1. Introduction
Urbanisation is driving rapid declines in species richness and abundance worldwide, driving changes in species assemblages, and leading to biotic homogenisation [
1,
2,
3]. However, it is known that urbanisation can increase or decrease species richness, depending on several variables [
4,
5,
6]. The impact of urbanisation on species abundance varies, depending on taxons and cities [
7,
8]. Urban habitats are reported to favour some species due to man-made habitats, wastes and substrates, which are exploited by animals as food, decreasing pressure from predators or competitors [
1,
9]. The main effect of urbanisation is the replacement of specialists with generalist species [
2,
4]. Nevertheless, some rare species live in urban environments [
10]. Species diversity within urban habitats can vary from very high to very low [
11,
12,
13]. Cities are heat islands [
14], and urban habitats at more northern latitudes can be similar to rural habitats at more southern latitudes [
15,
16]. However, urban sites can be less sunny and cooler than those in the suburbs [
17]. It is shown that climate strongly modifies the effects of urbanisation [
18]. The history of urbanisation, which is unequal in different parts of the world, also impacts its ecological consequences [
19]. Therefore, it is important to study the urbanisation effect in different regions. Now, people believe that it is important to conserve biodiversity in cities [
20], so we need to know patterns of the shaping of species assemblages in the urban environment.
Orthoptera are one of the most noticeable groups of insects due to their songs and hoppers. Some of them are pests, while others are endangered charismatic species [
21,
22,
23]. Orthopterans are frequently used as model organisms in ecological studies and modelling, because a large part of them have limited dispersal ability and specific habitats, and these insects can be relatively easily collected and identified [
24,
25,
26]. There are a significant number of papers about orthopterans in urban environments [
27,
28,
29,
30,
31,
32,
33,
34,
35,
36,
37,
38,
39,
40]. The earliest of them describe a decrease in diversity and abundance along urban gradients—from suburbs to city centres, where only some eurytopic species live. Recent surveys mainly focus on species traits. It was shown that species with low mobility and more specialised habitats have a higher probability of local extinction in the urban environment [
39]. Yet, in some cases, mobile species were more sensitive to urbanisation at larger scales than sedentary species [
33]. Urban habitats can be colonised by mobile species, as well as by habitat-specialised species that can live within small patches of habitat [
32]. As shown, the urban matrix has great significance for surviving orthopterans in cities [
40]. However, some surveys suggest that the vegetation type of the patch is more important than urban landscape parameters [
38]. Therefore, patterns of the influence of the urban environment on orthopterans are not well known. It is not clear what ecological traits of species (habitat preference, layer preference, dispersal ability) favour them to colonise urban habitats. In particular, some researchers found that cities are favourable to thermophile orthopterans [
31], but in light of the cited paper [
17], we should verify if it is a general pattern or not. Most studies on urbanisation effects on orthopterans have been performed in West, Central or South Europe and in West Siberia, while East Europe is underexplored in this aspect despite published species lists for some cities [
41,
42,
43,
44,
45]. Investigation of Orthoptera in the urban environment of East Europe would allow us to clarify general patterns of urbanisation impacting insects and assess the possibility of conservation of this group in cities.
In this study, we investigated Orthoptera in Kaluga, a typical middle-Russian city in the broadleaved and mixed coniferous-broadleaved forest zone. Our questions were as follows. (1) What species of Orthoptera colonise urban habitats, and what are ecological traits (habitat preference, layer preference, dispersal ability and phenology) according to which the urban environment filters species from a regional species pool? (2) What species of Orthoptera colonise lawns in the urban environment? (3) What properties of urban habitats enhance their favourability to Orthoptera?
2. Materials and Methods
2.1. Study Area
Kaluga is located in the centre of the East-European plain, on the northwest margin of the Middle-Russian Upland, at the boundary of the Continental and Boreal biogeoregions [
46] or the mixed coniferous-broadleaved forests zone and the broadleaved forest zone [
47]. So, the primary vegetation of the area is oak-linden and spruce-oak-linden forests. But it is an old agricultural region, so meadows (pastures and haylands) have a long history. It is located on the Oka River, the largest right tributary of the Volga River, and the meadows in its valley are rich in forest-steppe plants—“the Oka flora” [
48].
Kaluga is a large urban settlement and urban district, but for our study, we should regard it as an urban landscape, a specific system of habitats for insects. Historically, the urban landscape of Kaluga developed on the left bank of the Oka River, between its tributaries, the Yachenka and Kaluzhka [
49].
The western suburb of Kaluga (to the west of the Yachenka River) is formed by alluvial plains with sandy sediments [
50], with pine forests and dry meadows which are located mainly along the Oka River and have many forest-steppe plants, with small swamps.
The southeastern suburb of Kaluga is formed by a fluvioglacial plain with loam sediments and is occupied mainly by fields. Small tall-grass meadows are located along the Kaluzhka River. Dry meadows are located along the Oka River and on some roads, as well as in broad clearings along electrical lines. Locally, there are lime quarries.
The northeast suburb of Kaluga is formed mainly by moraine plains with clay loam sediments. It is occupied by mesic or moist meadows, secondary deciduous forests (including those with birch or aspen, as well as linden, oak, and maple), gardens, and hamlets. This suburb is separated from the urban landscape by a broad belt of deciduous forests and a cluster of new high-tech factories.
The north and northwest city peripheries and suburbs do not have large meadows; in these areas, industrial and residential buildings are alternated with secondary deciduous forests, making it difficult to find suitable sample plots. Through their landforms and forest vegetation, these lands resemble the northeast suburbs. In the south, the Oka River is the natural boundary of the urban landscape. Large meadows along this river within the Kaluga city are absent; the bank is mainly populated by willows and box-elder. On the right bank of the river, there are also built areas of Kaluga, but we regard them as separate small urban landscapes and do not include them in this research.
2.2. Sample Plots
In total, we surveyed 174 individual plots in the urban landscape and in the suburbs: the western (W), the northeastern (NE), and the southeastern (SE). Each sample plot is characterised by a unique combination of geographical coordinates, has uniform vegetation cover and is not divided by roads or by other barriers for orthopterans. We divided them into the following types, primarily based on EUNIS [
51].
2.2.1. Meadows
Habitats dominated by grasses and herbs, with low anthropogenic pressure,
i.e., unmowed or mowed one or two times per summer. This type includes all grasslands outside built areas or roadsides. Within built areas or roadsides, the mowing frequency was estimated by regular direct observation and by observing unmowed herbs (relatively tall with seeds) during summer surveys of the insects.
Dry Meadows
Grasslands on sands, with bare ground, dominated by dry-tolerant grasses and herbs:
Festuca ovina L.,
Festuca vallesiaca Schleich. ex Gaudin,
Koeleria glauca (Spreng.) DC.,
Pilosella spp.,
Artemisia campestris L.,
Fragaria viridis Weston.
Mesic Meadows
Dominated by such grasses as:
Calamagrostis epigejos (L.) Roth,
Dactylis glomerata L.,
Festuca pratensis Huds.,
Elytrigia repens (L.) Nevski,
Phleum pratense L.,
Festuca rubra L.,
Poa pratensis L.,
Agrostis capillaris L.
Wet Meadows
Dominated by large sedges (
Carex and
Scirpus spp.) and grasses.
2.2.2. Lawns
Intensively-mowed grass sites with short herb layers, surrounded by sealed surfaces and buildings, usually with small areas or on roadsides. Lawns can be found along streets, between residential or business buildings, in squares and parks. Usually, “lawn” means cultivated grassy vegetation. But in Kaluga lawns are often degraded, with bare ground and small ruderal weeds. Most sites are dominated by grasses,
i.e.,
D. glomerata, somewhere by forbs, such as
Taraxacum officinale F.H.Wigg., somewhere by forbs, which are typical for shaded environment, such as
Aegopodium podagraria L. or
Glechoma hederacea L. Most of the lawns are shaded by trees and buildings. Specific subcategories of lawns are unshaded lawns (located between roads without buildings) and lawns with shrubs (hedgerows or decorative), sometimes with small patches of rarely mowed herbage around shrubs or artificial constructions.
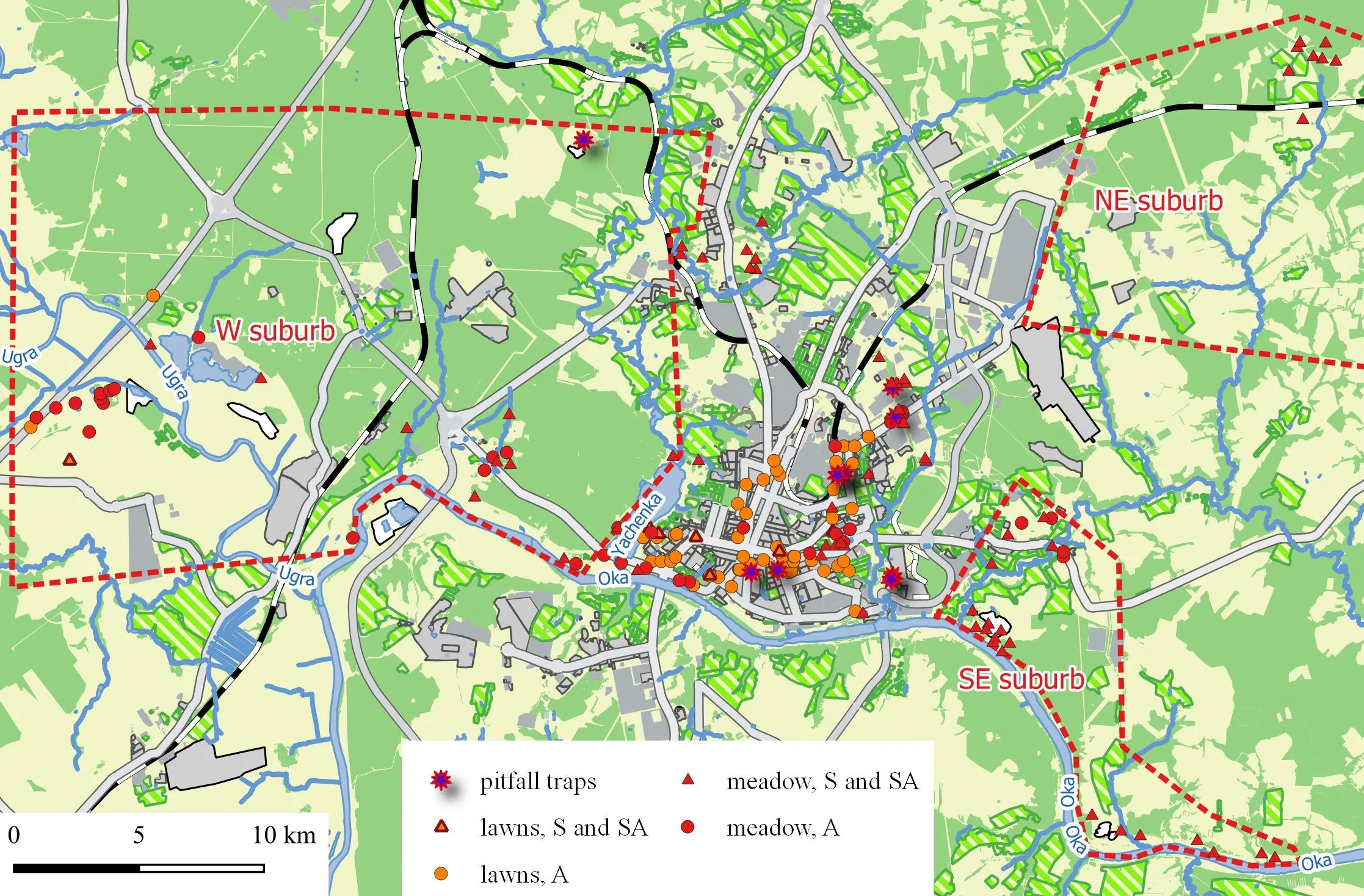
The distribution of these types of habitats in the urban landscape and the suburbs is presented in
. Sample plots are mapped in
. A list of sample plots with geographical coordinates can be found in supplementary materials.
.
Numbers of sample plots per type of habitats used for survey of Orthoptera in Kaluga, Russia.
| Method |
|
Numbers of Plots |
|
Total |
| Zone |
Lawns |
Meadows |
| Dry |
Mesic |
Wet |
| Sweepnet |
suburbs |
NE |
0 |
0 |
8 |
|
8 |
| SE |
0 |
3 |
17 |
|
20 |
| W |
1 |
3 |
7 |
1 |
12 |
| urban |
|
6 |
0 |
33 |
|
39 |
| Total |
|
7 |
6 |
66 |
|
79 |
| Acoustic and visual |
suburbs |
NE |
0 |
0 |
2 |
|
2 |
| SE |
0 |
3 |
8 |
|
11 |
| W |
2 |
5 |
15 |
|
22 |
| urban |
|
57 |
0 |
22 |
|
79 |
| Total |
|
59 |
8 |
47 |
|
114 |
| Pitfall traps |
urban |
|
3 |
0 |
6 |
1 |
10 |
. Map of the surveyed area. A—acoustic and visual observation, S—sweepnet.
For lawns in urban landscapes, we also measured the following properties. (1) Shading by trees or buildings during most part of the day: 0—unshaded, 1—shaded. (2) Density of the herbage: 0—sparse herb cover, with patches of bare ground, 1—dense herb cover, without visible patches of bare ground. (3) Presence of uncut patches—shrubs or tall ruderal or ornamental herbs (
i.e.,
D. glomerata and
Urtica dioica L.): 0—absent, 1—present. (4) Degree of isolation from a suburb or large urban meadow, assessed as a number of barriers, which are roads and large areas without grassland habitats: 1—one road with high traffic; 2—two roads with high traffic, or one such road and one array of built area of 100–200 m, 3—three roads or arrays of built area, 4—four or more roads with high traffic.
2.3. Survey of Orthoptera
Orthoptera were surveyed from 2002 to 2024. Detailed information on sampling events can be found in the datasets [
52,
53]. Information on sampling protocols for each plot is given in
Table S1.
We surveyed orthopterans using three methods: sweepnet, acoustic and visual observations, and pitfall traps.
Sweepnet was used on sunny days in late July–September when almost all species were adults. In some plots (No. 147, 156, 161, 162, 166 in
Table S1), orthopterans were also sampled by sweepnet in June–late July to find nymphs. Sweepnet in large meadows consisted of 100 strokes. Our path was a linear transect. Its directions were optimal to not scare away the insects before catching them. In small meadows and lawns, we passed all their areas; in linear habitats, it was also a linear path, and in squares, we went all lawns by a circle, so in such habitats, our sampling effort was 50–100 strokes.
Acoustic and visual observations have been conducted on sunny days in late July–September in 2023 and 2024. In 2023–2024, we went to most of the points surveyed previously using sweepnet to distinguish grasshoppers of the
Chorthippus biguttulus group. The duration of the acoustic survey was 15 min in lawns and small meadows and 30 min in large meadows. In late July–early August of 2023, we recorded grasshoppers of the
Chorthippus biguttulus group from plot No. 43 using Xiaomi Redmi 8. In other cases, orthopterans were auditively observed. In lawns, most of the specimens found by songs have also been seen (excluding
Tettigonia), so we note this method as “acoustic and visual observation”, and we usually did not need the sweepnet. In meadows, orthopterans have not been seen during acoustic observation, and we combined the sweepnet and acoustic observation.
Pitfall traps were applied to survey ground-dwelling insects and to quantitative assessment of abundance in urban areas. A trap was a 0.5 L transparent plastic cup with a mouth of 85 mm in diameter filled with 4% formalin solution, with a cover made of transparent polyethylene film. Every year, 15 traps were exposed during May–September, and insects were sampled every two weeks. In each plot, 8–15 traps were exposed according to their size. We calculated the activity density of insects—the number of specimens per 100 trap days.
Orthopterans were identified according to Bey-Bienko’s key [
54]. For
Omocestus and
Stenobothrus we used the book of Bey-Bienko and Mitshenko [
55]. Identification of specimens of the
Chorthippus biguttulus groups was based on recent works [
56,
57,
58,
59]. Nomenclature was given according to Orthoptera Species file [
60].
2.4. Species Traits and Statistical Analyses
The dispersal ability of species was accessed using a well-known paper on the Continental biogeoregion [
61] but with some corrections based on other works [
62,
63], which are consistent with our observation in Kaluga. By layer distribution, we divide species into several groups using literature data on life forms [
64,
65]: 1—insects spending all life on plants (specialised phytophiles); 2—insects which live on plants (grass-inhabitants) but lay eggs in the soil; 3—insects which keep both on plants and the ground; 4—insects living on the ground (geophiles and herpetobionts). By phenology, we divide species into two groups: “early”—emerging in June and early July; “late”—emerging in late July–August. By habitat preference, we classified species by their distribution in the suburbs of Kaluga into four groups: (1) grassland generalists (inhabit mesic, dry and moist meadows), (2) habitats of dry meadows, (3) habitats of moist to wet meadows, (4) forest species. For detailed information, see Table A1.
Our datasets consist of data on relative abundance (the number of specimens observed with a certain sampling effort). However, in this paper, we analyse only the presence–absence of species in each site, as the quantitative comparison of orthopterans observed at different times and in plots with unequal herbage heights appears to be problematic. Our data includes 1430 occurrences. One occurrence is a find of one species in one day in one site. These are rows of the datasets [
52,
53] belonging to Kaluga city and surroundings, excluding non-systematic (occasional) observations and data sampled using traps.
Data processing and statistical analyses were conducted using the R 4.1.3 [
66]. To compare frequencies of species and groups of species between suburbs and urban areas or between meadows and lawns, we used Fischer’s test. To assess the influence of the four above mentioned properties of lawns on the occurrence of insects, we used GLM (generalised linear models) with binary distribution and iteratively reweighted least squares. To compare groups of species according to mentioned species traits, we calculated a relative difference in species occurrences [
67] between suburbs and urban landscapes for each species and then obtained Kruskal-Wallis’s test for groups of species. To compare activity density between different types of habitats, we applied Kruskal-Wallis’s test. Maps were made in QGIS 3.
3. Results
3.1. Species Composition in Urban Landscape Compares to Suburbs
In total, we sampled 29 species of Orthoptera in meadows in Kaluga city and suburbs using sweepnet, acoustic and visual observations (
). Nine species of Orthoptera were found in 1–4 sites; hence, they are not applicable to comparison. Excluded two small ground-dwelling species of
Tetrix, we have 17 species of Orthoptera to assess the impact of urbanisation. For six species, we found a negative effect of the urban landscape on the number of occupied sites based on one or both methods. Five species did not decrease their occupancy in the urban landscape compared to suburbs, as estimated by both methods. They were:
Chorthippus apricarius (Linnaeus, 1758),
Ch. biguttulus (Linnaeus, 1758),
Chrysochraon dispar (Germar, 1834),
Conocephalus fuscus (Fabricius, 1793),
Decticus verrucivorus (Linnaeus, 1758). They may be regarded as tolerant of urbanisation. For other species, differences between urban and suburban habitats were not significant due to low frequencies. In meadows surveyed during June–late July, we found both adults and nymphs of such species as
C. fuscus,
D. verrucivorus,
Phaneroptera falcata (Poda, 1761),
Tettigonia cantans (Fuessly, 1775), the
Chorthippus biguttulus group,
Pseudochorthippus parallelus (Zetterstedt, 1821),
Tetrix spp. So, observed orthopterans are true inhabitants of the city.
Regarding species traits, we found that the urban environment negatively affected all three species of dry meadows and only one of ten grassland generalists (Fischer’s test
p = 0.014). Both forest (edge) species were influenced negatively.
For phenological groups, there were no significant differences between late and early species (the Kruskal-Wallis’s test
p = 0.489 for acoustic and visual observation and 0.496 for sweepnet). This was also true of the dispersal ability between high-movers and low-moderate-movers (
p = 0.489 and 0.496). Regarding layer preference, all groups are presented both among tolerant and among sensitive species (
p = 0.66 and 0.48).
.
Occurrences of Orthoptera in Kaluga city with suburbs estimated using sweepnet and acoustic and visual observation (AV).
| Species |
Percentage of Occupied Plots |
Fisher’s Test, p |
| Sweepnet |
AV |
| Suburb |
Urban |
Suburb |
Urban |
Sweepnet |
AV |
| Bicolorana bicolor (Philippi, 1830) |
32.5 |
23.1 |
20.0 |
1.3 |
ns |
ns |
| Calliptamus italicus (Linnaeus, 1758) |
5.0 |
0.0 |
11.4 |
0.0 |
ns |
0.0079 |
| Chorthippus apricarius (Linnaeus, 1758) |
47.5 |
46.2 |
17.1 |
15.2 |
ns |
ns |
| Chorthippus biguttulus (Linnaeus, 1758) |
72.5 |
71.8 |
71.4 |
73.4 |
ns |
ns |
| Chorthippus brunneus (Thunberg, 1815) |
5.0 |
0.0 |
14.3 |
0.0 |
ns |
0.0022 |
| Chorthippus dorsatus (Zetterstedt, 1821) |
80.0 |
53.8 |
57.1 |
36.7 |
ns |
ns |
| Chorthippus mollis (Charpentier, 1825) |
35.0 |
2.6 |
57.1 |
21.5 |
0.004 |
0.0001 |
| Chrysochraon dispar (Germar, 1834) |
17.5 |
20.5 |
8.6 |
1.3 |
ns |
ns |
| Conocephalus fuscus (Fabricius, 1793) |
45.0 |
61.5 |
25.7 |
30.4 |
ns |
ns |
| Decticus verrucivorus (Linnaeus, 1758) |
15.0 |
15.4 |
8.6 |
8.9 |
ns |
ns |
| Euthystira brachyptera (Ocskay, 1826) |
50.0 |
25.6 |
14.3 |
0.0 |
0.037 |
0.0022 |
| Gomphocerippus rufus (Linnaeus, 1758) |
5.0 |
0.0 |
0.0 |
0.0 |
ns |
ns |
| Leptophyes albovittata (Kollar, 1833) |
5.0 |
0.0 |
0.0 |
0.0 |
ns |
ns |
| Locusta migratoria (Linnaeus, 1758) |
0.0 |
0.0 |
2.9 |
0.0 |
ns |
ns |
| Metrioptera brachyptera (Linnaeus, 1761) |
7.5 |
0.0 |
0.0 |
0.0 |
ns |
ns |
| Oedipoda caerulescens (Linnaeus, 1758) |
7.5 |
0.0 |
14.3 |
0.0 |
ns |
0.0022 |
| Omocestus haemorrhoidalis (Charpentier, 1825) |
5.0 |
0.0 |
2.9 |
0.0 |
ns |
ns |
| Omocestus viridulus (Linnaeus, 1758) |
15.0 |
2.6 |
2.9 |
0.0 |
ns |
ns |
| Phaneroptera falcata (Poda, 1761) |
55.0 |
51.3 |
17.1 |
1.3 |
ns |
ns |
| Pholidoptera griseoaptera (De Geer, 1773) |
12.5 |
5.1 |
14.3 |
0.0 |
ns |
0.0022 |
| Poecilimon intermedius (Fieber, 1853) |
5.0 |
2.6 |
0.0 |
0.0 |
ns |
ns |
| Pseudochorthippus parallelus (Zetterstedt, 1821) |
72.5 |
69.2 |
25.7 |
7.6 |
ns |
ns |
| Roeseliana roeselii (Hagenbach, 1822) |
32.5 |
35.9 |
17.1 |
8.9 |
ns |
ns |
| Stenobothrus lineatus (Panzer, 1796) |
5.0 |
2.6 |
0.0 |
0.0 |
ns |
ns |
| Stethophyma grossum (Linnaeus, 1758) |
0.0 |
2.6 |
0.0 |
0.0 |
ns |
ns |
| Tetrix bipunctata (Linnaeus, 1758) |
2.5 |
0.0 |
0.0 |
0.0 |
ns |
ns |
| Tetrix subulata (Linnaeus, 1758) |
12.5 |
5.1 |
2.9 |
0.0 |
ns |
ns |
| Tetrix tenuicornis (Sahlberg, 1891) |
10.0 |
10.3 |
0.0 |
2.5 |
ns |
ns |
| Tettigonia cantans (Fuessly, 1775) |
27.5 |
20.5 |
20.0 |
11.4 |
ns |
ns |
3.2. Species Colonising Urban Lawns Compares to Meadows
In urban lawns, we found 11 species of Orthoptera, nine using acoustic and visual observation, eight using sweepnet (
), and five using pitfall traps (
). Almost all species were found in lawns rarer than in meadows. For four species, these differences were statistically significant. Based on acoustic and visual observation, the most frequent species in lawns were
Chorthippus biguttulus,
Ch. dorsatus, and
Conocephalus fuscus. These three species are late-emerging and have high dispersal ability.
.
Occurrences of Orthoptera in Kaluga city with suburbs estimated using sweepnet and acoustic and visual observation (AV).
| Species |
Number of Occupied Plots |
Fisher’s Test, p, Sweepnet |
| Sweepnet |
AV |
| Meadows |
Lawns |
Meadows |
Lawns |
| Bicolorana bicolor (Philippi, 1830) |
7 |
2 |
0 |
1 |
ns |
| Chorthippus apricarius (Linnaeus, 1758) |
17 |
1 |
5 |
6 |
ns |
| Chorthippus biguttulus (Linnaeus, 1758) |
22 |
4 |
19 |
33 |
0.0186 |
| Chorthippus dorsatus (Zetterstedt, 1821) |
16 |
5 |
7 |
17 |
ns |
| Chorthippus mollis (Charpentier, 1825) |
1 |
0 |
10 |
7 |
0.0042 |
| Chrysochraon dispar (Germar, 1834) |
8 |
0 |
1 |
0 |
ns |
| Conocephalus fuscus (Fabricius, 1793) |
22 |
2 |
7 |
13 |
ns |
| Decticus verrucivorus (Linnaeus, 1758) |
6 |
0 |
4 |
2 |
ns |
| Euthystira brachyptera (Ocskay, 1826) |
10 |
0 |
0 |
0 |
ns |
| Omocestus viridulus (Linnaeus, 1758) |
1 |
0 |
0 |
0 |
ns |
| Phaneroptera falcata (Poda, 1761) |
17 |
3 |
0 |
0 |
ns |
| Pholidoptera griseoaptera (De Geer, 1773) |
1 |
0 |
0 |
0 |
ns |
| Poecilimon intermedius (Fieber, 1853) |
1 |
0 |
0 |
0 |
ns |
| Pseudochorthippus parallelus (Zetterstedt, 1821) |
23 |
4 |
3 |
1 |
ns |
| Roeseliana roeselii (Hagenbach, 1822) |
14 |
0 |
4 |
0 |
0.0049 |
| Stenobothrus lineatus (Panzer, 1796) |
1 |
0 |
0 |
0 |
ns |
| Stethophyma grossum (Linnaeus, 1758) |
1 |
0 |
0 |
0 |
ns |
| Tetrix subulata (Linnaeus, 1758) |
1 |
0 |
0 |
0 |
ns |
| Tetrix tenuicornis (Sahlberg, 1891) |
2 |
1 |
1 |
0 |
ns |
| Tettigonia cantans (Fuessly, 1775) |
8 |
0 |
7 |
1 |
0.0001 |
Using pitfall traps, we found 15 species in urban meadows and five species in urban lawns (
). Total abundance and species richness of Orthoptera were significantly lower in lawns than in meadows. Three species were frequent in urban meadows but colonised no lawn.
Decticus verrucivorus and
Tetrix tenuicornis are ground-dwelling.
Chrysochraon dispar is regarded as a grass-inhabiting species that can lay eggs on plants. The most abundant species in lawns was
Ch. biguttulus.
.
Activity density of Orthoptera in urban habitats of Kaluga using pitfall traps.
| Species |
Activity Density, ex. per 100 Trap Days |
| Lawns |
Meadows |
| Median |
Min |
Max |
Median |
Min |
Max |
| Bicolorana bicolor (Philippi, 1830) |
0 |
0 |
0.1 |
0 |
0 |
0 |
| Chorthippus apricarius (Linnaeus, 1758) |
0 |
0 |
0.1 |
3.3 |
0 |
7.4 |
| Chorthippus biguttulus (Linnaeus, 1758) |
4.7 |
0.7 |
4.8 |
3.1 |
0.3 |
10.6 |
| Chorthippus dorsatus (Zetterstedt, 1821) |
0.2 |
0 |
0.3 |
0.1 |
0 |
0.8 |
| Chrysochraon dispar (Germar, 1834) |
0 |
0 |
0 |
0.1 |
0 |
0.3 |
| Conocephalus fuscus (Fabricius, 1793) |
0 |
0 |
0 |
0 |
0 |
0.2 |
| Decticus verrucivorus (Linnaeus, 1758) |
0 |
0 |
0 |
0.3 |
0 |
1.1 |
| Euthystira brachyptera (Ocskay, 1826) |
0 |
0 |
0 |
0 |
0 |
0.1 |
| Metrioptera brachyptera (Linnaeus, 1761) |
0 |
0 |
0 |
0 |
0 |
0.1 |
| Phaneroptera falcata (Poda, 1761) |
0 |
0 |
0 |
0 |
0 |
0.1 |
| Pseudochorthippus parallelus (Zetterstedt, 1821) |
0 |
0 |
0 |
0.1 |
0 |
5.1 |
| Roeseliana roeselii (Hagenbach, 1822) |
0 |
0 |
0 |
0 |
0 |
0.2 |
| Stenobothrus lineatus (Panzer, 1796) |
0 |
0 |
0 |
0 |
0 |
0.3 |
| Tetrix subulata (Linnaeus, 1758) |
0 |
0 |
0.1 |
0.2 |
0 |
0.8 |
| Tetrix tenuicornis (Sahlberg, 1891) |
0 |
0 |
0 |
5.9 |
0.6 |
10 |
| Total abundance |
4.7 |
1.1 |
5.1 |
14.8 |
6.7 |
24.0 |
| Species number per sample plot |
3 |
1 |
4 |
7 |
6 |
9 |
3.3. Occurrence of Orthopterans in the Lawns with Different Properties
For the most frequent species,
Chorthippus biguttulus, we found no correspondence with any lawn properties (
).
Chorthippus dorsatus tended to prefer lawns with sward herb cover.
Conocephalus fuscus was found more frequently in lawns with any unmowed patches—hedgerows, decorative shrubs, grasses or herbs, or ruderal weeds. Also, this species occurred more frequently in sites with sward herb cover. We did not find the impact of the degree of isolation from suburbs and large urban meadows on any orthopterans. For a single species of dry meadows,
Ch. mollis, we didn’t obtain adequate
glm due to the low number of occupied sites. Yet, it was found almost only in unshaded sites (Fisher’s test
p < 0.0001).
.
Influence of some properties of lawns on occurrences of the most frequent orthopterans (acoustic and visual observations, generalized linear models with binary distribution). Bold p-values are statisticaly significant.
| Species |
Parameters |
Gradation |
Estimate |
Std.Error |
z |
p-Value |
| C. fuscus (R2 = 0.35) |
(Intercept) |
|
−2.88 |
1.39 |
−2.08 |
0.0379 |
| Shading |
yes/no |
0.32 |
1.53 |
0.21 |
0.8315 |
| Herb density |
yes/no |
2.46 |
0.95 |
2.60 |
0.0094 |
| Unmowed patches |
yes/no |
4.03 |
1.02 |
3.97 |
0.0001 |
| Degree of isolation |
1 to 4 |
−0.10 |
0.55 |
−0.18 |
0.8575 |
| Ch. biguttulus (R2 = 0.11) |
(Intercept) |
|
17.71 |
1388.17 |
0.01 |
0.9900 |
| Shading |
yes/no |
−17.15 |
1388.17 |
−0.01 |
0.9900 |
| Herb density |
yes/no |
0.20 |
0.74 |
0.27 |
0.7850 |
| Unmowed patches |
yes/no |
−0.38 |
0.71 |
−0.54 |
0.5890 |
| Degree of isolation |
1 to 4 |
−0.10 |
0.35 |
−0.28 |
0.7780 |
| Ch. dorsatus (R2 = 0.11) |
(Intercept) |
|
0.65 |
0.99 |
0.66 |
0.5116 |
| Shading |
yes/no |
−0.61 |
1.03 |
−0.59 |
0.5557 |
| Herb density |
yes/no |
1.53 |
0.72 |
2.13 |
0.0331 |
| Unmowed patches |
yes/no |
0.11 |
0.79 |
0.14 |
0.8860 |
| Degree of isolation |
1 to 4 |
−0.38 |
0.36 |
−1.06 |
0.2893 |
4. Discussion
Colonisation of urban habitats by Orthoptera can be regarded as a combination of two processes: (1) colonisation of the urban landscape as the set of all habitats closed by dense buildings and sealed surfaces, and (2) colonisation of lawns as man-made and the most specific for city habitats. Alternatively, we can divide the recruitment of Orthoptera from the regional pool to the urban landscape into two steps: (1) introduction to urban area and (2) expansion to lawns as specific urban habitats.
- (1)
-
The species composition of Orthoptera in Kaluga city is diverse, but species of dry meadows are mainly eliminated, so the spectrum of habitat groups is narrower than in the suburbs. There are many sites with short and sparse herbage and bare ground, which are like dry sandy habitats in the suburbs. But they are generally shaded by buildings and trees, and frequent disturbances of their ground delay the normal development of orthopteran eggs and nymphs. This corresponds to knowledge about some ground-dweller arthropods, which sense urban parks as not heat but cold habitats [17]. A single dry-meadow-inhabitant which colonises urban habitats, Chorthippus mollis, has a high density in the suburbs, high dispersal ability and relatively small size, which allows its specimens to survive in small habitat patches. It prefers warm and sunny habitats [68]. It can eat a wide spectrum of plants [69]. In Kaluga city, it lives not only in Festuca-dominated sites like in the suburbs but also in Calamagrostis epigejos—Poa pratensis associations if they are not shaded and herbage is not too dense. So, we suggest that colonisation of urban habitats by this grasshopper is limited by their temperature and their radiation regime. Other dry-meadow specialists that did not develop the urban landscape—Oedipoda caerulescens and Calliptamus italicus—also have broad food niches [55,70].
- (2)
-
Lawns with frequent mowing of herbage are known to be unfavourable habitats for Orthoptera, especially for Ensifera [15,71]. Our results show that the environment of urban lawns makes the spectrum of layer groups narrower. Both strong plant-inhabitants (specialised phytophiles) and ground-dwellers (geophiles) are eliminated. It is caused by frequent disturbances in both the herb layer and soils, so only layer-mobile species can survive. Many layer-mobile orthopterans are also disadvantaged by lawns. The most noticeable katydid of the city, Tettigonia cantans, living in the crowns of trees and shrubs in the built area, is not an inhabitant of lawns because its eggs develop in the soil, and nymphs develop in the herb layer [72]. Life cycles of listed species of Tettigoniidae may be long (usually biennial) due to embryonic dormancy depending on soil temperatures [73,74], so they are sensitive to disturbances.
A single species of katydids which can be found quite frequently in urban lawns of Kaluga city is
Conocephalus fuscus. In other countries, it also inhabits road verges with tall grasses, urban wastelands, and ruderal sites [
75]. We propose that it has some preadaptations to the urban environment: (1) High dispersal ability due to long wings. (2) Short, annual life cycle because eggs develop without initial embryonic dormancy [
76]. (3) It lays eggs into shoots of grasses and sedges [
75], so it is less sensitive to soil disturbances. (4) It has a small gracile body so that it can keep on single stems of tall herbs or shrubs for a long time. In Kaluga city, some specimens have been observed in one place for a month or longer (see dataset [
53], occurrenceID 919). The short time of movement of its specimens minimises the probability of them being destroyed by cars and other machines. We did not exclude the possibility that the lower quality of urban habitats caused by mowing can be compensated for by a lower pressure of predators, e.g., insect-eating birds, in urban environments [
62].
Do orthopterans live on a lawn? It depends on the quality of a lawn, including herb density and the presence of uncut patches. Our data now does not allow us to decide if urban lawns are enough for the realisation of the life cycles of orthopterans or not. It is possible that it is not even for the most common orthopterans (
Chorthippus biguttulus,
Ch. dorsatus, and
Conocephalus fuscus). Some sites can be inhabited only by adult specimens. This can be one of the explanations for the fact that all lawn-tolerant species are late-emerging and high-dispersal.
The results show that of the 20 species observed in the urban landscape, only one species inhabits lawns of different quality. It is
Chorthippus biguttulus. Its association with mowed habitats was also noted in western parts of the Continental biogeoregion [
77]. High mobility probably allows it to survive in severe urban environments [
78].
The recruitment of listed species to the most transformed habitats of Kaluga looks to be expected based on papers about Orthoptera of other regions of Europe [
30,
32,
38,
40].
Fragmentation is known to be an important factor in the diversity of orthopterans [
72,
79]. In Kaluga, we didn’t find the negative influence of the isolation of lawns for any species. The key role of habitat quality in the shaping of urban assemblages of Orthoptera corresponds to the paper [
38].
It is known that leaving uncut refuges increases the diversity of Orthoptera [
15,
79,
80]. Our results support this notion and show that some species in cities need warm sites with sparse vegetation. This means that we can support a relatively high level of orthopteran diversity in the urban landscape using simple management practices. We should provide contrasting habitats: regularly mowed dry, unshaded sites and rarely-mowed sites with tall herbs and shrubs.
5. Conclusions
(1) There is a diverse complex of Orthoptera within the urban landscape of Kaluga, but it is almost deprived of dry meadow species. (2) Lawns are deprived of ground-dwelling species and have low species numbers and low total abundance of Orthoptera. Lawns are colonised by species with high dispersal ability. There are no species which prefer lawns to meadows in Kaluga. (3) Presence of Orthoptera in lawns is influenced by the density of the herb layer and the presence of uncut patches.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/568,
Table S1: List of sample plots with geographical coordinates.
Appendix A. Ecological Traits of Orthopteran Species Accepted for Kaluga City
Table A1.
Ecological traits of orthopteran species accepted for Kaluga city. For designation see Section 2.4.
| Species |
Habitats in Suburb |
Phenology |
Dispersal Ability |
Layer Group |
| Barbitistes constrictus Brunner von Wattenwyl, 1878 |
forest |
late |
1 |
2 |
| Bicolorana bicolor (Philippi, 1830) |
dry meadow |
early |
2 |
2 |
| Calliptamus italicus (Linnaeus, 1758) |
dry meadow |
late |
2 |
3 |
| Chorthippus apricarius (Linnaeus, 1758) |
meadow |
early |
2 |
2 |
| Chorthippus biguttulus (Linnaeus, 1758) |
meadow |
late |
3 |
2 |
| Chorthippus brunneus (Thunberg, 1815) |
forest |
early |
3 |
2 |
| Chorthippus dorsatus (Zetterstedt, 1821) |
meadow |
late |
3 |
2 |
| Chorthippus mollis (Charpentier, 1825) |
dry meadow |
late |
3 |
2 |
| Chrysochraon dispar (Germar, 1834) |
meadow (moist) |
early |
2 |
1 |
| Conocephalus fuscus (Fabricius, 1793) |
meadow |
late |
3 |
1 |
| Decticus verrucivorus (Linnaeus, 1758) |
meadow |
early |
1 |
3 |
| Euthystira brachyptera (Ocskay, 1826) |
meadow |
early |
2 |
1 |
| Gomphocerippus rufus (Linnaeus, 1758) |
forest |
late |
2 |
2 |
| Leptophyes albovittata (Kollar, 1833) |
no data |
late |
3 |
2 |
| Locusta migratoria (Linnaeus, 1758) |
no accessed |
late |
3 |
3 |
| Metrioptera brachyptera (Linnaeus, 1761) |
meadow |
early |
2 |
1 |
| Oedipoda caerulescens (Linnaeus, 1758) |
dry meadow |
late |
3 |
4 |
| Omocestus haemorrhoidalis (Charpentier, 1825) |
dry meadow |
early |
1 |
3 |
| Omocestus viridulus (Linnaeus, 1758) |
meadow (moist) |
early |
2 |
2 |
| Phaneroptera falcata (Poda, 1761) |
meadow |
late |
3 |
1 |
| Pholidoptera griseoaptera (De Geer, 1773) |
forest |
late |
1 |
3 |
| Poecilimon intermedius (Fieber, 1853) |
meadow |
early |
1 |
2 |
| Pseudochorthippus parallelus (Zetterstedt, 1821) |
meadow |
early |
2 |
2 |
| Roeseliana roeselii (Hagenbach, 1822) |
dry meadow |
early |
1 |
2 |
| Stenobothrus lineatus (Panzer, 1796) |
dry meadow |
early |
2 |
2 |
| Stethophyma grossum (Linnaeus, 1758) |
meadow (wet) |
late |
2 |
2 |
| Tetrix bipunctata (Linnaeus, 1758) |
no data |
overwinter |
2 |
4 |
| Tetrix subulata (Linnaeus, 1758) |
riparian |
overwinter |
1 |
4 |
| Tetrix tenuicornis (Sahlberg, 1891) |
meadow |
overwinter |
3 |
4 |
| Tettigonia cantans (Fuessly, 1775) |
meadow |
early |
1 |
2 |
Acknowledgments
I am grateful to Dmitry Gusarov, Polina Shmeleva, Artem Markov and other students of the Regional eco-biological centre who helped me to collect the insects.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Research data are available in the datasets [
52,
53].
Funding
This research received no external funding.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
McIntyre NE. The ecology of urban arthropods: A review and a call to action.
Ann. Entomol. Soc. Am. 2000,
93, 825–835.
[Google Scholar]
-
2.
McKinney ML. Urbanization as a major cause of biotic homogenization.
Biol. Conserv. 2006,
127, 247–260.
[Google Scholar]
-
3.
Fenoglio MS, Calviño A, González E, Salvo A, Videla M. Urbanisation drivers and underlying mechanisms of terrestrial insect diversity loss in cities.
Ecol. Entomol. 2021,
46, 757–771.
[Google Scholar]
-
4.
McKinney ML. Effects of urbanization on species richness: a review of plants and animals.
Urban Ecosyst. 2008,
11, 161–176.
[Google Scholar]
-
5.
Luck GW, Smallbone LT. Species diversity in urban landscapes: Patterns, drivers and implications. In Urban Ecology; Gaston K, Ed.; Cambridge University Press & British Ecological Society: London, UK, 2010; pp. 88–119.
-
6.
Concepción ED, Moretti M, Altermatt F, Nobis MP, Obrist MK. Impacts of urbanisation on biodiversity: The role of species mobility, degree of specialisation and spatial scale.
Oikos 2015,
124, 1571–1582.
[Google Scholar]
-
7.
Adams LW. Urban wildlife ecology and conservation: A brief history of the discipline.
Urban Ecosyst. 2005,
8, 139–156.
[Google Scholar]
-
8.
Chiari C, Dinetti M, Licciardello C, Licitra G, Pautasso M. Urbanization and the more-individuals hypothesis.
J. Anim. Ecol. 2010,
79, 366–371.
[Google Scholar]
-
9.
Meffert PJ, Dziock F. The influence of urbanisation on diversity and trait composition of birds.
Landsc. Ecol. 2013,
28, 943–957.
[Google Scholar]
-
10.
Ives CD, Lentini PE, Threlfall CG, Ikin K, Shanahan DF, Garrard GE, et al. Cities are hotspots for threatened species.
Glob. Ecol. Biogeogr. 2016,
25, 117–126.
[Google Scholar]
-
11.
Rebele F. Urban ecology and special features of urban ecosystems.
Glob. Ecol. Biogeogr. Lett. 1994,
4, 173–187.
[Google Scholar]
-
12.
Robinson WH. Urban Insects and Arachnids: A Handbook of Urban Entomology; Cambridge University Press: Cambridge, UK, 2005; 476p.
-
13.
Angold PG, Adler JP, Hill MO, Pullin A, Rushton S, Austin K, et al. Biodiversity in urban habitat patches.
Sci. Total Environ. 2006,
360, 196–204.
[Google Scholar]
-
14.
Kim HH. Urban heat island.
Int. J. Remote Sens. 1992,
13, 2319–2336.
[Google Scholar]
-
15.
Klausnitzer B. Ökologie der Großstadtfauna; G. Fischer: Jena, Germany, 1993; 454p.
-
16.
Banaszak-Cibicka W. Are urban areas suitable for thermophilic and xerothermic bee species (Hymenoptera: Apoidea: Apiformes)?
Apidologie 2014,
45, 145–155.
[Google Scholar]
-
17.
Bujan J, Bertelsmeier C, Ješovnik A. Insects in temperate urban parks face stronger selection pressure from the cold than the heat.
Ecol. Evol. 2024,
14, e11335.
[Google Scholar]
-
18.
Szabó B, Korányi D, Gallé R, Lövei GL, Bakonyi G, Batáry P. Urbanization decreases species richness, and increases abundance in dry climates whereas decreases in wet climates: a global meta-analysis.
Sci. Total Environ. 2023,
859, 160145.
[Google Scholar]
-
19.
Sultana M, Corlatti L, Storch I. Are the same factors determining biodiversity in cities across different regions? Comparing drivers of urban bird richness patterns in Southern Asia
vs. Western Europe.
Urban Ecosyst. 2023,
26, 1545–1557.
[Google Scholar]
-
20.
Dearborn DC, Kark S. Motivations for conserving urban biodiversity.
Conserv. Biol. 2010,
24, 432–440.
[Google Scholar]
-
21.
Samways MJ, Lockwood JA. Orthoptera conservation: pests and paradoxes.
J. Insect Conserv. 1998,
2, 143–149.
[Google Scholar]
-
22.
Latchininsky A, Sword G, Sergeev M, Cigliano MM, Lecoq M. Locusts and grasshoppers: Behavior, ecology, and biogeography.
Psyche 2011,
2011, 578327.
[Google Scholar]
-
23.
Byerly AR, Firneno TJ, Beard R, Larson EL. The role of community science in orthopteran research.
J. Orthoptera Res. 2023,
32, 109–114.
[Google Scholar]
-
24.
Fartmann T, Krämer B, Stelzner F, Poniatowski D. Orthoptera as ecological indicators for succession in steppe grassland.
Ecol. Indic. 2012,
2, 337–344.
[Google Scholar]
-
25.
Hochkirch A, Nieto A, García Criado M, Cálix M, Braud Y, Buzzetti FM, et al. European Red List of Grasshoppers, Crickets and Bush-Crickets; Publications Office of the European Union: Luxembourg, 2016; 88p.
-
26.
Song H. Biodiversity of orthoptera.
Insect Biodivers. Sci. Soc. 2018,
2, 245–279.
[Google Scholar]
-
27.
Schweiger H. Die Insektenfauna des Wiener Stadtgebietes als Beispiel einer Kontinentalen Grossstadtfauna. In XI Internationaler Kongress für Entomologie (1960); Organisationskomittee des XI. Internationalan Kongresses für Entomologie: Wien, Austria, 1962; Bd. 3, pp. 184–193.
-
28.
Ingrisch S. Zur Orthopterenfauna der Stadt Giessen (Hessen) (Saltatoria, Dermaptera und Blattoptera).
Entomol. Z. 1980,
90, 275–280.
[Google Scholar]
-
29.
Klausnitzer B, Klausnitzer U. Bemerkungen zur Faldheuschrecken–Fauna des Stadzentrums von Leipzig (Caelifera, Acrididae).
Entomol. Nachr. Ber. 1982,
26, 133–134.
[Google Scholar]
-
30.
Richter K, Klausnitzer B. On the orthopteran fauna (Blattaria, Dermaptera, Ensifera, Caelifera) of rural places with different urban influence in Leipzig (Investigations on an urban AE gradient, 5th contribution).
Entomol. Nachrichten Und Berichte 1987,
31, 163–167.
[Google Scholar]
-
31.
Sergeev MG. Patterns of Orthoptera communities formation in urbocoenoses.
Zhurnal Obs. Biol. 1987,
48, 230–237.
[Google Scholar]
-
32.
Nagy B. Orthoptera species and assemblages in the main habitat types of some urban areas in the Carpathian Basin.
Biologia 1997,
52, 233–240.
[Google Scholar]
-
33.
Penone C, Kerbiriou C, Julien J-F, Julliard R, Machon N, Le Vio I, et al. Urbanisation effect on orthoptera: which scale matters?
Insect Conserv. Divers. 2013,
6, 319–327.
[Google Scholar]
-
34.
Sergeev MG. Orthopteran assemblage transformations in urban and agricultural landscapes of extra-tropical Eurasia.
Euroasian Entomol. J. 2014,
13, 6–10.
[Google Scholar]
-
35.
Cherrill A. Large–scale spatial patterns in species richness of orthoptera in the greater London area, United Kingdom: relationships with land cover.
Landsc. Res. 2015,
40, 476–485.
[Google Scholar]
-
36.
Melliger RL, Rusterholz HP, Baur B. Habitat- and matrix-related differences in species diversity and trait richness of vascular plants, Orthoptera and Lepidoptera in an urban landscape.
Urban Ecosyst. 2017,
20, 1095–1107.
[Google Scholar]
-
37.
Piano E, Souffreau C, Merckx T, Baardsen LF, Backeljau T, Bonte D, et al. Urbanization drives cross–taxon declines in abundance and diversity at multiple spatial scales.
Glob. Chang. Biol. 2020,
26, 1196–1211.
[Google Scholar]
-
38.
Huchler K, Pachinger B, Kropf M. Management is more important than urban landscape parameters in shaping orthopteran assemblages across green infrastructure in a metropole.
Urban Ecosyst. 2022,
26, 209–222.
[Google Scholar]
-
39.
Ancillotto L, Labadessa R. Functional traits drive the fate of Orthoptera in urban areas.
Insect Conserv. Divers. 2024,
17, 304–311.
[Google Scholar]
-
40.
Pernat N, Buchholz S, Schirmel J. Urbanization reduces Orthoptera diversity and changes community structure towards mobile species.
Insect Conserv. Divers. 2024,
17, 259–272.
[Google Scholar]
-
41.
Chernyakhovsky ME. The fauna of katydids and grasshoppers of Moscow. In Rastitelnost i zhivotnoje naselenije Moscvy i Podmoscovya; Moscow State University: Moscow, Russia, 1978; pp. 35–36.
-
42.
Chernyakhovsky ME. Species composition and Orthoptera Insect Communities of the Moscow City. In Urboecosystems: Problems and Prospects of Development: Proceedings of the V International Scientific Conference; Ishim State Pedagogical Institute: Ishim, Russia, 2010; pp. 176–178.
-
43.
Schkarlet KY, Likhovid AA. The fauna and the population or orthopterans (Orthoptera) of Stavropol.
Vestn. Sev. –Kavk. Gos. Tekhnicheskogo Univ. 2010,
22, 40–44.
[Google Scholar]
-
44.
Tereschenko DA. Orthopterans (Orthoptera) of the Donetsk city and surroundings. In Donetskie Chteniya (Donetsk, May 2016); Southern Federal University: Rostov-on-Don, Russia, 2016; pp. 162–165.
-
45.
Aleksanov VV. Inventory of the Orthoptera (Insecta, Orthoptera) of Kaluga City. In Studies in Biological Diversity of Kaluga Region; OOO “TPS” Publ.: Tambov, Russia, 2019; pp. 101–119.
-
46.
Biogeographical Regions Europe 2016. 3. Release Date: 2016–1–08. Available online: https://www.eea.europa.eu/data–and–maps/data/biogeographical–regions–europe–3 (accessed on 10 September 2020).
-
47.
Lavrenko EM. The principles and units of geobotanical regionalization. In Geobotanicheskoye Raionirovaniye SSSR; Nauka: Moscow, Russia; Saint Petersburg, Russia, 1947; pp. 9–13.
-
48.
Reshetnikova NM, Mayorov SR, Skvortsov AK, Krylov AV, Voronkina NV, Popchenko MI, et al. Flora of Kaluga Oblast: Annotated List of Vascular Plants of Kaluga Oblast; KMK: Moscow, Russia, 2010; 548p.
-
49.
Pukhov VA. History of Kaluga City; Zolotaya Alleya: Kaluga, Russia, 2006; 160p.
-
50.
Barashkova ZK, Lavrovich ON, Shuleshkina EN. The Geological Maps of the Quaternary Sediments in Kaluga Region. Scale 1:500,000; Central Geological Center: Moscow, Russia, 1998; 1p.
-
51.
EUNIS Habitat Type Hierarchical View (Marine Version 2022 & Terrestrial Version 2021). Available online: https://eunis.eea.europa.eu/habitats–code–browser–revised.jsp (accessed on 18 September 2024).
-
52.
Aleksanov V, Karmazina I, Shulaev N, Ruchin A, Lukiyanov S, Lobachev E, et al. Orthoptera and Mantodea in the Continental Biogeographical Region and Adjacent Areas of European Russia. Version 1.15. Occurrence Dataset. Available online: https://doi.org/10.15468/xtpy3y (accessed on 9 July 2024).
-
53.
Aleksanov V. Orthoptera Sampled in Kaluga Region (European Russia) in 2024. Version 1.1. Occurrence Dataset. Available online: https://doi.org/10.15468/b4t6rx (accessed on 16 February 2025).
-
54.
Bey-Bienko GJ. Orthoptera. In Keys to Insects of European Part of USSR; Nauka: Moscow, Russia; Saint Petersburg, Russia, 1964; pp. 205–284.
-
55.
Bey-Bienko GJ, Mistshenko LL. Locusts and Grasshoppers of the USSR and Adjacent Countries; USSR Academy of Science Publ.: Moscow, Russia; Saint Petersburg, Russia, 1951; Volume 1–2; 667p.
-
56.
Bukhvalova MA. Acoustic signals and morphological features of some grasshoppers of the Chorthippus biguttulus group (Orthoptera, Acrididae) of Russia and adjacent territories.
Entomol. Rev. 1995,
74, 56–67.
[Google Scholar]
-
57.
Benediktov AA. To little-known taxa of
Chorthippus biguttulus group (Orthoptera, Acrididae, Gomphocerinae).
Vestn. Mosk. Univ. Seriya XVI Biol. 1999,
1, 42–45.
[Google Scholar]
-
58.
Willemse F, von Helversen O, Odé B. A review of
Chorthippus species with angled pronotal lateral keels from Greece with special reference to transitional populations between some Peloponnesean taxa (Orthoptera, Acrididae).
Zool. Meded. 2009,
83, 319–508.
[Google Scholar]
-
59.
Tarasova T, Tishechkin D, Vedenina V. Songs and morphology in three species of the Chorthippus biguttulus group (Orthoptera, Acrididae, Gomphocerinae) in Russia and adjacent countries.
ZooKeys 2021,
1073, 21–53.
[Google Scholar]
-
60.
Cigliano MM, Braun H, Eades DC, Otte D. Orthoptera Species File. Version 5.0/5.0. Available online: http://Orthoptera.SpeciesFile.org (accessed on 30 June 2024).
-
61.
Reinhardt K, Köhler G, Maas S, Detzel P. Low dispersal ability and habitat specificity promote extinctions in rare but not in widespread species: the Orthoptera of Germany.
Ecography 2005,
28, 593–602.
[Google Scholar]
-
62.
Gardiner T, Hill J. Directional dispersal patterns of Chorthippus parallelus (Orthoptera: Acrididae) in patches of grazed pastures.
J. Orthoptera Res. 2004,
13, 135–141.
[Google Scholar]
-
63.
Löffler F, Poniatowski D, Fartmann T. Orthoptera community shifts in response to land–use and climate change—Lessons from a long–term study across different grassland habitats.
Biol. Conserv. 2019,
236, 315–323.
[Google Scholar]
-
64.
Pravdin FN. Ecological Geography of the Insects of Central Asia; Orthopteroids. Nauka: Moscow, Russia, 1978; 272p.
-
65.
Storozhenko SY. Long–Horned Orthopterans (Orthoptera: Ensifera) of the Asiatic Part of Russia; Dal’nauka: Vladivostok, Russia, 2004; 280p.
-
66.
R Version 4.1.3. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: https://www.R–project.org/ (accessed on 22 April 2024).
-
67.
Wang WJ, Thompson FR III, He HS, Fraser JS, Dijak WD, Jones-Farrand T. Climate Change and Tree Harvest Interact to Affect Future Tree Species Distribution Changes.
J. Ecol. 2019,
107, 1901–1917.
[Google Scholar]
-
68.
Dvořák T, Hadrava J, Knapp M. The ecological niche and conservation value of Central European grassland orthopterans: A quantitative approach.
Biol. Conserv. 2022,
265, 109406.
[Google Scholar]
-
69.
Köhler G, Held M. Eine erfolgreiche Zucht des Verkannten Grashüpfers, Chorthippus mollis (Charpentier), am Gemeinen Beifuß,
Artemisia vulgaris L.
Articulata 2000,
15, 211–215.
[Google Scholar]
-
70.
Sergeev MG, Childebaev MK, Vankova IA, Gapparov FA, Kambulin VE, Kokanova E, et al. Italian Locust Calliptamus italicus (Linnaeus, 1758): Morphology, Distribution, Ecology, Population Management; FAO: Rome, Italy, 2016; xviii + 333p.
-
71.
Braschler B, Marini L, Thommen GH, Baur B. Effects of small–scale grassland fragmentation and frequent mowing on population density and species diversity of orthopterans: a long–term study.
Ecol. Entomol. 2009,
34, 321–329.
[Google Scholar]
-
72.
Chernyakhovsky ME, Volkova LB, Sobolev NA. Tettigonia cantans. In Red Data Book of the City of Moscow, 3rd ed.; Sobolev NA, Ed.; Moscow Department of Environmental Resources Management: Moscow, Russia, 2022; pp. 298–300.
-
73.
Ingrisch S. The plurennial life cycles of the European Tettigoniidae (Insecta: Orthoptera).
Oecologia 1986,
70, 606–616.
[Google Scholar]
-
74.
Ortis G, Mazzon L, Marini L. Effect of summer temperature on prolonged diapause of Tettigoniidae (Orthoptera) under realistic field conditions.
J. Insect Physiol. 2023,
146, 104499.
[Google Scholar]
-
75.
Kleukers RMJC, Decleer K, Haes ECM, Kolshorn P, Thomas B. The recent expansion of
Conocephalus discolor (Thunberg) (Orthoptera: Tettigoniidae) in western Europe.
Entomol. Gaz. 1996,
47, 37–50.
[Google Scholar]
-
76.
Eötvös CB, Magura T, Lövei GL. A meta–analysis indicates reduced predation pressure with increasing urbanization.
Landsc. Urban Plan. 2018,
180, 54–59.
[Google Scholar]
-
77.
Hochkirch A, Papen M. Behaviour–related microhabitat utilization in
Chorthippus apricarius (Linné, 1758) and
Chorthippus biguttulus (Linné, 1758).
Mitteilungen Dtsch. Ges. Allg. Angew. Entomol. 2001,
13, 343–346.
[Google Scholar]
-
78.
Rech F, Narimanov N, Bauer T, Schirmel J. Urbanization increases fluctuating asymmetry and affects behavioral traits of a common grasshopper.
Ecol. Evol. 2022,
12, e9658.
[Google Scholar]
-
79.
Kenyeres Z, Szentirmai I. Effects of different mowing regimes on orthopterans of Central-European mesic hay meadows.
J. Orthoptera Res. 2017,
26, 29–37.
[Google Scholar]
-
80.
Kaláb O, Šipoš J, Kočárek P. Leaving uncut refuges during meadow harvesting increases the functional diversity of Orthoptera.
Entomol. Sci. 2020,
23, 95–104.
[Google Scholar]