1. Introduction
Individual body condition has significant impacts on ecology, life history, mating success, and many other aspects that contribute to an organism’s fitness [
1,
2,
3,
4]. Calculating body condition indices for individuals within populations potentially enables health assessments at the individual- and population-level [
5,
6,
7]. Essentially, measures of body condition seek to isolate the aspect of body mass that is related to energy reserves, such as fats, from aspects of body mass that are related to body size [
8]. Direct measures of body condition for animals are often destructive or invasive, such as those outlined in ÓSkarsson [
9]. However, a variety of indirect (
i.e., non-destructive) methods to assess body condition have been published for various taxa [
5,
10,
11,
12,
13].
Body condition has been shown to have an impact on movement and survival in amphibian species as it is related to energy reserves [
1,
14]. Additionally, developmental stress in amphibians has also been shown to be a contributing factor to lower body condition values [
15]. Assessing body condition may also reveal the effects of multiple population stressors, such as climatic warming [
16], before they are detectable as demographic changes [
17]. Because of the potential for body condition to respond to poor conditions and influence animal health and behavior, assessing measures of population health, such as body condition, is particularly critical for informing recovery measures for endangered amphibians. The Houston toad (
Bufo [=
Anaxyrus]
houstonensis) is an exemplar of such an amphibian species, with a small geographic range and high risks of extinction.
The Houston toad’s range has been reduced from thirteen counties to nine in central and southeast Texas, USA: Austin, Bastrop, Burleson, Colorado, Lavaca, Lee, Leon, Milam and Robertson [
18,
19]. Habitat loss is a primary driver of decline and population extirpation in the species [
18,
20,
21]. For instance, populations in Bastrop County reached critically low numbers that verged on extirpation after stand-replacing wildfires impacted large portions of primary recovery habitat in 2011[
22]. Consistent with the aforementioned population decline, reproduction in the wild has been observed infrequently since then [
23]. This species is endangered, yet our understanding of the proximal causes of the decline remains ambiguous. By documenting and tracking trends in body condition, particularly for juveniles, we can evaluate the effectiveness of habitat management actions, assess the influence of annual weather (e.g., rainfall or temperature), and seek to understand how our management actions might improve the body condition of these populations. For example, potential influences of body condition on reproductive behavior (
i.e., chorus tenure) have been suggested in male
Bufo houstonensis [
24]. Gaining annual or even seasonal data on body condition can enable evaluation of habitat management (e.g., prescribed fire) and potentially link successful reproduction with improvements to body condition.
While measurements of body size and condition indices for adult life stages have been previously generated [
24,
25], such data remain very limited for juvenile
B. houstonensis. Greuter [
26] estimated survival from egg to 12 weeks post-emergence (0.04) and estimated survival to reproductive maturity at 0.003 [
27]; however, the survival of juveniles to older life stages has not been directly estimated [
27]. Gaining insights into variation in juvenile survival is a critical requirement in ensuring the long-term persistence of
B. houstonensis. Hatfield et al. [
28] used a population viability analysis to determine that an increase in the number of subpopulations and maintaining juvenile survivorship at >1% were among the most important factors in lowering extinction probabilities for
B. houstonensis. However, gathering data on juvenile health has proved difficult because of a low underlying abundance that has resulted in fewer juvenile observations. Additionally, juvenile detection is complicated by the lack of ecological knowledge of the juvenile life stage, members of which are only present during a narrow daily temporal window [
26]. Because juvenile
B. houstonensis are so poorly understood and difficult to detect, understanding the ecology of this life stage presents a particular challenge to species recovery efforts.
Management actions are being implemented on the Griffith League Scout Ranch (hereafter “the GLR” or just “GLR”) to increase the number of Houston toads within critical habitat. These recovery actions primarily include habitat restoration and population supplementation [
29]. Population supplementation has sought to boost juvenile survivorship through the addition of captively propagated egg strands and tadpoles at recovery sites. The Houston Zoo, in collaboration with Texas State University, Texas Parks and Wildlife Department (TPWD), and the U.S. Fish and Wildlife Service (USFWS), began supplementing
B. houstonensis populations on the GLR in 2007. Expansion of supplementation efforts of
B. houstonensis took place from 2014 to 2021, with the added involvement of both the Fort Worth and Dallas Zoos alongside the San Marcos Aquatic Resources Center. The number of chorusing
B. houstonensis at the primary recovery site has increased annually [
23,
30]. Supplementation and concomitant habitat restoration efforts as well as an increase in rainfall that has lifted drought conditions, may be likely causes of an observed increase of approximately 283
B. houstonensis detections per year [
31]. This increase in the size of breeding aggregates of
B. houstonensis has also enabled an increase in the number of emergent juvenile post-metamorphic toads (or toadlets).
Juvenile emergence remains cohort-based, with all of a given group of tadpoles completing metamorphosis and dispersing within a day or a few days. Moreover, once terrestrial, the individuals are only observed for a few hours just after sunrise on the subsequent days. The collection of individual morphological data on juveniles is further limited by the ability to predict the timing of emergence, which tends to happen in a short window of days. This presents the need to reduce time spent on handling and rapidly collect data for a statistically reasonable number of individual juveniles. Handling these very small individuals likely induces stress [
32], and the traditional caliper-based method of obtaining body measurements can be cumbersome and time consuming. A reduction in measurement time would allow for an increase in the number of toadlets processed while potentially mitigating handling stress applied to individuals [
32].
Historically, cameras have served a wide array of purposes within wildlife research. Camera-based measurement systems have been employed to calculate basic body measurements and construct body condition indices (BCI) in multiple taxa [
33,
34,
35,
36,
37]. Previous studies of population health in mammals [
33], reptiles [
35,
38], amphibians [
39] and fish [
40] have demonstrated that image-based measurements are a viable substitution for manually acquired measurements (
i.e., from calipers) across taxa [
34,
38]. It has been shown that image-based measurement techniques were adequate to obtain body length and girth [
33], snout-hindlimb length and snout-vent length in Western fence lizards (
Sceloporus occidentalis) [
38], total body length in brown tree snakes (
Boiga irregularis) [
35], and estimations of body mass from image-derived measurements of dorsal body area in tadpoles [
39]. A similar study was conducted with salamanders using the segmented line tool in ImageJ to account for curvature in the body, and no significant differences were observed between image-based and caliper-based measurements [
40]. Furthermore, Mott, Albert, Steffen and Uzzardo [
37] found that using photos to obtain measurements has a 38% to 46% reduction in handling time for
Ambystoma salamanders. In addition to the proposed benefits to juveniles, extending image-based measurements to adults represents potential reductions in handling time for breeding individuals. It would minimize disturbances to reproduction while likely enabling data on individual health through condition indices.
Here, we compare the utility of image-based and manual methods of obtaining body measurements in adult and juvenile life stages of
B. houstonensis to estimate body condition indices. We aim to determine whether there is a significant difference in measurements from either approach while comparing the handling time associated with each method. We sought to derive and test for differences in condition indices for each method of measurement to assess their efficacy in determining individual health. Furthermore, we secondarily sought to compare the accuracy of two commonly used condition indices, SMI and OLS, in assessing health for
B. houstonensis.
2. Materials and Methods
2.1. Study Site and Collection of Individual Toads
We sampled adults at the primary recovery site within critical habitat designated for
B. houstonensis [
22]. This recovery site, named the Griffith League Ranch (GLR), is a 1948-hectare ranch in Bastrop, Texas, owned by the Boy Scouts of America (Austin, TX, USA). GLR has an overstory that primarily consists of loblolly pine (
Pinus taeda), oaks (
Quercus sp.), and red cedar (
Juniperus virginiana). The understory consists of American beautyberry (
Callicarpa americana), yaupon holly (
Ilex vomitoria), and farkleberry (
Vaccinium arboretum). The substrate is primarily deep sandy soil, and anuran breeding is observed at permanent ponds (
n = 3), semipermanent ponds (
n = 10), and ephemeral ponds (
n > 10).
We partnered with Texas zoos that maintain a captive breeding population of
B. houstonensis intended to supplement wild breeding events within the primary recovery habitat through the release of egg strands, tadpoles, and juveniles. Toadlets hatched at the Fort Worth Zoo (Fort Worth, TX, USA) were transported for release at the Griffith League Ranch shortly after emergence in 2023. These toadlets were subjected to both the caliper-based and image-based methods and subsequently released within the understory adjacent to a permanent pond on the GLR. We conducted nocturnal, audio surveys of ponds on the GLR during the peak breeding season (
i.e., January to May) of
B. houstonensis in 2024. We surveyed ponds for a 5-minute interval during suitable weather conditions and subsequently exhaustively searched positive chorusing sites for breeding
B. houstonensis. Adult toads were captured by hand and identified as
B. houstonensis based on the presence of elongated bean-shaped parotoid glands, dark colored vocal sac in males, speckled ventral surface, and an “L” shape ridge in between the cranial crests [
41]. These characters immediately distinguish this species from the only co-occurring congener,
Bufo (=
Incilius)
nebulifer. Data was then collected from the captured individuals, after which they were released close to the site of capture. Here, we present data from male
B. houstonensis from which we collected measurements via caliper- and image-based methods. We only included male toads in our assessment of body condition since reproductive females may have contained egg masses at the time of capture, which would likely bias our determination of body condition.
2.2. Caliper-Based and Image-Based Measurement
A total of 37 adult and 94 juvenile
Bufo houstonensis individuals were subjected to both measurement trials. We measured Snout-to-Urostyle length (SUL in mm) and Head Width (HW in mm) of toads with calipers to the nearest millimeter. We photographed the same individuals using a digital smartphone camera (iPhone 14 Plus, Apple Inc., Cupertino, CA, USA) on top of either a 7 mm or 10 mm grid sheet for scale (
). We oriented toads on the grid sheet with their dorsal side facing upwards to potentially reduce handling time, as that is their natural stance. For juveniles, we recorded the start and stop time of physical handling per-individual (
i.e., from the moment of first contact for measurement to completion for the individual) for both caliper-based and image-based measurements using a smartphone stopwatch app. In the caliper trial, a second researcher was present to record the measurements taken. Adults and juveniles were weighed (in g) using a digital scale. Individual identification/collector numbers were assigned to each toad and recorded with the corresponding images or measurements to enable individual identification of all toads in later analyses. In adults, the longest toe on the right hind limb was partially clipped to enable DNA and disease testing and provide cohort-level identification of individuals to prevent repeated measures in the event of recapture.
. Image-based measurements of Houston toads (<i>Bufo</i> [=<i>Anaxyrus</i>] <i>houstonensis</i>) were used to assess individual health in 37 adults and 94 juveniles. Adults (<b>A</b>) and juveniles (<b>B</b>) were positioned on either a 10 mm (<b>A</b>) or 7 mm (<b>B</b>) grid sheet for scale and then photographed using a digital smartphone camera (iPhone 14 Plus, Apple Inc.). We assigned collector numbers to each toad to enable individual identification. We obtained measurements from images of juveniles and adults using the straight line tool in ImageJ.
We processed images of juvenile and adult
B. houstonensis in ImageJ [
42]. The straight line tool of the software was used to extract measures of SUL and HW by applying measurement vectors from that tool to the same points that they would have been measured using calipers. The scale was calibrated, for each image, in an mm/pixel format using grid squares in which the toads were photographed. We repeated the calibration step for each image to account for variation in camera position among photographs. For both caliper-based and image-based measurements in each life stage, observer bias was mitigated by ensuring the same observer (SS) collected both sets of measurements.
2.3. Statistical Analyses
We conducted all statistical analyses in R, version 4.3.0 [
43]. Measurement data were grouped by life stage, and statistical differences in SUL and HW were assessed by method of measurement within each life stage. We applied linear mixed effects models in the package lme4 [
44] to account for individual heterogeneity and non-independence of repeated measures, by including random variation in intercepts among individual IDs/collector numbers. For toadlets, differences in handling time were assessed by method using linear mixed effects models in the package lme4 [
44], while enabling random variation in intercepts among individual IDs/collector numbers. We estimated the significance of fixed factors in our linear mixed effects regressions via the package lmerTest [
45]. Analyses of handling time are only presented for toadlets as handling time for adults was not recorded.
2.4. Scaled Mass Index
We subsequently sought to assess differences in body condition using different measurement methods. For this reason, we grouped measurement data within each life stage by method of measurement (
i.e., adults measured by calipers, adults measured from images, juveniles measured by calipers, and juveniles measured from images). We computed Scaled Mass Indices (SMI) per the method outlined by Peig and Green [
13]. We first quantified the scaling exponent
b from the function $$M a s s = a l e n g t h^{b}$$ and then calculated scaled body masses for a given length of studied individuals. We performed standardized major axis (SMA) regressions of lnMass on lnSUL in package smatr [
46]. SMI was calculated for the SUL corresponding to the average SUL of all toads within a treatment group. Condition indices calculated within a life stage were then compared among methods of measurement using linear mixed effects regressions, and subsequently compared between life stages, while including random variation in intercepts among individual IDs/collector numbers [
44,
45]. Furthermore, we performed correlation tests between values of SMI and SUL within juvenile and adult life stages.
2.5. Residual Body Condition Indices
We also employed the “residuals’ method to express body condition [
10]. In this method, an ordinary least squares regression of lnMass on lnSUL is used to establish the theoretical body weight of an individual of a given length [
47]. Subsequently, this predicted body weight of individuals of a given length is subtracted from the body weight measured for that individual. Individuals with positive residuals are considered to present good body condition, while individuals with negative residual values likely have low energy reserves, though it could be due to other factors such as desiccation. We computed body condition by method of measurement within each life stage resulting in four test groups, as above. We used linear mixed effects regressions to test for statistical differences in residual condition indices by method within each life stage, while including random variation of intercepts among individual IDs/collector numbers. Similarly, we compared residual condition indices among life stages via linear mixed effects models, while accounting for random variation in intercepts among individual IDs/collector numbers [
44,
45]. We performed correlation tests between values of residual condition indices and SUL within juvenile and adult life stages.
3. Results
3.1. Comparisons of HW and SUL for Juvenile and Adult Toads
Juvenile results are from 94 zoo-sourced juveniles. Adult results are from 37 male
B. houstonensis captured over three nights between February to April 2024. We saw no difference in measurements of SUL generated via manual and image-based measurements in juveniles ($$\beta$$ = 0.089, SE = 0.133, $$t_{93}$$ = 0.669,
p = 0.505) or in adults ($$\beta$$ = 0.460, SE = 0.409, $$t_{36}$$ = 1.122,
p = 0.269). There was more variation among individual juveniles (SD = 2.577) relative to the juvenile residual standard deviation (SD = 0.913) and among individual adults (SD = 3.274) relative to the adult residual standard deviation (SD = 1.761). Similarly, we saw no difference in measurements of head width generated via manual and image-based measurements in juveniles ($$\beta$$ = −0.042, SE = 0.076, $$t_{93}$$ = −0.548,
p = 0.585) or in adults ($$\beta$$ = −0.149, SE = 0.285, $$t_{36}$$ = −0.521,
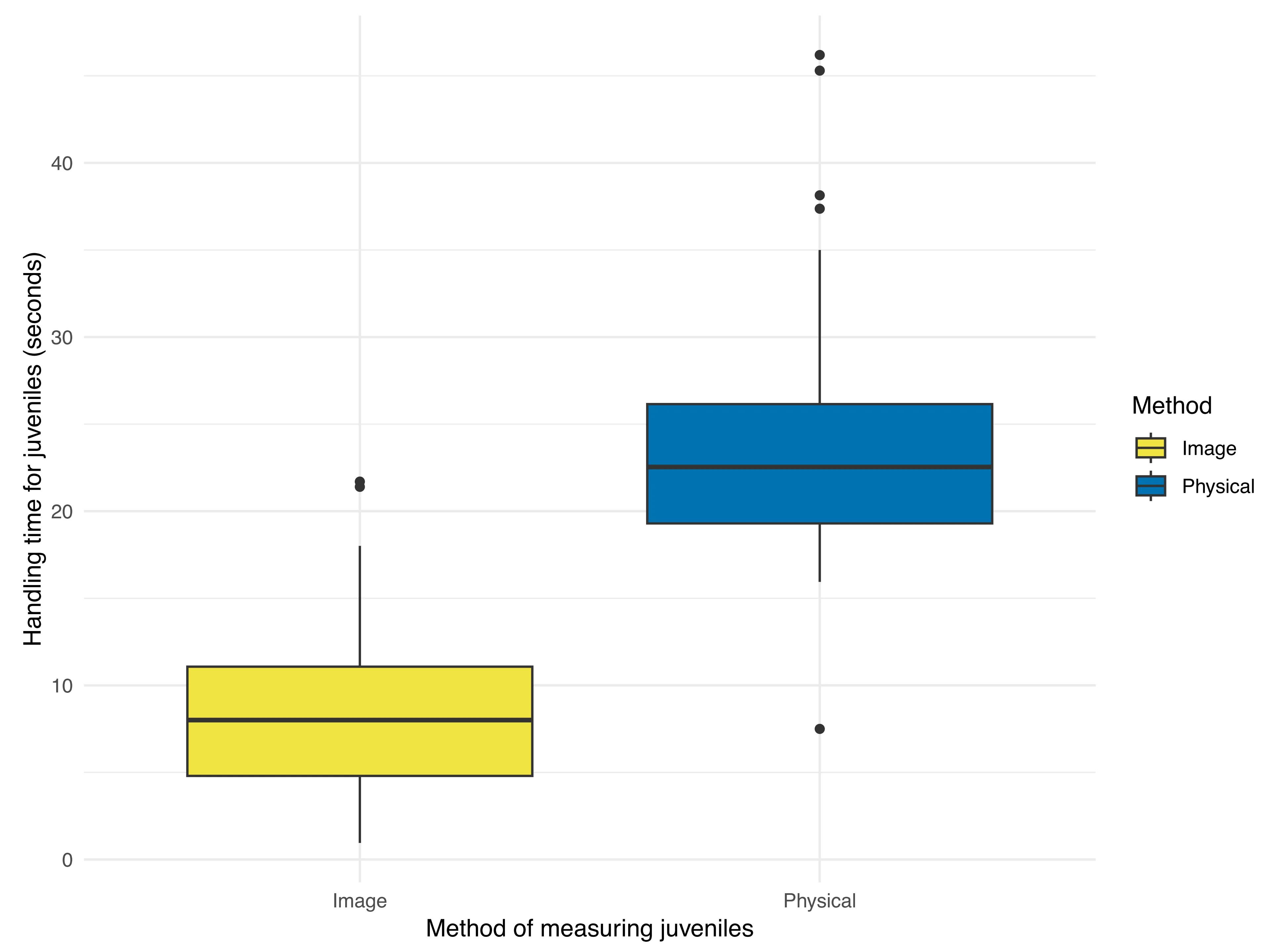
p = 0.606). More variation was present among individual juveniles (SD = 1.066) relative to the residual standard deviation (SD = 0.519). Conversely, we observed less variation among individual adult toads (SD = 0.964) relative to the residual standard deviation (SD = 1.227). We observed a significant difference in the duration for which toadlets were handled based on the method of measurement ($$\beta$$ = 15.056, SE = 0.730, $$t_{93}$$ = 20.640,
p < 0.0001). Handling time for toadlets via image-based measurements (8.282 ± 4.724 seconds;
) was significantly lower than handling time for caliper-based measurements (23.338 ± 6.141 seconds;
). Less variation was observed among individual juveniles (SD = 2.237) relative to the residual standard deviation (SD = 5.001).
. Box plot representing handling time in seconds to obtain caliper-based (physical) measurements and images of juvenile Houston toads (<i>Bufo</i> [=<i>Anaxyrus</i>] <i>houstonensis</i>). We observed that calliper-based measurements required a significantly longer duration of handling relative to the duration required to obtain images.
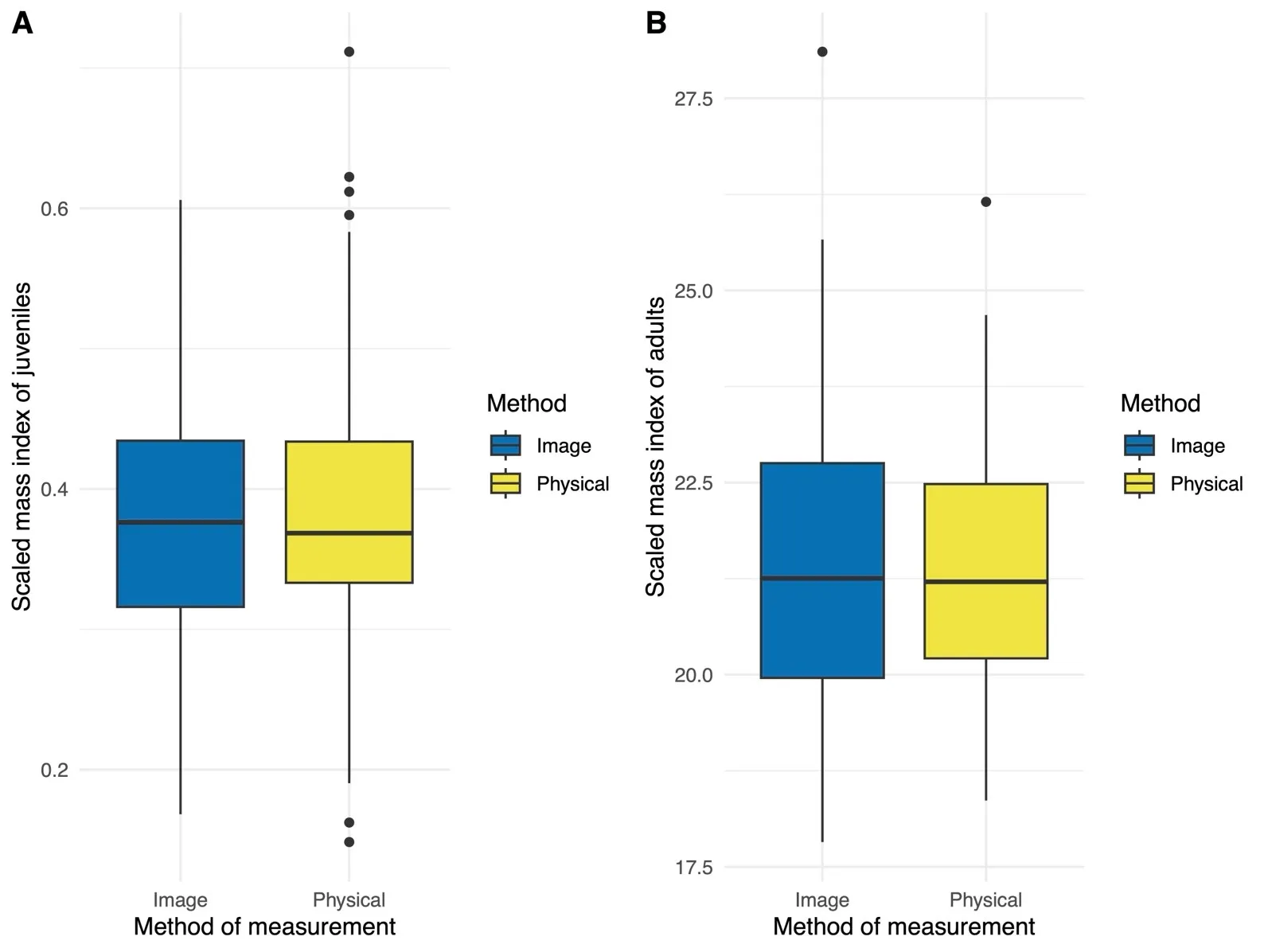
We observed no significant difference in SMI values by method of measurement within juvenile toadlets ($$\beta$$ = 0.002, SE = 0.011, $$t_{93}$$ = 0.208,
p = 0.836;
A). Less variation was observed among individual juveniles (SD = 0.055) relative to the residual standard deviation (SD = 0.073). A strongly negative correlation was present between SMI values and body length ($$r_{186}$$= −0.235,
p = 0.001;
Figure S1) in juvenile toads.
. Box plot representing scaled mass indices (SMI) obtained across image- and caliper-based (physical) methods in juvenile (<b>A</b>) and adult (<b>B</b>) Houston toads (<i>Bufo</i> [=<i>Anaxyrus</i>] <i>houstonensis</i>). We did not observe a significant difference in condition indices among our methods of measurement for either life stage.
We observed no significant difference in SMI values by method of measurement within adult
B. houstonensis ($$\beta$$ = −0.076, SE = 0.348, $$t_{36}$$ = −0.219,
p = 0.828;
B). Less variation was observed among individual adults (SD = 1.42) relative to the residual standard deviation (SD = 1.50). A strongly negative correlation was present between SMI values and body length in ($$r_{72}$$= −0.317,
p = 0.006;
Figure S2) adult toads.
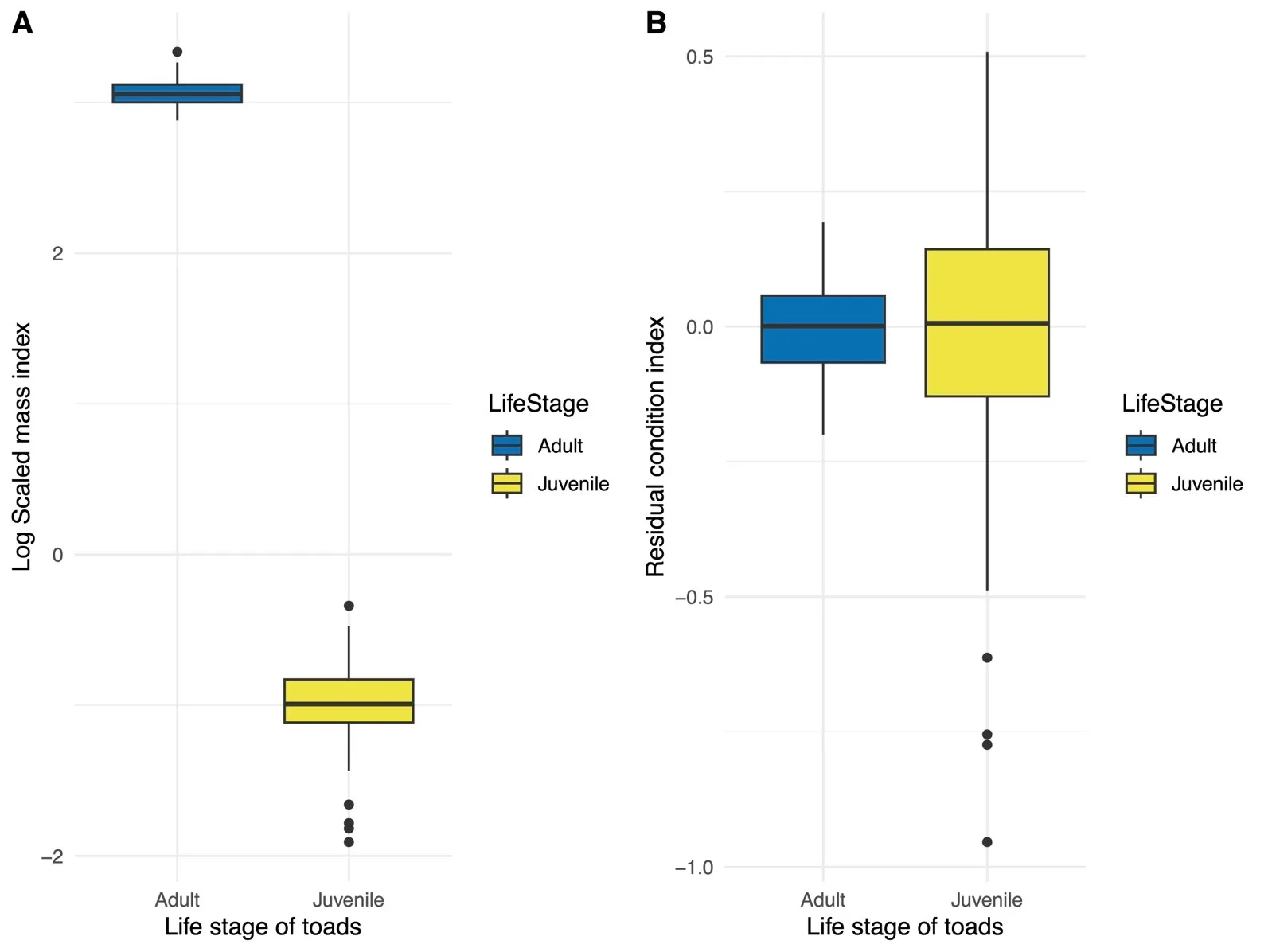
SMI values varied significantly by life stage ($$\beta$$ = −21.079, SE = 0.182, $$t_{128 . 9}$$ = −115.600,
p < 0.0001;
A). SMI values were significantly larger for adults (21.463 ± 2.051) relative to juvenile
B. houstonensis (0.385 ± 0.090). We observed less variation among individuals (SD = 0.757) relative to the residual standard deviation (SD = 0.787).
. Box plot representing a comparison of log transformed scaled mass indices (SMI; (<b>A</b>)) and residual condition indices (<b>B</b>) among juvenile and adult Houston toads (<i>Bufo</i> [=<i>Anaxyrus</i>] <i>houstonensis</i>). These indices include measurements from both image- and caliper-based trials. We observed SMI values were significantly higher in adults relative to juveniles, while residual condition indices did not differ significantly among life stages. For this reason, residual condition indices appear to be a measure of individual health that is independent of body size.
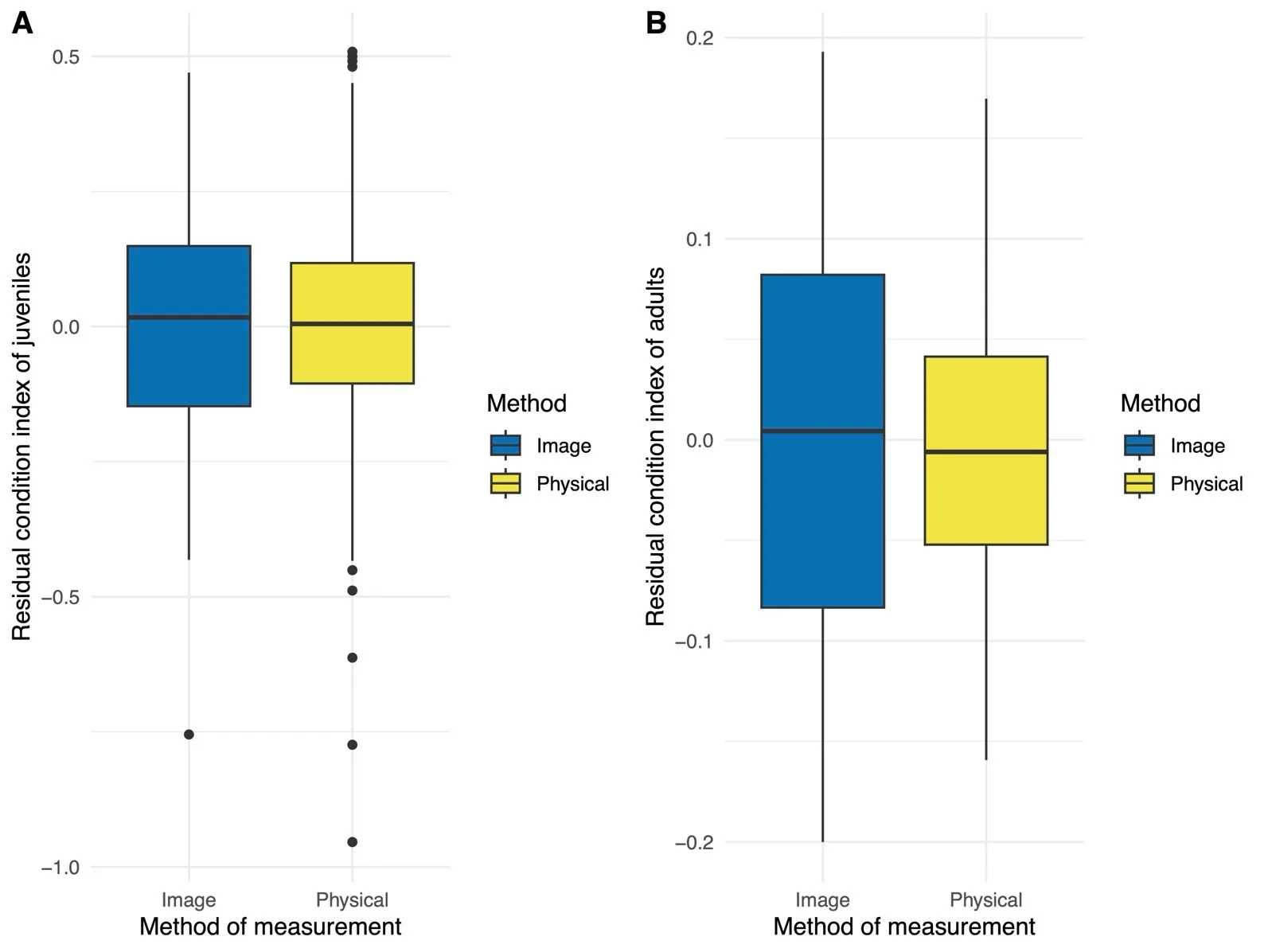
We observed no significant difference in residual condition indices by method of measurement within juvenile toadlets ($$\beta$$ = 0.289 × 10
−16, SE = 0.023, $$t_{93}$$ < 0.0001,
p > 0.999;
A). More variation was observed among individual juveniles (SD = 0.174) relative to the residual standard deviation (SD = 0.160). We observed virtually no correlation between residual condition indices and body length ($$r_{186}$$= 0.003,
p = 0.968;
Figure S3) in juvenile toads.
. Box plot representing residual condition indices obtained across image- and caliper-based (physical) methods in juvenile (<b>A</b>) and adult (<b>B</b>) Houston toads (<i>Bufo</i> [=<i>Anaxyrus</i>] <i>houstonensis</i> ). We did not observe a significant difference in residual condition indices among our methods of measurement for either life stage.
We observed no significant difference in residual condition indices by method of measurement within adult
B. houstonensis ($$\beta$$ = 0.3376 × 10
−17, SE = 0.013, $$t_{36}$$ < 0.0001,
p > 0.999;
B). More variation was observed among individual adults (SD = 0.068) relative to the residual standard deviation (SD = 0.058). There existed virtually no correlation between residual condition indices and body length in ($$r_{72}$$= 0.005,
p = 0.967;
Figure S4) adult toads.
We found no statistical difference in residual condition indices among life stages ($$\beta$$ = 0.147 × 10
−16, SE = 0.035, $$t_{129}$$ < 0.0001,
p > 0.999;
B). Residual condition indices for adults (
−0.867 × 10
−18 ± 0.089) and juvenile
B. houstonensis (−0.176 × 10
−16 ± 0.236) were similar. We observed more variation among individuals (SD = 0.153) relative to the residual standard deviation (SD = 0.140).
4. Discussion
We compared measurements of 94 juvenile toads that were obtained via caliper-based and image-based methods while also assessing the duration that we handled juveniles for each method of measurement. We obtained paired measurements from each method for 37 adult
B. houstonensis. Within each life stage, we found no significant difference between measurements of snout urostyle length and head width between image-based and caliper-based methods. We found that the handling time of juvenile toads was significantly lower in the image-based measurement trials than in the caliper-based measurement trials. Additionally, we found no difference in either SMI or residual condition indices between methods, for each life stage. Thus, image-based measurements offer a viable alternative to caliper-based measurements in assessing the health of juvenile
B. houstonensis. This image-based method also enables processing a statistically reasonable number of individuals within a short span of time, which potentially facilitates health assessments for a life stage that has a narrow daily and seasonal window of detection.
Image-based measurements may also be less susceptible to some of the limitations of caliper-based measurements. Arismendi, Bury, Zatkos, Snyder and Lindley [
40] found that digital camera images of Coastal cutthroat trout (
Oncorhynchus clarkii clarkii) and Coastal giant salamanders (
Dicamptodon tenebrosus) in groups of up to 20 individuals were adequate to gain accurate measurements of SVL and total body length of aquatic vertebrates. They sedated study subjects to promote a linear position when taking manual measurements and used the segmented line tool in ImageJ to account for movement and contortions in the Image-based measurements, and found no significant difference in the measurements taken between methods. While Arismendi, Bury, Zatkos, Snyder and Lindley [
40] did not encounter measurement error derived from spine curvature in aquatic vertebrates, Mott, Albert, Steffen, and Uzzardo [
37] found that there was a significant difference in measurements of body lengths between caliper-based and image-based methods when measuring
Ambystoma salamanders. They hypothesized that the snout-to-vent lengths derived from images were likely more accurate than those from calipers as they used the “bent line” tool in ImageJ to account for curvature in adults. Because salamanders do not maintain a linear body position under the stress of handling, and calipers are intended to calculate linear measurements, the calipers were suggested to be less accurate. The findings from Arismendi, Bury, Zatkos, Snyder and Lindley [
40] corroborate this suggestion, as they used a sedative to aid in physical measurements of larval salamanders to ensure that the subjects remained linear during measurement and found no significant differences between the image-based and manual body measurements.
We found no literature on post-metamorphic anurans being measured digitally. Because of this, we reference salamanders heavily in our literature review, but are cognizant of the differences in body plan. We likely did not encounter the same extent of body contortions in
B. houstonensis, as their body plan is less facilitative to spine curvature than that of a caudate. This enabled the use of the straight line tool in ImageJ for SUL measurements. We acknowledge that there is still a possibility for position-related error in measurement, such as an individual not being completely flat on the grid sheet. We took measures to minimize this error by ensuring all toads were lying flat and still before taking the photo. However, we did observe greater variation among individuals relative to the residual standard deviation when comparing SUL among methods of measurement using the package lme4, meaning that there is significant variation in SUL within both life stages. We speculate that varying levels of body contortions among individuals resulted in the observed individual-level heterogeneity.
We believe that this image measurement technique has the potential to serve as a valuable method of gathering body condition data more efficiently without sacrificing measurement accuracy. The decrease in handling time potentially reduces handling stress [
32]. The 15 s saved per individual would compound quickly, allowing for a higher volume of juveniles to be sampled during the narrow temporal window when they are present. This would help address a crucial knowledge gap within juvenile ecology that has previously limited the recovery of
B. houstonensis. We have observed an increase in the number of detections of
B. houstonensis on the landscape, which is likely correlated with recovery measures such as population supplementation and habitat restoration, as well as improvements in drought conditions [
31]. Maximizing the number of juveniles we can measure within the narrow window of their emergence is crucial to enable inferences on ecology and life history. Specifically, this could facilitate an understanding of the influence of environmental factors on individual health and the consequences of variation in individual health on juvenile dispersal and survival. This, paired with more efficient data collection, would enable more informed management plans for the juvenile life stage of
B. houstonensis. This difference in handling time could also be particularly advantageous to assessments of B. houstonensis adults, given that large (>100 individuals) breeding choruses of
B. houstonensis have been observed over recent years [
30,
31]. Adult handling time was not recorded for this study, as the detection of adult toads is not limited in the same temporal manner as juvenile detection. Therefore, decreasing handling time is not as critical for adults as it is for juveniles. As such, changes in adult handling time are based on assumptions extrapolated from the effects on juvenile handling time. Based on average handling times specific to each method that we estimated with juveniles, the total handling time for 10 adult
B. houstonensis would stand to be reduced from 3.9 minutes using a manual (
i.e., caliper) method of measurement to a little over 1.38 minutes using an image-based approach. This would minimize the duration of handling and thus mitigate stress at the individual level [
32] and the duration for which adults are removed from the reproductive bout. Applying this image-based approach of measurements to adults would therefore minimize disruption of breeding choruses while also enabling health assessments for reproductively active
B. houstonensis.
Along with the reduction in handling time, the image-based technique only requires one researcher, while caliper measurements require two (one to measure the animal and the other to record those measurements). This allows us to gain measurements efficiently using half the personnel. We note, however, that the time to record measurements did not factor into the handling time in the caliper-trial as it was done simultaneously by a second researcher. As such, the difference between the average handling time of the two measurement trials would likely be more drastic if only one individual were obtaining and recording caliper measurements. When using the image-based approach, the handling time only accounts for the time required to obtain the photo. It does not include the time spent to estimate measurements via ImageJ from scaled images. Thus, there is a possibility that using mm/pixel software to deduce body measurements may increase the overall investigator effort required to obtain body measurements for each individual. This trade-off is worthwhile as it provides the potential to mitigate stress and disruption to natural behavior as a result of decreasing individual handling time. Just as importantly, the time saved in the field maximizes the time available for additional individuals to be assessed during each sampling event.
We computed SMI and residual condition indices to express body condition in juvenile and adult
B. houstonensis. Scaled mass indices have found increased application as a measure of individual health in amphibians [
47,
48,
49]. However, condition factors must be independent of body size and remove the effect of ontogenetic growth on the weight-length relationship [
5,
47]. To enable meaningful comparison among individuals of different size- and age-classes, condition indices should be uncorrelated with body size [
5]. However, in our use of
bSMA (the scaling factor for SMI calculations) to calculate SMI [
5], we observed a significant negative correlation between SMI and SUL within each life-stage. This suggests that our SMI values were not independent of body size, thus violating a key assumption for a valid condition index. Furthermore, comparisons among life-stages showed that adults had a significantly higher SMI relative to juvenile
B. houstonensis. For this reason, we suggest that the indirectly calculated
bSMA does not provide an estimate of SMI that is independent of the structural size of Houston toads.
Other literature that expresses body condition and related measures of health to environmental factors has shown that
bSMA is non-independent of body size. For example, Zhelev and Minchev [
48] also observed a negative but non-significant (
p = 0.054) correlation between SMI values and body size in marsh frogs (
Pelophylax ridibundus). Brodeur, Damonte, Vera Candioti, Poliserpi, D'Andrea, and Bahl [
47] also show that an indirectly calculated
bSMA was significantly negatively correlated with body size in South American common frogs (
Leptodactylus latrans) and instead recommended using a scaling exponent from a non-linear regression, which yielded size-independent SMI estimates. To resolve this issue, we instead used condition indices expressed as residual values from OLS regressions. The OLS residual values satisfied the requirement of independence from body size within each life-stage and showed no difference among juvenile and adult life-stages, suggesting that residuals from an OLS regression may be more appropriate than those from an SMI in constructing body condition indices in the Houston toad. Brodeur, Damonte, Vera Candioti, Poliserpi, D’Andrea, and Bahl [
47] also corroborate this finding in their study of
L. latrans but maintained that residual condition indices were a much more variable parameter than SMI, possibly since the former provide no scaling of the obtained data [
48]. Furthermore, Estrada, Medina, Gratwicke, Ibanez, and Belden [
49] used SMI values to express changes in body condition, skin microbiome composition, and disease status of captive-born Limosa harlequin frogs (
Atelopus limosus) after release in the wild and in relation to wild conspecifics. However, to our knowledge, no tests of independence of SMI from body size were performed in this study. We reiterate Peig and Green’s [
13] statement of caution that no single method of estimating body condition is ideal and that users of condition indices must adopt a ‘healthy skepticism’ while attempting to validate the assumptions of estimated condition indices.
5. Conclusions
Our aim was to assess a digital measurement technique as an alternative to physical measurement techniques to obtain linear body measurements of B. houstonensis and apply those measurements to body condition indices. We found no significant difference between measurements of body size, irrespective of method, for juvenile or adult life stages. We also found that juveniles measured with image-based techniques had significantly lower handling time than those measured with physical techniques. Image-based measurements, therefore, show strong potential to minimize stress and handling time of juveniles while maximizing the number of individuals sampled during the narrow window of their emergence and dispersal, facilitating data collection on the health of a statistically reasonable number of juveniles. Extending image-based methods of measurements to adult B. houstonensis would similarly mitigate stress and disruption of reproductive behavior while allowing assessments of condition in reproductively active individuals. We found no difference in condition indices estimated from digital and physical methods of measurement. This is a crucial advantage to assessing health for a poorly documented life-stage of an endangered amphibian species, which would enable subsequent recovery actions that can optimize juvenile survival and potentially benefit in reducing extinction risks for B. houstonensis.
In our comparison of body condition indices, we suggest caution in using scaled mass index values that are indirectly calculated from scaling exponents of standardized major axis regressions. We observed that SMI values generated in this manner were not independent of body size, which is a fundamental violation of a condition index. Our results differ from the proposed advantage that Scaled Mass Index is less correlated with individual body size. SMI is mathematically outlined to be used in controlling for growth effects in linear measures of body size that may not be a direct measure, such as head width or beak length. It could be that our use of SUL, which is a more encompassing measure of body size that tends to correlate well with overall mass, caused an overcorrection via the scaling factor. Another possibility is that the relationship between the increase in length and increase in weight in Anurans is not adequately represented by the scaling factor used in the scaled mass index. The use of OLS residuals resolved this issue in B. houstonensis, though we cannot assert that it is a superior method without further testing. We can only conclude that it was the more appropriate method for our measurement data due to the lack of correlation between the residual value and body measurements. Further investigation would be needed to deduce the causality of the correlation in SMI. We encourage users of condition indices to test violations of independence prior to an assumption that such measures reflect ‘true condition’ in species of interest.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/583,
Figure S1: Plot representing correlation of scaled mass index (SMI) values and snout urostyle length (SUL) in juvenile Houston toads (
Bufo [=
Anaxyrus]
houstonensis);
Figure S2: Plot representing correlation of scaled mass index (SMI) values and snout urostyle length (SUL) in adult Houston toads (
Bufo [=
Anaxyrus]
houstonensis );
Figure S3: Plot representing correlation of residual condition indices and snout urostyle length (SUL) in juvenile Houston toads (
Bufo [=
Anaxyrus]
houstonensis);
Figure S4: Plot representing correlation of residual condition indices and snout urostyle length (SUL) in adult Houston toads (
Bufo [=
Anaxyrus]
houstonensis.
Acknowledgments
We thank the United States Fish and Wildlife Service, Texas Department of Transportation, Texas State University, Houston Zoo Inc., Fort Worth Zoo, and Dallas Zoo for their collaborative efforts in enabling recovery measures for Houston toads. We would also like to thank Ferris Zughaiyir, Jim Bell, and DJ Stout for their assistance with and facilitation of field work. Finally, we thank Saunders Drukker for analytical tips.
Author Contributions
Conceptualization, M.R.J.F., S.S., and J.M, Data Collection, S.S. and J.M., Data Analysis, S.S. and A.F., Data Curation, S.S. and J.M., Writing—Original Draft Preparation, A.F., Writing—Review & Editing, A.F., S.S., M.R.J.F., and J.M., Visualization, S.S., Supervision, M.R.J.F., Project Administration, M.R.J.F., Funding Acquisition, M.R.J.F. All authors have read and agreed to publish the manuscript.
Ethics Statement
Sample collection from toads was authorized under a USFWS permit (TE039544-0) and a TPWD scientific research permit (SPR-0102-191). Our experimental protocol was approved on 10 September 2024 by the Texas State University Institutional Animal Care and Use Committee (Approval number: 35141129; Protocol number: 9698). We adhered to the Animals in Research: Reporting On Wildlife (ARROW) guidelines by taking all measures to mitigate stress.
Informed Consent Statement
Not Applicable.
Data Availability Statement
R scripts used in analyses will be made available by the corresponding author upon reasonable request.
Funding
This research was funded by United States Fish and Wildlife Service as well as Texas Department of Transportation.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Lowe WH. Linking Dispersal to Local Population Dynamics: A Case Study Using a Headwater Salamander System.
Ecology 2003,
84, 2145–2154. doi:10.1890/0012-9658(2003)084[2145:Ldtlpd]2.0.Co;2.
[Google Scholar]
-
2.
Guy Morrison RI, Davidson NC, Wilson JR. Survival of the fattest: body stores on migration and survival in red knots
Calidris canutus islandica.
J. Avian Biol. 2007,
38, 479–487. doi:10.1111/j.0908-8857.2007.03934.x.
[Google Scholar]
-
3.
Scott DE, Casey ED, Donovan MF, Lynch TK. Amphibian lipid levels at metamorphosis correlate to post-metamorphic terrestrial survival.
Oecologia 2007,
153, 521–532. doi:10.1007/s00442-007-0755-6.
[Google Scholar]
-
4.
Tsvey A, Bulyuk VN, Kosarev V. Influence of body condition and weather on departures of first-year European robins,
Erithacus rubecula, from an autumn migratory stopover site.
Behav. Ecol. Sociobiol. 2007,
61, 1665–1674. doi:10.1007/s00265-007-0397-z.
[Google Scholar]
-
5.
Peig J, Green AJ. The paradigm of body condition: a critical reappraisal of current methods based on mass and length.
Funct. Ecol. 2010,
24, 1323–1332. doi:10.1111/j.1365-2435.2010.01751.x.
[Google Scholar]
-
6.
Mckinney MA, Atwood T, Dietz R, Sonne C, Iverson SJ, Peacock E. Validation of adipose lipid content as a body condition index for polar bears.
Ecol. Evol. 2014,
4, 516–527.
[Google Scholar]
-
7.
Stevenson RD, Woods WA. Condition indices for conservation: new uses for evolving tools.
Integr. Comp. Biol. 2006,
46, 1169–1190.
[Google Scholar]
-
8.
Green AJ. Mass/Length Residuals: Measures of Body Condition or Generators of Spurious Results?
Ecology 2001,
82, 1473–1483. doi:10.1890/0012-9658(2001)082[1473:Mlrmob]2.0.Co;2.
[Google Scholar]
-
9.
ÓSkarsson GJ. Variation in body condition, fat content and growth rate of Icelandic summer‐spawning herring
Clupea harengus L.
J. Fish Biol. 2008,
72, 2655–2676. doi:10.1111/j.1095-8649.2008.01886.x.
[Google Scholar]
-
10.
Bancila RA, Hartel T, Plăiaşu R, Smets J, Cogălniceanu D. Comparing three body condition indices in amphibians: a case study of yellow-bellied toad
Bombina variegata.
Amphibia-Reptilia 2010,
31, 558–562. doi:10.1163/017353710X518405Short.
[Google Scholar]
-
11.
Jakob EM, Marshall SD, Uetz GW. Estimating Fitness: A Comparison of Body Condition Indices.
Oikos 1996,
77, 61–67.
[Google Scholar]
-
12.
Falk BG, Snow RW, Reed RN. A validation of 11 body-condition indices in a giant snake species that exhibits positive allometry.
PLoS One 2017,
12, e0180791. doi:10.1371/journal.pone.0180791.
[Google Scholar]
-
13.
Peig J, Green AJ. New perspectives for estimating body condition from mass/length data: the scaled mass index as an alternative method.
Oikos 2009,
118, 1883–1891. doi:10.1111/j.1600-0706.2009.17643.x.
[Google Scholar]
-
14.
Lowe WH, Nislow KH, Likens GE. Forest structure and stream salamander diets: Implications for terrestrial-aquatic connectivity.
SIL Proceedings, 1922-2010 2005,
29, 279–286. doi:10.1080/03680770.2005.11902014.
[Google Scholar]
-
15.
Davis TM, Maerz JC. Spot symmetry predicts body condition in spotted salamanders,
Ambystoma maculatum.
Appl. Herpetol. 2007,
4, 195–205. doi:10.1007/978-3-540-34533-6_8.
[Google Scholar]
-
16.
Moldowan PD, Tattersall GJ, Rollinson N. Climate-associated decline of body condition in a fossorial salamander.
Glob. Chang Biol. 2022,
28, 1725–1739. doi:10.1111/gcb.15766.
[Google Scholar]
-
17.
MacCracken JG, Stebbings JL. Test of a Body Condition Index with Amphibians.
J. Herpetol. 2012,
46, 346–350. doi:10.1670/10-292.
[Google Scholar]
-
18.
Brown LE. Natural Hybridization and Trend toward Extinction in Some Relict Texas Toad Populations.
Southwest. Nat. 1971,
16, 185–199.
[Google Scholar]
-
19.
Forstner MRJ, Dixon JR. Houston Toad (Bufo houstonensis) 5-Year Review: Summary and Evaluation. Submitted to TPWD and USFWS March 30, 2010. 64 pgs. Available online: https://ecosphere-documents-production-public.s3.amazonaws.com/sams/public_docs/species_nonpublish/1891.pdf (accessed on 23 June 2025).
-
20.
Sirsi S, Rodriguez D, Forstner MRJ. Using genome-wide data to ascertain taxonomic status and assess population genetic structure for Houston toads (
Bufo [=
Anaxyrus]
houstonensis).
Sci. Rep. 2024,
14, 3306. doi:10.1038/s41598-024-53705-w.
[Google Scholar]
-
21.
Brown LE, Mesrobian A. Houston Toads and Texas Politics. In Amphibian Declines; University of California Press: Berkeley, CA, USA, 2005; pp. 150–167.
-
22.
Duarte A, Brown DJ, Forstner MRJ. Documenting Extinction in Real Time: Decline of the Houston Toad on a Primary Recovery Site.
J. Fish Wildlife Manag. 2014,
5, 363–371. doi:10.3996/112013-jfwm-071.
[Google Scholar]
-
23.
Forstner MRJ, Crump P. Houston toad population supplementation in Texas, USA. In Global Re-introduction Perspectives: 2011, Soorae PS, Ed.; IUCN/SSC Re-introduction Specialist Group & Abu Dhabi Environment Agency: Gland, Switzerland, 2011; pp. 71–76.
-
24.
Duarte A, Brown DJ, Forstner MRJ. Estimating Abundance of the Endangered Houston Toad on a Primary Recovery Site.
J. Fish Wildlife Manag. 2011,
2, 207–215. doi:10.3996/072011-jfwm-041.
[Google Scholar]
-
25.
Jones MC, Dixon JR, Forstner MRJ. Is Bigger Always Better? Mate Selection in the Houston Toad (
Bufo houstonensis).
J. Herpetol. 2011,
45, 455–456.
[Google Scholar]
-
26.
Greuter KL. Early Juvenile Ecology of the Endangered Houston Toad, Bufo houstonensis (Anura: Bufonidae). Master’s thesis, Texas State University, San Marcos, TX, USA, 2004.
-
27.
Swannack TM, Grant WE, Forstner MRJ. Projecting population trends of an endangered amphibian species in the face of uncertainty: A pattern-oriented approach.
Ecol. Model. 2009,
220, 148–159.
[Google Scholar]
-
28.
Hatfield JS, Price AH, Diamond DD, True CD. Houston toad (Bufo houstonensis) in Bastrop County, Texas: need for protecting multiple subpopulations. In Species Conservation and Management: Case Studies; Akcakaya HR, Burgman MA, Kindvall O, Wood CC, Sjogren-Gulve P, Hatfield JS, McCarthy CA, Eds.; Oxford University Press: Oxford, UK, 2004; pp. 292–298.
-
29.
Forstner MRJ, McHenry D, Gaston M, Villalobos L, Crump P, McCracken S, et al. The Houston toad 2007: annual research and monitoring. Submitted to the USFWS and collaborating partners. 20pgs.
-
30.
Bell J, Bassett LG, Zughaiyir FZ, Stout DJ, Forstner MRJ. Houston Toad 2021 Final Report for the Griffith League Ranch and Lost Pines Scout Reserve, Bastrop County, Texas; Privately published: Seguin, TX, USA, 2021.
-
31.
Zughaiyir FZ, Sirsi S, Rodriguez D, McHenry DJ, Forstner MRJ. Measuring interspecific admixture in endangered Houston toads (
Bufo [=
Anaxyrus]
houstonensis) at a main recovery site in the Lost Pines ecosystem of Texas, USA.
Conserv. Genet. 2024,
25, 1337–1351. doi:10.1007/s10592-024-01645-3.
[Google Scholar]
-
32.
Bliley JM, Woodley SK. The effects of repeated handling and corticosterone treatment on behavior in an amphibian (Ocoee salamander:
Desmognathus ocoee).
Physiol. Behav. 2012,
105, 1132–1139. doi:10.1016/j.physbeh.2011.12.009.
[Google Scholar]
-
33.
Meise K, Mueller B, Zein B, Trillmich F. Applicability of single-camera photogrammetry to determine body dimensions of pinnipeds: Galapagos sea lions as an example.
PLoS One 2014,
9, e101197. doi:10.1371/journal.pone.0101197.
[Google Scholar]
-
34.
Ramos EA, Landeo-Yauri S, Castelblanco-Martínez N, Arreola MR, Quade AH, Rieucau G. Drone-based photogrammetry assessments of body size and body condition of Antillean manatees.
Mammal. Biol. 2022,
102, 765–779. doi:10.1007/s42991-022-00228-4.
[Google Scholar]
-
35.
Siers SR. Allometric Regression of Snake Body Length from Head Image Measurements.
Wildlife Soc. Bull. 2021,
45, 538–545. doi:10.1002/wsb.1213.
[Google Scholar]
-
36.
Weatherhead PJ, Brown GP. Measurement versus estimation of condition in snakes.
Can. J. Zool. 1996,
74, 1617–1621. doi:10.1139/z96-179.
[Google Scholar]
-
37.
Mott CL, Albert SE, Steffen MA, Uzzardo JM. Assessment of digital image analyses for use in wildlife research.
Wildlife Biol. 2010,
16, 93–100. doi:10.2981/09-010.
[Google Scholar]
-
38.
Lambert MR, Yasuda CM, Todd BD. Evaluation of a photographic technique for estimating body size in lizards from a distance.
Herpetological Conserv. Biol. 2012,
7, 83–88.
[Google Scholar]
-
39.
Davis AK, Connell LL, Grosse A, Maerz JC. A Fast, Non-invasive Method of Measuring Growth in Tadpoles Using Image Analysis.
Herpetol. Rev. 2008,
39, 56–58.
[Google Scholar]
-
40.
Arismendi I, Bury G, Zatkos L, Snyder J, Lindley D. A method to evaluate body length of live aquatic vertebrates using digital images.
Ecol. Evol. 2021,
11, 5497–5502. doi:10.1002/ece3.7444.
[Google Scholar]
-
41.
McHenry D. Genetic Variation and Population Structure in the Endangered Houston Toad in Contrast to its Common Sympatric Relative, the Coastal Plain Toad. PhD Thesis, University of Missouri-Columbia, Columbia, MO, USA, 2010.
-
42.
Schneider CA, Rasband WS, Eliceiri KW. NIH Image to ImageJ: 25 years of image analysis.
Nat. Method. 2012,
9, 671–675.
[Google Scholar]
-
43.
Team RC. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023.
-
44.
Bates D, Maechler M, Bolker B, Walker S. Linear Mixed-Effects Models Using 'Eigen' and S4; 2025. Available online: https://cran.r-project.org/web/packages/lme4/index.html (accessed on 23 June 2025).
-
45.
Kuznetsova A, Brockhoff PB, Christensen RHB. lmerTest Package: Tests in Linear Mixed Effects Models.
J. Stat. Software 2017,
82, 1–26. doi:10.18637/jss.v082.i13.
[Google Scholar]
-
46.
Warton D, Duursma R, Falster D, Taskinen S. (Standardised) Major Axis Estimation and Testing Routines, 2022. Available online: https://cran.r-project.org/web/packages/smatr/smatr.pdf (accessed on 23 June 2025).
-
47.
Brodeur JC, Damonte MJ, Vera Candioti J, Poliserpi MB, D'Andrea MF, Bahl MF. Frog body condition: Basic assumptions, comparison of methods and characterization of natural variability with field data from Leptodactylus latrans.
Ecol. Indicators 2020,
112, 106098–106098. doi:10.1016/j.ecolind.2020.106098.
[Google Scholar]
-
48.
Zhelev ZM, Minchev DS. Body condition based on scaled mass index in adult
Pelophylax ridibundus (Amphibia: Ranidae) inhabiting a polluted area in Southern Bulgaria.
Biologia 2023,
79, 201–213. doi:10.1007/s11756-023-01521-4.
[Google Scholar]
-
49.
Estrada A, Medina D, Gratwicke B, Ibanez R, Belden LK. Body condition, skin bacterial communities and disease status: insights from the first release trial of the limosa harlequin frog,
Atelopus limosus.
Proc. Biol. Sci. 2022,
289, 20220586. doi:10.1098/rspb.2022.0586.
[Google Scholar]