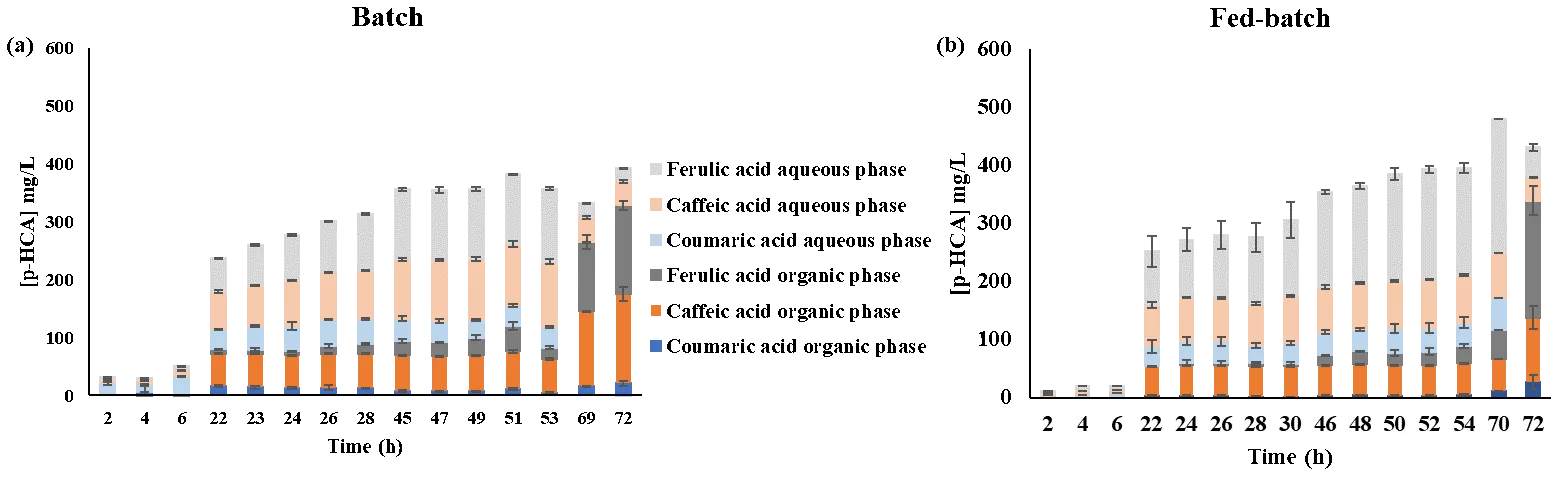1. Introduction
Ferulic acid (FA) is a secondary metabolite of the phenolic acid group that is commonly found in plant tissues. It is most commonly found in whole grains, spinach, parsley, grapes, rhubarbs, and cereal seeds. It is present in plants both in its free form and cross-linked to plant cell wall polysaccharides, lignin, and hydroxy fatty acids [
1]. FA is a precursor of a wide variety of chemicals, such as coniferyl alcohol, vanillin, sinapic acid, and curcumin [
2,
3]. Moreover, FA is an antioxidant that plays a potent role as a free radical scavenger in plants. FA has numerous applications in the food, cosmetic, and pharmaceutical industries due to its wide range of biological activities, including antimicrobial, antiallergic, and anticarcinogenic effects, as well as modulation of enzyme activity and hepatoprotection [
1,
4,
5].
Although FA can be extracted from biomass by either chemical or enzymatic hydrolysis, both approaches have some drawbacks, such as the use of hazardous solvents and difficult challenges for enzymatic extraction because most of the ferulic contents in crops are covalently linked to other biopolymers [
6]. Not only do microbial production approaches provide a promising alternative to both chemical synthesis and extraction from plant sources, but they also allow ferulic acid to be classified as a natural product [
7]. However, fermentation presents several challenges, such as the need to fine-tune the expression levels of pathway enzymes, optimize the availability of redox cofactors and metabolic precursors, and depend on the external addition of aromatic amino acids—costly components that limit industrial scalability [
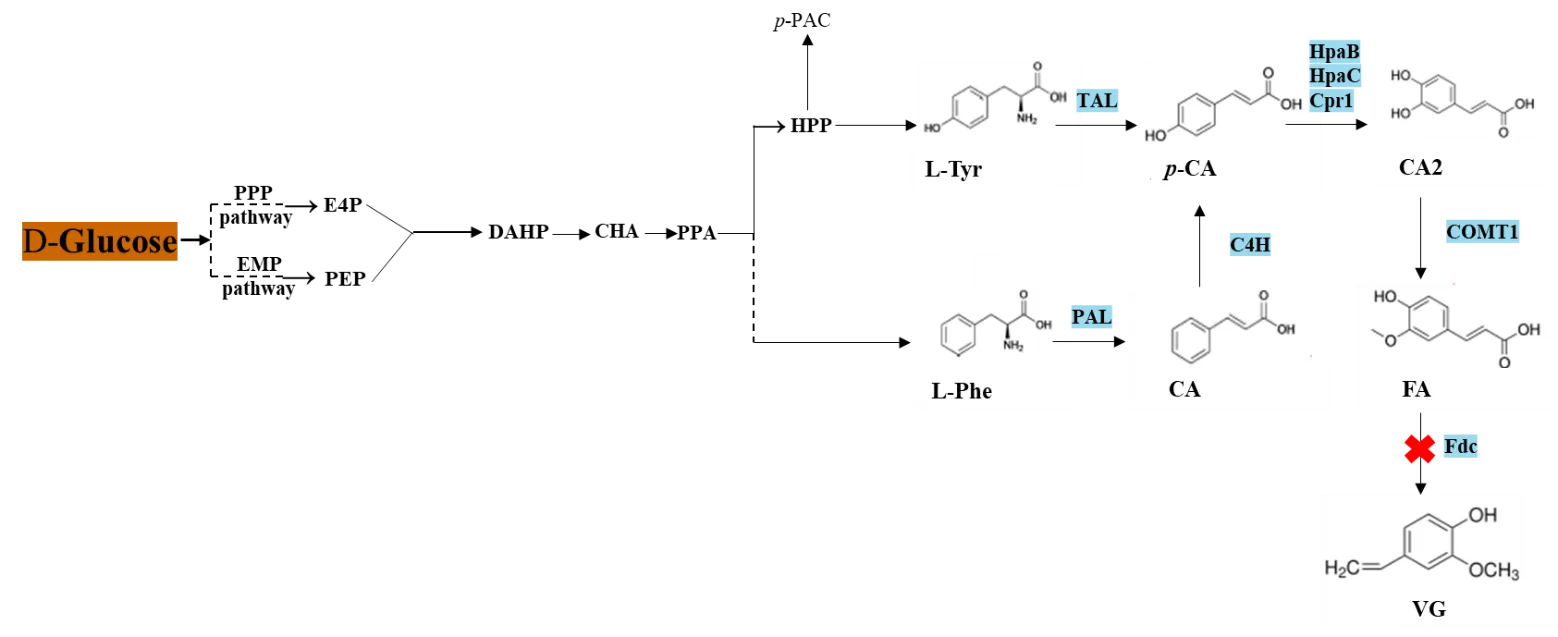
8]. FA is synthesized from
L-phenylalanine, which is converted to cinnamic acid, followed by
p-coumaric acid, caffeic acid, and finally to FA by a cascade of enzymes, namely phenylalanine ammonia lyase (PAL), cinnamate 4-hydroxylase (C4H), 4-hydroxyphenylacetate 3-hydroxylase oxygenase (HpaB), 4-hydroxyphenylacetate 3-hydroxylase reductase (HpaC), cytochrome P450 reductase (Cpr), and caffeic acid
O-methyltransferase (COMT), respectively (
). FA can also be biosynthesized from
L-tyrosine by tyrosine ammonia lyase (TAL), which catalyses the conversion of
L-tyrosine to
p-coumaric acid, which is then hydroxylated to caffeic acid, and finally, caffeic acid is methylated into ferulic acid (
).
. Schematic illustration of the ferulic acid biosynthetic pathway in <em>S. cerevisiae </em>ABG20 strain. PPP pathway: pentose phosphate pathway; EMP pathway: Embden−Meyerhof−Parnas pathway; E4P: erythrose 4-phosphate; PEP: phosphoenolpyruvate; DAHP: 3-deoxy-D-arabinoheptulosonic acid-7-phosphate; CHA: chorismic acid; PPA: prephenate; HPP: para-hydroxyphenylpyruvate; <em>p</em>-PAC: para-hydroxy-acetaldehyde; <span style="font-variant-caps:small-caps;">L</span>-Tyr: <span style="font-variant-caps:small-caps;">L</span>-tyrosine; <span style="font-variant-caps:small-caps;">L</span>-Phe: <span style="font-variant-caps:small-caps;">L</span>-phenylalanine; CA: cinnamic acid; <em>p</em>-CA: <em>p</em>-coumaric acid; CA2: caffeic acid; FA: ferulic acid; VG: vinylguaiacol; Fdc: ferulic acid decarboxylase; TAL: tyrosine ammonia-lyase; PAL: phenylalanine ammonia-lyase; C4H: cinnamate-4-hydroxylase; HpaB & HpaC: 4-hydroxyphenylacetate 3-hydroxylases; Cpr1: cytochrome P450 reductase; COMT1: caffeic acid <em>O</em>-methyltransferase.
While previous studies have made substantial progress in demonstrating the feasibility of microbial production of FA in
E. coli, the biosynthetic efficiency is still significantly hindered by challenges related to heterologous gene expression, redox factors, and the host cell’s primary precursor productivity. For example, Choi et al. reported the production of 7.1 mg·L
−1 of FA by expressing three genes:
Tal,
Sam5, and
Comt, which encode tyrosine ammonia-lyase, 4-coumarate 3-hydroxylase, and
O-methyltransferase, respectively [
8]. Independently, Kang et al. produced 196 mg·L
−1 of FA from simple carbon sources combined with
Tal codon optimization [
9]. Recently, Lv et al. increased FA production to 212 mg·L
−1 by tuning the promoter strength [
10]. Although this biotechnological production method is promising and offers opportunities for further optimization in microorganism engineering, there is a lack of research focused on optimizing the fermentation process and improving the downstream processing of FA. In this study, a yeast microorganism was used due to its status as an aromatic amino acid prototroph, allowing ferulic acid production to be achieved using only
d-glucose as the substrate without requiring any additional supplementation. Additionally, the intensification of ferulic acid production was achieved by implementing a liquid-liquid extraction method combined with a fed-batch fermentation strategy.
In a previous study on
S. cerevisiae, we reported the production of
p-coumaric acid (
p-CA), another plant phenolic acid with antimicrobial properties and low solubility, using a batch biphasic fermentation system with oleyl alcohol as an extractant [
11]. This system provides two major bioprocess benefits: (1) detoxifying the broth, thus increasing microorganism productivity and yield due to a decrease in product inhibition, and (2) overcoming the solubility limits of the synthesized products. This previous study was conducted using 2X Yeast Nitrogen Base (YNB) without amino acids [
11] in batch mode. However, this medium is expensive for fermentation, making medium optimization critical for large-scale FA production. To perform such optimization, one commonly used method is the Design of Experiments (DoE) and Response Surface Methodology (RSM), which are statistical tools that allow the determination of the optimum value for each process parameter evaluated [
12]. In addition, one of the key steps in designing a cost-effective bioprocess, the fed-batch cultivation strategy, is widely used in industrial bioproduction and is considered the first choice to achieve high productivity levels [
13]. Limited feed profiles of
d-glucose for
S. cerevisiae fed-batch cultivation have been explored to maximize the carbon flux from
d-glucose to the targeted product, reducing
d-glucose repression and the Crabtree effect [
14]. For example, fed-batch fermentation using
d-glucose resulted in a two-fold increase in resveratrol concentration compared to batch fermentation (415.6 mg·L
−1 vs. 235.6 mg·L
−1, respectively) [
15]. Similarly, Scalcinati et al. described the production of α-Santalene in a recombinant
S. cerevisiae strain using a fed-batch process, allowing the production of 92 mg·L
−1 [
16]. To the best of our knowledge, there has been no study on the production of FA from only
d-glucose using
S. cerevisiae and a fed-batch extractive fermentation strategy.
The objective of this study was to produce ferulic acid (FA) from
d-glucose using an engineered
Saccharomyces cerevisiae ABG20 strain, with the goal of developing a more economically viable medium and fermentation process. Given the hydrophobic nature of FA, which limits its accumulation in the aqueous phase, we initially evaluated FA production in both defined (YNB) and complex (YEPD) media. Finally, an appropriate bioprocessing approach was implemented, involving the cultivation of S. cerevisiae using both batch and fed-batch biphasic fermentation strategies. This work proposes a medium formulation for the heterologous production of FA from
d-glucose and demonstrates its application in extractive fed-batch fermentation.
2. Materials and Methods
2.1. Strain and Culture Media
2.1.1. ABG20: Engineered
S. cerevisiae Strain
The engineered strain ABG20 was constructed and supplied by Abolis Biotechnology (Évry, France). This strain was engineered to produce
de novo ferulic acid. ABG20 strain was derived from the
Saccharomyces cerevisiae parental strain FY1679 (genotype: MATα ura3-52/ura3-52 trp1Δ63/TRP1 leu2Δ1/LEU2 his3Δ200/HIS3 GAL2/GAL2). Strain ABG20 corresponds to an auxotrophic α-type haploid with the following genotype: MATα ura3-52 trp1Δ63 leu2Δ1 his3Δ200 fdc1Δ. The
Fdc1 gene encoding ferulic acid decarboxylase was deleted, and one copy of the following genes was introduced under the strong promoters:
PaHpaB-
SeHpaC-URA encoding 4-hydroxyphenylacetate 3-hydroxylase oxygenase from
Pseudomonas aeruginosa (
PaHpaB-PKG21040.1) and 4-hydroxyphenylacetate 3-hydroxylase reductase from
Salmonella enterica (
SeHpaC-GAR62209.1), was inserted into chromosome XI under the HHF1 and CCW12 promoters, respectively, using URA3 as selection marker.
TAL4mopti-
palev-
C4Hev-TRP encoding tyrosine ammonia lyase (
TAL-AGZ04575.1) and phenylalanine ammonia lyase (
PAL-NP_190894.1), cinnamate 4-hydroxylase (
C4H-NP_180607.1), was inserted into chromosome X under TEF1, CCW12, and TDH3 promoters, respectively, using TRP1 as the marker.
Cpr1-LEU2, which encodes cytochrome P450 reductase (
CPR1-CAA49446.1), was integrated into chromosome XII under the TEF1 promoter, using LEU2 as the marker. Caffeic acid
O-methyltransferase (
COMT-At5g54160) was integrated into chromosome XV under the TEF1 promoter, using HIS as the marker. The final genotypes of ABG20 were Δ
Fdc1,
paHpaB-
SeHpaC-URA,
TAL4mopti-
palev-
C4Hev-
TRP,
Cpr1-LEU, and
COMT1-HIS.
shows a schematic illustration of the modifications carried out in the ABG20 strain.
2.1.2. Media
Yeast Nitrogen Base medium without amino acids (YNB
w/
o aa) was bought from Sigma Aldrich,
Saint-Quentin-Fallavier, France. A 10X stock solution (67 g·L
−1) was prepared according to the manufacturer’s instructions and sterilized by filtration using 0.2 μm filters. The final concentration of the YNB in the medium was 2X (13.4 g·L
−1) with 20 g·L
−1 of
d-glucose (Fischer Scientific, anhydrous, 99%), 0.04 M of KH
2PO
4 (Fisher Scientific, Illkirch Graffenstaden, France) and 0.04 M of K
2HPO
4 (Alfa Aesar, Schiltigheim, France). The Yeast Peptone Dextrose (YEPD) medium consisted of 10 g·L
−1 of peptone (Fisher Scientific), 10 g·L
−1 of yeast extract (Fisher Scientific) and 20 g·L
−1 of
d-glucose (Fisher Scientific).
2.2. FA Production in Flasks
YNB and YEPD Media
Shake flask cultures were carried out in 500-mL baffled flasks containing 50 mL of either YNB 2X medium or YEPD medium and incubated at 30 °C with stirring (180 rpm) for 72 h. Samples were taken every 24 h to monitor OD
600 nm, and follow
p-coumaric, caffeic and ferulic acids production.
2.3. Batch and Fed-Batch Biphasic Mode in Bioreactor for FA Production
For every experiment, the strain was cultivated in a 500-mL baffled Erlenmeyer flask containing 80 mL of YEPD medium overnight at 30 °C and 180 rpm from an inoculum preserved at −80 °C. Bioreactors were inoculated with pre-culture at an initial optical density (OD) of 0.5 at 600 nm (OD
600 nm). Experiments were performed in duplicate.
Batch biphasic experiments were performed in 1.5 L bioreactors (Global Process Concept (GPC), La Rochelle, France) with a PRO-LAB
TM controller unit and C-BIO2
TM control software as previously described [
11]. The working volume was 500 mL of medium fermentation, and 500 mL of oleyl alcohol (
Z-octadec-9-en-1-ol, Merck KGaA, Darmstadt, Germany). In order to avoid a water/oil emulsion, a sintered-microsparger (GPC, La Rochelle, France) was used for aeration at a rate of 0.1 L·min
−1. Agitation speed was fixed at 75 rpm, and 0.004% (
v/
v) of Tween 80 was added when necessary (4 h after the beginning of the fermentation process). A pH level of 6.5 was maintained using KOH 1 M or H
2SO
4 0.5 M. ABG20 strain was cultivated in a medium containing 20 g·L
−1 of
d-glucose (Fisher Scientific, anhydrous, 99%), 10 g·L
−1 of peptone (from Fisher Scientific), 10 g·L
−1 of yeast extract (Fisher Scientific), 1 g·L
−1 of (NH
4)
2SO
4, 4 g·L
−1 of KH
2PO
4 (Fisher Scientific, France) and 2 g·L
−1 of K
2HPO
4 (Alfa Aesar, France).
Fed-batch biphasic experiments were conducted using the same physical parameters as batch experiments. Feeding with the fresh medium was started only after residual ethanol produced from
d-glucose consumption phase was totally consumed. Fresh medium was composed of 200 g·L
−1 of
d-glucose, 10 g·L
−1 of (NH
4)
2SO
4, 40 g·L
−1 of KH
2PO
4 and 20 g·L
−1 of K
2HPO
4. A flow rate of 2 g·h
−1 was used. At the end of the fermentation, 100 mL of fed-batch solution was used. As the working volume of the fermenters was 1 L, we reduced the initial working volume for the fermentation medium from 500 mL to 400 mL and the same volume of oleyl alcohol (400 mL) was added at the beginning of cultivation. In order to recover efficiently the ferulic acid in the solvent phase, acidification of the medium was performed after 70 h of cultivation.
2.4. Analytical Methods
2.4.1. Quantification of Dry Matter
Biomass was quantified by measuring cell dry weight (CDW). 5-mL samples of culture medium were centrifuged at 4500×
g for 10 min. Pellets were washed once with one volume of distilled water to remove any trace of the medium. They were then dried overnight at 105 °C in the oven, and their cell dry weight was measured. Yeast growth was also determined by OD
600 nm using a spectrophotometer Cary 60 UV-Vis from Agilent (Marcy l’Etoile, France).
2.4.2. Quantification of Substrate and Metabolites in Fermentation Broth
2 mL of samples were collected every 2 h during working hours for 72 h. Ferulic,
p-CA and caffeic acids were quantified using high-performance liquid chromatography (HPLC) (Thermo scientific, Illkirch Graffenstaden, France, Ultimate 3000) coupled with an ultraviolet-diode detector (UV-DAD) and the elution method was used as previously described [
17]. Sinapic acid was used as an internal standard. A 100 mg·L
−1 of sinapic acid solution in acetonitrile was prepared and kept at −20 °C and was added at 50% (
v/
v) to every aqueous sample before centrifugation (10,000×
g, 10 min). Supernatants were filtered through regenerated cellulose (RC) 0.2 µm filters. Calibration curves with five points and internal standards for ferulic,
p-CA and caffeic acids were prepared to allow their quantification.
On biphasic fermentations, organic phase samples were prepared as follows: 970 µL of methanol was added to 30 µL of the sample containing 250 µL of internal standard and then the samples were filtered and analysed by HPLC. Calibration curves for ferulic,
p-CA and caffeic acids with five points and internal standards were also performed in these conditions allowing for quantify them.
d-Glucose and ethanol were quantified in aqueous phases using HPLC (Thermo scientific, Ultimate 3000) equipped with a Shodex refractive index detector and a Thermo scientific ultraviolet detector. For more information on analytical methods, it can be useful to see our previous work [
17].
2.4.3. Statistical Analysis
The data set size in each experiment is
n = 2, so the range was used to demonstrate statistical dispersion.
3. Results
3.1. Growth and FA Production in YNB and YEPD Media
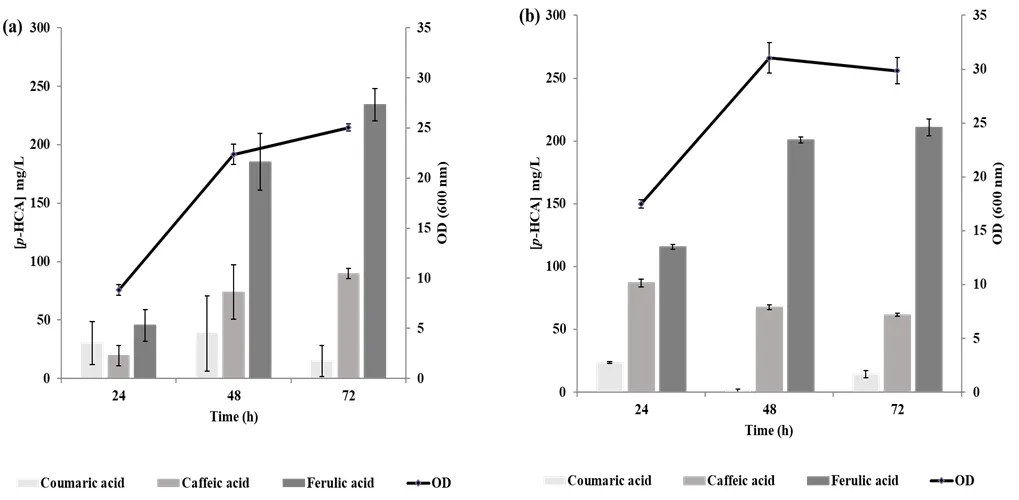
The growth and production of FA and their intermediates,
p-coumaric acid (
p-CA) and caffeic acid, by the ABG20 strain, were compared in both YNB and YEPD media. The optimal growth of ABG20 was observed in YEPD medium after 48 h of cultivation (
b), while in YNB medium, the strain continued to grow until 72 h of cultivation (
a). After 48 h of cultivation, FA production accounted for 185.0 mg·L
−1 in YNB medium and 200.1 mg·L
−1 in YEPD medium (
a,b), with corresponding productivity levels of 3.8 mg·L
−1·h
−1 and 4.2 mg·L
−1·h
−1, respectively. After 72 h of cultivation, FA production in YEPD medium showed a slight increase, while in YNB medium, FA production reached 234 mg·L
−1, though the productivity remained lower than that observed at 48 h. These findings indicate that the rich YEPD medium increases FA productivity within the first 48 h of cultivation. Notably, over the same 48-h cultivation period, about 48% of the produced
p-CA in YNB and 90% in YEPD were converted into caffeic acid. Additionally, approximately 60% of the caffeic acid in YNB and 66% in YEPD were converted into FA (
a,b).
. Strain production dynamics in YNB and YEPD media in flasks after 24, 48 and 72 h of cultivation. (<strong>a</strong>) <em>p</em>-coumaric, caffeic and ferulic acid productions in mg·L<sup>−1</sup> and OD<sub>600 nm</sub> in YNB medium. (<strong>b</strong>) <em>p</em>-coumaric, caffeic and ferulic acid productions in mg·L<sup>−1 </sup>and OD<sub>600 nm</sub> in YEPD medium. The results presented are the mean values ± SD for two independent biological replicates.
In addition to the composition of the YEPD medium, which yielded the highest ferulic acid production, we chose to supplement the formulation with (NH₄)₂SO₄ and a phosphate buffer, both of which previously demonstrated a positive impact on p-coumaric acid production in our earlier study.
3.2. Batch and Fed-Batch Biphasic Fermentation for Ferulic Acid Production
To overcome the solubility limits and the antimicrobial effect of this compound, a liquid-liquid extraction coupled with fermentation was performed. Oleyl alcohol was selected as the extractant due to its compatibility, as previously established.
illustrates the outcomes of this approach.
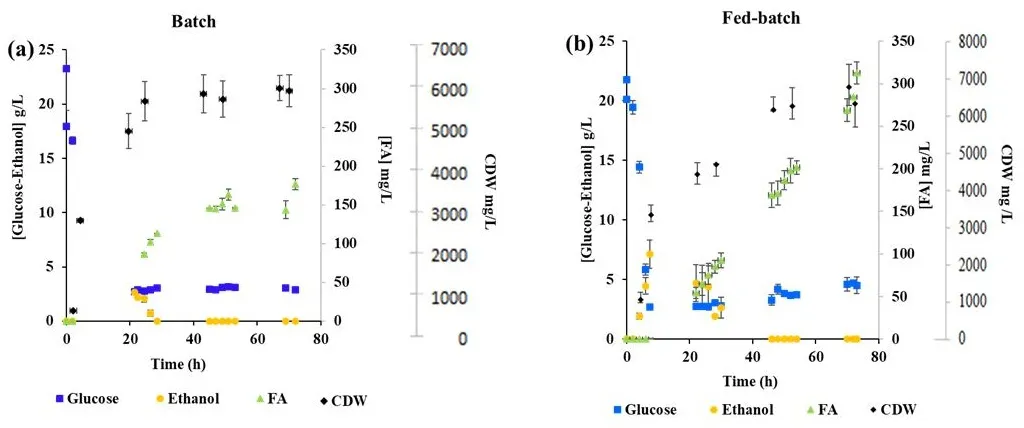
. Total production concentrations of ferulic acid (FA) (obtained from mass balance calculation in each phase and expressed per volume of broth), <span style="font-variant-caps:small-caps;">d</span>-glucose consumption, ethanol and cell dry weight (CDW) productions in batch (<strong>a</strong>) and fed-batch (<strong>b</strong>) biphasic fermentations using optimized medium. All data represent the mean of <em>n</em> = 2 independent replicates and error bars show standard deviation.
After 72 h of cultivation, the results demonstrated that FA production correlated with the growth of ABG20 in the medium, using both fermentation strategies as depicted in
a,b. Unlike the batch mode, the fed-batch mode showed no stationary phase or decline in FA production, indicating a slower metabolic rate with this approach. The final biomass levels were 6.1 g·L
−1 and 6.3 g·L
−1 for batch and fed-batch modes, respectively (
a,b). Interestingly, there was no difference in biomass production using the two fermentation strategies. However, the use of the optimized medium with yeast extract and peptone significantly improved
S. cerevisiae growth compared to the YNB medium from our previous study on
p-CA production, where only 3.5 g/L of biomass was obtained after 72 h of cultivation [
17].
In batch biphasic mode, the maximum concentration of FA reached was 176.7 mg·L
−1 (
a). FA accumulation primarily occurred after
d-glucose depletion in the batch fermentation (
a). To enhance FA production, we performed fed-batch cultivation by supplying additional
d-glucose after initial consumption of 20 g·L
−1 during the batch phase. After the complete depletion of residual ethanol produced during
d-glucose consumption, controlled feeding of
d-glucose was initiated.
Significantly higher FA production occurred during the respiratory phase, roughly between 24 h and 45 h in batch experiments and from 22 h to 72 h in fed-batch experiments, culminating in a final FA concentration of 312.6 mg·L
−1 in the fed-batch biphasic process (
b). Thus, compared to the batch process, the fed-batch approach resulted in a twofold increase in FA production. During fed-batch fermentation, the respiratory growth phase closely aligned with the feeding profile, resulting in negligible ethanol accumulation in the medium. Minimal quantities of acetic acid and glycerol were detected, while no succinate was observed throughout the fed-batch fermentation period.
The distribution of FA and related intermediates (
p-CA and caffeic acid) across batch and fed-batch fermentations is illustrated in
.
. Ferulic acid distribution in biphasic experiments in aqueous and organic phases in (<b>a</b>) batch and (<b>b</b>) fed-batch fermentations. All data represent the mean of <i>n</i> = 2 independent replicates and error bars show standard deviation.
As depicted in
, pH plays a crucial role in influencing the extraction of
p-HCAs from the fermentation broth. Throughout the initial 70 h of cultivation, regardless of the fermentation strategy,
p-HCAs (such as
p-CA, caffeic acid, and FA) were predominantly found in the aqueous phase,
i.e., the broth, when maintained at pH 6.5 (
a,b). After 70 h of cultivation, lowering the pH to 4.5 resulted in the broth maintaining very low concentrations of
p-HCAs such as
p-CA, caffeic acid, and FA (0.0, 41.7 and 23.0 mg·L
−1, respectively, in batch fermentation, and 0.0, 40.0 and 52.4 mg·L
−1, respectively, in fed-batch fermentation, represented by full bullets in
a,b.
4. Discussion
There is a growing demand for ferulic acid (FA) across various industries, such as pharmaceuticals, cosmetics, and food. Microbial fermentation offers an attractive route for FA production from inexpensive substrates, and employing suitable strategies holds significant promise. In this study, FA production in ABG20 strain was initially evaluated using two different media, a classic defined medium, 2X Yeast Nitrogen Base medium (YNB) without amino acids, and a complex rich medium, Yeast Extract Peptone Dextrose (YEPD). Each medium presents its own set of advantages and disadvantages. The YNB medium, being chemically defined, offers better control and monitoring of the fermentation process due to its consistent composition. However, its higher cost and longer development time are significant drawbacks [
18,
19]. On the other hand, the YEPD medium is widely used for fermentation processes despite its inherent variability between batches [
18,
20]. The improved growth and productivity observed in the YEPD medium can be attributed to the presence of precursors that significantly enhance the strain’s growth by conserving metabolic energy, a benefit not seen in defined media like YNB [
21]. The accumulation of caffeic acid in the ABG20 strain is likely due to the overexpression of
PaHpaB and
SeHpaC, which encode the subunits of 4HPA3H (4-hydroxyphenylacetate 3-hydroxylase), an enzyme that hydroxylates
p-CA into caffeic acid. Hernandez-Chavez and Lin have reported that the overexpression of 4HPA3H enables the efficient conversion of
p-coumaric acid to caffeic acid [
22,
23]. However, the incomplete conversion of caffeic acid suggests that the
COMT1 gene, encoding caffeate-
O-methyltransferase [
22], is a limiting factor in FA production.
In order to avoid the solubility limits and the antimicrobial properties of FA, batch and fed-batch biphasic fermentations strategies were employed. Fed-batch approach resulted in a 2-fold increase in FA production compared to the batch strategy from 22 h to 72 h. Studies have indicated that fermentative metabolism in
S. cerevisiae redirects carbon flux towards the pentose phosphate pathway, thereby reducing the availability of substrates like erythrose-4-phosphate and phosphoenolpyruvate in the shikimate pathway, crucial for aromatic amino acid synthesis [
24]. This alteration in carbon flux potentially explains the trends observed in FA production before the 22-h mark of cultivation. Fed-batch cultivation offers distinct advantages over batch cultivation by enabling operation at lower residual substrate concentrations, thereby mitigating the inhibitory effects of substrate accumulation. Published research has demonstrated significant improvements in product yields using fed-batch fermentation strategies. For instance, engineered strains of
S. cerevisiae achieved a two-fold increase in resveratrol production [
15], while another study reported a 98% enhancement in α-santalene production (1.45 mg·L
−1 vs. 92.00 mg·L
−1) through fed-batch strategy [
16].
Regarding the distribution of ferulic acid (FA) and its related intermediates, such as
p-CA and caffeic acid,n during, batch and fed-batch fermentations, the acidic pH facilitated the migration of
p-HCAs predominantly into the organic phase, enabling effective detoxification of the broth and efficient recovery of p-HCAs. Increased solubility of
p-HCAs in the fatty alcohol phase can be attributed to their protonation when the pH is lower than their respective pK
1 values, which are 4.65, 4.62 and 4.56, for
p-CA, caffeic acid, and FA, respectively.
In this study, we demonstrated the feasibility of
de novo microbial production of FA from
d-glucose using an engineered strain of
S. cerevisiae (ABG20). Through optimization of the culture medium, continuous extraction of FA, and application of fed-batch processing, we achieved a production yield of 312.6 mg·L
−1 of FA, a production that has not yet been reported in the literature.
Acknowledgments
The authors gratefully acknowledge Arnaud Haudrechy for proofreading the article.
Author Contributions
B.G.: Formal analysis, Investigation, Data curation; J.C.: Formal analysis, investigation; F.B. (Fanny Brunissen) & F.B. (Fanny Brunois): HPLC data acquisition and analysis; C.F.: Investigation; F.A.: Conceptualization, Project administration, Funding acquisition, Review and Editing; M.L.: Supervision. N.I.: Conceptualization, Methodology, Supervision, Formal analysis, Investigation, Writing—original draft, Writing—review & editing.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data will be made available on request.
Funding
This work was supported by the Région Grand Est (France), Conseil Départemental de la Marne (France), and Grand Reims (France). The Agence Nationale de la Recherche (ANR, Grant ANR-17-CE07-0046) funded strain construction.
Declaration of Competing Interest
The authors declare that they have no competing financial interests or personal relationships that could have appeared to influence the work reported here.
References
-
1.
Kumar N, Pruthi V. Potential Applications of Ferulic Acid from Natural Sources.
Biotechnol. Rep. 2014,
4, 86–93. doi:10.1016/j.btre.2014.09.002.
[Google Scholar]
-
2.
Rodrigues JL, Araújo RG, Prather KLJ, Kluskens LD, Rodrigues LR. Production of Curcuminoids from Tyrosine by a Metabolically Engineered
Escherichia coli Using Caffeic Acid as an Intermediate.
Biotechnol. J. 2015,
10, 599–609. doi:10.1002/biot.201400637.
[Google Scholar]
-
3.
Braga A, Faria N. Bioprocess Optimization for the Production of Aromatic Compounds with Metabolically Engineered Hosts: Recent Developments and Future Challenges.
Front. Bioeng. Biotechnol. 2020,
8, 96. doi:10.3389/fbioe.2020.00096.
[Google Scholar]
-
4.
Borges A, Ferreira C, Saavedra MJ, Simões M. Antibacterial Activity and Mode of Action of Ferulic and Gallic Acids against Pathogenic Bacteria.
Microb. Drug Resist. 2013,
19, 256–265. doi:10.1089/mdr.2012.0244.
[Google Scholar]
-
5.
Pei K, Ou J, Huang J, Ou S. P-Coumaric Acid and Its Conjugates: Dietary Sources, Pharmacokinetic Properties and Biological Activities.
J. Sci. Food Agric. 2016,
96, 2952–2962. doi:10.1002/jsfa.7578.
[Google Scholar]
-
6.
Flourat AL, Combes J, Bailly-Maitre-Grand C, Magnien K, Haudrechy A, Renault JH, et al. Accessing P-Hydroxycinnamic Acids: Chemical Synthesis, Biomass Recovery, or Engineered Microbial Production?
ChemSusChem 2021,
14, 118–129. doi:10.1002/cssc.202002141.
[Google Scholar]
-
7.
Serra S, Fuganti C, Brenna E. Biocatalytic Preparation of Natural Flavours and Fragrances.
Trends Biotechnol. 2005,
23, 193–198. doi:10.1016/j.tibtech.2005.02.003.
[Google Scholar]
-
8.
Choi O, Wu CZ, Kang SY, Ahn JS, Uhm TB, Hong YS. Biosynthesis of Plant-SpeciWc Phenylpropanoids by Construction of an ArtiWcial Biosynthetic Pathway in
Escherichia coli.
J. Ind. Microbiol. Biotechnol. 2011,
38, 1657–1665. doi:10.1007/s10295-011-0954-3.
[Google Scholar]
-
9.
Kang SY, Choi O, Lee JK, Hwang BY, Uhm TB, Hong YS. Artificial Biosynthesis of Phenylpropanoic Acids in a Tyrosine Overproducing
Escherichia coli Strain.
Microb. Cell Fact. 2012,
11, 1–9. doi:10.1186/1475-2859-11-153.
[Google Scholar]
-
10.
Lv H, Zhang Y, Shao J, Liu H, Wang Y. Ferulic Acid Production by Metabolically Engineered
Escherichia coli.
Bioresour. Bioprocess. 2021,
8, 70. doi:10.1186/s40643-021-00423-0.
[Google Scholar]
-
11.
Combes J, Imatoukene N, Couvreur J, Godon B, Brunissen F, Fojcik C, et al. Intensification of P-Coumaric Acid Heterologous Production Using Extractive Biphasic Fermentation.
Bioresour. Technol. 2021,
337, 125436. doi:10.1016/j.biortech.2021.125436.
[Google Scholar]
-
12.
Singh V, Haque S, Niwas R, Srivastava A, Pasupuleti M, Tripathi CKM. Strategies for Fermentation Medium Optimization: An in-Depth Review.
Front. Microbiol. 2017,
7, 2087. doi:10.3389/fmicb.2016.02087.
[Google Scholar]
-
13.
Villadsen J, Nielsen J, Lidén G. Principles, Bioreaction Engineering; Springer Science & Business Media: Lyngby, Denmark: 2011; ISBN 9781441996879.
-
14.
Postma E, Verduyn C, Scheffers WA, Van Dijken JP. Enzymic Analysis of the Crabtree Effect in Glucose-Limited Chemostat Cultures of Saccharomyces Cerevisiae.
Appl. Environ. Microbiol. 1989,
55, 468–477. doi:10.1128/aem.55.2.468-477.1989.
[Google Scholar]
-
15.
Li M, Kildegaard KR, Chen Y, Rodriguez A, Borodina I, Nielsen J.
De Novo Production of Resveratrol from Glucose or Ethanol by Engineered Saccharomyces Cerevisiae.
Metab. Eng. 2015,
32, 1–11. doi:10.1016/j.ymben.2015.08.007.
[Google Scholar]
-
16.
Scalcinati G, Knuf C, Partow S, Chen Y, Maury J, Schalk M, et al. Dynamic Control of Gene Expression in Saccharomyces Cerevisiae Engineered for the Production of Plant Sesquitepene α-Santalene in a Fed-Batch Mode.
Metab. Eng. 2012,
14, 91–103. doi:10.1016/j.ymben.2012.01.007.
[Google Scholar]
-
17.
Combes J, Imatoukene N, Couvreur J, Godon B, Fojcik C, Allais F, et al. An Optimized Semi-Defined Medium for p-Coumaric Acid Production in Extractive Fermentation.
Process Biochem. 2022,
122, 357–362. doi:10.1016/j.procbio.2022.10.021.
[Google Scholar]
-
18.
Dahod SK, Greasham R, Kennedy M. Raw Materials Selection and Medium Development for Industrial Fermentation Processes.
Man. Ind. Microbiol. Biotechnol. 2014, 659–668. doi:10.1128/9781555816827.ch46.
[Google Scholar]
-
19.
Zhang J, Greasham R. Chemically Defined Media for Commercial Fermentations.
Appl. Microbiol. Biotechnol. 1999,
51, 407–421. doi:10.1007/s002530051411.
[Google Scholar]
-
20.
Morais V, Suárez N. Economic Evaluation of Streptococcus Pneumoniae Culture Media.
Am. J. Biochem. Biotechnol. 2016,
12, 133–138. doi:10.3844/ajbbsp.2016.133.138.
[Google Scholar]
-
21.
Hahn-Hägerdal B, Karhumaa K, Larsson CU, Gorwa-Grauslund M, Görgens J, van Zyl WH. Role of Cultivation Media in the Development of Yeast Strains for Large Scale Industrial Use.
Microb. Cell Fact. 2005,
4, 1–16. doi:10.1186/1475-2859-4-31.
[Google Scholar]
-
22.
Hernández-Chávez G, Martinez A, Gosset G. Metabolic Engineering Strategies for Caffeic Acid Production in
Escherichia coli.
Electron. J. Biotechnol. 2019,
38, 19–26. doi:10.1016/j.ejbt.2018.12.004.
[Google Scholar]
-
23.
Lin Y, Yan Y. Biosynthesis of Caffeic Acid in
Escherichia coli Using Its Endogenous Hydroxylase Complex.
Microb. Cell Fact. 2014,
11, 3–11. doi:10.1186/1475-2859-11-42.
[Google Scholar]
-
24.
Frick O, Wittmann C. Characterization of the Metabolic Shift between Oxidative and Fermentative Growth in Saccharomyces Cerevisiae by Comparative 13C Flux Analysis.
Microb. Cell Fact. 2005,
4. doi:10.1186/1475-2859-4-30.
[Google Scholar]