1. Introduction
The Th17/IL-17 signaling pathway is a crucial regulator of immune and inflammatory function. Th17 cells play a key role in host immunity, particularly in combating infection and maintaining mucosal tissue barrier functions [
1,
2,
3,
4,
5]. During activation of the Th17/IL-17 pathway, Th17 cells produce IL-17 family cytokines that bind specifically to downstream IL-17 receptors, such as IL-17RA and IL-17RC, leading to the release of other inflammatory cell chemokines and stimulating further inflammatory cascades. Thus, the Th17/IL-17 pathway plays an important role in immune activation; however, pathologically excessive activation of the Th17/IL-17 response has been implicated in autoimmune and inflammatory disorders such as psoriasis, ankylosing spondylitis, multiple sclerosis, and systemic inflammatory response syndrome (SIRS) [
1].
Although the Th17/IL-17 signaling cascade has been extensively implicated in various physiological and pathological contexts, the regulatory mechanisms of the Th17/IL-17 pathway remain unclear. Further delineation of the regulatory mechanism of the Th17/IL-17 signaling pathway is necessary to guide the clinical management of IL-17 mediated inflammation and autoimmunity.
In this study, we unexpectedly found that spermidine synthase (SRM) and spermine oxidase (SMOX) are highly expressed in
Il17a+ cells. SRM and SMOX are the most important catalytic enzymes for the synthesis of spermidine from putrescine and spermine, respectively [
6,
7]. Spermidine is a polyaliphatic amine with anti-inflammatory properties [
8,
9,
10,
11,
12]. Recent reports highlighted that spermidine plays important roles in regulating T-cell lineage differentiation [
13,
14,
15]. However, the direct impact of spermidine on Th17 cytokine production has not been carefully examined. In addition, spermidine has been linked to various anti-inflammatory mechanisms but has not been directly linked to the regulatory pathway for Th17/IL-17 cytokine secretion prior to this study.
Here we establish that spermidine directly regulates the Th17/IL-17 signaling pathway in already differentiated T cells, including memory T cells and gamma-delta T cells. The regulatory roles of spermidine and its inhibitors in the Th17/IL-17 pathway were characterized
in vitro and
in vivo. We found that spermidine can directly target T cells and downregulate the expression of Th17/IL-17 signaling pathway products, thereby inhibiting IL-17 cytokine-induced inflammation. Conversely, inhibition of spermidine synthesis can reverse its suppressive effects on inflammatory cytokine release in the Th17/IL-17 pathway. Furthermore, single-cell RNA-seq (scRNA-Seq) results revealed that spermidine’s regulation of cytokine production depends on the antioxidant activity of peroxiredoxin 1 (PRDX1) in Th17 cells. The results of this study establish spermidine’s regulatory role in Th17/IL-17 signaling, providing further guidance for the potential pharmacologicl management of inflammation and hyper-immunity.
2. Materials and Methods
2.1. Mouse Models
All the mice used in this project were of wildtype strains and purchased from Jackson Lab (Cat# 000664; Bar Harbor, ME, USA) [
16]. Animals were maintained in pathogen-free conditions at the core animal facility at the University of Pittsburgh Medical Center. Mice aged 6–8 weeks were used to set up pre-designed models, with approval from the University of Pittsburgh Institutional Animal Care and Use Committee, Protocol #: 21028745, 2/2/2021.
2.2. Mouse Peritoneal Neutrophil Induction and Treatment
4% sterile Thioglycollate (Sigma, Cat# 70157; St. Louis, MO, USA) in ddH
2O was applied to induce mice peritonitis, 100 mL per mouse with intraperitoneal [
17] injection. 5 h later, mice were sacrificed by CO
2 euthanasia. Peritoneal lavage samples were collected by injecting sterile PBS into the peritoneal cavity with a 10 mL syringe. Peritoneal lavage cells were isolated and stimulated overnight at 37 °C with 50 ng/ mL m-IL-1β (Biolegend, Cat# 575002; San Diego, CA, USA) plus m-IL-23 (Biolegend, Cat# 589002) in 10% FBS-IMDM media. Cells were collected for scRNA-seq analysis.
2.3. Mouse Lung Inflammation Induction and Treatment
Either LPS (InvivoGen, #vac-3pelps; San Diego, CA, USA) or P.A. (ATCC
® 10145GFP™) were administered by oropharyngeal aspiration. Spermidine (Sigma, Cat# S2626-1G) or the spermidine synthesis inhibitor DL-α-Difluoromethylornithine (DFMO, Cayman Chemical, Cat# 16889; Ann Arbor, MI, USA) were administered at the time of inoculation. Three modules were designed in this project: LPS administration (5 mg/g body weight) + spermidine treatment (5 mg/g body weight); P.A. infection (1 × 10
6 cfu/mouse) + spermidine treatment (5 mg/g body weight); LPS administration (5 mg/g body weight) + DFMO treatment (1.5 mg/g body weight). For spermidine treatment modules, mice were injected intraperitoneally with a spermidine solution or PBS as a control, then anesthetized with isoflurane and challenged with LPS or PA intratracheally (IT). For DFMO treatment experiments, mice were pretreated with DFMO or PBS intraperitoneally for 1 h, after which all mice were anesthetized with isoflurane and intratracheally inoculated with LPS. 18 h after inoculation, mice from all modules were euthanized with CO
2 for further processing.
2.4. BAL and Lung Tissue Processing
Before harvesting the lungs, mouse BAL was retrieved with sterile PBS through the trachea. Cells were obtained by centrifuging the BAL fluid at 300×
g for 10 min at 4 °C. Cytospin slides were prepared from fresh BAL and stained for inflammatory cell differential counts (Cytospin 4, Thermo Scientific, Kwik-Dif Stains kitFisher Scientific, #99-907-00; Waltham, MA, USA). The right superior and inferior lobes were homogenized in 1 mL sterile PBS using the gentleMACS Octo Dissociator (Miltenyi Biotec; Bergisch Gladbach, Germany). A 100 mL aliquot of homogenate was reserved for the bacteria colony forming unit (CFU) assay. The remaining homogenate was centrifuged at 4 °C over 10,000×
g for 10 min, and the supernatant was stored at −80 °C for further use in ELISA and LDH assays. The right middle lobe was cut down and incubated in a 65 °C incubator for 48 h, and the wet/dry weight ratio as well as the edema levels were measured. The left inferior lobe was immersed in RLT buffer with 2-mercaptoethanol for RNA extraction. The left superior lobe was fixed with 10% neutral buffered formalin (Sigma, Cat# HT501128-4L) at 4 °C for over 24 h and subsequently subjected to H&E staining.
2.5. LDH Detection
LDH Cytotoxicity Assay (Promega, Cat# G1780; Madison, WI, USA) was used to quantify LDH levels as a function of cytotoxicity. The procedure was performed according to the manufacturer’s protocol.
2.6. Bactericidal Experiments with Spermidine
Live bacteria including KP396, KP-NDM, PA and PA14 were treated with 0, 0.5, 1, 2, 3 and 4mM spermidine for 4 and 24 h respectively. After the given time points, bactericidal activity was determined by using a CFU assay.
2.7. Bacteria CFU Assay
This assay was used to accurately assess bacteria burden. Briefly, the primary sample was serially diluted. A 10 mL aspirate from each diluted sample was transferred to a LB agar plate in technical triplicate. The agar plates were incubated at 37 °C overnight, and the resulting bacterial colonies were counted.
2.8. Primary T Cell Isolation
Fresh T cells from the spleen and draining lymph node in the lung were used for
in vitro experiments. Spleen and lung draining lymph nodes were harvested from WT mice after CO
2 euthanasia. Single cell suspensions from spleen and lymph node tissue were collected separately by crushing the organs with sterile, chilled PBS through a 70 mm filter and a syringe.
A mouse T cell isolation kit (STEMCELL, Cat# 19851; Vancouver, BC, Canada) was used to enrich T cells from harvested spleen cells. Unwanted cells were negatively purified from spleen cell suspensions by incubating spleen cells with biotinylated antibodies directed against non-T cells. The unwanted cells were magnetically isolated from the total spleen cells with a streptavidin-coated magnetic bead, enriching the spleen cell suspension with T cells.
2.9. Primary Cell Stimulation
Enriched spleen T cells and lung draining lymph node cells were plated (3–4 × 10
6 cells) in a 24-well plate, using 0.5 mL IMEM, 10% FBS, and standard Penicillin-Streptomycin antibiotics in each well. Cells were treated with various interventional conditions, as follows: 50 ng/mL m-IL-1β/m-IL-23; 50 ng/mL m-IL-1β/m-IL-23+3 mM spermidine; 50 ng/mL m-IL-1β/m-IL-23+20 mM DFMO; 50 ng/mL m-IL-1β/m-IL-23+3 mM spermidine+10 mM Ade (ChemFaces, Cat# CFN99215; Wuhan, China). For DFMO experiments, DFMO and control PBS were added to the cells 4 h before IL-1β/IL-23 stimulation to block spermidine synthesis. For other experimental groups, all treatments were administered to the cells at the time of stimulation. At the 18 h time point, cells were harvested for RNA-seq, spermidine quantification by high-performance liquid chromatography (HPLC), or lysed in RLT buffer with 2-mercaptoethanol for gene expression analysis. The supernatant was used for ELISA experiments.
2.10. RNA Extraction and cDNA Synthesis
Cell and lung tissue RNA was extracted with RNeasy Miniprep Kit (QIAGEN, Cat# 74136; Hilden, Germany), in accordance with the manufacturer’s instructions. cDNA was synthesized using qScriptTM cDNA Synthesis Kits (Quantabio, Cat# 95047-100; Beverly, MA, USA).
2.11. Real-Time PCR
Real-time PCR was constructed and read by the Bio-Rad CFX96 system using TaqMan PCR Master Mix (Bio-Rad, Cat# 1725284; Hercules, CA, USA) and pre-mixed probe sets:
Il17a (Mm00439618_m1),
Il17f (Mm00521423_m1),
Il22 (Mm01226722_g1),
Ifng (Mm01168134_m1),
Srm (Mm00726089_s1),
Smox (Mm01275475_m1),
Prdx1 (Mm01621996_s1),
Gapdh (Mm99999915_g1) and
Hprt (Mm03024075_m1), from Thermo Scientific.
2.12. ELISA
ELISA kits were used to measure mouse IL-17A (BioLegend, Cat# 432506) and mouse IFN-γ (BioLegend, Cat# 430804). The procedures were strictly employed according to the manufacturer’s instructions.
2.13. ScRNA-Seq, RNA-Seq and Total-Seq
ScRNA-seq libraries were constructed and sequenced following the “Single Cell 3′ Reagent Kits v2 User Guide” (10× Genomics, Pleasanton, CA, USA), and cell viabilities between all samples ranged from 58.5% to 79.4%. The RNA-seq methodology was developed in close reference to a published study [
18]. Similar to ScRNA-seq, Total-seq was performed following the cell hashing protocol (BioLegend). Briefly, five groups of cells were pre-labelled with mouse HashTag oligos (HTO) on cell surface: spleen T cell control group (SPT ctrl) with hashtag 1 (BioLegend, Cat# 155801); spleen T cell IL-1β/IL-23 stimulation group (SPT stim) with hashtag 2 (BioLegend, Cat# 155803); spleen T cell IL-1β/IL-23 plus spermidine treatment group (SPT sper) with hashtag 3 (BioLegend, Cat# 155805); lymph node cell IL-1β/IL-23 stimulation group (LN stim) with hashtag 4 (BioLegend, Cat# 155807); lymph node cell IL-1β/IL-23 plus spermidine treatment group (LN sper) with hashtag 5 (BioLegend, Cat# 155809). We targeted 4000 cells per sample, totalling 20,000 cells. The five cell populations were mixed and barcoded, and barcoded cDNA was synthesized via reverse transcription. Further cDNA libraries were constructed by cDNA amplification. HTO-derived cDNAs (<180 bp) and mRNA-derived cDNAs (>300 bp) were separated using a magnetic beads selection methodology. An HTO sequencing library was generated for sequencing, followed by another round of cDNA amplification. The mRNA-derived cDNA library was constructed following the conventional scRNA-seq protocol. The HTO library and DNA library were then sequenced on an Illumina HiSeq sequencer (MedGenome, Inc.; Foster City, CA, USA). Sequencing data were initially analyzed using Cell Ranger (10× Genomics) and downstream analysis was performed using R. The R package Seurat (v5.0.0) was used to perform HTO demultiplexing, doublet, negative, and low quality cell filtration, as well as unsupervised clustering. Differential gene expression was performed using the Wilcoxon method and with the “findmarkers” function in Seurat.
2.14. Statistics
All data analyses were achieved and graphed by Prism 8.0 (GraphPad). The one-way ANOVA test was performed to compare differential gene expression among four groups. Paired Student’s
t-test was applied for other comparisons between the two paired groups.
3. Results
3.1. Enzymes for Spermidine Synthesis (SRM, SMOX) Are Upregulated, and Spermidine Is Downregulated upon Th17/IL-17 Pathway Activation in Th17 Cells
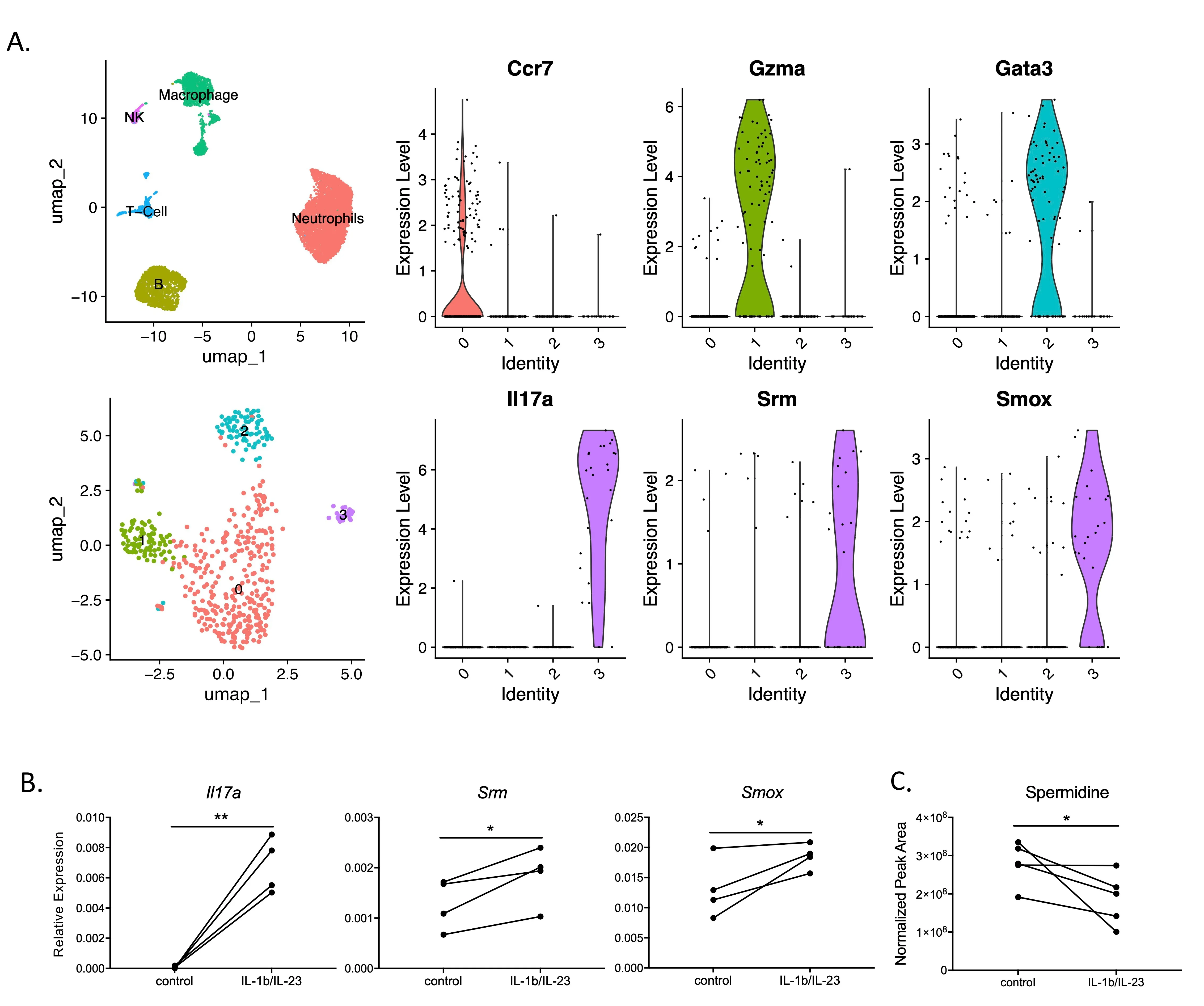
To determine the molecular pathways implicated in Th17/IL-17 signaling, we carefully analysed a dataset where mouse peritoneal lavage cells were stimulated with IL-1β/IL-23 and assessed with scRNA-Seq, with a focus on four subsets of T-cells (
A). Surprisingly,
Srm and
Smox genes were found to be upregulated in IL-17+ expressing cells (
A). Spermidine synthase (SRM) and spermine oxidase (SMOX) are both catalytic enzymes for spermidine synthesis. To further explore the relationship between spermidine and the Th17/IL-17 pathway, we assessed
Srm and
Smox expression levels with qPCR in splenic T cells derived from adult mice. Adult mice were used for their mature immune systems which already differentiated Th17 cells, allowing for the investigation of spermidine’s direct role on T cells independent of the differentiation regulatory pathways.
Srm and
Smox were highly expressed in IL-17 producing cells upon stimulation by IL-1β/IL-23 (
B). Next, to further explore the link between spermidine and Th17 cytokine production, we directly measured the levels of spermidine and other polyamines (spermine and putrescine) in T cells after activation of the Th17/IL-17 pathway. Despite the upregulation of spermidine synthesis enzymes, lower spermidine levels were observed after Th17/IL-17 pathway stimulation (
C), whereas other polyamines remained unchanged (results not shown). The
Srm and
Smox gene expression profiles, as well as changes in spermidine levels in mature T cells suggest that spermidine may be involved in regulating the Th17/IL-17 pathway independently, due to its role in T cell differentiation.
. Polyamine metabolism is involved in IL-17 production. (<strong>A</strong>) Thioglycollate-induced mouse peritoneal lavage cells were collected and stimulated with IL-1b/IL-23, and scRNA data confirmed IL-17-producing cells had high <em>Srm</em> and <em>Smox</em> expression. (<strong>B</strong>,<strong>C</strong>) Spleen T cells were stimulated with mouse IL-1b+IL-23 overnight, (<strong>B</strong>) <em>Il17a</em>, <em>Srm</em> and <em>Smox</em> were highly induced, gene expression relative to <em>Gapdh </em>(graphs displayed in panel B are representative ones from four repeated experiments, N = 2–4 in each experiment); (<strong>C</strong>) Polyamine in the cytoplasm of T cells were measured by HPLC system. <sup>∗</sup><em>p</em> < 0.05, <sup>∗∗</sup> <em>p</em> < 0.01 by paired Student <em>t</em>-test.
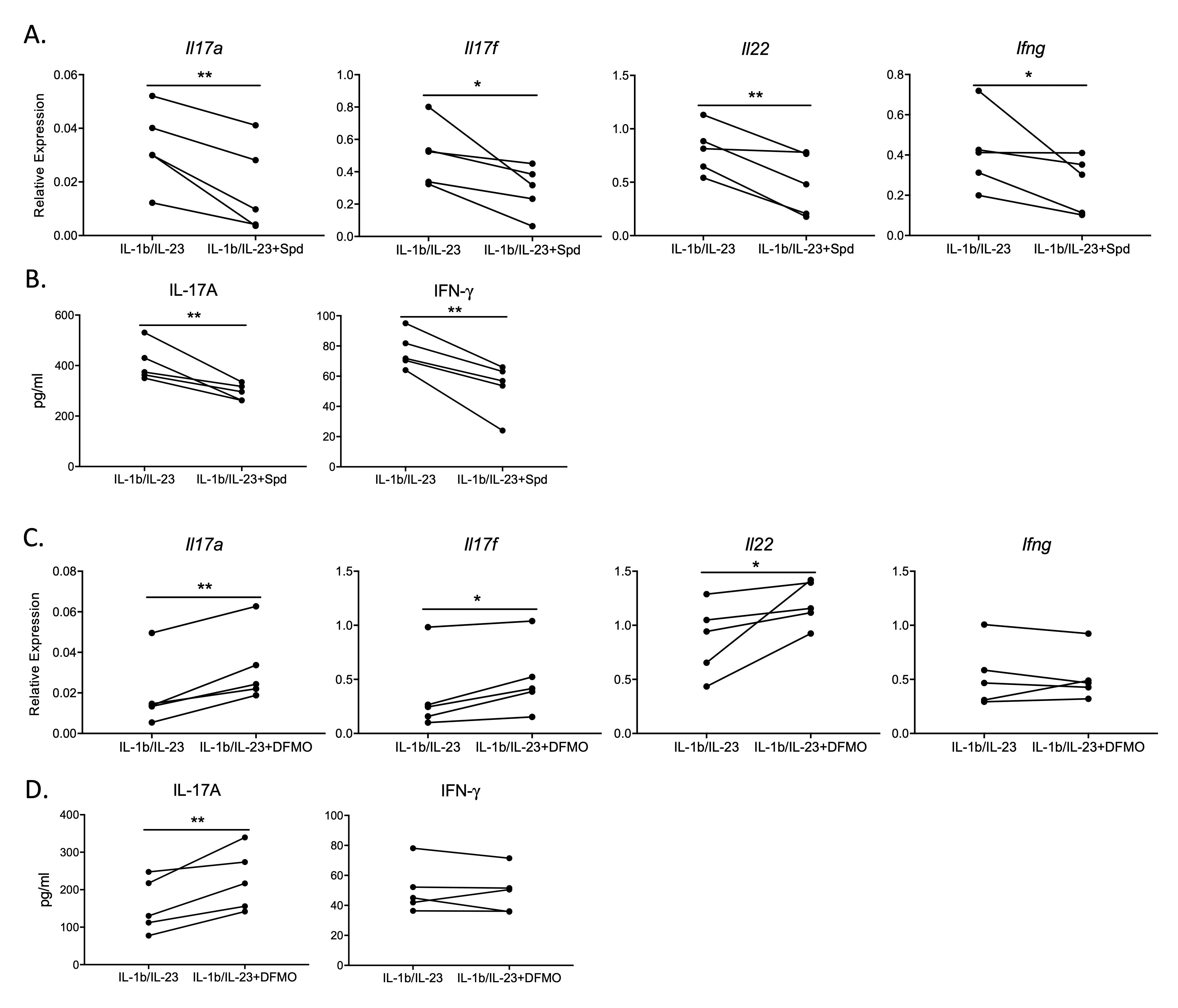
To determine the direct effect of spermidine on the Th17/IL-17 pathway, the cytokine expression profile of activated T cells was measured in the presence of exogenous spermidine. We monitored cytokine regulation by measuring mRNA expression and protein levels for Th17 effector cytokines.
Upon administration of spermidine, both the gene expression and protein levels of Th17/IL-17 cytokine products were downregulated in T cells stimulated with IL-1β/IL-23 (
A,B). We also observed downregulated gene expression and cytokine levels for the Th1 effector cytokine IFN-γ, which is typically activated in conjunction with the Th17/IL-17 pathway during the fight against foreign infections [
19,
20]. These results indicate that spermidine acts directly on T cells in downregulating Th17/IL-17 pathway products.
Upon administration of difluoromethylornithine (DFMO), an inhibitor of spermidine synthesis [
21], the expression of Th17/IL-17 pathway product genes
Il17a,
Il17f,
Il22 and IL-17A protein was enhanced above IL-1β/IL-23 stimulation alone (
C,D). However, DFMO did not affect the IFN-γ pathway (
C,D). This result reinforces that spermidine may be one of the key native regulatory points specific to the Th17/IL-17 pathway.
. Spermidine downregulates Th17/IL-17 by directly targeting T cells <em>in vitro</em>. Spleen T cells were stimulated with IL-1b/IL-23 in the presence or absence of spermidine or DFMO treatment overnight. This is a representative graph of five experiments, N = 2–4 in each experiment. (<strong>A</strong>) <em>Il17a</em>, <em>Il17f</em>, <em>Il22</em> and <em>Ifng</em> gene expression was measured by qPCR (relative expression to <em>Hprt</em>). (<strong>B</strong>) IL-17A and IFN-g proteins expression tested by ELISA. (<strong>C</strong>): <em>Il17a</em>, <em>Il17f</em>, <em>Il22</em> and <em>Ifng</em> gene expression was measured by qPCR (relative expression to <em>Hprt)</em>. (<strong>D</strong>): IL-17A and IFN- g proteins were tested by ELISA. <sup>∗</sup> <em>p</em> < 0.05, <sup>∗∗</sup> <em>p</em> < 0.01 by paired Student <em>t</em>-test.
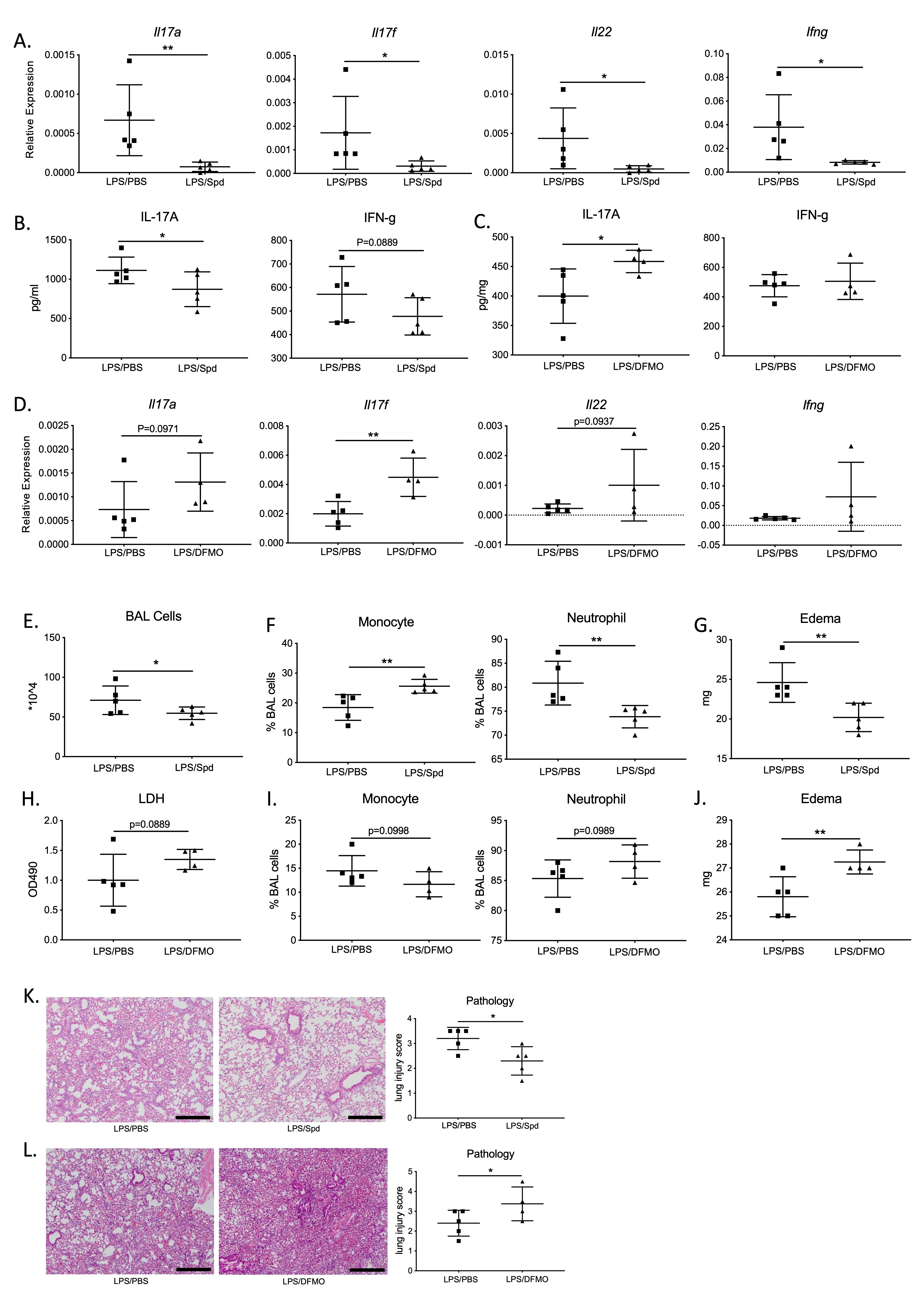
With spermidine’s role in the negative regulation of Th17/IL-17 signaling established
in vitro, we assessed spermidine’s effects on Th17/IL-17 signaling using an acute lung injury (ALI) mouse model. Bacterial infections damage the host lung through two primary mechanisms: (1) uncontrolled bacterial overgrowth, and (2) excessive inflammation that leads to self-inflicted tissue damage. The ALI model is based on the context of bacterial pneumonia, where WT mice are inoculated with LPS to induce lung inflammation and injury, with and without coadministration of spermidine. In this acute injury mouse model, spermidine downregulated the Th17/IL-17 pathway expression, including
Il17a,
Il17f and
Il22 genes, as well as IL-17A cytokine levels (
A,B). At the same time, the IFN-γ pathway, which is closely associated with the Th17/IL-17 pathway as previously described, was also inhibited by spermidine, resulting in decreased expression of the
Ifng gene (
A,B). Inhibition of native spermidine synthesis with DFMO resulted in the upregulation of Th17/IL-17 pathway genes, such as
Il17a,
Il17f, and
Il22, as well as IL-17A protein expression (
C,D). Consistent with the corresponding
in vitro results, DFMO did not affect IFN-γ levels
in vivo (
C,D).
. Spermidine dampens inflammation through inhibiting IL-17 production in an acute lung injury (ALI) <em>in vivo</em> model. (<strong>A</strong>,<strong>B</strong>) Lung tissue <em>Il17a</em> and <em>Ifng</em> mRNA (<strong>C</strong>) and protein (<strong>D</strong>) were obtained by ELISA and qPCR (relative expression to <em>Hprt), </em>N = 5 in both PBS and spermidine group. (<strong>C</strong>,<strong>D</strong>) Expression of <em>Il17, Ifng</em> and <em>Il22</em> in ALI model after DFMO treatment, N = 5 in PBS group; N = 4 in DFMO group. (<strong>E</strong>–<strong>G</strong>) Assessment of inflammation by BAL cellularity and edema quantification, in PBS and spermidine treated lungs. (<strong>H</strong>–<strong>J</strong>) Evaluation of inflammatory parameters after blocking spermidine with DFMO. (<strong>K</strong>,<strong>L</strong>) Lung tissue histology injury score assessment in ALI model after spermidine or DFMO. Scale bars: 300 μm. <sup>∗</sup> <em>p</em> <0.05, <sup>∗∗</sup> <em>p</em> < 0.01 by unpaired Student <em>t</em>-test.
We assessed the degree of lung inflammation and damage in this mouse model. We found that spermidine-treated mice had less severe inflammation and lung damage compared to the PBS-treated group. There was an overall fewer number of immune cells in the bronchoalveolar lavage (BAL) fluid of the spermidine-treated group compared to the controls (
E). Immune cells from spermidine-administered lungs had higher proportions of monocytes and lower proportions of neutrophils (
F). In addition, edema levels were reduced in spermidine- treated groups compared to the PBS-treated group (
G). Collectively, these measurements indicate lower levels of inflammation in the lungs as a result of spermidine treatment. Not only did spermidine result in lower degrees of inflammation, but it also protected against lung damage according to lung histology (
K).
Inhibition of endogenous spermidine synthesis with DFMO promoted lung inflammation and injury, resulting in higher cytotoxicity, proportionally higher neutrophil counts, and elevated levels of lung edema (
H,J). Elevated levels of injury and inflammatory infiltration in DFMO-treated lungs were also evident in lung histology (
L). Overall, the combination of lower expression of Th17/IL-17 pathway products and protected lung phenotypes suggests that spermidine protects against lung inflammation and injury by downregulating the Th17/IL-17 signaling pathway.
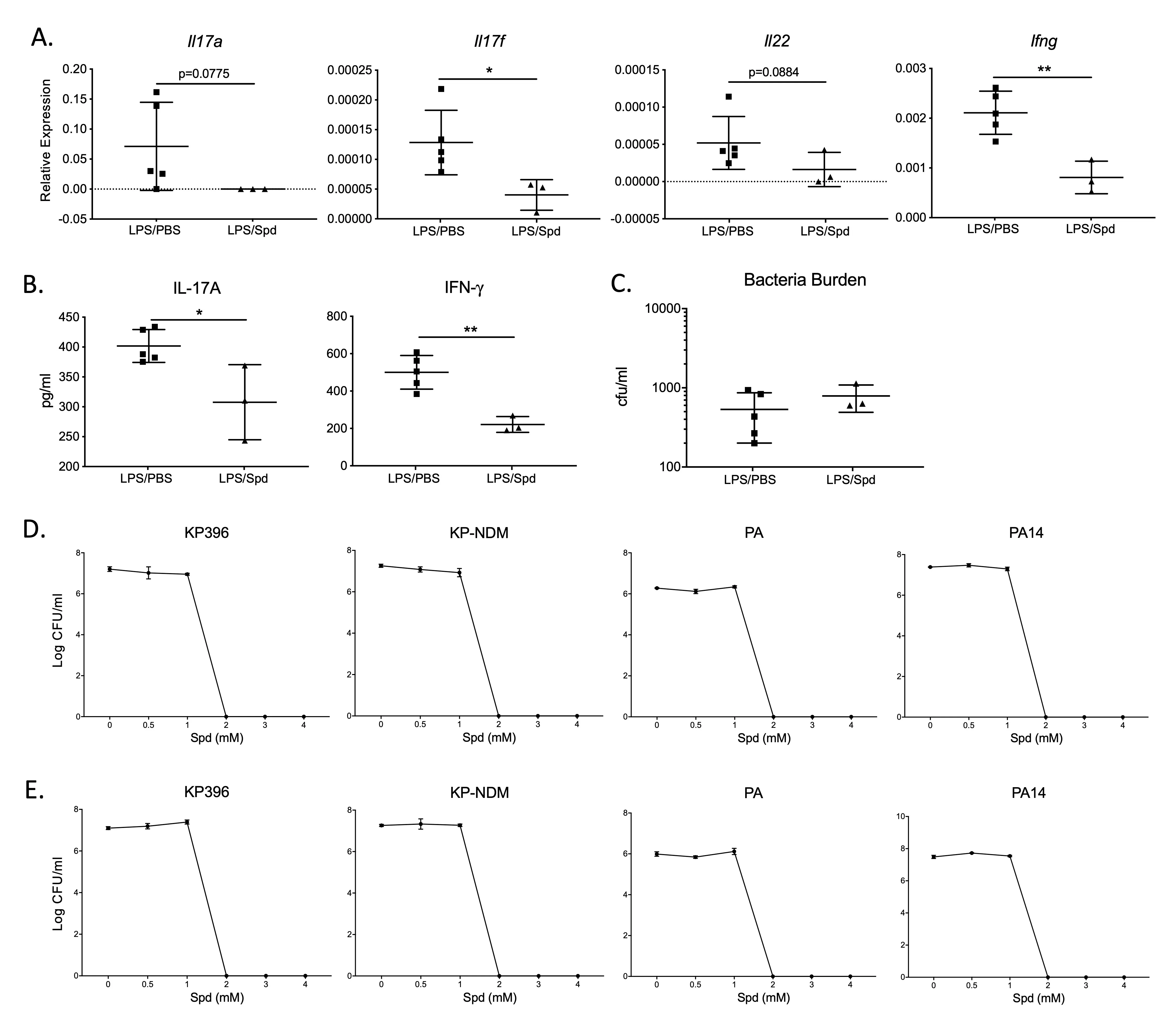
3.4. Spermidine Downregulates the Th17/IL-17 Pathway in an Acute Live Bacterial Infection Mouse Model
A live bacterial infection mouse model was used to assess lung bacterial clearance upon spermidine modulation of the Th17/IL-17 pathway. It is well established that the body’s response to KP (
Klebsiella pneumoniae) and PA (
Pseudomonas aeruginosa), two gram-negative bacilli, is mainly mediated by the Th17/IL-17 pathway [
22,
23]. In a PA infection model, the spermidine- treated group inhibited Th17/IL-17 pathway expression, but it did not affect PA bacterial clearance (
A–C). Spermidine down-regulated Th17/IL-17 pathway expression, including
Il17a,
Il17f and
Il22 genes and IL-17A protein expression (
A,B). There was no difference in BAL protein concentration between the spermidine-treatment group and the control group (results not shown).
Upregulation of the Th17/IL-17 pathway expression is expected to enhance bacterial clearance, while inhibiting Th17/IL-17 signaling can lead to bacterial retention [24,25]. Inhibiting the expression of the Th17/IL-17 pathway with spermidine may therefore hinder bacterial clearance in mice, posing a potential concern for elevated infection risk during inflammation management. However, the quantitative measurement of residual bacterial burden in the lungs showed that spermidine did not adversely affect the clearance of PA bacteria (Figure 4C). To further investigate this unimpaired bacterial clearance, we directly administered spermidine to different subtypes of KP (KP396, KP-NDM) and PA (PA ATCC10145, PA14)
in vitro. Both KP and PA were completely eradicated after 4 h of spermidine treatment at spermidine concentrations of ≥2 mM (Figure 4D), and the bactericidal effect of spermidine after 24 h of treatment was the same as that of after 4h (Figure 4E). The direct bactericidal effect of spermidine may aid in bacterial clearance, maintaining mucosal barrier homeostasis despite the downregulation of Th17/IL-17 pathway expression.
To further determine whether spermidine targets T cells to reduce inflammation, we used T cell knockout mice to investigate whether spermidine acts on other immune cells. We established LPS-induced acute lung injury and live bacterial infection models in
Rag2/II2rg double gene knockout mice, which are deficient in T, B and NK cells. The Th17 cytokines were completely absent in these mice according to RT-PCR (data not shown), indicating that T cell function was knocked out. As shown, spermidine did not protect against inflammation in
Rag2/II2rg mice, when these were subjected to ALI and PA14 infection (
A–C and
D–F respectively). All markers of inflammation were elevated with or without spermidine administration, including the number of BAL immune cells, the proportion of monocytes and neutrophils, BAL total protein, markers of lung edema, and lung histopathological score (
). This stark loss in spermidine’s anti-inflammatory effect supports the fact that spermidine is likely acting upon Th17 T cells in the Th17/IL-17 pathway and not through other immune cells.
. Spermidine treatment suppresses Th17/IL-17 pathway in a live PA bacteria infected mouse model. (<strong>A</strong>,<strong>B</strong>) Th17/IL-17 pathway-dependent genes in PA-infected mice. N = 5 in PBS group; N = 3 in Spermidine (Spd) group. Gene expression relative expression to <em>Hprt</em>. (<strong>C</strong>) Lung bacterial burden after PA infection (18h post-infection). (<strong>D</strong>,<strong>E</strong>) Bactericidal activity of varying concentrations of spermidine. Live bacteria including KP396, KP-NDM, PA (as PA ATCC10145) and PA14 were treated with 0, 0.5, 1, 2, 3 and 4 mM spermidine for 4 h (<strong>D</strong>) and 24 h respectively (<strong>E</strong>). <sup>∗</sup> <em>p</em> < 0.05, <sup>∗∗</sup> <em>p</em> < 0.01 by unpaired Student <em>t</em>-test.
. Lymphocyte- depleted <em>Rag2/Il2rg</em> double KO mice do not respond to spermidine. <em>Rag2/II2rg</em> double knockout mice were injected with Spermidine (50 μg/g) or PBS i.p twice a day. (<strong>A</strong>–<strong>C</strong>) Spermidine or PBS pre-treated mice were inoculated with 5 μg/g LPS by intratracheal instillation (IT) to develop ALI. Mice were assessed for inflammatory markers: cellular recruitment, protein, edema and pathology score. Scale bars: 300 μm. (<strong>D</strong>–<strong>F</strong>) Spermidine or PBS pre-treated mice were infected with live PA14 by IT. Mice were assessed for inflammatory markers: cellular recruitment, protein, edema and pathology score. Scale bars: 300 μm.
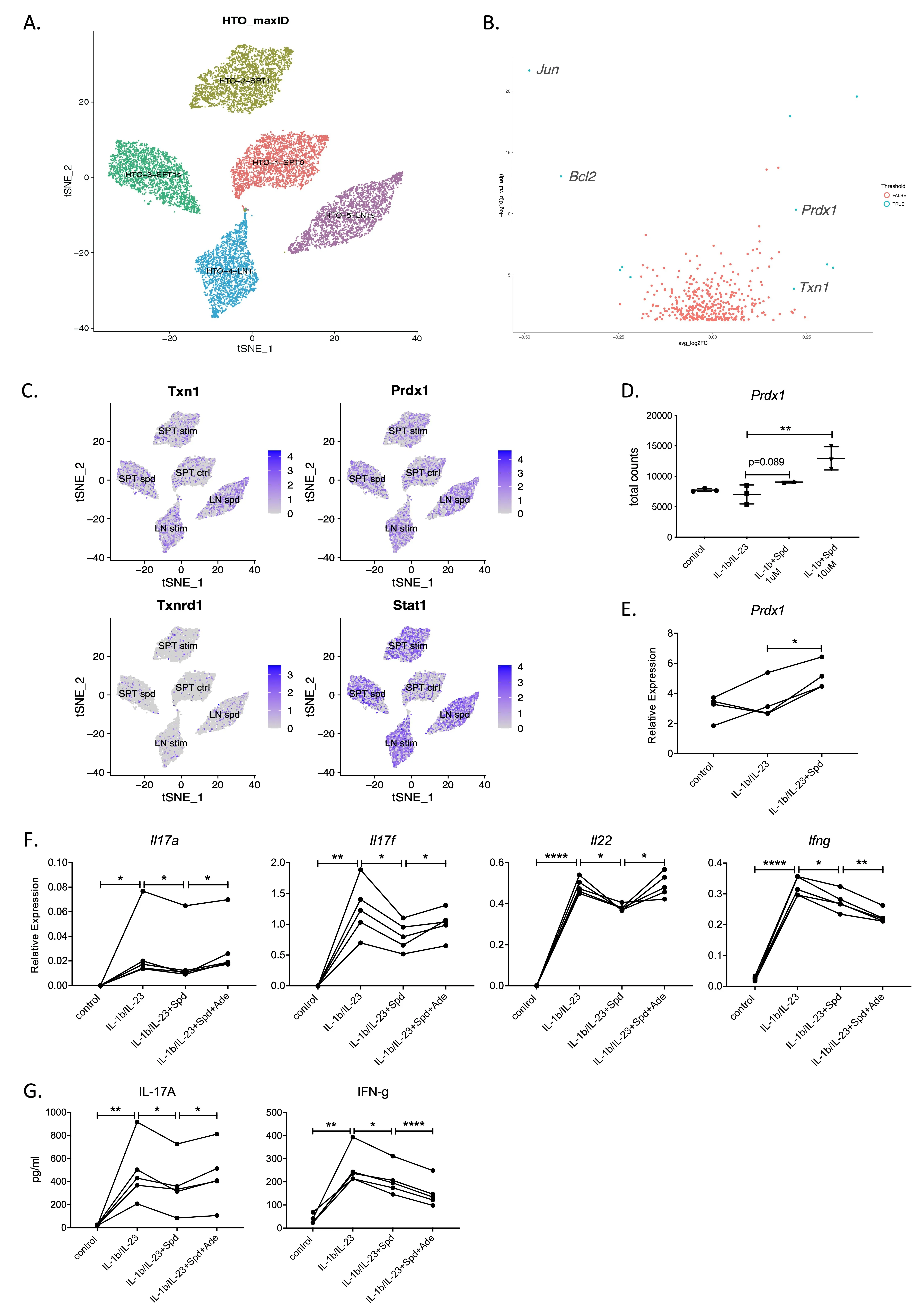
To determine the intrinsic mechanism by which effector Th17 cytokines are downregulated by spermidine in T cells, we performed scRNA-Seq and bulk mRNA sequencing analysis on IL-1β/IL-23 activated splenic T cells (SPT) and lymph node (LN) cells treated with spermidine. To minimize batch effects between samples, we multiplexed all samples using the cell hashing method [
26] for spermidine treatment (SPT sper and LN sper) and untreated groups (SPT stim and LN stim) (
A). After de-multiplexing [
27], the results showed that compared with the untreated T cell control group (SPT stim), the expression of the
Txn1 and
Prdx1 genes was significantly increased in the spermidine treatment group (
B,C).
Prdx1 encodes peroxidase 1 (PRDX1), which is an antioxidant enzyme that protects the cell by reducing hydrogen peroxide retention [
28,
29]. Coincidentally, thioredoxin (TXN1) is an oxidoreductase also with antioxidant activities [
30,
31]. Other genes, such as
Txnrd1 and
Stat1 did not change significantly. We further confirmed this using mRNA-Seq data that the expression of the
Txn1 and
Prdx1 genes in T cells in the spermidine treatment group increased directly in proportion to the concentration of spermidine (
D). In the subsequent RT-PCR verification, we observed that only the
Prdx1 gene was highly expressed in the spermidine treatment group (
E).
Prdx1 expression had a strong association with spermidine treatment, raising the possibility of PRDX1 being a key spermidine effector.
To determine whether spermidine regulates Th17 cytokine production through PRDX1, adenanthin was co-administered with spermidine into activated Th17 cells. Adenanthin is a recently discovered inhibitor of antioxidant enzymes, including PRDX1 [
32,
33]. As previously observed, IL-1β/IL-23 activated the Th17/IL-17 and IFN-γ pathways, while spermidine significantly down-regulated Th17/IL-17 and IFN-γ products (
F). Compared to spermidine treatment alone, the addition of adenanthin enhanced the gene expression of
Il17a,
Il17f,
Il22 as well as the levels of Th17/IL-17 products (
F,G). Adenanthin successfully reversed the inhibitory effect of spermidine on the Th17/IL-17 pathway, suggesting that spermidine downregulates the Th17/IL-17 pathway by regulating PRDX1 activity. Interestingly, the combination of adenanthin and spermidine treatment further inhibited the gene and protein expression of IFN-γ (
F,G), which needs to be further explored in future investigations.
Adenanthin and DFMO are both inhibitors of spermidine’s action against Th17/IL-17 signaling. DFMO inhibits the upstream of spermidine during spermidine synthesis, while adenanthin inhibits the downstream of spermidine by inhibiting PDRX1. Administration of DFMO reverses the inhibitory effect of spermidine on the Th17/IL-17 pathway and enhances the expression of Th17/IL-17 pathway products. In contrast, coadministration of adenanthin with spermidine did not completely restore the expression of Th17/IL-17 pathway products. These data suggest that spermidine’s activity is not completely inhibited by suppressing PDRX1. These results suggest that spermidine may inhibit the expression of Th17/IL-17 products through additional mechanisms, which require further investigation.
Our data establish that spermidine directly suppresses IL-17 production in differentiated Th17/memory T cells by upregulating peroxiredoxin-1 (PRDX1). The complete reversal of spermidine-mediated IL-17 inhibition upon PRDX1 blockade supports this conclusion. While the precise molecular mechanism requires further investigation, PRDX1’s well-characterized role as an antioxidant enzyme suggests that spermidine may mitigate redox-sensitive signaling pathways (e.g., NF-κB/STAT3) triggered by IL-1β/IL-23 stimulation. This pathway operates independently of T-cell differentiation and represents a novel metabolic checkpoint for IL-17 regulation.
. Spermidine inhibits IL-17 production in a <em>Prdx1</em> dependent manner. (<strong>A</strong>) scRNA-Seq integration and cell cluster label of fresh lymph node cells and enriched spleen T cells, stimulated with IL-1b/IL-23 with or without 3 μM Spermidine (Spd) overnight. Five major cell populations were identified: spleen T cell control group (SPT ctrl); spleen T cell IL-1b/IL-23 stimulation group (SPT stim); spleen T cell IL-1b/IL-23 plus spermidine treatment group (SPT sper); lymph node cell IL-1b/IL-23 stimulation group (LN stim); lymph node cell IL-1b/IL-23 plus spermidine treatment group (LN sper). (<strong>B</strong>) Volcano plots representing differentially expressed genes between IL-1b/IL-23 group vs IL-1b/IL-23 plus spermidine treatment group (cutoffs: LogFC = 0.2 and padj = 0.001). (<strong>C</strong>) <em>Txn1</em> and <em>Prdx1</em> gene expression after spermidine treatment of IL-1b/IL-23—activated IL-17-producing cells. (<strong>D</strong>,<strong>E</strong>) Both mRNA-seq data (<strong>D</strong>) and RT-PCR (<strong>E</strong>) revealed <em>Prdx1</em> as a potential target for spermidine. N = 3. (<strong>F</strong>,<strong>G</strong>) Enriched spleen T cells were stimulated with IL-1b/IL-23, IL-1b/IL-23 + Spd, IL-1b/IL-23 + 3uM Spd + 10uM Adenanthin (Ade) separately, incubating overnight. Ade treatment reversed the suppression of Th17/IL-17 pathway induced by spermidine. <em>Il17a</em>, <em>Il17f</em>, <em>Il22</em> and <em>Ifng</em> gene expression was measured by qPCR (<strong>F</strong>), relative expression to <em>Hprt</em>; IL-17A and IFN-g proteins were tested by ELISA (<strong>G</strong>). This figure is a representative of four experiments, N = 2–3 in each experiment. <sup>∗</sup> <em>p</em> < 0.05, <sup>∗∗</sup> <em>p</em> < 0.01, <sup>∗∗∗∗</sup> <em>p</em> < 0.001, by Student <em>t</em>-tests.
4. Discussion
The pivotal role of the Th17/IL-17 signaling pathway in the body’s immune system highlights the importance of studying the regulatory mechanism of this pathway. Understanding the regulatory mechanisms of the Th17/IL-17 signaling pathway, especially the regulatory factors of Th17 cells, have always been of great interest for immunology research. Th17 cells have been reported to be inhibited by cytokines secreted by most other types of T cells such as Th1 cells and Th2 cells, which play important roles in controlling inflammatory cascades and hyper-immunity [
34,
35,
36]. Regulatory T cells play a role in monitoring and regulating Th17 cells, but the specific regulatory mechanisms for the Th17/IL-17 signaling pathway have been elusive [
37,
38].
Polyamines, especially spermidine and spermine, have been implicated with inflammation, with elevated levels of polyamines found in the inflammatory sites of infection, trauma, tumor and autoimmune diseases [
39,
40]. Prior to this study, the mechanism by which polyamines enact their anti-inflammatory effects had not yet been established. In this study, we identified a key regulatory mechanism of spermidine in the Th17/IL-17 signaling pathway. Based on both
in vitro and
in vivo models, we found that spermidine directly acts on T cells to downregulate the Th17/IL-17 signaling pathway. Based on RNA sequencing data, we determined that spermidine downregulates the Th17/IL-17 signaling pathway by controlling PRDX1 protein activity in T cells. PRDX1 protein is primarily involved in antioxidant functions, helping to remove reactive oxygen species (ROS) within cells. PRDX1 activity is crucial during periods of cellular oxidative stress, such as in inflammation, where excessive amounts of reactive oxygen species are produced in the cell [
41,
42]. Our observation that spermidine acts through PRDX1 is consistent with the literature, which indicates that spermidine is involved in controlling oxidative stress [
43,
44,
45]. In addition, prior studies have suggested that Spd and PRDX1 can both inhibit the inflammatory master transcription factor NF-kB. Spd inhibits NF-kB by blocking its translocation into the nucleus [
9], PRDX1 regulates upstream signal transduction by degrading the ligase TRAF6 [
46]. Taken together, the effects of Spd and PRDX1 represent a much stronger double inhibition of the NF-kB transcription factor, which is responsible for multiple key inflammatory genes such as IL-1B and TNF.
We also found that spermidine treatment, despite its immune suppressive role, does not negatively affect lung bacterial clearance in mice. In addition to its anti-inflammatory and immunomodulatory effects, spermidine was found to have a direct bactericidal impact against PA and KP. These results suggest that spermidine may be a key factor in the host-infection interface, maintaining mucosal barrier homeostasis by controlling infection without detrimental levels of inflammation. A further understanding of how spermidine endogenously regulates the dynamic balance between Th17/IL-17 suppression and infection will guide clinical strategies against infection-induced hyper-immunity.
Further investigations are necessary to explore other molecular mechanisms by which spermidine inhibits the Th17/IL-17. Our comparison of DFMO and adenanthin, inhibitors of spermidine synthesis and effectors, respectively, indicates that spermidine may have additional Th17/IL-17 pathway effectors besides PRDX1. In addition, whether other polyamines, which have also been implicated with anti-inflammatory effects, engage in a similar regulatory mechanism as spermidine must also be investigated [
36,
37]. Furthering the mechanistic understanding of Th17/IL-17 regulation by endogenous and exogenous spermidine may aid efforts to clinically manage debilitating cases of infection and autoimmune diseases.
Acknowledgements
This project is supported by HL137709, HL154231 and AI153549 from the National Institute of Health.
Author Contributions
Conceptualization, G.T.-N. and K.C.; Methodology, J.L., Y.J.K., S.C., X.A., L.F., C.E., D.L., A.A.F., Y.P., G.T.-N., K.C.; Validation, S.C. and L.F.; Formal Analysis, J.L., S.C. and D.L.; Investigation, X.X.; Resources, X.X.; Data Curation, X.X.; Writing—J.L., G.T.-N., K.C.; Writing—Review & Editing, J.L., Y.J.K., S.C., D.L., Y.P., G.T.-N., K.C.; Visualization, S.C., D.L.; Supervision, G.T.-N. and K.C.; Funding Acquisition, G.T.-N. and K.C.
Ethics Statement
All the mice were used with approval from the University of Pittsburgh Institutional Animal Care and Use Committee, Protocol #: 21028745, 2/2/2021.
Informed Consent Statement
Not applicable.
Data Availability Statement
R code used for RNA-seq analysis is available upon request. The sequencing data reported in this paper will be deposited in the Gene Expression Omnibus.
Funding
This project is supported by HL137709 (to K.C.), HL154231 (to G.T.-N.) and AI153549 (to G.T.-N.)
Declaration of Competing Interest
None of authors have any conflict of interest to report.
References
-
1.
Pappu R, Ramirez-Carrozzi V, Sambandam A. The interleukin-17 cytokine family: critical players in host defence and inflammatory diseases.
Immunology 2011,
134, 8–16. doi:10.1111/j.1365-2567.2011.03465.x.
[Google Scholar]
-
2.
Beisswenger C, Honecker A, Kamyschnikow A, Bischoff M, Tschernig T, Bals R. Moxifloxacin modulates inflammation during murine pneumonia.
Respir. Res. 2014,
15, 82. doi:10.1186/1465-9921-15-82.
[Google Scholar]
-
3.
Xu X, Shao B, Wang R, Zhou S, Tang Z, Lu W, et al. Role of Interleukin-17 in defense against pseudomonas aeruginosa infection in lungs.
Int. J. Clin. Exp. Med. 2014, 7, 809–816.
[Google Scholar]
-
4.
Dubin PJ, Kolls JK. IL-23 mediates inflammatory responses to mucoid Pseudomonas aeruginosa lung infection in mice.
Am. J. Physiol. Lung Cell. Mol. Physiol. 2007,
292, L519–528. doi:10.1152/ajplung.00312.2006.
[Google Scholar]
-
5.
Dubin PJ, Martz A, Eisenstatt JR, Fox MD, Logar A, Kolls JK. Interleukin-23-mediated inflammation in Pseudomonas aeruginosa pulmonary infection.
Infect. Immun. 2012,
80, 398–409. doi:10.1128/IAI.05821-11.
[Google Scholar]
-
6.
Pegg AE. Mammalian polyamine metabolism and function.
IUBMB Life 2009,
61, 880–894. doi:10.1002/iub.230.
[Google Scholar]
-
7.
Pegg AE. The function of spermine.
IUBMB Life 2014,
66, 8–18. doi:10.1002/iub.1237.
[Google Scholar]
-
8.
Morón B, Spalinger M, Kasper S, Atrott K, Frey-Wagner I, Fried M, et al. Activation of protein tyrosine phosphatase non-receptor type 2 by spermidine exerts anti-inflammatory effects in human THP-1 monocytes and in a mouse model of acute colitis.
PLoS ONE 2013,
8, e73703. doi:10.1371/journal.pone.0073703.
[Google Scholar]
-
9.
Choi YH, Park HY. Anti-inflammatory effects of spermidine in lipopolysaccharide-stimulated BV2 microglial cells.
J. Biomed. Sci. 2012,
19, 31. doi:10.1186/1423-0127-19-31.
[Google Scholar]
-
10.
Jeong JW, Cha HJ, Han MH, Hwang SJ, Lee DS, Yoo JS, et al. Spermidine Protects against Oxidative Stress in Inflammation Models Using Macrophages and Zebrafish.
Biomol. Ther. 2018,
26, 146–156. doi:10.4062/biomolther.2016.272.
[Google Scholar]
-
11.
Yang Q, Zheng C, Cao J, Cao G, Shou P, Lin L, et al. Spermidine alleviates experimental autoimmune encephalomyelitis through inducing inhibitory macrophages.
Cell Death Differ. 2016,
23, 1850–1861. doi:10.1038/cdd.2016.71.
[Google Scholar]
-
12.
Paul S, Kang SC. Natural polyamine inhibits mouse skin inflammation and macrophage activation.
Inflamm. Res. 2013,
62, 681–688. doi:10.1007/s00011-013-0620-5.
[Google Scholar]
-
13.
Carriche GM, Almeida L, Stüve P, Velasquez L, Dhillon-LaBrooy A, Roy U, et al. Regulating T-cell differentiation through the polyamine spermidine.
J. Allergy Clin. Immunol. 2021,
147, 335–348.e311. doi:10.1016/j.jaci.2020.04.037.
[Google Scholar]
-
14.
Puleston DJ, Baixauli F, Sanin DE, Edwards-Hicks J, Villa M, Kabat AM, et al. Polyamine metabolism is a central determinant of helper T cell lineage fidelity.
Cell 2021,
184, 4186–4202.e4120. doi:10.1016/j.cell.2021.06.007.
[Google Scholar]
-
15.
Wagner A, Wang C, Fessler J, DeTomaso D, Avila-Pacheco J, Kaminski J, et al. Metabolic modeling of single Th17 cells reveals regulators of autoimmunity.
Cell 2021,
184, 4168–4185.e4121. doi:10.1016/j.cell.2021.05.045.
[Google Scholar]
-
16.
Pennec X, Cachier P, Ayache N. Understanding the “Demon’s Algorithm”: 3D Non-rigid Registration by Gradient Descent; Springer: Berlin/Heidelberg, Germany, 1999.
-
17.
Moghbeli K, Lipp MA, Bueno M, Craig A, Rojas M, Abbas M, et al. NKG2D blockade impairs tissue-resident memory T cell accumulation and reduces chronic lung allograft dysfunction.
JCI Insight 2025,
10, e184048. doi:10.1172/jci.insight.184048.
[Google Scholar]
-
18.
Ray M, Horne W, McAleer JP, Ricks DM, Kreindler JL, Fitzsimons MS, et al. RNA-seq in Pulmonary Medicine: How Much Is Enough?
Am. J. Respir. Crit. Care Med. 2015,
192, 389–391. doi:10.1164/rccm.201403-0475LE.
[Google Scholar]
-
19.
Shi Y, Liu XF, Zhuang Y, Zhang JY, Liu T, Yin Z, et al. Helicobacter pylori-induced Th17 responses modulate Th1 cell responses, benefit bacterial growth, and contribute to pathology in mice.
J. Immunol. 2010,
184, 5121–5129. doi:10.4049/jimmunol.0901115.
[Google Scholar]
-
20.
Wang CF, Yang WT, Yue LM, Qiu JY, Zhang LJ, Wang C, et al. Prominent contribution of Th1, Th17, and Tregs to the host response during M. neoaurum infection.
Genet. Mol. Res. 2016,
15. doi:10.4238/gmr.15038989.
[Google Scholar]
-
21.
Gamble LD, Purgato S, Murray J, Xiao L, Yu DM, Hanssen KMD, et al. Inhibition of polyamine synthesis and uptake reduces tumor progression and prolongs survival in mouse models of neuroblastoma.
Sci. Transl. Med. 2019,
11,
eaau1099. doi:10.1126/scitranslmed.aau1099.
[Google Scholar]
-
22.
Chen K, Eddens T, Trevejo-Nunez G, Way EE, Elsegeiny W, Ricks DM, et al. IL-17 Receptor Signaling in the Lung Epithelium Is Required for Mucosal Chemokine Gradients and Pulmonary Host Defense against K. pneumoniae.
Cell Host Microbe 2016,
20, 596–605. doi:10.1016/j.chom.2016.10.003.
[Google Scholar]
-
23.
Bayes HK, Ritchie ND, Evans TJ. Interleukin-17 Is Required for Control of Chronic Lung Infection Caused by Pseudomonas aeruginosa.
Infect. Immun. 2016,
84, 3507–3516. doi:10.1128/IAI.00717-16.
[Google Scholar]
-
24.
Ye P, Garvey PB, Zhang P, Nelson S, Bagby G, Summer WR, et al. Interleukin-17 and lung host defense against Klebsiella pneumoniae infection.
Am. J. Respir. Cell Mol. Biol. 2001,
25, 335–340. doi:10.1165/ajrcmb.25.3.4424.
[Google Scholar]
-
25.
Ye P, Rodriguez FH, Kanaly S, Stocking KL, Schurr J, Schwarzenberger P, et al. Requirement of interleukin 17 receptor signaling for lung CXC chemokine and granulocyte colony-stimulating factor expression, neutrophil recruitment, and host defense.
J. Exp. Med. 2001,
194, 519–527. doi:10.1084/jem.194.4.519.
[Google Scholar]
-
26.
Stoeckius M, Zheng S, Houck-Loomis B, Hao S, Yeung BZ, Mauck III WM, et al. Cell Hashing with barcoded antibodies enables multiplexing and doublet detection for single cell genomics.
Genome Biol. 2018,
19, 224. doi:10.1186/s13059-018-1603-1.
[Google Scholar]
-
27.
Xin H, Lian Q, Jiang Y, Luo J, Wang X, Erb C, et al. GMM-Demux: sample demultiplexing, multiplet detection, experiment planning, and novel cell-type verification in single cell sequencing.
Genome Biol. 2020,
21, 188. doi:10.1186/s13059-020-02084-2.
[Google Scholar]
-
28.
Wu C, Dai H, Yan L, Liu T, Cui C, Chen T, et al. Sulfonation of the resolving cysteine in human peroxiredoxin 1: A comprehensive analysis by mass spectrometry.
Free Radic. Biol. Med. 2017,
108, 785–792. doi:10.1016/j.freeradbiomed.2017.04.341.
[Google Scholar]
-
29.
El Eter E, Al-Masri AA. Peroxiredoxin isoforms are associated with cardiovascular risk factors in type 2 diabetes mellitus.
Braz. J. Med. Biol. Res. 2015, 48, 465–469. doi:10.1590/1414-431X20144142.
[Google Scholar]
-
30.
Nordberg J, Arner ES. Reactive oxygen species, antioxidants, and the mammalian thioredoxin system.
Free Radic. Biol. Med. 2001,
31, 1287–1312. doi:10.1016/s0891-5849(01)00724-9.
[Google Scholar]
-
31.
Mohammadi F, Soltani A, Ghahremanloo A, Javid H, Hashemy SI. The thioredoxin system and cancer therapy: a review.
Cancer Chemother. Pharmacol. 2019,
84, 925–935. doi:10.1007/s00280-019-03912-4.
[Google Scholar]
-
32.
Liu CX, Yin QQ, Zhou HC, Wu YL, Pu JX, Xia L, et al. Adenanthin targets peroxiredoxin I and II to induce differentiation of leukemic cells.
Nat. Chem. Biol. 2012,
8, 486–493. doi:10.1038/nchembio.935.
[Google Scholar]
-
33.
Siernicka M, Winiarska M, Bajor M, Firczuk M, Muchowicz A, Bobrowicz M, et al. Adenanthin, a new inhibitor of thiol-dependent antioxidant enzymes, impairs the effector functions of human natural killer cells.
Immunology 2015,
146, 173–183. doi:10.1111/imm.12494.
[Google Scholar]
-
34.
Park H, Li Z, Yang XO, Chang SH, Nurieva R, Wang YH, et al. A distinct lineage of CD4 T cells regulates tissue inflammation by producing interleukin 17.
Nat. Immunol. 2005,
6, 1133–1141. doi:10.1038/ni1261.
[Google Scholar]
-
35.
Sutton C, Brereton C, Keogh B, Mills KH, Lavelle EC. A crucial role for interleukin (IL)-1 in the induction of IL-17-producing T cells that mediate autoimmune encephalomyelitis.
J. Exp. Med. 2006,
203, 1685–1691. doi:10.1084/jem.20060285.
[Google Scholar]
-
36.
Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM, et al. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages.
Nat. Immunol. 2005,
6, 1123–1132. doi:10.1038/ni1254.
[Google Scholar]
-
37.
Ichiyama K, Yoshida H, Wakabayashi Y, Chinen T, Saeki K, Nakaya M, et al. Foxp3 inhibits RORgammat-mediated IL-17A mRNA transcription through direct interaction with RORgammat.
J. Biol. Chem. 2008,
283, 17003–17008. doi:10.1074/jbc.M801286200.
[Google Scholar]
-
38.
Zhou L, Lopes JE, Chong MM, Ivanov II, Min R, Victora GD, et al. TGF-beta-induced Foxp3 inhibits T(H)17 cell differentiation by antagonizing RORgammat function.
Nature 2008,
453, 236–240. doi:10.1038/nature06878.
[Google Scholar]
-
39.
Merentie M, Uimari A, Pietilä M, Sinervirta R, Keinänen TA, Vepsäläinen J, et al. Oxidative stress and inflammation in the pathogenesis of activated polyamine catabolism-induced acute pancreatitis.
Amino Acids 2007,
33, 323–330. doi:10.1007/s00726-007-0522-3.
[Google Scholar]
-
40.
Thomas TJ, Gunnia UB, Seibold JR, Thomas T. Defective signal-transduction pathways in T-cells from autoimmune MRL-lpr/lpr mice are associated with increased polyamine concentrations.
Biochem. J. 1995,
311, 175–182. doi:10.1042/bj3110175.
[Google Scholar]
-
41.
Brüne B, Dehne N, Grossmann N, Jung M, Namgaladze D, Schmid T, et al. Redox control of inflammation in macrophages.
Antioxid. Redox Signal. 2013,
19, 595–637. doi:10.1089/ars.2012.4785.
[Google Scholar]
-
42.
Mills EL, O’Neill LA. Reprogramming mitochondrial metabolism in macrophages as an anti-inflammatory signal.
Eur. J. Immunol. 2016,
46, 13–21. doi:10.1002/eji.201445427.
[Google Scholar]
-
43.
Tkachenko AG, Akhova AV, Shumkov MS, Nesterova LY. Polyamines reduce oxidative stress in Escherichia coli cells exposed to bactericidal antibiotics.
Res. Microbiol. 2012,
163, 83–91. doi:10.1016/j.resmic.2011.10.009.
[Google Scholar]
-
44.
Chattopadhyay MK, Tabor CW, Tabor H. Polyamines protect Escherichia coli cells from the toxic effect of oxygen.
Proc. Natl. Acad. Sci. USA 2003,
100, 2261–2265. doi:10.1073/pnas.2627990100.
[Google Scholar]
-
45.
Tkachenko AG, Fedotova MV. Dependence of protective functions of Escherichia coli polyamines on strength of stress caused by superoxide radicals.
Biochemistry 2007,
72, 109–116. doi:10.1134/s0006297907010130.
[Google Scholar]
-
46.
Min Y, Kim MJ, Lee S, Chun E, Lee KY. Inhibition of TRAF6 ubiquitin-ligase activity by PRDX1 leads to inhibition of NFKB activation and autophagy activation.
Autophagy 2018,
14, 1347–1358. doi:10.1080/15548627.2018.1474995.
[Google Scholar]