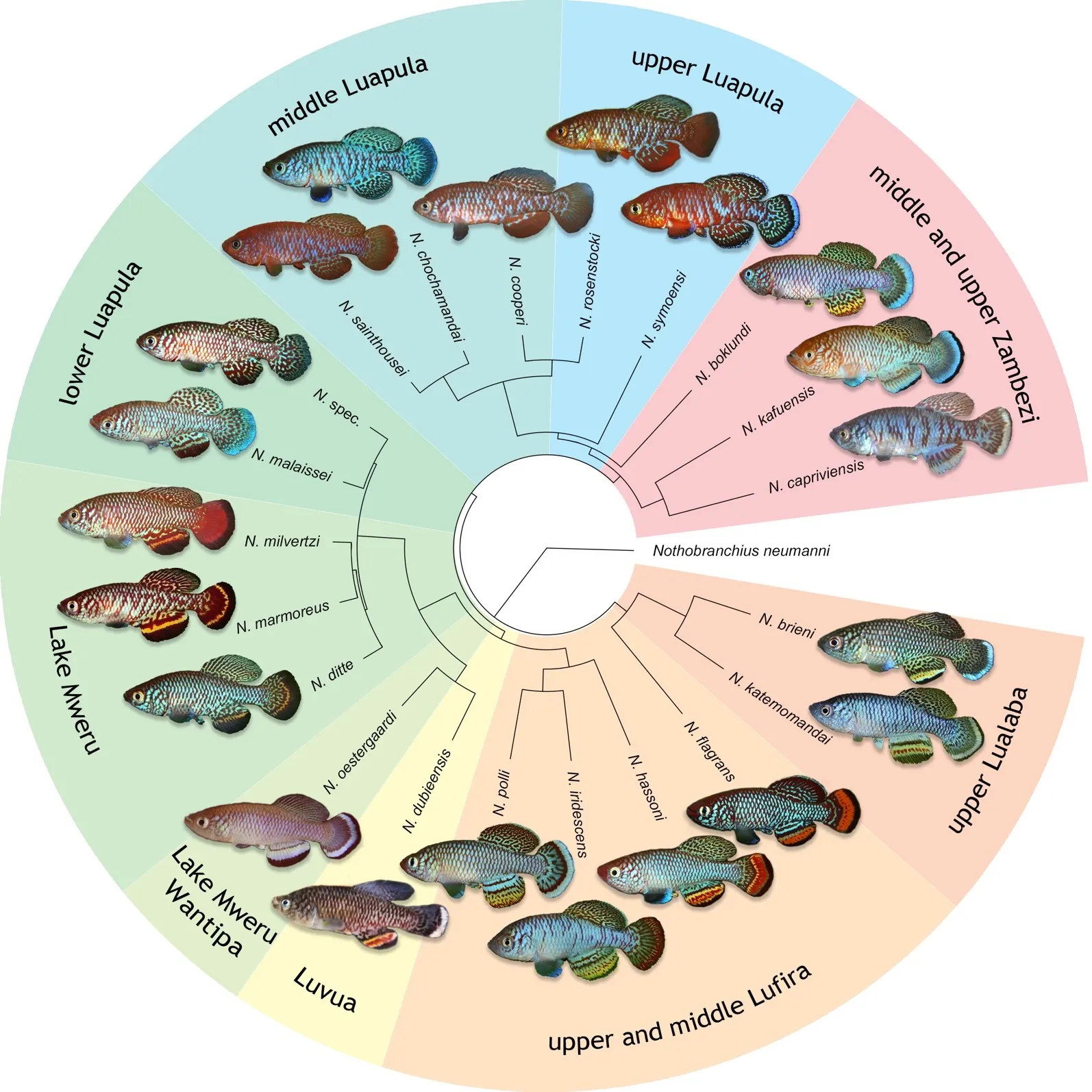
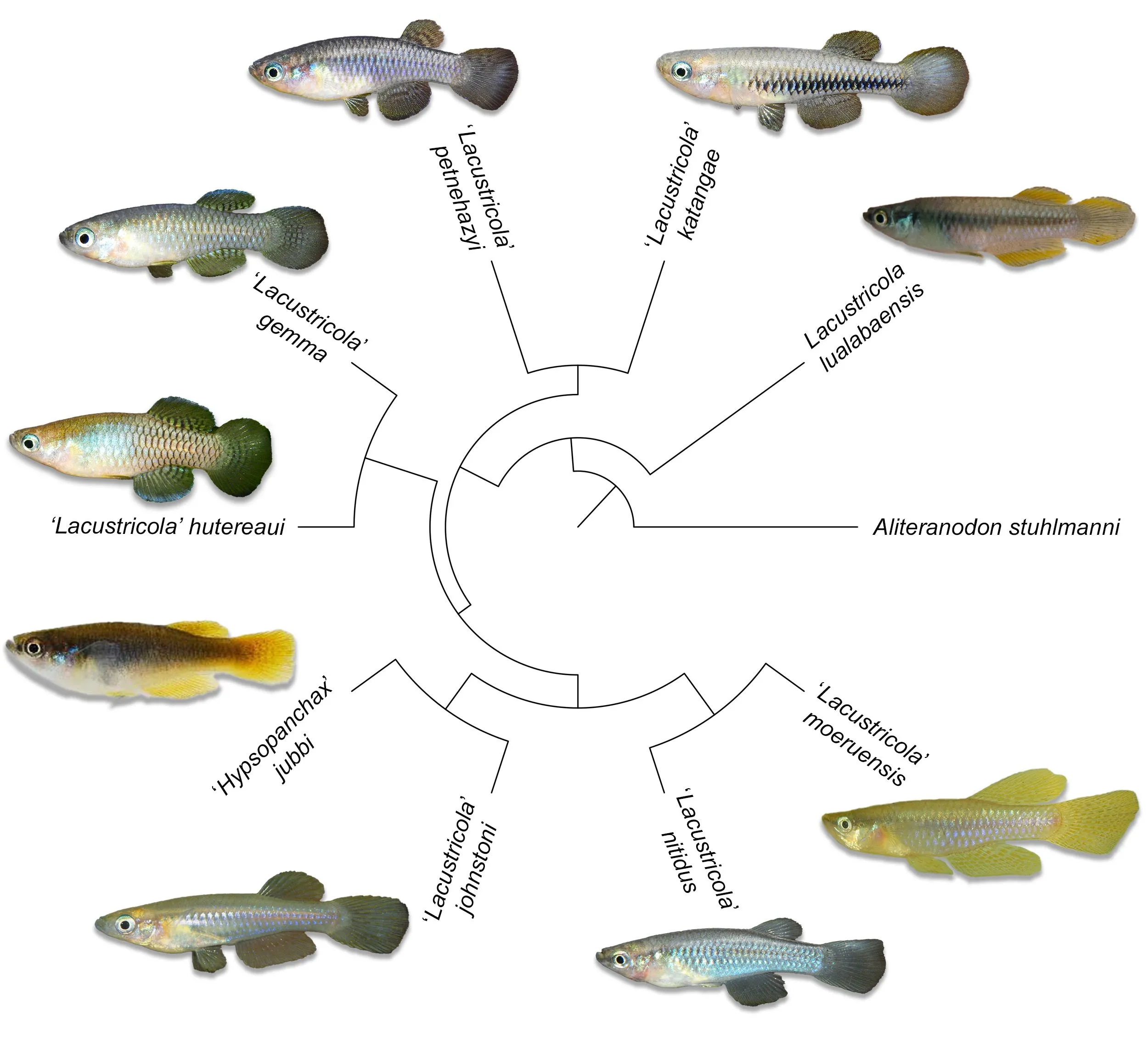
Within the cyprinodotiform fish families in the Bangweulu–Mweru and the upper Lualaba freshwater ecoregions in the upper Congo drainage, 20 species and one putative species of the family Nothobranchiidae are recovered, all belonging to the
Nothobranchius brieni species group within the genus
Nothobranchius (
). All species can be identified based on the colour pattern of males, morphometrics, and molecular analysis.
There are nine species of the family Procatopodidae known in the uppermost Congo drainage, one belonging to the genus
Lacustricola and eight species belonging to the undescribed genus ‘
Lacustricola’, including one species currently referred to as ‘
Hypsopanchax’ (
). These species can be identified based on the colour pattern of males, meristics, morphometrics, and molecular analysis
3.1. Morphological Analysis
3.1.1. Nothobranchiidae
Divergence of morphological characters between males of species belonging to the
N.
brieni species group was found. Populations of the new species show diagnostic, non-overlapping morphometric characters from the most closely related species.
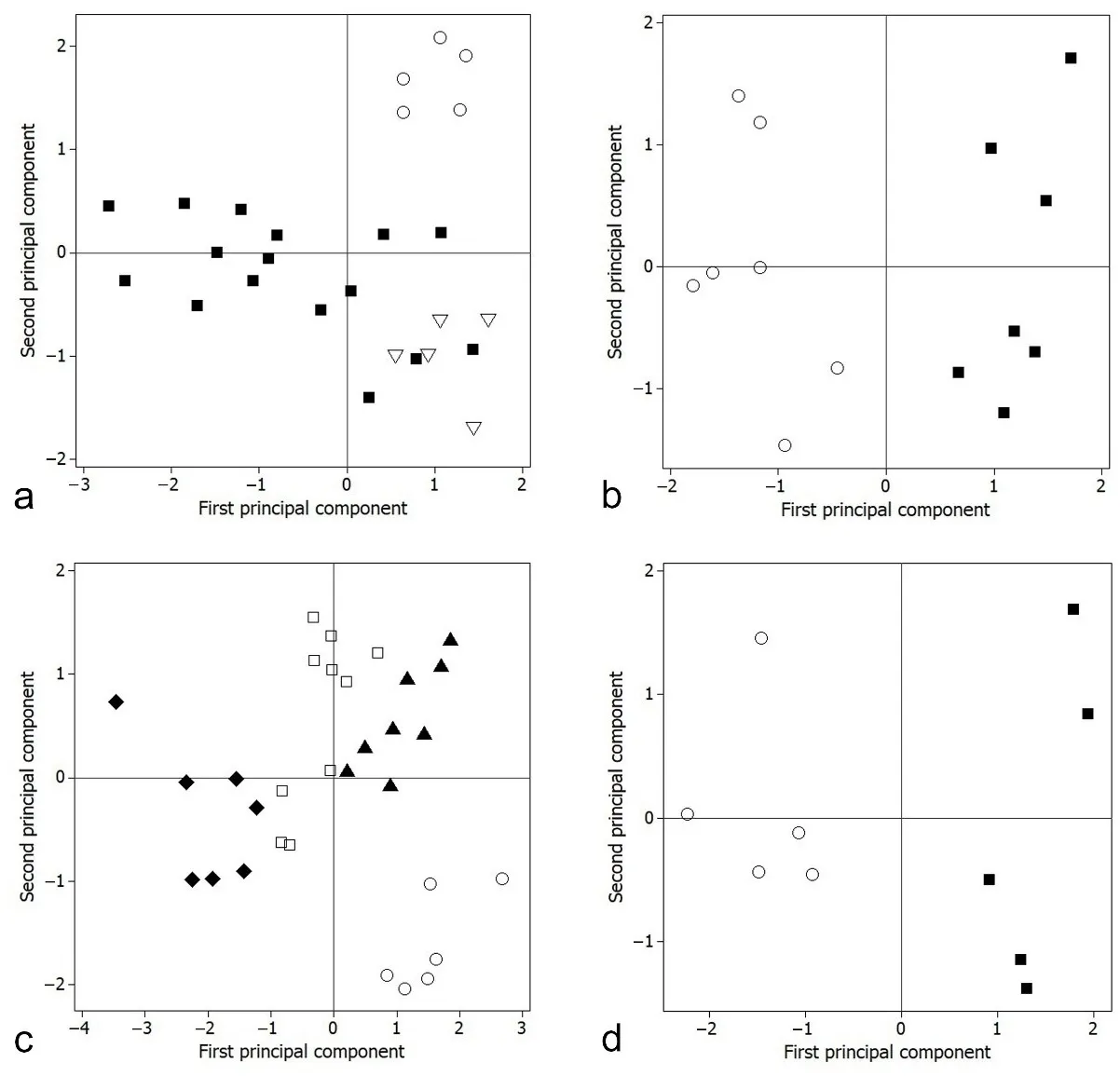
In order to highlight the biometric differences, the PCA technique was employed on the correlation matrices of the most distinctive morphometric characters, in order to corroborate the hypotheses of species delimitation. The best subsets regression resulted in distinctive subsets of the most significant predictors that are shown in
,
,
and
. The first two principal components were retained for each analysis, supported by the eigenvalue-one criterion and proportion of the components in total variance. The proportion of variance explained by the retained principal components and factor loadings of each variable is also shown in
,
,
and
. The series of analyses showed that the new members of the
N.
brieni species group cluster separately from other most similar and closely related species in the space of the first and second principal components (
a–d).
Nothobranchius iridescens was discriminated from
N.
hassoni and
N. polli on the axis of the second principal component [PC2] (
a), shown by differences mainly associated with caudal peduncle length and anal-fin base length (
).
Nothobranchius katemomandai was distinguished from
N. brieni on PC1 (
b), reflecting differences mainly in postorbital length and dorsal-fin base length (
).
Nothobranchius marmoreus was differentiated from
N.
ditte,
N.
malaissei, and
N.
milvertzi on PC2 (
c); associated with differences mainly in head width; whereas also on PC1 from
N.
ditte and
N.
malaissei, associated with caudal peduncle length, predorsal length in % of preanal length, and head length (
).
Nothobranchius dubieensis was discriminated from
N. oestergaardi on PC1 (
d), representing mainly morphometric differences in snout length and caudal peduncle length in % of its depth (
).
The non-overlapping clusters of specimens recovered by the PCA support the hypothesis that the new species of the
N.
brieni species group can also be separated by morphological characters.
. Comparative morphometry in males of the <em>Nothobranchius brieni </em>species group. Score plots of principal component analysis on best subsets of distinctive morphometric characters, first component <em>vs.</em> second component: (<strong>a</strong>). <em>N. iridescens</em> (○) <em>vs.</em> <em>N. hassoni</em> (■) and <em>N. polli</em> (▽); (<strong>b</strong>). <em>N. katemomandai</em> (○) <em>vs.</em> <em>N. brieni</em> (■); (<strong>c</strong>). <em>N. marmoreus</em> (○) vs <em>N. ditte</em> (□), <em>N. malaissei</em> (♦) and <em>N. milvertzi</em> (▲); and (<strong>d</strong>). <em>N. dubiensis</em> (○) <em>vs.</em> <em>N. oestergaardi</em> (■).
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the log-transformed data of most distinctive combination of four measurement characters, identified by best subset regression for Nothobranchius iridescens and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Morphometric characters |
|
|
| Caudal peduncle length in % of its depth |
0.187 |
0.881 |
| Caudal-fin length |
0.683 |
0.023 |
| Postorbital length |
−0.522 |
−0.078 |
| Anal-fin base length |
0.475 |
−0.466 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
1.7158 |
1.0259 |
| Explained variance (% of total variance) |
42.9 |
25.6 |
| Cumulative variance (%) |
|
68.5 |
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the log-transformed data of most distinctive combination of three measurement characters, identified by best subset regression for Nothobranchius katemomandai and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Morphometric characters |
|
|
| Caudal peduncle depth |
0.088 |
0.974 |
| Postorbital length |
0.711 |
0.084 |
| Dorsal-fin base length |
−0.698 |
0.208 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
1.7309 |
1.0282 |
| Explained variance (% of total variance) |
57.7 |
34.3 |
| Cumulative variance (%) |
|
92.0 |
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the log-transformed data of most distinctive combination of four measurement characters, identified by best subset regression for Nothobranchius marmoreus and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Morphometric characters |
|
|
| Head width |
0.141 |
0.915 |
| Caudal peduncle length in % of its depth |
0.589 |
−0.235 |
| Predorsal length in % of preanal length |
0.570 |
−0.221 |
| Head length |
−0.555 |
−0.244 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
2.0847 |
1.0663 |
| Explained variance (% of total variance) |
52.1 |
26.7 |
| Cumulative variance (%) |
|
78.8 |
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the log-transformed data of the most distinctive combination of four measurement characters, identified by best subset regression for Nothobranchius dubieensis and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Morphometric characters |
|
|
| Preanal length |
−0.348 |
−0.779 |
| Snout length |
0.606 |
0.173 |
| Caudal peduncle length in % of its depth |
0.560 |
−0.228 |
| Body depth at pelvic-fin origin |
−0.445 |
0.558 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
2.4876 |
1.0613 |
| Explained variance (% of total variance) |
62.2 |
26.5 |
| Cumulative variance (%) |
|
88.7 |
3.1.2. Procatopodidae
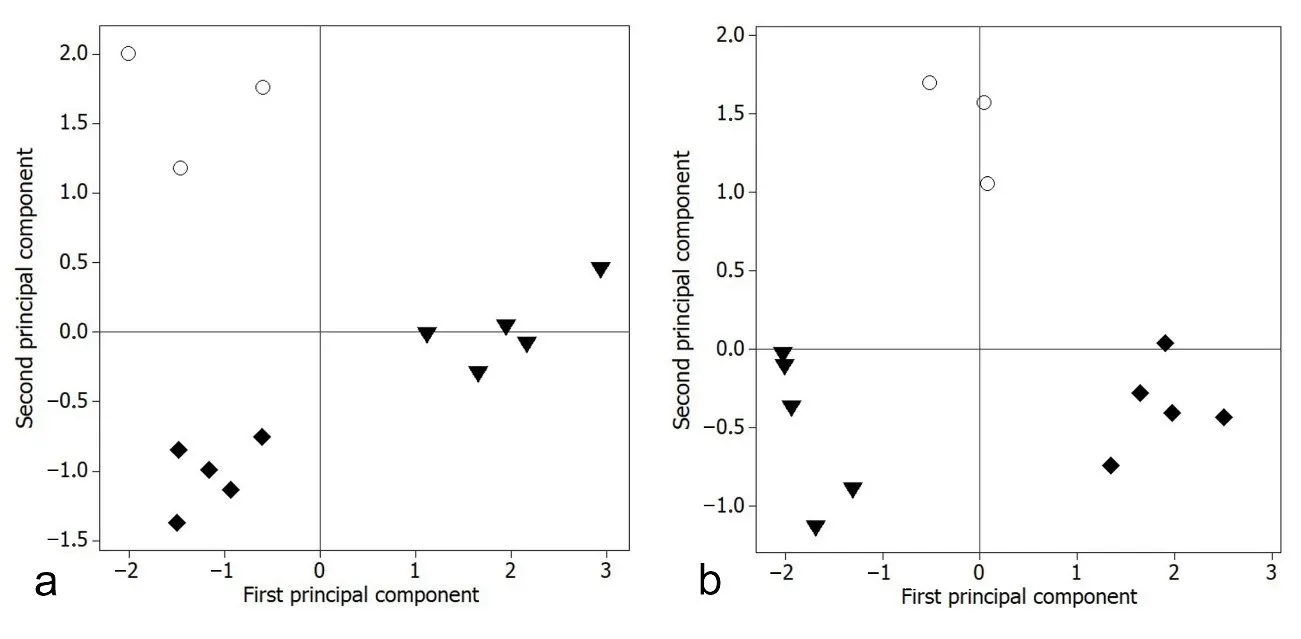
Divergence of morphological characters between populations of the new species and the most similar and closely related species have been found, showing diagnostic, non-overlapping morphometric characters.
The PCA reveals that the new population, here identified as a new species and named ‘
Lacustricola’
gemma, groups separately on score plots for PC1
vs. PC2 (
a,b) from ‘
L.’
chobensis and ‘
L.’
hutereaui. For meristic characters (
a), the first principal component is mostly associated with the scales in the circumpeduncular series, whereas the second component accounts for the scales in the transverse series and dorsal-fin rays (
). For the morphometric characters (
b), the first principal component explains much of the variation among specimens in Head width in % of its depth and caudal fin length, whereas the second principal component is associated mainly with head width and body depth (
).
. Comparative meristics and morphometry in males of ‘<em>Lacustricola</em>’ <em>gemma</em> (○), ‘<em>L</em>.’ <em>chobensis</em> (♦) and ‘<em>L</em>.’ <em>hutereaui</em> (▼). Score plot of principal component analysis on first component <em>vs.</em> second component: (<strong>a</strong>). six meristic characters; (<strong>b</strong>). best subset of four distinctive morphometric characters.
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the square-root transformed data of meristic characters for ‘Lacustricola’ gemma and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Meristic characters |
|
|
| Scales transverse |
0.268 |
−0.792 |
| Dorsal-fin rays |
−0.140 |
0.657 |
| Scales circumpeduncular |
0.558 |
0.021 |
| Anal-fin rays |
0.460 |
0.527 |
| Scales mid-longitudinal series |
0.472 |
−0.250 |
| Dorsal fin to anal fin relative position |
0.428 |
0.178 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
2.4876 |
1.0613 |
| Explained variance (% of total variance) |
62.2 |
26.5 |
| Cumulative variance (%) |
|
88.7 |
.
Factor loadings and proportions of variance explained by the selected first two principal components (PC1 and PC2) of a principal component analysis (PCA) carried out on the log-transformed data of the most distinctive combination of four measurement characters, identified by best subset regression for ‘Lacustricola’ gemma and the comparative material. The most important loading values are in bold.
| Variables and Eigenanalysis |
PC1 |
PC2 |
| Morphometric characters |
|
|
| Head width in % of its depth |
0.377 |
0.822 |
| Caudal peduncle length in % of its depth |
0.565 |
−0.148 |
| Body depth at pelvic-fin origin |
−0.562 |
−0.058 |
| Caudal-fin length |
−0.473 |
0.547 |
| Eigenanalysis of the correlation matrix |
|
|
| Eigenvalue |
2.9202 |
1.0050 |
| Explained variance (% of total variance) |
73.0 |
20.1 |
| Cumulative variance (%) |
|
93.1 |
3.2. Molecular Analysis
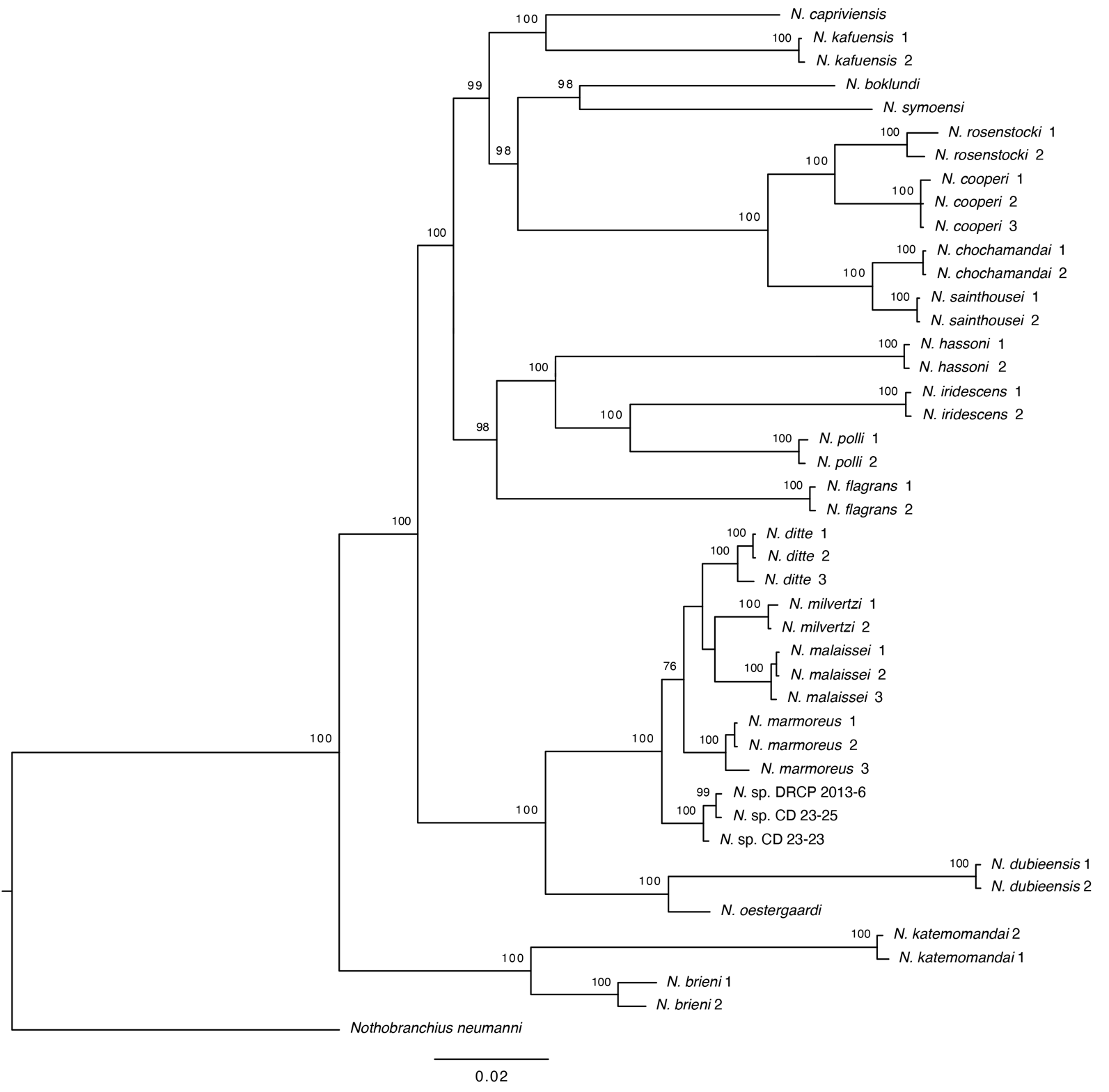
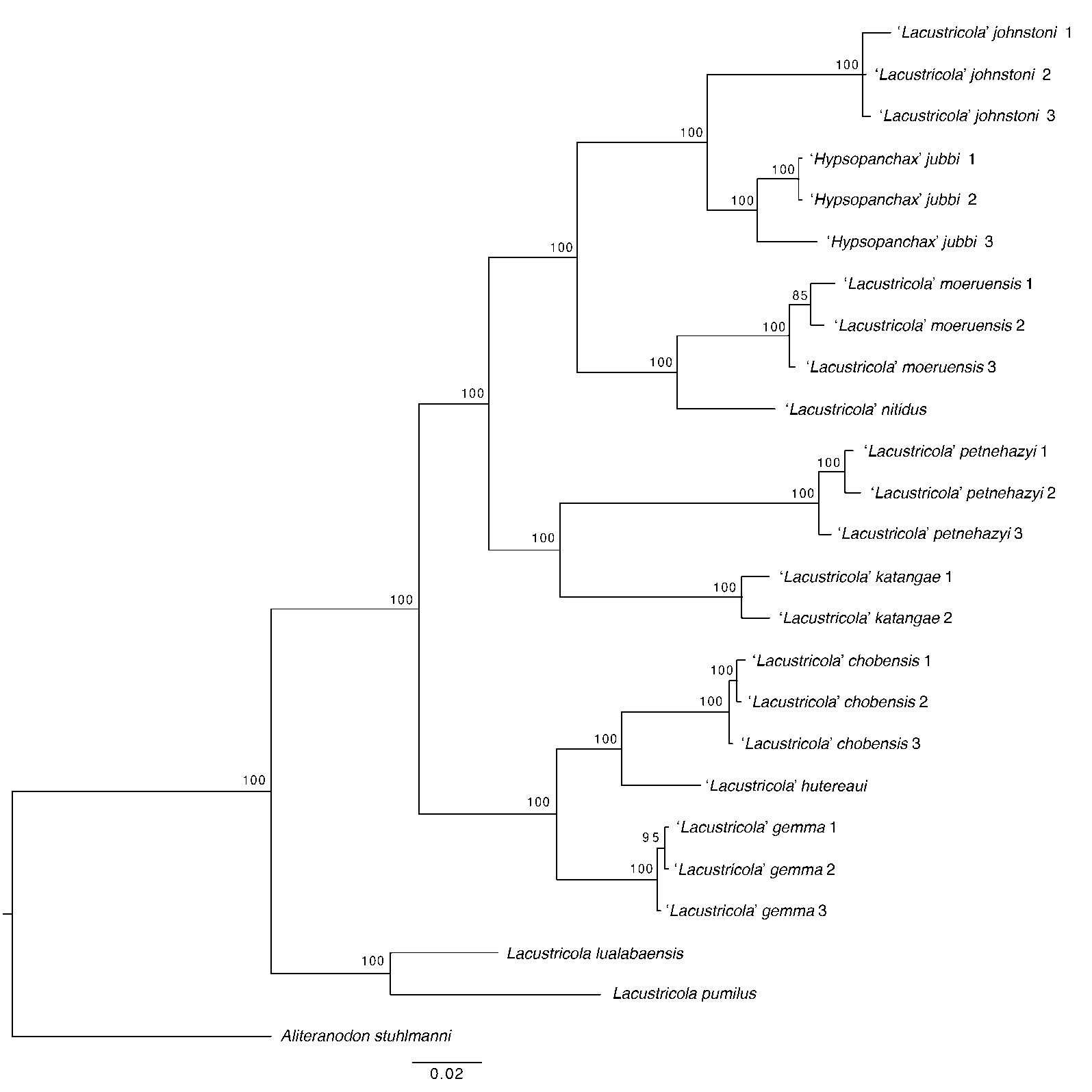
DNA sequence data from the mitochondrial COI genes were generated and utilized for all taxa, complementing the sequences available from previous analyses. The BI and ML analyses recovered similar topologies. For Nothobranchiids, a full BI phylogeny including all analysed taxa is presented in
, while a simplified ML phylogeny, with one terminal per species and accompanying specimen images to illustrate species diversity, is shown in
. For Procatopodids, the corresponding BI and simplified ML phylogenies are presented in
and
, respectively.
. Phylogenetic tree of the <em>Nothobranchius brieni</em> species group, based on analysis of the mitochondrial molecular marker Cytochrome oxidase subunit I (COI), using Bayesian inference. Support values at nodes represent Bayesian posterior probability. Relevant data regarding specimens analysed for this tree are presented in .
. Colour images of representative male specimens of all members of the <em>Nothobranchius brieni</em> species group, plotted on a phylogenetic tree based on maximum likelihood analysis of the mitochondrial molecular marker Cytochrome Oxidase Subunit I (COI), and associated drainage system distribution information. Photographs by B. Nagy, except <em>N. kafuensis</em> by Cs. Nagy, <em>N. oestergaardi</em> by S. Valdesalici and <em>N. dubieensis</em> by A. Kalumba.
. Phylogenetic tree of the Procatopodid species to examine the phylogenetic structure of the family in the upper Congo drainage, based on analysis of the mitochondrial molecular marker Cytochrome oxidase subunit I (COI), using Bayesian inference. Support values at nodes represent Bayesian posterior probability. Relevant data regarding specimens analysed for this tree are presented in .
. Colour images of representative male specimens of the Procatopodid species in the upper Congo drainage, plotted on phylogenetic tree based on maximum likelihood analysis of the mitochondrial molecular marker Cytochrome Oxidase Subunit I (COI). Photographs by B. Nagy, except ‘<em>H</em>.’ <em>jubbi</em> by SAIAB, <em>L. lualabaensis</em> by B. Katemo Manda, and ‘<em>L</em>.’ <em>johnstoni</em> by B. Watters.
3.2.1. Nothobranchiidae
The BI phylogeny (
) retrieved the
N. brieni species group in four, geographically segregated clades:
Nothobranchius brieni and
N. katemomandai from the upper Lualaba drainage in a basal position; the species from lower Luapula drainage and Lake Mweru basin (referred to as Lake Mweru complex in [
36]) as well as containing
N. oestergaardi from Lake Mweru Wantipa and
N. dubieensis from nearby Luvua system in a clade; the species from the upper and middle Lufira drainage in a clade; and the species from the middle and upper Luapula drainage and the Zambezi drainage were retrieved in a clade.
The ML phylogeny retrieved a similar topology, except that
N. brieni and
N. katemomandai were in a sister position to
N. flagrans. The phylogeny based on ML reveals strong geographic segregation, with clades organized by the host river drainages. The ML phylogeny, including one terminal per species, and illustrated with colour pictures of each taxon, is represented in
.
In both analyses, the four new species of the
N. brieni species group are retrieved as phylogenetically distinct lineages. The results of molecular analyses of COI gene sequences support the genetic distinction of the four new species and confirm their position, together with all known members in the
N. brieni species group.
3.2.2. Procatopodidae
The BI phylogeny (
) retrieved the family in the upper Congo drainage in the following clades:
Lacustricola lualabaensis from the upper Congo forms a clade with
L. pumilus from the Lake Tanganyika basin; followed by clades likely belonging to the undescribed genus ‘
Lacustricola’, generally recognized as the ‘
L.’
johnstoni group, the ‘
L.’
katangae group, and the ‘
L.’
hutereaui group. The new populations were found to belong to the latter group.
The ML phylogeny retrieved a similar topology. The phylogeny based on ML, including one terminal per species from the target study area, and illustrated with colour pictures of each taxon, is represented in
.
The new populations are retrieved as a phylogenetically distinct lineage, and the analyses confirm the position within the ‘
Lacustricola’ genus.
3.3. Taxonomy
3.3.1.
Nothobranchius iridescens, New Species
http://zoobank.org/urn:lsid:zoobank.org:act: 2CDB599E-DAFC-4FF6-BDEA-D9D4760E9049
Nothobranchius spec. ‘Lutandula’: Nagy 2024b: 73 [
82].
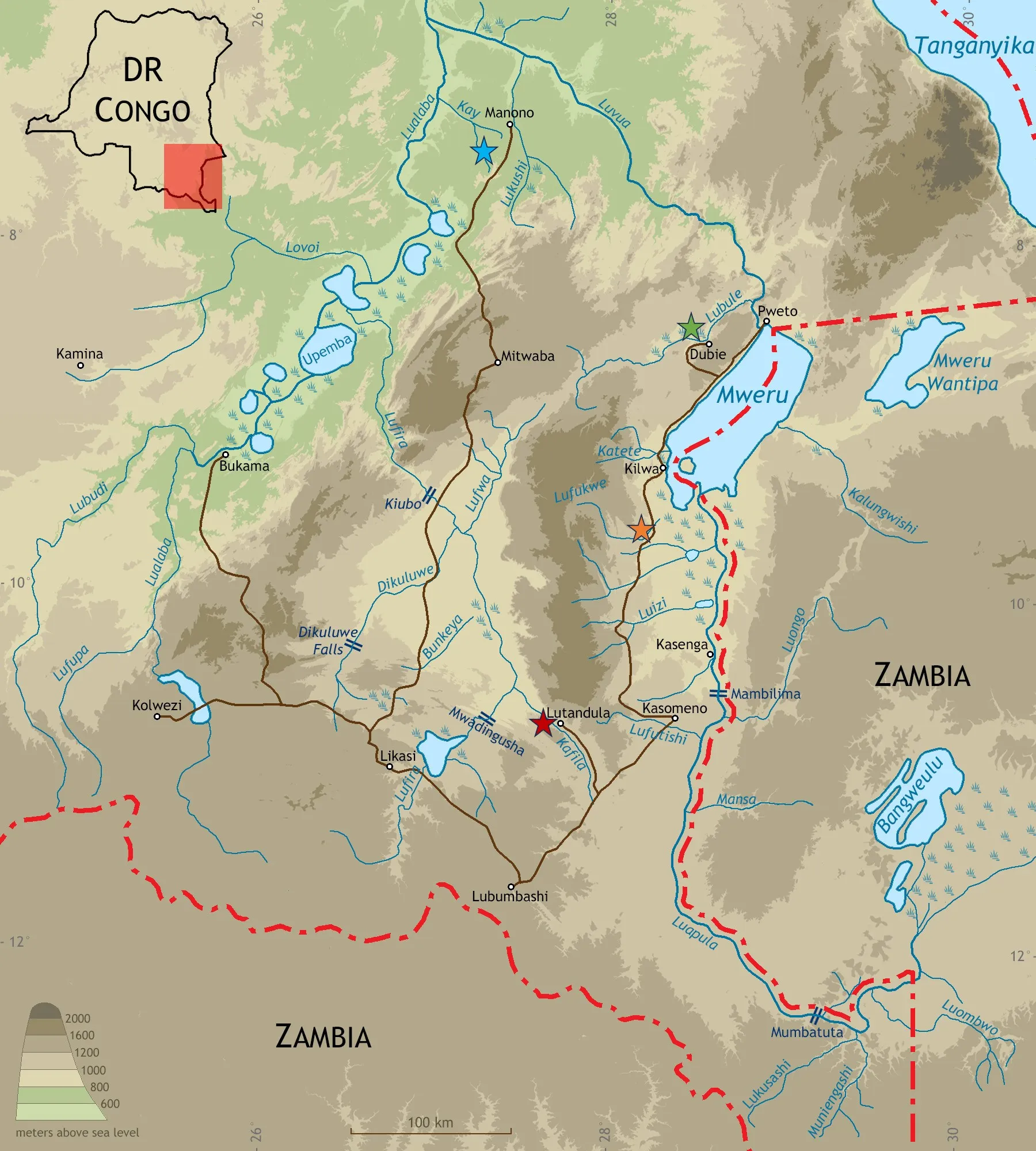
Holotype. BE_RMCA_VERT.2025.008.P.0003, male, 41.8 mm SL; DR Congo: Kafila system: Lufira drainage: shallow remnant pools in seasonal floodplain, 8 km northwest of Lutandula village, 10°49′44″ S, 27°48′09″ E, 1062 m alt.; B. Nagy, A. Chocha Manda & A. Kalumba, 19 April 2023 (field code: CD 23-21).
Paratypes. BE_RMCA_VERT.2025.008.P.0004, male, 43.0 mm SL; collected with the holotype. — BE_RMCA_VERT.2025.008.P.0005–0010, 3 males, 29.2–36.2 mm SL & 3 females, 27.0–29.9 mm SL; DR Congo: Kafila system: Lufira drainage: shallow remnant pools in seasonal floodplain, 11 km northwest of Lutandula village, 10°48′36″ S, 27°47′19″ E; A. Kalumba, 28 May 2023 (field code: CD 23-30).
Diagnosis. Nothobranchius iridescens is distinguished from all other species of the genus by the unique combination in male colouration of having anal fin with irregular red-brown spots and stripes, creating two irregular submedial and medial bands and with broad yellow subdistal band; and caudal fin with a slender light blue subdistal band, densely marked with irregular red spots, and narrow dark brown distinct distal margin.
Further,
N. iridescens is distinguished from the most closely related
N. polli by greater caudal peduncle length (153–161 in % of its depth
vs. 133–139); and greater eye diameter (31–34% HL
vs. 23–26); and from
N. hassoni by smaller predorsal length (52.4–55.6% SL
vs. 55.7–59.5); and greater caudal peduncle length (153–161 in % of its depth
vs. 133–143).
Description. General body features are illustrated in
. Morphometric and meristic characters of the holotype and paratypes are summarised in
. A medium sized
Nothobranchius species (maximum observed size: 43.0 mm SL in males and 29.9 mm SL in females).
Males: General body shape is robust, laterally compressed, and deep. Greatest body depth at vertical in front of pelvic-fin origin. Greatest body width at pectoral-fin base, with body progressively narrowing towards caudal-fin base. Dorsal profile straight from tip of snout to nape and convex to base of last dorsal-fin ray, straight to slightly concave on caudal peduncle. Ventral profile convex from lower jaw to base of last anal-fin ray, straight to slightly concave on caudal peduncle. Caudal peduncle shallow, length 1.5–1.6 times of its depth. Anus situated directly in front of anal-fin origin.
Head short, laterally compressed, deeper than wide. Snout slightly pointed, smaller than eye diameter. Mouth supraterminal, slightly oblique in profile. Jaws subequal, lower jaw longer than upper, posterior end of rictus at same level or slightly ventral to centre of eye. Premaxilla and dentary with many irregularly distributed conical, slightly curved teeth in the outer row of lower and upper jaws. Orbit large, in the anterior half of the head, in the dorsal portion of the head side. The branchiostegal membrane projects posteriorly from the operculum.
Dorsal-fin origin anterior to anal-fin origin, with both fins originating posterior to mid-length of the body. Overall distal part of dorsal and anal fins rounded, with small contact organs in form of papillae on fin rays and distal margin with short filamentous rays. Posterior tip/margin of dorsal fin reaching caudal-fin base. Pectoral fin subtriangular, insertion at about vertical or slightly posterior to margin of opercular opening, base slightly oblique, with 17–18 rays, upper fin rays placed slightly anteriorly to lower fin rays, tip reaching or slightly overlapping base of pelvic fin. Pelvic fin subabdominal, origin at about mid-length of body, short, bases medially separated, tip reaching urogenital papilla. Caudal fin rounded, with 17–18 branched rays, plus 2 to 3 unbranched, smaller rays at dorsal and ventral origins.
Scales cycloid. Body and head entirely scaled, except for the ventral surface of the head.
. <i>Nothobranchius iridescens</i>: (<b>a</b>). BE_RMCA_VERT.2025.008.P.0003, holotype, preserved male, 41.8 mm SL; (<b>b</b>). holotype live; (<b>c</b>). BE_RMCA_VERT.2025.008.P.0004, paratype, live male, 43.0 mm SL; (<b>d</b>). type locality; DR Congo: Kafila system: Lufira drainage: shallow remnant pools in seasonal floodplain, 8 km northwest of Lutandula village, 10°49′44″ S, 27°48′09″ E, 1062 m alt.; B. Nagy, A. Chocha Manda & A. Kalumba, 19 April 2023 (field code: CD 23-21). Photographed by B. Nagy (<b>a</b>–<b>c</b>) and A. Kalumba (<b>d</b>).
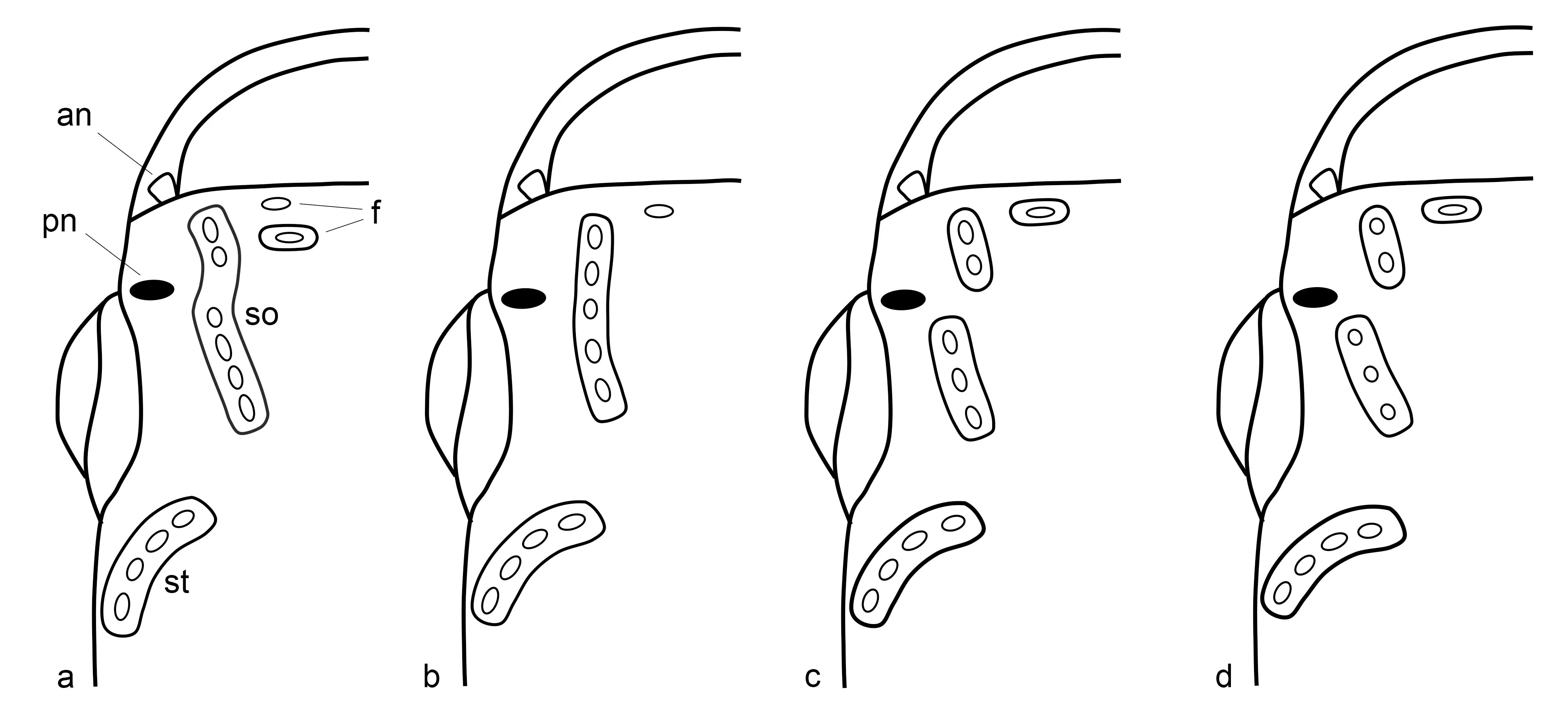
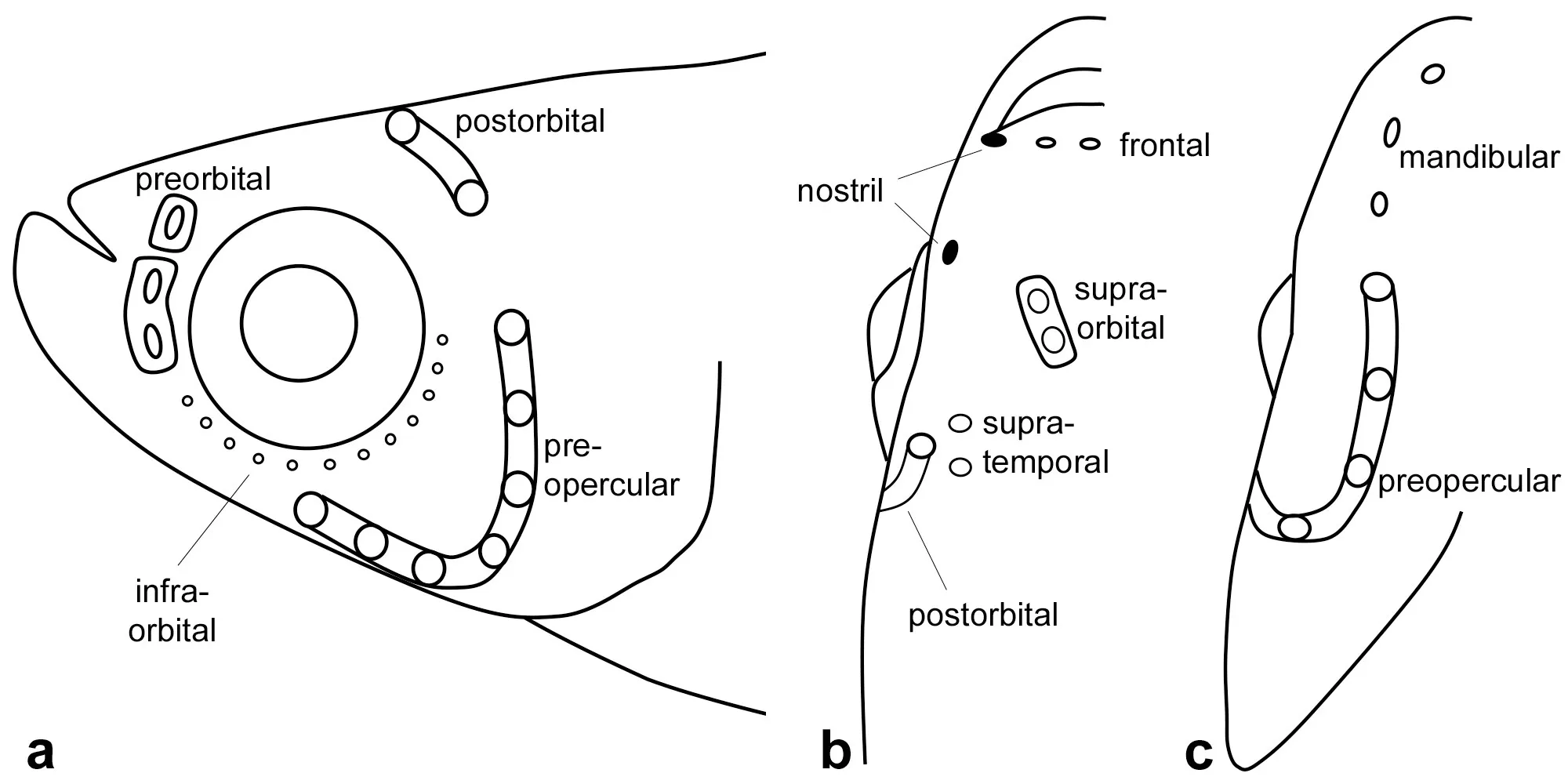
Cephalic squamation pattern variable. Anterior nostril at the anterior tip of the snout, tubular opening lateral to the upper lip. Posterior nostril in front of the orbit, with oblique oval aperture. Frontal neuromasts are in a shallow groove. Cephalic sensory system at supraorbital level in a continuous, curved shallow groove, with two exposed neuromasts in the anterior part and four exposed neuromasts in the posterior part, whereas at supratemporal level in a curved groove, with four exposed neuromasts (
a). Preorbital canal in shallow groove with two exposed neuromasts; postorbital canal in shallow groove with one exposed neuromast; infraorbital series with about eight neuromasts at ventral margin of eye. Mandibular canal is in a shallow groove with a series of small neuromasts, curved to the lateral midline in front. One neuromast is on each scale along the trunk mid-longitudinal series.
Females: Smaller than males. Body generally similar but less laterally compressed and slightly more slender than in males (body depth at pelvic fin origin 27.4–28.1% SL
vs. 29.1–32.3; head width in % of its depth greater than in males 77–82
vs. 64–72). Anal fin subtriangular, tip rounded, central rays longer and more rigid (
vs. anal fin rounded in males). Anal, dorsal and pelvic fins positioned more posteriorly than in males (61.5–63.5% SL
vs. 56.1–59.6; 57.5–58.0% SL
vs. 52.4–55.6; 49.5–50.7% SL
vs. 44.5–47.8, respectively). Anal-fin and dorsal-fin base lengths are smaller than in males (23.1–25.9% SL
vs. 26.1–29.0; 17.7–18.1% SL
vs. 22.3–23.5, respectively). Caudal peduncle is more slender than in males (161–164 in % of its depth
vs. 153–161). Branchiostegal membrane not projecting distally (
vs. projecting distally in males). No papillae or epidermal tissue present on dorsal and anal fins (
vs. both present in males).
. Diagrammatic representation of cephalic structure, dorsal view of head in: (<strong>a</strong>). <em>Nothobranchius iridescens</em>; (<strong>b</strong>). <em>N. katemomandai</em>; (<strong>c</strong>). <em>N. marmoreus</em>; and (<strong>d</strong>). <em>N. dubieensis</em>. Key to label abbreviations: an, anterior nostril; pn, posterior nostril; f, frontal neuromast; so, supraorbital canal; st, supratemporal canal.
Live colouration of males (
b,c): scales on head and trunk light blue to green-blue with irregular red-brown posterior margin, forming irregular vertical, chevron-formed striped pattern on body. Snout, frontal and dorsal portion of head red-brown, throat light blue. Posterior scale margins on post-orbital portion of the operculum create three to four red-brown, anteriorly lowering, oblique bars. Exposed part of the branchiostegal membrane is light blue. Iris light yellow, with dark grey-black spots, especially on upper and lower-most parts, creating a dark vertical bar through the centre of the eye. Dorsal fin yellow to light blue with irregular red-brown stripes, perpendicular to fin rays proximally, parallel to fin rays medially and distally, fin tips light blue. Anal fin light blue, with irregular red-brown spots and stripes, creating two irregular submedial and medial bands, with broad yellow subdistal band, with irregular red-brown spots and narrow dark brown distal margin. Caudal fin yellow to light blue, with irregular red-brown spots and stripes parallel to fin rays proximally and medially, followed by an irregular narrow red-brown band and a slender light blue subdistal band, with irregular red spots, depending on the density, creating a red-brown band, and a narrow dark brown distinct distal margin. Pelvic fin light blue, with narrow red-brown medial band and yellow distal band. Pectoral fin hyaline with blue posterior distal margin.
Live colouration of females: scales on trunk and head light brown with narrow dark brown posterior margin, forming a slightly visible reticulated pattern on body. Overall light brown colour of scales, darker on dorsum and lighter to silvery on venter. Blue iridescence on the opercular region and anterior midlateral half of the trunk. Iris yellow. All fins are hyaline.
.
Morphometric and meristic data of holotype and paratypes of Nothobranchius iridescens. Holotype values included in ranges, mean and standard deviation of males. H, holotype; SD, standard deviation.
| Morphometric and Meristic Characters |
|
Males |
Females |
|
(n = 5) |
(n = 3) |
| H |
Range |
Mean |
SD |
Range |
Mean |
SD |
| Standard length |
41.8 |
29.2–43.0 |
|
|
27.0–29.9 |
|
|
| Percent of standard length |
|
|
|
|
|
|
|
|
Total length |
123.0 |
123.0–125.7 |
124.7 |
1.1 |
125.4–127.8 |
126.4 |
1.2 |
|
Body depth at pelvic-fin origin |
32.3 |
29.1–32.3 |
30.8 |
1.3 |
27.4–28.1 |
27.8 |
0.4 |
|
Head length |
30.6 |
27.6–31.9 |
29.8 |
1.7 |
31.4–33.3 |
32.3 |
1.0 |
|
Preanal length |
59.6 |
56.1–59.6 |
57.1 |
1.4 |
61.5–63.5 |
62.2 |
1.2 |
|
Predorsal length |
54.8 |
52.4–55.6 |
54.1 |
1.3 |
57.5–58.0 |
57.8 |
0.2 |
|
Prepelvic length |
47.8 |
44.5–47.8 |
46.4 |
1.4 |
49.5–50.7 |
50.1 |
0.6 |
|
Prepectoral length |
30.9 |
28.8–31.9 |
30.4 |
1.4 |
31.8–33.7 |
32.7 |
1.0 |
|
Caudal peduncle length |
21.5 |
19.8–22.1 |
21.0 |
0.9 |
20.0–20.8 |
20.4 |
0.4 |
|
Caudal peduncle depth |
13.4 |
12.8–14.1 |
13.4 |
0.5 |
12.2–12.7 |
12.6 |
0.3 |
|
Dorsal-fin base length |
26.1 |
26.1–29.0 |
27.5 |
1.2 |
23.1–25.9 |
24.2 |
1.5 |
|
Anal-fin base length |
23.0 |
22.3–23.5 |
22.8 |
0.5 |
17.7–18.1 |
18.0 |
0.3 |
|
Caudal-fin length |
23.0 |
23.0–25.7 |
24.7 |
1.1 |
25.4–29.3 |
26.9 |
2.0 |
| Percent of head length |
|
|
|
|
|
|
|
|
Head width |
50 |
53–59 |
56.8 |
2.5 |
55–59 |
56.3 |
1.9 |
|
Head depth |
87 |
82–87 |
84.7 |
2.6 |
69–72 |
70.9 |
1.5 |
|
Interorbital width |
39 |
36–40 |
38.3 |
1.9 |
36–39 |
37.1 |
1.5 |
|
Postorbital length |
50 |
50–55 |
51.7 |
2.0 |
48–52 |
50.2 |
2.2 |
|
Suborbital depth |
24 |
19–24 |
21.0 |
2.0 |
17–21 |
18.5 |
2.4 |
|
Eye diameter |
31 |
31–34 |
31.7 |
1.2 |
34–37 |
35.3 |
1.8 |
|
Snout to eye end length |
50 |
45–50 |
48.3 |
2.0 |
48–52 |
49.8 |
2.2 |
|
Snout length |
20 |
17–20 |
18.6 |
1.7 |
17–19 |
17.8 |
1.2 |
| Other morphometric ratios |
|
|
|
|
|
|
|
|
Head width in % of its depth |
68 |
64–72 |
67.1 |
3.1 |
77–82 |
79.5 |
2.6 |
|
Caudal peduncle length in % of its depth |
161 |
153–161 |
156.4 |
2.7 |
161–164 |
162.7 |
1.9 |
|
Predorsal length in % of preanal length |
92 |
92–98 |
94.8 |
2.8 |
91–94 |
92.9 |
2.1 |
| Meristics |
|
range |
mode |
|
range |
mode |
|
|
Dorsal-fin rays |
16 |
16–17 |
16 |
|
16–17 |
16 |
|
|
Anal-fin rays |
17 |
17–18 |
17 |
|
17–18 |
17 |
|
|
Scales mid-longitudinal series |
29 |
28–30 |
29 |
|
28–29 |
29 |
|
|
Scales transverse |
10 |
10 |
10 |
|
10 |
10 |
|
|
Scales circumpeduncular |
12 |
12 |
12 |
|
12 |
12 |
|
Distribution. Nothobranchius iridescens is endemic to seasonal freshwater habitats of the upper Congo drainage in south-eastern DR Congo. It is currently known from ephemeral pools and marshes on floodplains associated with the Kafila river in the Lufira system (
). The Kafila is a major right bank affluent entering the middle Lufira just below the Mwadingusha Falls, which isolates the middle from the upper Lufira.
. Localities of new species described in present paper: <em>Nothobranchius iridescens</em> (red star), <em>N. katemomandai</em> and ‘<em>Lacustricola</em>’ <em>gemma</em> (blue star), <em>N. marmoreus</em> (orange star), and <em>N. dubieensis</em> (green star). Individual symbols may represent more than one locality or population. Map prepared by B. Nagy.
The climate of the ecoregion is tropical and moist, with a mean annual rainfall of around 1000 mm [
83]. The rivers in the Lufira system flood seasonally in response to the rains. The waters are the highest during the rainy season and, from February to April and driest between September and January [
83,
84].
At the type locality,
Nothobranchius iridescens was the only species of the genus observed. The accompanying fish fauna consisted of non-annual species belonging to the families Cichlidae and Cyprinidae. The type locality on 19 April 2023 was an ephemeral pool formed in the floodplain of the Kafila River (
d). The pool was about 50 cm deep at its deepest point, the water was turbid, and it covered an area that was partly overgrown with grass. The habitat principally belongs to category 1.2 as defined by Watters (2015a) [
85], representing a pool and a flooded grassy area on a floodplain. However, the anthropogenic impact is evident, as the local population altered the natural habitat by placing fences and fish traps into the biotope.
Aquarium maintenance of selected specimens was undertaken in the facilities of the University of Lubumbashi for observation of breeding behaviour and biology.
Nothobranchius iridescens has a mode of reproduction that is common for all known annual congeners under aquarium conditions.
Conservation status. Nothobranchius iridescens is recommended to be assessed as Endangered. The species is currently known only from the area of the type locality, two sites situated in a limited section of the Kafila system of the Lufira drainage. It might exist at some other sites within the drainage system of the Kafila, but its distribution will remain restricted, and any potential additional subpopulations are expected to be fragmented and not in contact or having limited contact with each other. The species is expected to be restricted to ephemeral wetlands within the Kafila drainage system. The section of the drainage in the Kafila system where the species is currently known is 67 km
2, whereas the estimated maximum extent of occurrence (EOO) would be less than 1000 km
2, the area of occupancy (AOO) with a maximum estimate of less than 100 km
2 and expected maximum of five threat-based locations. Currently known sites, including the type locality, are close to human populations. Phases in the seasonal life cycle of this species underscore the vulnerabilities of the ecological processes in the ephemeral habitats, as the survival of the species is dependent on suitable conditions during both dry and wet seasons. The author observed the expansion of agriculture in the area and, consequently, increasing anthropogenic pressures on the land and water use. The resultant habitat changes are likely to modify the habitats in ways that render them unsuitable and thus degraded when considering their support of the seasonal life cycle of the species, and thus represent an important extinction risk. Additionally, the natural habitat at the type locality has been altered by the local population through the installation of fences and the placement of fish traps within the biotope. Using IUCN (2012) principles [
86],
N.
iridescens meets the criteria B1ab(iii)+2ab(iii) for Endangered, considering upper estimates of the EOO, AOO, and number of locations, as well as the risk of continuous decline of the known wetland habitats in which it is known to live.
Nevertheless, at present, no conservation/protection measures are in place for this species. Further, not a single protected area covers at least part of its presently known distribution. As such, there is a need for habitat protection at the type locality and a second close-by site, currently the only sites from which the species has been recorded to date. Further field surveys targeting additional suitable habitats should be conducted in the floodplains of the drainage system, in order better to document the full geographic distribution of this species and thus better envision how to elaborate on appropriate conservation measures.
Etymology. The specific epithet
iridescens is a Latin participial adjective deriving from the ancient Greek word iris (ἶρις), rainbow, referring to the colourful appearance of the males, containing different colours of the rainbow, such as yellow and red in the fins, reflective blue on the body slightly shifting hue depending on the angle of light.
3.3.2.
Nothobranchius katemomandai, New Species
http://zoobank.org/urn:lsid:zoobank.org:act: 22C69DA3-5C13-4C51-BEAF-F10BC311AB27
Nothobranchius spec. CD 23-26: Nagy 2024a: 2 [
87].
Nothobranchius spec. ‘Manono’: Nagy 2024b: 72 [
82]; Watters & Nagy 2025: 426 [
88].
Holotype. BE_RMCA_VERT.2025.008.P.0011, male, 31.1 mm SL; DR Congo: Kay system: upper Lualaba drainage: shallow remnant pool in small seasonal riverbed, 0.5 km west of Lubule village, 07°31′47″ S, 27°13′24″ E, 634 m alt.; B. Katemo Manda, 17 April 2023 [field code: CD 23-26].
Paratypes. BE_RMCA_VERT.2025.008.P.0012–0020, 6 males, 30.3–43.2 mm SL & 3 females, 27.0–28.8 mm SL; collected with the holotype.
Diagnosis. Nothobranchius katemomandai is distinguished from all other species of the genus by the unique combination in male colouration of having anal fin with narrow brown submedial band, followed by a slender yellow band, a slender red-brown band and a slender dark brown distal band; and caudal fin with brown spots proximally and medially, and with slender white to light blue subdistal band and a narrow dark grey distal band.
Further,
N. katemomandai is distinguished from most closely related
N. brieni by greater dorsal-fin base length (25.7–28.0% SL
vs. 24.1–25.4); smaller postorbital length (49–51% HL
vs. 53–58); and smaller head width (68–74 in % of its depth
vs. 76–83).
Description. General body features are illustrated in
. Morphometric and meristic characters of holotype and paratypes are summarised in
. A medium sized
Nothobranchius species (maximum observed size: 43.2 mm SL in males and 28.8 mm SL in females).
Males: General body shape is robust, laterally compressed, and deep. Greatest body depth at vertical in front of pelvic-fin origin. Greatest body width at pectoral-fin base, with body progressively narrowing towards caudal-fin base. Dorsal profile slightly concave to straight from tip of snout to nape and convex to base of last dorsal-fin ray, straight to slightly concave on caudal peduncle. Ventral profile convex from lower jaw to base of last anal-fin ray, straight to slightly concave on caudal peduncle. Caudal peduncle moderately shallow, length 1.3–1.4 times of its depth. Anus situated directly in front of anal-fin origin.
Head short, laterally compressed, deeper than wide. Snout slightly pointed, smaller than eye diameter. Mouth supraterminal, slightly oblique in profile. Jaws subequal, lower jaw longer than upper, posterior end of rictus at same level or slightly ventral to centre of eye. Premaxilla and dentary with many irregularly distributed conical, slightly curved teeth at the outer row of lower and upper jaws. Orbit large, almost entirely in the anterior half of the head, in the dorsal portion of the head side. Branchiostegal membrane projects posteriorly from the operculum.
Dorsal-fin origin anterior to anal-fin origin, with both fins originating posterior to mid-length of the body. Overall distal part of dorsal and anal fins rounded, with small contact organs in form of papillae on fin rays and distal margin with short filamentous rays. Posterior tip/margin of dorsal fin reaching caudal-fin base. Pectoral fin subtriangular, insertion at about vertical or slightly posterior to margin of opercular opening, base slightly oblique, with 15–16 rays, upper fin rays placed slightly anteriorly to lower fin rays, tip reaching or slightly overlapping base of pelvic fin. Pelvic fin subabdominal, origin at about mid-length of body, short, bases medially separated, tip reaching urogenital papilla. Caudal fin rounded, with 16–17 branched rays, plus 3 to 4 unbranched, smaller rays at dorsal and ventral origins.
Scales cycloid. Body and head entirely scaled, except for the ventral surface of the head.
Cephalic squamation pattern variable. Anterior nostril at the anterior tip of the snout, tubular opening lateral to the upper lip. Posterior nostril in front of the orbit, with oblique oval aperture. Frontal neuromast in shallow groove. Cephalic sensory system at supraorbital level in a continuous, slightly curved shallow groove, with five exposed neuromasts, whereas at supratemporal level in a curved groove, with four exposed neuromasts (
b). Preorbital canal in shallow groove with two exposed neuromasts; postorbital canal in shallow groove with one exposed neuromast; infraorbital series with about twelve small buttons at ventral margin of eye. Mandibular canal is in a shallow groove with a series of small neuromasts, curved to the lateral midline in front. One neuromast is on each scale along the trunk mid-longitudinal series.
Females: Smaller than males. Body generally similar but less laterally compressed and slightly more slender than in males (body depth at pelvic fin origin 27.1–28.6% SL
vs. 29.0–35.6; head depth 77–82% of its depth
vs. 68–74). Anal fin subtriangular, tip rounded, central rays longer and more rigid (
vs. anal fin rounded in males). Anal fin positioned more posteriorly than in males (62.3–65.6% SL
vs. 57.9–60.5). Dorsal-fin base length smaller than in males (22.2–23.8% SL
vs. 25.7–28.0). Branchiostegal membrane not projecting distally (
vs. projecting distally in males). No papillae or epidermal tissue present on dorsal and anal fins (
vs. both present in males).
.
Morphometric and meristic data of holotype and paratypes of Nothobranchius katemomandai. Holotype values included in ranges, mean and standard deviation of males. H, holotype; SD, standard deviation.
| Morphometric and Meristic Characters |
|
Males |
Females |
|
(n = 7) |
(n = 3) |
| H |
Range |
Mean |
SD |
Range |
Mean |
SD |
| Standard length |
31.1 |
30.3–43.2 |
|
|
27–28.8 |
|
|
| Percent of standard length |
|
|
|
|
|
|
|
|
Total length |
125.7 |
121.5–125.7 |
124.5 |
1.4 |
122.6–127.1 |
124.7 |
2.3 |
|
Body depth at pelvic-fin origin |
31.5 |
29.0–35.6 |
31.1 |
2.2 |
27.1–28.6 |
27.7 |
0.8 |
|
Head length |
32.2 |
28.9–32.2 |
30.6 |
1.3 |
29.6–32.2 |
30.6 |
1.4 |
|
Preanal length |
59.5 |
57.9–60.5 |
59.7 |
0.9 |
62.3–65.6 |
63.9 |
1.6 |
|
Predorsal length |
57.6 |
56.2–59.0 |
57.6 |
1.0 |
58.9–59.7 |
59.2 |
0.5 |
|
Prepelvic length |
47.9 |
43.5–48.8 |
46.5 |
1.8 |
50.4–51.0 |
50.7 |
0.3 |
|
Prepectoral length |
33.1 |
29.2–33.1 |
31.5 |
1.3 |
29.6–32.6 |
31.4 |
1.6 |
|
Caudal peduncle length |
20.3 |
18.3–20.6 |
19.5 |
0.8 |
17.4–20.1 |
18.7 |
1.4 |
|
Caudal peduncle depth |
15.1 |
13.4–15.1 |
14.3 |
0.6 |
12.6–14.2 |
13.5 |
0.8 |
|
Dorsal-fin base length |
28.0 |
25.7–28.0 |
27 |
0.8 |
22.2–23.8 |
23.2 |
0.9 |
|
Anal-fin base length |
21.9 |
21.0–22.9 |
21.8 |
0.6 |
17.7–21.5 |
19.4 |
1.9 |
|
Caudal-fin length |
25.7 |
21.5–25.7 |
24.5 |
1.4 |
22.6–27.1 |
24.7 |
2.3 |
| Percent of head length |
|
|
|
|
|
|
|
|
Head width |
58 |
58–63 |
59.6 |
1.5 |
58–61 |
59.5 |
1.6 |
|
Head depth |
84 |
80–88 |
84.7 |
2.6 |
75–76 |
75.6 |
0.6 |
|
Interorbital width |
43 |
40–46 |
42.8 |
1.9 |
40–44 |
41.4 |
2.1 |
|
Postorbital length |
51 |
49–51 |
50.0 |
1.0 |
49–52 |
50.7 |
1.8 |
|
Suborbital depth |
22 |
17–22 |
18.9 |
1.9 |
14–16 |
15.0 |
1.3 |
|
Eye diameter |
31 |
31–40 |
35.2 |
2.7 |
35–36 |
35.8 |
0.5 |
|
Snout to eye end length |
49 |
49–52 |
50.0 |
1.0 |
48–51 |
49.3 |
1.8 |
|
Snout length |
19 |
15–19 |
18.0 |
1.8 |
17–21 |
18.6 |
2.3 |
| Other morphometric ratios |
|
|
|
|
|
|
|
|
Head width in % of its depth |
69 |
68–74 |
70.4 |
2.5 |
77–82 |
78.7 |
2.6 |
|
Caudal peduncle length in % of its depth |
134 |
131–139 |
136.3 |
2.7 |
138–142 |
139.2 |
2.0 |
|
Predorsal length in % of preanal length |
97 |
96–98 |
96.5 |
0.8 |
90–95 |
92.7 |
2.5 |
| Meristics |
|
range |
mode |
|
range |
mode |
|
|
Dorsal-fin rays |
16 |
15–17 |
16 |
|
16–18 |
16 |
|
|
Anal-fin rays |
17 |
15–17 |
17 |
|
16–18 |
18 |
|
|
Scales mid-longitudinal series |
27 |
26–29 |
27 |
|
27–28 |
27 |
|
|
Scales transverse |
10 |
10 |
10 |
|
10 |
10 |
|
|
Scales circumpeduncular |
12 |
12 |
12 |
|
12 |
12 |
|
. <em>Nothobranchius katemomandai</em>: (<strong>a</strong>). BE_RMCA_VERT.2025.008.P.0011, holotype, preserved male, 31.1 mm SL; (<strong>b</strong>). live wild-caught male; (<strong>c</strong>). live wild-caught female; (<strong>d</strong>). type locality; DR Congo: Kay system: upper Lualaba drainage: shallow remnant pool in small seasonal riverbed, 0.5 km west of Lubule village, 07°31′47″ S, 27°13′24″ E, 634 m alt.; B. Katemo Manda, 17 April 2023 (field code: CD 23-26). Photographed by B. Nagy (<strong>a</strong>–<strong>c</strong>) and B. Katemo Manda (<strong>d</strong>).
Live colouration of males (
b–c): scales on head and trunk light blue with irregular red-brown posterior margin, forming irregular reticulation and oblique barred pattern on body. Snout, frontal and dorsal portion of head light grey, throat light blue to silver. Posterior scale margins on post-orbital portion of operculum forming two to three red-brown, anteriorly lowering, oblique bars. Exposed part of branchiostegal membrane white. Iris light yellow, with dark grey-black spots, creating a dark ring around the centre of the eye. Dorsal fin yellow to light blue with brown spots, forming in part irregular stipes perpendicular to fin rays, fin tips dark grey. Anal fin base light blue to yellow-blue, occasionally with irregular brown spots proximally, with narrow brown submedial band, followed by a slender yellow band, a slender red-brown band and a slender dark brown distal band. Caudal fin light blue, densely marked with brown spots proximally and medially, with slender white to light blue subdistal band and a narrow dark grey distal band. Pelvic fin transparent brown with yellow and light blue bands medially and distally, and with red-brown dots. Pectoral fin hyaline with blue posterior distal margin.
Live colouration of females: scales on trunk and head light brown with narrow dark brown posterior margin, forming reticulated pattern on body. Overall light brown colour of scales, darker on dorsum and lighter to silvery on venter. Blue iridescence on the opercular region and anterior midlateral half of the trunk. Iris yellow. All fins are hyaline.
Distribution. Nothobranchius katemomandai is endemic to seasonal freshwater habitats of the upper Congo drainage in south-eastern DR Congo. It is currently known from remnant pools in small ephemeral riverbeds associated with the Kay system in the upper Lualaba drainage (
). The Kay is a major right bank affluent entering the upper Lualaba below Lake Upemba.
Ecology and biology. The area of the upper Lualaba experiences a mean annual rainfall of around 1000 mm, with December the wettest month. In the Upemba System, the waters of the Lualaba are high from February-April and low from August to October [
83]. The dry season lasts for around 160 days.
At the type locality,
N.
katemomandai was the only
Nothobranchius species observed. The accompanying fish fauna consisted of the non-annual ‘
Lacustricola’. The type locality on 17 April 2023 was a remnant pool formed in the riverbed of the seasonal Lubule River (
d). The habitat was 10–15 m in diameter, about 1 m deep at its deepest point, and connected to the 1–2 m large shallow seasonal riverbed. The edges of both the pool and the ephemeral riverbed were overgrown by grass. The water was turbid. The water temperature around noon was 27.3 °C, the pH was 7.2, and it contained 120 ppm of total dissolved solids. The habitat belongs to category 1.4.1 as defined by Watters (2015a), representing a remnant pool in a seasonal stream system on a floodplain.
Water parameters, measured by B. Katemo Manda at five different sites in the Kay system, where the existence of
N. katemomandai has been proven, showed average values of: temperature of 28 °C; pH of 6.4; total dissolved solids of 36 ppm.
Aquarium maintenance of selected specimens was undertaken for observation of breeding behaviour and biology.
Nothobranchius katemomandai has an annual mode of reproduction, the same as all of its known congeners. Under captive conditions, peat moss was used successfully as an artificial spawning substrate. A plastic container with the substrate was placed in the aquarium and removed after a period of two weeks, and dried to a slightly moist state. An embryonic development period of four to five months was observed at about 22–24 °C. Eggs are slightly oval, measuring 1.21 mm long and 1.13 mm wide on average (
n = 10), with short adhesive filaments of up to 40 µm on the chorion, slightly attaching the eggs to the substrate.
Conservation status. Nothobranchius katemomandai is recommended to be assessed as Vulnerable. The species is currently known only from the area of the type locality, several sites in the Kay system of the Lualaba drainage. It might exist at some other sites within the drainage system of the upper Lualaba River, but its distribution will remain restricted, and any potential additional subpopulations are expected to be fragmented and not necessarily in contact with each other. The entire drainage area of the Kay system has an extent of 1142 km
2, and the species is expected to be found from a maximum of ten threat-based locations. Some of the known sites at present, including the type locality, are directly adjacent to human populations. There is an expansion of agriculture in seasonal wetland habitats of the known distribution area and, consequently, an increase in anthropogenic pressures on land and water use. Using IUCN (2012) principles [
86],
N.
katemomandai meets the criteria B1ab(iii) for Vulnerable, considering the drainage area as maximum EOO and the number of estimated maximum threat-based locations, as well as the risk of continuous decline of the known wetland habitats in which it is known to live.
There are currently no conservation or protection measures in place for this species. Additionally, it is unknown whether any protected areas would partially encompass its known distribution. Therefore, habitat protection is needed at the type locality and other sites within the Kay system where the species has been recorded. Further field surveys should target additional suitable habitats within the drainage area to document the full geographic range of the species more accurately. This would help in developing appropriate conservation strategies.
Etymology. The specific epithet is given in honour of friend Bauchet Katemo Manda, professor at the University of Lubumbashi, the discoverer of this species, for his dedication to the research of the ichthyofauna of the Democratic Republic of Congo. The exciting adventures and numerous challenges faced during joint research expeditions will always remain vivid memories.
3.3.3.
Nothobranchius marmoreus, New Species
http://zoobank.org/urn:lsid:zoobank.org:act: AE267C54-9106-4BCA-A793-3D205C5ECF42
Nothobranchius spec. ‘Mokobe’: van der Merwe et al. 2021: 10 [
36].
Nothobranchius spec. ‘Mukobe’: Nagy 2024b: 75 [
82].
Holotype. BE_RMCA_VERT.2025.008.P.0021, male, 36.3 mm SL; DR Congo: Lake Mweru basin: Lufukwe system: ephemeral pool in the floodplain of the seasonal river, 1.5 km west of Mukobe village, 09°41′32.9″ S, 28°12′42.5″ E, 962 m alt.; B. Nagy, A. Chocha Manda & A. Kalumba, 12 April 2023 [field code: CD 23-9].
Paratypes. BE_RMCA_VERT.2025.008.P.0022–0027, 3 males, 34.2–41.2 mm SL & 3 females, 31.4–33.9 mm SL; collected with the holotype. — BE_RMCA_VERT.2025.008.P.0028–0030, 2 males, 35.1–36.7 mm SL & 1 female, 31.7 mm SL; same locality; B. Nagy, A. Chocha Manda & G. Petneházy, 10 April 2016 [field code: CD 16-12].
Diagnosis. Nothobranchius marmoreus is distinguished from all other species of the genus by the unique combination in male colouration of having a body with irregular red-brown patches and stripes, forming a marble-like mottled pattern; and anal and caudal fins with slender yellow to amber subdistal band and broad dark brown distal band.
Further,
N. marmoreus is distinguished from most closely related
N. ditte by smaller head depth (80–84% HL
vs. 87–94); and greater caudal peduncle length (152–159 in % of its depth
vs. 130–140); from
N. malaissei by smaller head length (25.5–30.9% SL
vs. 32.7–35.4); and greater caudal peduncle length (152–159 in % of its depth
vs. 124–131); and from
N. milvertzi by smaller prepelvic length (45.5–46.7% SL
vs. 46.9–50.8); and smaller head width (65.5–72.9 in % of its depth
vs. 73.5–82.6).
Description. General body features are illustrated in
. Morphometric and meristic characters of holotype and paratypes are summarised in
. A medium sized
Nothobranchius species (maximum observed size: 41.2 mm SL in males and 33.9 mm SL in females).
Males: General body shape is robust, laterally compressed, and deep. Greatest body depth at vertical in front of pelvic-fin origin. Greatest body width at pectoral-fin base, with body progressively narrowing towards caudal-fin base. Dorsal profile slightly concave to straight from tip of snout to nape and convex to base of last dorsal-fin ray, straight to slightly concave on caudal peduncle. Ventral profile convex from lower jaw to base of last anal-fin ray, straight to slightly concave on caudal peduncle. Caudal peduncle shallow, length 1.5–1.6 times its depth. Anus situated directly in front of anal-fin origin.
. <em>Nothobranchius marmoreus</em>: (<strong>a</strong>). BE_RMCA_VERT.2025.008.P.0021, holotype, preserved male, 36.3 mm SL; (<strong>b</strong>). live wild-caught male; (<strong>c</strong>). live wild-caught female; (<strong>d</strong>). type locality; DR Congo: Lake Mweru basin: Lufukwe system: ephemeral pool in the floodplain of the seasonal river, 1.5 km west of Mukobe village, 09°41′32.9″ S, 28°12′42.5″ E, 962 m alt.; B. Nagy, A. Chocha Manda & A. Kalumba, 12 April 2023 [field code: CD 23-9]. Photographed by B. Nagy.
.
Morphometric and meristic data of holotype and paratypes of Nothobranchius marmoreus. Holotype values included in ranges, mean and standard deviation of males. H, holotype; SD, standard deviation.
| Morphometric and Meristic Characters |
|
Males |
Females |
|
(n = 6) |
(n = 4) |
| H |
Range |
Mean |
SD |
Range |
Mean |
SD |
| Standard length |
36.3 |
32.6–41.2 |
|
|
31.4–33.9 |
|
|
| Percent of standard length |
|
|
|
|
|
|
|
|
Total length |
125.6 |
120.2–125.6 |
123.2 |
1.8 |
119.2–124.5 |
122.4 |
2.4 |
|
Body depth at pelvic–fin origin |
30.0 |
26.7–32.0 |
28.3 |
2.2 |
24.5–27.4 |
25.5 |
1.3 |
|
Head length |
30.9 |
25.5–30.9 |
29.4 |
2.0 |
28.7–32.5 |
31.0 |
1.6 |
|
Preanal length |
56.5 |
56.1–59.1 |
57.3 |
1.0 |
59.6–64.4 |
62.1 |
2.0 |
|
Predorsal length |
55.9 |
54.2–58.2 |
56.2 |
1.4 |
56.7–61.9 |
59.0 |
2.2 |
|
Prepelvic length |
45.5 |
45.5–46.7 |
46.1 |
0.5 |
44.2–50.2 |
47.3 |
2.7 |
|
Prepectoral length |
31.1 |
25.7–31.1 |
29.7 |
2.1 |
28.7–33.7 |
31.8 |
2.2 |
|
Caudal peduncle length |
19.3 |
19.3–20.9 |
20.0 |
0.6 |
19.8–22.3 |
20.7 |
1.2 |
|
Caudal peduncle depth |
12.1 |
12.1–13.6 |
12.8 |
0.5 |
10.3–11.8 |
11.0 |
0.6 |
|
Dorsal–fin base length |
26.4 |
23.9–29.9 |
26.5 |
2.5 |
21.7–23.9 |
23.1 |
1.0 |
|
Anal–fin base length |
22.9 |
21.6–24.0 |
22.7 |
1.0 |
18.5–19.5 |
19.0 |
0.4 |
|
Caudal–fin length |
25.6 |
20.2–25.6 |
23.2 |
1.8 |
19.2–24.5 |
22.4 |
2.4 |
| Percent of head length |
|
|
|
|
|
|
|
|
Head width |
56 |
55–59 |
56.4 |
1.5 |
53–58 |
54.7 |
2.2 |
|
Head depth |
83 |
80–84 |
82.2 |
1.3 |
68–72 |
70.2 |
1.8 |
|
Interorbital width |
42 |
41–45 |
43.4 |
1.9 |
40–47 |
42.5 |
2.9 |
|
Postorbital length |
49 |
49–56 |
51.5 |
2.7 |
51–56 |
53.5 |
2.7 |
|
Suborbital depth |
14 |
14–17 |
15.8 |
1.3 |
12–17 |
14.4 |
2.2 |
|
Eye diameter |
38 |
33–38 |
35.0 |
2.0 |
31–33 |
32.2 |
0.6 |
|
Snout to eye end length |
51 |
44–51 |
48.5 |
2.7 |
44–49 |
46.5 |
2.7 |
|
Snout length |
17 |
15–20 |
17.4 |
1.9 |
13–19 |
16.3 |
2.6 |
| Other morphometric ratios |
|
|
|
|
|
|
|
|
Head width in % of its depth |
68 |
66–73 |
68.7 |
2.7 |
76–81 |
78.0 |
2.7 |
|
Caudal peduncle length in % of its depth |
159 |
152–159 |
155.5 |
2.8 |
185–191 |
188.7 |
2.5 |
|
Predorsal length in % of preanal length |
99 |
97–100 |
98.1 |
1.3 |
94–96 |
95.0 |
0.9 |
| Meristics |
|
range |
mode |
|
range |
mode |
|
|
Dorsal–fin rays |
17 |
15–17 |
17 |
|
16–17 |
17 |
|
|
Anal–fin rays |
17 |
16–19 |
17 |
|
17–19 |
17 |
|
|
Scales mid–longitudinal series |
29 |
28–30 |
29 |
|
28–29 |
28 |
|
|
Scales transverse |
11 |
10–11 |
11 |
|
10–11 |
10 |
|
|
Scales circumpeduncular |
12 |
12–14 |
12 |
|
10 |
10 |
|
Head short, laterally compressed, deeper than wide. Snout slightly pointed, smaller than eye diameter. Mouth supraterminal, slightly oblique in profile. Jaws subequal, lower jaw longer than upper, posterior end of rictus at same level or slightly ventral to centre of eye. Premaxilla and dentary with many irregularly distributed conical, slightly curved teeth in the outer row of lower and upper jaws. Orbit large, almost entirely in the anterior half of the head, in the dorsal portion of the head side. Branchiostegal membrane projects posteriorly from the operculum.
Dorsal-fin origin anterior to anal-fin origin, with both fins originating posterior to mid-length of body. Overall distal part of dorsal and anal fins rounded, with small contact organs in form of papillae on fin rays and distal margin with short filamentous rays. Posterior tip/margin of dorsal fin reaching caudal-fin base. Pectoral fin subtriangular, insertion at about vertical or slightly posterior to margin of opercular opening, base slightly oblique, with 18–19 rays, upper fin rays placed slightly anteriorly to lower fin rays, tip reaching or slightly overlapping base of pelvic fin. Pelvic fin subabdominal, origin at about mid-length of body, short, bases medially separated, tip reaching urogenital papilla. Caudal fin rounded, with 17–18 branched rays, plus 3 to 4 unbranched smaller rays at dorsal and ventral origins.
Scales cycloid. Body and head entirely scaled, except for the ventral surface of the head.
Cephalic squamation pattern variable. Anterior nostril at the anterior tip of the snout, tubular opening lateral to the upper lip. Posterior nostril in front of the orbit, with oblique oval aperture. Frontal neuromast in shallow groove. Cephalic sensory system at supraorbital level in a shallow groove fragmented into two portions, with two and three exposed neuromasts, respectively, whereas at the supratemporal level, it is in a curved groove, with four exposed neuromasts (
c). Preorbital canal in shallow groove with three to four exposed neuromasts; postorbital canal in shallow groove with one exposed neuromast; infraorbital series with about twelve small buttons at ventral margin of eye. Mandibular canal is in a shallow groove with a series of small neuromasts, curved to the lateral midline in front. One neuromast is located on each scale along the mid-longitudinal series of the trunk.
Females: Smaller than males. Body generally similar but somewhat less laterally compressed and slightly more slender than in males: head width greater (76–81% of its depth
vs. 66–73), caudal peduncle more slender 185–191% of its depth
vs. 152–159. Anal fin subtriangular, tip rounded, central rays longer and more rigid (
vs. anal fin rounded in males). Dorsal-fin and anal-fin base lengths are smaller than in males (21.7–23.9% SL
vs. 24.0–29.9, 18.5–19.5% SL
vs. 21.6–24.0, respectively). Branchiostegal membrane not projecting distally (
vs. projecting distally in males). No papillae or epidermal tissue present on dorsal and anal fins (
vs. both present in males).
Colouration. Live colouration of males (
b): scales on head and trunk light green-blue to silver with irregular red-brown posterior margin, forming irregular marble-like patches and striped pattern on body. Snout, frontal and dorsal portion of head red-brown, throat light green-blue to silver. Posterior scale margins on post-orbital portion of operculum creating three red-brown, oblique, and vertical bars. Exposed part of branchiostegal membrane silver. Iris light yellow, with dark grey-black spots, especially on upper and lower-most parts, creating a dark vertical bar through the centre of the eye. Dorsal fin yellow to light green-blue with irregular red-brown stripes, perpendicular to fin rays proximally, parallel to fin rays medially and distally, fin tips with narrow dark brown distal band. Anal fin yellow to light green-blue, with irregular red-brown spots proximally, followed by a slender red-brown submedial band, a yellow to amber medial band and a broad dark brown distal band. Caudal fin base yellow to light green-blue, with irregular red-brown spots proximally, merging into a broad red-brown band medially, followed by an irregular narrow yellow to amber subdistal band and a broad dark brown distal band. Pelvic fin yellow to light green-blue, with red-brown proximal band and dark brown distal band. Pectoral fin hyaline with blue posterior distal margin.
Live colouration of females (
c): scales on trunk and head light brown with narrow dark grey posterior margin, forming a slightly visible reticulated pattern on body. Overall light brown colour of scales darker on dorsum and lighter to silvery on venter. Blue iridescence on the opercular region and on the trunk. Iris yellow. All fins are hyaline.
Distribution. Nothobranchius marmoreus is endemic to seasonal freshwater habitats of the upper Congo drainage in south-eastern DR Congo. It is currently known only from the type locality, a remnant pool associated with the Lufukwe system in the Lake Mweru basin (
). The Lufukwe is an affluent entering the lake from a south-western direction.
Ecology and biology. The area of the southern Lake Mweru basin experiences a mean annual rainfall of 1020–1120 mm [
83]. The waters are the highest during the rainy season and, from March to May and driest between September and January [
83,
84].
At the type locality,
N.
marmoreus was the only
Nothobranchius species observed. The type locality on 12 April 2023 was a large ephemeral pool formed in the floodplain of the seasonal Lufukwe River (
d). The habitat was 25–30 m in diameter, about 1.5 m deep at its deepest point. Shallower parts and edges of the pool were overgrown by grass. The water was turbid. The water temperature late morning was 24.0 °C, the pH was 7.4, and it contained 35 ppm of total dissolved solids. Water parameters, measured at the same location during the previous visit of 10 April 2016, showed the following values of: temperature of 27.8 °C in mid-afternoon; pH of 7.36; total dissolved solids of 42 ppm. The habitat primarily belongs to category 1.2 as defined by Watters (2015a) [
85], representing a pool and flooded grassy area on a floodplain. However, the habitat also shows the influence of human activity: the pool lies where the road crosses the seasonal floodplain, and the natural pool has been excavated to make it larger and prevent flooding on the road. Therefore, the habitat falls into the secondary category of 2.1, modified by human activities related to road construction.
Aquarium maintenance of selected specimens was undertaken for observation of breeding behaviour and biology.
Nothobranchius marmoreus has an annual mode of reproduction, the same as all of its known congeners. Under captive conditions, peat moss has been successfully used as an artificial spawning substrate. A plastic container with the substrate was placed in the aquarium and removed after a period of two weeks. It was then dried to a slightly moist state. An embryonic development period of three to four months was observed at about 22–24 °C. Eggs are slightly oval, measuring 1.26 mm long and 1.21 mm wide on average (
n = 10), with short adhesive filaments of 40–90 µm on chorion, slightly attaching the eggs to the substrate.
Conservation status. Nothobranchius marmoreus is recommended to be assessed as Endangered. The species is currently known only from the type locality, a site situated in the Lufukwe system in the Lake Mweru basin. It might exist at some other sites within the drainage system of the Lufukwe, but its distribution will remain restricted, and any potential additional subpopulations are expected to be fragmented and have little or no contact with each other. The species is expected to be confined to ephemeral wetlands within the Lufukwe system, which has a drainage area of 559 km
2, representing the maximum estimated extent of occurrence. Its area of occupancy is estimated to be less than 100 km
2, with a likely maximum of five threat-based locations. The type locality, currently the only known site for the species, is located in close proximity to a human population. Phases in the seasonal life cycle of this species underscore the vulnerabilities of the ecological processes in the ephemeral habitats, as the survival of the species is dependent on suitable conditions during both dry and wet seasons. The author observed the expansion of agriculture in the general area and, consequently, increasing anthropogenic pressures on the land and water use. The resultant habitat changes are likely to modify the habitats in ways that render them unsuitable and thus degraded when considering their support of the seasonal life cycle of the species, and thus represent an important extinction risk. Further, the natural habitat at the type locality has been altered by the local human population through excavation to retain water for longer periods. This modification affects the natural rhythm and duration of the dry and wet seasons in the biotope. Additionally, specimens collected from the type locality during a first visit in 2016 have exhibited clinical signs, such as lesions suggestive of infection with
Aphanomyces spp, and the fish were in a generally weak condition, with mortality within a few hours after capture. Using IUCN (2012) principles [
86],
N.
marmoreus meets the criteria B1ab(iii)+2ab(iii) for Endangered, considering upper estimates of the EOO, AOO, and number of locations, as well as the risk of continuous decline of the known wetland habitats in which it is known to live.
There is currently no conservation or protection measure in place for this species. Additionally, its only known locality is not part of any protected area. Therefore, habitat protection is needed at the type locality. Further field surveys should target additional suitable habitats within the drainage area to document the full geographic range of the species more accurately. This would help in developing appropriate conservation strategies.
Etymology. The specific epithet
marmoreus is a Latin adjective deriving from the ancient Greek word marmaros (μάρμαρος) in the meaning of resembling marble, marblelike, and in reference to the irregularly mottled marbled colour pattern on the body of the males.
3.3.4.
Nothobranchius dubieensis, New Species
http://zoobank.org/urn:lsid:zoobank.org:act: DF33ED09-C5E6-499B-A478-7BEFC2732115
Holotype. BE_RMCA_VERT.2025.008.P.0031, male, 40.6 mm SL; DR Congo: Tambe stream: Lubule system: Luvua drainage: shallow remnant pool in small seasonal riverbed, 5.5 km west of Dubie village, 08°35′31″ S, 28°28′51″ E, 987 m alt.; A. Kalumba, 20 May 2023 (field code: CD 23-29).
Paratypes. BE_RMCA_VERT.2025.008.P.0032–0038, 4 males, 35.2–41.4 mm SL & 3 females, 29.5–37.2 mm SL; collected with the holotype.
Diagnosis. Nothobranchius dubieensis is distinguished from all other species of the genus by the unique combination in male colouration of having anal fin with narrow dark brown submedial band, narrow yellow and orange medial bands, narrow white subdistal band, and slender dark brown distal band; and caudal fin with irregular red-brown spots and stripes proximally and medially, followed by irregular narrow red-brown subdistal band and slender white distal band, with interrupted red-brown fin tips.
Further,
N. dubieensis is distinguished from the most closely related
N. oestergaardi by greater body depth (31.9–35.5% SL
vs. 28.0–31.5); smaller snout length (14–16% HL 21–31); and smaller caudal peduncle length (119–125 in % of its depth
vs. 125–140).
Description. General body features are illustrated in
. Morphometric and meristic characters of holotype and paratypes are summarised in
. A medium sized
Nothobranchius species (maximum observed size: 41.4 mm SL in males and 37.2 mm SL in females).
. <em>Nothobranchius dubieensis</em>: (<strong>a</strong>). BE_RMCA_VERT.2025.008.P.0031, holotype, preserved male, 36.3 mm SL; (<strong>b</strong>,<strong>c</strong>). wild male photographed after capture; (<strong>d</strong>). type locality; DR Congo: Tambe stream: Lubule system: Luvua drainage: shallow remnant pool in small seasonal riverbed, 5.5 km west of Dubie village, 08°35′31″ S, 28°28′51″ E, 987 m alt.; A. Kalumba, 20 May 2023 (field code: CD 23-29). Photographed by B. Nagy (<strong>a</strong>) and A. Kalumba (<strong>b</strong>–<strong>d</strong>).
Males: General body shape is robust, laterally compressed, and deep. Greatest body depth at vertical in front of pelvic-fin origin. Greatest body width at pectoral-fin base, with body progressively narrowing towards caudal-fin base. Dorsal profile slightly concave to straight from tip of snout to nape and convex to base of last dorsal-fin ray, straight to slightly concave on caudal peduncle. Ventral profile convex from lower jaw to base of last anal-fin ray, straight to slightly concave on caudal peduncle. Caudal peduncle shallow, length 1.2–1.3 times its depth. Anus situated directly in front of anal-fin origin.
Head short, laterally compressed, deeper than wide. Snout slightly pointed, smaller than eye diameter. Mouth supraterminal, slightly oblique in profile. Jaws subequal, lower jaw longer than upper, posterior end of rictus at same level or slightly ventral to centre of eye. Premaxilla and dentary with many irregularly distributed conical, slightly curved teeth in the outer row of lower and upper jaws. Orbit large, in the anterior half of the head, in the dorsal portion of the head side. Branchiostegal membrane projects posteriorly from operculum.
.
Morphometric and meristic data of holotype and paratypes of Nothobranchius dubieensis. Holotype values included in ranges, mean and standard deviation of males. H, holotype; SD, standard deviation.
| Morphometric and Meristic Characters |
|
Males |
Females |
|
(n = 5) |
(n = 3) |
| H |
Range |
Mean |
SD |
Range |
Mean |
SD |
| Standard length |
40.6 |
35.2–41.4 |
|
|
29.5–37.2 |
|
|
| Percent of standard length |
|
|
|
|
|
|
|
|
Total length |
123.2 |
120.5–125.4 |
123.4 |
1.9 |
122.8–128.5 |
125.3 |
2.9 |
|
Body depth at pelvic-fin origin |
32.5 |
31.9–35.5 |
33.5 |
1.7 |
27.9–29.8 |
28.6 |
1.0 |
|
Head length |
31.8 |
30.9–32.4 |
31.7 |
0.7 |
29.2–30.4 |
29.8 |
0.6 |
|
Preanal length |
65.0 |
59.7–65.6 |
63.4 |
2.3 |
65.3–67.8 |
66.6 |
1.2 |
|
Predorsal length |
61.1 |
58.5–64.5 |
61.3 |
2.2 |
62.1–65.8 |
64.3 |
2.0 |
|
Prepelvic length |
48.3 |
47.0–51.0 |
49.1 |
1.5 |
50.8–53.0 |
51.9 |
1.1 |
|
Prepectoral length |
34.2 |
31.9–34.2 |
32.9 |
1.0 |
29.8–31.5 |
30.5 |
0.8 |
|
Caudal peduncle length |
18.2 |
17.1–19.1 |
18.2 |
0.7 |
17.3–19.1 |
18.3 |
0.9 |
|
Caudal peduncle depth |
15.3 |
13.7–16.1 |
15.0 |
0.9 |
11.9–12.6 |
12.3 |
0.4 |
|
Dorsal-fin base length |
23.6 |
23.6–28.4 |
25.9 |
2.0 |
18.4–23.7 |
21.6 |
2.8 |
|
Anal-fin base length |
17.5 |
17.5–23.7 |
19.8 |
2.4 |
15.0–16.4 |
15.8 |
0.7 |
|
Caudal-fin length |
23.2 |
20.5–25.4 |
23.4 |
1.9 |
22.8–28.5 |
25.3 |
2.9 |
| Percent of head length |
|
|
|
|
|
|
|
|
Head width |
62 |
62–66 |
63.3 |
1.5 |
57–63 |
60.5 |
2.9 |
|
Head depth |
95 |
90–96 |
92.6 |
2.4 |
76–78 |
77.2 |
1.1 |
|
Interorbital width |
43 |
43–48 |
46.2 |
1.8 |
44–48 |
45.8 |
1.7 |
|
Postorbital length |
58 |
56–59 |
57.3 |
1.1 |
56–60 |
57.3 |
2.5 |
|
Suborbital depth |
16 |
13–18 |
15.8 |
1.9 |
11–15 |
12.9 |
2.1 |
|
Eye diameter |
28 |
28–34 |
30.2 |
2.2 |
30–35 |
31.9 |
3.0 |
|
Snout to eye end length |
42 |
42–44 |
42.7 |
1.1 |
40–44 |
42.7 |
2.5 |
|
Snout length |
16 |
14–16 |
14.8 |
0.6 |
11–15 |
12.6 |
2.1 |
| Other morphometric ratios |
|
|
|
|
|
|
|
|
Head width in % of its depth |
66 |
66–71 |
68.3 |
1.9 |
75–81 |
78.4 |
3.0 |
|
Caudal peduncle length in % of its depth |
119 |
119–125 |
121.6 |
3.0 |
146–151 |
147.8 |
2.9 |
|
Predorsal length in % of preanal length |
94 |
94–98 |
96.8 |
1.8 |
95–98 |
96.7 |
1.5 |
| Meristics |
|
range |
mode |
|
range |
mode |
|
|
Dorsal-fin rays |
15 |
15–16 |
15 |
|
15–16 |
16 |
|
|
Anal-fin rays |
16 |
16–17 |
16 |
|
16–17 |
16 |
|
|
Scales mid-longitudinal series |
30 |
28–30 |
29 |
|
28–29 |
29 |
|
|
Scales transverse |
10 |
10–11 |
11 |
|
11 |
11 |
|
|
Scales circumpeduncular |
10 |
10–12 |
12 |
|
12 |
12 |
|
Dorsal-fin origin anterior to anal-fin origin, with both fins originating posterior to mid-length of the body. Overall distal part of dorsal and anal fins rounded, with small contact organs in form of papillae on fin rays and distal margin with short filamentous rays. Posterior tip/margin of dorsal fin reaching caudal-fin base. Pectoral fin subtriangular, insertion at about vertical or slightly posterior to margin of opercular opening, base slightly oblique, with 18–19 rays, upper fin rays placed slightly anteriorly to lower fin rays, tip reaching or slightly overlapping base of pelvic fin.
Pelvic fin subabdominal, origin at about mid-length of body, short, bases medially separated, tip reaching urogenital papilla. Caudal fin rounded, with 17–18 branched rays, plus 3 to 4 unbranched smaller rays at dorsal and ventral origins.
Scales cycloid. Body and head entirely scaled, except for the ventral surface of the head.
Cephalic squamation pattern variable. Anterior nostril at the anterior tip of the snout, tubular opening lateral to the upper lip. Posterior nostril in front of the orbit, with oblique oval aperture. Frontal neuromast in shallow groove. Cephalic sensory system at supraorbital level in a shallow groove fragmented into two portions, with two and three exposed neuromasts, respectively, whereas at supratemporal level in a curved groove, with four exposed neuromasts (
d). Preorbital canal in shallow groove with two to three exposed neuromasts; postorbital canal in shallow groove with one exposed neuromast; infraorbital series not visible on most specimens. Mandibular canal in a shallow groove with a series of about a dozen small buttons, curved to the lateral midline in front. One neuromast is on each scale along the trunk mid-longitudinal series.
Females: Smaller than males. Body generally similar but less laterally compressed and slightly more slender than in males (body depth at pelvic fin origin 27.9–29.8% SL
vs. 31.9–35.5 in males; head width 75–81% of its depth
vs. 66–71; caudal peduncle length 146–151% of its depth
vs. 119–125). Head length is smaller than in males, with 29.2–30.4% SL
vs. 30.9–32.4. Anal fin subtriangular, tip rounded, central rays longer and more rigid (
vs. anal fin rounded in males). Anal-fin base length smaller than in males (15.0–16.4% SL
vs. 17.5–23.7). Branchiostegal membrane not projecting distally (
vs. projecting distally in males). No papillae or epidermal tissue present on dorsal and anal fins (
vs. both present in males).
Colouration. Live colouration of males (
c): scales on head and trunk light blue with red-brown posterior margin, forming irregular vertical, curved line pattern on body. Snout, frontal and dorsal portion of head red-brown to grey, throat light grey. Posterior scale margins on post-orbital portion of operculum create three red-brown, anteriorly lowering, oblique bars. Exposed part of branchiostegal membrane light blue. Iris light yellow, with dark grey-black spots, especially on upper and lower-most parts, creating a dark vertical bar through centre of eye. Dorsal fin light blue to light yellow with irregular red-brown stripes, perpendicular to fin rays proximally, parallel to fin rays medially and distally. Anal fin light blue, with irregular red-brown spots proximally, followed by narrow dark brown submedial band, narrow yellow and narrow orange medial bands, narrow white subdistal band, and slender dark brown distal band. Caudal fin light blue to yellow, with irregular red-brown spots and stripes proximally and medially, followed by irregular narrow red-brown subdistal band and slender white distal band, with interrupted red-brown fin tips. Pelvic fin with yellow and orange bands. Pectoral fin hyaline with blue posterior distal margin.
Females: scales on trunk and head light brown with narrow dark brown posterior margin, forming a slightly visible reticulated pattern on the body. Overall light brown colour of scales, darker on dorsum and lighter to silvery on venter. Blue iridescence on the opercular region and anterior midlateral half of the trunk. Iris yellow. All fins are hyaline.
Distribution. Nothobranchius dubieensis is endemic to seasonal freshwater habitats of the upper Congo drainage in south-eastern DR Congo. It is currently known from ephemeral marshes and ponds associated with the Lubule system in the Luvua drainage (
). The Lubule is a left affluent entering the Luvua from the southern direction.
Ecology. The area around the Lake Mweru basin experiences a mean annual rainfall of 1020–1120 mm [
83]. The waters are the highest during the rainy season and this from March to May, and driest between September and January [
83,
84].
At the type locality,
N.
dubieensis was the only
Nothobranchius species observed. The accompanying fish fauna consisted of non-annual ‘
Lacustricola’ species. The type locality on 20 May 2023 was a remnant, culvert-like pool, connecting to an ephemeral riverbed and a large swampy area, as part of the Tambe stream in the seasonal Lubule system (
d). The habitat was about 10 m long and 1 m wide, and about 50 cm deep at its deepest point. The edges of the biotope were overgrown by grass. The water was turbid, but the bottom was visible in shallower, undisturbed parts. The habitat principally belongs to category 1.2 as defined by Watters (2015a) [
85], representing a pool and flooded grassy area on a floodplain. The habitat is close to human population, and the anthropogenic impact is evident, as there are fish trap constructions in the biotope and trash left behind in the biotope.
Conservation status. Nothobranchius dubieensis is recommended to be assessed as Vulnerable. The species is currently known only from the area of the type locality, several sites in the Lubule system of the Luvua drainage. It might exist at some other sites within the drainage system of the Luvua, but its distribution will remain restricted, and any potential additional subpopulations are expected to be fragmented and not necessarily in contact with each other. The area of the Lubule system where the species is found measures about 3600 km
2, which is expected to be the maximum extent of occurrence. It is expected to inhabit a maximum of ten threat-based locations. Presently known sites, including the type locality, are directly adjacent to human populations. There is an expansion of agricultural seasonal wetland habitats in the known distribution area, and consequently, an increase in anthropogenic pressures on land and water use. Using IUCN (2012) principles [
86],
N.
dubieensis meets the criteria B1ab(iii) for Vulnerable, considering upper estimates of the EOO and number of threat-based locations, as well as the risk of continuous decline of the known wetland habitats in which it is known to live.
There are currently no conservation or protection measures in place for this species. Additionally, it is not known if any protected areas would partly encompass its known distribution. Therefore, habitat protection is needed at the type locality and other sites within the Lubule system where the species has been recorded. Further field surveys should target additional suitable habitats within the drainage area to document the full geographic range of the species more accurately. This would help in developing appropriate conservation strategies.
Etymology. The specific name is given in reference to Dubie township in south-eastern DR Congo, near which place this species is found, and the type locality is situated. An adjective derived from the geographical name.
3.3.5.
‘Lacustricola’ gemma, New Species
http://zoobank.org/urn:lsid:zoobank.org:act: 5C225ABD-CE65-4450-B8F8-041415B5A95E
Holotype. BE_RMCA_VERT.2025.008.P.0039, male, 22.5 mm SL; DR Congo: Kay system: upper Lualaba drainage: shallow remnant pool in small seasonal riverbed, 0.5 km west of Lubule village, 07°31′47″ S, 27°13′24″ E; B. Katemo Manda, 17 April 2023 (field code: CD 23-26).
Paratypes. BE_RMCA_VERT.2025.008.P.0040–0043, male, 23.0 mm SL & 3 females, 20.7–22.4 mm SL; collected with the holotype. — BE_RMCA_VERT.2025.008.P.0044, male, 16.8 mm SL; DR Congo: Kay system: upper Lualaba drainage: Lake Bowe, 07°40′53″ S, 27°08′46″ E; B. Katemo Manda, 17 December 2022.
Diagnosis. ‘Lacustricola’ gemma is distinguished from all other species of the genus by the unique combination in live colouration: in males, a pattern of iridescent, diamond-shaped, light blue spots in scale centres, especially evident below mid-longitudinal series on posteroventral portion of flank, and larger iridescent blue blotches on dorsum, creating an irregular reflective pattern on body; median fins yellow to hyaline, dorsal and anal fins with six to eight dark grey stripes perpendicular to fin rays basally and posteriorly, with light blue iridescence basally; dorsal with light blue distinct margin; anal with dark grey margin; caudal fin with four to five dark grey stripes perpendicular to fin rays; as well as in females, dorsal and anal fins hyaline with irregular brown stripes, fin tips with narrow light blue margin, caudal hyaline without markings.
Further,
‘Lacustricola’ gemma is distinguished from both ‘
L.’
hutereaui and ‘
L.’
chobensis by the following morphometric characters in males: body depth at pelvic-fin origin 25.6–27.4 (
vs. 21.1–24.1 and 30.0–32.3, respectively); anal-fin base length 18.7–19.6 (
vs. 14.2–17.1 and 20.7–25.2); smaller head depth 79–83 (
vs. 86–91 and 86–90); greater head width in % of its depth 83–87 (
vs. 76–83 and 70–74); and caudal peduncle length in % of its depth 111–114 (
vs. 150–156 and 98–105).
Description. General body features are illustrated in
. Morphometric and meristic characters of holotype and paratypes are summarised in
.
Males: general body shape laterally compressed and moderately deep. Small species, maximum observed size 23.0 mm SL. Greatest vertical body depth in front of pelvic-fin origin and shallowest at mid-portion of caudal peduncle. Greatest body width at pectoral-fin base, with body progressively narrowing towards caudal-fin base. Dorsal profile slightly convex from snout to base of dorsal fin and straight to slightly concave from base of dorsal fin to caudal fin. Ventral profile convex from lower jaw to base of last anal-fin ray, straight to slightly concave on ventral midline of caudal peduncle. Caudal peduncle relatively deep, length 111–114 in % of its depth. Anus directly in front of anal-fin origin.
Head short, laterally compressed, deeper than wide (head width 83–87 in % of its depth). Snout rounded, smaller than eye diameter. Mouth superior, oblique in profile. Jaws not equal, lower jaw longer than upper, posterior end of corner of mouth at the same level as the centre of eye. Premaxilla and dentary with many irregularly distributed conical teeth. Orbit large (38–41% HL).
Dorsal fin set rearwards, origin posterior to anal-fin origin, both fins originating posterior to mid-length of body. Dorsal and anal fins rounded. Dorsal fin, 9; anal fin, 13–14. Pectoral fin subtriangular, insertion relatively high and posterior to margin of opercular opening; base oblique, upper fin rays placed anteriorly to lower fin rays, 10–11 rays. Pelvic fin sub-abdominal; its origin slightly posterior to mid-length between insertions of pectoral and anal fins, almost reaching origin of anal fin. Caudal fin large and truncate, with 14–15 branched rays, plus five or six dorsal and ventral procurrent rays.
Scales cycloid, body and head entirely scaled, except for the ventral surface of the head. Scales in mid-longitudinal series, 20–23, plus two or three small scales on caudal-fin base. Transverse rows of scales in front of dorsal-fin origin, 6; scale rows around caudal peduncle, 10.
. ‘<i>Lacustricola</i>’ gemma: (<b>a</b>). BE_RMCA_VERT.2025.008.P.0039, holotype, preserved male, 22.5 mm SL; (<b>b</b>). holotype live male; (<b>c</b>). live wild-caught female; (<b>d</b>). type locality; DR Congo: Kay system: upper Lualaba drainage: shallow remnant pool in small seasonal riverbed, 0.5 km west of Lubule village, 07°31′47″ S, 27°13′24″ E, 634 m alt.; B. Katemo Manda, 17 April 2023 (field code: CD 23-26). Photographed by B. Nagy (<b>a</b>–<b>c</b>) and B. Katemo Manda (<b>d</b>).
Two frontal neuromasts in a shallow groove. Cephalic sensory system at preorbital level in two discontinuous shallow grooves, with one and two neuromasts, respectively; infraorbital series with about a dozen small buttons; postorbital canal tubular with two pores; preopercular sensory systems tubular with seven pores (
a). Cephalic sensory system at supraorbital level in a short groove, with two neuromasts; whereas at supratemporal level, two to three exposed neuromasts (
b). Mandibular level with three to four exposed neuromasts parallel to the outer margin of lower jaw (
c).
Females: Body depth smaller than in males (27.6–29.5% SL
vs. 25.6–27.4; head less laterally compressed than in males (head width 87–91% of its depth
vs. 83–87); preanal, predorsal and prepelvic lengths greater than in male (63.8–64.7% SL
vs. 59.1–62.2; 70.0–71.5% SL
vs. 66.1–69.3; and 44.7–48.8% SL
vs. 41.7–44.0); anal-fin base length smaller than in male (15.5–16.6% SL
vs. 18.7–19.6); and caudal peduncle shallower than in males (caudal peduncle depth 15.2–16.4% SL
vs. 16.7–17.4; caudal peduncle length 120–124% of its depth
vs. 111–114).
. Diagrammatic representation of the cephalic sensory system in ‘<i>Lacustricola</i>’ <i>gemma</i>; BE_RMCA_VERT.2025.008.P.0039, holotype, male, 22.5 mm SL; (<b>a</b>). lateral view of head; (<b>b</b>). dorsal view of head; (<b>c</b>). ventral view of head.
.
Morphometric and meristic data of holotype and paratypes of ‘Lacustricola’ gemma. Holotype values included in ranges, mean and standard deviation of males. H, holotype; SD, standard deviation.
| Morphometric and Meristic Characters |
|
Males |
Females |
|
(n = 3) |
(n = 3) |
| H |
Range |
Mean |
SD |
Range |
Mean |
SD |
| Standard length |
22.5 |
16.8–23.0 |
|
|
20.7–22.4 |
|
|
| Percent of standard length |
|
|
|
|
|
|
|
|
Total length |
133.3 |
129.6–133.3 |
130.9 |
2.1 |
130.4–133.8 |
132.4 |
1.8 |
|
Body depth at pelvic-fin origin |
26.7 |
25.6–27.4 |
26.6 |
0.9 |
27.6–29.5 |
28.3 |
1.0 |
|
Head length |
28.0 |
24.3–28.0 |
25.8 |
1.9 |
25.3–29.0 |
26.7 |
2.0 |
|
Preanal length |
62.2 |
59.1–62.2 |
60.7 |
1.5 |
63.8–64.7 |
64.2 |
0.5 |
|
Predorsal length |
69.3 |
66.1–69.3 |
67.5 |
1.7 |
70.0–71.5 |
70.7 |
0.7 |
|
Prepelvic length |
44.0 |
41.7–44.0 |
42.9 |
1.2 |
44.7–48.8 |
46.6 |
2.1 |
|
Prepectoral length |
30.7 |
26.1–30.7 |
27.8 |
2.5 |
26.7–30.0 |
27.8 |
1.8 |
|
Caudal peduncle length |
18.7 |
18.7–19.6 |
19.1 |
0.5 |
18.8–19.8 |
19.3 |
0.5 |
|
Caudal peduncle depth |
16.9 |
16.7–17.4 |
17.0 |
0.4 |
15.2–16.4 |
15.9 |
0.7 |
|
Dorsal-fin base length |
14.2 |
14.2–16.7 |
15.7 |
1.3 |
12.9–14.5 |
13.9 |
0.9 |
|
Anal-fin base length |
18.7 |
18.7–19.6 |
19.1 |
0.5 |
15.5–16.6 |
16.2 |
0.6 |
|
Caudal-fin length |
33.3 |
29.6–33.3 |
30.9 |
2.1 |
30.4–33.8 |
32.4 |
1.8 |
| Percent of head length |
|
|
|
|
|
|
|
|
Head width |
67 |
67–71 |
69.0 |
2.4 |
70–73 |
71.7 |
1.5 |
|
Head depth |
79 |
79–83 |
81.6 |
2.0 |
78–84 |
80.4 |
2.8 |
|
Interorbital width |
52 |
52–57 |
55.0 |
2.4 |
56–59 |
57.2 |
1.2 |
|
Postorbital length |
41 |
41–43 |
41.7 |
1.0 |
44–48 |
45.6 |
2.4 |
|
Suborbital depth |
17 |
14–18 |
15.3 |
1.8 |
15–19 |
16.7 |
2.2 |
|
Eye diameter |
38 |
38–41 |
38.7 |
1.6 |
38–42 |
40.5 |
1.9 |
|
Snout to eye end length |
59 |
57–59 |
58.3 |
1.0 |
52–56 |
54.4 |
2.4 |
|
Snout length |
22 |
19–22 |
20.3 |
1.7 |
17–18 |
17.4 |
0.8 |
| Other morphometric ratios |
|
|
|
|
|
|
|
|
Head width in % of its depth |
84 |
83–87 |
84.6 |
2.1 |
87–91 |
89.2 |
2.2 |
|
Caudal peduncle length in % of its depth |
111 |
111–114 |
112.4 |
1.9 |
120–124 |
121.4 |
1.9 |
|
Predorsal length in % of preanal length |
111 |
109–113 |
111.2 |
2.2 |
109–112 |
110.1 |
1.7 |
| Meristics |
|
range |
mode |
|
range |
mode |
|
|
Dorsal-fin rays |
9 |
9 |
9 |
|
9 |
9 |
|
|
Anal-fin rays |
13 |
13–14 |
13 |
|
13 |
13 |
|
|
Dorsal fin to anal fin relative position |
6 |
5–6 |
6 |
|
6 |
6 |
|
|
Scales mid-longitudinal series |
21 |
21–23 |
23 |
|
20–22 |
20 |
|
|
Scales transverse |
6 |
6 |
6 |
|
6 |
6 |
|
|
Scales circumpeduncular |
10 |
10 |
10 |
|
10 |
10 |
|
Coloration. Live males and females (
): Scales on trunk with a pattern of discrete iridescent, diamond-shaped, light blue spots, especially evident below mid-longitudinal series on posteroventral portion of flank, creating an irregular reflective pattern on body. Scales on dorsum grey-brown, with light blue iridescent blotches in males. Scales on the abdomen from opercle to pelvic fin are white to silver. Head yellow to light brown-grey, snout and dorsal portion of head grey-brown, throat silver. Exposed branchiostegal membrane white. Iris silver, light blue iridescent in the upper portion. Iridescent silver to blue blotches on postorbital opercular region. Iridescent silver to blue humeral blotch on post-opercular region in males. Median fins yellow to hyaline, dorsal and anal fins with six to eight dark grey stripes perpendicular to fin rays basally and posteriorly, with light blue iridescence basally; dorsal with light blue distinct margin; anal with dark grey margin; caudal fin with four to five dark grey stripes perpendicular to fin rays. Dorsal and anal fin in female hyaline with irregular brown stripes, with fin tips with narrow light blue margin, caudal hyaline without markings. Pelvic fin yellow in males, hyaline in females.
Distribution. ‘
Lacustricola’
gemma is endemic to freshwater habitats of the upper Congo drainage in south-eastern DR Congo. It is currently known from waters associated with the Kay system in the upper Lualaba drainage (
). The Kay is a major right bank affluent entering the upper Lualaba below Lake Upemba.
Ecology and biology. The area of the upper Lualaba experiences a mean annual rainfall of around 1000 mm, with December the wettest month. In the Upemba System, the waters of the Lualaba are high from February–April and low from August–October [
83]. The dry season lasts for around 160 days.
At the type locality, ‘
Lacustricola’
gemma was the only procatopodid species observed. The accompanying fish fauna consisted of the seasonal
Nothobranchius katemomandai. The type locality on 17 April 2023 was a remnant pool formed in the riverbed of the seasonal Lubule River (
d). The habitat was a drying ephemeral riverbed about 1–2 m wide, the edges of which were overgrown by grass. The water was turbid. The water temperature around noon was 27.3 °C, the pH was 7.2, and it contained 120 ppm of total dissolved solids. The habitat belongs to category 1.4.1 as defined by Watters (2015a) [
85], representing a remnant pool in a seasonal stream system on a floodplain.
Water parameters, measured by B. Katemo Manda at another confirmed site in Lake Bowe, showed a temperature of 28.4 °C; pH of 7.2; total dissolved solids of 30 ppm.
Aquarium maintenance of selected specimens was undertaken for observation of breeding behaviour and biology. ‘
Lacustricola’
gemma has a non-annual mode of reproduction, the same as all of its known congeners. Under captive conditions, a spawning mop prepared from acrylic yarn was used successfully as an artificial spawning medium, simulating submerged vegetation. Eggs were picked from the mop and stored in plastic containers with shallow water for development. An embryonic development period of about two to three weeks was observed at about 22–24 °C. Eggs are relatively large, round, measuring 1.6 mm in diameter on average (
n = 10).
Conservation status. ‘
Lacustricola’
gemma is recommended to be assessed as Vulnerable. The species is currently known only from the area of the type locality, sites in the Kay system of the Lualaba drainage. It might exist at some other sites within the drainage system of the upper Lualaba River, but its distribution will remain restricted. The entire drainage area of the Kay system has an extent of 1142 km
2 and the species is expected to be found from a maximum of ten threat-based locations. Some of the known sites at present, including the type locality, are directly adjacent to human populations. There is an expansion of agriculture in seasonal wetland habitats of the known distribution area and, consequently, an increase in anthropogenic pressures on the land and water use. Using IUCN (2012) principles [
86], ‘
L.’
gemma meets the criteria B1ab(iii) for Vulnerable, considering the drainage area as maximum EOO and the number of estimated maximum threat-based locations, as well as the risk of continuous decline of the known wetland habitats in which it is known to live.
There are currently no conservation or protection measures in place for this species. Additionally, it is unknown whether any protected areas would partially encompass its known distribution. Therefore, habitat protection is needed at the type locality and other sites within the Kay system where the species has been recorded. Further field surveys should target additional suitable habitats within the drainage area to document the full geographic range of the species more accurately. This would help in developing appropriate conservation strategies.
Etymology. The specific epithet
gemma is derived from the ancient Greek word γέμω, originally meaning “precious stone” or “gem”. It refers to the pattern of the body scales, which are adorned with numerous iridescent, diamond-shaped markings, evoking the appearance of tiny gems. The name also alludes to the relatively small adult size of the species. It is treated as a noun in the nominative singular, standing in apposition to the generic name.
4.1. Nothobranchiidae
4.1.1. History of
Nothobranchius brieni Species Group
The first documented species of
Nothobranchius from the south-eastern upper Congo drainage was
N. brieni, described from the Lualaba River by Poll (1938) [
17]. This was followed by Wildekamp’s (1978) redescription of
N. brieni and the description of three additional species [
6]. Since then, the recognised species richness within the group has grown to sixteen, followed by the identification of four more species in the present study (see
). This expanding body of research reflects the remarkable complexity and diversity of the
N. brieni species group and underscores the importance of continued exploration to better understand its evolutionary relationships and ecological significance better.
4.1.2. Phylogenetic Analysis of
Nothobranchius brieni Species Group
Molecular phylogenetic reconstructions in different studies confirmed that species of the
N. brieni species group form a well-supported monophyletic group (e.g., [
3,
8,
9,
36,
55,
68,
89]), that is resolved as part of the geographically-structured Inland clade (
sensu [
68]). All
Nothobranchius species found in the Katanga Province of the Congo, Zambia, and the Zambezi Region of Namibia have been found to comprise a distinct clade, referred to as the Kalahari Clade in [
36]. These species constitute the
N. brieni species group.
In this study, a phylogenetic analysis based on the COI gene sequence was conducted, demonstrating sufficient discriminative power for species resolution. The analysis included an expanded dataset of the
N. brieni species group, with the primary aim of evaluating the internal structure of the group and identifying new species as phylogenetically distinct lineages. The author acknowledges the limitations of single-gene analyses but presents this study to offer insights in comparison to previous research. It serves as an additional element among multiple corroborating lines of evidence in species delimitation. The phylogeny retrieved in this study is largely congruent with previous analyses.
Within the Lake Mweru complex, in addition to the described members of the genus, a putative species from the upper Lubi system is represented by three terminals:
N. spec. DRCP 2013-06 (also known as
N. spec. Kasenga), and
N. spec. CD 23-23 and
N. spec. CD 23-25. The description of that putative species from the upper Lubi drainage is in review, and it is represented here for clarity [
11].
4.1.3. Biogeography of
Nothobranchius brieni Species Group
Species in the
N. brieni group show allopatric distributions, each restricted to separate drainage systems or isolated parts of the same system, often divided by physical barriers such as mountains or waterfalls (e.g., [
10,
13,
16,
18];
). Their limited dispersal ability and strong habitat fidelity have led to speciation patterns closely aligned with historical drainage reorganisations across the south-central African plateau [
90].
Phylogenetic patterns and landscape history suggest that ancient hydrological connections facilitated early diversification, later disrupted by river capture and drainage shifts [
36,
91]. Given these mechanisms, further exploration of remote floodplains in south-eastern DR Congo and northern Zambia is likely to uncover further, as yet undescribed species.
4.1.4. Conservation Aspects
All
Nothobranchius fishes in the upper Congo drainage are subject to a high level of threat and belong to one of the threatened Red List categories [
18]. The importance of the discoveries from conservation aspects is illustrated by the numerous increasing anthropogenic impacts, and requires the need for active protection of the freshwater wetland habitats in which these species occur. The impact of human activities was evident in three out of four types of localities, whereas the fourth type of locality in the Kay system is adjacent to human population.
Molecular analyses revealed evidence of fish-pathogenic oomycetes
Aphanomyces invadans and
A. laevis on samples collected in 2023, which represent a first-time record for the host of the fish genus
Nothobranchius [
87]. Observations involve at least two of the new species in this study. Specimens of
N. marmoreus were observed by the present author in 2016 with lesions suggestive of infection with
Aphanomyces spp, whereas molecular analysis revealed
Aphanomyces laevis on
N. katemomandai collected in 2023. Evidence of infection with oomycete pathogens documented in Nagy (2024a) [
87] represents an important additional threat to the unique seasonal aquatic biodiversity of the region.
Conservation efforts for
Nothobranchius species must focus on preserving their unique ephemeral wetland habitats and addressing the multiple human-induced threats they face. Their conservation is not only crucial for maintaining biodiversity but also for preserving valuable scientific research opportunities for aging studies.
4.2. Procatopodidae
4.2.1. History of Procatopodid Species in Upper Congo
Different forms of lampeyes in the upper Congo region have been long associated with three early-described species and typically lumped into them:
Haplochilus johnstoni [
47];
Haplochilus katangae [
48]; and
Haplochilus hutereaui [
46]. These species were largely characterized by basic external features, such as elongated body and unmarked fins
vs. a zigzag black stripe along the flank
vs. barred median fins, respectively. Based on this simple characterization, many populations across the southern part of Africa were associated with each of these species, creating species groups. The status of an additional, long-described species,
Haplochilus moeruensis [
54], was not clear and was often associated with different species around the Lake Mweru drainage, until Seegers (1996) [
92] selected a lectotype and published a photograph of it.
In parallel, the alpha taxonomy of lampeye cyprinodontiform fishes of Africa has been mostly neglected for a long time. Their phylogenetic relationships were poorly researched, and different hypotheses yielded inconsistent results among various authors [
40,
93,
94,
95,
96].
Aforementioned species were assigned to several genera during several decades, namely to
Aplocheilichthys (e.g., [
22,
45]), to
Micropanchax (e.g., [
40,
56,
97]) or to
Lacustricola (e.g., [
96,
98]), whereas currently they are regarded as belonging to an undescribed genus ‘
Lacustricola’ [
41].
4.2.2. Phylogenetic Analysis of Procatopodid Species in Upper Congo
Phylogenetic analysis performed by Bragança & Costa (2019) [
41] revealed evidence that the genus
Lacustricola is polyphyletic, with included species occurring in two separate, geographically disjunct clades. The eastern African
Lacustricola clade comprises species that are largely confined to coastal rivers and lakes in East Africa, including the type species of the genus
L. pumilus, from the Lake Tanganyika basin. Another clade, comprising species predominantly distributed in southern Africa, represents an undescribed genus. The species of this clade are referred to in this paper as ‘
Lacustricola’. The species from the upper Congo drainage mentioned above belong to this group (
). Additionally, three other members of Procatopodidae found in the upper Congo drainage also belong to this genus, such as ‘
L.’
petnehazyi [
56], ‘
L.’
nitidus [
55], as well as a form referred to herein as ‘
L.’ cf. jubbi, initially described in the genus
Hypsopanchax by Poll & Lambert (1965) [
53] from the upper Zambezi (
).
However, one species described from the upper Congo basin was found to belong to the genus
Lacustricola (
sensu stricto) as
L. lualabaensis was revealed to be part of the same phylogenetic clade as
L. pumilus.
4.2.3. Geographical Distribution and Biogeography
The complex geological history of the Congo drainage and the diverse and changing aquatic environments it supports, likely provided numerous ecological niches for Procatopodids to adapt to and speciate. Periods of isolation due to forest fragmentation and changing water levels may have promoted allopatric and peripatric speciation in Procatopodid species [
49,
50].
The complex network of tributaries and rivers in the Congo drainage probably facilitated the dispersal and diversification of Procatopodid species. It is likely that many undiscovered or unidentified new species exist within the drainage system. To fully understand the evolution of Procatopodid species in the Upper Congo drainage, further research focusing specifically on these fish in the region is necessary.
4.2.4. Conservation Aspects
The conservation of southern African procatopodid species faces significant challenges due to the vulnerability of their habitats. During the rainy season, lampeyes typically migrate to shallow, flooded areas for breeding, highlighting the critical importance of these ephemeral aquatic environments. This seasonal behavioural pattern dependence makes them especially susceptible to habitat alterations and degradation. The conservation of these species is further complicated by the multitude of threats facing wetland ecosystems in southern Africa. Human activities such as agricultural expansion, water extraction, urban development, and industrial pollution, particularly from mining operations, pose severe risks to these delicate habitats. Recent taxonomic studies indicate that the Procatopodidae family is much more diverse than previously thought. In addition to some widely distributed species, it also includes narrowly distributed endemics [
51]. As the majority of wetlands in southern Africa are unprotected, this underscores the urgent need for comprehensive conservation strategies. Preserving the integrity of these wetlands throughout all seasons is crucial for maintaining the ecological balance and safeguarding the distinctive seasonal freshwater biodiversity they support, including the vulnerable procatopodid species.
4.3. Comparative Material
‘
Lacustricola’
chobensis: SAIAB 30007, holotype, male, 24.7 mm SL; SAIAB 30008, 4 paratypes, males, 11.6–16.4 mm SL; Botswana: Chobe River: Kasane, 17°49′ S, 25°08′ E.
‘L
acustricola’
hutereaui: RMCA P-1822-3, syntypes, 2 males, 22.1–25.1 mm SL; DR Congo: Uele River: Dungu, 03°41′ N, 29°08′ E. — RMCA 73-23-P-10550-865, 3 males, 27.1–31.7 mm SL; DR Congo: Uele River: Buta, 02°47′ N, 24°46′ E.
Nothobranchius brieni: RMCA 50016, lectotype, male, 46.3 mm SL; RMCA 50018-27, paralectotypes, 4 males, 44.4–46.5 mm SL; DR Congo: Lualaba drainage: Bukama, 09°12′ S, 25°51′ E. — RMCA B4-019-P-0013-4, 2 males, 30.3–30.4 mm SL; DR Congo: Lualaba drainage: Bukama, 09°11′ S, 25°51′ E.
N. ditte: RMCA 2016-027-P-0001, holotype, male, 33.0 mm SL; RMCA 2016-027-P-0002, paratype, male, 38.9 mm SL; RMCA 2016- 027-P-0029, paratype, male, 42.4 mm SL; RMCA 2016-027-P-0004-10, 7, paratypes, males, 31.1–38.9 mm SL; DR Congo: Lake Mweru basin: Kilwa, 09°12′ S, 28°17′ E.
N. hassoni: MSNG 51837, holotype, male, 41.1 mm SL; MSNG 51838-9, paratypes, 2 males, 34.9–37.2 mm SL; MSNG 51840, paratype, male, 33.9 mm SL; RMCA A3-028- P-0003, paratype, male, 41.0 mm SL; DR Congo: middle Lufira drainage: Lufwa system: Kanvungwe, 09°29′ S, 27°17′ E. — RMCA A3-028-P-0004, paratype, male, 32.0 mm SL; DR Congo: middle Lufira drainage: Lwipa system, 09°08′ S, 27°20′ E. — RMCA B3-028-P- 0001-4, 4 males, 27.9–35.2 mm SL; RMCA B4-019-P-0012, male, 38.5 mm SL; DR Congo: middle Lufira drainage: Bunkeya, 10°25′ S, 26°58′ E. — RMCA B0-013-P-0001-0014, 5 males, 40.1–43.7 mm SL; DR Congo: middle Lufira drainage: Mwashya, 10°42′ S, 27°21′ E.
N. malaissei: RMCA 73-24-P-952, holotype, male, 33.0 mm SL; RMCA 73-24-P-914-919, RMCA 73-24-P-947-951, 6 males, 36.4–43.2 mm SL; DR Congo: Luapula drainage: Kabiasha, 10°17′ S, 28°09′ E.
N. milvertzi: RMCA B2-027-P-0001, holotype, male, 36.9 mm SL; RMCA B2-027-P-0002, 1, male, 36.0 mm SL; RMCA B2-027-P-0003-16, 14, 6 males, 30.5–35.9 mm SL; Zambia: Lake Mweru basin, Lushiba Marsh: Chienge, 08°36′ S, 29°07′ E.
N. oestergaardi: BMNH 2010.12.6.1, holotype, male, 31.8 mm SL; RMCA 2010-33-P-4, paratype, male, 30.4.0 mm SL; MSNG 56047A-B-C, paratype, male, 42.7 mm SL; paratype, male, 29.0 mm SL; SAIAB 98224, paratype, male, 26.4 mm SL; Zambia, Lake Mweru Wantipa basin: Kalaba, 8°25′ S, 29°50′ E.
N. polli: RMCA 192399, holotype, male, 28.9 mm SL; DR Congo: Katanga province: upper Lufira drainage: Dilungu swamp, 10°45′ S, 27°15′ E. — RMCA B4-019-P-0015, male, 34.1 mm SL; DR Congo: Katanga province: upper Lufira drainage, 11°02′ S, 27°18′ E. — RMCA B4-019-P-0016-8, 3 males, 26.1–35.1 mm SL; DR Congo: Katanga province: upper Lufira drainage: Kyembe, 11°02′ S, 27°18′ E.
I am grateful to my late friend, Auguste Chocha Manda, from the University of Lubumbashi, for his long-standing support in research expeditions and the immense pleasure always for the time spent together in the field; to Bauchet Katemo Manda and Augustin Kalumba, both from the University of Lubumbashi, and to Gábor Petneházy for assistance in the field, and to Stefano Valdesalici for providing preserved material for molecular analysis; to Augustin Kalumba, Bauchet Katemo Manda, Brian Watters, Csenge Nagy, Stefano Valdesalici and SAIAB staff for fish photographs, and to Emmanuel Vreven for his support on permits and collection management.
The study was conducted in accordance with the ethical standards and Congolese legislation, according to the guidelines of the Declaration of Helsinki, authorized by the University of Lubumbashi and approved on 20 March 2025 by the Ministry of Environment and Sustainable Development, Kinshasa, DR Congo (n°13 SG/EDD/BTB/ANCCB-RDC/03/2025).
Not applicable.
Specimens of the type series and examined comparative material are deposited in ichthyology collections, whereas genetic sequences are deposited in GenBank. Additional data to support the findings of this study are available from the author upon reasonable request.
This research received no external funding.
The author declares that he has no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.