1. Introduction
Collagen, a principal component of the extracellular matrix (ECM) [
1], constitutes ~30% of the human body’s total protein content. It represents 70% of the skin’s dry weight, and ~90% of the protein in tendons and corneal tissue [
2]. The collagen family comprises 28 types, with molecular isomerism underpinning functional and structural diversity () [
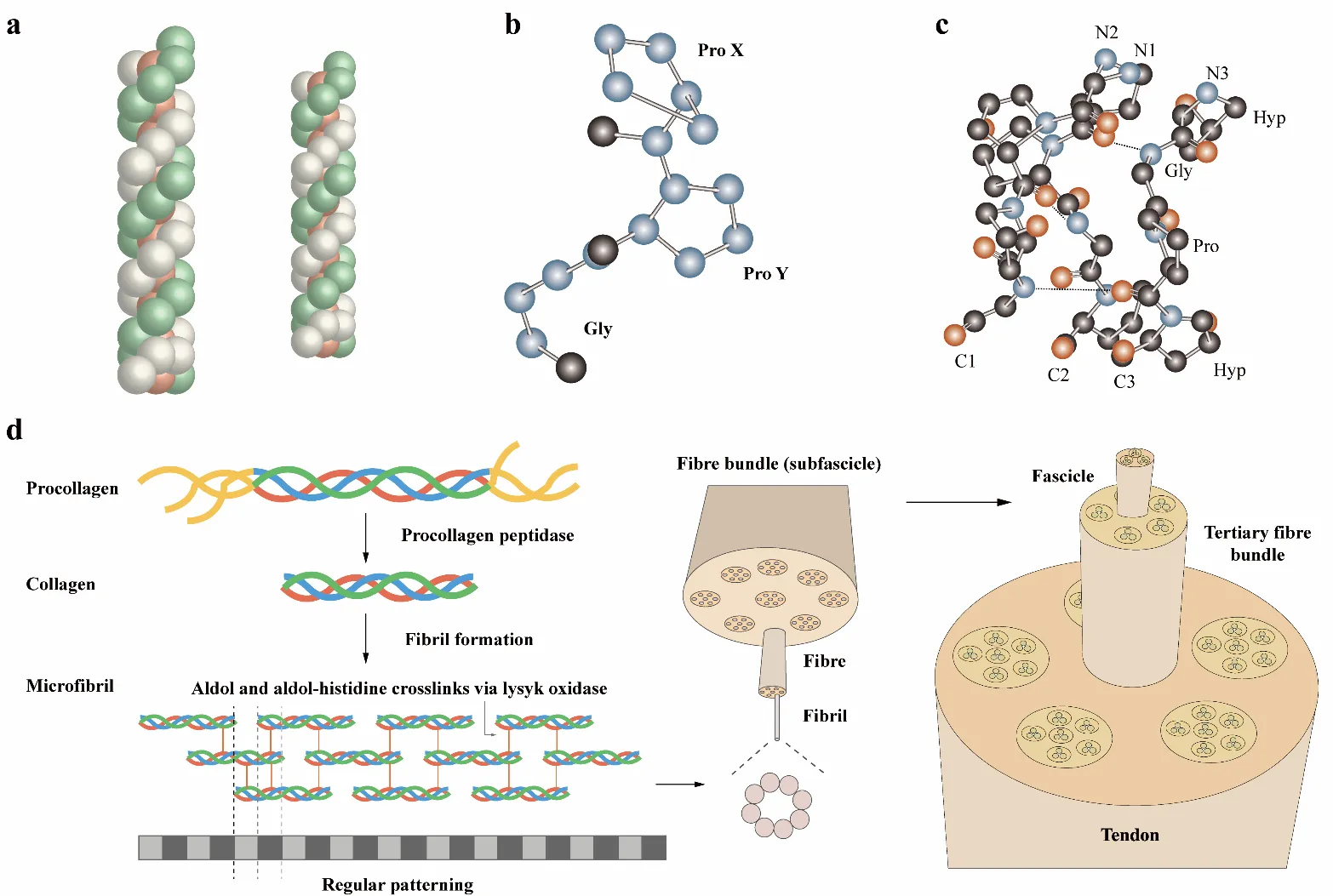
3]. This heterogeneity governs collagen’s physicochemical characteristics. Some types of collagen have a triple-helix structural feature (). These chains intertwine into a superhelix (). The repeating (Gly-X-Y)ₙ motif facilitates tight packing of the helices, where X and Y are frequently proline and hydroxyproline. Hydrogen bonds form between the carbonyl group of the X-position residue in one chain and the amide nitrogen of glycine in an adjacent chain [
4], creating a lattice parallel to the polypeptide backbone and perpendicular to the helical axis [
5].
.
The diversity of the collagen family.
| Collagen Type |
Extraction Difficulty |
Function |
Category |
Collagen Subtypes |
| Collagen I |
No |
Provide tensile strength [6] |
Fibrillar collagen |
Banded fibrils |
| Collagen II |
No |
Provide compressive strength and elasticity [7] |
Fibrillar collagen |
Banded fibrils |
| Collagen III |
No |
Provides support in soft tissues and maintains elasticity [8] |
Fibrillar collagen |
Banded fibrils |
| Collagen IV |
Strong ECM interactions |
Provide scaffolding for epithelial and endothelial layers [9] |
Non-fibrillar collagen |
Network-forming collagens |
| Collagen V |
Strong ECM interactions, abundant in the embryonic stage |
Co-assemble with type I collagen in tissues like skin and placenta [10] |
Fibrillar collagen |
Banded fibrils |
| Collagen VI |
Low abundance, strong ECM interaction |
Mechanical support, cytoprotective function, promotion of tumor growth and progression [11] |
Non-fibrillar collagen |
Beaded microfibrils |
| Collagen VII |
Low abundance, strong ECM interaction |
Provide stability to the dermal-epidermal adhesion [12] |
Non-fibrillar collagen |
Anchoring fibrils |
| Collagen VIII |
Low abundance, strong ECM interaction |
Active roles in angiogenesis and ECM remodeling [13] |
Non-fibrillar collagen |
Short chain collagens |
| Collagen IX |
Low abundance, strong ECM interaction |
Support cartilage integrity and stability [14] |
Non-fibrillar collagen |
FACIT |
| Collagen X |
Low abundance, strong ECM interaction |
Regulate matrix mineralization and compartmentalizing matrix components [15] |
Non-fibrillar collagen |
Fibril-associated collagens |
| Collagen XI |
Abundant in embryonic stage, strong ECM |
Regulate collagen fibrillogenesis [16] |
Fibrillar collagen |
Fibril-forming collagens |
| Collagen XII |
Low abundance, strong ECM interaction |
Stabilize type I collagen fibrils [17] |
Non-fibrillar collagen |
FACIT |
| Collagen XIII |
Transmembrane nature |
Function as an adhesion molecule [18] |
Non-fibrillar collagen |
Transmembrane collagen |
| Collagen XIV |
Abundant in the embryonic stage, strong ECM interactions |
Regulate early stages of fibrillogenesis [19] |
Non-fibrillar collagen |
FACIT |
| Collagen XV |
Low abundance |
Structural link between producing cells and connective tissues [20] |
Non-fibrillar collagen |
Multiplexing |
| Collagen XVI |
Abundant in the embryonic stage, strong ECM interactions |
Support interaction of connective tissue cells with their ECM [21] |
Non-fibrillar collagen |
FACIT |
| Collagen XVII |
Transmembrane nature |
Facilitate epidermal-dermal attachment, a niche for hair follicle stem cells [22] |
Non-fibrillar collagen |
Transmembrane collagen |
| Collagen XVIII |
Abundant in the embryonic stage |
Control blood vessel formation [23] |
Non-fibrillar collagen |
Multiplexing |
| Collagen XIX |
Low abundance, strong ECM interaction |
Unknown functions. Suggest the regulation of cardiac extracellular matrix structure [24] |
Non-fibrillar collagen |
FACIT |
| Collagen XX |
Low abundance, strong ECM interaction |
Unknown functions. It may serve as a biomarker of solid tumors [25] |
Non-fibrillar collagen |
FACIT |
| Collagen XXI |
Low abundance, strong ECM interaction |
Unknown functions. May contribute to the extracellular matrix assembly [26] |
Non-fibrillar collagen |
FACIT |
| Collagen XXII |
Low abundance, strong ECM interaction |
Act as a cell adhesion ligand for skin epithelial cells and fibroblasts [27] |
Non-fibrillar collagen |
FACIT |
| Collagen XXIII |
Transmembrane nature |
Unknown functions. An important biomarker for lung cancer [28] |
Non-fibrillar collagen |
Transmembrane collagen |
| Collagen XXIV |
Low abundance, limited in developing bone, and strong ECM interactions |
Promote fibrillogenesis in bone and cornea [29] |
Fibrillar collagen |
Fibril-forming collagen |
| Collagen XXV |
Low abundance, strong ECM interaction |
Promote fusion of myoblasts into myofibers [30] |
Non-fibrillar collagen |
Transmembrane collagen |
| Collagen XXVI |
Low abundance |
Unknown functions. Suggest to support testis and ovary development [31] |
Non-fibrillar collagen |
FACIT |
| Collagen XXVII |
Abundant in the embryonic stage |
Support calcification of cartilage and the transition of cartilage to bone [32] |
Fibrillar collagen [33] |
Fibril-forming collagen |
| Collagen XXVIII |
Low abundance |
May contribute to neuron protection and support [34] |
Non-fibrillar collagen |
Fibril-forming collagen |
. Fibrillar collagen assembly and the collagen triple helix: (<b>a</b>) Structure of triple helix, consisting of the repeating (ProHypGly)4-(ProHypAla)-(ProHypGly)5 sequence [
35]. (<b>b</b>) ProX-ProY-Gly chain in collagen triple helix [
36]. (<b>c</b>) Three strands in the collagen triple helix stagger together and form a ladder-like pattern of hydrogen bonds. (<b>d</b>) The biosynthesis of collagen begins with procollagen. Collagen molecules assemble into microfibrils in the extracellular matrix [
37,
38].
Collagen plays a pivotal role in mediating interactions between cell and the ECM [
39]. A triple-helical domain characterizes fibrillar collagen. Other collagen types, such as fibril-associated collagen with interrupted triple helices (FACIT), demonstrate the interspersion of triple-helical domains within non-collagenous (NC) domains. These NC domains are crucial for structural assembly and confer biological activity to collagen [
40].
1.1. Fibrillar Collagen
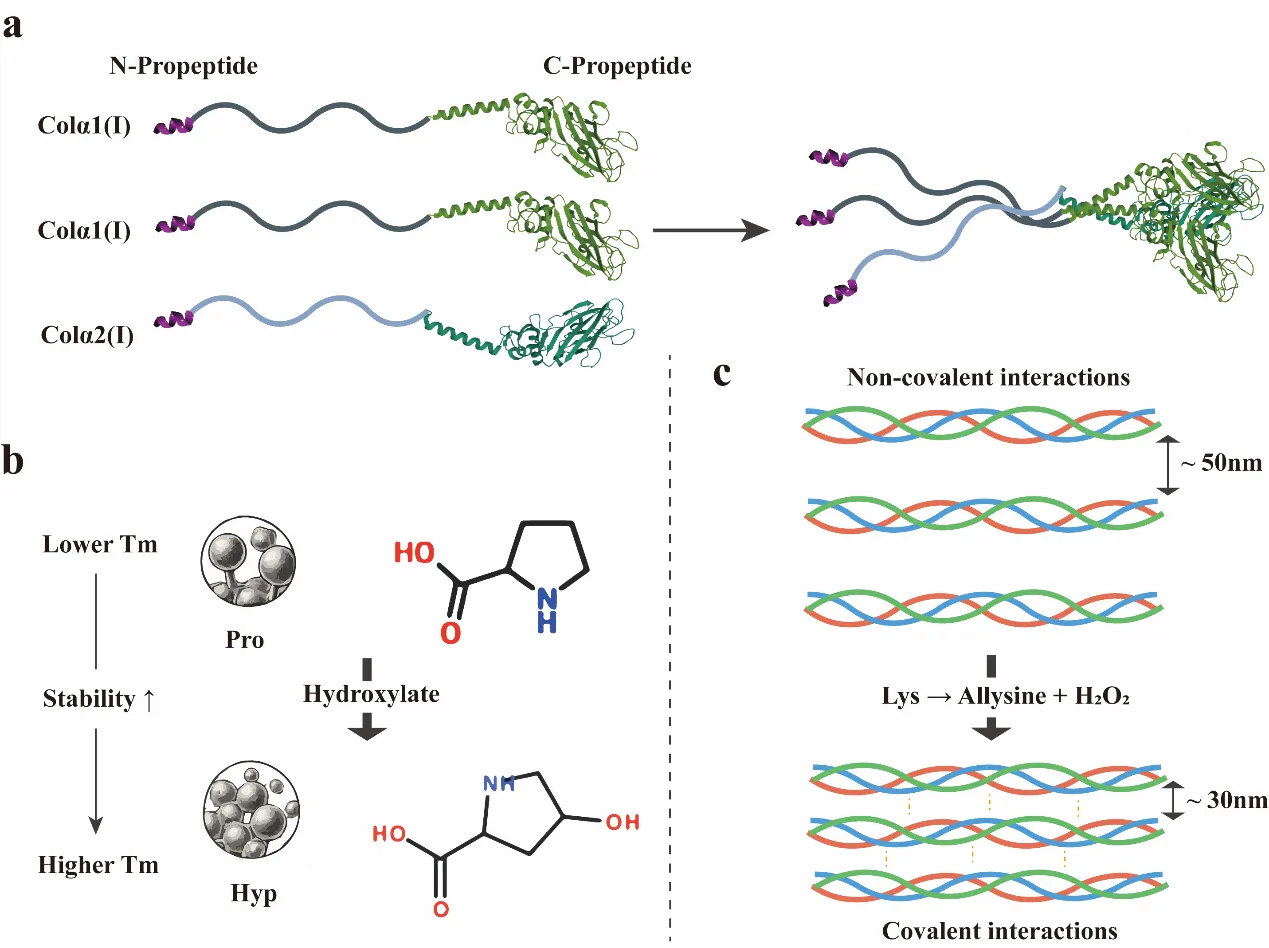
Fibrillar collagen exhibits a hierarchical structure wherein parallel staggered collagen molecules self-assemble into fibrous nanostructures that further aggregate into higher-order assemblies [
41]. Synthesis begins with the production of procollagen precursors containing carboxy-terminal propeptides and signal sequences that direct trafficking to the rough endoplasmic reticulum (ER) [
42]. Within the ER, propeptides undergo hydroxylation of lysine and proline residues by lysyl and prolyl hydroxylases, respectively [
42]. Proline and hydroxyproline play a crucial role in stabilizing the triple helix [
43,
44]. Following lysyl hydroxylation and O-linked glycosylation, α-chains trimerize into procollagen, a process initiated by the C-propeptide domain. This domain recognizes α-chains via a specialized mechanism, forming a stable core that drives triple helix assembly [
45,
46,
47].
The formation of collagen microfibers involves the nucleation, organization, and unidirectional elongation of short primary nanofibers () [
48,
49], which subsequently merge to form microfibrils exhibiting increased longitudinal and axial dimensions [
50,
51]. These supramolecular assemblies undergo stabilization via covalent crosslinking [
52], a process initiated by extracellular lysine oxidases that catalyze the oxidative deamination of lysine residues in target peptides [
53]. Within the microfibrils, the N- and C-termini of adjacent collagen monomers interact and are covalently cross-linked by lysyl oxidase, further enhancing structural stability [
53,
54]. Ultimately, the assembly of collagen microfibers depends on the complex interplay of chemical and physical interactions among its components [
55].
1.2. Non-Fibrillar Collagen
The collagen family also includes non-fibrillar members, such as network-forming collagen IV, which assemble into sheet-like networks rather than fibers [
56]. These collagen IV networks have been vital to the evolution of multicellular organisms [
57]. Collagen IV differs from fibrillar collagen in two key aspects of its higher-order assembly [
58]. First, the C-terminal non-collagenous (NC1) domain of collagen IV is retained during assembly and plays a central role in network formation [
9,
59]. Second, collagen IV features a sequence with a discontinuity in the (Gly-X-Y)n repeat, where glycine is absent or replaced in one of the three residues. This disrupts the formation of continuous triple-helical regions and creates local structural instability [
60], a feature also seen in other non-fibrillar collagens [
4].
The assembly of collagen IV scaffolds is a complex process. Intracellular enzymes collaborate to construct the heterotrimer, but the assembly into a three-dimensional (3D) scaffold occurs extracellularly. After secretion, protomers align via their NC1 and 7S domains, forming critical junctions at the protofibril ends. The triple helix then undergoes superhelical formation through lateral interactions [
61]. During this process, functional molecules are incorporated into the collagen triple helix by embedding binding sites along the protofibril length. These network-forming collagens act as “smart” scaffolds, especially as components of the basement membrane, supporting the development and function of multicellular tissues [
9].
1.3. Biological Function of Collagen
Collagen is vital in modulating a range of signaling pathways essential for development, regeneration, and tissue repair, thus maintaining tissue homeostasis. Its interaction with cells is primarily mediated by specific receptors facilitating bidirectional transmission of mechanical and biochemical signals through cytoskeleton-mediated processes (). Key receptors include integrins and discoidin domain receptors (DDR1 and DDR2) [
62,
63].
Integrins are heterodimeric receptors present on nearly all cell types and serve as major mediators for extracellular matrix components, including collagen. They play pivotal roles in regulating cell signaling, migration, survival, and differentiation [
63,
64]. Four collagen-binding integrins, α1β1, α2β1, α10β1, and α11β1, have been identified within the integrin α1 domain subgroup [
65,
66,
67]. Although they share the function of collagen receptors, they are expressed in different cell types and mediate distinct biochemical signals. For example, α1β1 integrin interacting with collagen triggers biological responses such as Grb2 recruitment, MAPK activation for cell proliferation, FAK phosphorylation facilitating fibroblast-to-myofibroblast differentiation, and activation of the Shc-mediated pathway in skin regeneration.
Receptor tyrosine kinases, especially DDR1 and DDR2, are also activated upon collagen binding [
68,
69]. DDR1 is mainly expressed in epithelial cells, while DDR2 is found in fibroblasts and mesenchymal cells. Unlike integrins, DDRs mediate ECM signaling unidirectionally. DDR2’s interaction with collagen II is indirectly regulated by integrin/cytokine pathways and AGE-mediated signaling [
70,
71]. DDR2-bound collagen in the ECM activates the JNK/MAPK and PI3K/Akt pathways, influencing cell proliferation, survival, and gene expression. DDR1 binding to collagen activates JNK, NF-κB, p38, ERK1/2 MAPKs, and PI3K/Akt signaling pathways. DDR1 inactivation leads to interactions with E-cadherin, promoting cell-cell contact. Both collagen-mediated cell-matrix communication and collagen-independent cell-cell interactions influence the triggering of diverse signaling pathways through DDRs [
72].
.
Receptors related to Collagen.
| Receptor |
Type |
Distribution |
Biological Regulation |
Ligand Binding Specificity |
| Integrins |
α1β1 |
Fibroblasts,
Mesenchymal tissues
|
Wound healing; Regulates the proliferation of living cells, MMP expression, and collagen synthesis; Fibroblast to myofibroblast differentiation; Invasion and growth of hepatocellular carcinoma |
Collagens I, III, IV, IX, XIII, XVI, and the collagen IV chain–derived peptide arresten [73,74,75] |
| α2β1 |
Platelets, epithelium, Fibroblasts, and Mesenchymal tissues |
Platelet adhesion to collagen; Hepatocellular carcinoma invasion and growth; Wound healing |
Collagens I, III, IV, V, XI, XVI, and XXIII [76,77,78] |
| α10β1 |
Cartilage and Chondrocytes |
Chondrogenic differentiation; Cartilage repair, Skeletal growth |
Collagen II and IX [78] |
| α11β1 |
Periodontal ligaments |
Wound healing; Cell migration; Mediates the contraction of collagen lattices; Myofibroblast differentiation |
Collagen I and XIII [79,80] |
| Receptor Tyrosine Kinases (DDR) |
DDR1 |
Epithelial cells, Smooth muscle cells, Fibroblasts, Oligodendrocytes and Macrophages |
Development and growth of organs; Cell proliferation, survival, homing, and colonization; Inhibits tumor growth |
Collagen I–V [81] |
| DDR2 |
Chondrocytes |
Development and growth of organs; Development of bone and cartilages; Pathological process of arthritis, wound healing, dwarfism, and tumor |
Collagen I–III, and V[81] |
| Immunoglobulin Receptor |
GPVI |
Megakaryocytes and Platelets |
Wound healing |
Collagen I–III [82] |
| OSCAR |
A wide range of myeloid cells |
Osteoclast growth induction for bone resorption; Osteoclast differentiation |
Collagen I and II [83] |
| Leukocyte Receptor Complex (LRC) |
LRC |
Immune cells |
Autoimmunity; Antiviral immunity; Graft tolerance; Regulates osteoclast differentiation. |
Collagen I and III [84] |
| Other Receptors |
Fibronectin |
Extracellular matrix, Plasma, and Cell surface |
Tissue growth; Wound repair; Fibroblast migration; Nerve regeneration stabilization; Extracellular matrix and embryogenesis; cell-to-cell adhesion |
Collagen I and III [85] |
| Vitronectin |
Extracellular matrix, Blood serum, Platelets, and Bone. |
Cell proliferation; Adhesion; Immune defense; Hemostasis; Fibrinolysis |
Collagen I [86] |
| uPARAP |
Mesenchymal cell surface, Osteocytes, and Osteoblasts |
Fibroblast migration; Primary adhesion of collagen to fibroblasts |
Collagen I, II, IV, and V [87] |
2. From Native Collagen to Protein Engineering
The isolation of collagen from natural tissues laid the foundation for its application in biomedicine, yet the inherent limitations of native collagen extraction have catalyzed the evolution toward engineered alternatives. Pioneering work by Lister and Macewen utilizing sheep intestinal collagen sutures [
4,
88] established collagen’s biocompatibility, while its subsequent use as cell culture matrices [
89] revealed structural dependency on tissue-specific supramolecular assemblies [
90,
91].
Traditional extraction protocols predominantly utilize mammalian sources like bovine, pig skin, and rat tail tendon, and marine byproducts like fish through acidic/alkaline hydrolysis or enzymatic digestion [
92,
93,
94]. Collagen from bovine Achilles tendon [
95,
96] and porcine skin [
97] dominated early biomaterial development [
98,
99,
100].
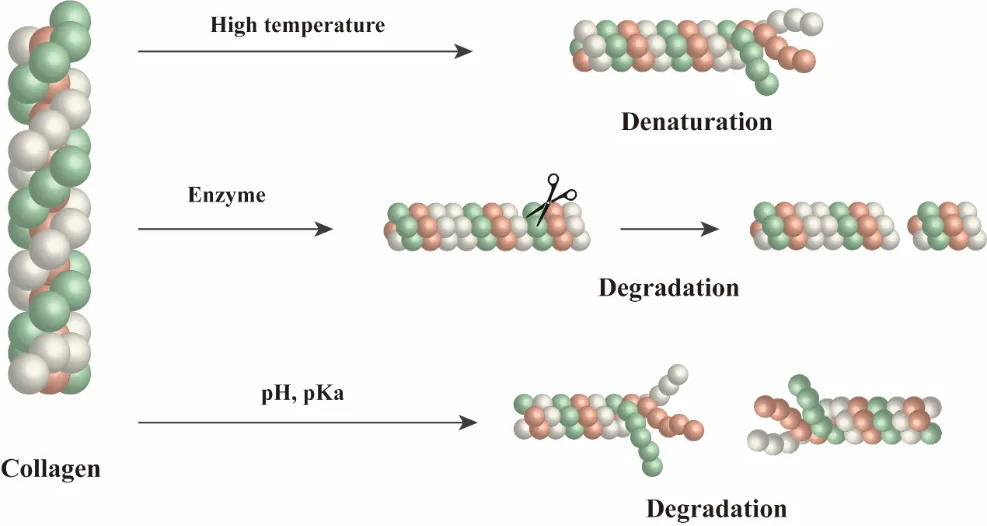
Acid and alkaline extraction methodologies inevitably perturb these critical structural determinants through three primary mechanisms (). First, thermal denaturation at temperatures exceeding 40℃ induces unwinding of the triple helix into disordered random coils, leading to an 85% reduction in tensile strength compared to native fibrillar collagen [
90]. Second, enzymatic hydrolysis using proteases such as pepsin preferentially cleaves non-helical telopeptide regions, generating fragmented polypeptides (3–6 kDa) with compromised cell-binding RGD motifs essential for integrin-mediated signaling [
101,
102,
103]. Third, the hydrolysis of lysyl oxidase-catalyzed pyridinoline crosslinks, as evidenced by the decrease in Young’s modulus [
99].
. Structural Degradation During Natural Collagen Extraction: There are three principal mechanisms that compromise collagen’s structural integrity during the extraction process: thermal denaturation, enzymatic hydrolysis, and cross-link disruption.
To overcome these limitations, collagen mimetic peptides (CMPs) have emerged as engineered alternatives that recapitulate core triple-helical motifs while enabling programmable functionality. Recent advances demonstrate that CMPs can be rationally designed to resist thermal denaturation through strategic incorporation of unnatural amino acids or covalent cross-linking [
104]. Notably, π-system end-capping strategies stabilize short CMPs with only 3–6 repeats, achieving melting temperatures up to 76 °C [
105]. Furthermore, functional domains (e.g., GFOGER for integrin binding) can be embedded without disrupting fibril morphology, restoring cell-adhesion activity lost in enzymatically hydrolyzed collagen fragments [
106,
107]. These approaches collectively enable the synthesis of chemically complex, well-controlled collagen mimetic biomaterials.
3. Biological Synthesis of Recombinant Collagen
The drawbacks of native collagen extraction, such as the risk of pathogen transmission, immunogenicity, structural heterogeneity, and limited scalability, have prompted a shift toward recombinant collagen production [
108] (
). Recombinant strategies enable control over amino acid sequence, post-translational modifications (PTMs), and molecular architecture, allowing the fabrication of collagen with tunable properties (
).
Escherichia coli is widely favored among expression systems for its rapid proliferation, low production cost, and high-density fermentation capability [
109]. However, the lack of endogenous prolyl-4-hydroxylase (P4H) in
E. coli impairs hydroxyproline synthesis, thereby limiting triple-helix thermal stability [
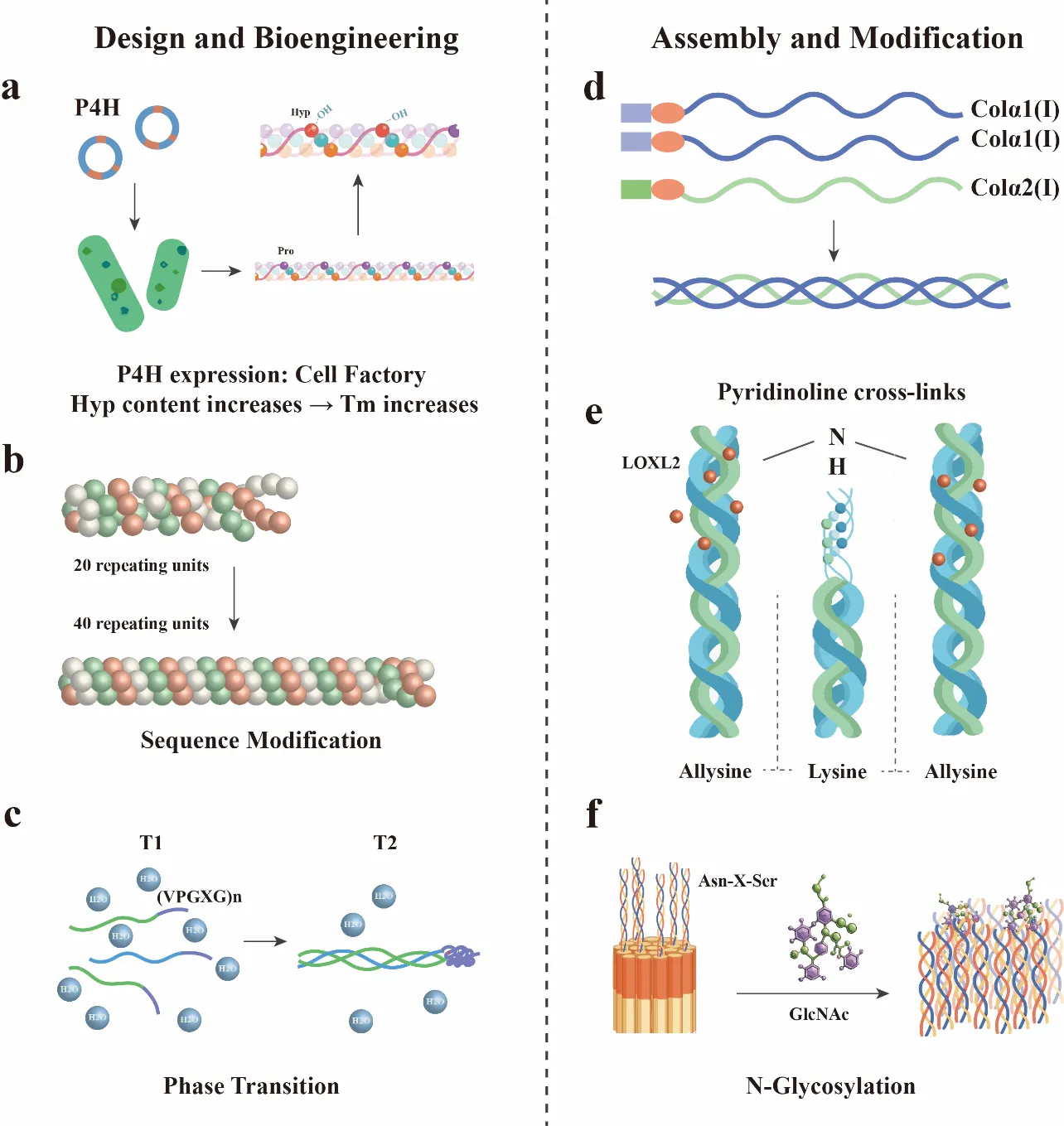
110]. To overcome this, researchers have implemented heterologous coexpression of P4HA and P4HB from
Caenorhabditis elegans, increasing hydroxyproline content to 15% and raising the collagen melting temperature (T
m) by approximately 7 °C [
111,
112,
113]. Further enhancements involve rational sequence engineering, extending Gly-X-Y repeats from 20 to 40 units to improve interchain packing, and the fusion of elastin-like polypeptides (ELPs) to the N-terminus [
114], enabling temperature-responsive trimerization with over 80% efficiency at physiological temperature [
115].
. Protein Engineering Strategies for Collagen Stabilization: (<b>a</b>) C-propeptide-mediated heterotrimeric assembly of type I collagen. (<b>b</b>) Hydroxyproline-mediated stabilization of triple-helix thermodynamics. (<b>c</b>) Enzymatic crosslinking enhances fibrillar shear resistance.
Yeast expression systems, particularly
Pichia pastoris, offer a more conducive environment for collagen biosynthesis with appropriate PTMs. Coexpression of human P4HA1/P4HB achieves hydroxyproline levels up to 88% of native collagen [
116,
117] while optimized dual-promoter constructs allow stoichiometric expression of pro-α1 (I) and pro-α2 (I) chains for heterotrimer formation [
118]. Nonetheless, purification of heterotrimers at >65% remains a technical challenge. Industrial-scale fed-batch fermentation faces challenges in dissolved oxygen management, where maintaining >30% saturation without oxidative stress requires fine-tuned bioreactor control. Additionally, the production process in yeast systems often requires strict control of methanol induction, pH stability, and osmolarity to avoid protein aggregation and ensure correct folding. These parameters are dynamically regulated through feedback-controlled fermentation systems integrating real-time methanol sensing, DO and pH monitoring, and osmotic pressure balancing. Strategies such as pulse-wise methanol feeding (1.0–2.0 g/L), pH stabilization at 5.0 ± 0.1, and osmolyte supplementation (e.g., sorbitol, betaine) have enhanced solubility and reduced aggregation [
117]. Additionally, additives like sodium pyruvate can boost TCA cycle flux, increasing collagen yield by over 20% and shortening induction time. Industrial-scale systems often employ high-density fed-batch protocols (>150 g DCW/L) with optimized agitation-oxygen transfer to maintain protein quality [
119]. Use of engineered strains with enhanced secretory pathways, or those co-expressing molecular chaperones, has shown promise in improving the yield and solubility of full-length collagen proteins. Recent progress also includes the coexpression of lysyl oxidase homologs (e.g., LOXL2) [
120] to introduce enzymatic crosslinking and targeted N-glycosylation at Asn-X-Ser motifs to improve proteolytic resistance and in vivo stability [
121]. Moreover, in vitro hydroxylation using purified P4H enzymes has emerged as a complementary strategy for microbial-expressed collagen, allowing post-expression modification of Hyp content without the need for endogenous enzymatic activity. (
) Mammalian expression systems such as CHO and HEK293 cells represent the advancements for producing recombinant collagen with near-native fidelity, particularly in PTMs and structural assembly [
122,
123,
124]. The production cost remains higher than that of microbial systems, limiting application in high-volume products such as scaffold matrices. Emerging alternatives include stem cell-derived extracellular matrix secretion without genetic modification, wherein extracellular niche modulation induces the synthesis of human ECM collagen (hCol) with native-like molecular weights (α1 ~132 kDa; α2 ~122 kDa), glycosylation profiles, and fibrillar periodicity [
124]. This strategy bypasses transgene-associated regulatory hurdles and offers immunologically safer products. Regarding function and safety, the residues of terminal peptides and non-human glycosylation may trigger an immune response. Studies have been conducted to remove N/C-terminal propeptide and telopeptide from recombinant type I collagen, and the endotoxin is controlled within 0.05 EU/mg to ensure the function and safety [
125,
126].
. Engineering Strategies for Collagen Biosynthesis. Design and engineering: (<b>a</b>) Cell factory for P4H expression; (<b>b</b>) Sequence Modification; (<b>c</b>) ELP phase separation. Assembly and Modification: (<b>d</b>) Chain Stoichiometry Control; (<b>e</b>) LOXL2-Mediated Crosslinking; (<b>f</b>) Directed N-Glycosylation.
Complementing these bioproduction systems, synthetic biology and AI-guided protein design rapidly advance collagen engineering. Platforms such as ColDiff and ColGen-GA apply diffusion-based modeling and genetic algorithms to design stable, self-assembling collagen variants with tailored biofunctions [
127]. AI-assisted strategies in collagen sequence design, structure prediction, and assembly optimization are illustrated in Section 4. These tools enable the rational construction of collagen domains for diverse biomedical applications [
128]. Beyond domain-level sequence design, AI-driven simulation platforms now integrate structural dynamics and protein interaction predictions to assess fibrillogenesis potential or improve integrin-binding affinity.
Collectively, recent breakthroughs such as the development of self-assembling recombinant collagen hydrogels without chemical crosslinkers [
129,
130], high-temperature-resistant triple helix structures stabilized by cold-adapted chaperones, and coexpression of folding enhancers have substantially addressed many technical barriers [
131,
132]. However, challenges remain in ensuring batch consistency, achieving uniform post-translational modifications, and meeting regulatory standards for clinical application.
.
The fabrication and biosynthesis of collagen.
| Type of Collagen |
Host |
Yield |
Comments |
Concerned Contaminants |
| Natural Collagen |
_ |
_ |
Obtained by enzymatic action; Immunogenicity, pathogen transmission issues; Human collagen extraction is limited and expensive |
Residual host proteins; chemical residues; pathogenic microorganisms [92] |
| Recombinant Human Collagen |
E. coli |
90 mg/L |
Coexpression with mimivirus prolyl and lysyl hydroxylases |
Endotoxins (from E. coli LPS); media residues [113] |
| P. pastoris |
0.6 g/L |
Coexpression with human prolyl hydroxylases in a bioreactor with constant oxygen supply |
Cell wall components (β-glucans, Mannans); media residues [117] |
| Mammalian cells (293-EBNA) |
0.5–80 mg/L |
Coexpression with P4H subunits is not required except for collagens X and VIII expression. Low yields of collagen V heterotrimers |
Mycoplasma contamination; Viral contaminants [121,127] |
| Plants (tobacco) |
30 mg/kg |
Coexpression with P4H subunits to obtain hydroxylated collagen |
Mycotoxins and contaminants from fungal infections; heavy metals; pesticides/herbicides [114,116] |
| Transgenic maize seeds |
4 mg/kg; 12 mg/kg |
Co-expressed with/without both the α- and β- subunits of a recombinant human P4H (rP4H). |
| Drosophila melanogaster S2 cells |
10–50 mg/L |
Production of collagen I and IX heterotrimers |
Mycoplasma contamination; distinct glycosylated products [133,134] |
4. AI-Assisted Design and Modeling of Collagen
4.1. Advances in AI-Driven Protein Engineering
Recent advances in deep learning have enabled the
de novo design of synthetic collagen with programmable properties. ProtSeed, a sequence-structure co-design framework, has shown a 3.2-fold increase in heterotrimer assembly efficiency over random sequences by optimizing charge complementarity and steric compatibility at glycine-X-Y junction [
135]. Progress in deep learning has transformed collagen structural prediction and design. The AlphaFold Multimer model has shown excellent performance in predicting protein complex structures, with prediction accuracies of 67% and 69% at the interface between hetero and homo oligomers, respectively, when tested on 4433 protein complexes [
136]. The progress in geometric deep learning and the application of models have enhanced our ability to predict and design collagen structures, leading to the development of new collagen-based materials and therapies with tailored properties.[
137,
138,
139]
4.2. AI-Assisting Design of Collagen Variants
Generative artificial intelligence (AI) technologies have opened new window for the
de novo design of collagen. Tools based on autoregressive models and Transformer architectures [
131,
132,
133], such as ProteinMPNN [
140] and ESM-IF [
141], have enhanced sequence design precision by integrating geometric features, such as torsion angles and backbone vectors, and geometric vector perceptrons (GVPs), particularly excelling in tasks requiring sub-nanoscale control, such as collagen fibril assembly [
142]. For instance, the diffusion model ColDiff, combined with a supervised learning strategy, extracts sequence features from human collagen multi-omics data to generate collagen-mimetic peptides (CMPs) with GXY repeat structures, achieving a Pearson correlation of up to 0.95 (natural collagen) and 0.8 (synthetic CMPs) between predicted and experimental melting temperatures (T
m) [
143]. Additionally, the synergistic application of genetic algorithms (GA) and deep learning, like ColGen-GA, enables rapid generation of homotrimeric type I collagen sequences (1000 sequences in 8 h), with T
m prediction errors less than 5%, outperforming traditional molecular dynamics simulations [
144].
Sequence design based on generative models requires modular strategies combined with experimental validation to optimize biological functionality. The adhesion module guides self-assembly into periodic banded fibers through hydrophobic or electrostatic interactions, while the functional module, based on Streptococcus Scl2 collagen-like protein fragments, introduces osteoblast-binding domains without disrupting fiber morphology [
145]. Coarse-grained simulations and atomic modeling elucidate the hierarchical assembly of collagen triple helices into fibrils via gap-overlap stacking, consistent with experimental observations [
146,
147]. Experimental validation shows that such synthetic collagen achieves tensile strength approaching 50 MPa comparable to natural type I collagen and promotes osteogenic precursor cell differentiation, increasing alkaline phosphatase activity by over 2-fold [
148]. Computational design strategies (e.g., reinforcement learning) enable the generation of collagen-mimetic peptides (CMPs) with high self-assembly propensity, though experimental validation remains critical [
149]. Short CMPs can form hydrogels at low concentrations, while long-chain variants require proline hydroxylation (up to 90% in yeast systems) for conformational stability [
143], highlighting the necessity of post-translational modifications in biomimetic design.
4.3. AI-Assisting Optimization of Collagen Protein
AI technologies enable quantitative optimization of collagen material properties through high-throughput screening and molecular engineering strategies. The ColGen-GA framework identifies key GXY triplets contributing to thermal stability by analyzing millions of generated sequences: sequences containing (GPO)
14 exhibit the lowest ΔT
m values (T
m reduction of 3 ℃) [
144]. Experimental validation further demonstrates that AI-designed recombinant CMPs achieve secretion efficiencies of 0.1–0.2 mg/mL in
Pichia pastoris, with characteristic CD spectral peaks at 220–222 nm and a 40% improvement in cell adhesion efficiency compared to traditional collagen. Moreover, AI-driven optimization of collagen extraction processes, such as enzyme concentration, temperature, and pH value, reduces energy consumption by 40% and waste by 45%, while enabling collagen recovery from waste materials such as fish scales and bovine hides, showcasing end-to-end efficiency from molecular design to industrial production [
150,
151]. Future efforts should focus on integrating multimodal data, sequence-structure-function, to enhance model generalizability and leveraging automated experimental platforms, such as iBioFoundry, to complete the “design-synthesis-testing” loop [
152].
5. Engineering on Collagen Biomaterials
Contemporary molecular engineering strategies enable the precise customization of collagen biomaterials through domain-specific modifications and advanced crosslinking architectures. These approaches have unlocked transformative potential for synthetic collagens across a range of biomedical applications (
). For example, incorporating cell-binding RGD motifs or MMP-sensitive cleavage sites into collagen scaffolds improves cell-material interactions, a fibrinogen-collagen hybrid hydrogel demonstrated a minimum wound closure rate of 83.3% compared to natural collagen with 69.4% [
153]. Similarly, VEGF peptide-functionalized scaffolds enhance vascular regeneration by increasing surface wettability, inducing VEGF receptor phosphorylation, and promoting HUVEC survival and proliferation [
154]. These scaffolds, fabricated via simple polymer mixing methods, hold promise for sustaining endothelial cell viability during vascular network formation, potentially improving transplanted tissue survival rates. Beyond regenerative medicine, engineered collagens are facilitating oncology research. Collagen-based 3D tumor models with tunable stiffness recapitulate tumor microenvironment mechanics, enhancing immune checkpoint marker expression and aligning drug response profiles with clinical observations [
155,
156]. The aberrant crosslinking in pathological matrices revealed by AGE mediated collagen fiber binding in liver cirrhosis, can impair remodeling ability. The fibers crosslinked by AGE form coarse bundles, with a 3-fold increase in diameter and a 60% decrease in macrophage remodeling efficiency, promoting fibrosis progression through cytoskeletal disorder and type II immune polarization [
157]. This emphasizes the necessity of precise crosslinking control in engineering supports.
Beyond its conventional biomedical applications, recombinant collagen emerges as a transformative material in intelligent drug delivery systems and tissue engineering. Recent technological advancements have capitalized on collagen’s programmable biodegradability to develop stimuli-responsive nanocarriers. pH-sensitive collagen nanocapsules have demonstrated remarkable tumor-specific drug release efficiency, achieving 90% drug release through lysosomal acidity-triggered dissolution at pH 5.5 [
158]. Light-responsive elastin-like peptide nanoparticles have enabled spatiotemporal control of targeted cellular delivery through near-infrared-induced phase transitions, enhancing targeting efficacy by two-fold [
159].
In the realm of tissue engineering, recombinant collagen has exhibited good biocompatibility and cell activity promotion in skin wound repair applications. The incorporation of recombinant collagen into GelMA (gelatin methacryloyl) has enhanced cellular activity and migration capacity, thereby accelerating wound healing processes [
160].
In vivo experiments demonstrated that wounds treated with recombinant collagen-modified GelMA achieved an 80% healing rate within 14 days, representing a 1.2-fold improvement compared to untreated diabetic mice. The mechanical and self-healing properties of collagen-based hydrogels can be optimized through the strategic network and chain topology design [
161,
162,
163]. The mechanical optimization carried out through network topology design now combines with a hyperelastic framework, with a self-recovery rate of>95% after 500 cycles, and a compression tension elastic ratio adjusted to 1.5, approaching natural tissue characteristics [
164].
.
Biomedical Applications of Collagen-based Biomaterials.
| Applications |
Tissue |
Origin |
Crosslinking Agents |
Bioharzard of Crosslinking Agents |
| Wound dressing |
Epidermal and dermal acellular scaffolds |
Synthetic/ Natural |
1,4-butanediol diglycidyl ether (BDDGE)/ EDC |
Contact dermatitis, allergic[120,121] |
| Hydrogels based on human-like collagen and carboxyl pullulan |
Synthetic |
Butanediol-diglycidyl ether (BDDE) |
| Tendon Repair |
Collagen-glycosaminoglycan scaffold |
_ |
Acrylonitrile butadiene styrene (ABS) |
Not an irritant [165] |
| Treatment of Intervertebral Disc Degeneration and Cartilage Repair |
Fibrillized jellyfish collagen and alginate hydrogel |
Natural |
EDC |
Skin irritant [122] |
| Self-assembled fibrocartilage |
Natural |
Lysyl oxidase like-2 (LOXL-2) |
Not an irritant [166] |
| Type II collagen-hyaluronic acid hydrogel |
Synthetic |
EDC |
Skin irritant [167] |
| Type II collagen scaffold/chondroitin sulfate composite gel |
Natural |
Genipin |
LD50: 237 mg/kg (oral route) for mice [168,169,170] |
| Drug Delivery |
Growth factor-conjugated fibrin microbeads |
Synthetic |
Genipin |
| Collagen-hydroxyapatite scaffolds/Collagen-chitosan-graphene oxide mixture |
Synthetic |
EDC/N-hydroxysuccinimide (NHS) |
An irritant/harmful by ingestion [171,172] |
| Treatment of Cardiovascular Diseases |
Bovine pericardial |
_ |
Dye-mediated photooxidation |
Not an irritant [173] |
| Decellularized carotids from a newborn calf |
Synthetic/ Natural |
Co-crosslinking with procyanidins and glutaraldehyde |
Skin irritant; may induce asthma [142] |
| Alginategelatin-polysaccharide scaffold |
Synthetic |
Glutaraldehyde |
Skin irritant; may induce asthma [174] |
| Bone Tissue Engineering |
Decellularized osteochondral plug from pigs |
Natural |
Epigallocatechin-3 gallate (EGCG) |
Not an irritant [137] |
| Recombinant peptide based on human collagen type I |
Synthetic/ Natural |
Hexamethylene diisocyanate/ Genipin |
Skin irritant/LD50: 237 mg/kg (oral route) for mice [138] |
| Collagen-glycosaminoglycan scaffold with/without mineral content |
Synthetic |
EDC/NHS |
An irritant/ harmful by ingestion [139] |
| Neural Tissue Engineering |
Collagen/Heparin sulfate scaffold |
_ |
UV light [175] |
_ |
| Type I collagen from tendons |
Natural/ Synthetic |
Genipin/Glutaraldehyde |
LD50: 237 mg/kg (oral route) for mice /skin irritant; may induce asthma [140,176] |
| Tissue Regeneration |
Type I/II collagen composite scaffold |
Synthetic |
EDC |
Skin irritant [141] |
| Collagen scaffold |
Synthetic |
Sulfosuccinimidyl
4-(N-maleimidomethyl) cyclohexane-1-carboxylate
|
Skin irritant [177] |
| type I collagen |
Natural |
Riboflavin 5′ monophosphate (FMN) |
Not an irritant [178] |
| Treatment of corneal diseases |
Enucleated rabbit corneas |
_ |
Rose bengal and green light [179] |
_ |
| Dentistry |
Demineralized human dentin |
Natural |
Plant-derived polyphenols [180] |
_ |
| Bovine collagen |
Natural |
Epicatechin |
Skin irritant [181] |
6. Summary and Perspectives
Collagen, the most abundant structural protein in the human body, has become a cornerstone of biomedical engineering. Its hierarchical architecture, biocompatibility, and multifunctional versatility make it highly valuable. This review traces collagen’s evolution from its native biological roles to its applications in engineered biomaterials, focusing on molecular design principles, synthetic strategies, and computational innovations that overcome the limitations of natural collagen extraction. Key advancements include developing recombinant expression systems with enhanced thermal stability and AI-driven design platforms exploring new biomaterials.
The integration of computational design, synthetic biology, and advanced manufacturing is pioneering collagen engineering. Next-generation collagen materials programmable in mechanics, biodegradation, and bioactivity will be transferred into regenerative medicine, oncology models, and smart drug delivery systems. However, achieving this vision requires interdisciplinary progress to align molecular-scale innovation with clinical and industrial requirements.
Acknowledgments
We also would like to thank the iBiofoundary and Core Facility of Institute for Intelligent Bio/Chem Manufacturing, and ZJU-Hangzhou Global Scientific and Technological Innovation Center, Zhejiang University.
Author Contributions
Conceptualization, Y.Y. and X.Z.; Software, Z.W. and C.Z.; Writing—Original Draft Preparation, X.Z.; Writing—Review & Editing, X.Z., Y.Y. and Q.Z., X.L., B.L.; Visualization, X.Z.; Supervision, Y.Y.; Project Administration, Y.Y.; Funding Acquisition, Y.Y.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The datasets generated during the current study are available from the corresponding author on reasonable request.
Funding
This research was supported by Zhejiang Provincial Natural Science Foundation of China under Grant (No. LR25C100001), ‘Pioneer’ and ‘Leading Goose’ R&D Program of Zhejiang Province (No. 2025C01100, No. 2025C01102), the National Natural Science Foundation of China General Program (No.32471372).
Declaration of Competing Interest
The authors declare that they have no competing interests.
References
-
1.
Granito RN, Custódio MR, Rennó ACM. Natural Marine Sponges for Bone Tissue Engineering: The State of Art and Future Perspectives.
J. Biomed. Mater. Res. B Appl. Biomater. 2017,
105, 1717–1727. doi:10.1002/jbm.b.33706.
[Google Scholar]
-
2.
Viguet-Carrin S, Garnero P, Delmas PD. The Role of Collagen in Bone Strength.
Osteoporos. Int. 2006,
17, 319–336. doi:10.1007/s00198-005-2035-9.
[Google Scholar]
-
3.
Ricard-Blum S. The Collagen Family.
Cold Spring Harb. Perspect. Biol. 2011,
3, a004978. doi:10.1101/cshperspect.a004978.
[Google Scholar]
-
4.
Shoulders MD, Raines RT. Collagen Structure and Stability.
Annu. Rev. Biochem. 2009,
78, 929–958. doi:10.1146/annurev.biochem.77.032207.120833.
[Google Scholar]
-
5.
Fallas JA, O’Leary LER, Hartgerink JD. Synthetic Collagen Mimics: Self-Assembly of Homotrimers, Heterotrimers and Higher Order Structures.
Chem. Soc. Rev. 2010,
39, 3510. doi:10.1039/b919455j.
[Google Scholar]
-
6.
Kannus P. Structure of the Tendon Connective Tissue.
Scand. J. Med. Sci. Sports 2000,
10, 312–320. doi:10.1034/j.1600-0838.2000.010006312.x.
[Google Scholar]
-
7.
Hu X, Wu Z, Zhang Z, Yao H, Wang D-A. Type II Collagen Scaffolds for Tissue Engineering.
Commun. Mater. 2024,
5, 149. doi:10.1038/s43246-024-00598-x.
[Google Scholar]
-
8.
Kuivaniemi H, Tromp G. Type III Collagen (COL3A1): Gene and Protein Structure, Tissue Distribution, and Associated Diseases.
Gene 2019,
707, 151–171. doi:10.1016/j.gene.2019.05.003.
[Google Scholar]
-
9.
Brown KL, Cummings CF, Vanacore RM, Hudson BG. Building Collagen IV Smart Scaffolds on the Outside of Cells.
Protein Sci. 2017,
26, 2151–2161. doi:10.1002/pro.3283.
[Google Scholar]
-
10.
Malak TM, Ockleford CD, Bell SC, Dalgleish R, Bright N, Macvicar J. Confocal Immunofluorescence Localization of Collagen Types I, III, IV, V and VI and Their Ultrastructural Organization in Term Human Fetal Membranes.
Placenta 1993,
14, 385–406. doi:10.1016/S0143-4004(05)80460-6.
[Google Scholar]
-
11.
Di Martino A, Cescon M, D’Agostino C, Schilardi F, Sabatelli P, Merlini L, et al. Collagen VI in the Musculoskeletal System.
Int. J. Mol. Sci. 2023,
24, 5095. doi:10.3390/ijms24065095.
[Google Scholar]
-
12.
Chung HJ, Uitto J. Type VII Collagen: The Anchoring Fibril Protein at Fault in Dystrophic Epidermolysis Bullosa.
Dermatol. Clin. 2010,
28, 93–105. doi:10.1016/j.det.2009.10.011.
[Google Scholar]
-
13.
Li Q, Tintut Y, Demer LL, Vazquez-Padron RI, Bendeck MP, Hsu JJ. Collagen VIII in Vascular Diseases.
Matrix Biol. 2024,
133, 64–76. doi:10.1016/j.matbio.2024.08.006.
[Google Scholar]
-
14.
Carlsen S, Nandakumar KS, Holmdahl R. Type IX Collagen Deficiency Enhances the Binding of Cartilage-Specific Antibodies and Arthritis Severity.
Arthritis Res. 2006,
8, R102.
[Google Scholar]
-
15.
Shen G. The Role of Type X Collagen in Facilitating and Regulating Endochondral Ossification of Articular Cartilage.
Orthod. Craniofac. Res. 2005,
8, 11–17. doi:10.1111/j.1601-6343.2004.00308.x.
[Google Scholar]
-
16.
Sun M, Luo EY, Adams SM, Adams T, Ye Y, Shetye SS, et al. Collagen XI Regulates the Acquisition of Collagen Fibril Structure, Organization and Functional Properties in Tendon.
Matrix Biol. 2020,
94, 77–94. doi:10.1016/j.matbio.2020.09.001.
[Google Scholar]
-
17.
Papanicolaou M, Parker AL, Yam M, Filipe EC, Wu SZ, Chitty JL, et al. Temporal Profiling of the Breast Tumour Microenvironment Reveals Collagen XII as a Driver of Metastasis.
Nat. Commun. 2022,
13, 4587. doi:10.1038/s41467-022-32255-7.
[Google Scholar]
-
18.
Heikkinen A, Tu H, Pihlajaniemi T. Collagen XIII: A Type II Transmembrane Protein with Relevance to Musculoskeletal Tissues, Microvessels and Inflammation.
Int. J. Biochem. Cell Biol. 2012,
44, 714–717. doi:10.1016/j.biocel.2012.01.024.
[Google Scholar]
-
19.
Tao G, Levay AK, Peacock JD, Huk DJ, Both SN, Purcell NH, et al. Collagen XIV Is Important for Growth and Structural Integrity of the Myocardium.
J. Mol. Cell. Cardiol. 2012,
53, 626–638. doi:10.1016/j.yjmcc.2012.08.002.
[Google Scholar]
-
20.
Bretaud S, Guillon E, Karppinen S-M, Pihlajaniemi T, Ruggiero F. Collagen XV, a Multifaceted Multiplexin Present across Tissues and Species.
Matrix Biol. Plus 2020,
6–7, 100023. doi:10.1016/j.mbplus.2020.100023.
[Google Scholar]
-
21.
Grässel S, Bauer RJ. Collagen XVI in Health and Disease.
Matrix Biol. 2013,
32, 64–73. doi:10.1016/j.matbio.2012.11.001.
[Google Scholar]
-
22.
Natsuga K, Watanabe M, Nishie W, Shimizu H. Life before and beyond Blistering: The Role of Collagen
XVII in Epidermal Physiology.
Exp. Dermatol. 2019,
28, 1135–1141. doi:10.1111/exd.13550.
[Google Scholar]
-
23.
Aikio M, Elamaa H, Vicente D, Izzi V, Kaur I, Seppinen L, et al. Specific Collagen XVIII Isoforms Promote Adipose Tissue Accrual via Mechanisms Determining Adipocyte Number and Affect Fat Deposition.
Proc. Natl. Acad. Sci. USA 2014,
111, E3043–E3052. doi:10.1073/pnas.1405879111.
[Google Scholar]
-
24.
Sadri G, Fischer AG, Brittian KR, Elliott E, Nystoriak MA, Uchida S, et al. Collagen Type XIX Regulates Cardiac Extracellular Matrix Structure and Ventricular Function.
Matrix Biol. 2022,
109, 49–69. doi:10.1016/j.matbio.2022.03.007.
[Google Scholar]
-
25.
Thorlacius-Ussing J, Jensen C, Madsen EA, Nissen NI, Manon-Jensen T, Chen IM, et al. Type XX Collagen Is Elevated in Circulation of Patients with Solid Tumors.
Int. J. Mol. Sci. 2022,
23, 4144. doi:10.3390/ijms23084144.
[Google Scholar]
-
26.
Chou M-Y, Li H-C. Genomic Organization and Characterization of the Human Type XXI Collagen (COL21A1) Gene.
Genomics 2002,
79, 395–401. doi:10.1006/geno.2002.6712.
[Google Scholar]
-
27.
Koch M, Schulze J, Hansen U, Ashwodt T, Keene DR, Brunken WJ, et al. A Novel Marker of Tissue Junctions, Collagen XXII.
J. Biol. Chem. 2004,
279, 22514–22521. doi:10.1074/jbc.M400536200.
[Google Scholar]
-
28.
Kehlet SN, Karsdal MA. Type XXIII Collagen. In Biochemistry of Collagens, Laminins and Elastin; Elsevier: Amsterdam, The Netherlands, 2016; pp 139–141. doi:10.1016/B978-0-12-809847-9.00023-4.
-
29.
Koch M, Laub F, Zhou P, Hahn RA, Tanaka S, Burgeson RE, et al. Collagen XXIV, a Vertebrate Fibrillar Collagen with Structural Features of Invertebrate Collagens.
J. Biol. Chem. 2003,
278, 43236–43244. doi:10.1074/jbc.M302112200.
[Google Scholar]
-
30.
Gonçalves TJM, Boutillon F, Lefebvre S, Goffin V, Iwatsubo T, Wakabayashi T, et al. Collagen XXV Promotes Myoblast Fusion during Myogenic Differentiation and Muscle Formation.
Sci. Rep. 2019,
9, 5878. doi:10.1038/s41598-019-42296-6.
[Google Scholar]
-
31.
Sato K, Yomogida K, Wada T, Yorihuzi T, Nishimune Y, Hosokawa N, et al. Type XXVI Collagen, a New Member of the Collagen Family, Is Specifically Expressed in the Testis and Ovary.
J. Biol. Chem. 2002,
277, 37678–37684. doi:10.1074/jbc.M205347200.
[Google Scholar]
-
32.
Hjorten R, Hansen U, Underwood RA, Telfer HE, Fernandes RJ, Krakow D, et al. Type XXVII Collagen at the Transition of Cartilage to Bone during Skeletogenesis.
Bone 2007,
41, 535–542. doi:10.1016/j.bone.2007.06.024.
[Google Scholar]
-
33.
Exposito J-Y, Valcourt U, Cluzel C, Lethias C. The Fibrillar Collagen Family.
Int. J. Mol. Sci. 2010,
11, 407–426. doi:10.3390/ijms11020407.
[Google Scholar]
-
34.
Grimal S, Puech S, Wagener R, Ventéo S, Carroll P, Fichard-Carroll A. Collagen XXVIII Is a Distinctive Component of the Peripheral Nervous System Nodes of Ranvier and Surrounds Nonmyelinating Glial Cells.
Glia 2010,
58, 1977–1987. doi:10.1002/glia.21066.
[Google Scholar]
-
35.
Bella J, Eaton M, Brodsky B, Berman HM. Crystal and Molecular Structure of a Collagen-Like Peptide at 1.9 Å Resolution.
Science 1994,
266, 75–81. doi:10.1126/science.7695699.
[Google Scholar]
-
36.
Berisio R, Vitagliano L, Mazzarella L, Zagari A. Crystal Structure of the Collagen Triple Helix Model [(Pro-Pro-Gly)
10]
3.
Protein Sci. 2002,
11, 262–270. doi:10.1110/ps.32602.
[Google Scholar]
-
37.
Fan D, Takawale A, Lee J, Kassiri Z. Cardiac Fibroblasts, Fibrosis and Extracellular Matrix Remodeling in Heart Disease.
Fibrogenesis Tissue Repair 2012,
5, 15. doi:10.1186/1755-1536-5-15.
[Google Scholar]
-
38.
Liu Y, Ramanath HS, Wang D-A. Tendon Tissue Engineering Using Scaffold Enhancing Strategies.
Trends Biotechnol. 2008,
26, 201–209. doi:10.1016/j.tibtech.2008.01.003.
[Google Scholar]
-
39.
Tuckwell DS, Ayad S, Grant ME, Takigawa M, Humphries MJ. Conformation Dependence of Integrin-Type II Collagen Binding: Inability of Collagen Peptides to Support Α2β1 Binding, and Mediation of Adhesion to Denatured Collagen by a Novel Α5β1-Fibronectin Bridge.
J. Cell Sci. 1994,
107, 993–1005. doi:10.1242/jcs.107.4.993.
[Google Scholar]
-
40.
Franzke C-W, Bruckner P, Bruckner-Tuderman L. Collagenous Transmembrane Proteins: Recent Insights into Biology and Pathology*.
J. Biol. Chem. 2005,
280, 4005–4008. doi:10.1074/jbc.R400034200.
[Google Scholar]
-
41.
Mouw JK, Ou G, Weaver VM. Extracellular Matrix Assembly: A Multiscale Deconstruction.
Nat. Rev. Mol. Cell Biol. 2014,
15, 771–785. doi:10.1038/nrm3902.
[Google Scholar]
-
42.
Myllyharju J. Intracellular Post-Translational Modifications of Collagens. In Collagen; Brinckmann J, Notbohm H, Müller PK, Eds.; Topics in Current Chemistry; Springer: Berlin/Heidelberg, Germany, 2005; Volume 247, pp. 115–147. doi:10.1007/b103821.
-
43.
Persikov AV, Ramshaw JAM, Kirkpatrick A, Brodsky B. Electrostatic Interactions Involving Lysine Make Major Contributions to Collagen Triple-Helix Stability.
Biochemistry 2005,
44, 1414–1422. doi:10.1021/bi048216r.
[Google Scholar]
-
44.
Brodsky B, Persikov AV. Molecular Structure of the Collagen Triple Helix. In Advances in Protein Chemistry; Elsevier: Amsterdam, The Netherlands, 2005; Volume 70, pp. 301–339. doi:10.1016/S0065-3233(05)70009-7.
-
45.
Boudko SP, Engel J, Bächinger HP. The Crucial Role of Trimerization Domains in Collagen Folding.
Int. J. Biochem. Cell Biol. 2012,
44, 21–32. doi:10.1016/j.biocel.2011.09.009.
[Google Scholar]
-
46.
McLaughlin SH, Bulleid NJ. Molecular recognition in procollagen chain assembly.
Matrix Biol. 1998,
16, 369–377. doi:10.1016/s0945-053x(98)90010-5.
[Google Scholar]
-
47.
Bourhis J-M, Mariano N, Zhao Y, Harlos K, Exposito J-Y, Jones EY, et al. S. Structural Basis of Fibrillar Collagen Trimerization and Related Genetic Disorders.
Nat. Struct. Mol. Biol. 2012,
19, 1031–1036. doi:10.1038/nsmb.2389.
[Google Scholar]
-
48.
Birk DE, Trelstad RL. Extracellular Compartments in Tendon Morphogenesis: Collagen Fibril, Bundle, and Macroaggregate Formation.
J. Cell Biol. 1986,
103, 231–240.
[Google Scholar]
-
49.
Canty EG, Lu Y, Meadows RS, Shaw MK, Holmes DF, Kadler KE. Coalignment of Plasma Membrane Channels and Protrusions (Fibripositors) Specifies the Parallelism of Tendon.
J. Cell Biol. 2004,
165, 553–563. doi:10.1083/jcb.200312071.
[Google Scholar]
-
50.
Bruckner P. Suprastructures of Extracellular Matrices: Paradigms of Functions Controlled by Aggregates Rather than Molecules.
Cell Tissue Res. 2010,
339, 7–18. doi:10.1007/s00441-009-0864-0.
[Google Scholar]
-
51.
Wenstrup RJ, Florer JB, Brunskill EW, Bell SM, Chervoneva I, Birk DE. Type V Collagen Controls the Initiation of Collagen Fibril Assembly.
J. Biol. Chem. 2004,
279, 53331–53337. doi:10.1074/jbc.M409622200.
[Google Scholar]
-
52.
Eyre DR, Weis MA, Wu J-J. Advances in Collagen Cross-Link Analysis.
Methods 2008,
45, 65–74. doi:10.1016/j.ymeth.2008.01.002.
[Google Scholar]
-
53.
Lucero HA, Kagan HM. Lysyl Oxidase: An Oxidative Enzyme and Effector of Cell Function.
Cell. Mol. Life Sci. 2006,
63, 2304–2316. doi:10.1007/s00018-006-6149-9.
[Google Scholar]
-
54.
Orgel JP, Wess TJ, Miller A. The
in Situ Conformation and Axial Location of the Intermolecular Cross-Linked Non-Helical Telopeptides of Type I Collagen.
Structure 2000,
8, 137–142. doi:10.1016/S0969-2126(00)00089-7.
[Google Scholar]
-
55.
O’Leary LER, Fallas JA, Bakota EL, Kang MK, Hartgerink JD. Multi-Hierarchical Self-Assembly of a Collagen Mimetic Peptide from Triple Helix to Nanofibre and Hydrogel.
Nat. Chem. 2011,
3, 821–828. doi:10.1038/nchem.1123.
[Google Scholar]
-
56.
Knupp C, Squire JM. Molecular Packing in Network-Forming Collagens.
Sci. World J. 2003,
3, 558–577. doi:10.1100/tsw.2003.40.
[Google Scholar]
-
57.
Fidler AL, Boudko SP, Rokas A, Hudson BG. The Triple Helix of Collagens—an Ancient Protein Structure That Enabled Animal Multicellularity and Tissue Evolution.
J. Cell Sci. 2018,
131,
jcs203950. doi:10.1242/jcs.203950.
[Google Scholar]
-
58.
Kadler KE, Hojima Y, Prockop DJ. Assembly of Collagen Fibrils de Novo by Cleavage of the Type I pC-Collagen with Procollagen C-Proteinase. Assay of Critical Concentration Demonstrates That Collagen Self-Assembly Is a Classical Example of an Entropy-Driven Process.
J. Biol. Chem. 1987,
262, 15696–15701. doi:10.1016/S0021-9258(18)47783-6.
[Google Scholar]
-
59.
Cummings CF, Pedchenko V, Brown KL, Colon S, Rafi M, Jones-Paris C, et al. Extracellular Chloride Signals Collagen IV Network Assembly during Basement Membrane Formation.
J. Cell Biol. 2016,
213, 479–494. doi:10.1083/jcb.201510065.
[Google Scholar]
-
60.
Bella J. A First Census of Collagen Interruptions: Collagen’s Own Stutters and Stammers.
J. Struct. Biol. 2014,
186, 438–450. doi:10.1016/j.jsb.2014.03.022.
[Google Scholar]
-
61.
Yurchenco PD, Ruben GC. Basement Membrane Structure in Situ: Evidence for Lateral Associations in the Type IV Collagen Network.
J. Cell Biol. 1987,
105, 2559–2568. doi:10.1083/jcb.105.6.2559.
[Google Scholar]
-
62.
Elango J, Hou C, Bao B, Wang S, Maté Sánchez De Val JE, Wenhui W. The Molecular Interaction of Collagen with Cell Receptors for Biological Function.
Polymers 2022,
14, 876. doi:10.3390/polym14050876.
[Google Scholar]
-
63.
Hu K, Hu M, Xiao Y, Cui Y, Yan J, Yang G, et al. Preparation Recombination Human-like Collagen/Fibroin Scaffold and Promoting the Cell Compatibility with Osteoblasts.
J. Biomed. Mater. Res. A 2021,
109, 346–353. doi:10.1002/jbm.a.37027.
[Google Scholar]
-
64.
Lowell CA, Mayadas TN. Overview: Studying Integrins In Vivo. In Integrin and Cell Adhesion Molecules; Shimaoka M, Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2011; Volume 757, pp. 369–397. doi:10.1007/978-1-61779-166-6_22.
-
65.
Velling T, Kusche-Gullberg M, Sejersen T, Gullberg D. cDNA Cloning and Chromosomal Localization of Human Α11 Integrin.
J. Biol. Chem. 1999,
274, 25735–25742. doi:10.1074/jbc.274.36.25735.
[Google Scholar]
-
66.
Briesewitz R, Epstein MR, Marcantonio EE. Expression of Native and Truncated Forms of the Human Integrin Alpha 1 Subunit.
J. Biol. Chem. 1993,
268, 2989–2996. doi:10.1016/S0021-9258(18)53871-0.
[Google Scholar]
-
67.
Iwamoto DV, Calderwood DA. Regulation of Integrin-Mediated Adhesions.
Curr. Opin. Cell Biol. 2015,
36, 41–47. doi:10.1016/j.ceb.2015.06.009.
[Google Scholar]
-
68.
Shrivastava A, Radziejewski C, Campbell E, Kovac L, McGlynn M, Ryan TE, et al. An Orphan Receptor Tyrosine Kinase Family Whose Members Serve as Nonintegrin Collagen Receptors.
Mol. Cell 1997,
1, 25–34. doi:10.1016/S1097-2765(00)80004-0.
[Google Scholar]
-
69.
Löffek S, Hurskainen T, Jackow J, Sigloch FC, Schilling O, Tasanen K, et al. Transmembrane Collagen XVII Modulates Integrin Dependent Keratinocyte Migration via PI3K/Rac1 Signaling.
PLoS ONE 2014,
9, e87263. doi:10.1371/journal.pone.0087263.
[Google Scholar]
-
70.
Yang S, Kim J, Ryu J-H, Oh H, Chun C-H, Kim BJ, et al. Hypoxia-Inducible Factor-2α Is a Catabolic Regulator of Osteoarthritic Cartilage Destruction.
Nat. Med. 2010,
16, 687–693. doi:10.1038/nm.2153.
[Google Scholar]
-
71.
Gonzalez DM, Medici D. Signaling Mechanisms of the Epithelial-Mesenchymal Transition.
Sci. Signal. 2014,
7,
re8. doi:10.1126/scisignal.2005189.
[Google Scholar]
-
72.
Ozaki I, Hamajima H, Matsuhashi S, Mizuta T. Regulation of TGF-?1-Induced Pro-Apoptotic Signaling by Growth Factor Receptors and Extracellular Matrix Receptor Integrins in the Liver.
Front. Physiol. 2011,
2, 78. doi:10.3389/fphys.2011.00078.
[Google Scholar]
-
73.
Madamanchi A, Santoro SA, Zutter MM. α2β1 Integrin.
Adv Exp Med Biol. 2014,
819, 41-60. doi:10.1007/978-94-017-9153-3_3.
[Google Scholar]
-
74.
Calderwood DA, Tuckwell DS, Eble J, Kühn K, Humphries MJ. The Integrin Α1 A-Domain Is a Ligand Binding Site for Collagens and Laminin.
J. Biol. Chem. 1997,
272, 12311–12317. doi:10.1074/jbc.272.19.12311.
[Google Scholar]
-
75.
Nykvist P, Tu H, Ivaska J, Käpylä J, Pihlajaniemi T, Heino J. Distinct Recognition of Collagen Subtypes by Α1β1 and α2β1Integrins.
J. Biol. Chem. 2000,
275, 8255–8261. doi:10.1074/jbc.275.11.8255.
[Google Scholar]
-
76.
Kern A, Eble J, Golbik R, Kühn K. Interaction of Type IV Collagen with the Isolated Integrins Α1β1 and Α2β1.
Eur. J. Biochem. 1993,
215, 151–159. doi:10.1111/j.1432-1033.1993.tb18017.x.
[Google Scholar]
-
77.
Jokinen J, Dadu E, Nykvist P, Käpylä J, White DJ, Ivaska J, et al. Integrin-Mediated Cell Adhesion to Type I Collagen Fibrils.
J. Biol. Chem. 2004,
279, 31956–31963. doi:10.1074/jbc.M401409200.
[Google Scholar]
-
78.
Käpylä J, Jäälinoja J, Tulla M, Ylöstalo J, Nissinen L, Viitasalo T, et al. The Fibril-Associated Collagen IX Provides a Novel Mechanism for Cell Adhesion to Cartilaginous Matrix.
J. Biol. Chem. 2004,
279, 51677–51687. doi:10.1074/jbc.M409412200.
[Google Scholar]
-
79.
Koivunen J, Tu H, Kemppainen A, Anbazhagan P, Finnilä MA, Saarakkala S, et al. Integrin Α11β1 Is a Receptor for Collagen XIII.
Cell Tissue Res. 2021,
383, 1135–1153. doi:10.1007/s00441-020-03300-y.
[Google Scholar]
-
80.
Zhang W-M, Käpylä J, Puranen JS, Knight CG, Tiger C-F, Pentikäinen OT, et al. Α11β1 Integrin Recognizes the GFOGER Sequence in Interstitial Collagens.
J. Biol. Chem. 2003,
278, 7270–7277. doi:10.1074/jbc.M210313200.
[Google Scholar]
-
81.
Orgel JPRO, Madhurapantula RS. A Structural Prospective for Collagen Receptors Such as DDR and Their Binding of the Collagen Fibril.
Biochim. Biophys. Acta BBA—Mol. Cell Res. 2019,
1866, 118478. doi:10.1016/j.bbamcr.2019.04.008.
[Google Scholar]
-
82.
Jung SM, Takemura Y, Imamura Y, Hayashi T, Adachi E, Moroi M. Collagen-Type Specificity of Glycoprotein VI as a Determinant of Platelet Adhesion.
Platelets 2008,
19, 32–42. doi:10.1080/09537100701609027.
[Google Scholar]
-
83.
Schultz HS, Guo L, Keller P, Fleetwood AJ, Sun M, Guo W, et al. OSCAR-collagen Signaling in Monocytes Plays a Proinflammatory Role and May Contribute to the Pathogenesis of Rheumatoid Arthritis.
Eur. J. Immunol. 2016,
46, 952–963. doi:10.1002/eji.201545986.
[Google Scholar]
-
84.
Son M. Understanding the Contextual Functions of C1q and LAIR-1 and Their Applications.
Exp. Mol. Med. 2022,
54, 567–572. doi:10.1038/s12276-022-00774-4.
[Google Scholar]
-
85.
Steffensen B, Xu X, Martin PA, Zardeneta G. Human Fibronectin and MMP-2 Collagen Binding Domains Compete for Collagen Binding Sites and Modify Cellular Activation of MMP-2.
Matrix Biol. 2002,
21, 399–414. doi:10.1016/S0945-053X(02)00032-X.
[Google Scholar]
-
86.
Sano K, Asanuma-Date K, Arisaka F, Hattori S, Ogawa H. Changes in Glycosylation of Vitronectin Modulate Multimerization and Collagen Binding during Liver Regeneration.
Glycobiology 2007,
17, 784–794. doi:10.1093/glycob/cwm031.
[Google Scholar]
-
87.
Melander MC, Jürgensen HJ, Madsen DH, Engelholm LH, Behrendt N. The Collagen Receptor uPARAP/Endo180 in Tissue Degradation and Cancer (Review).
Int. J. Oncol. 2015,
47, 1177–1188. doi:10.3892/ijo.2015.3120.
[Google Scholar]
-
88.
Chattopadhyay S, Raines RT. Collagen-based Biomaterials for Wound Healing.
Biopolymers 2014,
101, 821–833. doi:10.1002/bip.22486.
[Google Scholar]
-
89.
Huang X, Lv J, Li Y, Mao S, Li Z, Jing Z, et al. Programmable C-to-U
RNA Editing Using the Human
APOBEC 3A Deaminase.
EMBO J. 2020,
39, e104741. doi:10.15252/embj.2020104741.
[Google Scholar]
-
90.
Sorushanova A, Delgado LM, Wu Z, Shologu N, Kshirsagar A, Raghunath R, et al. The Collagen Suprafamily: From Biosynthesis to Advanced Biomaterial Development.
Adv. Mater. 2019,
31, 1801651. doi:10.1002/adma.201801651.
[Google Scholar]
-
91.
Parenteau-Bareil R, Gauvin R, Berthod F. Collagen-Based Biomaterials for Tissue Engineering Applications.
Materials 2010,
3, 1863–1887. doi:10.3390/ma3031863.
[Google Scholar]
-
92.
Silvipriya K, Kumar K, Bhat A, Kumar B, John A, Lakshmanan P. Collagen: Animal Sources and Biomedical Application.
J. Appl. Pharm. Sci. 2015, 123–127. doi:10.7324/JAPS.2015.50322.
[Google Scholar]
-
93.
Fan J, Zhuang Y, Li B. Effects of Collagen and Collagen Hydrolysate from Jellyfish Umbrella on Histological and Immunity Changes of Mice Photoaging.
Nutrients 2013,
5, 223–233. doi:10.3390/nu5010223.
[Google Scholar]
-
94.
Gelse K. Collagens—Structure, Function, and Biosynthesis.
Adv. Drug Deliv. Rev. 2003,
55, 1531–1546. doi:10.1016/j.addr.2003.08.002.
[Google Scholar]
-
95.
Lima CA, Campos JF, Filho JLL, Converti A, Da Cunha MGC, Porto ALF. Antimicrobial and Radical Scavenging Properties of Bovine Collagen Hydrolysates Produced by Penicillium Aurantiogriseum URM 4622 Collagenase.
J. Food Sci. Technol. 2015,
52, 4459–4466. doi:10.1007/s13197-014-1463-y.
[Google Scholar]
-
96.
Zhang Y, Olsen K, Grossi A, Otte J. Effect of Pretreatment on Enzymatic Hydrolysis of Bovine Collagen and Formation of ACE-Inhibitory Peptides.
Food Chem. 2013,
141, 2343–2354. doi:10.1016/j.foodchem.2013.05.058.
[Google Scholar]
-
97.
Choi D, Min S-G, Jo Y-J. Functionality of Porcine Skin Hydrolysates Produced by Hydrothermal Processing for Liposomal Delivery System.
J. Food Biochem. 2018,
42, e12464. doi:10.1111/jfbc.12464.
[Google Scholar]
-
98.
Dandagi GL, Byahatti SM. An Insight into the Swine-Influenza A (H1N1) Virus Infection.
Lung India 2011,
28, 34–38.
[Google Scholar]
-
99.
León-López A, Morales-Peñaloza A, Martínez-Juárez VM, Vargas-Torres A, Zeugolis DI, Aguirre-Álvarez G. Hydrolyzed Collagen—Sources and Applications.
Molecules 2019,
24, 4031. doi:10.3390/molecules24224031.
[Google Scholar]
-
100.
Gómez-Guillén MC, Giménez B, López-Caballero ME, Montero MP. Functional and Bioactive Properties of Collagen and Gelatin from Alternative Sources: A Review.
Food Hydrocoll. 2011,
25, 1813–1827. doi:10.1016/j.foodhyd.2011.02.007.
[Google Scholar]
-
101.
Skierka E, Sadowska M. The Influence of Different Acids and Pepsin on the Extractability of Collagen from the Skin of Baltic Cod (Gadus Morhua).
Food Chem. 2007,
105, 1302–1306. doi:10.1016/j.foodchem.2007.04.030.
[Google Scholar]
-
102.
Ketnawa S, Benjakul S, Martínez-Alvarez O, Rawdkuen S. Fish Skin Gelatin Hydrolysates Produced by Visceral Peptidase and Bovine Trypsin: Bioactivity and Stability.
Food Chem. 2017,
215, 383–390. doi:10.1016/j.foodchem.2016.07.145.
[Google Scholar]
-
103.
Hong H, Chaplot S, Chalamaiah M, Roy BC, Bruce HL, Wu J. Removing Cross-Linked Telopeptides Enhances the Production of Low-Molecular-Weight Collagen Peptides from Spent Hens.
J. Agric. Food Chem. 2017,
65, 7491–7499. doi:10.1021/acs.jafc.7b02319.
[Google Scholar]
-
104.
Hulgan SAH, Hartgerink JD. Recent Advances in Collagen Mimetic Peptide Structure and Design.
Biomacromolecules 2022,
23, 1475–1489. doi:10.1021/acs.biomac.2c00028.
[Google Scholar]
-
105.
Mukherjee S, Varshashankari V, Feba A, Ayyadurai N, Balamurugan K, Shanmugam G. Hyperstable and Fibril-Forming Collagen-Mimetic Peptides in Shortest Triple Helices: Empowering the Capping by π-Systems.
Biomacromolecules 2025,
26, 2171–2185. doi:10.1021/acs.biomac.4c01455.
[Google Scholar]
-
106.
Bourgot I, Primac I, Louis T, Noël A, Maquoi E. Reciprocal Interplay Between Fibrillar Collagens and Collagen-Binding Integrins: Implications in Cancer Progression and Metastasis.
Front. Oncol. 2020,
10, 1488. doi:10.3389/fonc.2020.01488.
[Google Scholar]
-
107.
Mhanna R, Öztürk E, Vallmajo-Martin Q, Millan C, Müller M, Zenobi-Wong M. GFOGER-Modified MMP-Sensitive Polyethylene Glycol Hydrogels Induce Chondrogenic Differentiation of Human Mesenchymal Stem Cells.
Tissue Eng. Part A 2014,
20, 1165–1174. doi:10.1089/ten.tea.2013.0519.
[Google Scholar]
-
108.
Zhao C, Xiao Y, Ling S, Pei Y, Ren J. Structure of Collagen.
Methods Mol. Biol. 2021,
2347, 17–25. doi:10.1007/978-1-0716-1574-4_2.
[Google Scholar]
-
109.
Zhang C, Fan D, Shang L, Ma X, Luo Y, Xue W, et al. Optimization of Fermentation Process for Human-like Collagen Production of Recombinant Escherichia Coli Using Response Surface Methodology.
Chin. J. Chem. Eng. 2010,
18, 137–142. doi:10.1016/s1004-9541(08)60334-1.
[Google Scholar]
-
110.
Tang Y, Yang X, Hang B, Li J, Huang L, Huang F, et al. Efficient Production of Hydroxylated Human-Like Collagen Via the Coexpression of Three Key Genes in Escherichia Coli Origami (DE3).
Appl. Biochem. Biotechnol. 2016,
178, 1458–1470. doi:10.1007/s12010-015-1959-6.
[Google Scholar]
-
111.
Nokelainen M, Tu H, Vuorela A, Notbohm H, Kivirikko KI, Myllyharju J. High-level Production of Human Type I Collagen in the Yeast
Pichia Pastoris.
Yeast 2001,
18, 797–806. doi:10.1002/yea.730.
[Google Scholar]
-
112.
Werkmeister JA, Ramshaw JAM. Recombinant Protein Scaffolds for Tissue Engineering.
Biomed. Mater. 2012,
7, 012002. doi:10.1088/1748-6041/7/1/012002.
[Google Scholar]
-
113.
Rutschmann C, Baumann S, Cabalzar J, Luther KB, Hennet T. Recombinant Expression of Hydroxylated Human Collagen in
Escherichia coli.
Appl. Microbiol. Biotechnol. 2014,
98, 4445–4455. doi:10.1007/s00253-013-5447-z.
[Google Scholar]
-
114.
Ruggiero F, Exposito J-Y, Bournat P, Gruber V, Perret S, Comte J, et al. Triple Helix Assembly and Processing of Human Collagen Produced in Transgenic Tobacco Plants.
FEBS Lett. 2000,
469, 132–136. doi:10.1016/S0014-5793(00)01259-X.
[Google Scholar]
-
115.
Xu X, Gan Q, Clough RC, Pappu KM, Howard JA, Baez JA, et al. Hydroxylation of Recombinant Human Collagen Type I Alpha 1 in Transgenic Maize Co-Expressed with a Recombinant Human Prolyl 4-Hydroxylase.
BMC Biotechnol. 2011,
11, 69. doi:10.1186/1472-6750-11-69.
[Google Scholar]
-
116.
Vuorela A. Assembly of Human Prolyl 4-Hydroxylase and Type III Collagen in the Yeast Pichia Pastoris: Formation of a Stable Enzyme Tetramer Requires Coexpression with Collagen and Assembly of a Stable Collagen Requires Coexpression with Prolyl 4-Hydroxylase.
EMBO J. 1997,
16, 6702–6712. doi:10.1093/emboj/16.22.6702.
[Google Scholar]
-
117.
Myllyharju’ J, Nokelainen M, Vuorela A. Expression of Recombinant Human Type 1-111 Collagens in the Yeast Pichia Pastoris.
Biochem. Soc. Trans. 2000,
28.
[Google Scholar]
-
118.
Toman PD, Chisholm G, McMullin H, Giere LM, Olsen DR, Kovach RJ, et al. Production of Recombinant Human Type I Procollagen Trimers Using a Four-Gene Expression System in the Yeast Saccharomyces Cerevisiae.
J. Biol. Chem. 2000,
275, 23303–23309. doi:10.1074/jbc.M002284200.
[Google Scholar]
-
119.
Liu X, Yang Y, Zhang W, Sun Y, Peng F, Jeffrey L, et al. Expression of Recombinant Protein Using
Corynebacterium Glutamicum: Progress, Challenges and Applications.
Crit. Rev. Biotechnol. 2016,
36, 652–664. doi:10.3109/07388551.2015.1004519.
[Google Scholar]
-
120.
Li X, Xue W, Zhu C, Fan D, Liu Y, Ma X. Novel Hydrogels Based on Carboxyl Pullulan and Collagen Crosslinking with 1, 4-Butanediol Diglycidylether for Use as a Dermal Filler: Initial in Vitro and in Vivo Investigations.
Mater. Sci. Eng. C 2015,
57, 189–196. doi:10.1016/j.msec.2015.07.059.
[Google Scholar]
-
121.
Kumar V, Kumar N, Gangwar AK, Singh H, Singh R. Comparative Histologic and Immunologic Evaluation of 1,4-Butanediol Diglycidyl Ether Crosslinked versus Noncrosslinked Acellular Swim Bladder Matrix for Healing of Full-Thickness Skin Wounds in Rabbits.
J. Surg. Res. 2015,
197, 436–446. doi:10.1016/j.jss.2015.04.080.
[Google Scholar]
-
122.
Pustlauk W, Paul B, Gelinsky M, Bernhardt A. Jellyfish Collagen and Alginate: Combined Marine Materials for Superior Chondrogenesis of hMSC.
Mater. Sci. Eng. C 2016,
64, 190–198. doi:10.1016/j.msec.2016.03.081.
[Google Scholar]
-
123.
Unsöld C, Pappano WN, Imamura Y, Steiglitz BM, Greenspan DS. Biosynthetic Processing of the Pro-Α1(V)2Pro-Α2(V) Collagen Heterotrimer by Bone Morphogenetic Protein-1 and Furin-like Proprotein Convertases.
J. Biol. Chem. 2002,
277, 5596–5602. doi:10.1074/jbc.M110003200.
[Google Scholar]
-
124.
Zhou H, Li W, Pan L, Zhu T, Zhou T, Xiao E, et al. Human Extracellular Matrix (ECM)-like Collagen and Its Bioactivity.
Regen. Biomater. 2024,
11,
rbae008. doi:10.1093/rb/rbae008.
[Google Scholar]
-
125.
Jia Y, Han Y, Zhang Y, Li L, Zhang B, Yan X. Multifunctional Type Lll Recombinant Human Collagen Incorporated Sodium Alginate Hydrogel with Sustained Release of Extra Cellular Vehicles for Wound Healing Multimodal Therapy in Diabetic Mice.
Regen. Ther. 2024,
27, 329–341. doi:10.1016/j.reth.2024.03.010.
[Google Scholar]
-
126.
Zhuang Z-M, Wang Y, Chen L, Wu Z-R, Zhang T, Bei H-P, et al. Triple-Molded, Reinforced Arrowhead Microneedle Patch of Dual Human-Derived Matrix for Integrated Management of Diabetic Wounds.
Biomaterials 2026,
324, 123520. doi:10.1016/j.biomaterials.2025.123520.
[Google Scholar]
-
127.
Wang Q, An Q, Wang Y, Yang J, Zhang X, Jiang S, et al. Development and Mechanistic Investigation of Recombinant Type III Humanized Collagen Gel for Mid-Facial Soft Tissue Repair.
Collagen Leather 2025,
7, 14. doi:10.1186/s42825-025-00196-8.
[Google Scholar]
-
128.
Munyemana JC, He H, Fu C, Fan Y, Sun X, Xiao J. Recombinant Collagen-Templated Biomineralized Synthesis of Biocompatible pH-Responsive Porous Calcium Carbonate Nanospheres.
ACS Omega 2023,
8, 30879–30887. doi:10.1021/acsomega.3c01467.
[Google Scholar]
-
129.
Zhang H, Wang J, Wei J, Fu X, Ma J, Chen J. Molecular Engineering of Recombinant Protein Hydrogels: Programmable Design and Biomedical Applications.
Gels 2025,
11, 579. doi:10.3390/gels11080579.
[Google Scholar]
-
130.
Vitale M, Ligorio C, Smith IP, Richardson SM, Hoyland JA, Bella J. Incorporation of Natural and Recombinant Collagen Proteins within Fmoc-Based Self-Assembling Peptide Hydrogels.
Gels 2022,
8, 254. doi:10.3390/gels8050254.
[Google Scholar]
-
131.
Gu J, He Y, He C, Zhang Q, Huang Q, Bai S, et al. Advances in the Structures, Mechanisms and Targeting of Molecular Chaperones.
Signal Transduct. Target. Ther. 2025,
10, 84. doi:10.1038/s41392-025-02166-2.
[Google Scholar]
-
132.
Masuda R, Phyu Thant KP, Kawahara K, Oki H, Kadonosono T, Kobayashi Y, et al. A Yeast Two-Hybrid System to Obtain Triple-Helical Ligands from Combinatorial Random Peptide Libraries.
J. Biol. Chem. 2024,
300, 107794. doi:10.1016/j.jbc.2024.107794.
[Google Scholar]
-
133.
Ruggiero F, Koch M. Making Recombinant Extracellular Matrix Proteins.
Methods 2008,
45, 75–85. doi:10.1016/j.ymeth.2008.01.003.
[Google Scholar]
-
134.
Li H-C, Huang C-C, Chen S-F, Chou M-Y. Assembly of Homotrimeric Type XXI Minicollagen by Coexpression of Prolyl 4-Hydroxylase in Stably Transfected Drosophila Melanogaster S2 Cells.
Biochem. Biophys. Res. Commun. 2005,
336, 375–385. doi:10.1016/j.bbrc.2005.08.018.
[Google Scholar]
-
135.
Shi C, Wang C, Lu J, Zhong B, Tang J. Protein Sequence and Structure Co-Design with Equivariant Translation.
arXiv 2023, arXiv:2210.08761.
[Google Scholar]
-
136.
Evans R, O’Neill M, Pritzel A, Antropova N, Senior A, Green T, et al. Protein Complex Prediction with AlphaFold-Multimer.
BioRxiv 2021. doi:10.1101/2021.10.04.463034.
[Google Scholar]
-
137.
Elder S, Clune J, Walker J, Gloth P. Suitability of EGCG as a Means of Stabilizing a Porcine Osteochondral Xenograft.
J. Funct. Biomater. 2017,
8, 43. doi:10.3390/jfb8040043.
[Google Scholar]
-
138.
Pawelec KM, Confalonieri D, Ehlicke F, Van Boxtel HA, Walles H, Kluijtmans SGJM. Osteogenesis and Mineralization of Mesenchymal Stem Cells in Collagen Type I-based Recombinant Peptide Scaffolds.
J. Biomed. Mater. Res. A 2017,
105, 1856–1866. doi:10.1002/jbm.a.36049.
[Google Scholar]
-
139.
Lee JC, Pereira CT, Ren X, Huang W, Bischoff D, Weisgerber DW, et al. Optimizing Collagen Scaffolds for Bone Engineering: Effects of Cross-Linking and Mineral Content on Structural Contraction and Osteogenesis.
J. Craniofac. Surg. 2015,
26, 1992–1996. doi:10.1097/SCS.0000000000001918.
[Google Scholar]
-
140.
Berndt M, Li Y, Seyedhassantehrani N, Yao L. Fabrication and Characterization of Microspheres Encapsulating Astrocytes for Neural Regeneration.
ACS Biomater. Sci. Eng. 2017,
3, 1313–1321. doi:10.1021/acsbiomaterials.6b00229.
[Google Scholar]
-
141.
Han L, Zhang Z, Wang B, Wen Z. Construction and Biocompatibility of a Thin Type I/II Collagen Composite Scaffold.
Cell Tissue Bank. 2018,
19, 47–59. doi:10.1007/s10561-017-9653-2.
[Google Scholar]
-
142.
Wang X, Ma B, Chang J. Preparation of Decellularized Vascular Matrix by Co-Crosslinking of Procyanidins and Glutaraldehyde.
Biomed. Mater. Eng. 2015,
26, 19–30. doi:10.3233/BME-151548.
[Google Scholar]
-
143.
Wang X, Xu K, Ma L, Sun R, Wang K, Wang R, et al. Diffusion Model Assisted Designing Self-Assembling Collagen Mimetic Peptides as Biocompatible Materials.
Brief. Bioinform. 2024,
26,
bbae622. doi:10.1093/bib/bbae622.
[Google Scholar]
-
144.
Khare E, Yu C-H, Gonzalez Obeso C, Milazzo M, Kaplan DL, Buehler MJ. Discovering Design Principles of Collagen Molecular Stability Using a Genetic Algorithm, Deep Learning, and Experimental Validation.
Proc. Natl. Acad. Sci. USA 2022,
119, e2209524119. doi:10.1073/pnas.2209524119.
[Google Scholar]
-
145.
Hu J, Li J, Jiang J, Wang L, Roth J, McGuinness KN, et al. Design of Synthetic Collagens That Assemble into Supramolecular Banded Fibers as a Functional Biomaterial Testbed.
Nat. Commun. 2022,
13, 6761. doi:10.1038/s41467-022-34127-6.
[Google Scholar]
-
146.
Köster S, Evans HM, Wong JY, Pfohl T. An In Situ Study of Collagen Self-Assembly Processes.
Biomacromolecules 2008,
9, 199–207. doi:10.1021/bm700973t.
[Google Scholar]
-
147.
Hafner AE, Gyori NG, Bench CA, Davis LK, Šarić A. Modeling Fibrillogenesis of Collagen-Mimetic Molecules.
Biophys. J. 2020,
119, 1791–1799. doi:10.1016/j.bpj.2020.09.013.
[Google Scholar]
-
148.
Li Z, Du T, Ruan C, Niu X. Bioinspired Mineralized Collagen Scaffolds for Bone Tissue Engineering.
Bioact. Mater. 2021,
6, 1491–1511. doi:10.1016/j.bioactmat.2020.11.004.
[Google Scholar]
-
149.
Shen H, Fallas JA, Lynch E, Sheffler W, Parry B, Jannetty N, et al. De Novo Design of Self-Assembling Helical Protein Filaments.
Science 2018,
362, 705–709. doi:10.1126/science.aau3775.
[Google Scholar]
-
150.
LeCun Y, Bengio Y, Hinton G. Deep Learning.
Nature 2015,
521, 436–444. doi:10.1038/nature14539.
[Google Scholar]
-
151.
Srinivasan A, Gupta A, Narayanamurthy V. A Comprehensive Review of AI-Based Collagen Valorization: Recent Trends, Innovations in Extraction, and Applications.
Green Anal. Chem. 2025,
12, 100212. doi:10.1016/j.greeac.2025.100212.
[Google Scholar]
-
152.
Li W, Peng J, Yao D, Rao B, Xia Y, Wang Q, et al. The Structural Basis for the Collagen Processing by Human P3H1/CRTAP/PPIB Ternary Complex.
Nat. Commun. 2024,
15, 7844. doi:10.1038/s41467-024-52321-6.
[Google Scholar]
-
153.
Qian Y, Xu C, Xiong W, Jiang N, Zheng Y, He X, et al. Dual Cross-Linked Organic-Inorganic Hybrid Hydrogels Accelerate Diabetic Skin Wound Healing.
Chem. Eng. J. 2021,
417, 129335. doi:10.1016/j.cej.2021.129335.
[Google Scholar]
-
154.
Yao T, Chen H, Wang R, Rivero R, Wang F, Kessels L, et al. Thiol-Ene Conjugation of a VEGF Peptide to Electrospun Scaffolds for Potential Applications in Angiogenesis.
Bioact. Mater. 2023,
20, 306–317. doi:10.1016/j.bioactmat.2022.05.029.
[Google Scholar]
-
155.
Conde J, Oliva N, Atilano M, Song HS, Artzi N. Self-Assembled RNA-Triple-Helix Hydrogel Scaffold for microRNA Modulation in the Tumour Microenvironment.
Nat. Mater. 2016,
15, 353–363. doi:10.1038/nmat4497.
[Google Scholar]
-
156.
Kubyshkin V. Stabilization of the Triple Helix in Collagen Mimicking Peptides.
Org. Biomol. Chem. 2019,
17, 8031–8047. doi:10.1039/C9OB01646E.
[Google Scholar]
-
157.
Lyu C, Kong W, Liu Z, Wang S, Zhao P, Liang K, et al. Advanced Glycation End-Products as Mediators of the Aberrant Crosslinking of Extracellular Matrix in Scarred Liver Tissue.
Nat. Biomed. Eng. 2023,
7, 1437–1454. doi:10.1038/s41551-023-01019-z.
[Google Scholar]
-
158.
Wang J, Wu Q, Wang Y, Xiang L, Feng J, Zhou Z, et al. Collagenase-Loaded pH-Sensitive Nanocarriers Efficiently Remodeled Tumor Stroma Matrixes and Improved the Enrichment of Nanomedicines.
Nanoscale 2021,
13, 9402–9414. doi:10.1039/D1NR00950H.
[Google Scholar]
-
159.
Le DHT, Ibrahimova V, Van Den Wildenberg SAH, Wu H, Fonseca A, Torres T, et al. Light-Responsive Elastin-Like Peptide-Based Targeted Nanoparticles for Enhanced Spheroid Penetration.
Angew. Chem. Int. Ed. 2023,
62, e202300511. doi:10.1002/anie.202300511.
[Google Scholar]
-
160.
Li Y, Xia Y, Liu X, Wang J, Sun Y, Huang J, et al. Rational Design of Bioengineered Recombinant Collagen-like Protein Enhances GelMA Hydrogel for Diabetic Wound Healing.
Int. J. Biol. Macromol. 2024,
280, 136012. doi:10.1016/j.ijbiomac.2024.136012.
[Google Scholar]
-
161.
Sun TL, Luo F, Kurokawa T, Karobi SN, Nakajima T, Gong JP. Molecular Structure of Self-Healing Polyampholyte Hydrogels Analyzed from Tensile Behaviors.
Soft Matter 2015,
11, 9355–9366. doi:10.1039/C5SM01423A.
[Google Scholar]
-
162.
Yu HC, Zheng SY, Fang L, Ying Z, Du M, Wang J, et al. Reversibly Transforming a Highly Swollen Polyelectrolyte Hydrogel to an Extremely Tough One and Its Application as a Tubular Grasper.
Adv. Mater. 2020,
32, 2005171. doi:10.1002/adma.202005171.
[Google Scholar]
-
163.
Meyers MA, McKittrick J, Chen P-Y. Structural Biological Materials: Critical Mechanics-Materials Connections.
Science 2013,
339, 773–779. doi:10.1126/science.1220854.
[Google Scholar]
-
164.
Su R, Ma C, Han B, Zhang H, Liu K. Proteins for Hyperelastic Materials.
Small 2025,
21, 2406388. doi:10.1002/smll.202406388.
[Google Scholar]
-
165.
Mozdzen LC, Rodgers R, Banks JM, Bailey RC, Harley BAC. Increasing the Strength and Bioactivity of Collagen Scaffolds Using Customizable Arrays of 3D-Printed Polymer Fibers.
Acta Biomater. 2016,
33, 25–33. doi:10.1016/j.actbio.2016.02.004.
[Google Scholar]
-
166.
Hadidi P, Cissell DD, Hu JC, Athanasiou KA. Temporal Development of Near-Native Functional Properties and Correlations with qMRI in Self-Assembling Fibrocartilage Treated with Exogenous Lysyl Oxidase Homolog 2.
Acta Biomater. 2017,
64, 29–40. doi:10.1016/j.actbio.2017.09.035.
[Google Scholar]
-
167.
Priyadarshani P, Li Y, Yang S, Yao L. Injectable Hydrogel Provides Growth-permissive Environment for Human Nucleus Pulposus Cells.
J. Biomed. Mater. Res. A 2016,
104, 419–426. doi:10.1002/jbm.a.35580.
[Google Scholar]
-
168.
Zhou X, Wang J, Fang W, Tao Y, Zhao T, Xia K, et al. Genipin Cross-Linked Type II Collagen/Chondroitin Sulfate Composite Hydrogel-like Cell Delivery System Induces Differentiation of Adipose-Derived Stem Cells and Regenerates Degenerated Nucleus Pulposus.
Acta Biomater. 2018,
71, 496–509. doi:10.1016/j.actbio.2018.03.019.
[Google Scholar]
-
169.
Bryś M, Urbańska K, Olas B. Novel Findings Regarding the Bioactivity of the Natural Blue Pigment Genipin in Human Diseases.
Int. J. Mol. Sci. 2022,
23, 902.
[Google Scholar]
-
170.
Vardar E, Larsson HM, Allazetta S, Engelhardt EM, Pinnagoda K, Vythilingam G, et al. Microfluidic Production of Bioactive Fibrin Micro-Beads Embedded in Crosslinked Collagen Used as an Injectable Bulking Agent for Urinary Incontinence Treatment.
Acta Biomater. 2018,
67, 156–166. doi:10.1016/j.actbio.2017.11.034.
[Google Scholar]
-
171.
Quinlan E, Thompson EM, Matsiko A, O’Brien FJ, López-Noriega A. Functionalization of a Collagen–Hydroxyapatite Scaffold with Osteostatin to Facilitate Enhanced Bone Regeneration.
Adv. Healthc. Mater. 2015,
4, 2649–2656. doi:10.1002/adhm.201500439.
[Google Scholar]
-
172.
Liu T, Dan W, Dan N, Liu X, Liu X, Peng X. A Novel Grapheme Oxide-Modified Collagen-Chitosan Bio-Film for Controlled Growth Factor Release in Wound Healing Applications.
Mater. Sci. Eng. C 2017,
77, 202–211. doi:10.1016/j.msec.2017.03.256.
[Google Scholar]
-
173.
Baird CW, Myers PO, Piekarski B, Borisuk M, Majeed A, Emani SM, et al. Photo-Oxidized Bovine Pericardium in Congenital Cardiac Surgery: Single-Centre Experience.
Interact. Cardiovasc. Thorac. Surg. 2016,
24, 240–244. doi:10.1093/icvts/ivw315.
[Google Scholar]
-
174.
Rosellini E, Zhang YS, Migliori B, Barbani N, Lazzeri L, Shin SR, et al. Protein/Polysaccharide-based Scaffolds Mimicking Native Extracellular Matrix for Cardiac Tissue Engineering Applications.
J. Biomed. Mater. Res. A 2018,
106, 769–781. doi:10.1002/jbm.a.36272.
[Google Scholar]
-
175.
Chen C, Zhao M, Zhang R, Lu G, Zhao C, Fu F, et al. Collagen/Heparin Sulfate Scaffolds Fabricated by a 3D Bioprinter Improved Mechanical Properties and Neurological Function after Spinal Cord Injury in Rats.
J. Biomed. Mater. Res. A 2017,
105, 1324–1332. doi:10.1002/jbm.a.36011.
[Google Scholar]
-
176.
Siriwardane ML, DeRosa K, Collins G, Pfister BJ. Controlled Formation of Cross-Linked Collagen Fibers for Neural Tissue Engineering Applications.
Biofabrication 2014,
6, 015012. doi:10.1088/1758-5082/6/1/015012.
[Google Scholar]
-
177.
Li Y, He Q, Hu X, Liu Y, Cheng X, Li X, et al. Improved Performance of Collagen Scaffolds Crosslinked by Traut’s Reagent and Sulfo-SMCC.
J. Biomater. Sci. Polym. Ed. 2017,
28, 629–647. doi:10.1080/09205063.2017.1291296.
[Google Scholar]
-
178.
Bell A, Kofron M, Nistor V. Multiphoton Crosslinking for Biocompatible 3D Printing of Type I Collagen.
Biofabrication 2015,
7, 035007. doi:10.1088/1758-5090/7/3/035007.
[Google Scholar]
-
179.
Fadlallah A, Zhu H, Arafat S, Kochevar I, Melki S, Ciolino JB. Corneal Resistance to Keratolysis After Collagen Crosslinking with Rose Bengal and Green Light.
Investig. Opthalmology Vis. Sci. 2016,
57, 6610. doi:10.1167/iovs.15-18764.
[Google Scholar]
-
180.
Moreira MA, Souza NO, Sousa RS, Freitas DQ, Lemos MV, De Paula DM, et al. Efficacy of New Natural Biomodification Agents from Anacardiaceae Extracts on Dentin Collagen Cross-Linking.
Dent. Mater. 2017,
33, 1103–1109. doi:10.1016/j.dental.2017.07.003.
[Google Scholar]
-
181.
Lim E, Lim M-J, Min K-S, Kwon Y-S, Hwang Y-C, Yu M-K, et al. Effects of Epicatechin, a Crosslinking Agent, on Human Dental Pulp Cells Cultured in Collagen Scaffolds.
J. Appl. Oral Sci. 2016,
24, 76–84. doi:10.1590/1678-775720150383.
[Google Scholar]