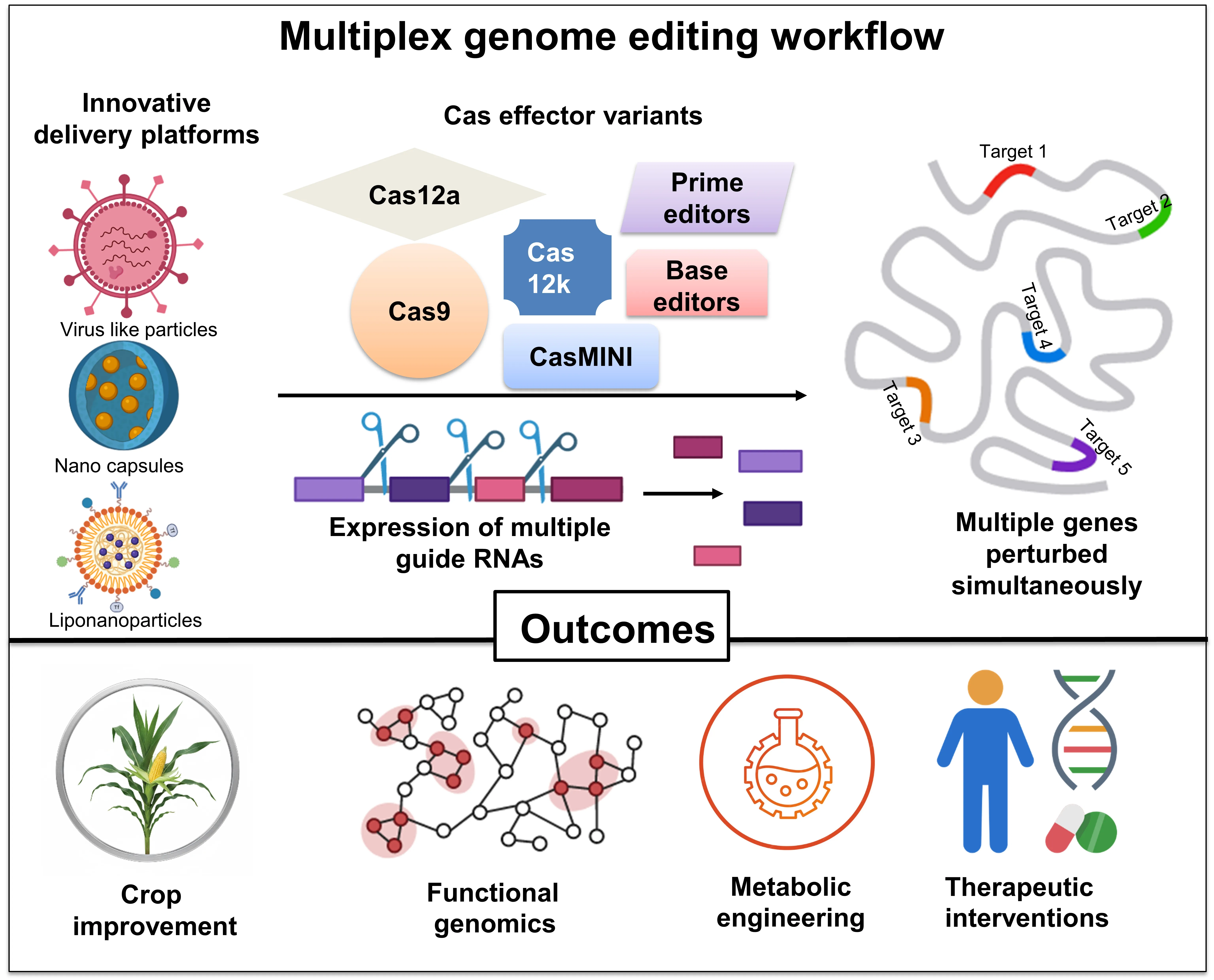
23]. This expansion of capabilities is attributed to extensive reengineering of well-characterized Cas proteins and the ongoing discovery of orthologous systems with complementary features. The concerted efforts were made on reducing the size of Cas effectors, fusing Cas proteins with other functional domains (such as transcriptional regulation or base editing), and engineering crRNAs for improved precision and versatility. Additionally, attempts have been made to increase the fidelity of CRISPR/Cas systems to minimize off-target effects and expand their targeting scope by broadening PAM recognition or developing new Cas variants. The application of different CRISPR variants with respect to perturbing multiple loci was fortified further by progress in four interconnected areas (schematically shown in ). First, efficient crRNA expression and processing strategies, including synthetic CRISPR arrays, novel RNA cleavage mechanisms, and chemically modified or truncated guide RNAs, have enabled the simultaneous expression of a large number of unique crRNAs, thereby enhancing target specificity and guide stability. Second, engineering of existing Cas proteins and exploration of compact and orthogonal CRISPR systems, such as Cas12f and Cas12j and phage-derived variants, has broadened PAM compatibility, reduced cargo size, and made available systems with intrinsic multiplexing capabilities (diversity of Cas proteins, their features, and mechanisms are summarized in ). Third, the development of base and prime editors offers unique capabilities that can mitigate DSB-related toxicity and enable complex single nucleotide corrections in a therapeutic context. Finally, the availability of innovative delivery platforms for efficient and cell-specific delivery of editing components in plant and mammalian cells, including lipid nanoparticles, polymeric carriers, and exosome-based systems, has expanded genome editing into challenging cell types and in
in vivo environments. These enabling advances have catalyzed impactful MGE applications in multitude of organisms.
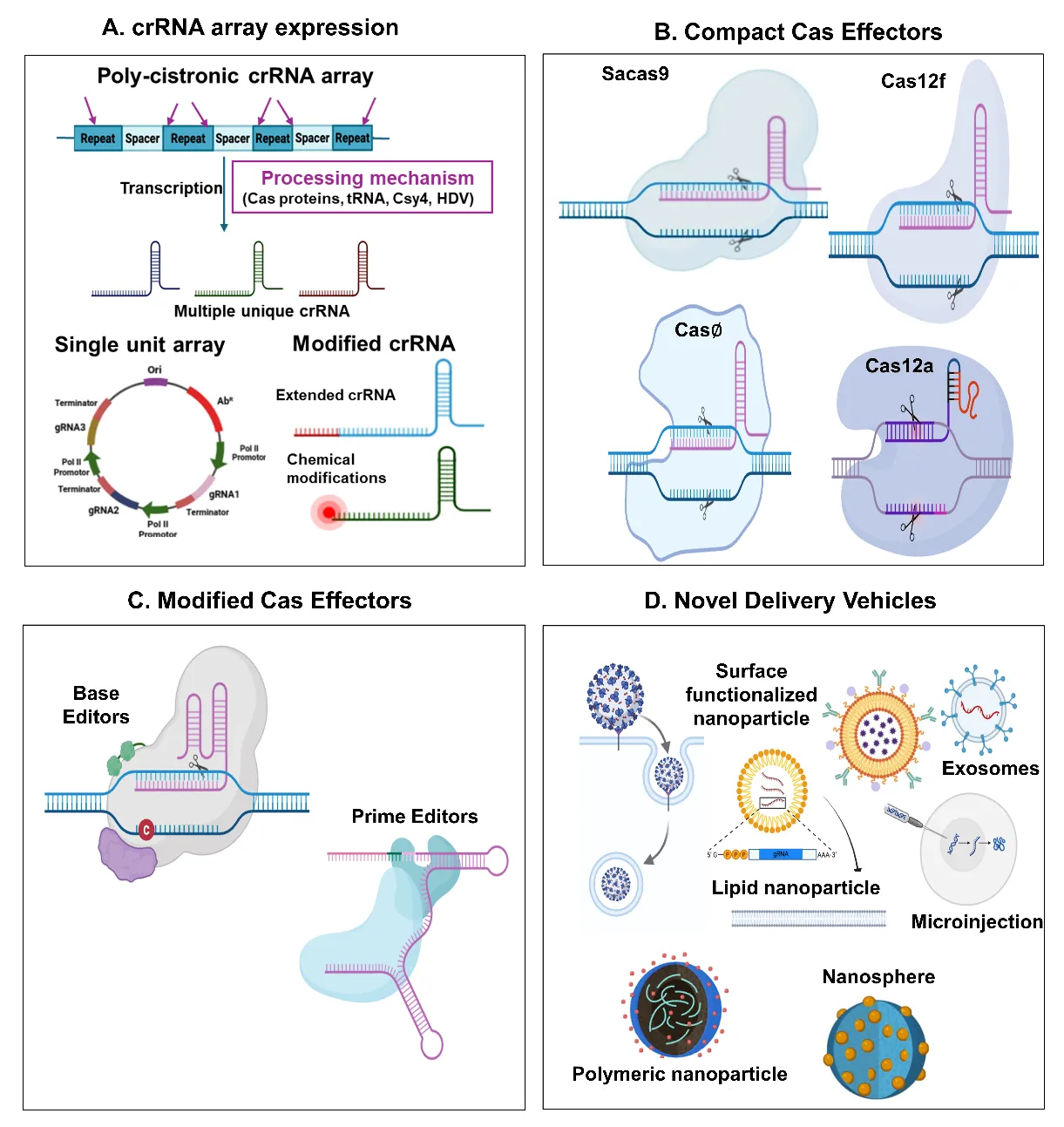
. Snapshot of technological advancements driving multiplex genome editing. (<b>A</b>) Various strategies for crRNA expression, processing, and stability are shown. This includes synthetic polycistronic or single unit arrays driven by RNA polymerase promoters. Pink arrows show processing sites acted by different cleavage mechanisms (Cas protein intrinsic processing activity, tRNA = tRNA sequence between individual crRNA, csy4 = endoribonuclease cleaves stem loop RNA sequence between crRNA, HDV = hepatitis delta virus ribozymes). The figure also depicts modified crRNA, such as extended spacer length or chemically modified guide RNAs (red dots indicate different chemical modifications in crRNA), to enhance stability and function. (<b>B</b>) Compact Cas effectors (e.g., SaCas9, Cas12a, Cas12f, and Casɸ) with unique crRNA processing abilities and PAM requirements are represented. (<b>C</b>) Modified Cas effectors, such as base editors and prime editors that enable precise nucleotide changes without generating double strand-strand breaks and donor templates are highlighted. (<b>D</b>) Novel delivery systems including lipid nanoparticles (LNPs), polymeric nanoparticles, surface functionalized vesicles, nanospheres, and exosomes that facilitate effective and cell-specific delivery of MGE components, are depicted.
Multiplex editing in microbial system has been instrumental in efficient metabolic reprogramming and optimization of the strain. For instance, in
E. coli and
Corynebacterium glutamicum, researchers have employed CRISPR/Cas9 and Cas12a to edit multiple genes regulating central carbon metabolism in a single transformation event, resulting in the construction of high-yielding strains for biofuel and amino acid production [
24,
25]. Similarly, studies in
Saccharomyces cerevisiae have demonstrated multiplexed base editing for pathway rewiring, including the optimization of isobutanol and β-carotene biosynthesis, by targeting up to 10 genes simultaneously using multiplexed gRNA arrays [
26,
27]. In plants, CRISPR-based multiplexing has revolutionized crop breeding by enabling the simultaneous modification of agronomic traits that are polygenic or quantitatively inherited in one generation. Numerous reports have demonstrated multiplex editing to improve yield, abiotic/biotic stress resistance, nutritional quality, and to domesticate crops, particularly in rice, maize, wheat, soya bean, and tomato plants [
28,
29,
30,
31,
32]. In mammalian systems, multiplex editing has provided a new tool for modelling complex diseases in cell lines or mouse models and enabled the design of modalities that are useful for clinical interventions. In cell lines, combinatorial gene knockouts to investigate synthetic lethality by drugs, immune evasion, drug discovery, and antiviral responses have been extensively studied using MGE approaches [
33,
34,
35,
36]. In mice and zebrafish, multiple mutations in tumor suppressors and oncogenes in a single generation have been created to study cancer progression and genetic interactions [
37,
38]. Despite this progress, MGE faces notable challenges, including efficient delivery of large effectors such as SpCas9, limited PAM recognition, variable editing efficiency across multiple targets, and risks of off-target activity or DNA damage responses. These limitations have stimulated the development of smaller Cas variants, engineered high-fidelity nucleases, and alternative systems such as base and prime editors that enable precise modifications without creating DSBs. Complementary innovations in delivery technology and computational guide RNA design further accelerate the practical application of multiplex CRISPR tools.
Recent literature has covered MGE, but with a strong focus on specific areas such as, trait stacking and crop improvement in plants [
39,
40,
41], microbial strain engineering [
42,
43], and the development of advanced CRISPR/Cas effectors like Cas12a and engineered Cas9 variants [
44,
45]. A comprehensive perspective that highlights common challenges and technological convergences across plant, microbes, and animal systems, is lacking. This review aims to address that gap, and enhance our understanding of shared challenges and solutions across different domains of life, thereby offering a unified viewpoint on multiplex genome editing.
.
Summary of the major CRISPR-based systems employed for multiplex genome editing.
| System |
Mechanism |
Variants |
Advantages |
Disadvantages |
References |
| Cas9 |
- Induces blunt ended DSBs,
- Intrinsic crRNA processing is not present in the absence of tracrRNA
- Guided by 20bp sgRNA
-Complex and long repeat sequence
- Canonical NGG PAM
|
SpCas9
SaCas9
eSpCas9 1.1
SpCas9-HF1
HiFi Cas9
zCas9i
SpRYCas9
StCas9
|
High editing activity
well characterized
flexible targeting
wide host range
|
Off-target effects
limited PAM (NGG)
Large Cas protein size
toxicity
reduced efficiency with >4 targets
variable HDR rates
|
[46,47,48,49,50,51] |
| Cas12a (Cpf1) |
- Induces staggered DSBs,
- Capable of processing its crRNA
- shorter repeat sequence
- Canonical TTTV PAMs
|
AsCas12a
LbCas12a
enAsCas12a
ultraAsCas12a
|
Intrinsic multiplexing
T-rich PAM
shorter crRNA
No reported toxicity
Easy to express
|
limited PAM (TTTV)
large Cas protein
variable activity across targets
|
[52,53,54,55,56] |
| Cas12f |
- Compact effector with DSB activity,
- Requires scaffold stabilization and helper proteins
- Highly efficient
|
Un1Cas12f1
CasMINI
enAsCas12f
|
- Ultra compact
- suitable for viral delivery
- diverse PAMs
|
Lower expression
requires intensive engineering and optimization for editing in eukaryotes
|
[56,57,58] |
| Phage-derived Cas variants |
- Diverse cleavage products such as DSBs, nickase, trans cleavage)
- Expanded PAM range
|
CasΦ (Cas12j2)
Cas12k
Casλ
|
Small size
diverse PAMs
high fidelity
Applicable to non-model organisms
|
variable activity
limited characterization in non-model host systems
|
[59,60,61,62] |
| Base Editors (CBEs, ABEs) |
- Cas9 nickase fused to cytidine or adenosine deaminase to mediate single base C→T or A→G without DSBs
- Paired sgRNAs
|
BE3
BE4
ABE7.1
ABE8e
Target-AID
TadCBE
TadDE
|
Precise
Efficient
no DSBs
low indels
|
Limited to specific transitions
bystander/off-target effects
Large size of the fused protein
|
[62,63,64,65,66] |
| Prime Editors (PEs) |
-Combines Cas9 nickase with a reverse transcriptase,
- Prime editing guide RNA (pegRNA) to direct targeted insertions, deletions, and substitutions without DSBs or donor DNA templates.
|
PE1
PE2
PE3
PE5max
twinPE
-PE7
|
Versatile editing (all mutation types)
no DSBs
no requirement for donor DNA template
Less toxicity
|
complex design
lower efficiency
delivery constraints of bulky protein
longer pegRNA
|
[67,68,69,70] |
2. Cas Effectors for Multigene Editing
2.1. Advancement in Cas9 Based Effectors
The CRISPR/Cas9 system, originally derived from the adaptive immune system of
Streptococcus pyogenes, consists of the Cas9 endonuclease which is guided by a synthetic guide RNA (sgRNA), to the target and generates DSBs at specific genomic loci. This system rapidly gained prominence due to its versatility and its adaptability for a wide range of genetic manipulations across diverse organisms. Since its initial deployment for single gene modifications in microbes [
22] and human cell lines [
71]. Cas9-based editing has rapidly evolved to address more complex biological questions. One of the key trajectories was the demonstration of MGE capability in microbes, where simultaneous targeting of two targets in a single experiment with 75% efficiency could be achieved for the first time [
72]. After this demonstration, hundreds of studies explored the multiplexing capabilities in industrially relevant prokaryotic and eukaryotic microorganisms as well as in mammalian cells/tissues. Moreover, Cas9 has been extensively used to reconstruct plant-based natural biosynthetic pathways in yeast, enabling robust functional genomics [
26]. In these multiplexing efforts, different sgRNAs were provided as multiple individual expression cassettes, each transcribed from its own promoter, or as a single polycistronic cassette. In the latter, RNA cleavage sequences are incorporated between individual sgRNA, which get processed post expression, releasing multiple unique crRNAs. The overview of these approaches is discussed in the later part of this review.
The utility of Cas9 has also been extended to multiplexed editing in non-model organisms, for example, deletion of multiple genes involved in carotenoid synthesis (
car2 and
ura3) in
Rhodotorula toruloides, with high efficiency, showcasing the versatility of the Cas9-based multiplexed editing platforms [
73]. The system has also been employed in a diversity of dicot and monocot crop plants for numerous applications such as crop domestication, quality and yield improvement, elimination of anti-nutritional compounds, and abiotic/biotic stress resistance. One such study reported the introduction of 20 mutations in γ- and ω-gliadin genes of wheat to develop a gluten-free wheat variety [
74]. In past years, CRISPR based MGE has become a successful strategy for creating new crop genotypes and to incorporate agriculturally useful traits in a single generation. Beyond its success in plants, multiplexed CRISPR/Cas9 editing has been extended to other cell types and organisms. One notable example is the use of multiplexed editing to generate porcine endogenous retrovirus-inactivated pigs, addressing a major safety concern of transplanting pig organs into humans [
75]. A plethora of studies have reported use of Cas9 for cancer therapy, with most strategies encompasses such as modulation of immune response, repair of mutations, inactivation of oncogenes, target screening, creation of disease models, and drug discovery [
76,
77,
78].
While Cas9-based multiplex genome, editing presents a remarkable opportunity for advancing genetic research, but its broader adoption faces significant challenges. One of the foremost issues is the considerable size of the Cas9 protein (1368 aa residues) itself, which complicates its delivery into cells, particularly for
in vivo editing. Intensive efforts have been made to engineer Cas9 protein or characterize smaller systems from different sources. The well characterized smaller Cas9 systems are identified from
Staphylococcus aureus (SauCas9, 1053 aa residues) [
48] and
Campylobacter jejuni (CjCas9, 984 aa residues) [
79]. While smaller Cas9 variants provide a potential alternative, they are often limited by low efficiency and specificity. Also, most of them recognize elongated PAMs, e.g., SauCas9 NNGRRT (R = A or G) and CjCas9 NNNNRYAC (Y = C or T), which reduces the number of uniquely addressable target sites in the genome. To address these limitations, synthetic RNA-guided nucleases (sRGNs), which are smaller, highly active, specific, and recognize a smaller PAM, were obtained through protein engineering. These systems have demonstrated robust
in vivo editing capabilities in both murine models and non-human primates, independent of their delivery mode, making them well-suited candidates for gene therapy applications [
80]. The other challenge with the system is the inefficient processing of multiple sgRNA simultaneously, as Cas9 does not process its own crRNA. This has been addressed by introducing various artificial processing mechanisms in the crRNA array, which are thoroughly discussed in Section 2. Alternatively, several modifications in sgRNA have been explored; one such report utilized 5′-end truncated sgRNAs to achieve multiplex single-nucleotide-resolution genome editing in
E. coli [
25]. Apart from this, toxicity and off-target effects pose a multi-fold problem in MGE, particularly in eukaryotic genomes. This has been addressed by developing more efficient high fidelity SpCas9 variants such as enhanced specificity Cas9 (1.1) (eSpCas9(1.1)), SpCas9-HF2, and HypaCas9, which have shown reduced off-target effects while maintaining high editing efficiency [
81,
82,
83,
84]. The above-mentioned issues are further exacerbated by the possibility of eliciting a p53 mediated DNA damage response in mammalian cells, where introduction of simultaneous cleavages in DNA leads to cellular senescence or apoptosis [
85]. This issue is addressed by the development of newer CRISPR precision editing technologies, such as base editing and prime editing, that do not involve DSBs (discussed in detail in Section 2.3). Alternative CRISPR types, such as type V Cas12 and its variants with unique and complementary features to Cas9, have been discussed in Section 2.2.
2.2. Advancements in Cas12-Based Effectors
The type V CRISPR/Cas12 family has emerged as a versatile tool for genome editing across prokaryotic and eukaryotic systems. While SpCas9 was the initial focus of genome editing research, there has been a remarkable surge of interest in identifying novel CRISPR systems with complementary features to meet diverse editing outcomes with minimum off targets. The CRISPR/Cas12a system, which was first studied for genome manipulation in the type V family, is composed of a short crRNA and a Cas12a endonuclease protein. Cas12a harbours separate RNase and DNase activity, and can process primary crRNA transcript expressed from a single promoter into multiple crRNAs, making it better suited for multiplexed editing [
52]. Cas12a is relatively smaller in size than the Cas9 system, and it recognizes T-rich PAM instead of G-rich PAM, expanding the range of target sites. A system with capability of autonomously processing its own crRNA arrays, distinct PAM utilization, lower cytotoxicity, a simplified crRNA architecture, reduced off-target effects, and robust activity would be an ideal choice for effective MGE [
86]. Cas12a, particularly the variant derived from
Acidaminococcus sp., has been extensively engineered for genome editing applications, including MGE [
87]. Early advancements in the application of Cas12a-mediated multiplexing have been documented in the model organism
Arabidopsis thaliana and rice [
88,
89], and in human cell lines [
90], exhibiting a range of multiplex editing efficiencies ranging from 50–70%. The unique features of the Cas12 system have encouraged the scientific community to explore its potential for MGE based therapeutic interventions, particularly for diseases involving multiple gene targets. This approach holds promise in treating complex ocular disorders such as age-related macular degeneration, glaucoma, diabetic retinopathy, retinitis pigmentosa, and dry eye syndrome [
91,
92,
93]. The comparatively substantial size of the Cas12a protein (~1200–1300 amino acids), which restricted complex manipulations and constrained packaging for
in vivo delivery, hindered its applications. This spurred efforts to develop compact Cas effectors with enhanced multiplexing capabilities and to design tools with higher editing efficiency and reduced off-target effects [
94,
95]. The various members of the Cas12 family have been explored and optimized for gene editing
in vivo. Cas12a (1200–1500 aa) is the prototypical variant, while smaller orthologues like Cas12m (~600 aa), Cas12n (~500 aa), Cas12c and Cas12f (400–700 aa) have emerged as compact alternatives, broadening the CRISPR genome editing toolkit [
96,
97,
98,
99]. These Cas12 variants differ in size, structural features, PAM recognition, efficiency and cleavage mechanisms [
100]. The published reviews have detailed classification and mechanistic diversity of Cas12 variants. Thus, this review discusses Cas12 variants specifically engineered or adapted for MGE applications across different host systems.
The Cas12f (also known as Cas14,) variants which were smaller in size ranging from 400–700 aa were initially characterized as single stranded nucleases. These variants were engineered to cleave double-stranded DNA in a PAM-dependent manner but showed limited cleavage activity in mammalian systems [
53]. Protein engineering further advanced these to develop compact synthetic variants such as CasMINI (529 aa). CasMINI supports robust gene activation, base editing, and nuclease activity in mammalian cells with high specificity and minimal off-target effects. MGE utilizing CasMINI has demonstrated efficiency comparable to or exceeding that of Cas12a in mammalian systems. It provides superior delivery compatibility due to its small size, even after fusion with different domains [
58]. An additional variant of Cas12f, un1Cas12f1 exhibited successful editing capabilities in organisms such as
E. coli, Bacillus anthracis, and
Klebsiella pneumoniae, as well as in the
in vivo murine models, wherein the editing efficiency at the
hpd locus attained up to 80% when administered through AAV vectors. In
Streptomyces coelicolor, un1Cas12f1 enabled the concurrent deletion of two genes, representing a significant step towards multiplex bacterial genome editing [
101]. In addition to the Cas12f, other Cas12 variants were engineered for higher efficiency and robustness. For instance, the utilization of 3'-truncated gRNAs of Cas12a in conjunction with low-temperature recovery conditions facilitated single-nucleotide precision editing of three distinct loci in
E. coli [
102]. Additionally, FnCas12a derived from
Francisella novicida, in conjunction with non-homologous end joining (NHEJ) proteins sourced from
Mycobacterium tuberculosis, facilitated donor-free gene knockout and cluster deletion within the rice pathogen
Xanthomonas oryzae pv. oryzae (Xoo). MGE alongside plasmid curing was effectively demonstrated in this strain, thereby establishing this system as a highly amenable platform for pathogen functional genomics [
103]. To further enhance efficiency within the human context, the Cas12a-REDIT strategy which combines AsCas12a with ten GCN4 peptides, while simultaneously expressing a RecT recombinase tethered to an ScFv-GCN4 domain, was developed. This configuration effectively recruited RecT to loci of Cas12a activity, thereby augmenting the editing efficiency at four human loci (AAVS1, DYNLT1, EMX1, DNMT1) by ~four-fold. The observed editing efficiencies ranged from 40–70% and were contingent upon the specific locus and cell type [
104]. In plant systems, the Mb3Cas12a effector was optimized for high-throughput multiplexing through incorporation of up to 13 crRNAs within a single transcript in
Arabidopsis thaliana, culminating in efficient editing and stable inheritance of multiple edited alleles with minimal off-target effects, as validated by whole-genome sequencing [
105]. This system was also utilized for simultaneous editing of eight target sites using a single crRNA cassette in the cotton plant, achieving an efficiency of over 90% at almost all target sites. Additionally, the Mb2Cas12a system revealed effective editing at more relaxed PAM VTTV (V = A, C and G) sites in the cotton genome, which expanded the genome editing range by approximately 2.6-fold than the wildtype LbCas12a [
106]. Another variant called CRISPR/Cas12i3, recently attracted attention due to its smaller protein size and less restricted canonical “TTN” PAM. However, due to its relatively lower editing efficiency, Cas12i3 was improved by fusing with 5' exonucleases. The system was used to edit multiple genes in rice with 82.76% efficiency, highlighting the application of miniature CRISPR-Cas proteins for robust genome editing [
107]. Overall, these variants broadened the editing repertoire for crop improvement, predominantly in the context of polygenic trait enhancement, which requires a large number of genes in unrelated pathways to be edited simultaneously with greater efficiency.
The exploration of complementary CRISPR systems was further extended to bacteriophages that included Cas12j2 (CasΦ), a highly compact nuclease which has demonstrated significant editing in rice plant protoplast. This multiplex editing platform achieved an efficiency range of 15–50% across four target loci in rice; however, the rates of stable transformation in differentiated tissues remained suboptimal. Nonetheless, Cas12j systems signify a critical advancement towards the development of highly compact CRISPR effectors [
108]. Another development in the field, is Cas12k, an enzymatically inactive effector linked to CRISPR-associated transposase system (e.g., ShCAST), which has been repurposed into a novel tool designated as C12KGET. In
Sinorhizobium meliloti, this system facilitated efficient large fragment insertions and multiplex editing. Targeting genes within the vitamin B12 biosynthesis pathway led to an enhancement in metabolic output, thereby underscoring the utility of Cas12k in the domain of microbial metabolic engineering [
109]. In a nutshell, several functional advantages, such as intrinsic CRISPR array processing, smaller protein size, simplified PAM preferences, delivery compatibility, and engineering flexibility, drive the interest in miniaturized CRISPR systems and make them better suited for MGE. The progression of the CRISPR-Cas12 system from the earlier Cas12a-based multiplex editors to the compact effectors such as CasMINI, Cas12j2, and Cas12k represents a crucial milestone towards the realization of scalable, multi-gene editing platforms. These systems not only overcome delivery challenges but also offer both architectural and functional enhancements fuelled by parallel advancements in structure-guided design, protein engineering and guide RNA optimization. The compact Cas12 systems with multiplex efficiencies varying from 50–85% across bacterial, plant, and human cell systems, and capabilities that include gene activation, base editing, large DNA insertion, and gene repression, are positioned to accelerate multigene control over intricate biological systems spanning across therapeutic, agricultural, and microbial platforms.
2.3. Emergence of Base and Prime Editors
The advent of base editors (BEs) and prime editors (PEs) has redefined the landscape of genome engineering by enabling precise and predictable base conversions without introducing DSBs and donor templates. Unlike classical CRISPR/Cas9 systems, which rely on the cellular repair pathways, error-prone NHEJ or HDR for modifications, BEs and PEs facilitate programmable single-nucleotide substitutions or small insertions/deletions without DSBs. A seed sequence in crRNA defines the editing site, referred to as the editing window, and these platforms open avenues for a vast range of precise genetic perturbations [
110].
2.3.1. Multiplex Genome Editing with Base Editors
Base editors are chimeric proteins combining catalytically nuclease impaired dCas9 with different nucleotide deaminases, allowing C→T (cytidine base editors, CBEs) or A→G (adenine base editors, ABEs) conversions [
63,
111]. Many human genetic diseases and desirable traits in crops and livestock are caused by multiple distinct point mutations occurring at defined genetic locus. Now, with a multiplexed base editing approach, a spectrum of single nucleotide changes can be edited in mammalian systems and plants. These tools are highly precise, flexible with reduced unintended activity. The simultaneous base conversions (A•T to G•C) at three independent loci in rice were demonstrated using ABE with editing efficiencies of 14–39% [
112,
113]. The multiple edited sites were inherited across generations, resulting in mutant line generation in a short time frame. Similarly, simultaneous base editing at the OsWaxy and OsSPL14 loci using evolved ABEs in rice reduced amylose content and altered plant morphology [
114,
115], apart from several studies which has utilized BEs for improving rice grain yield, abiotic stress tolerance, grain quality, disease, and herbicide resistance [
116,
117,
118,
119]. In mammalian systems, C-to-T and A-to-G edits were introduced in 45 endogenous gene loci of HEK293 cells with minimal off-target effects [
120]. Another study has reported combinatorial
in situ mutagenesis utilizing dCas12a base editor variant (MultiduBE) [
121]. Further, concurrent A-to-G and C-to-T substitutions within a single editing window have been reported, which uses CRISPR/Cas9-based dual-deaminase base editor called SPACE (synchronous programmable adenine and cytosine editor) with minimal off-target effects [
122]. This approach circumvents the need for multiple editor constructs and enhances multiplex fidelity. Over the years, BEs have evolved tremendously, ranging from BE1, BE2, BE3, Target-AID, HF-BE3, TadCBE3, and many more derivatives to improve expression and efficiency in different host ranges. Still, certain limitations are fundamentally tide to them, for instance, they can induce only 4 out of 12 possible base conversions, operate within a narrow editing window limited to a few nucleotides, and depends on strict PAM availability, although this constraint has been partially alleviated through the engineering of near PAM less Cas9 variants. To overcome these challenges, a novel approach known as prime editing has been developed, which utilizes a distinct mechanism to replace sequences of varying lengths.
2.3.2. Multiplexed Editing with Prime Editors
Prime editors (PE) extend the versatility of CRISPR-based editing by enabling targeted insertions, deletions, and all possible 12 base substitutions without requiring DSBs or donor templates. The system comprises a Cas9 nickase fused to a reverse transcriptase (RT) and a prime editing guide RNA (pegRNA). The pegRNA contains a target specific guide RNA, a primer binding sequence (PBS), and a reverse transcription template (RTT) sequence, which encodes the targeted modifications [
68]. The mechanism of prime editing is less sensitive to cellular replication state and endogenous HDR-based DNA repair pathways. Multiplex prime editing has yielded promising results in rice, where edits were successfully introduced at multiple loci, including OsALS and OsEPSPS, thereby conferring herbicide resistance [
123]. However, the editing efficiency at individual loci varied considerably, ranging from 0.2% to 7.5%. This study has demonstrated the feasibility of delivering multiple pegRNAs simultaneously, establishing multiplex prime editing as a viable strategy for enabling precise genome modifications in plant systems [
123]. These findings are consistent with broader developments in rice multiplex prime editing. For instance, a modular assembly-based multiplex prime editing system enabled simultaneous editing of up to four distinct genes, two of them conferring herbicide tolerance (OsALS1 and OsEPSPS1) and the other two conferring resistance to bacterial blight (TFIIAγ5 and OsSWEET11a). In this system, co-editing efficiencies reached up to 43.5% in the T₀ generation for all the four targets [
124]. In general, prime editing components exhibit poor expression in plants, restricting their applicability and also currently limiting its successful implementation in model monocot species such as rice and maize. Ongoing advancements are focused on optimizing pegRNA expression strategies, modifying reverse transcriptase components for enhanced activity, and extending editing windows to broaden applicability across diverse plant species. The original PPE (plant prime editor) was upgraded by introducing a V223A substitution into reverse transcriptase resulting in ePPEplus. This engineered plant prime editor was able to edit four to ten genes in protoplasts and up to eight genes in regenerated wheat plants at frequencies up to 74.5%, thus expanding the applicability of prime editors for stacking of multiple agronomic traits in hexaploid wheat plants [
29]. PE systems have achieved multiplex editing in human cell lines using pegRNA arrays or co-delivery of multiple pegRNAs with orthogonal nicking sgRNAs [
125]. These developments underscore the potential of PEs for combinatorial disease modelling and complex modifications where multiple mutations must be corrected simultaneously. Recent innovations, such as engineered pegRNAs (epegRNAs) with structured RNA motifs, have enhanced prime editing efficiency and fidelity [
126]. Moreover, the emergence of twin prime editors (twinPEs) and all-in-one vectors with self-processing ribozymes or tRNA scaffolds has improved the efficiency of overall MGE [
127,
128].
Both BEs and PEs exhibit high target specificity, reduced off-target activity, and programmable flexibility, making them better alternatives for MGE compared to traditional CRISPR/Cas systems. However, the efficiency of multiplex editing is influenced by factors such as target site accessibility, delivery, and competition among guide RNAs. While base editors are preferred for precise conversions within narrow windows, prime editors offer broader editing potential, albeit with lower efficiency and higher construct complexity. The engineered drive-and-process (DAP) CRISPR array architectures allow multiplex base-editing (MBE) and multiplex prime-editing (MPE) in human cells, enabling the study of complex functional genomics and the treatment of polygenic diseases [
129]. The integration of machine learning tools for pegRNA design, improved delivery systems (e.g., nanoparticles, viral vectors), and compact Cas variants (e.g., Cas12f, CasMINI) promises to enhance the practical applicability of BEs and PEs based MGE.
3. crRNA Expression Strategies and Innovations for Multiplexed Genome Editing
The core of the MGE platforms is the efficient synthesis and processing of multiple crRNAs, which guides corresponding Cas nucleases to targeted genomic regions. The different CRISPR systems (e.g., Cas9, Cas12a, Cas13, or Cas12a2,
etc.), host organisms, and desired editing complexity all influence the ways in which crRNAs are expressed and processed. The expression of crRNA has evolved through various innovative strategies since the first report of CRISPR-based multiplexed gene editing appeared way back in 2013 [
8]. Most of the improvements have focused at enhancing efficiency, specificity, and ease of construction. These strategies encompass the use of different CRISPR systems, use of single and multiple transcriptional units, promoter optimizations/selection, and innovative RNA processing techniques, and facilitate the efficient editing of multiple genomic targets in a wide range of host systems. In this review, we provide an overview of most significant and recent methods utilized for multiple crRNA expression (
).
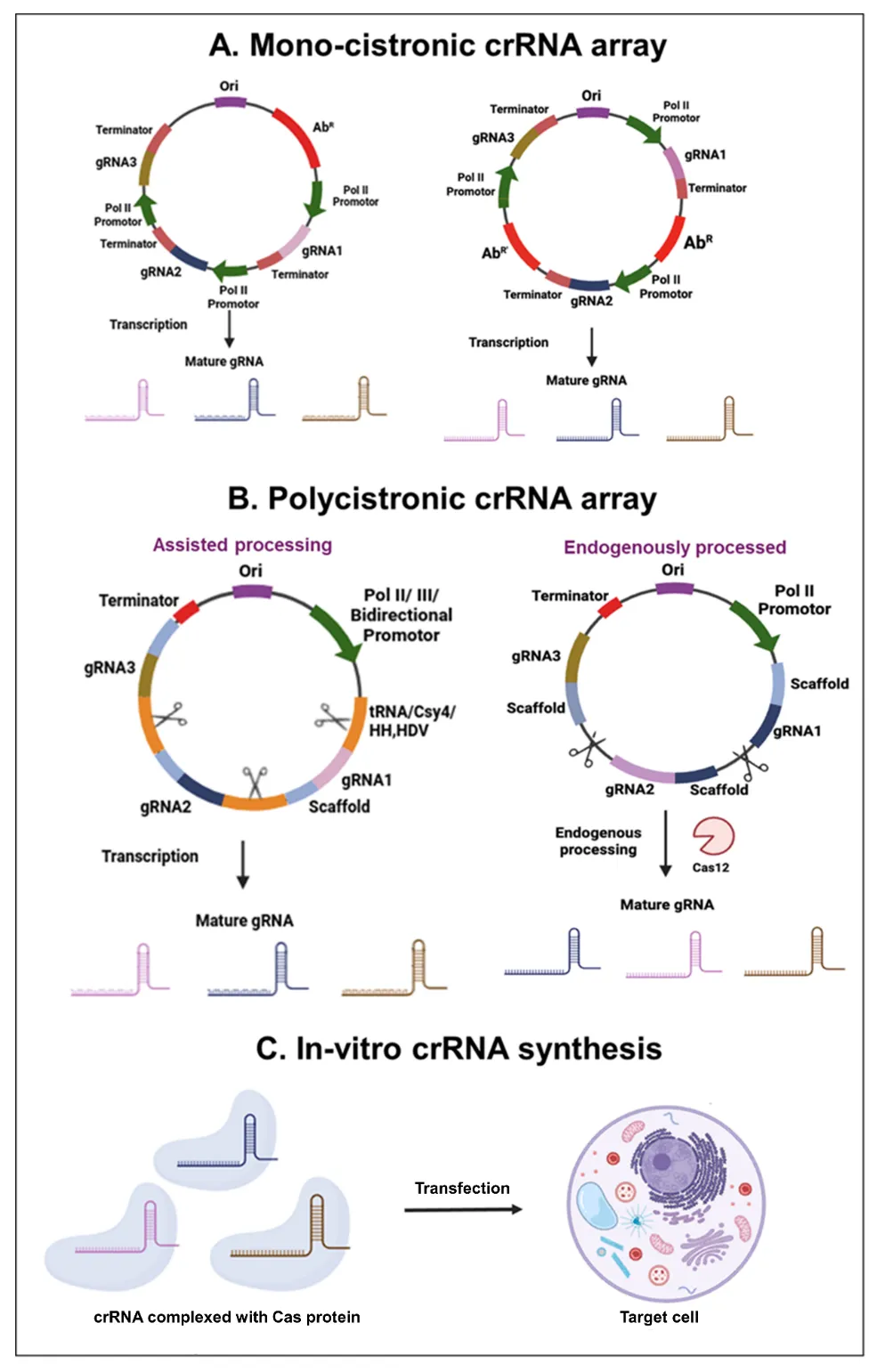
. Strategies for multiplex gRNA expression in genome editing: A mono-cistronic gRNA array, each carrying an individual promoter and terminator arranged in tandem or separated by ori (origin of replication) and antibiotic resistance gene (<b>A</b>). Polycistronic expression array is processed to generate individual crRNA via a native CRISPR processing mechanism or by artificially inserting RNA cleavage sites (tRNA, HDV, siRNA) in a synthetic gRNA array (<b>B</b>). <i>In vitro</i> transcribed mature crRNAs are directly complexed with Cas proteins and injected into target cells as RNP (<b>C</b>).
There are two main strategies for
in vivo expression of multiple crRNAs. The first is to design a mono-cistronic array which consists of multiple gRNAs encoded as a single transcription unit under an individual promoter (typically RNA Pol III promoters, e.g., U6, H1) and terminator. The multiple individual cassettes can be combined into a single vector in a continuous fashion or interspaced by stuffer sequences such as different antibiotic resistance genes to prevent internal recombination of similar genetic elements (
A). The utilization of a single transcriptional unit allows for the streamlined expression of multiple gRNAs with minimum 3’end modifications, reducing the overall complexity of construct designing. The assembly of different crRNA expression units in a single binary vector is facilitated by different molecular biology approaches like Golden Gate Assembly, Gibson assembly, and the use of type IIS restriction enzymes. Consequently, various innovative approaches utilizing these techniques have been developed to efficiently assemble multiple individual expression cassettes within a single vector [
130,
131]. One such approach is the single-step multiplexing technique known as STAgR (String Assembly gRNA Cloning), which stands out as a highly effective method for constructing vectors that can accommodate a diverse array of gRNAs. STAgR leverages N20 targeting sequences as essential homologous regions for Gibson assembly. This system exhibits a high degree of customization, enabling users to selectively choose and mix vector backbones, gRNA configuration, number, and promoters to meet their distinct experimental requirements [
132]. However, all these tools realistically support only a limited number of sgRNA expression cassettes, and the construction of vectors containing ultra-multiplex sgRNA expression cassettes is both difficult and inefficient. Another report advanced crRNA expression by assembling 40 sgRNA in a single binary vector, leading to high co-editing efficiency of multiple traits in rice [
133]. This is particularly useful for dissecting complex polygenic traits like yield, which involves many small-effect genes and redundant regulatory networks. However, these approaches of crRNA array expression commonly suffer from challenges such as repeated usage of the same sets of promoters and terminators that sometimes lead to internal recombination, requirement of multiple selection markers, lengthy constructs, expression burden on host cell, and, more importantly, labour intensive construction of expression cassettes and plasmids.
The second strategy uses polycistronic crRNA architecture, in which multiple crRNA sequences are transcribed as a single RNA using a single promoter (RNA pol II/III or bidirectional) and subsequently processed into individual mature crRNAs [
134] (
B). This is particularly well suited for Class 2 CRISPR systems (e.g., Cas12a/Cpf1), which naturally process precursor crRNAs (pre-crRNAs) via their intrinsic RNase activity [
52]. These arrays can be easily customized using direct repeat–spacer units, enabling compact and scalable multiplexing [
87]. The utility of a polycistronic array is limited for Cas9, as it is unable to process its synthetic crRNA arrays. It thus depends on heterologous RNA cleavage sequences between individual spacer repeat units. This motivated development of artificial processing strategies which involves tRNA based spacers, ribozyme flanking sequences, Csy4-mediated cleavage and intron processing-based systems. In tRNA processing strategy, host tRNA-processing enzymes (e.g., RNase P and RNase Z) are utilized to precisely cleave tRNA–crRNA fusions [
135,
136]. This strategy has proven effective in both plant and mammalian systems, enabling multiplexing of up to 10 targets. Originally demonstrated in rice and maize, this tRNA–gRNA strategy enables multiple gRNAs to be processed efficiently from a single transcript in multiple host systems [
137]. The main advantage lies in its design simplicity and compact vector size. The other modification in crRNA expression was introduced by flanking it with self-cleaving ribozymes such as hammerhead (HH) and hepatitis delta virus (HDV) domain, resulting in autonomous processing of guide RNAs post-transcription [
138,
139]. The self-cleavage activities of ribozymes enable coordinated expression of crRNA without additional poly U sequences at the 3′ terminal, significantly improving gene editing efficiency. However, the decrease in processing activity as the number of gRNAs increases is a potential disadvantage of ribozyme-mediated gene editing [
140]. Another innovative approach involves Csy4 based RNase system from the
Pseudomonas aeruginosa, which specifically recognizes and cleaves at engineered hairpin structures flanking crRNAs, allowing precise processing and reduced crosstalk [
134,
141]. However, this approach requires co-expression of the Csy4 protein, adding to the expression burden and delivery constraints. Alternatively, strategies utilizing endogenous microRNA processing machinery, such as Drosha-DGCR8 complex attempted to reduce the burden of expressing additional components to achieve MGE. In this approach, gRNAs are embedded in miRNA or shRNA scaffolds, and upon their transcription, Drosha cleaves to release functional gRNAs. However, processing efficiency can vary depending on transcript architecture and Drosha specificity, which may limit gRNA design [
142]. As an alternative to
in vivo expression-based methods,
in vitro transcribed crRNAs can be pre-complexed with Cas9 protein to form ribonucleoprotein (RNP) complexes, which are subsequently introduced into mammalian cells or primary cell types by transfection or viral vectors (depicted in
C). This enables transient expression and genome editing while bypassing the need for endogenous processing of crRNAs. For example, targeting PDX1 exon 1 and non-coding DNA into primary human islet cells revealed genetic targets for diabetes regulation [
143,
144].
The further improvement in efficiency of crRNA expression is brought about by optimizing promoter selection, for example, RNA Pol III promoters (e.g., U6, H1) are preferred for short, defined transcripts with minimal 3′ end modifications. In contrast, RNA Pol II promoters are used in conjunction with ribozymes or tRNA-processing motifs for generating mature crRNAs [
48,
139]. For these modified crRNA expression designs, RNA pol II promoters allow tissue-specific and inducible expression, thus overcoming the disadvantage of constitutive expression of CRISPR/Cas components [
145,
146]. Efforts to increase transcript stability include engineering direct repeat sequences, optimizing secondary structures to resist degradation, and incorporating chemical modifications for enhanced nuclear retention. Some of these strategies also contribute to minimizing off target binding. Notably, chemical modification in crRNA structure has proven to be an effective strategy to either stabilize or improve the expression of crRNA. Incorporation of 2-aminoadenine (base Z) in crRNAs was shown to increase the binding affinity between crRNA and its complementary DNA target, thus improving on-target editing efficiency while reducing off-target effects [
147]. In another strategy, crRNA was stabilized by fusing it with U4AU4 3′-overhang and was used to show enhanced editing efficiency by,AsCpf1 [
148]. These advances are further complemented by synthetic biology approaches such as design and synthesis of programmable, modular arrays using standardized linker elements, oligo annealing approaches, Gibson assembly, and Golden Gate cloning methods [
146]. However, crRNAs arrays are difficult to generate due to recurring repeat sequences. A versatile one-pot, modular platform CRATES (CRISPR Array Trimmed-Element Synthesis), was reported for constructing CRISPR arrays and libraries using defined junctions within trimmed spacer regions [
149]. It enabled efficient assembly of arrays for Cas9, Cas12a, and Cas13a, supporting multiplexed DNA/RNA targeting in cell-free systems,
E. coli, and
S. cerevisiae. This method allowed composite arrays to be processed by multiple nucleases simultaneously despite differences in their ability to process crRNA, facilitating array-based co-expression of gRNAs for different systems from a single transcript. Notably, processing efficiency is influenced by terminal repeat processing and crRNA secondary structure, which can affect the efficiency of individual guide RNA [
58,
70]. Further, integration of artificial intelligence and machine learning algorithms in designing gRNAs and optimizing editing strategies is anticipated further to enhance the capabilities of MGE [
150,
151,
152].
In general, editing efficiency is context-dependent and may vary based on target loci and experimental conditions. As genome editing progresses toward broader clinical and agricultural applications, ensuring specificity, and minimizing off-target effects of crRNA are crucial innovations combining synthetic biology with endogenous processing mechanisms that mostly influence future innovations. AI-assisted tools for gRNA design and optimization and synthetic promoters tailored to target species or cell type, will further enhance the efficacy and precision of multiplexed editing systems.
4. AI-Driven Advances in Multiplex Genome Editing
Artificial intelligence (AI) plays a transformative role in several aspects of CRISPR research and applications, including multiplex genome editing. The integration of AI and genome editing has enabled high throughput and precise genome editing approaches to address complex biological problems. Following the successful implementation of CRISPR-Cas9 based genome editing in higher eukaryotes, many laboratories initiated high throughput multiplex editing experiments, leading to accumulation of large-scale experimental data on specific sgRNA activity and validated off targets. At the same time, advancements in machine learning and cloud computing have led to more accurate predictions of on-target and off-target effects, thereby reducing unintended genetic modifications and enhancing the safety of gene-editing approaches. The landmark study in 2014 by Doench et al., reported a predictive machine learning model based on 1841 sgRNAs to improve sgRNA design for gene editing and genetic screens [
153]. Subsequently, CHOPCHOP was launched, which uses efficient sequence alignment algorithms to rigorously predict off-target binding of sgRNAs. In addition, it predicts restriction sites and primer candidates for each possible target site, facilitating a streamlined pipeline of mutant generation and validation. The ease-of-use and speed of CHOPCHOP made it a valuable tool for designing guide RNAs for different CRISPR modalities [
154]. These early models served as a foundational step for selecting target sites and optimizing CRISPR/Cas9 high throughput screens and MGE. Further advancement was brought about by large-scale pre-trained machine learning models. For example, synthetic gRNA-target paired libraries and next-generation sequencing (NGS) were used to develop a powerful machine learning model to evaluate multiple Cas9 variants to predict the optimal variant and its corresponding gRNA targeting human genome sites with minimum off-target effects [
155]. The rapid advancement of diverse CRISPR toolkit modalities has generated a pressing need for specialized computational tools tailored to the unique requirements of each system. For instance, deep transformer-based models like DTMP-Prime have been developed specifically to predict prime editing efficiency by analyzing extensive pegRNA and NGS datasets, thereby extracting effective predictive features [
156]. The widespread adoption of deep learning and reinforcement learning techniques has also enabled sophisticated analysis of large-scale CRISPR screening data, integrating multi-omics information such as transcriptomics, proteomics, and epigenomics to construct predictive models that assess editing performance and risks across varied cellular contexts. AI-driven computational tools now support every step in the genome editing pipeline, from enhancing target selection accuracy and designing highly specific guide RNAs (gRNAs) to precise off-target prediction and big data processing. This has accelerated the discovery of novel CRISPR-associated proteins, facilitated drug discovery, and therapy optimization. Large language models (LLMs), including ProGen2 and OpenCRISPR-1, have been instrumental in generating millions of novel Cas protein sequences predicted to maintain critical functional domains as their natural counterparts while also designing optimized sgRNAs tailored to AI-predicted Cas effectors [
157]. AI-powered models such as DeepCRISPR, CRISTA, and DeepHF analyze comprehensive genomic datasets to predict effective and specific gRNAs considering factors like genomic context, Cas protein type, and editing goals, thereby improving targeting accuracy and minimizing unintended effects [
158]. Furthermore, advanced LLM agent systems like CRISPR-GPT automate and enhance gene-editing design and data analysis by integrating domain expertise, retrieval methods, and external tools to assist experiment planning, gRNA design, delivery method selection, protocol drafting, assay design, and data analysis, effectively serving as AI co-pilots in genome engineering [
159].
5. Delivery Strategies for CRISPR Based Genome Editing: Innovations and Challenges
The development of efficient and targeted delivery systems remains a central challenge in realizing the full potential of multiplexed CRISPR-based genome editing tools. The therapeutic and translational potential of CRISPR/Cas technology is contingent upon the safe and effective delivery of various editing components. The cargo can be plasmid DNA, messenger RNA (mRNA), or ribonucleoprotein (RNP) complexes, which need to enter target cells and tissues specifically and have good bioavailability. A broad spectrum of delivery modalities has been developed to meet these criteria, encompassing viral vectors, non-viral platforms, and hybrid systems. Viral vectors such as lentiviruses, adeno-associated viruses (AAVs), and baculoviruses have been widely used owing to their high transduction efficiency and stable expression [
160]. However, limitations such as immunogenicity, cytotoxicity, limited cargo capacity, random genomic integration, and difficulties in large-scale production in GMP setup have hindered their broader application in therapeutic [
161,
162]. These constraints have accelerated interest in non-viral delivery systems, which offer transient expression, enhanced safety profiles, and greater flexibility in cargo composition and size [
163]. Among these, nanoparticle-based systems including lipid nanoparticles (LNPs), polymeric nanoparticles, DNA nanoclew, Cell penetrating peptides (CPPs), gold nanoparticles, mesoporous silica nanoparticles, and biomimetic vesicles have emerged as promising platforms [
164]. These platforms have been optimized for delivery efficiency, endosomal escape, cellular uptake, and biocompatibility by fine-tuning their physicochemical parameters (e.g., size, charge, surface ligands). A recent research interest is growing in stimuli-responsive nanocarriers, which are engineered to release their cargo in response to specific intracellular or environmental triggers such as pH gradients, redox conditions, temperature, or light. These carriers enhance spatial and temporal control over CRISPR delivery, reducing off-target effects and improving overall specificity and effectiveness [
165]. In addition, ligand-functionalized nanoparticles have demonstrated remarkable potential for targeted delivery. These formulations are designed to interact with specific receptors on target cells, thereby facilitating receptor-mediated endocytosis or transcytosis. For example, the α7 nicotinic acetylcholine receptor, predominantly expressed in endothelial and neuronal cells, has been harnessed for targeted delivery. Another example includes αvβ3 integrin for enhanced tumor-specific delivery of CRISPR components as it is commonly over expressed in tumors [
166].
Multiple comprehensive reviews have previously summarized the various delivery platforms, discussing the design principles, advantages, and limitations of viral vectors, lipid-based carriers, polymeric systems, cell-penetrating peptides, extracellular vesicles, and virus-like particles. These reports have also evaluated key parameters such as delivery efficiency, immunogenicity, cargo capacity, and targeting specificity across diverse biological contexts [
167,
168,
169,
170,
171]. Building upon these recent and significant advances with a particular focus on the integration of synthetic biology, nanotechnology, and biomaterials science have been discussed. The focus is on the development of next-generation carriers that are suited for
in vivo MGE applications.
First such example includes metal-organic frameworks (MOFs), a class of porous crystalline materials, characterized by high surface area, tunable pore sizes, and chemical versatility that enable efficient encapsulation and intracellular release of nucleic acids and proteins. These vesicles protect components from enzymatic degradation and facilitate endosomal escape, resulting in enhanced editing efficiency in mammalian cells [
172,
173,
174]. Extracellular vesicle-based delivery systems have also attracted attention due to their potential to transport functional proteins and RNAs across cellular membranes in a physiologically compatible manner. ARRDC1-mediated micro vesicles (ARMMs) have demonstrated efficacy in delivering macromolecules [
175]. A recent study demonstrated significantly ameliorated acetaminophen-induced liver injury in mice using ARMMs loaded with Cas9 and sgRNAs against
Cyp2e1 gene. Notably, ARMMs enable packaging of genome editing components and tissue-specific delivery, although their performance remains limited to selected cell types and targets [
176]. Another report showed the design and use of VLPs based on Two-Photon Polymerization (2PP), termed as Nanoscribes. These have shown the capability to edit myoblasts, hiPSCs, and hiPSC-derived hematopoietic stem cells with an efficiency up to 25% [
177]. Another recent innovation involved using liposomal-coated mesoporous silica nanoparticles (lipoMSNs), which have been optimized for CRISPR-Cas9 plasmid and RNP delivery. Using this platform, researchers simultaneously edited key regulators of lipid metabolism (PCSK9, APOC3, and ANGPTL3) in murine hepatocytes, achieving synergistic reductions in serum cholesterol and triglycerides. This study demonstrates the potential of MGE for polygenic disorders, although further work is needed to assess long-term safety and off-target effects [
178]. Amphiphilic cell-penetrating peptides, such as RALA, represent another promising non-viral vector delivery system. RALA peptides self-assemble into nanoparticles through electrostatic interactions with negatively charged molecules before delivering them across the cell membrane. This facilitates cargo delivery to hard-to-transfect cells, including human mesenchymal stem cells. These platforms have shown low cytotoxicity and high editing efficiency, making them suitable for
ex vivo gene editing in regenerative medicine applications [
179]. Another alternative system that utilizes deconstructed wheat dwarf virus (dWDV)-based autonomously replicating vehicle was used to show high-efficiency editing in rice. This system enabled precise editing of the
OsEPSPS gene without compromising yield, underscoring its utility for plant genome engineering [
180].
The efficient delivery of CRISPR components in plants is more challenging due to their larger and more complex genome structures and rigid cell walls around the plant cell. The traditional
Agrobacterium mediated delivery has been complemented by engineering of other plant viruses such as Tobacco rattle virus (TRV) [
181], Sonchus yellow net rhabdovirus (SYNV) [
182] and Barley stripe mosaic virus (BMSV) [
183]. These viruses do not integrate in plant genomes and have higher cargo capacity; therefore, they are widely employed for DNA-free editing in plants [
184]. The particle bombardment is another most common alternative for transforming plants, which uses high-velocity particles to deliver CRISPR components into a wide range of difficult to transfect monocots and dicots [
185]. In addition to this, protoplast transfection offers a direct method for DNA-free genome editing and a good transient system to evaluate the activity of gene-editing reagents before moving into a more labor intensive transformation pipeline. Unfortunately, this is limited to the species with efficient protoplast regeneration protocols. In contrast to these classical methods, different classes of nanomaterials (carbon nanotubes, MSNs, polymeric NPs, Inorganic NPs, carbon dots, AuNPs) with functionalized surfaces have been used for genome editing applications in various plants such as
Zea mays,
Nicotina,
Triticum. aestivum,
Allium cepa, and
Arabidopsis [
186]. Despite these advances, achieving efficient delivery in the context of complex tissues, such as the immunosuppressive tumor microenvironment, remains an obstacle for engineered immune cell therapies. Stability and scalability of delivery formulations, particularly for clinical translation, also need to be addressed. Moreover, regulatory considerations around non-viral systems, particularly regarding repeat dosing, batch variability, and long-term genomic integrity pose additional hurdles. Continued efforts in material engineering, synthetic biology, and computational design will likely yield next-generation delivery systems that combine precision, safety, and flexibility across a broad range of applications.
6. Concluding Remarks
The advancements in CRISPR technology have significantly transformed the landscape of genome editing, particularly through multiplexing capabilities. The innovations in orthologous Cas9 enzymes or other RNA guided nucleases such as Cas12a enable researchers to target multiple genes simultaneously, enhancing the efficiency and precision of combinatorial CRISPR screens, revealing novel and complex genetic interactions. As the field continues to evolve, the potential applications of MGE span various domains, including agriculture, medicine, and biotechnology. In crop improvement avenues, DNA-free MGE is of immense importance for the commercialization of gene-edited crops because the resulting plants would not contain foreign DNA. Alongside these scientific advancements, ethical considerations and regulatory frameworks surrounding CRISPR-based genome editing also demand equal attention. Concerns related to biosafety, unintended off-target effects, ecological impacts, germline editing, and equitable access to these technologies are continuously evolving as our ability to modify genomes becomes more sophisticated. Importantly, these ethical discussions are not limited to multiplex applications but are broadly applicable to all CRISPR-based approaches. Addressing these aspects through inclusive dialogue and robust regulatory frameworks is crucial to ensure that scientific progress is aligned with societal needs and values. A recently published article describes a more detailed discussion of these aspects [
187]. In conclusion, the ongoing research and refinement focused on guide design and expression with an impetus from synthetic biology approaches, delivery, controlled expression of CRISPR elements, and innovative plasmid assembly techniques promise to unlock new avenues in biomedical research and crop resilience, ultimately contributing to a sustainable future.
Acknowledgments
We thank Smruti Ranjan Panda for assistance in the preparation of the manuscript. This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author Contributions
Conceptualization, D.R.; Writing—Original Draft Preparation, D.R. and G.B.; Writing—Review & Editing, D.R. and G.B.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
No data was used for the research described in the article.
Funding
This research received no external funding.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Pickar-Oliver A, Gersbach CA. The next generation of CRISPR–Cas technologies and applications.
Nat. Rev. Mol. Cell Biol. 2019,
20, 490–507. doi:10.1038/s41580-019-0131-5.
[Google Scholar]
-
2.
Doudna JA, Charpentier E. The new frontier of genome engineering with CRISPR-Cas9.
Science 2014,
346, 1258096. doi:10.1126/science.1258096.
[Google Scholar]
-
3.
Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, Zhang F. Genome engineering using the CRISPR-Cas9 system.
Nat. Protoc. 2013,
8, 2281–2308. doi:10.1038/nprot.2013.143.
[Google Scholar]
-
4.
Xue C, Greene EC. DNA repair pathway choices in CRISPR-Cas9-mediated genome editing.
Trends Genet. 2021,
37, 639–656. doi:10.1016/j.tig.2021.02.008.
[Google Scholar]
-
5.
Maruyama T, Dougan SK, Truttmann MC, Bilate AM, Ingram JR, Ploegh HL. Increasing the efficiency of precise genome editing with CRISPR-Cas9 by inhibition of nonhomologous end joining.
Nat. Biotechnol. 2015,
33, 538–542. doi:10.1038/nbt.3190.
[Google Scholar]
-
6.
Reuven N, Adler J, Broennimann K, Myers N, Shaul Y. Recruitment of DNA repair MRN complex by intrinsically disordered protein domain fused to Cas9 improves efficiency of CRISPR-mediated genome editing.
Biomolecules 2019,
9, 584. doi:10.3390/biom9100584.
[Google Scholar]
-
7.
Li G, Zhang X, Wang H, Liu D, Li Z, Wu Z, et al. Increasing CRISPR/Cas9-mediated homology-directed DNA repair by histone deacetylase inhibitors.
Int. J. Biochem. Cell Biol. 2020,
125, 105790. doi:10.1016/j.biocel.2020.105790.
[Google Scholar]
-
8.
Cong L, Ran FA, Cox D, Lin S, Barretto R, Habib N, et al. Multiplex genome engineering using CRISPR/Cas systems.
Science 2013,
339, 819–823. doi:10.1126/science.1231143.
[Google Scholar]
-
9.
Zhang Y, Buchholz F, Muyrers JP, Stewart AF. A new logic for DNA engineering using recombination in Escherichia coli.
Nat. Genet. 1998,
20, 123–128. doi:10.1038/2417.
[Google Scholar]
-
10.
Wang HH, Isaacs FJ, Carr PA, Sun ZZ, Xu G, Forest CR, et al. Programming cells by multiplex genome engineering and accelerated evolution.
Nature 2009,
460, 894–898. doi:10.1038/nature08187.
[Google Scholar]
-
11.
Bao Z, Cobb RE, Zhao H. Accelerated genome engineering through multiplexing.
Wiley Interdiscip Rev. Syst. Biol. Med. 2016,
8, 5–21. doi:10.1002/wsbm.1319.
[Google Scholar]
-
12.
Barbieri EM, Muir P, Akhuetie-Oni BO, Yellman CM, Isaacs FJ. Precise editing at DNA replication forks enables multiplex genome engineering in eukaryotes.
Cell 2017,
171, 1453–1467.e13. doi:10.1016/j.cell.2017.10.034.
[Google Scholar]
-
13.
Ronda C, Pedersen LE, Sommer MO, Nielsen AT. CRMAGE: CRISPR optimized mage recombineering.
Sci. Rep. 2016,
6, 19452. doi:10.1038/srep19452.
[Google Scholar]
-
14.
Nyerges Á, Csörgő B, Nagy I, Bálint B, Bihari P, Lázár V, et al. A highly precise and portable genome engineering method allows comparison of mutational effects across bacterial species.
Proc. Nat. Acad. Sci. USA 2016,
113, 2502–2507, Mar. doi:10.1073/pnas.1520040113.
[Google Scholar]
-
15.
García-Otín AL, Guillou F. Mammalian genome targeting using site-specific recombinases.
Front. Biosci. 2006,
11, 1108–1136.
[Google Scholar]
-
16.
Tian X, Zhou B. Strategies for site-specific recombination with high efficiency and precise spatiotemporal resolution.
J. Biol. Chem. 2021,
296, doi:10.1016/j.jbc.2021.100509.
[Google Scholar]
-
17.
Durrant MG, Perry NT, Pai JJ, Jangid AR, Athukoralage JS, Hiraizumi M, et al. Bridge RNAs direct programmable recombination of target and donor DNA.
Nature 2024,
630, 984–993. doi:10.1038/s41586-024-07552-4.
[Google Scholar]
-
18.
Hiraizumi M, Perry NT, Durrant MG, Soma T, Nagahata N, Okazaki S, et al. Structural mechanism of bridge RNA-guided recombination.
Nature 2024,
630, 994–1002. doi:10.1038/s41586-024-07570-2.
[Google Scholar]
-
19.
Partridge SR, Hall RM. The IS 1111 family members IS 4321 and IS 5075 have subterminal inverted repeats and target the terminal inverted repeats of Tn 21 family transposons.
J. Bacteriol. 2003,
185, 6371–6384. doi:10.1128/JB.185.21.6371-6384.2003.
[Google Scholar]
-
20.
Choi S, Ohta S, Ohtsubo E. A novel IS element, IS 621, of the IS 110/IS 492 family transposes to a specific site in repetitive extragenic palindromic sequences in Escherichia coli.
J. Bacteriol. 2003,
185, 4891–4900. doi:10.1128/JB.185.16.4891-4900.2003.
[Google Scholar]
-
21.
Gaj T, Gersbach CA, Barbas CF. ZFN, TALEN, and CRISPR/Cas-based methods for genome engineering.
Trends Biotechnol. 2013,
31, 397–405. doi:10.1016/j.tibtech.2013.04.004.
[Google Scholar]
-
22.
Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A programmable dual-RNA–guided DNA endonuclease in adaptive bacterial immunity.
Science 2012,
337, 816–821. doi:10.1126/science.1225829.
[Google Scholar]
-
23.
Pacesa M, Pelea O, Jinek M. Past, present, and future of CRISPR genome editing technologies.
Cell 2024,
187, 1076–1100. doi:10.1016/j.cell.2024.01.042.
[Google Scholar]
-
24.
Yin L, Xi D, Shen Y, Ding N, Shao Q, Qian Y, et al. Rewiring metabolic flux in Corynebacterium glutamicum using a CRISPR/dCpf1-based bifunctional regulation system.
J. Agric. Food Chem. 2024,
72, 3077–3087. doi:10.1021/acs.jafc.3c08529.
[Google Scholar]
-
25.
Lim SR, Lee HJ, Kim HJ, Lee SJ. Multiplex single-nucleotide microbial genome editing achieved by CRISPR-Cas9 using 5′-end-truncated sgRNAs.
ACS Synth. Biol. 2023,
12, 2203–2207. doi:10.1021/acssynbio.3c00323.
[Google Scholar]
-
26.
Utomo JC, Hodgins CL, Ro DK. Multiplex genome editing in yeast by CRISPR/Cas9–a potent and agile tool to reconstruct complex metabolic pathways.
Front. Plant Sci. 2021,
12, 719148. doi:10.3389/fpls.2021.719148.
[Google Scholar]
-
27.
Ronda C, Maury J, Jakočiu̅nas T, Baallal Jacobsen SA, Germann SM, Harrison SJ, et al. CrEdit: CRISPR mediated multi-loci gene integration in Saccharomyces cerevisiae.
Microb. Cell Factor. 2015,
14, 97. doi:10.1186/s12934-015-0288-3.
[Google Scholar]
-
28.
Lorenzo CD, Debray K, Herwegh D, Develtere W, Impens L, Schaumont D, et al. BREEDIT: a multiplex genome editing strategy to improve complex quantitative traits in maize.
Plant Cell 2023,
35, 218–238. doi:10.1093/plcell/koac243.
[Google Scholar]
-
29.
Ni P, Zhao Y, Zhou X, Liu Z, Huang Z, Ni Z, et al. Efficient and versatile multiplex prime editing in hexaploid wheat.
Genome Biol. 2023,
24, 156. doi:10.1186/s13059-023-02990-1.
[Google Scholar]
-
30.
Bai M, Yuan J, Kuang H, Gong P, Li S, Zhang Z, et al. Generation of a multiplex mutagenesis population via pooled CRISPR-Cas9 in soya bean.
Plant Biotechnol. J. 2020,
18, 721–731. doi:10.1111/pbi.13239.
[Google Scholar]
-
31.
Lu Y, Ye X, Guo R, Huang J, Wang W, Tang J, et al. Genome-wide targeted mutagenesis in rice using the CRISPR/Cas9 system.
Mol. Plant 2017,
10, 1242–1245. doi:10.1016/j.molp.2017.06.007.
[Google Scholar]
-
32.
Jacobs TB, Zhang N, Patel D, Martin GB. Generation of a collection of mutant tomato lines using pooled CRISPR libraries.
Plant Physiol. 2017,
174, 2023–2037. doi:10.1104/pp.17.00489.
[Google Scholar]
-
33.
Han K, Jeng EE, Hess GT, Morgens DW, Li A, Bassik MC. Synergistic drug combinations for cancer identified in a CRISPR screen for pairwise genetic interactions.
Nat. Biotechnol. 2017,
35, 463–474. doi:10.1038/nbt.3834.
[Google Scholar]
-
34.
Gao Z, Fan M, Das AT, Herrera-Carrillo E, Berkhout B. Extinction of all infectious HIV in cell culture by the CRISPR-Cas12a system with only a single crRNA.
Nucleic Acids Res. 2020,
48, 5527–5539. doi:10.1093/nar/gkaa226.
[Google Scholar]
-
35.
Thompson DB, Aboulhouda S, Hysolli E, Smith CJ, Wang S, Castanon O, et al. The Future of Multiplexed Eukaryotic Genome Engineering.
ACS Chem. Biol. 2018,
13, 313–325. doi:10.1021/acschembio.7b00842.
[Google Scholar]
-
36.
Yates KB, Tonnerre P, Martin GE, Gerdemann U, Al Abosy R, Comstock DE, et al. Epigenetic scars of CD8+ T cell exhaustion persist after cure of chronic infection in humans.
Nat. Immunol. 2021,
22, 1020–1029. doi:10.1038/s41590-021-00979-1.
[Google Scholar]
-
37.
Tang K, Zhou L, Tian X, Fang SY, Vandenbulcke E, Du A, et al. Cas12a-knock-in mice for multiplexed genome editing, disease modelling and immune-cell engineering.
Nat. Biomed. Eng. 2025. doi:10.1038/s41551-025-01371-2.
[Google Scholar]
-
38.
Guo S, Gao G, Zhang C, Peng G. Multiplexed genome editing for efficient phenotypic screening in Zebrafish.
Vet. Sci. 2022,
9, 92. doi:10.3390/vetsci9020092.
[Google Scholar]
-
39.
Singh J, Sharma D, Brar GS, Sandhu KS, Wani SH, Kashyap R, et al. CRISPR/Cas tool designs for multiplex genome editing and its applications in developing biotic and abiotic stress-resistant crop plants.
Mol. Biol. Rep. 2022,
49, 11443–11467. doi:10.1007/s11033-022-07741-2.
[Google Scholar]
-
40.
Abdelrahman M, Wei Z, Rohila JS, Zhao K. Multiplex genome-editing technologies for revolutionizing plant biology and crop improvement.
Front. Plant Sci. 2021,
12, 721203. doi:10.3389/fpls.2021.721203.
[Google Scholar]
-
41.
Hu Y, Patra P, Pisanty O, Shafir A, Belew ZM, Binenbaum J, et al. Multi-Knock—a multi-targeted genome-scale CRISPR toolbox to overcome functional redundancy in plants.
Nat. Plants 2023,
9, 572–587. doi:10.1038/s41477-023-01374-4.
[Google Scholar]
-
42.
Adiego-Perez B, Randazzo P, Daran JM, Verwaal R, Roubos JA, Daran-Lapujade P, et al. Multiplex genome editing of microorganisms using CRISPR-Cas.
FEMS Microbiol. Lett. 2019,
366,
fnz086. doi:10.1093/femsle/fnz086.
[Google Scholar]
-
43.
Lim SR, Lee SJ. Multiplex CRISPR-Cas genome editing: next-generation microbial strain engineering.
J. Agr. Food Chem. 2024,
72, 11871–11884. doi:10.1021/acs.jafc.4c01650.
[Google Scholar]
-
44.
Alok A, Sandhya D, Jogam P, Rodrigues V, Bhati KK, Sharma H, et al. The rise of the CRISPR/Cpf1 system for efficient genome editing in plants.
Front. Plant Sci. 2020,
11, 264. doi:10.3389/fpls.2020.00264.
[Google Scholar]
-
45.
Paul B, Montoya G. CRISPR-Cas12a: Functional overview and applications.
Biomed. J. 2020,
43, 8–17. doi:10.1016/j.bj.2019.10.005.
[Google Scholar]
-
46.
Walton RT, Christie KA, Whittaker MN, Kleinstiver BP. Unconstrained genome targeting with near-PAMless engineered CRISPR-Cas9 variants.
Science 2020,
368, 290–296. doi:10.1126/science.aba8853.
[Google Scholar]
-
47.
Nishimasu H, Cong L, Yan WX, Ran FA, Zetsche B, Li Y, et al. Crystal structure of Staphylococcus aureus Cas9.
Cell 2015,
162, 1113–1126. doi:10.1016/j.cell.2015.08.007.
[Google Scholar]
-
48.
Ran FA, Cong L, Yan WX, Scott DA, Gootenberg JS, Kriz AJ, et al. In vivo genome editing using Staphylococcus aureus Cas9.
Nature 2015,
520, 186–191. doi:10.1038/nature14299.
[Google Scholar]
-
49.
Jinek M, Jiang F, Taylor DW, Sternberg SH, Kaya E, Ma E, et al. Structures of Cas9 endonucleases reveal RNA-mediated conformational activation.
Science 2014,
343, 1247997. doi:10.1126/science.1247997.
[Google Scholar]
-
50.
Vakulskas CA, Dever DP, Rettig GR, Turk R, Jacobi AM, Collingwood MA, et al. A high-fidelity Cas9 mutant delivered as a ribonucleoprotein complex enables efficient gene editing in human hematopoietic stem and progenitor cells.
Nat Med. 2018,
24, 1216–1224. doi:10.1038/s41591-018-0137-0.
[Google Scholar]
-
51.
Agudelo D, Carter S, Velimirovic M, Duringer A, Rivest JF, Levesque S, et al. Versatile and robust genome editing with Streptococcus thermophilus CRISPR1-Cas9.
Genome Res. 2020,
30, 107–117. doi:10.1101/gr.255414.119.
[Google Scholar]
-
52.
Zetsche B, Gootenberg JS, Abudayyeh OO, Slaymaker IM, Makarova KS, Essletzbichler P, et al. Cpf1 is a single RNA-guided endonuclease of a class 2 CRISPR-Cas system.
Cell 2015,
163, 759–771. doi:10.1016/j.cell.2015.09.038.
[Google Scholar]
-
53.
Hino T, Omura SN, Nakagawa R, Togashi T, Takeda SN, Hiramoto T, et al. An AsCas12f-based compact genome-editing tool derived by deep mutational scanning and structural analysis.
Cell 2023,
186, 4920–4935. doi:https://doi.org/10.1016/j.cell.2023.08.031.
[Google Scholar]
-
54.
Zhang L, Zuris JA, Viswanathan R, Edelstein JN, Turk R, Thommandru B, et al. AsCas12a ultra nuclease facilitates the rapid generation of therapeutic cell medicines.
Nat. Commun. 2021,
12, 3908. doi:10.1038/s41467-021-24017-8.
[Google Scholar]
-
55.
Kleinstiver BP, Sousa AA, Walton RT, Tak YE, Hsu JY, Clement K, et al. Engineered CRISPR–Cas12a variants with increased activities and improved targeting ranges for gene, epigenetic and base editing.
Nat. Biotechnol. 2019,
37, 276–282. doi:10.1038/s41587-018-0011-0.
[Google Scholar]
-
56.
Hu Y, Han L, Mo Q, Du Z, Jiang W, Wu X, et al. Engineering miniature CRISPR-Cas Un1Cas12f1 for efficient base editing.
Mol. Ther. Nucleic Acids 2024,
35, 102201. doi:10.1016/j.omtn.2024.102201.
[Google Scholar]
-
57.
Chen L, Zhou X, Huang C, Zhang Y, Xin C, Hong J, et al. Engineered Un1Cas12f1 with boosted gene-editing activity and expanded genomic coverage.
Proc. Nat. Acad. Sci. USA 2025,
122, e2501292122. doi:10.1073/pnas.2501292122.
[Google Scholar]
-
58.
Xu X, Chemparathy A, Zeng L, Kempton HR, Shang S, Nakamura M, et al. Engineered miniature CRISPR-Cas system for mammalian genome regulation and editing.
Mol. Cell 2021,
81, 4333–4345. doi:https://doi.org/10.1016/j.molcel.2021.08.008.
[Google Scholar]
-
59.
Pausch P, Soczek KM, Herbst DA, Tsuchida CA, Al-Shayeb B, Banfield JF, et al. DNA interference states of the hypercompact CRISPR–CasΦ effector.
Nat. Struct. Mol. Biol. 2021,
28, 652–661. doi:10.1038/s41594-021-00632-3.
[Google Scholar]
-
60.
Pausch P, Al-Shayeb B, Bisom-Rapp E, Tsuchida CA, Li Z, Cress BF, et al. CRISPR-CasΦ from huge phages is a hypercompact genome editor.
Science 2020,
369, 333–337. doi:10.1126/science.abb1400.
[Google Scholar]
-
61.
Al-Shayeb B, Skopintsev P, Soczek KM, Stahl EC, Li Z, Groover E, et al. Diverse virus-encoded CRISPR-Cas systems include streamlined genome editors.
Cell 2022,
185, 4574–486. doi:10.1016/j.cell.2022.10.020.
[Google Scholar]
-
62.
Gaudelli NM, Lam DK, Rees HA, Solá-Esteves NM, Barrera LA, Born DA, et al. Directed evolution of adenine base editors with increased activity and therapeutic application.
Nat. Biotechnol. 2020,
38, 892–900. doi:10.1038/s41587-020-0491-6.
[Google Scholar]
-
63.
Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage.
Nature 2016,
533, 420–424. doi:10.1038/nature17946.
[Google Scholar]
-
64.
Neugebauer ME, Hsu A, Arbab M, Krasnow NA, McElroy AN, Pandey S, et al. Evolution of an adenine base editor into a small, efficient cytosine base editor with low off-target activity.
Nat. Biotechnol. 2023,
41, 673–685. doi:10.1038/s41587-022-01533-6.
[Google Scholar]
-
65.
Nishida K, Arazoe T, Yachie N, Banno S, Kakimoto M, Tabata M, et al. Targeted nucleotide editing using hybrid prokaryotic and vertebrate adaptive immune systems.
Science 2016,
353,
aaf8729. doi:10.1126/science.aaf8729.
[Google Scholar]
-
66.
Richter MF, Zhao KT, Eton E, Lapinaite A, Newby GA, Thuronyi BW, et al. Phage-assisted evolution of an adenine base editor with improved Cas domain compatibility and activity.
Nat. Biotechnol. 2020,
38, 883–891. doi:10.1038/s41587-020-0453-z.
[Google Scholar]
-
67.
Anzalone AV, Gao XD, Podracky CJ, Nelson AT, Koblan LW, Raguram A, et al. Programmable deletion, replacement, integration and inversion of large DNA sequences with twin prime editing.
Nat. Biotechnol. 2022,
40, 731–740. doi:10.1038/s41587-021-01133-w.
[Google Scholar]
-
68.
Anzalone AV, Randolph PB, Davis JR, Sousa AA, Koblan LW, Levy JM, et al. Search-and-replace genome editing without double-strand breaks or donor DNA.
Nature 2019,
576, 149–157. doi:10.1038/s41586-019-1711-4.
[Google Scholar]
-
69.
Yan J, Oyler-Castrillo P, Ravisankar P, Ward CC, Levesque S, Jing Y, et al. Improving prime editing with an endogenous small RNA-binding protein.
Nature 2024,
628, 639–647. doi:10.1038/s41586-024-07259-6.
[Google Scholar]
-
70.
Chen PJ, Hussmann JA, Yan J, Knipping F, Ravisankar P, Chen PF, et al. Enhanced prime editing systems by manipulating cellular determinants of editing outcomes.
Cell 2021,
184, 5635–5652. doi:10.1016/j.cell.2021.09.018.
[Google Scholar]
-
71.
Mali P, Yang L, Esvelt KM, Aach J, Guell M, DiCarlo JE, et al. RNA-guided human genome engineering via Cas9.
Science 2013,
339, 823–826. doi:10.1126/science.1232033.
[Google Scholar]
-
72.
Jiang W, Bikard D, Cox D, Zhang F, Marraffini LA. RNA-guided editing of bacterial genomes using CRISPR-Cas systems.
Nat. Biotechnol. 2013,
31, 233–239. doi:10.1038/nbt.2508.
[Google Scholar]
-
73.
Otoupal PB, Ito M, Arkin AP, Magnuson JK, Gladden JM, Skerker JM. Multiplexed CRISPR-Cas9-based genome editing of Rhodosporidium toruloides.
Msphere 2019,
4, 10–128. doi:10.1128/msphere.00099-19.
[Google Scholar]
-
74.
Sánchez-León S, Marín-Sanz M, Guzmán-López MH, Gavilán-Camacho M, Simón E, Barro F. CRISPR/Cas9-mediated multiplex gene editing of gamma and omega gliadins: paving the way for gliadin-free wheat.
J. Exp. Botany 2024,
75, 7079–7095. doi:10.1093/jxb/erae376.
[Google Scholar]
-
75.
Niu D, Wei HJ, Lin L, George H, Wang T, Lee IH, et al. Inactivation of porcine endogenous retrovirus in pigs using CRISPR-Cas9.
Science 2017,
357, 1303–1307. doi:10.1126/science.aan4187.
[Google Scholar]
-
76.
Ratan ZA, Son YJ, Haidere MF, Uddin BM, Yusuf MA, Zaman SB, et al. CRISPR-Cas9: a promising genetic engineering approach in cancer research.
Ther. Adv. Med. Oncol. 2018,
10, 1758834018755089. doi:10.1177/1758834018755089.
[Google Scholar]
-
77.
Rabaan AA, AlSaihati H, Bukhamsin R, Bakhrebah MA, Nassar MS, Alsaleh AA, et al. Application of CRISPR/Cas9 technology in cancer treatment: a future direction.
Curr. Oncol. 2023,
30, 1954–1976. doi:10.3390/curroncol30020152.
[Google Scholar]
-
78.
Chehelgerdi M, Chehelgerdi M, Khorramian-Ghahfarokhi M, Shafieizadeh M, Mahmoudi E, Eskandari F, et al. Comprehensive review of CRISPR-based gene editing: mechanisms, challenges, and applications in cancer therapy.
Mol. Cancer 2024,
23, 9. doi:10.1186/s12943-023-01925-5.
[Google Scholar]
-
79.
Kim E, Koo T, Park SW, Kim D, Kim K, Cho HY, et al. In vivo genome editing with a small Cas9 orthologue derived from Campylobacter jejuni.
Nat. Commun. 2017,
8, 14500. doi:10.1038/ncomms14500.
[Google Scholar]
-
80.
Schmidt MJ, Gupta A, Bednarski C, Gehrig-Giannini S, Richter F, Pitzler C, et al. Improved CRISPR genome editing using small highly active and specific engineered RNA-guided nucleases.
Nat. Commun. 2021,
12, 4219. doi:10.1038/s41467-021-24454-5.
[Google Scholar]
-
81.
Slaymaker IM, Gao L, Zetsche B, Scott DA, Yan WX, Zhang F. Rationally engineered Cas9 nucleases with improved specificity.
Science 2016,
351, 84–88. doi:10.1126/science.aad5227.
[Google Scholar]
-
82.
Kleinstiver BP, Pattanayak V, Prew MS, Tsai SQ, Nguyen NT, Zheng Z, et al. High-fidelity CRISPR–Cas9 nucleases with no detectable genome-wide off-target effects.
Nature 2016,
529, 490–495. 2016, doi:10.1038/nature16526.
[Google Scholar]
-
83.
Chen JS, Dagdas YS, Kleinstiver BP, Welch MM, Sousa AA, Harrington LB, et al. Enhanced proofreading governs CRISPR–Cas9 targeting accuracy.
Nature 2017,
550, 407–410. doi:10.1038/nature24268.
[Google Scholar]
-
84.
Huang X, Yang D, Zhang J, Xu J, Chen YE. Recent advances in improving gene-editing specificity through CRISPR–Cas9 nuclease engineering.
Cells 2022,
11, 2186. doi:10.3390/cells11142186.
[Google Scholar]
-
85.
Wang JY, Doudna JA. CRISPR technology: A decade of genome editing is only the beginning.
Science 2023,
379,
eadd8643. doi:10.1126/science.add8643.
[Google Scholar]
-
86.
Bayat H, Modarressi MH, Rahimpour A. The conspicuity of CRISPR-Cpf1 system as a significant breakthrough in genome editing.
Curr. Microbiol. 2018,
75, 107–115.
[Google Scholar]
-
87.
Zetsche B, Heidenreich M, Mohanraju P, Fedorova I, Kneppers J, DeGennaro EM, et al. Multiplex gene editing by CRISPR–Cpf1 using a single crRNA array.
Nat. Biotechnol. 2017,
35, 31–34. doi:10.1038/nbt.3737.
[Google Scholar]
-
88.
Li H, Zhu Z, Li S, Li J, Yan L, Zhang C, et al. Multiplex precision gene editing by a surrogate prime editor in rice.
Mol. Plant 2022,
15, 1077–1080. doi:https://doi.org/10.1016/j.molp.2022.05.009.
[Google Scholar]
-
89.
Zhang Y, Ren Q, Tang X, Liu S, Malzahn AA, Zhou J, et al. Expanding the scope of plant genome engineering with Cas12a orthologs and highly multiplexable editing systems.
Nat. Commun. 2021,
12, 1944. doi:10.1038/s41467-021-22330-w.
[Google Scholar]
-
90.
Li T, Zhu L, Xiao B, Gong Z, Liao Q, Guo J. CRISPR-Cpf1-mediated genome editing and gene regulation in human cells.
Biotechnol. Adv. 2019,
37, 21–27. doi:https://doi.org/10.1016/j.biotechadv.2018.10.013.
[Google Scholar]
-
91.
Pardue MT, Allen RS. Neuroprotective strategies for retinal disease.
Progr. Retinal Eye Res. 2018,
65, 50–76. doi:https://doi.org/10.1016/j.preteyeres.2018.02.002.
[Google Scholar]
-
92.
Sepah YJ, Velez G, Tang PH, Yang J, Chemudupati T, Li AS, et al. Proteomic analysis of intermediate uveitis suggests myeloid cell recruitment and implicates IL-23 as a therapeutic target.
Am. J. Ophthalmol. Case Rep. 2020,
18, 100646. doi:https://doi.org/10.1016/j.ajoc.2020.100646.
[Google Scholar]
-
93.
Bigini F, Lee SH, Sun YJ, Sun Y, Mahajan VB. Unleashing the potential of CRISPR multiplexing: Harnessing Cas12 and Cas13 for precise gene modulation in eye diseases.
Vision Res. 2023,
213, 108317. doi:https://doi.org/10.1016/j.visres.2023.108317.
[Google Scholar]
-
94.
Xuan Q, Wang J, Nie Y, Fang C, Liang W. Research progress and application of miniature CRISPR-Cas12 system in gene editing.
Int. J. Mol. Sci. 2024,
25, 12686. doi:10.3390/ijms252312686.
[Google Scholar]
-
95.
Wu H, Sun Y, Wang Y, Luo L, Song Y. Advances in miniature CRISPR-Cas proteins and their applications in gene editing.
Arch. Microbiol. 2024,
206, 231. doi:10.1007/s00203-024-03962-0.
[Google Scholar]
-
96.
Chen P, Zhou J, Wan Y, Liu H, Li Y, Liu Z, et al. A Cas12a ortholog with stringent PAM recognition followed by low off-target editing rates for genome editing.
Genome Biol. 2020,
21, 78. doi:10.1186/s13059-020-01989-2.
[Google Scholar]
-
97.
Wu WY, Mohanraju P, Liao C, Adiego-Pérez B, Creutzburg SC, Makarova KS, et al. The miniature CRISPR-Cas12m effector binds DNA to block transcription.
Mol. Cell 2022,
82, 4487–4502. doi:10.1016/j.molcel.2022.11.003.
[Google Scholar]
-
98.
Chen W, Ma J, Wu Z, Wang Z, Zhang H, Fu W, et al. Cas12n nucleases, early evolutionary intermediates of type V CRISPR, comprise a distinct family of miniature genome editors.
Mol. Cell 2023,
83, 2768–2780. doi:10.1016/j.molcel.2023.06.014.
[Google Scholar]
-
99.
Awan MJ, Amin I, Mansoor S. CRISPR-Cas12c: a noncleaving DNA binder with minimal PAM requirement.
Trends Biotechnol. 2022,
40, 1141–1143. doi:10.1016/j.tibtech.2022.07.005.
[Google Scholar]
-
100.
Badon IW, Oh Y, Kim HJ, Lee SH. Recent application of CRISPR-Cas12 and OMEGA system for genome editing.
Mol. Ther. 2024,
32, 32–43. doi:https://doi.org/10.1016/j.ymthe.2023.11.013.
[Google Scholar]
-
101.
Kim DY, Lee JM, Moon SB, Chin HJ, Park S, Lim Y, et al. Efficient CRISPR editing with a hypercompact Cas12f1 and engineered guide RNAs delivered by adeno-associated virus.
Nat. Biotechnol. 2022,
40, 94–102. doi:10.1038/s41587-021-01009-z.
[Google Scholar]
-
102.
Lim SR, Kim HJ, Lee SJ. Efficient CRISPR-Cas12f1-mediated multiplex bacterial genome editing via low-temperature recovery.
J. Microbiol. Biotechnol. 2024,
34, 1522. doi:10.4014/jmb.2403.03033.
[Google Scholar]
-
103.
Yan F, Wang J, Zhang S, Lu Z, Li S, Ji Z, et al. CRISPR/FnCas12a-mediated efficient multiplex and iterative genome editing in bacterial plant pathogens without donor DNA templates.
PLoS Pathogens. 2023,
19, e1010961. doi:10.1371/journal.ppat.1010961.
[Google Scholar]
-
104.
Wang C, Xia Q, Zhang Q, Qu Y, Su S, Cheng JK, et al. CRISPR-Cas12a System With Synergistic Phage Recombination Proteins for Multiplex Precision Editing in Human Cells.
Front. Cell Dev. Biol. 2022,
9, 719705. doi:10.3389/fcell.2021.719705.
[Google Scholar]
-
105.
Jordan WT, Currie S, Schmitz RJ. Multiplex genome editing in Arabidopsis thaliana using Mb3Cas12a.
Plant Direct. 2021,
5, e344. doi:10.1002/pld3.344.
[Google Scholar]
-
106.
Hui F, Tang X, Li B, Alariqi M, Xu Z, Meng Q, et al. Robust CRISPR/Mb2Cas12a genome editing tools in cotton plants.
Imeta 2024,
3, e209. doi:10.1002/imt2.209.
[Google Scholar]
-
107.
Wang W, Li S, Yang J, Li J, Yan L, Zhang C, et al. Exploiting the efficient Exo: Cas12i3-5M fusions for robust single and multiplex gene editing in rice.
J. Integr. Plant Biol. 2025,
67, 1246–1253. doi:10.1111/jipb.13850.
[Google Scholar]
-
108.
Liu S, Sretenovic S, Fan T, Cheng Y, Li G, Qi A, et al. Hypercompact CRISPR–Cas12j2 (CasΦ) enables genome editing, gene activation, and epigenome editing in plants.
Plant Commun. 2022,
3, 100453. doi:10.1016/j.xplc.2022.100453.
[Google Scholar]
-
109.
Cui Y, Dong H, Tong B, Wang H, Chen X, Liu G, et al. A versatile Cas12k-based genetic engineering toolkit (C12KGET) for metabolic engineering in genetic manipulation-deprived strains.
Nucleic Acids Res. 2022,
50, 8961–8973. doi:10.1093/nar/gkac655.
[Google Scholar]
-
110.
Averina OA, Kuznetsova SA, Permyakov OA, Sergiev PV. Current knowledge of base editing and prime editing.
Mol. Biol. 2024,
58, 571–587. doi:10.31857/S0026898424040013.
[Google Scholar]
-
111.
Gaudelli NM, Komor AC, Rees HA, Packer MS, Badran AH, Bryson DI, et al. Programmable base editing of A• T to G• C in genomic DNA without DNA cleavage.
Nature 2017,
551, 464–471. doi:10.1038/nature24644.
[Google Scholar]
-
112.
Yan D, Ren B, Liu L, Yan F, Li S, Wang G, et al. High-efficiency and multiplex adenine base editing in plants using new TadA variants.
Mol. Plant 2021,
14, 722–731. doi:https://doi.org/10.1016/j.molp.2021.02.007.
[Google Scholar]
-
113.
Mishra R, Joshi RK, Zhao K. Genome editing in rice: recent advances, challenges, and future implications.
Front. Plant Sci. 2018,
9, 1361. doi:10.3389/fpls.2018.01361.
[Google Scholar]
-
114.
Sun Y, Jiao G, Liu Z, Zhang X, Li J, Guo X, et al. Generation of high-amylose rice through CRISPR/Cas9-mediated targeted mutagenesis of starch branching enzymes.
Front. Plant Sci. 2017,
8, 298. doi:10.3389/fpls.2017.00298.
[Google Scholar]
-
115.
Xu Y, Lin Q, Li X, Wang F, Chen Z, Wang J, et al. Fine-tuning the amylose content of rice by precise base editing of the Wx gene.
Plant Biotechnol. J. 2021,
19, 11–13. doi:10.1111/pbi.13433.
[Google Scholar]
-
116.
Li J, Sun Y, Du J, Zhao Y, Xia L. Generation of targeted point mutations in rice by a modified CRISPR/Cas9 system.
Mol. Plant 2017,
10, 526–529. doi:10.1016/j.molp.2016.12.001.
[Google Scholar]
-
117.
Liu X, Qin R, Li J, Liao S, Shan T, Xu R, et al. A CRISPR-Cas9-mediated domain-specific base-editing screen enables functional assessment of ACCase variants in rice.
Plant Biotechnol. J. 2020,
18, 1845. doi:10.1111/pbi.13348.
[Google Scholar]
-
118.
Tabassum J, Ahmad S, Hussain B, Mawia AM, Zeb A, Ju L. Applications and potential of genome-editing systems in rice improvement: current and future perspectives.
Agronomy 2021,
11, 1359. doi:10.3390/agronomy11071359.
[Google Scholar]
-
119.
Monsur MB, Shao G, Lv Y, Ahmad S, Wei X, Hu P, et al. Base editing: The ever expanding clustered regularly interspaced short palindromic repeats (CRISPR) tool kit for precise genome editing in plants.
Genes 2020,
11, 466. doi:10.3390/genes11040466.
[Google Scholar]
-
120.
Xie J, Huang X, Wang X, Gou S, Liang Y, Chen F, et al. ACBE, a new base editor for simultaneous C-to-T and A-to-G substitutions in mammalian systems.
BMC Biol. 2020,
18, 131. doi:10.1186/s12915-020-00866-5.
[Google Scholar]
-
121.
Wu Y, Li Y, Liu Y, Xiu X, Liu J, Zhang L, et al. Multiplexed in-situ mutagenesis driven by a dCas12a-based dual-function base editor.
Nucleic Acids Res. 2024,
52, 4739–4755. doi:10.1093/nar/gkae228.
[Google Scholar]
-
122.
Grünewald J, Zhou R, Lareau CA, Garcia SP, Iyer S, Miller BR, et al. A dual-deaminase CRISPR base editor enables concurrent adenine and cytosine editing.
Nat. Biotechnol. 2020,
38, 861–864. doi:10.1038/s41587-020-0535-y.
[Google Scholar]
-
123.
Butt H, Rao GS, Sedeek K, Aman R, Kamel R, Mahfouz M. Engineering herbicide resistance via prime editing in rice.
Plant Biotechnol. J. 2020,
18, 2370. doi:10.1111/pbi.13399.
[Google Scholar]
-
124.
Gupta A, Liu B, Raza S, Chen QJ, Yang B. Modularly assembled multiplex prime editors for simultaneous editing of agronomically important genes in rice.
Plant Commun. 2024,
5. 100741. doi:https://doi.org/10.1016/j.xplc.2023.100741.
[Google Scholar]
-
125.
Doman JL, Sousa AA, Randolph PB, Chen PJ, Liu DR. Designing and executing prime editing experiments in mammalian cells.
Nat Protocols 2022,
17, 2431–2468. doi:10.1038/s41596-022-00724-4.
[Google Scholar]
-
126.
Nelson JW, Randolph PB, Shen SP, Everette KA, Chen PJ, Anzalone AV, et al. Engineered pegRNAs improve prime editing efficiency.
Nat. Biotechnol. 2022,
40, 402–410. doi:10.1038/s41587-021-01039-7.
[Google Scholar]
-
127.
Awan MJ, Ali Z, Amin I, Mansoor S. Twin prime editor: seamless repair without damage.
Trends Biotechnol. 2022,
40, 374–376. doi:10.1016/j.tibtech.2022.01.013.
[Google Scholar]
-
128.
Wu K, Sánchez-Rivera FJ. It's prime time for multiplexed prime editing.
Cell Genom. 2025,
5, 100852. doi:10.1016/j.xgen.2025.100852.
[Google Scholar]
-
129.
Yuan Q, Gao X. Multiplex base-and prime-editing with drive-and-process CRISPR arrays.
Nat. Commun. 2022,
13, 2771. doi:10.1038/s41467-022-30514-1.
[Google Scholar]
-
130.
Zhao X, Yang L, Li P, Cheng Z, Jia Y, Luo L, et al. High-accuracy crRNA array assembly strategy for multiplex CRISPR.
Mol. Ther. Nucleic Acids 2025,
36, 102428. doi:10.1016/j.omtn.2024.102428.
[Google Scholar]
-
131.
Wang JW, Wang A, Li K, Wang B, Jin S, Reiser M, et al. CRISPR/Cas9 nuclease cleavage combined with Gibson assembly for seamless cloning.
Biotechniques 2015,
58, 161–170. doi:10.2144/000114261.
[Google Scholar]
-
132.
Breunig CT, Durovic T, Neuner AM, Baumann V, Wiesbeck MF, Köferle A, et al. One step generation of customizable gRNA vectors for multiplex CRISPR approaches through string assembly gRNA cloning (STAgR).
PloS ONE 2018,
13, e0196015. doi:10.1371/journal.pone.0196015.
[Google Scholar]
-
133.
Wu S, Kyaw H, Tong Z, Yang Y, Wang Z, Zhang L, et al. A simple and efficient CRISPR/Cas9 system permits ultra-multiplex genome editing in plants.
Crop J. 2024,
12, 569–582. doi:10.1016/j.cj.2024.01.010.
[Google Scholar]
-
134.
Hsieh-Feng V, Yang Y. Efficient expression of multiple guide RNAs for CRISPR/Cas genome editing.
Abiotech 2020,
1, 123–134. doi:10.1007/s42994-019-00014-w.
[Google Scholar]
-
135.
Dong F, Xie K, Chen Y, Yang Y, Mao Y. Polycistronic tRNA and CRISPR guide-RNA enables highly efficient multiplexed genome engineering in human cells.
Biochem. Biophys. Res. Commun. 2017,
482, 889–895. doi:10.1016/j.bbrc.2016.11.129.
[Google Scholar]
-
136.
Xie K, Minkenberg B, Yang Y. Boosting CRISPR/Cas9 multiplex editing capability with the endogenous tRNA-processing system.
Proc. Nat. Acad. Sci. USA 2015,
112, 3570–3575. doi:10.1073/pnas.1420294112.
[Google Scholar]
-
137.
Jiang C, Geng L, Wang J, Liang Y, Guo X, Liu C, et al. Multiplexed gene engineering based on dCas9 and gRNA-tRNA array encoded on single transcript.
Int. J. Mol. Sci. 2023,
24, 8535. doi:10.3390/ijms24108535.
[Google Scholar]
-
138.
Gao Z, Herrera-Carrillo E, Berkhout B. Improvement of the CRISPR-Cpf1 system with ribozyme-processed crRNA.
RNA Biol. 2018,
15, 1458–1467. doi:10.1080/15476286.2018.1551703.
[Google Scholar]
-
139.
Wang L, Han H. Strategies for improving the genome-editing efficiency of class 2 CRISPR/Cas system.
Heliyon 2024,
10, e38588. doi:10.1016/J.HELIYON.2024.E38588.
[Google Scholar]
-
140.
Xu L, Zhao L, Gao Y, Xu J, Han R. Empower multiplex cell and tissue-specific CRISPR-mediated gene manipulation with self-cleaving ribozymes and tRNA.
Nucleic acids Res. 2017,
45, e28. doi:10.1093/nar/gkw1048.
[Google Scholar]
-
141.
Haurwitz RE, Sternberg SH, Doudna JA. Csy4 relies on an unusual catalytic dyad to position and cleave CRISPR RNA. The
EMBO J. 2012,
31, 2824–2832. doi:10.1038/emboj.2012.107.
[Google Scholar]
-
142.
Yan Q, Xu K, Xing J, Zhang T, Wang X, Wei Z, et al. Multiplex CRISPR/Cas9-based genome engineering enhanced by Drosha-mediated sgRNA-shRNA structure.
Sci. Rep. 2016,
6, 38970. doi:10.1038/srep38970.
[Google Scholar]
-
143.
Bevacqua RJ, Zhao W, Merheb E, Kim SH, Marson A, Gloyn AL, et al. Multiplexed CRISPR gene editing in primary human islet cells with Cas9 ribonucleoprotein.
iScience 2023,
27, 108693. doi:10.1016/J.ISCI.2023.108693.
[Google Scholar]
-
144.
Bevacqua RJ, Zhao W, Merheb E, Kim SH, Marson A, Gloyn AL, et al. Multiplexed CRISPR gene editing in primary human islet cells with Cas9 ribonucleoprotein.
bioRxiv 2023, doi:10.1101/2023.09.16.558090.
[Google Scholar]
-
145.
Nijkamp E, Ruffolo JA, Weinstein EN, Naik N, Madani A. Progen2: exploring the boundaries of protein language models.
Cell Syst. 2023,
14, 968–978. doi:10.1016/j.cels.2023.10.002.
[Google Scholar]
-
146.
McCarty NS, Graham AE, Studená L, Ledesma-Amaro R. Multiplexed CRISPR technologies for gene editing and transcriptional regulation.
Nat. Commun. 2020,
11, 1281. doi:10.1038/s41467-020-15053-x.
[Google Scholar]
-
147.
Xun G, Zhu Z, Singh N, Lu J, Jain PK, Zhao H. Harnessing noncanonical crRNA for highly efficient genome editing.
Nat. Commun. 2024,
15, 3823. doi:10.1038/s41467-024-48012-x.
[Google Scholar]
-
148.
Bin Moon SU, Lee JM, Kang JG, Lee NE, Ha DI, Kim DY, et al. Highly efficient genome editing by CRISPR-Cpf1 using CRISPR RNA with a uridinylate-rich 3′-overhang.
Nat. Commun. 2018,
9, 3651. doi:10.1038/s41467-018-06129-w.
[Google Scholar]
-
149.
Liao C, Ttofali F, Slotkowski RA, Denny SR, Cecil TD, Leenay RT, et al. Modular one-pot assembly of CRISPR arrays enables library generation and reveals factors influencing crRNA biogenesis.
Nat. Commun. 2019,
10, 2948. doi:10.1038/s41467-019-10747-3.
[Google Scholar]
-
150.
Erdoğan S. Integration of artificial intelligence and genome editing system for determining the treatment of genetic disorders.
Balkan Med. J. 2024,
41, 419. doi:10.4274/balkanmedj.galenos.2024.2024-080824.
[Google Scholar]
-
151.
Chen L, Liu G, Zhang T. Integrating machine learning and genome editing for crop improvement.
Abiotech 2024,
5, 262–277. doi:10.1007/s42994-023-00133-5.
[Google Scholar]
-
152.
Chia BS, Seah YF, Wang B, Shen K, Srivastava D, Chew WL. Engineering a New Generation of Gene Editors: Integrating Synthetic Biology and AI Innovations.
ACS Synth. Biol. 2025,
14, 636–647. doi:10.1021/acssynbio.4c00686.
[Google Scholar]
-
153.
Doench JG, Hartenian E, Graham DB, Tothova Z, Hegde M, Smith I, et al. Rational design of highly active sgRNAs for CRISPR-Cas9–mediated gene inactivation.
Nat. Biotechnol. 2014,
32, 1262–1267. doi:10.1038/nbt.3026.
[Google Scholar]
-
154.
Montague TG, Cruz JM, Gagnon JA, Church GM, Valen E. CHOPCHOP: a CRISPR/Cas9 and TALEN web tool for genome editing.
Nucleic Acids Res. 2014,
42, W401–W407. doi:10.1093/nar/gku410.
[Google Scholar]
-
155.
Li J, Wu P, Cao Z, Huang G, Lu Z, Yan J, et al. Machine learning-based prediction models to guide the selection of Cas9 variants for efficient gene editing.
Cell Rep. 2024,
43, 113765. doi:10.1016/j.celrep.2024.113765.
[Google Scholar]
-
156.
Alipanahi R, Safari L, Khanteymoori A. DTMP-prime: A deep transformer-based model for predicting prime editing efficiency and PegRNA activity.
Mol. Ther. Nucleic Acids 2024,
35, 102370. doi:10.1016/j.omtn.2024.102370.
[Google Scholar]
-
157.
Ruffolo JA, Nayfach S, Gallagher J, Bhatnagar A, Beazer J, Hussain R, et al. Design of highly functional genome editors by modelling CRISPR–Cas sequences.
Nature 2025,
645, 518–525. doi:10.1038/s41586-025-09298-z.
[Google Scholar]
-
158.
Kim MG, Go MJ, Kang SH, Jeong SH, Lim K. Revolutionizing CRISPR technology with artificial intelligence.
Exp. Mol. Med. 2025,
57, 1419–1431. doi:10.1038/s12276-025-01462-9.
[Google Scholar]
-
159.
Qu Y, Huang K, Yin M, Zhan K, Liu D, Yin D, et al. CRISPR-GPT for agentic automation of gene-editing experiments.
Nat. Biomed. Eng. 2025, 1–4. doi:10.1038/s41551-025-01463-z.
[Google Scholar]
-
160.
Du Y, Liu Y, Hu J, Peng X, Liu Z. CRISPR/Cas9 systems: Delivery technologies and biomedical applications.
Asian J. Pharm. Sci. 2023,
18, 100854. doi:10.1016/j.ajps.2023.100854.
[Google Scholar]
-
161.
Wang D, Tai PW, Gao G. Adeno-associated virus vector as a platform for gene therapy delivery.
Nat. Rev. Drug discov. 2019,
18, 358–378. doi:10.1038/s41573-019-0012-9.
[Google Scholar]
-
162.
Lundstrom K. Viral vectors in gene therapy: where do we stand in 2023?
Viruses 2023,
15, 698. doi:10.3390/v15030698.
[Google Scholar]
-
163.
Sioson VA, Kim M, Joo J. Challenges in delivery systems for CRISPR-based genome editing and opportunities of nanomedicine.
Biomed. Eng. Lett. 2021,
11, 217–233. doi:10.1007/s13534-021-00199-4.
[Google Scholar]
-
164.
Lino CA, Harper JC, Carney JP, Timlin JA. Delivering CRISPR: a review of the challenges and approaches.
Drug Deliv. 2018,
25, 1234–1257. doi:10.1080/10717544.2018.1474964.
[Google Scholar]
-
165.
Fang T, Cao X, Ibnat M, Chen G. Stimuli-responsive nanoformulations for CRISPR-Cas9 genome editing.
J. Nanobiotechnol. 2022,
20, 354. doi:10.1186/s12951-022-01570-y.
[Google Scholar]
-
166.
Zhang J, Zhou Y, Qiao J, Liu Y. Recent advances in spatiotemporal control of the CRISPR/Cas9 system.
Colloid. Surf. B Biointerf. 2025,
248, 114474. doi:https://doi.org/10.1016/j.colsurfb.2024.114474.
[Google Scholar]
-
167.
Clarissa EM, Karmacharya M, Choi H, Kumar S, Cho YK. Nature Inspired Delivery Vehicles for CRISPR-Based Genome Editing.
Small 2025, 2409353. doi:10.1002/smll.202409353.
[Google Scholar]
-
168.
Lee H, Rho WY, Kim YH, Chang H, Jun BH. CRISPR-Cas9 gene therapy: non-viral delivery and stimuli-responsive nanoformulations.
Molecules 2025,
30, 542. doi:10.3390/molecules30030542.
[Google Scholar]
-
169.
Tashima T. Non-Invasive Delivery of CRISPR/Cas9 Ribonucleoproteins (Cas9 RNPs) into Cells via Nanoparticles for Membrane Transport.
Pharmaceutics 2025,
17, 201. doi:10.3390/pharmaceutics17020201.
[Google Scholar]
-
170.
Kaupbayeva B, Tsoy A, Safarova Y, Nurmagambetova A, Murata H, Matyjaszewski K, et al. Unlocking genome editing: Advances and obstacles in CRISPR/Cas delivery technologies.
J. Funct. Biomat. 2024,
15, 324. doi:10.3390/jfb15110324.
[Google Scholar]
-
171.
Shaikhutdinov IH, Ilyasov PV, Gribkova OV, Limareva LV. Non-viral systems for intracellular delivery of genome editing tools.
Vavilov J. Genet.s Breeding 2024,
28, 239. doi:10.18699/vjgb-24-28.
[Google Scholar]
-
172.
Alyami MZ, Alsaiari SK, Li Y, Qutub SS, Aleisa FA, Sougrat R, et al. Cell-type-specific CRISPR/Cas9 delivery by biomimetic metal organic frameworks.
J. Am. Chem. Soc. 2020,
142, 1715–120. doi:10.1021/jacs.9b11638.
[Google Scholar]
-
173.
Kangarshahi BM, Javazm AB, Naghib SM. CRISPR-integrated metal-organic frameworks for biosensing applications: Recent advances and future perspective.
Sensing Bio-Sensing Res. 2025,
47, 100736. doi:https://doi.org/10.1016/j.sbsr.2025.100736.
[Google Scholar]
-
174.
Yu J, Tang M, Zhou Z, Wei Z, Wan F, Hou S, et al. Biologically produced and metal-organic framework delivered dual-cut CRISPR/Cas9 system for efficient gene editing and sensitized cancer therapy.
Acta Biomat. 2024,
178, 296–306. doi:10.1016/j.actbio.2024.02.030.
[Google Scholar]
-
175.
Wang Q, Yu J, Kadungure T, Beyene J, Zhang H, Lu Q. ARMMs as a versatile platform for intracellular delivery of macromolecules.
Nat. Commun. 2018,
9, 960. doi:10.1038/s41467-018-03390-x.
[Google Scholar]
-
176.
Chen Z, Wang Q, Lu Q. Engineering ARMMs for improved intracellular delivery of CRISPR-Cas9.
Extracell. Vesicle 2025,
5, 100082. doi:https://doi.org/10.1016/j.vesic.2025.100082.
[Google Scholar]
-
177.
Halegua T, Risson V, Carras J, Rouyer M, Coudert L, Jacquier A, et al. Delivery of Prime editing in human stem cells using pseudoviral NanoScribes particles.
Nat. Commun. 2025,
16, 397. doi:10.1038/s41467-024-55604-0.
[Google Scholar]
-
178.
LaBauve AE, Saada EA, Jones IK, Mosesso R, Noureddine A, Techel J, et al. Lipid-coated mesoporous silica nanoparticles for anti-viral applications via delivery of CRISPR-Cas9 ribonucleoproteins.
Sci. Rep. 2023,
13, 6873. doi:10.1038/s41598-023-33092-4.
[Google Scholar]
-
179.
Graham JP, Castro JG, Werba LC, Amitrano J, Fardone L, Francis KP, et al. Versatile Cell Penetrating Peptide for Multimodal CRISPR Gene Editing in Primary Stem Cells.
Adv. Funct. Mat. 2025, e01532. doi:10.1101/2024.09.23.614499.
[Google Scholar]
-
180.
Borphukan B, Khatun M, Fartyal D, James D, Reddy MK. A Gemini Virus-Derived Autonomously Replicating System for HDR-Mediated Genome Editing of the EPSP Synthase Gene in Indica Rice.
Plants 2025,
14, 477. doi:10.3390/plants14030477.
[Google Scholar]
-
181.
Guo G, Li MJ, Lai JL, Du ZY, Liao QS. Development of tobacco rattle virus-based platform for dual heterologous gene expression and CRISPR/Cas reagent delivery.
Plant Sci. 2022,
325, 111491. doi:10.1016/j.plantsci.2022.111491.
[Google Scholar]
-
182.
Daròs JA, Pasin F, Merwaiss F. CRISPR-Cas-based plant genome engineering goes viral.
Mol. Plant 2023,
16, 660–661. doi:10.1016/j.molp.2023.03.010.
[Google Scholar]
-
183.
Hu J, Li S, Li Z, Li H, Song W, Zhao H, et al. A barley stripe mosaic virus-based guide RNA delivery system for targeted mutagenesis in wheat and maize.
Mol. Plant Pathol. 2019,
20, 1463–1474. doi:10.1111/mpp.12849.
[Google Scholar]
-
184.
Mahmood MA, Naqvi RZ, Rahman SU, Amin I, Mansoor S. Plant virus-derived vectors for plant genome engineering.
Viruses 2023,
15, 531. doi:10.3390/v15020531.
[Google Scholar]
-
185.
Panda D, Karmakar S, Dash M, Tripathy SK, Das P, Banerjee S, et al. Optimized protoplast isolation and transfection with a breakpoint: accelerating Cas9/sgRNA cleavage efficiency validation in monocot and dicot.
Abiotech 2024,
5, 151–168. doi:10.1007/s42994-024-00139-7.
[Google Scholar]
-
186.
Vats S, Kumawat S, Brar J, Kaur S, Yadav K, Magar SG, et al. Opportunity and challenges for nanotechnology application for genome editing in plants.
Plant Nano Biol. 2022,
1, 100001. doi:10.1016/j.plana.2022.100001.
[Google Scholar]
-
187.
Bindal G, Misra CS, Singh M, Rangu SS, Pandey N, Rath D. High throughput CRISPR approaches: new solutions for challenging problems.
TrAC Trends Analyt. Chem. 2025,
189, 118245. doi:10.1016/J.TRAC.2025.118245.
[Google Scholar]
 Devashish Rath
1,2,*
Devashish Rath
1,2,*