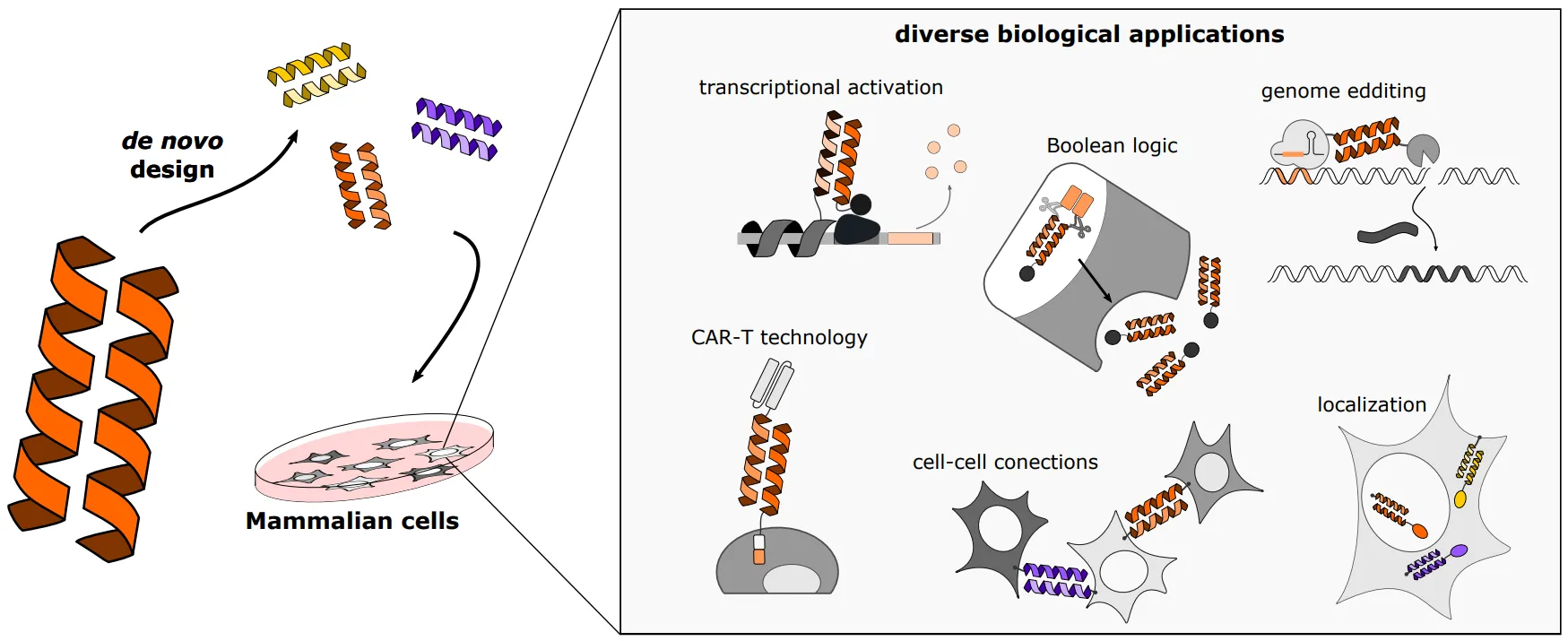
1. Introduction
Cells are complex systems with numerous processes running in parallel, regulated at different levels and by different mechanisms. Mammalian cells, are of interest to us because they govern our life and diseases. They are capable of maintaining homeostasis and responding to diverse signals, however in some cases these natural systems are not sufficient. Thus, scientists strive to engineer the properties of mammalian cells, allowing them to be better understood and treat diverse pathological conditions using advanced gene and cell therapy. In these cases, there is a need to introduce external control or new types of regulation of selected cellular processes, which are compatible and, preferably, orthogonal to the existing processes. This can be achieved by introducing several types of protein–protein interactions. Designed interactions can be governed by natural protein–protein (e.g., protein-anti/nano-body) or protein–peptide interaction domains like WW, SH3, PH, PDZ [
1,
2,
3,
4,
5,
6] and synthetically designed interaction domains such as coiled coil domains. *De novo* designed coiled coils have several advantages in comparison to naturally occurring dimerization domains. Due to well understood amino acid sequence to structure relationship in coiled coils, we are able to fine tune their affinities, stability and most importantly their orthogonality. They can be used to direct selected biological molecules to specific subcellular locations, activate or inactivate selected biological functions, and regulate gene transcription based on internal or external biological, chemical or physical signals or their combinations and many more. Many excellent examples of efficient engineering of mammalian cells have been demonstrated, using diverse ectopic or designed modules [
7,
8]. The availability of well-understood modular tools of synthetic biology facilitates their incorporation into diverse biological systems. Designed coiled coils, which are the topic of this review, thus represent a highly versatile tool for engineered mammalian (and other) cells.
2. Coiled Coils
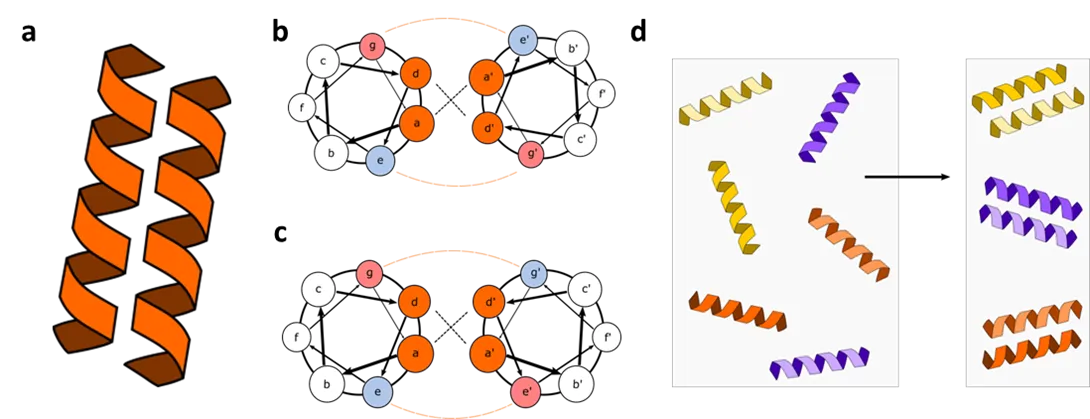
First described by Crick in 1953 [
9], coiled coils (CC) (a) are widespread structural motifs in natural proteins. Approximately 2–3% of all protein residues are in coiled coil segments and as such, they are involved in diverse organizational and regulatory processes. They perform both structural and functional roles and act in protein–protein interaction as well as in DNA-binding domains [
10]. The most common type of coiled coils is left-handed, where each helix has seven amino acid residues in a repeat (also termed heptad repeat) [
11]. In these motifs, hydrophobic (H) residues are usually alternately spaced three and four residues apart and separated by polar (P) residues, creating the pattern (HPPHPPP) [
12]. This repeat is usually denoted *a-b-c-d-e-f-g* in one helix and *a'-b'-c'-d'-e'-f'-g'* in the other with anywhere from two (in designed coiled coils) [
13] to 200 [
14] of heptad repeats in proteins. Such motifs fold in amphipathic helices, which assemble into bundles where their hydrophobic faces are buried from solvent. Helices within the coiled coil dimer can be positioned in either parallel (b) or antiparallel (c) orientation. Amino acids at positions *a* and *d* are typically non-polar residues, which form the core of a coiled coil stabilized by hydrophobic and van der Waals interactions. Positions *e* and *g* are solvent exposed, polar or charged residues which contribute to the specificity of pairing predominantly through electrostatic interactions [
15]. The remaining *b*, *c* and *f* positions are solvent-exposed and usually present amino acids contributing to the helical propensity [
16]. Amino acid sequence variations within a coiled coil pair enable the generation of a large number of designs [
16,
17]. Due to the well-understood rules governing their folding and specificity [
15,
16] they are suitable for building modular protein scaffolds [
18], enzymes [
19], and transcription factors [
20].
. Structure and orthogonality of coiled coils. (a) Schematic representation of coiled coil dimers. (b,c) Interactions between CC-forming peptides defining specificity and orientation in parallel (b) and antiparallel (c) coiled coils. (d) Schematic representation of an orthogonal set of coiled coils. Peptides interact only with the designed interaction partner, forming a coiled coil.
Based on the relatively straightforward structure-activity relationship of CC dimers, and the negative design [
16,
18,
21,
22], *de novo* heterodimeric coiled coils have been designed, using the combinatorial arrangement of interacting heptads, which can be achieved by introduction of an Asn residue at the *a* position of parallel CC dimers (b,c). This destabilizes CC interaction due to the Asn–Asn interaction in the desired pair and it also destabilizes the non-desired Asn–Leu interaction, thereby increasing the orthogonality (d) among the peptide set members. In addition to the electrostatic interactions between positions *e* and *g*, stability could be tuned by the α-helical propensity at non-interacting *b*, *c* and *f* residues, which enables tuning of the affinity within several orders of magnitude, while maintaining the specificity. Several CC pairs have been designed and analyzed experimentally as well as bioinformatically. They showed high orthogonality and were used for diverse purposes. A set of validated CC pairs developed and tested in our group is shown here, including parallel (b) as well as antiparallel (c) heterodimers, with a range of affinities (). Several other sets are available, developed notably by Keating [
23], Mason [
5], and Woolfson [
12] groups.
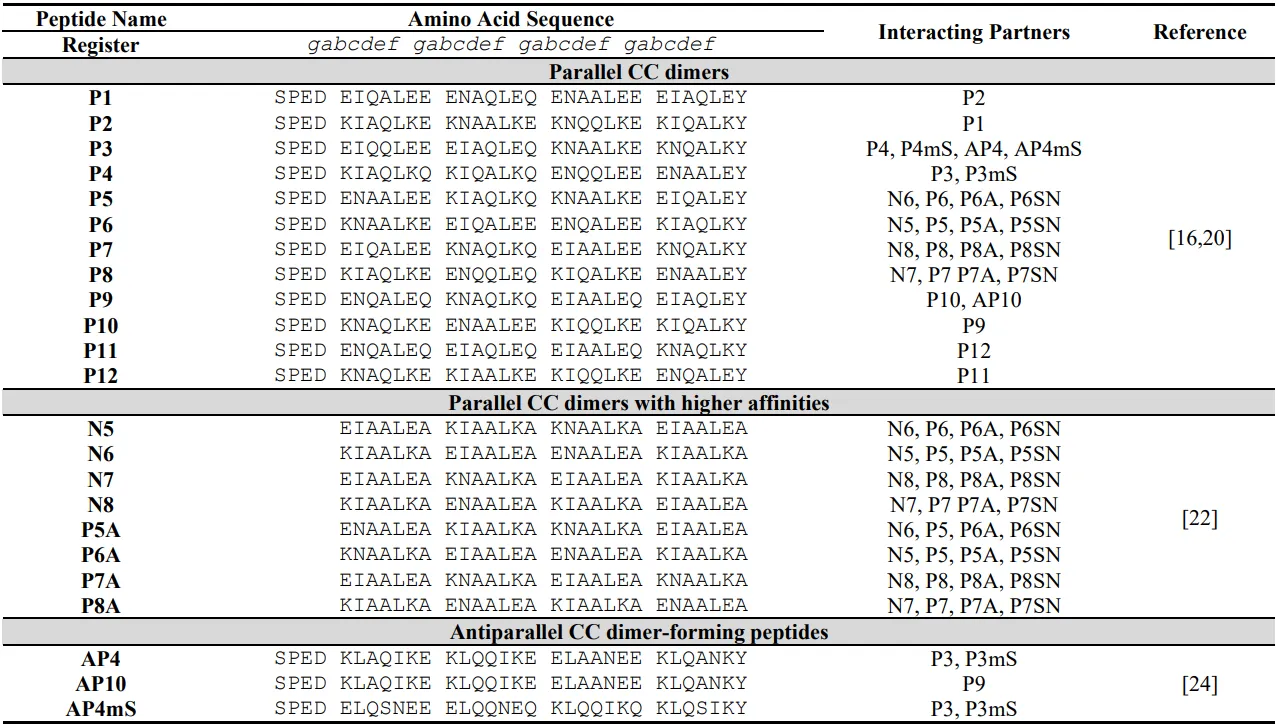
. Sequences of a set of orthogonal designed coiled coil dimers validated in mammalian cells.
3. Designed Regulation of Mammalian Cells
The key features of coiled coils include the orthogonality, their ability to bind to their designed partner, and the minimal cross interactions [
16], which makes them ideal as units for dimerization in the complex environment of mammalian cells. Manipulation of the local helical propensity of interacting peptides can be used to tune the stabilities of coiled coil dimers over a wide range.
Adjusting CC dimer stability, while maintaining the specificity, can be achieved by altering intramolecular charge-charge interactions and helical propensity [
21], thus enabling fine-tuning of CC stability. Importantly, the range of stabilities of CC dimers can be tuned across several orders of magnitude, from micromolar to a nanomolar range. This is achieved by modifying the length of pairs (number of heptad repeats) or concatenation of CC modules and by modifying residues at interacting as well as at noninteracting sites [
20,
21,
22,
25,
26]. In addition to a small genetic footprint of CC modules, we can also specify the orientation of CC dimers (parallel vs. antiparallel), which allows precise positioning within the context of protein assemblies and is particularly important for the design of coiled coil protein origami cages [
18]. These features were demonstrated by Plaper et al. [
22] where a split luciferase reporter was used to determine the stability of CC pairs in mammalian cells (a). *In vitro* determined KD values of designed coiled coil dimers were in the nanomolar range (1–20 nM) for the strongest binding pairs (). Based on the NICP set of coiled coils, an expanded set of orthogonal coiled coils was designed () and experimentally evaluated via split luciferase reporter assembly (a) as well as TALE and VP16 mediated association for transcription regulation (c) assessing the possibility to use them in mammalian cells. By pairing coiled coils with complementary electrostatic motifs but different helical propensities at non-interacting *b*, *c*, and *f* positions, CC pairs can be disrupted by the addition of a peptide with higher affinity and thus displaced by an interacting partner (d).
An important part of the designed regulation of mammalian cells is the introduction of synthetic transcriptional regulators that induce or inhibit gene expression. Several transcriptional regulation systems are based on native coiled coil forming domains such as Fos/Jun [
23]. One key advantage of *de novo* designed proteins is the reduced risk of undesired interferences with the cell’s natural processes, which has been investigated both by bioinformatics as well as by experimental testing in mammalian cells [
20]. Edgell et al. [
27] designed coiled coils in order to mediate protein–protein interactions in *Escherichia coli*. A newly designed parallel homodimer was successful in the transcription repression assay based on the reconstitution of the Lac repressor. By using a set of orthogonal coiled coils we can improve the response of mammalian cells to external signals. Lebar et al. [
20] tested the orthogonality of a set of six orthogonal coiled coil pairs, termed NICP (d) in mammalian cells using synthetic transcriptional regulators, comprising peptide fused to a DNA binding TALE [
28,
29] protein and a second peptide fused to the VP16 activation domain [
30,
31] (c). The NICP set generated a toolbox for a tunable level of protein–protein interactions. The interaction affinity can be further strengthened by concatenation of several CC peptides. Coiled coil orthogonality in mammalian cells further provides the possibility of simultaneous regulation of multiple protein–protein interactions or localization multiplexing (b).
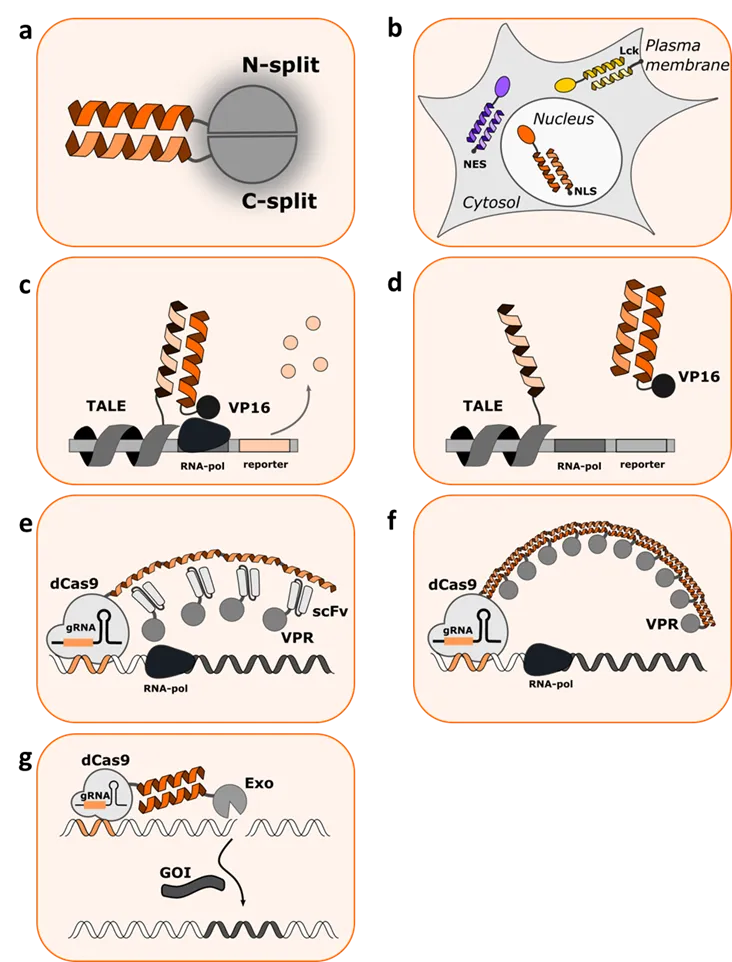
. Coiled coil mediated regulation of mammalian cells. (a) Coiled coils mediated reconstitution of the reporter protein, such as split firefly luciferase or SEA. Interaction between CC-forming peptides enables sufficient vicinity for split protein domains to reconstitute and regain catalytic activity. (b) A toolbox of orthogonal CC-forming peptides can serve as specific cellular anchors for distinct subcellular localization of diverse target proteins. NES—nuclear export signal; NLS—nuclear localization signal; Lck—signal peptide for plasma membrane anchoring. (c) Depiction of CC-peptide fused to a DNA binding TALE protein and a second peptide fused to VP16 activation domain. Interaction between the two peptides brings the DNA binding domain (TALE) and activation domain (VP16) into proximity, allowing transcription of the reporter protein. (d) Addition of a displacer peptide perturbs the proximity of TALE and VP16 resulting in interruption of transcription. (e) Repeats of parallel homodimeric GCN4 peptide array (SunTag) fused to dCas9 protein. A single-chain variable fragment (scFv) antibody against GCN4 fused to VPR fusion, enabled recruitment of VPR to dCas-GCN4 peptide and thus transcriptional activation. (f) Repeats of peptide P3 (CCC-tag) fused to dCas9. Peptide P4 fused to VPR domains enabled recruitment of VPR to dCas-P3 and thus transcriptional activation. (g) Recruitment of exonuclease (Exo) to Cas9/gRNA via coiled coil interaction (CCExo) for genome editing. GOI–gene of interest.
A highly versatile system for the transcriptional regulation of mammalian cells is based on the adaptive bacterial immunity CRISPR system [
32,
33] using a nuclease-deficient mutant of the CRISPR effector protein Cas9 (dCas9) as a DNA binding domain [
34]. Together with a sequence-specific small guide RNAs (sgRNAs), dCas9 can be directed to any sequence in the mammalian genome. The direct fusion of dCas to herpes virus-derived transcription activation domains (VP16, VP64, VPR) or transcription inhibition domain KRAB produced potent transcriptional regulators [
35]. Several derivatives from CRISPR–dCas9 system have been developed since 2016 [
36]. Transcription of a gene can be further enhanced by recruiting multiple copies of transcriptional activators to promoter regions. One possibility for such regulation is a sequential fusion of several coiled coil forming peptides on the DNA-binding protein. Tanenbaum et al. [
37] developed a protein scaffold, by repeating a parallel homodimeric GCN4 peptide array, termed SunTag (e). A single-chain variable fragment (scFv) antibody against GCN4 was used to target VP64 fusion protein to GCN4 peptide in Suntag. Fusion of Suntag to dCas9 allowed the recruitment of several VP64 proteins to the promotor region, thus enhancing the level of dCas9 transcriptional activation of endogenous genes. Similar concatenation of coiled coil peptides, was developed by Lebar et al. through several fused repeats of peptide P3 [
20], where transcriptional regulation domains were recruited via CC peptide, rather than scFv domains (f). Here, dCas9 protein was fused to up to ten repeats of peptide P3 (CCC-tag) from the orthogonal toolbox of *de novo* designed coiled coils. Peptide P4 was genetically fused to VP46 or VPR domains, facilitating the recruitment of transcriptional activation domains to the promotor region. The use of orthogonal heterodimeric peptides P3 and P4 has a clear advantage over homodimeric GCN4 peptide. While GCN4 peptide allows the formation of homodimeric interactions, and therefore formation of ill-defined stoichiometry, due to the possibility of either intra- or intermolecular interactions, the heterodimeric P3/P4 coiled coil forms only intermolecular interactions, which leads to highly potent CRISPR–dCas9-based transcriptional activators. CCC-tag-based CRISPR–dCas9 system demonstrated up to several thousand-fold increased levels of reporter gene transcription including efficient upregulation of genomic targets.
The small size of coiled coils enables their use as protein tags, while not interfering with the structure of the protein of interest. SunTag by Tanenbaum et al. [
37] allowed the efficient recruitment of multiple proteins to a single genomic locus, labeling telomeres. Alternatively, an orthogonal set of coiled coil pairs can simultaneously direct proteins to different subcellular localizations, as shown in Lebar et al. [
20]. In this case, each of the orthogonal peptides was directed to plasma membrane, nucleus, or cytosol by fusion of an appropriate localization signal. The desired protein cargos (fluorescent proteins) were recruited to the appropriate location by specific CC interactions via the complementary coiled coil peptide fused to the cargo protein. The orthogonality of used peptide pairs enabled fluorescent proteins to be discretely localized, specifically in the cell area with the appropriate anchor (b).
Further expanding the versatility of coiled coil peptides, Lainšček et al. [
38] designed an enhanced gene editing method by recruiting exonuclease to Cas9/gRNA via coiled coil interaction, termed CCExo (g). This method robustly increased gene knock-out due to progressive DNA strand recession at the cleavage site, decreasing the re-ligation of the non-edited DNA. Implementation of CCExo increased deletion size and enhanced gene inactivation efficiency in the context of several different DNA targets, gRNA selection, Cas variants, tested cell lines, and types of delivery. Coiled coil dimer forming modules are also a useful tool as a screening method for the detection of effective split proteins (a). Golob-Urbanc et al. [
39] demonstrated the detection of functional split superantigen using coiled coil modules, where the split superantigen staphylococcal enterotoxin A (SEA) regained its activity only by fusion to heterodimeric coiled coil forming peptides (a).
4. Coiled Coil-Mediated Signaling Circuits
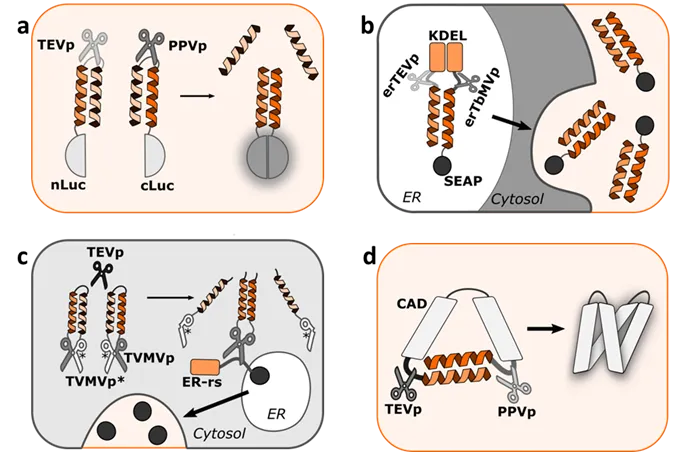
Coiled coil based regulation in mammalian cells was also explored by Fink et al. [
24] where coiled coils were used to introduce a fast response of mammalian cells based on posttranslational proteolytic modifications. With strategically positioned protease cleavage sites and peptides from the orthogonal set of coiled coils with a tunable affinity for competitive displacement, they designed modular and scalable signaling pathways. This platform allowed a generation of split-protease-cleavable orthogonal-CC-based (SPOC) logic circuits demonstrating Boolean logic in mammalian cells (a). Praznik et al. [
40] showcased an endoplasmic reticulum retention signal-based lumER system (b). They adapted SPOC designed constructs to ER lumen (lumER) and designed a system that allowed retention of proteins of interest (POI) in the ER. Coiled coils in this study enabled reconstitution of split protease upon activation by other proteases as well as retention of POI inside ER lumen to facilitate faster secretion or membrane localization of proteins, generated from the pre-synthesized pool of proteins. This enabled modular construction of A nimply B as well as AND Boolean logic gate. Similarly, Vlahos et al. [
41] created a modular, generalizable design called Retained Endoplasmic Cleavable Secretion (RELEASE) (c). They engineered proteins that were retained in the endoplasmic reticulum and displayed or secreted in response to specific proteases. Reconstitution of their designed split proteases was based on coiled coil interaction.
Using coiled coils for domain inhibition and proteolytic regulation, Jazbec et al. [
42] reported an innovative genetically engineered protease-activated Orai calcium channels activator called PACE—a genetically engineered mediator between proteolytic activity and calcium influx (d). In this study, a part of STIM protein (which together with Orai1 channels modulates calcium entry) termed CAD was engineered. Self-dimerization and activation of CAD was inhibited with an intramolecular coiled coil forming peptide pair. Strategically positioned protease cleavage sites in CAD constructs enabled its processing and subsequent activation of the Orai1 channel. This created a highly regulated intracellular calcium entry and downstream signal processing.
. Coiled coil-regulated signaling circuits. (a) Protease-cleavable orthogonal-CC-based (SPOC) logic circuits demonstrating Boolean logic in mammalian cells. (b) Endoplasmic reticulum (ER) retention signal-based lumER system which allows retention of protein of interest (POI) in the ER. Coiled coils enable reconstitution of split protease which facilitates fast secretion of POI. (c) Modular, generalizable design termed Retained Endoplasmic Cleavable Secretion (RELEASE). Coiled coils enable the reconstitution of split protease which enables the release of retained protein. (d) Genetically engineered protease-activated Orai calcium channels activator PACE, a genetically engineered mediator between proteolytic activity and calcium influx. Proteolytic cleavage enables the release of CC-peptides and activation of PACE. TEVp—tobacco etch virus protease; PPVp—plum pox virus protease; nLuc—N-terminal firefly luciferase domain; cLuc—C-terminal firefly luciferase domain; KDEL—endoplasmic reticulum retention signal; TVMVp—Tobacco vein mottling virus protease; ER-rs—endoplasmic reticulum retention signal; ER—endoplasmic reticulum; CAD—CRAC activating domain.
5. Coiled Coil Facilitated Cell–Cell Interactions
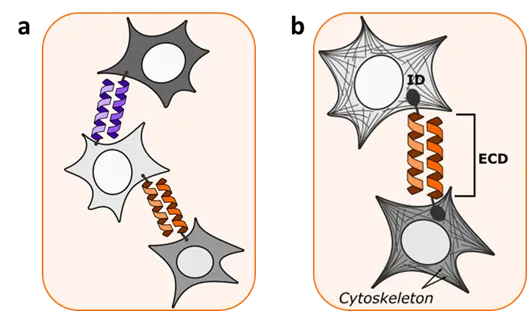
In nature, cell–cell interactions or interactions between cells and extracellular matrix occur through cell adhesion molecules (CAM), which plays a central role in normal tissue structure and function [
43]. Chao et al. [
44] showed that by engineering synthetic CAMs based on coiled coils, called “helixCAM”, specific programmable interactions between cells and patterned aggregation could be achieved in both bacteria and human cells (a). By expressing orthogonal coiled coil forming polypeptides fused to a transmembrane domain on a mammalian cell surface, they constructed three-dimensional cell structures and managed to specifically target suspension cells to adherent cells. Stevens et al. [
45] systematically explored the modularity of CAMs. They prepared several synthetic CAMs (synCAMs) by fusing orthogonal extracellular binding domains (ECD) to endogenous CAM intracellular domains from native adhesion molecules (b). Contrary to helixCAM, the synthetic CAMs contained a preserved intracellular domain. In one of the designs they prepared a homotypic synCAM where self-dimerizing Aph (computationally designed) or IF1 (bovine ATPase inhibitor IF1) coiled coils were used as ECD. Those were fused to the extracellular domain of fibronectin, providing additional separation from the cell membrane, and then linked to transmembrane and intracellular domain from ICAM-1. Mixed cells expressing orthogonal synCAMs formed self-organizing structures in a modular manner which can be predicted according to expressed synCAM.
. Cell–cell interactions mediated by coiled coils. (a) Programmable cell assembly with “helixCAM”. Coiled coils are fused to a transmembrane domain and expressed on a cell surface thus enabling cell–cell connections. (b) “synCAMs” are composed of extracellular binding domain (ECD) based on coiled coil fused with endogenous CAM intracellular domain (ID) originated from native adhesion molecules.
6. CAR Receptor Designs with Coiled Coils
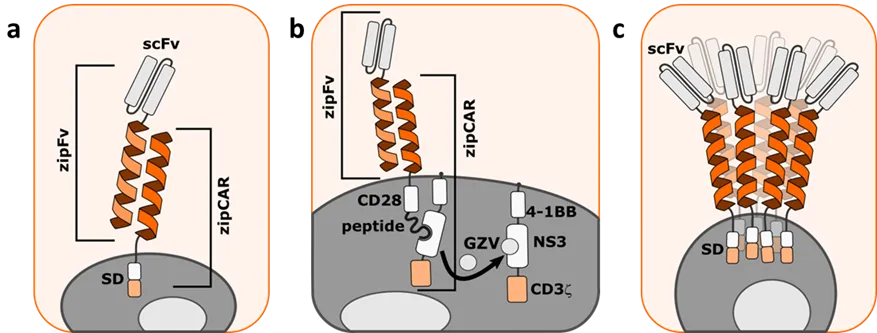
Chimeric antigen receptor (CAR) T cells are an emerging new cancer immunotherapy. Clinical trials with CD19 CAR T cells have resulted in remarkably effective and durable clinical responses in severe forms of advanced B-cell malignancies [
46]. There are still some limitations of the efficacy, safety, and widespread use of current CAR-T cell therapies. New generations of CART cells are emerging to circumvent these issues and reduce the adverse effects. By splitting the CAR receptor at the extracellular region and incorporating a complementary coiled coil forming peptide into both components, Cho et al. [
47] developed a split, universal, and programmable (SUPRA) CAR receptor (a). It consists of an adapter molecule, which is scFv fused with a coiled coil forming peptide (also called leucine zipper) (zipFv), and a universal receptor composed of intracellular signaling domains, a transmembrane domain, and an extracellular cognate coiled coil forming peptide (zipCAR) that binds specifically to the zipFv coiled coil forming peptide. In this design scFv is prepared externally and can be dissociated from the signaling motif of the CAR, allowing a panel of different antigens to be targeted without re-engineering immune cells for each new target. In addition, adjusting zipFv concentration and affinity to target antigen, coiled coil affinity for each other, and zipCAR expression levels enable modulating the output level of SUPRA CAR cells. Authors have further exploited the SUPRA CAR and demonstrated broad applicability on adaptive and innate immune cells [
48].
. CAR T cell therapies regulated by coiled coils. (a) SUPRA CAR system consists of universal zipFv (scFv targeting tumor antigen fused to a leucine zipper) and zipCAR (cognate extracellular leucine zipper fused with intracellular signaling domains (SD)). (b) ON-OFF VIPER CAR system consists of zipFv (scFv targeting tumor antigen fused to a leucine zipper) and zipCAR divided into two parts enabling regulation of signaling by small molecule grazoprevir (GZV). (c) Schematic representation of oligomeric CAR receptor with incorporated pentameric coiled coils from COMP. Coiled coils enable connections between scFv and signaling domains (SD).
Li et al. [
49] developed a collection of versatile protease regulated (VIPER) CAR receptors using viral non-structural protein 3 (NS3) with proteolytic activity, and an exogenous control with clinically approved antiviral drugs (b). To create a universal CAR receptor that could be switched ON and OFF on the demand, they applied the OFF VIPER CAR design to a SUPRA CAR technology. The three-component system is composed of zipFv, and similar to SUPRA CAR, meanwhile, intracellular zipCAR is divided into two parts: the first, consists of an extracellular coiled coil, transmembrane domain, CD28 costimulatory domain, and an NS3-binding peptide; the second is also a membrane-tethered component containing a DAP10 ectodomain, costimulatory 4-1BB domain, catalytically inactive NS3, and CD3ζ signaling domain. Only when all three components are present and the tumor antigen is recognized by zipFv, the CAR receptor is activated. With the addition of the small molecule grazoprevir, that disturbs the binding of NS3-binding peptide and NS3, the intracellular parts of zipCAR are dissociated and the signaling is inhibited. SUPRA CAR and VIPER CAR demonstrate how coiled coils can be a powerful tool for designing universal and tunable CAR receptors.
In addition to dimerization, coiled coil motif represents one of the most common structural motifs in proteins and is responsible for protein oligomerization in a highly specific manner. Therefore coiled coil motifs can also be used to enhance activation through higher-order oligomers. Cordoba et al. [
50] incorporated pentameric coiled coil from cartilage oligomeric matrix protein (COMP) between scFv-CD22 and transmembrane domain of CAR receptor and showed an increased sensitivity to target tumor antigen CD22 in comparison to a hinge domain from human IgG1 (c).
7. Discussion and Conclusions
Coiled coil dimers are small, versatile, and tunable building modules that can be used to regulate the assembly or disassembly of proteins of interest. This in turn leads to the heart of many biological processes. Numerous approaches have been conceived, mainly for combinations with genetically fused protein domains, to perform diverse functions and implement different logical functions both inside and outside the cells. Although this review focuses on applications in mammalian cells, similar strategies could equally be used in any type of biological or biomimetic systems, for example, to construct designed protein nanostructures [
18,
51]. CCs have a small genetic footprint (about 100 base pairs per interaction module in most useful set comprising 4 heptads) which are subtle and typically expressed well, soluble and have favorable folding properties. The largest set of orthogonal CCs measures around 10–20 pairs [
20,
52]. This is substantially less than nucleic acid-based complementarity but is sufficient for most applications, and may potentially also be used for cellular barcoding. Although coiled coil segments span from a few to tens of heptads, currently the most commonly used one is comprised of 4 heptads. Shorter coiled coils have substantially weaker affinity and enable smaller orthogonal sets, while on the other hand, it is still a challenge to design longer CC pairs with sufficient selectivity. Coiled coils can be designed to pair in a parallel or antiparallel orientation, which may be useful or even essential for some fused protein domains, autoinhibition or for the proper reconstitution of some split enzymes.
In addition to an already impressive collection of applications, we can expect that coiled coils could be used for many more implementations in mammalian cell regulation, such as the formation of protein condensates or the regulation of protein functions.
Author Contributions
Writing—Original Draft Preparation, all authors; Writing—Review & Editing, all authors.; Funding Acquisition, R.J.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This research was funded by the Slovenian Research Agency grants number P4-0176, J1-2481, and J1-1711 and European Research Council (ERC AdG MaCChines 787115).
Declaration of Competing Interest
The authors are co-inventors of patent applications on the regulation of mammalian cells based on coiled coils.
References
-
1.
Wenz MT, Bertazzon M, Sticht J, Aleksić S, Gjorgjevikj D, Freund C, et al. Target Recognition in Tandem WW Domains: Complex Structures for Parallel and Antiparallel Ligand Orientation in h-FBP21 Tandem WW.
J. Chem. Inf. Model. 2022,
62, 6586–6601.
[Google Scholar]
-
2.
Kurochkina N, Guha U. SH3 domains: Modules of protein–protein interactions.
Biophys. Rev. 2013,
5, 29–39.
[Google Scholar]
-
3.
Scheffzek K, Welti S. Pleckstrin homology (PH) like domains—Versatile modules in protein-protein interaction platforms.
FEBS Lett. 2012,
586, 2662–2673.
[Google Scholar]
-
4.
Haslam NJ, Shields DC. Peptide-Binding Domains: Are Limp Handshakes Safest?
Sci. Signal. 2012,
5, pe40.
[Google Scholar]
-
5.
Crooks RO, Lathbridge A, Panek AS, Mason JM. Computational Prediction and Design for Creating Iteratively Larger Heterospecific Coiled Coil Sets.
Biochemistry 2017,
56, 1573–1584.
[Google Scholar]
-
6.
Lee H-J, Zheng JJ. PDZ domains and their binding partners: structure, specificity, and modification.
Cell Commun. Signal 2010,
8, 8.
[Google Scholar]
-
7.
Ausländer S, Fussenegger M. Engineering Gene Circuits for Mammalian Cell–Based Applications.
Cold Spring Harb. Perspect. Biol. 2016,
8, a023895.
[Google Scholar]
-
8.
Kojima R, Fussenegger M. Synthetic Biology: Engineering Mammalian Cells To Control Cell‐to‐Cell Communication at Will.
ChemBioChem 2019,
20, 994–1002.
[Google Scholar]
-
9.
Crick FHC. The packing of α-helices: Simple coiled-coils.
Acta Crystallogr. 1953,
6, 689–697.
[Google Scholar]
-
10.
Lupas AN, Gruber M. The Structure of α-Helical Coiled Coils. In Advances in Protein Chemistry; Elsevier: Oxford, UK, 2005; Volume 70, pp. 37–38.
-
11.
Mason JM, Arndt KM. Coiled Coil Domains: Stability, Specificity, and Biological Implications.
ChemBioChem 2004,
5, 170–176.
[Google Scholar]
-
12.
Moutevelis E, Woolfson DN. A Periodic Table of Coiled-Coil Protein Structures.
J. Mol. Biol. 2009,
385, 726–732.
[Google Scholar]
-
13.
Burkhard P, Meier M, Lustig A. Design of a minimal protein oligomerization domain by a structural approach.
Protein Sci. 2000,
9, 2294–2301.
[Google Scholar]
-
14.
Kohn WD, Mant CT, Hodges RS. α-Helical Protein Assembly Motifs.
J. Biol. Chem. 1997,
272, 2583–2586.
[Google Scholar]
-
15.
Negron C, Keating AE. A Set of Computationally Designed Orthogonal Antiparallel Homodimers that Expands the Synthetic Coiled-Coil Toolkit.
J. Am. Chem. Soc. 2014,
136, 16544–16556.
[Google Scholar]
-
16.
Gradišar H, Jerala R. De novo design of orthogonal peptide pairs forming parallel coiled-coil heterodimers.
J. Pept. Sci. 2011,
17, 100–106.
[Google Scholar]
-
17.
Taylor WR, Chelliah V, Hollup SM, MacDonald JT, Jonassen I. Probing the “Dark Matter” of Protein Fold Space.
Structure 2009,
17, 1244–1252.
[Google Scholar]
-
18.
Ljubetič A, Lapenta F, Gradišar H, Drobnak I, Aupič J, Strmšek Ž, et al. Design of coiled-coil protein-origami cages that self-assemble in vitro and in vivo.
Nat. Biotechnol. 2017,
35, 1094–1101.
[Google Scholar]
-
19.
Shekhawat SS, Porter JR, Sriprasad A, Ghosh I. An Autoinhibited Coiled-Coil Design Strategy for Split-Protein Protease Sensors.
J. Am. Chem. Soc. 2009,
131, 15284–15290.
[Google Scholar]
-
20.
Lebar T, Lainšček D, Merljak E, Aupič J, Jerala R. A tunable orthogonal coiled-coil interaction toolbox for engineering mammalian cells.
Nat. Chem. Biol. 2020,
16, 513–519.
[Google Scholar]
-
21.
Drobnak I, Gradišar H, Ljubetič A, Merljak E, Jerala R. Modulation of Coiled-Coil Dimer Stability through Surface Residues while Preserving Pairing Specificity.
J. Am. Chem. Soc. 2017,
139, 8229–8236.
[Google Scholar]
-
22.
Plaper T, Aupič J, Dekleva P, Lapenta F, Keber MM, Jerala R, et al. Coiled-coil heterodimers with increased stability for cellular regulation and sensing SARS-CoV-2 spike protein-mediated cell fusion.
Sci. Rep. 2021,
11, 9136.
[Google Scholar]
-
23.
Newman JRS, Keating AE. Comprehensive Identification of Human bZIP Interactions with Coiled-Coil Arrays.
Science 2003,
300, 2097–2101.
[Google Scholar]
-
24.
Fink T, Lonzarić J, Praznik A, Plaper T, Merljak E, Leben K, et al. Design of fast proteolysis-based signaling and logic circuits in mammalian cells.
Nat. Chem. Biol. 2019,
15, 115–122.
[Google Scholar]
-
25.
Smith AJ, Thomas F, Shoemark D, Woolfson DN, Savery NJ. Guiding Biomolecular Interactions in Cells Using de Novo Protein–Protein Interfaces.
ACS Synth. Biol. 2019,
8, 1284–1293.
[Google Scholar]
-
26.
Thomas F, Boyle AL, Burton AJ, Woolfson DN. A Set of De Novo Designed Parallel Heterodimeric Coiled Coils with Quantified Dissociation Constants in the Micromolar to Sub-nanomolar Regime.
J. Am. Chem. Soc. 2013,
135, 5161–5166.
[Google Scholar]
-
27.
Edgell CL, Smith AJ, Beesley JL, Savery NJ, Woolfson DN. De Novo Designed Protein-Interaction Modules for In-Cell Applications.
ACS Synth. Biol. 2020,
9, 427–436.
[Google Scholar]
-
28.
Zhang F, Cong L, Lodato S, Kosuri S, Church GM, Arlotta P. Efficient construction of sequence-specific TAL effectors for modulating mammalian transcription.
Nat. Biotechnol. 2011,
29, 149–153.
[Google Scholar]
-
29.
Geißler R, Scholze H, Hahn S, Streubel J, Bonas U, Behrens SE, et al. Transcriptional activators of human genes with programmable DNA-specificity.
PLoS ONE 2011,
6, 1–7.
[Google Scholar]
-
30.
Hall DB, Struhl K. The VP16 Activation Domain Interacts with Multiple Transcriptional Components as Determined by Protein-Protein Cross-linking in vivo.
J. Biol. Chem. 2002,
277, 46043–46050.
[Google Scholar]
-
31.
Sadowski I, Ma J, Triezenberg S, Ptashne M. GAL4-VP16 is an unusually potent transcriptional activator.
Nature 1988,
335, 563–564.
[Google Scholar]
-
32.
Perez-Pinera P, Kocak DD, Vockley CM, Adler AF, Kabadi AM, Polstein LR, et al. RNA-guided gene activation by CRISPR-Cas9–based transcription factors.
Nat. Methods 2013,
10, 973–976.
[Google Scholar]
-
33.
Jinek M, Chylinski K, Fonfara I, Hauer M, Doudna JA, Charpentier E. A Programmable Dual-RNA–Guided DNA Endonuclease in Adaptive Bacterial Immunity.
Science 2012,
337, 816–821.
[Google Scholar]
-
34.
Qi LS, Larson MH, Gilbert LA, Doudna JA, Weissman JS, Arkin AP, et al. Repurposing CRISPR as an RNA-Guided Platform for Sequence-Specific Control of Gene Expression.
Cell 2013,
152, 1173–1183.
[Google Scholar]
-
35.
Thakore PI, Black JB, Hilton IB, Gersbach CA. Editing the epigenome: technologies for programmable transcription and epigenetic modulation.
Nat. Methods 2016,
13, 127–137.
[Google Scholar]
-
36.
Chavez A, Tuttle M, Pruitt BW, Ewen-Campen B, Chari R, Ter-Ovanesyan D, et al. Comparison of Cas9 activators in multiple species.
Nat. Methods 2016,
13, 563–567.
[Google Scholar]
-
37.
Tanenbaum ME, Gilbert LA, Qi LS, Weissman JS, Vale RD. A protein-tagging system for signal amplification in gene expression and fluorescence imaging.
Cell 2014,
159, 635–646.
[Google Scholar]
-
38.
Lainšček D, Forstnerič V, Mikolič V, Malenšek Š, Pečan P, Benčina M, et al. Coiled-coil heterodimer-based recruitment of an exonuclease to CRISPR/Cas for enhanced gene editing.
Nat. Commun. 2022,
13, 3604.
[Google Scholar]
-
39.
Golob-Urbanc A, Rajčević U, Strmšek Ž, Jerala R. Design of split superantigen fusion proteins for cancer immunotherapy.
J. Biol. Chem. 2019,
294, 6294–6305.
[Google Scholar]
-
40.
Praznik A, Fink T, Franko N, Lonzarić J, Benčina M, Jerala N, et al. Regulation of protein secretion through chemical regulation of endoplasmic reticulum retention signal cleavage.
Nat. Commun. 2022,
13, 1323.
[Google Scholar]
-
41.
Vlahos AE, Kang J, Aldrete CA, Zhu R, Chong LS, Elowitz MB, et al. Protease-controlled secretion and display of intercellular signals.
Nat. Commun. 2022,
13, 912.
[Google Scholar]
-
42.
Jazbec V, Jerala R, Benčina M. Proteolytically Activated CRAC Effectors through Designed Intramolecular Inhibition.
ACS Synth. Biol. 2022,
11, 2756–2765.
[Google Scholar]
-
43.
Honig B, Shapiro L. Adhesion Protein Structure, Molecular Affinities, and Principles of Cell-Cell Recognition.
Cell 2020,
181, 520–535.
[Google Scholar]
-
44.
Chao G, Wannier TM, Gutierrez C, Borders NC, Appleton E, Chadha A, et al. helixCAM: A platform for programmable cellular assembly in bacteria and human cells.
Cell 2022,
185, 3551–3567.e39.
[Google Scholar]
-
45.
Stevens AJ, Harris AR, Gerdts J, Kim KH, Trentesaux C, Ramirez JT, et al. Programming multicellular assembly with synthetic cell adhesion molecules.
Nature 2023,
614, 144–152.
[Google Scholar]
-
46.
-
47.
Cho JH, Collins JJ, Wong WW. Universal Chimeric Antigen Receptors for Multiplexed and Logical Control of T Cell Responses.
Cell 2018,
173, 1426–1438.e11.
[Google Scholar]
-
48.
Cho JH, Okuma A, Sofjan K, Lee S, Collins JJ, Wong WW. Engineering advanced logic and distributed computing in human CAR immune cells.
Nat. Commun. 2021,
12, 792.
[Google Scholar]
-
49.
Li H-S, Wong NM, Tague E, Ngo JT, Khalil AS, Wong WW. High-performance multiplex drug-gated CAR circuits.
Cancer Cell 2022,
40, 1294–1305.e4.
[Google Scholar]
-
50.
Cordoba S, Onuoha S, Thomas S, Pignataro DS, Hough R, Ghorashian S, et al. CAR T cells with dual targeting of CD19 and CD22 in pediatric and young adult patients with relapsed or refractory B cell acute lymphoblastic leukemia: A phase 1 trial.
Nat. Med. 2021,
27, 1797–1805.
[Google Scholar]
-
51.
Gradišar H, Božič S, Doles T, Vengust D, Hafner-Bratkovič I, Mertelj A, et al. Design of a single-chain polypeptide tetrahedron assembled from coiled-coil segments.
Nat. Chem. Biol. 2013,
9, 362–366.
[Google Scholar]
-
52.
Clifford Boldridge W, Ljubetič A, Kim H, Lubock N, Szilágyi D, Lee J, et al. A Multiplexed Bacterial Two-Hybrid for Rapid Characterization of Protein–Protein Interactions and Iterative Protein Design.
bioRxiv 2020.
https://doi.org/10.1101/2020.11.12.377184.
 Anja Golob-Urbanc
1,†
Tjaša Plaper
1,†
Anja Golob-Urbanc
1,†
Tjaša Plaper
1,†