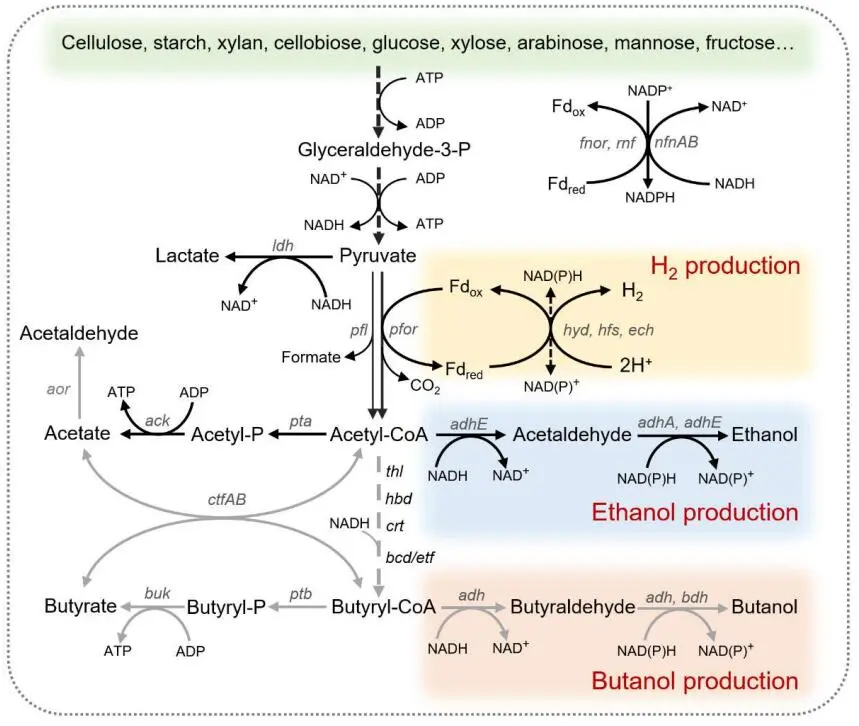
1. Introduction
Fossil fuel, consumed worldwidely, is the non-renewable energy, the exhaustion of which has inspired the interest in the environmentally friendly energy—biofuels [
1,
2]. The usage of biofuels such as bioethanol can reduce fossil fuel consumption, cut down the emission of SO
2 and NO
X, and consequently do good to the environment [
3]. Lignocellulose is seen as an abundant, sustainable and low-cost resource on the earth for biofuels production, which can not only avoid the conflict between food and fuels, but also help to deal with the globally abundant agricultural residues to protect the local environment [
4,
5]. Lignocelluloses such as wheat straw, rice straw and sugarcane bagasse were adopted for biofuels production [
6,
7,
8], suggesting bio-processing of biomass is widely accepted due to the good perspective [
9]. Unfortunately, biofuel engineers must face the fact that cellulosic ethanol still cannot match the low cost of petro-chemicals and first-generation (1G) ethanol (usually produced from cereal grains, sugar and amylaceous plants, popularly in USA and Brazil) [
5,
10]. The second and third-generation ethanol is obtained from lignocellulosic materials and algae, respectively, while the latter is not well developed due to some limited factors like the technology for extracting bioethanol from seaweed [
11]. The conversion of lignocellulosic feedstock into biofuels at competitive prices depends on the efficient conversion of lignocellulosic biomass. In order to achieve this purpose, changes need to be done urgently, such as reducing the enzyme costs, application of enzyme-free saccharification processes, enforcing the co-fermentation of mixed sugars for biofuels production [
5,
12,
13,
14].
Generally, lignocellulose mainly contained lignin (15–20%), cellulose (40–50%) and hemicellulose (25–35%) [
15], which could be decomposed to monosaccharides and disaccharides then followed by biofuels conversion [
16]. Traditional method for lignocellulose pretreatment aims at obtaining glucose by cellulases, including endoglucanase, exoglucanase and β-glucosidase. However, complete hydrolysis of lignocellulose faces many challenges in biomass application [
17,
18]: (1) β-glucosidase is the rate limiting enzyme for cellulose hydrolysis and large amount usage of β-glucosidase will increase the cost; (2) Glucose in the hydrolysate is an inhibitor of cellulase, which leads to low efficiency of complete hydrolysis of cellulose; (3) The presence of glucose in the complete hydrolysate can not only easily cause the microbial pollution, but also induce the carbon catabolite repression (CCR), prolonging the fermentation period. Therefore, pretreatment with incomplete hydrolysate of lignocellulose began to emerge [
19,
20].
Among microorganisms that can ferment biomass to biofuels, thermophilic anaerobic bacteria, with an optimal growth temperature range of 45–70 °C, have some advantages over mesophilic bacteria [
21], such as the ability to co-utilize glucose and xylose by *Thermoanaerobacter* species [
22,
23] (they are further subdivided into the genus *Thermoanaerobacter* and the genus *Thermoanaerobacterium* due to some differences such as the absence or presence of the nitrogenase genes [
24]), lower risk of the contamination for high temperature fermentation [
25], less heat exchange following pretreatment and higher economy to recover ethanol at temperatures over 50 °C by continuous distillation [
26]. Another common and well-studied thermophile is *Clostridium thermocellum*, which could degrade cellulose but the titer of biofuels produced was low [
27,
28,
29]. Here, we mainly focus on *Thermoanaerobacter* species, which might be the potential chassis microorganisms for lignocellulosic biorefinery with the ability to directly utilize hemicellulose or even cellulose [
30,
31]. This review will comprehensively discuss the utilization of substrates, genetic operation platforms constructed and the strategies employed for enhancing the biofuel production of *Thermoanaerobacter* species (). Additionally, the potential of *Thermoanaerobacter* species co-cultured with *C. thermocellum* for consolidated bioprocessing (CBP) will be also addressed.
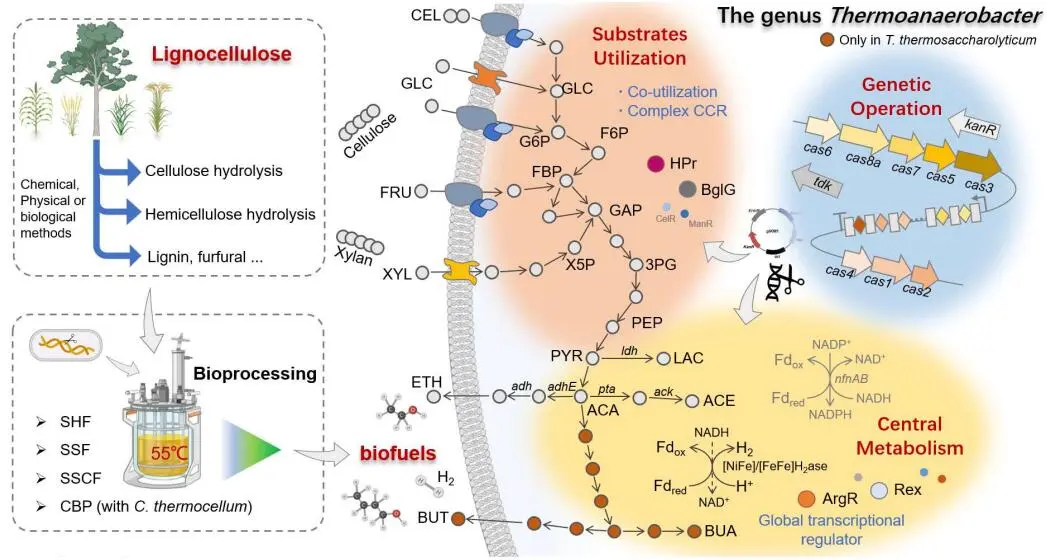
. Diagram of different substrates utilization, genetic operation and main metabolic pathways in *Thermoanaerobacter* species for biofuels fermentation under various bioprocesses. CEL, cellobiose; GLC, glucose; FRU, fructose; XYL, xylose; ETH, ethanol; BUT, butanol; G6P, glucose-6-phosphate; F6P, fructose-6-phosphate; FBP, fructose-1,6-biphosphate; GAP, glyceraldehyde-3-phosphate; X5P, xylulose-5-phosphate; 3PG, 3-phosphoglycerate; PEP, phosphoenolpyruvate; PYR, pyruvate; LAC, lactate; ACA, acetyl-CoA; ACE, acetate; BUA, butyrate.
2. Excellent Performance on Substrates Utilization
*2.1. Xylan and Xylose Metabolism in Thermoanaerobacter Species*
Xylan is the second most abundant plant polysaccharide on the earth, and the D-xylose is the primary ingredient of the hemicellulose from lignocellulosic biomass, both of which are not the common or preferred substrates for most microorganisms [
32]. Xylose metabolism appears to occur exclusively via the traditional xylose isomerase-xylulokinase-pentose phosphate pathway in *Thermoanaerobacter* species [
33].
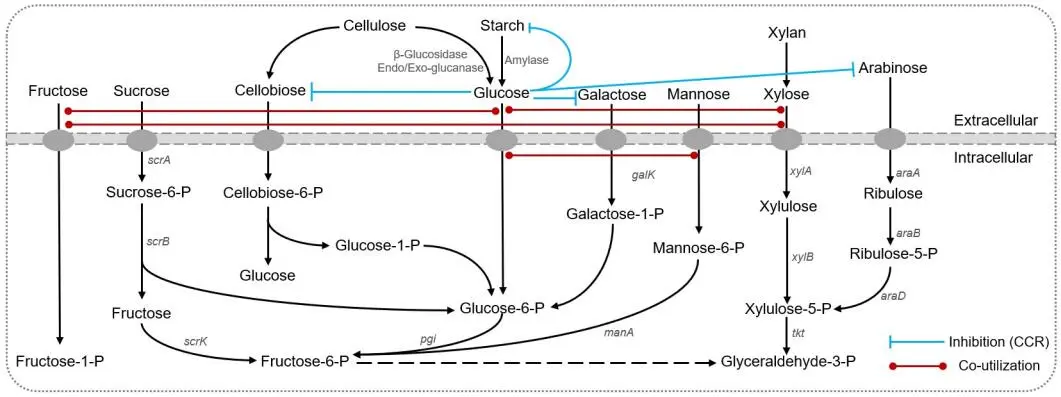
As shown in , the genus *Thermoanaerobacter* has been reported to be an excellent utilizer of xylan and xylose [
30,
34], the feature of which is of particular industrial interest. In *Thermoanaerobacterium aotearoense* SCUT27 (SCUT27), results of the genome annotation revealed 10 genes related to xylose utilization, including one xylose isomerase, one xylulose kinase, and 8 ABC-type xylose transporters [
35]. Compared to *T. thermosaccharolyticum* DSM571, strain SCUT27 possesses two more D-xylose ABC transporter-related genes for xylose utilization. Strain SCUT27 also can degrade xylan directly and four genes related to xylan degradation, such as one gene for β-1,4-xylanase, one for β-xylosidase, and the other 2 for xylanase/chitin deacetylase were found in the genome [
35]. In strain SCUT27, xylanase and β-xylosidase that belong to the glycoside hydrolase family 10 (GH10) have been cloned and characterized [
36,
37]. The function of *Tsac_1445* and *Tsac_1464* for xylan/xylose metabolism was also characterized in *T. saccharolyticum* [
38]. Additionally, xylose delays cell lysis by sustaining coenzyme and ion metabolism, thus enhancing the viability of the thermophiles [
23]. Most of mesophilic strains use glucose preferentially to xylose owing to CCR. However, *Thermoanaerobacter* species can co-utilize xylose and glucose without CCR [
22,
39] as discussed below.
. Main sugar metabolism in genus *Thermoanaerobacter*.
*2.2. Glucose and Xylose Co-Fermentation in Thermoanaerobacter Species*
When cultured with mixed carbon source, microorganisms would selectively use preferred carbon sources by preventing the expression of enzymes participating in secondary substrates [
19]. This regulation is called CCR, which benefits microorganisms for competitions in natural environments but prolongs the fermentation periods and increases the costs for industrial applications. Co-fermentation of multiple carbon sources for the production of high value-added biofuels is a promising production strategy. Among the abundant microorganisms in nature, thermophilic anaerobic bacteria such as *Thermoanaerobacter* sp. X514, *T. saccharolyticum* and strain SCUT27 were reported to have the native ability to utilize glucose and xylose simultaneously [
22,
23,
39]. To reveal molecular mechanism of the glucose and xylose co-utilization in *Thermoanaerobacter* sp. X514, the glycobiome was performed and analyzed via two-dimensional transcriptome sequencing and results revealed that the functional modules of glucose and xylose metabolism were independent and regulated by different transcriptional anti-terminators BglGs, which was quite different from xylose transporters regulated by XylR in *Escherichia coli*, *Bacillus subtilis* and *Clostridium tyrobutyricum* [
23]. However, the functional modules of glucose and xylose metabolism regulated by BglGs independently in *Thermoanaerobacter* sp. X514 could not perfectly explain the release of CCR between glucose and xylose. It’s reported that BglG family transcription antiterminators are key players in the control of carbohydrate catabolism in bacteria [
40,
41]. These antiterminators are generally composed of three modules, an N-terminal RNA-binding domain (CAT) followed by two homologous regulation modules PRD1 and PRD2 that control the RNA binding activity of the effector via phosphorylation on the conserved histidines [
41]. LicT is one member of BglG family transcription antiterminator. The phosphorylation of LicT in PRD1 is catalyzed by P~EIIBBgl in the absence of β-glucosides and inhibits the activity of LicT. In contrast, when β-glucosides are present, P~EIIBBgl is preferably transferring the phosphate group to β-glucosides with the PRD1 of LicT unphosphorylated. However, this format of LicT is also non-functional and requires P~His-HPr to phosphorylate PRD2, in the absence of preferred carbon sources such as glucose. When His-100 and His-169 in PRD1 are unphosphorylated, and one or both conserved His in PRD2 are phosphorylated, LicT is activated and binds bound to RNA on the ribonucleic antiterminator to stabilize an RNA secondary structure for the successful transcription as shown in . The transcription of operons mediated by BglG is also regulated by the preferred carbon sources. In addition to BglG, both MtlR and LevR, as the transcriptional activators, are also activated by P~His-HPr in the absence of preferred carbon sources [
39,
42,
43], which could explain the catabolite control protein A (CcpA)-independent CCR between glucose and cellobiose in strain SCUT27 [
20].
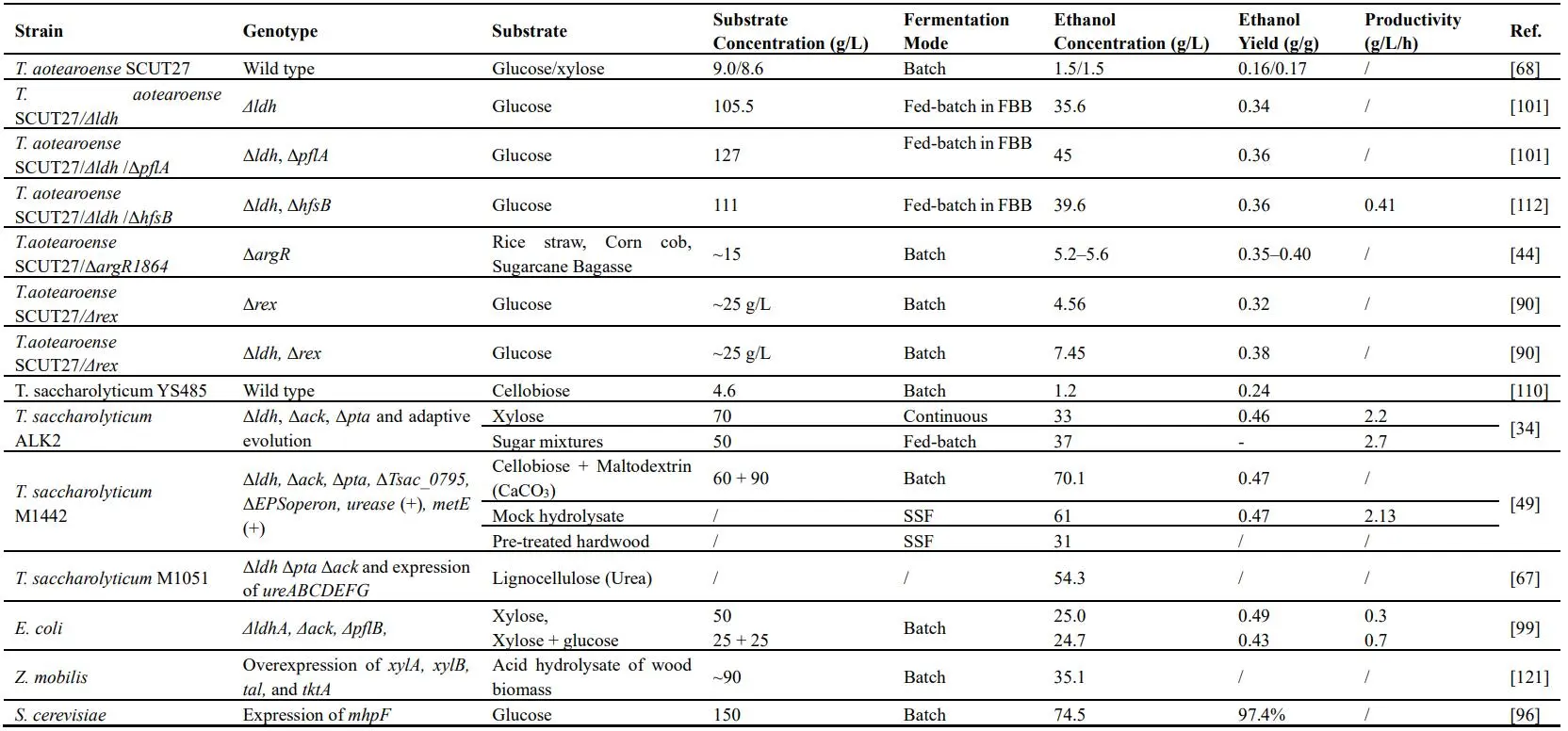
In addition, some other sugars can also be co-utilizaed during fermentation (). With the lignocellulosic hydrolysate as substrate, once glucose and xylose concentrations approached zero, mannose and galactose were consumed simultaneously, the concentration of which were decreased by 98% and 92%, respectively. Finally, 37 g/L ethanol was produced from mixed sugars with an average ethanol productivity of 1.5 g/L/h in *T. saccharolyticum* engineered strain (ALK2) [
34]. Although strain SCUT27 could co-utilize glucose and xylose, the presence of glucose still repressed xylose catabolism in some extent [
44]. Therefore, new microorganism should be obtained by further screening and metabolic engineering. Thermophilic strain like TG57 was selected under the pressure of 2-deoxyglucose with *ccr* and *xylR* deletion/silence and thus was able to simultaneously ferment glucose, xylose and arabinose to produce butanol (7.33 g/L) as the sole solvent [
45]. Other strategies for the enhancing utilization of substrates were also discussed below.
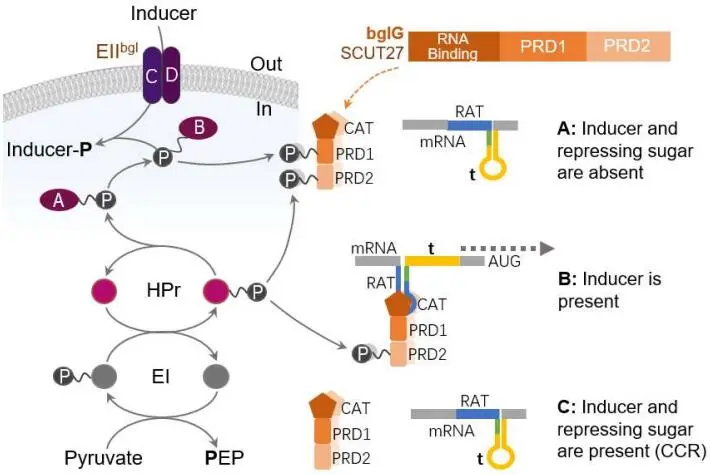
. Mechanism of the transcriptional antiterminator BglG regulation [
40]. (A) In the absence of the inducer, transcription of PTS-encoding genes is inhibited via the formation of a terminator structure (t, yellow) on the mRNA upstream from the start codon. Under this condition, the antiterminator cannot bind to its RNA target, RAT (blue), because the EIIB is mainly phosphorylated and transfers its phosphoryl group to PRD1 of the antiterminator. The absence of a repressing sugar is also expected to allow phosphorylation at the activating domain (PRD2) by P~His-HPr. However, the negative effect of phosphorylation at PRD1 is dominant. In most antiterminator-controlled PTS operons, the two sites RAT and t overlap (green), and the formation of the terminator therefore prevent the formation of the RAT stem-loop. (B) If only an inducer is present, the EIIB as well as PRD1 of the corresponding antiterminator will be present mainly in an unphosphorylated form. Antiterminator-controlled PTSs are usually the sugar transporters with low-capacity and could be phosphorylated at PRD2 by sufficient P~His-HPr in the absence of repressing sugar. The activated antiterminator binds to RAT and thus favors the formation of the RAT stem-loop (in green), thereby preventing the formation of the terminator. (C) If, in addition to the inducing sugar, a repressing carbohydrate is present, the amount of P~His-HPr will be low in the cells. In firmicutes, P~Ser-HPr will also be formed, which further lowers the amount of P~His-HPr. These conditions prevent the phosphorylation at PRD2, resulting in inactivation of most antiterminators.
*2.3. Starch Metabolism in Thermoanaerobacter*
Starch is the abundant carbon source in the nature and high-temperature tolerant amylase is urgently needed to degrade the starch for industrial applications. Many types of thermostable enzymes have been studied for decades and one of them is amylopullulanase (APU), which possesses hydrolytic activity towards α-1,4 and α-1,6-glucosidic linkages and has been well-studied for industrial applications [
46,
47].
*Thermoanaerobacter* species such as *T. saccharolyticum* and strain SCUT27 can use starch, suggesting that thermostable amylases could be obtained from them [
22,
38,
48]. It’s reported that the uitlization of starch was strongly inhibited by 2-deoxyglucose (an analogue of glucose), indicating that the CCR existed between starch and glucose () [
22]. The CCR related genes, *ccpA*, *ptsH* and *hprK* were confirmed in strain SCUT27 and the high affinity between CcpA and phosphorylated HPr was proved via affinity chromatography [
22]. Currie et al. also studied key hydrolases and transporters (Tsac_1344-1349) employed by *T. saccharolyticum* for starch metabolism, and genes expressions among starch, glucose or xylan were compared, showing a host of differences in terms of transporters, sensory genes, and hydrolases [
38]. The highest ethanol titer achieved was 70 g/L in batch culture with a mixture of 90 g/L maltodextrin and 60 g/L cellobiose by *T. saccharolyticum* [
49].
*2.4. Intracellular Cellobiose Metabolism in Thermoanaerobacter*
Cellulose is the major component in most of the lignocelluloses and can be hydrolyzed to glucose by cellulases, including endoglucanase, exoglucanase and β-glucosidase. Cellobiose is abundant in partial hydrolysis of lignocellulosic hydrolysates due to the weak activity of β-glucosidase, and the rate-limiting enzyme in the enzymatic hydrolysis [
50]. There are three types of industrial hosts with the capability to use cellulose: (1) uptaking glucose from cellulose with cellulase pretreatment; (2) utilizing cellobiose extracellularly from partial hydrolysis of cellulose with the ability to produce β-glucosidases; (3) assimilating cellobiose intracellularly from partial cellulose hydrolysis with transporters and intracellular β-glucosidases [
51]. Several critical barriers existed with the complete extracellular hydrolysis of cellulose, such as high costs for the supplementation of β-glucosidase, CCR caused by the released glucose during hydrolysis and energy waste for glucose metabolism instead of cellobiose [
17,
52].
The genus *Thermoanaerobacter* was engineered to overcome these obstacles for intracellular assimilation of cellobiose. Like strain SCUT27, it can assimilate cellobiose intracellularly with CCR existing between cellobiose and glucose [
20,
22]. Cel2 was the only intact operon for cellobiose utilization including integral PTS transport elements and glycoside hydrolase, the transcription of which was regulated by σ54-dependent transcription activator CelRcel2. By replacing the σ54-dependent promoter of Cel2 with the strong promoter from gene *adhE*, the mutant obtained the ability to co-utilize glucose and cellobiose [
20]. In hemicellulose-degrading *T. saccharolyticum*, the biochemical and structure of a glucose-tolerant β-glucosidase was analyzed by Kim et al and results showed that TsaBgl had the unique conformation compared with other structurally known Bgls, and it also could convert cellobiose as well as other soluble cello-oligomers into glucose, providing energy sources for living organisms [
53]. In *T. saccharolyticum*, 41 g/L ethanol was achieved from enzymatic hydrolysate of hardwood containing 96 g/L mixture of glucose and cellobiose [
54] and 70 g/L ethanol was obtained in batch fermentation with a mixture of cellobiose and maltodextrin [
49].
*2.5. Traits about Metabolism of Other Substrates*
In addition to the characteristics of sugar metabolism mentioned above, there are some other traits about sugar metabolism in the genus *Thermoanaerobacter*. In *T. saccharolyticum*, it was found that the utilization of mannose and arabinose was started before glucose and xylose were exhausted, owing to some thermostable enzymes efficient in Thermoanaerobater, such as β-mannanase [
34,
55]. However, in strain SCUT27, the metabolism of galactose and arabinose was inhibited by glucose, while mannose could be co-fermented with glucose [
22]. In *T. saccharolyticum*, arabinose was completely utilized even at high concentrations, the metabolism of which was suppressed significantly at the presence of glucose or mannose. Directed mutations of *hpr* resulted in a reversal of carbon source preferences, and the mutant (His15Asp) utilized arabinose first, then followed by glucose, slowly and incompletely [
54]. Notably, in *T. saccharolyticum*, CCR of galactose, arabinose and cellobiose in the presence of 2-deoxyglucose could be relieved by the HPr (His15Asp) mutation [
39].
The central carbon metabolic network of *T. saccharolyticum* was established based on isotopic tracing and results showed that the metabolism of glucose was exclusively depended on EMP pathway, and both the Entner-Doudoroff (ED) pathway and the oxidative pentose phosphate pathway (oxPPP) were inactive [
56]. Additionally, some thermophiles such as *T. pseudethanolicus* 39E and *T. thermosaccharolyticum* are capable of metabolizing sucrose [
33]. Interestingly, *T. butyriciformans* sp. nov. USBA-019T and *T. thermosaccharolyticum* MJ1, presented different products composition under various substrates [
57,
58]. The main fermentation products of *T. butyriciformans* sp. nov. USBA-019T under glucose, lactose, cellobiose, galactose, arabinose, xylose, starch and xylan were acetate and butyrate. Other products, such as ethanol and lactic acid, also could be produced under starch and cellobiose [
57]. Mannitol and glycerol were the more reduced substrates than glucose, and *T. mathranii* could use mannitol and glycerol to acquire higher ethanol yield [
59]. Recently, it was reported that *T. kivui* as an acetogenic model organism could reduce CO
2 with electrons derived from H
2 or CO, or from organic substrates in the Wood–Ljugdahl pathway (WLP) [
60,
61], further broadening the substrate spectrum of Thermoanaerobater.
3. Genetic Operation Platforms
*3.1. Vectors for Gene Overexpression*
This section would provide a comprehensive description about the current genetic tools and techniques for modification of the genus *Thermoanaerobacter*, which belongs to low-G+C thermophilic and obligate anaerobes in the class *Clostridia*, and is considered to be resistant to genetic engineering due to the difficulty of introducing foreign DNA to cells. Shaw et al. reported the evidence of natural genetic competence in 13 kinds of *Thermoanaerobacter* and *Thermoanaerobacterium* strains, which were previously believed to be difficult to be transformed or genetically recalcitrant [
62]. And in *T. saccharolyticum* JW/SL-YS485, natural competence-mediated DNA incorporation occurs during the exponential growth phase with both replicating plasmid and circular or linear DNA [
62]. The transformation protocol and methods for transformants selection of *T. saccharolyticum* are described by Hon et al [
48].
The earliest and most common vector for transformation and gene-overexpression in the genus *Thermoanaerobacter* such as *Thermoanaerobacterium* sp. strain JW/SL-YS485 was the plasmid pIKM1 containing kanamycin resistance gene (resistance gene of high-temperature kanamycin, *htk*) [
54,
63]. This replicating plasmid was originated from a 2.4 kb thermophilic replicating plasmid in *T. saccharolyticum* strain B6A-RI with KanR, AmpR, and a pUC origin, and then developed to create a convenient cloning vector [
54]. Additionally, pTE16 was constructed based on the *E. coli*-*Clostridium perfringens* shuttle vector pJIR715 with a thermostable chloramphenicol (Cm) resistance cassette. The electro-transformation of T. ethanolicus JW200 was performed with pTE16 and the transformation efficiency was 50 ± 7.4 transformants per μg plasmid DNA [
64]. Based on the hyperthermophilic *Caldicellulosiruptor bescii* replicon pBAS2, Joseph et al. constructed vectors which could be transformed into some thermophiles at 60 °C and maintained in multiple copy with the replication initiation protein RepL [
65]. Le et al. successfully constructed an *E. coli*-*T. ethanolicus* shuttle expression vector pBlu10-*htk* for effective heterologous expression [
66]. Some heterologous genes expression gave the strains expected performances successfully. For example, a heterologous gene gldA encoding an NAD
+-dependent glycerol dehydrogenase was expressed to facilitate NADH regeneration for enhancing ethanol formation in *T. mathranii* [
59]. Genes encoding the urease were integrated in *T. saccharolyticum* to hydrolyze urea [
67].
*3.2. Gene Knockout by Homologous Recombination*
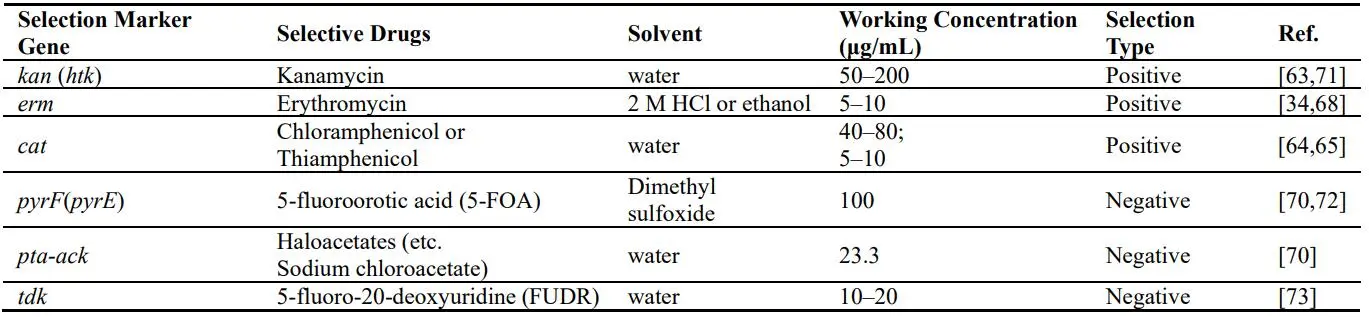
Kanamycin, erythromycin, chloramphenicol and thiamphenicol are four common antibiotics that can be used for the positive selection of the transformants from the genus *Thermoanaerobacter* () [
34,
64]. Kanamycin and thiamphenicol are generally preferred due to the lower background and high efficiency of transformation. Notably, thiamphenicol or chloramphenicol is not used for *T. thermosaccharolyticum*, which carries acyltransferase gene like thiolase (*thl*). And these antibiotics can work in conditions that are suitable for the optimal growth of the genus *Thermoanaerobacter* (i.e., temperature and pH) under the responding working concentration as shown in [
34,
64,
68]. Haloacetates, 5-fluoroorotic acid (5-FOA) and 5-fluoro-2’-deoxyuridine (5-FUDR) are the negative selective drugs and can be adopted for the selection of transformants from genus *Thermoanaerobacter* with the corresponding endogenous negative selection markers genes, *pta-ack*, *pryF* and *tdk* deleted, respectively [
48,
69]. Several kinds of editing ways such as deletions, insertions, or a combination of a deletion and an insertion by homologous recombination in the genome could be achieved by the transformation of the suicide plasmid (circular DNA) like pBluescript II SK(+) or PCR products (linear DNA). Markerless gene deletion strategies were developed based on the counter-selectable markers, such as *pyrF*, *tdk* and *pta*/*ack* genes, which could eliminate the constraints imposed by the small number of positive selection markers for *T. saccharolyticum* [
70]. The design of the suicide vector for markerless editing was also described in several studies [
48,
69,
70].
. List of the practicable selection marker genes and the responding agents for *Thermoanaerobacter* species genetically engineering.
*3.3. Application of CRISPR/Cas Genome Editing System in Thermophiles*
Recently, there are some application of Clustered Regularly-Interspaced Short Palindromic Repeat (CRISPR)/cas (CRISPR associated) system in thermophiles. Compared with traditional makerless editing, the advantage of the CRISPR system is to make the markerless editing easier for just one step. Mougiakos et al. identified and characterized ThermoCas9 from the thermophilic bacterium *Geobacillus thermodenitrificans* T12, which was active between 20 and 70 °C *in vitro* [
74]. And thermoCas9-based engineering tools for gene editing such as gene deletion and transcriptional silencing at 55 °C in *Bacillus smithii* were performed successfully [
75]. Le et al. developed a thermostable Cas9-based system for genome editing of *T. ethanolicus*, in which three genes (*tdk*, *adhE* and *rsp*) were edited successfully [
66].
However, the heterologous Cas9 generally was poisonous to the cells [
76,
77]. Therefore, the native CRISPR/Cas system of some thermophiles was developed. For *Thermoanaerobacter* species, Dai et al. developed the endogenous type I-B CRISPR/Cas system with thymidine kinase gene (*tdk*) as a negative selection marker for multiplex genome editing in strain SCUT27, and the engineered strain SCUT27/Δ*tdk*/Δ*ldh*/Δ*argR* exhibited the significant improvement over the wild type on ethanol production and xylose utilization [
76]. In *C. thermocellum*, Walker et al. also repurposed the native Type I–B CRISPR/cas systems for genome editing [
78]. Additionally, three thermophilic Cas9 variants (heterologous Type II) were tested and results showed that only GeoCas9, isolated from *Geobacillus stearothermophilus*, was active in *C. thermocellum* [
78]. To overcome the limitation of homologous recombination between the repair template and the genome for both the Type I–B and the Type II GeoCas9 system, three novel thermophilic recombinases were tested, and only the exo/beta homologs, isolated from Acidithiobacillus caldus, exhibited the activity in *C. thermocellum* and led to higher genome editing efficiency [
78].
The CRISPR-based transcriptional regulation of repression and activation (CRISRPi & CRISRPa) is a simple and high-throughput genome engineering tool and has been successfully performed in the genus *Clostridium* but rarely in thermophiles [
79,
80,
81,
82]. Nowadays, Ganguly et al. developed a ThermoCas9-based CRISPR interference (CRISPRi) system, which was applied to repress the transcription of lactate dehydrogenase (LDH) and phosphotransacetylase (PTA) genes in T. thermocellum with 67% and 62% enzymatic activity loss, respectively, without gene damage [
83]. In addition to the heterologous Type II CRISPRi system, native Type I system was developed in *Haloferax volcanii*, *E. coli*, *Listeria monocytogenes* and *S. thermophilus* as well as in plants and even human cells [
84,
85,
86], implying the potential application of endogenous type I CRISRPi or CRISRPa system in the genus *Thermoanaerobacter*.
4. Strategies for Enhancing the Biofuel Production
*4.1. Enhancement of Substrates Utilization*
The performance of the utilization of various substrates in *Thermoanaerobacter* has been reviewed above (section 2). However, there are not so many researches about the localization and class of each sugar transporters in *Thermoanaerobacter*, as well as the responding modification. To improve the tolerance to substrates, the strain Δ*ldh*/Δ*pta*-*ack* of *T. saccharolyticum* was continuously cultured for approximately 3000 h with progressively increased feed xylose concentrations and finally the mutant exhibited a greater capacity for xylose consumption. More than 99% of the feed xylose was metabolized at concentrations up to 70 g/L with a mean ethanol yield of 0.46 g/g in continuous culture at pH of 5.2–5.4 without base addition [
34]. In addition to the adaption evolution under high concentration substrates, genetic modification is also a key strategy to improve substrate utilization. Four enzymes from *Herbinix spp.* strain LL1355 which could hydrolyze almost all of glucuronoarabinoxylan linkages were successfully expressed in *T. thermosaccharolyticum*, allowing it to consume more substrates than that of the parent strain (78% vs. 53%) with ethanol yield improved by 24% from corn fiber [
87]. Arginine repressors (ArgRs), the transcriptional repressors of the arginine biosynthesis, were reported to be involved in several key metabolic pathways and might be the global regulator [
88,
89]. In strain SCUT27, the deficiency of *argR* (V518_1864) could greatly improve the co-utilization of glucose and xylose, due to the enhanced activity of xylose isomerase, xylulokinase and the higher energy level [
44]. Deletion of the redox-sensing transcriptional repressor (Rex) also improved the ability of sugar consumption of SCUT27 about 74–131% with the improvement of total ATP concentration and the cofactors NADH and NAD+ concentrations [
90].
Consequently, further work can be carried out to improve the substrates utilization from the following three aspects: (i) Figure out and strengthen the transporters of sugars in *Thermoanaerobacter*; (ii) Understand the components and effectors of CCR in *Thermoanaerobacter*, and realize the co-utilization among the sugars and even expand the substrate spectra; (iii) High substrate tolerant strain should be screened or engineered by adaptive evolution and random mutation.
*4.2. Enforcement of the Metabolic Flux to Biofuels Production*
Biofuels are emerged as a promising alternative to fossil fuels for meeting the rapid increases of energy consumption and accumulation of greenhouse gases (climate change) [
14]. For the genus *Thermoanaerobacter*, there are mainly three biofuels products: ethanol, hydrogen and butanol. Ethanol and hydrogen are principal products for majority of the *Thermoanaerobacter* species while butanol is only produced in a part of the genus *Thermoanaerobacter* like *T. thermosaccharolyticum* [
91], which has the metabolic pathways of butyrate and butanol (). In addition to biofuels, other bioproducts like lactic acid and acetic acid were rarely reported to be produced by *Thermoanaerobacter* species [
60,
61,
92]. So, here we mainly discuss the research of *Thermoanaerobacter* species on biofuels production.
4.2.1. Ethanol Production
Bioethanol is one of the most attractive biofuels as it can serve as a blendstock for gasoline [
93]. As a result, several countries, including the USA, Brazil, China, Canada, India and many EU members, have already proclaimed commitments to reduce their dependence on fossil fuels towards developing bioethanol production [
94]. Currently, bioethanol is commonly produced by *Saccharomyces cerevisiae* [
95,
96], *Zymomonas mobilis* [
97,
98] and *E. coli* [
96,
99]. *Thermoanaerobacter* species are good ethanol producers at 55 °C (the temperature more economical to recover ethanol by continuous distillation) [
54,
100,
101]. Strategies for enhancing the ethanol production mainly include strengthening the pathway of ethanol production, lowering the pathway of by-products, regulating the intracellular cofactor regeneration system as well as ATP level, or adaptive evolution ().
Both pyruvate formate lyase (PFL) and pyruvate ferredoxin oxidoreductase (PFOR) were found to participate in acetyl-CoA formation from pyruvate in genus *Thermoanaerobacter* [
102]. PFOR is the key enzyme for high ethanol production in genus *Thermoanaerobacter*, because it is not inhibited by accumulation of ethanol or NADH while PFOR of *C. thermocellum* is [
103]. Acetyl-CoA would be catalyzed by three major *adh* genes (*adhA*, *adhB*, and *adhE*) for ethanol production [
33]. In addition, a few thermophilic bacterial species within the genus *Thermoanaerobacter* harbored a copy of aldehyde:ferredoxin oxidoreductase (*aor*), which reduced a variety of organic acids to their corresponding aldehydes, showing the AOR-ADH pathway in strains like *Thermoanaerobacter* sp. strain X514, *T. brockii* ssp. Finnii and *T. pseudethanolicus* [
104,
105,
106]. AdhE is the bifunctional enzyme including alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) activities, the deletion of which would reduce the ethanol yield by ~95% [
107]. NADH was the preferred cofactor for both ALDH and ADH activities [
108]. It was reported that deletion of *adhE* reduced NADH-dependent ADH activity by 98%, which did not affect the NADPH-dependent ADH activity [
72]. Notably, some mutants with high ethanol production, showed the increased NADPH-dependent ALDH and ADH activities, in which deletion of *adhA* dramatically reduced NADPH-dependent ADH activity and ethanol production [
72,
108]. Moreover, multiple gene deletions of all annotated alcohol dehydrogenase genes showed that AdhA and AdhE were the two main alcohol dehydrogenases involved in ethanol production in *Thermoanaerobacter* specie [
72]. Thus, AdhA is responsible for the NADPH-ADH activity in *Thermoanaerobacterium* specie, especially in high-ethanol-producing strains, and may also serve as an additional acetaldehyde scavenger [
72]. However, the overexpression of bifunctional or monofunctional ADH genes in wild type or engineered strains seems no improvement in ethanol titer [
54].
. The metabolic pathway for H2, ethanol and butanol synthesis in *Thermoanaerobacter* species. The H2 and ethanol formation pathway in *Thermoanaerobacterium* species while the butanol formation pathway in *T. thermosaccharolyticum* only. *ldh*, lactate dehydrogenase; Fdox, pyruvate:ferredoxin oxidoreductase oxidized; Fdred, pyruvate:ferredoxin oxidoreductase reduced; *hyd*, *hfs* and *ech*, three hydrogenases; *pta*, phosphotrans acetylase; *ack*, acetate kinase; *adh* (*adhE*, *adhhA*), alcohol/aldehyde dehydrogenase; *bdh*, butanol dehydrogenase; *buk*, butyrate kinase; *ptb*, butyryl-CoA dehydrogenase; *ctfAB*, CoA transferase; *thl*, thiolase; *hbd*, β-hydroxybutyryl-CoA dehydrogenase; *crt*, crotonase; *bcd*, butyryl-CoA dehydrogenase; *etf*, electron transferring flavoprotein; *nfnAB*, NADH-dependent reduced ferredoxin:NADP+ oxidoreductase; *pfl*, pyruvate formate lyase; *pfor*, pyruvate ferredoxin oxidoreductase. *aor*, aldehyde:ferredoxin oxidoreductase. The grey arrows show the pathways only existed in a part of strains.
For higher ethanol production by redistribution of carbon flow, metabolic engineering of the genus *Thermoanaerobacter* began with the disruption of the lactate production pathway [
100,
109]. Compared with the wild type, the biomass of mutant Δ*ldh* without lactic acid production, was increased by 31.0–31.4% under glucose or xylose as well as 2.1 to 2.4-fold increases in the yield of ethanol (mole/mole substrate) [
100]. Shaw et al. knocked out the genes involved in organic acid formation (acetate kinase, phosphate acetyltransferase, and L-lactate dehydrogenase) and the resultant strain was able to produce ethanol as the sole detectable product (90% of the theoretical maximum yield) and 37 g/L ethanol was produced from mixed sugars with a maximum ethanol productivity of 2.7 g/L/h [
34]. Shaw et al. investigated three putative hydrogenase enzyme systems in *T. saccharolyticum* and found that the strain with specified *hfs* and *ldh* deletions exhibited an increased ethanol yield from consumed carbohydrates [
110,
111]. The deletion of *hfsB* was also performed in strain SCUT27 and results indicated the enhanced ethanol production was obtained, due to the increased NADH supply and alcohol dehydrogenase activity [
112]. Pyruvate formate lyase-activating protein (PflA) could assist the formation of pyruvate formate lyase (Pfl), which catalyzed pyruvate into acetyl-CoA and formate. The formation of formate also affected the ethanol production by changing the reducing power supply [
101]. In strain SCUT27, the mutant Δ*ldh*/Δ*pflA* was constructed and it could produce 45 g/L ethanol during immobilized cell fermentation, owing to the increased electrons available for ethanol production [
101]. The enhanced ethanol/acetate ratio was also important for ethanol production, which contributed to the resistant to 5% (*v*/*v*) exogenous ethanol [
101]. To decrease amino acid secretion and reduce ammonium assimilation in *C. thermocellum*, type I glutamine synthetase gene (*glnA*) was knocked out and the levels of secreted valine and total amino acids were reduced by 53% and 44%, respectively, along with ethanol yields increased by 53% [
27].
Additionally, reducing equivalents are key factors of alcohol production in *Thermoanaerobacter* species. There are three types of ferredoxin:NAD(P)H oxidoreductase (FNOR) reactions: uncoupled FNOR reaction, proton- or Na+-translocating FNOR reaction (RNF) and NADH-dependent FNOR reaction (NFN) [
113]. NADH-FNOR (Tsac_1705) which transfers electrons from the reduced ferredoxin to NAD+, is the main enzyme responsible for ferredoxin oxidization in the NADH-based ethanol pathway in *T. saccharolyticum* [
113]. In the high-ethanol-producing mutant (Δ*ack*/*pta*), NADPH-FNOR activity was increased compared with that of the wild type, which presumably led to the increased NADPH production [
72,
114]. The electron-bifurcating enzyme complex NfnAB (one of NADH-dependent reduced ferredoxin:NADP+ oxidoreductases, NFN), which transfers electrons from the reduced ferredoxin and NADH to 2 NADP+, is thought to play a key role in linking NAD(P)(H) metabolism with ferredoxin metabolism and has been hypothesized to be the main enzyme for ferredoxin oxidization in mutants of *T. saccharolyticum* for increasing ethanol production [
114]. In strain SCUT27, the NADH/NAD+ ratio in the ∆*nfnAB* mutant was increased [
68]. Interestingly, *adhA* is directly located in the upstream *nfnAB* in *T. saccharolyticum*, showing the potential link between reducing power supply [
114]. Two different mechanisms for stoichiometric yield of ethanol were proposed in *T. saccharolyticum*. One was based on NADPH and NfnAB, and the other was dependent on NADH and a yet undescribed NADH-FNOR (like unknown *rnf* complexes) [
114]. The redox-sensing transcriptional repressor Rex, which was widely reported as the targeting *adhE* by sensing the redox state, was knocked out in strain SCUT27. Results showed that the ethanol yield of strain SCUT27/Δrex was increased by 75–91% within different carbon sources, along with the improvement of total ATP, cofactors concentration as well as the activities of alcohol dehydrogenase [
90]. To regenerate cofactor, *T. mathranii* was fed with mannitol to increase ethanol yield [
59]. To further facilitate NADH regeneration for ethanol formation, a heterologous gene *gldA* encoding a NAD+-dependent glycerol dehydrogenase was overexpressed in *T. mathranii*, and the resultant mutant showed increased the ethanol yield in the presence of glycerol undr xylose [
59]. However, it was also reported that a wide range of genetic modifications for the mixed acid fermentation pathways did not significantly alter the NAD(P)H/NAD(P)+ ratios as well as their concentrations in *T. saccharolyticum* [
115]. It appeared that for each species, the ratios were maintained in a somewhat narrow range: 0.4–0.7 (NADH/NAD+) and 0.5–0.9 (NADPH/NADP+) [
115].
Moreover, adaptive evolution under exogenous ethanol did not always result in mutants of *T. saccharolyticum* that could produce more ethanol and in fact, the mutant obtained usually produced lower titer of ethanol than the wild type [
54]. A strain of *T. pseudethanolicus* with enhanced ethanol tolerance showed high yields of ethanol but lower titer than the parent strain, and NADH-dependent ADH, ALDH, and FNOR activities of the mutant were lost, but NADPH-dependent activities remained [
116]. Interestingly, addition of acetate showed an unexpected stimulatory effect on ethanolic fermentation by *T. ethanolicus* strain 39E and strain X514 while lactate was in general inhibitory to ethanolic fermentation. In *T. saccharolyticum* M2886, the highest ethanol titer achieved was 70 g/L (also the maximun ethanol tolerance) in batch culture under the mixture of cellobiose and maltodextrin by removing exopolysaccharide synthesis gene (*perR*) and replacing the gene of methyglyoxal synthase with *pta*/*ack* genes, after multiple rounds of adaptation and selection [
49]. Furthermore, it suggested that vitamin B12 de novo synthesis in strain X514 had a significant positve effect on ethanol yields in both mono- and coculture [
33]. The vitamin B12 pathway may appear to promote ethanologenesis through ethanolamine and 1,2-propanediol [
23]. Additionally, conversion of acetic acid to acetone in engineered *T. saccharolyticum* resulted in the improved ethanol productivity and titer, and was an attractive low-cost solution to acetic acid inhibition [
117].
To sum up, ethanol production in *Thermoanaerobacter* species could be further improved by conducting more carbon flux to ethanol and weakening the reducing power consumed by the branch-pathways for enhancing ethanol production. In addition, high-ethanol-tolerant strain should be screened by adaptive evolution or random mutation. Finally, future studies on ethanol production would focus on the cheaper and less toxic feedstocks to improve their application value.
4.2.2. H
2 Production
Hydrogen produced from biomass feedstocks is considered as an effective solution toward a decarbonized economy [
118]. Through the circular economy framework, it is possible to produce the sustainable biohydrogen and other bioproducts while address the issues such as waste management [
119]. In *Thermoanaerobacter* species, hydrogen is a necessary product for the disposal of the reducing equivalents. Strains like *T. tengcongensis* are able to convert the reducing equivalents generated during the fermentation from glucose to the acetate and CO
2 by reducing H+ to H
2 (). A unique combination of hydrogenases, a ferredoxin-dependent [NiFe] hydrogenase and an NADH-dependent Fe-only hydrogenase, was found to be responsible for H
2 formation in *T. tengcongensis* [
120].
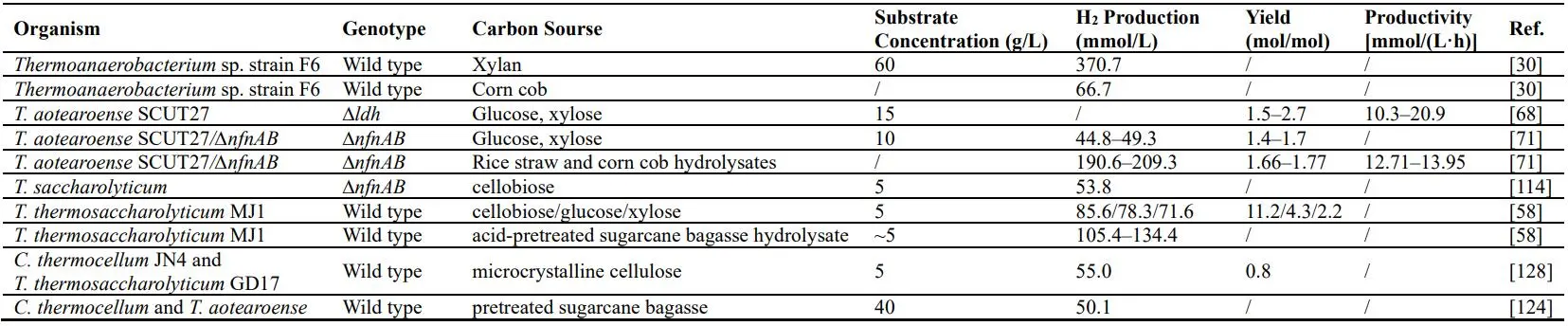
. Summary of the ethanol production by *Thermoanaerobacter* species and common ethanologenic strains.
Three putative hydrogenase systems were identified in *T. saccharolyticum*: A four-gene operon containing two [FeFe]-hydrogenase genes (hfsBD), provisionally termed hfs (hydrogenase-Fe-S), of which the deletion resulted in a growth defect [
111], was found to be the main enzymatic catalyst of hydrogen production; A second hydrogenase gene cluster, a bifurcating NAD(P)H-linked [FeFe]-hydrogenase (hyd), exhibited methyl viologen-linked hydrogenase enzymatic activity and was specific to the transfer of electrons from NAD(P)H to H+; A third gene cluster, a putative [NiFe]-hydrogenase with homology to the ech genes, did not exhibit the hydrogenase activity [
110,
112]. Strains with a deletion of the hfs genes exhibited a 95% decrease in both hydrogen and acetic acid production, and hfsB seemed as the most important gene of the operon hfsABCD [
110]. The genes hyd, ech and hfsABCD, highly homologic to that of *T. saccharolyticum* and *T. tengcongensis*, were also found in strain SCUT27 and proved to be related to H
2 production by gene knockout [
112].
To redirect the NADH flow to hydrogen production, the gene encoding L-lactate dehydrogenase was knocked out, resulting in 2 and 2.5-folds increase of H
2 yield and production rate, respectively [
69]. In the wild type of *T. saccharolyticum*, deletion of nfnAB showed a 46% increase in H
2 formation owing to the enhanced NADH generatation [
114]. Also, in strain SCUT27, the concentration of hydrogen production was increased by 41.1% due to the enhanced NADH/NAD+ ratio in the ∆*nfnAB* mutant [
68]. Additionally, deletion of PFL, potential H
2-uptake (NAD(P)H-dependent) H
2ases and ethanol producing pathways and overexpression of Fd-dependent H
2ases could also increase H
2 yields and production [
122]. However, when H
2 was accumulated in the closed system, the fermentation was partially shifted to ethanol production, due to the high hydrogen partial pressure [p(H
2)] leading to the low NADH-dependent hydrogenase activity [
121].
Furthermore, a number of H
2-producing *Thermoanaerobacter* species were screened and co-cultured with other strains for higher H
2 production. Hu et al. isolated a new *T. thermosaccharolyticum* named MJ1 from paper sludge, whose hydrogen yields could reach 11.18 mol‑H
2/mol cellobiose at an initial sugar concentration of 5 g/L, and the hydrogen production was 111.8 mM under sugarcane bagasse hydrolysate pretreated with 60% dilute‑acid (1%, g/v) [
58]. Li & Zhu took a biphasic fermentation approach for the production of ethanol and hydrogen from cassava pulp. The glucose generated by co-culturing of *C. thermocellum* and strain SCUT27 under cassava pulp was utilized rapidly by subsequently inoculated *S. cerevisiae*, and the final productions of 8.8 g/L ethanol and 4.1 mmol/L hydrogen were achieved [
123]. *C. thermocellum* and strain SCUT27 were also co-cultured with 40 g/L alkali-pretreated sugarcane bagasse (SCB) for bio-hydrogen production, with a final hydrogen production of ~50 mmol/L [
124].
For enhanced thermophilic biohydrogen production with cellulosic wastes, future work can be carried out from the following aspects: First, digging out new thermophilic strains which can naturally produce high level of hydrogen efficiently. Second, providing new strategies for genetic modifications on *Thermoanaerobacter* species for higher hydrogen production, such as developing favorable thermophilic biohydrogen pathways and strengthening enzymes stability. Third, co-cultureing with other microorganisms for further improving the substrates utilization or product yields. Finally, enhancing the biohydrogen production technology under the dark fermentation mode with the implementation of agroindustrial wastes as potential substrates, and exploring the suitable temperature and pH which are the imperative parameters during the dark fermentation.
4.2.3. Butanol Production
Except for the general products of hydrogen and ethanol, some *Thermoanaerobacter* species like *T. thermosaccharolyticum* also have great potential to produce butanol (), which is a renewable alternative to gasoline in vehicles without any modifications in engine system [
2] due to its higher boiling point [
125].
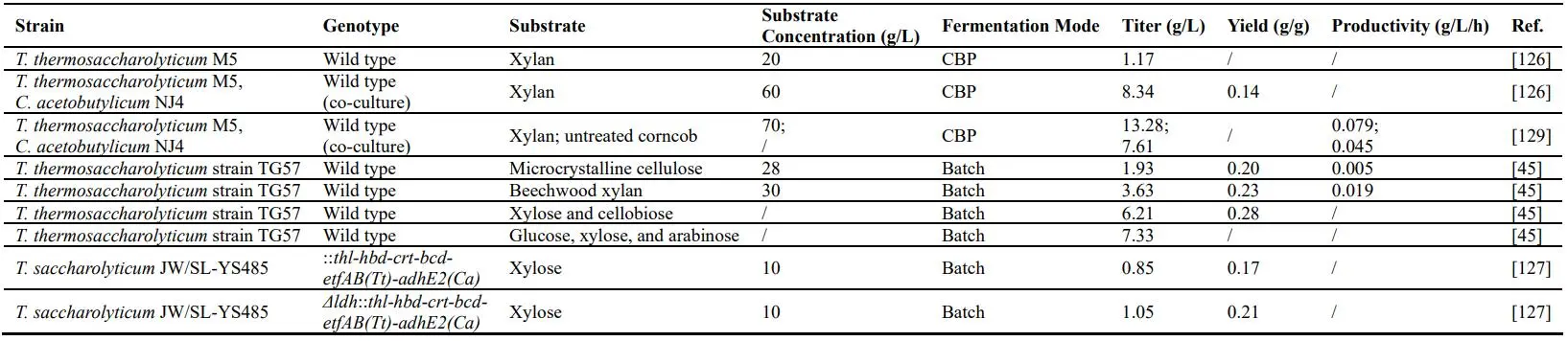
A novel thermophilic and butanogenic *T. thermosaccharolyticum* M5 was isolated and could directly produce butanol from hemicellulose via a unique ethanol–butanol (EB) pathway, with the efficient expression of xylanase, β-xylosidase and the bifunctional alcohol/aldehyde dehydrogenase (AdhE) [
91]. In addition, process optimization based on the characteristic of AdhE was carried out, which further improved the final butanol titer to 1.17 g/L from xylan [
126]. Furthermore, the co-culture of *Thermoanaerobacterium* sp. M5 and *C. acetobutylicum* NJ4 was established to improve the butanol titer to 8.34 g/L from xylan through CBP [
126]. Moreover, Li et al. reported a wild-type *T. thermosaccharolyticum* strain TG57, which was capable of converting microcrystalline cellulose directly to butanol (1.93 g/liter) as the only final product. And the strain TG57 was also able to simultaneously ferment glucose, xylose, and arabinose to produce 7.33 g/L butanol [
45]. Integration of heterologous components of n-butanol pathway (*thl*-*hbd*-*crt*-*bcd*-*etfAB*) from *T. thermosaccharolyticum* DSM 571 resulted in 0.85 g/L butanol from 10 g/L xylose in *T. saccharolyticum* JW/SL-YS485 (21% of the theoretical maximum yield), and resulted in 1.05 g/L butanol from 10 g/L xylose in lactate deficient strain (26% of the theoretical maximum yield) [
127].
In general, the titer of butanol produced by metabolic engineering strategies needs to be improved due to the high toxicity of butanol and the low catalytic activity of enzymes. The following works can be adopted to optimize the butanol biosynthesis: (i) eliminate the byproduct like ethanol to improve the yield of butanol; (ii) obtain the efficient and thermotolerant enzymes by rational engineering or direct evolution for butanol producing; (iii) improve cell tolerance to butanol via transporter engineering or evolution engineering; (iv) screen novel thermophilic butanol producers that could produce much higher butanol titer than that of *C. acetobutylicum*.
. Performance comparison of *Thermoanaerobacter* species for biohydrogen production.
. Butanol production by *Thermoanaerobacter* species.
*4.3. Improvement of the Tolerance to Lignocellulose-Derived Inhibitors*
*T. saccharolyticum* was able to utilize most of the carbohydrates in the hydrolysates of pretreated hardwood, indicating that the strain possessed all the proteins required for efficiently breaking down hemicellulose and could produce twice more ethanol from detoxified hydrolysate than that of untreated hydrolysate (31 g/L vs. 16 g/L) in fermenters [
54]. It’s reported that lignocellulose-derived inhibitors could trigger the accumulation of reactive oxygen species (ROS), resulting in cell damage. Some researches also suggested that inhibitors presented a redox challenge to cells [
54,
130]. Although the inhibitors such as HMF and furfural could be metabolized by strains like *T. saccharolyticum*, some inhibitors such as furfural could delay growth, and others such as vanillin could affect cell yield [
54,
76].To make full use of the lignocellulose, detoxification of lignocellulosic hydrolysates was usually necessary.
Activated carbon was used to reduce the toxicity of both the solid and the liquid feeds [
49]. The treatment of the hydrolysate with diafiltration and/or 1% activated carbon resulted in much higher ethanol titers [
54]. Tolerance of the strains to inhibitors was greatly improved by the presence of cysteine in the medium [
54]. Additionally, acetic acid was also a kind of inhibitor. The ethanol production was stopped when acetic acid levels reached 10–14g/L [
54]. Overliming was demonstrated as an effective method to detoxify polymeric hemicellulose [
54]. An excess of calcium carbonate provided excellent buffering at a pH of 5.5, which is close to the optimum pH for *T. saccharolyticum* [
49]. Since many inhibitors had greater negative effect in minimal media than in yeast extract containing media, addition of yeast extract seemed a simple way for alleviating the suppression [
24,
49].
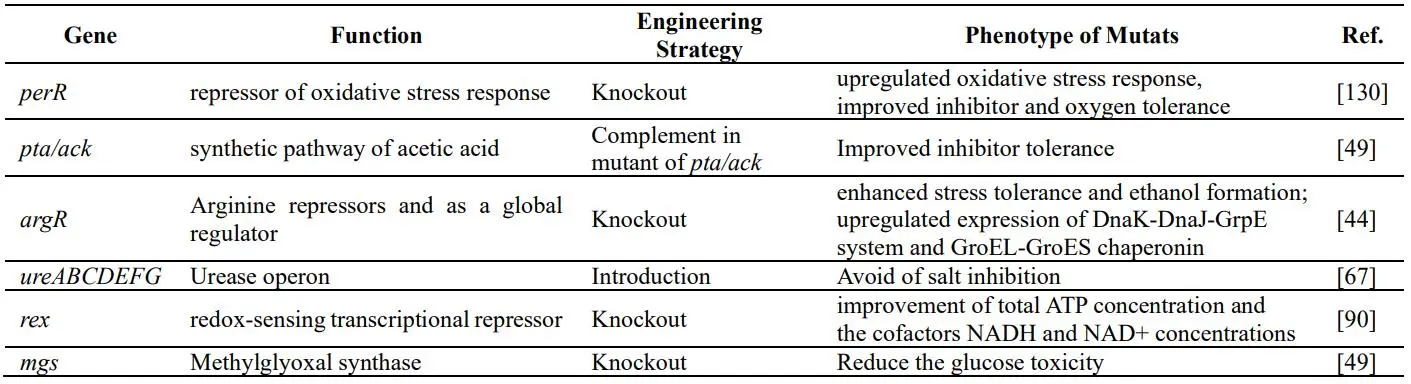
Moreover, genetic modification also played a vital role in strengthening *Thermoanaerobacter* species against lignocellulose-derived inhibitors, as shown in . Elimination of the gene *perR* in *T. saccharolyticum* led to dramatical improvement in inhibitor tolerance and higher oxygen tolerance as well as higher ethanol titer [
49,
130]. The *pta*/*ack* genes seemed the key genes for the inhibitor tolerance phenotype and the dramatic inhibitor tolerance was conferred by restoring the *pta*/*ack* genes into a *pta*/*ack*(−) strain [
49]. In strain SCUT27/ΔargR1864, the improved concentration of intracellular ATP and NAD(H) provided more energy to respond the stresses, which gave the mutant the better cell viability to utilize lignocellulosic hydrolysates for enhancing ethanol production [
44]. In addition, the transcriptome data between SCUT27/ΔargR1864 and wild type showed that ArgR1864 was the negative regulator for chaperonin synthesis. With argR1864 knockout, the mutant showed the upregulated expression of DnaK-DnaJ-GrpE system and GroEL-GroES chaperonin, which not only endowed the strain with the enhanced ability to scavenge the reactive oxygen species (ROS), but also gave the mutant the capability to maintain better cell growth, xylose consumption, acetic acid and ethanol production under the stress of various lignocellulose-derived inhibitors [
131].
It was reported that *T. saccharolyticum* was inhibited by monosaccharides at concentrations higher than 40 g/L, and with methylglyoxal synthase deletion, the mutant could grow at 100 g/L glucose and produce more ethanol from hardwood hydrolysate [
49]. Furthermore, during continuous cultures of *T. thermosaccharolyticum*, the cessation of growth and fermentation at feed exceeding 70 g/L xylose seemed to be attributed to salt formed by neutralizing organic acid rather than the toxicity of ethanol [
132]. Notably, salt inhibition could be avoided by introduction of the urease operon [
67].
. Gene sites engineered for enhanced tolerance against inhibitors in *Thermoanaerobacter* species.
5. Applications of Thermoanaerobacter Species for Various Bioprocessing
When enzymatic hydrolysis is adopted for cellulosic ethanol production, different levels of process integration can be visualized: (i) separate (or sequential) hydrolysis and fermentation (SHF), where the enzymes (cellulases) are used separately from fermentation tanks; (ii) simultaneous saccharification and fermentation (SSF), which consolidates enzymatic hydrolysis with the fermentation of hexose and pentose sequentially; (iii) simultaneous saccharification and cofermentation (SSCF), which further combines the fermentation of hexose and pentose together; and (iv) consolidated bioprocessing (CBP), where cellulases and ethanol are produced in a single reactor [
33]. For *Thermoanaerobacter* species, cellulose hydrolysis is the main limiting factor in the progress of fermentation [
34].
A fed-batch fermentation with *T. saccharolyticum* M2886 was performed and 50 g/L ethanol after 60 h cultivation under pretreated hardwood was obtained in SHF mode [
49]. 37 g/L ethanol was produced by strain ALK2 in SHF at 50°C which had a 2.5-fold decrease in cellulase loading compared with that of *S. cerevisiae* at 37 °C [
34]. For SSF, mesophiles ferment well at ≤37 °C and thus require substantially higher cellulase loadings compared with the thermophiles cultured under 50~60 °C (the optimal temperature for both enzymes activity and the growth of *Thermoanaerobacter*) [
34,
54]. Pre-hydrolysis for a long time facilitates the cellulose conversion, but the high initial sugar concentrations >50 g/L (usually due to high initial enzyme dose) would inhibit the cells growth [
54]. SSF of avicel with *T. saccharolyticum* in batch mode at an initial concentration of 50 g/L after pretreatment with a commercial cellulase showed the better performance than that that of yeast (2.5-fold increase of cellulase loading). And further reductions in cellulase loading could be anticipated as more thermostable cellulases are developed [
34,
133]. Under the optimal substrate “mock hydrolysate” and conditions such as temperature, pH-control strategy, inoculum size, cellulase mixture and feeding regimen, *T. saccharolyticum* could produce over 61 g/L ethanol in thermophilic SSF [
54]. Moreover, 31 g/L ethanol was produced in SSF of pre-treated hardwood and 26 g/L ethanol was obtained with a hardwood hemicellulose extract [
49]. Fermentation conditions were also developed to get the highest possible ethanol titer in a SSCF configuration under the conditions expected from pre-treated hardwood (100 g/L purified cellulose, 10 g/L acetic acid, 35 g/L xylose and 20 g/L glucose), and more than 60 g/L ethanol (greater than 90% yield) was achieved at 93 h cultivation by strain M1442 [
49]. Additionally, cell lysis behavior was observed when the strain grew too fast, but it could be prevented under mixed sugars with a slower growth rate, or in the medium with high concentration of magnesium or low pH [
54].
A preferred CBP system can directly convert both cellulose and hemicellulose into target products without the costly hydrolytic enzyme cocktail. Cellulolytic strains like *C. thermocellum* are necessary to be used to hydrolyze and ferment all of the sugars in the broth with *Thermoanaerobacter* species. It was reported that the coculture of *C. thermocellum* JN4 and *T. thermosaccharolyticum* GD17 could significantly improve the efficiency of cellulosic biofuel [
134]. Biodegradation of sugarcane bagasse (SCB) was remarkably improved with the addition of non-ionic surfactant (etc. Triton X-100) to the CBP system with *C. thermocellum* [
135]. The applications of CBP for hydrogen production have been shown above (section 4.2.2). Lynd et al. also listed some detailed advances in CBP using *C. thermocellum* and *T. saccharolyticum* [
136]. While these production schemes represent increasing levels of simplification through process consolidation, consolidation of multiple steps often results in a loss of process efficiency. Improving the efficiency of individual step, such as cellulose hydrolysis and ethanol fermentation, is still the important task for the development of economically feasible cellulosic biofuel.
6. Conclusions
In this paper, the traits of *Thermoanaerobacter* species on substrate utilization, their current platforms of genetic operation, and the strategies for enhanced biofuels (ethanol, hydrogen and butanol) production were reviewed. Numerous ways have been developed to enhance the sugar consumption and biofuel production in *Thermoanaerobacter* species. However, the production of ethanol is far less than that of *S. cerevisiae* and *Z. mobilis*. Achievement of biobutanol with high titer is still difficult due to the overwhelming metabolic stress. Further research should be performed as follows to establish efficient microbial cell factories for the genus *Thermoanaerobacter* to produce biofuels or other bioproducts:
(i) Better understand the genes and pathways involved in carbohydrate transport and metabolism, and address the mechanism of the co-utilization between glucose and xylose, which can shorten the fermentation period and make full use of the lignocellulosic materials.
(ii) Further develop effective elements of genetic operation such as useable fluorescent proteins for *Thermoanaerobacter* species, and technologies in large-scale engineering like CRISPRi and CRISPRa, for conveniently analyzing metabolic flux, engineering the high-producing strains and exploring the physiological mechanism of the bacteria.
(iii) The whole-cell models of *Thermoanaerobacter* species should be built to analyze the cellular networks and find key bottlenecks in biofuels production. Then the corresponding metabolic engineering strategies could be used for enhancing titer and yield of the products.
(iv) Tolerance to the inhibitors, high concentration of substrates and products, always poses a great challenge for *Thermoanaerobacter* species. It is necessary to reveal the mechanism and lay the fundation to improve the resistance of cells to several inhibitors for higher biofuels production.
(v) Optimize the fermentation process and construct microbial consortia to reduce the metabolic burden. More strategies and techniques should be practiced on perspective applications of *Thermoanaerobacter* species for consolidated bioprocessing.
(vi) Other valuable biochemicals and bioproducts such as poly-3-hydroxybutyrate (PHB), polylactic acid (PLA) and fatty acid esters could be further developed in *Thermoanaerobacter* species once getting the responding thermostable enzymes.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This work was supported by the National Natural Science Foundation of China (21878103) and the Natural Science Foundation of Guangdong Province of China (2018A030310368).
Declaration of Competing Interest
The authors declare that they have no competing interests.
References
-
1.
El-Dalatony MM, Saha S, Govindwar SP, Abou-Shanab RAI, Jeon BH. Biological Conversion of Amino Acids to Higher Alcohols.
Trends Biotechnol. 2019,
37, 855–869.
[Google Scholar]
-
2.
Riaz S, Mazhar S, Abidi SH, Syed Q, Abbas N, Saleem Y, et al. Biobutanol production from sustainable biomass process of anaerobic ABE fermentation for industrial applications.
Arch. Microbiol. 2022,
204, 672.
[Google Scholar]
-
3.
Busic A, Mardetko N, Kundas S, Morzak G, Belskaya H, Ivancic Santek M, et al. Bioethanol Production from Renewable Raw Materials and Its Separation and Purification: A Review.
Food Technol. Biotechnol. 2018,
56, 289–311.
[Google Scholar]
-
4.
Broda M, Yelle DJ, Serwańska K. Bioethanol Production from Lignocellulosic Biomass-Challenges and Solutions.
Molecules 2022,
27, 8717.
[Google Scholar]
-
5.
Abo BO, Gao M, Wang Y, Wu C, Ma H, Wang Q. Lignocellulosic biomass for bioethanol: An overview on pretreatment, hydrolysis and fermentation processes.
Rev. Environ. Health 2019,
34, 57–68.
[Google Scholar]
-
6.
Guo Y, Liu Y, Guan M, Tang H, Wang Z, Lin L, et al. Production of butanol from lignocellulosic biomass: recent advances, challenges, and prospects.
RSC Adv. 2022,
12, 18848–18863.
[Google Scholar]
-
7.
Kaparaju P, Serrano M, Thomsen AB, Kongjan P, Angelidaki I. Bioethanol, biohydrogen and biogas production from wheat straw in a biorefinery concept.
Bioresour. Technol. 2009,
100, 2562–2568.
[Google Scholar]
-
8.
Kim JS, Park SC, Kim JW, Park JC, Park SM, Lee JS. Production of bioethanol from lignocellulose: Status and perspectives in Korea.
Bioresour. Technol. 2010,
101, 4801–4805.
[Google Scholar]
-
9.
Awasthi MK, Sarsaiya S, Patel A, Juneja A, Singh RP, Yan B, et al. Refining biomass residues for sustainable energy and bio-products: An assessment of technology, its importance, and strategic applications in circular bio-economy.
Renew. Sustain. Energy Rev. 2020,
127, 109876.
[Google Scholar]
-
10.
Parascanu MM, Sanchez N, Sandoval-Salas F, Carreto CM, Soreanu G, Sanchez-Silva L. Environmental and economic analysis of bioethanol production from sugarcane molasses and agave juice.
Environ. Sci. Pollut. Res. Int. 2021,
28, 64374–64393.
[Google Scholar]
-
11.
Aparicio E, Rodríguez-Jasso RM, Pinales-Márquez CD, Loredo-Treviño A, Robledo-Olivo A, Aguilar CN, et al. High-pressure technology for Sargassum spp biomass pretreatment and fractionation in the third generation of bioethanol production.
Bioresour. Technol. 2021,
329, 124935.
[Google Scholar]
-
12.
Kim SR, Ha SJ, Wei N, Oh EJ, Jin YS. Simultaneous co-fermentation of mixed sugars: a promising strategy for producing cellulosic ethanol.
Trends Biotechnol. 2012,
30, 274–282.
[Google Scholar]
-
13.
Kazemi Shariat Panahi H, Dehhaghi M, Dehhaghi S, Guillemin GJ, Shiung Lam S, Aghbashlo M, et al. Engineered bacteria for valorizing lignocellulosic biomass into bioethanol.
Bioresour. Technol. 2021,
344, 126212.
[Google Scholar]
-
14.
Liu Y, Cruz-Morales P, Zargar A, Belcher MS, Pang B, Englund E, et al. Biofuels for a sustainable future.
Cell 2021,
184, 1636–1647.
[Google Scholar]
-
15.
Dietrich K, Dumont MJ, Del Rio LF, Orsat V. Sustainable PHA production in integrated lignocellulose biorefineries.
New Biotechnol. 2019,
49, 161–168.
[Google Scholar]
-
16.
Zhang C, Wen H, Chen C, Cai D, Fu C, Li P, et al. Simultaneous saccharification and juice co-fermentation for high-titer ethanol production using sweet sorghum stalk.
Renew. Energy 2019,
134, 44–53.
[Google Scholar]
-
17.
Parisutham V, Chandran SP, Mukhopadhyay A, Lee SK, Keasling JD. Intracellular cellobiose metabolism and its applications in lignocellulose-based biorefineries.
Bioresour. Technol. 2017,
239, 496–506.
[Google Scholar]
-
18.
Zhao T, Tashiro Y, Zheng J, Sakai K, Sonomoto K. Semi-hydrolysis with low enzyme loading leads to highly effective butanol fermentation.
Bioresour. Technol. 2018,
264, 335–342.
[Google Scholar]
-
19.
Gorke B, Stulke J. Carbon catabolite repression in bacteria: many ways to make the most out of nutrients.
Nat. Rev. Microbiol. 2008,
6, 613–624.
[Google Scholar]
-
20.
Qu CY, Zhang Y, Dai KQ, Fu HX, Wang JF. Metabolic engineering of
Thermoanaerobacterium aotearoense SCUT27 for glucose and cellobiose co-utilization by identification and overexpression of the endogenous cellobiose operon.
Biochem. Eng. J. 2021,
167, 107922.
[Google Scholar]
-
21.
Rahayu F, Kawai Y, Iwasaki Y, Yoshida K, Kita A, Tajima T, et al. Thermophilic ethanol fermentation from lignocellulose hydrolysate by genetically engineered Moorella thermoacetica.
Bioresour. Technol. 2017,
245, 1393–1399.
[Google Scholar]
-
22.
Zhu M, Lu Y, Wang J, Li S, Wang X. Carbon Catabolite Repression and the Related Genes of ccpA, ptsH and hprK in
Thermoanaerobacterium aotearoense.
PloS ONE 2015,
10, e0142121.
[Google Scholar]
-
23.
Lin L, Song H, Tu Q, Qin Y, Zhou A, Liu W, et al. The
Thermoanaerobacter glycobiome reveals mechanisms of pentose and hexose co-utilization in bacteria.
PLoS Genet. 2011,
7, e1002318.
[Google Scholar]
-
24.
Currie DH, Raman B, Gowen CM, Tschaplinski TJ, Land ML, Brown SD, et al. Genome-scale resources for
Thermoanaerobacterium saccharolyticum.
BMC Syst. Biol. 2015,
9, 30.
[Google Scholar]
-
25.
Zhou J, Ouyang J, Xu Q, Zheng Z. Cost-effective simultaneous saccharification and fermentation of l-lactic acid from bagasse sulfite pulp by Bacillus coagulans CC17.
Bioresour. Technol. 2016,
222, 431–438.
[Google Scholar]
-
26.
Taylor MP, Eley KL, Martin S, Tuffin MI, Burton SG, Cowan DA. Thermophilic ethanologenesis: Future prospects for second-generation bioethanol production.
Trends Biotechnol. 2009,
27, 398–405.
[Google Scholar]
-
27.
Rydzak T, Garcia D, Stevenson DM, Sladek M, Klingeman DM, Holwerda EK, et al. Deletion of Type I glutamine synthetase deregulates nitrogen metabolism and increases ethanol production in
Clostridium thermocellum.
Metab. Eng. 2017,,
41, 182–191.
[Google Scholar]
-
28.
Holwerda EK, Olson DG, Ruppertsberger NM, Stevenson DM, Murphy SJL, Maloney MI, et al. Metabolic and evolutionary responses of
Clostridium thermocellum to genetic interventions aimed at improving ethanol production.
Biotechnol. Biofuels 2020,
13, 40.
[Google Scholar]
-
29.
Mazzoli R, Olson DG, Lynd LR. Construction of lactic acid overproducing
Clostridium thermocellum through enhancement of lactate dehydrogenase expression.
Enzyme Microb. Technol. 2020,
141, 109645.
[Google Scholar]
-
30.
Jiang Y, Lu J, Lv Y, Wu R, Dong W, Zhou J, et al. Efficient hydrogen production from lignocellulosic feedstocks by a newly isolated thermophlic
Thermoanaerobacterium sp. strain F6.
Int. J. Hydrog. Energy 2019,
44, 14380–14386.
[Google Scholar]
-
31.
O-Thong S, Prasertsan P, Karakashev D, Angelidaki I. Thermophilic fermentative hydrogen production by the newly isolated
Thermoanaerobacterium thermosaccharolyticum PSU-2.
Int. J. Hydrog. Energy 2008,
33, 1204–1214.
[Google Scholar]
-
32.
Deutschmann R, Dekker RF. From plant biomass to bio-based chemicals: latest developments in xylan research.
Biotechnol. Adv. 2012,
30, 1627–1640.
[Google Scholar]
-
33.
Hemme CL, Fields MW, He Q, Deng Y, Lin L, Tu Q, et al. Correlation of genomic and physiological traits of thermoanaerobacter species with biofuel yields.
Appl. Environ Microbiol. 2011,
77, 7998–8008.
[Google Scholar]
-
34.
Shaw AJ, Podkaminer KK, Desai SG, Bardsley JS, Rogers SR, Thorne PG, et al. Metabolic engineering of a thermophilic bacterium to produce ethanol at high yield.
Proc. Natl. Acad. Sci. USA 2008,
105, 13769–13774.
[Google Scholar]
-
35.
Ai H, Zhang J, Yang M, Yu P, Li S, Zhu M, et al. Draft Genome Sequence of an Anaerobic, Thermophilic Bacterium,
Thermoanaerobacterium aotearoense SCUT27, Isolated from a Hot Spring in China.
Genome Announc. 2014,
2, e00041-14.
[Google Scholar]
-
36.
Huang X, Li Z, Du C, Wang J, Li S. Improved Expression and Characterization of a Multidomain Xylanase from
Thermoanaerobacterium aotearoense SCUT27 in Bacillus subtilis.
J. Agric. Food Chem. 2015,
63, 6430–6439.
[Google Scholar]
-
37.
Xu T, Huang X, Li Z, Lin CS, Li S. Enhanced Purification Efficiency and Thermal Tolerance of
Thermoanaerobacterium aotearoense β-Xylosidase through Aggregation Triggered by Short Peptides.
J. Agric. Food Chem. 2018,
66, 4182–4188.
[Google Scholar]
-
38.
Currie DH, Guss AM, Herring CD, Giannone RJ, Johnson CM, Lankford PK, et al. Profile of secreted hydrolases, associated proteins, and SlpA in
Thermoanaerobacterium saccharolyticum during the degradation of hemicellulose.
Appl. Environ. Microbiol. 2014,
80, 5001–5011.
[Google Scholar]
-
39.
Tsakraklides V, Shaw AJ, Miller BB, Hogsett DA, Herring CD. Carbon catabolite repression in
Thermoanaerobacterium saccharolyticum.
Biotechnol. Biofuels 2012,
5, 85.
[Google Scholar]
-
40.
Deutscher J, Francke C, Postma PW. How phosphotransferase system-related protein phosphorylation regulates carbohydrate metabolism in bacteria.
Microbiol. Mol. Biol. Rev. 2006,
70, 939–1031.
[Google Scholar]
-
41.
Deutscher J, Ake FM, Derkaoui M, Zebre AC, Cao TN, Bouraoui H, et al. The bacterial phosphoenolpyruvate:carbohydrate phosphotransferase system: regulation by protein phosphorylation and phosphorylation-dependent protein-protein interactions.
Microbiol. Mol. Biol. Rev. 2014,
78, 231–256.
[Google Scholar]
-
42.
Galinier A, Deutscher J. Sophisticated Regulation of Transcriptional Factors by the Bacterial Phosphoenolpyruvate: Sugar Phosphotransferase System.
J. Mol. Biol. 2017,
429, 773–789.
[Google Scholar]
-
43.
Nie X, Yang B, Zhang L, Gu Y, Yang S, Jiang W, et al. PTS regulation domain-containing transcriptional activator CelR and sigma factor σ54 control cellobiose utilization in
Clostridium acetobutylicum.
Mol. Microbiol. 2016,
100, 289–302.
[Google Scholar]
-
44.
Qu C, Chen L, Fu H, Wang J. Engineering
Thermoanaerobacterium aotearoense SCUT27 with argR knockout for enhanced ethanol production from lignocellulosic hydrolysates.
Bioresour. Technol. 2020,
310, 123435.
[Google Scholar]
-
45.
Li T, Zhang C, Yang KL, He J. Unique genetic cassettes in a
Thermoanaerobacterium contribute to simultaneous conversion of cellulose and monosugars into butanol.
Sci. Adv. 2018,
4, e1701475.
[Google Scholar]
-
46.
Lin HY, Chuang HH, Lin FP. Biochemical characterization of engineered amylopullulanase from
Thermoanaerobacter ethanolicus 39E-implicating the non-necessity of its 100 C-terminal amino acid residues.
Extremophiles 2008,
12, 641–650.
[Google Scholar]
-
47.
Vieille C, Zeikus GJ. Hyperthermophilic enzymes: Sources, uses, and molecular mechanisms for thermostability.
Microbiol. Mol. Biol. Rev. 2001,
65, 1–43.
[Google Scholar]
-
48.
Hon S, Tian L, Zheng T, Cui J, Lynd LR, Olson DG. Methods for Metabolic Engineering of
Thermoanaerobacterium saccharolyticum.
Methods Mol. Biol. 2020,
2096, 21–43.
[Google Scholar]
-
49.
Herring CD, Kenealy WR, Joe Shaw A, Covalla SF, Olson DG, Zhang J, et al. Strain and bioprocess improvement of a thermophilic anaerobe for the production of ethanol from wood.
Biotechnol. Biofuels 2016,
9, 125.
[Google Scholar]
-
50.
Singhania RR, Patel AK, Sukumaran RK, Larroche C, Pandey A. Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production.
Bioresour. Technol. 2013,
127, 500–507.
[Google Scholar]
-
51.
Vinuselvi P, Lee SK. Engineering Escherichia coli for efficient cellobiose utilization.
Appl. Microbiol. Biotechnol. 2011,
92, 125–132.
[Google Scholar]
-
52.
Parisutham V, Kim TH, Lee SK. Feasibilities of consolidated bioprocessing microbes: From pretreatment to biofuel production.
Bioresour. Technol. 2014,
161, 431–440.
[Google Scholar]
-
53.
Kim IJ, Bornscheuer UT, Nam KH. Biochemical and Structural Analysis of a Glucose-Tolerant β-Glucosidase from the Hemicellulose-Degrading
Thermoanaerobacterium saccharolyticum.
Molecules 2022,
27, 290.
[Google Scholar]
-
54.
Herring CD, Kenealy WR, Shaw AJ, Raman B, Tschaplinski TJ, Brown SD, et al. Final Report on Development of Thermoanaerobacterium saccharolyticum for the Conversion of Lignocellulose to Ethanol; U.S. Department of Energy OSTI: Oak Ridge, TN, USA, 2012; Technical Report.
-
55.
Zhu M, Zhang L, Yang F, Cha Y, Li S, Zhuo M, Huang S, Li J. A Recombinant β-Mannanase from
Thermoanaerobacterium aotearoense SCUT27: Biochemical Characterization and Its Thermostability Improvement.
J. Agric. Food Chem. 2020,
68, 818–825.
[Google Scholar]
-
56.
Jacobson TB, Korosh TK, Stevenson DM, Foster C, Maranas C, Olson DG, Lynd LR, Amador-Noguez D. In Vivo Thermodynamic Analysis of Glycolysis in
Clostridium thermocellum and
Thermoanaerobacterium saccharolyticum Using (13)C and (2)H Tracers.
mSystems 2020,
5, e00736-19.
[Google Scholar]
-
57.
Lopez G, Canas-Duarte SJ, Pinzon-Velasco AM, Vega-Vela NE, Rodriguez M, Restrepo S, Baena S. Description of a new anaerobic thermophilic bacterium,
Thermoanaerobacterium butyriciformans sp. nov.
Syst. Appl. Microbiol. 2017,
40, 86–91.
[Google Scholar]
-
58.
Hu BB, Zhu MJ. Direct hydrogen production from dilute-acid pretreated sugarcane bagasse hydrolysate using the newly isolated T
hermoanaerobacterium thermosaccharolyticum MJ1.
Microb. Cell Factories 2017,
16, 77.
[Google Scholar]
-
59.
Yao S, Mikkelsen MJ. Metabolic engineering to improve ethanol production in hermoanaerobacter mathranii.
Appl. Microbiol. Biotechnol. 2010,
88, 199–208.
[Google Scholar]
-
60.
Katsyv A, Jain S, Basen M, Muller V. Electron carriers involved in autotrophic and heterotrophic acetogenesis in the thermophilic bacterium
Thermoanaerobacter kivui.
Extremophiles 2021,
25, 513–526.
[Google Scholar]
-
61.
Weghoff MC, Muller V. CO Metabolism in the Thermophilic Acetogen
Thermoanaerobacter kivui.
Appl. Environ. Microbiol. 2016,
82, 2312–2319.
[Google Scholar]
-
62.
Shaw AJ, Hogsett DA, Lynd LR. Natural competence in
Thermoanaerobacter and
Thermoanaerobacterium species.
Appl. Environ. Microbiol. 2010,
76, 4713–4719.
[Google Scholar]
-
63.
Mai V, Lorenz WW, Wiegel J. Transformation of
Thermoanaerobacterium sp. strain JW/SL-YS485 with plasmid pIKM1 conferring kanamycin resistance.
FEMS Microbiol. Lett. 1997,
148, 163–167.
[Google Scholar]
-
64.
Peng H, Fu B, Mao Z, Shao W. Electrotransformation of
Thermoanaerobacter ethanolicus JW200.
Biotechnol. Lett. 2006,
28, 1913–1917.
[Google Scholar]
-
65.
Groom J, Chung D, Olson DG, Lynd LR, Guss AM, Westpheling J. Promiscuous plasmid replication in thermophiles: Use of a novel hyperthermophilic replicon for genetic manipulation of
Clostridium thermocellum at its optimum growth temperature.
Metab. Eng. Commun. 2016,
3, 30–38.
[Google Scholar]
-
66.
Le Y, Fu Y, Sun J. Genome Editing of the Anaerobic Thermophile
Thermoanaerobacter ethanolicus Using Thermostable Cas9.
Appl. Environ Microbiol. 2020,
87, e01773-20.
[Google Scholar]
-
67.
Shaw AJ, Covalla SF, Miller BB, Firliet BT, Hogsett DA, Herring CD. Urease expression in a
Thermoanaerobacterium saccharolyticum ethanologen allows high titer ethanol production.
Metab. Eng. 2012,
14, 528–532.
[Google Scholar]
-
68.
Li Y, Hu J, Qu C, Chen L, Guo X, Fu H, et al. Engineered
Thermoanaerobacterium aotearoense with nfnAB knockout for improved hydrogen production from lignocellulose hydrolysates.
Biotechnol. Biofuels 2019,
12, 214.
[Google Scholar]
-
69.
Li S, Lai C, Cai Y, Yang X, Yang S, Zhu M, et al. High efficiency hydrogen production from glucose/xylose by the ldh-deleted
Thermoanaerobacterium strain.
Bioresour. Technol. 2010,
101, 8718–8724.
[Google Scholar]
-
70.
Shaw AJ, Covalla SF, Hogsett DA, Herring CD. Marker removal system for
Thermoanaerobacterium saccharolyticum and development of a markerless ethanologen.
Appl. Environ. Microbiol. 2011,
77, 2534–2536.
[Google Scholar]
-
71.
Basen M, Geiger I, Henke L, Müller V, Elliot MA. A Genetic System for the Thermophilic Acetogenic Bacterium
Thermoanaerobacter kivui.
Appl. Environ Microbiol. 2018,
84, e02210-17.
[Google Scholar]
-
72.
Zheng T, Olson DG, Murphy SJ, Shao X, Tian L, Lynd LR. Both
adhE and a Separate NADPH-Dependent Alcohol Dehydrogenase Gene,
adhA, Are Necessary for High Ethanol Production in
Thermoanaerobacterium saccharolyticum.
J. Bacteriol. 2017,
199, e00542-16.
[Google Scholar]
-
73.
Shao X, Zhou J, Olson DG, Lynd LR. A markerless gene deletion and integration system for
Thermoanaerobacter ethanolicus.
Biotechnol. Biofuels 2016,
9, 100.
[Google Scholar]
-
74.
Mougiakos I, Mohanraju P, Bosma EF, Vrouwe V, Finger Bou M, Naduthodi MIS, et al. Characterizing a thermostable Cas9 for bacterial genome editing and silencing.
Nat. Commun. 2017,
8, 1647.
[Google Scholar]
-
75.
Song Y, He S, Jopkiewicz A, Setroikromo R, van Merkerk R, Quax WJ. Development and application of CRISPR-based genetic tools in Bacillus species and Bacillus phages.
J. Appl. Microbiol. 2022,
133, 2280–2298.
[Google Scholar]
-
76.
Dai K, Fu H, Guo X, Qu C, Lan Y, Wang J. Exploiting the Type I-B CRISPR Genome Editing System in Thermoanaerobacterium aotearoense SCUT27 and Engineering the Strain for Enhanced Ethanol Production.
Appl. Environ Microbiol. 2022,
88, e0075122.
[Google Scholar]
-
77.
Zhou X, Wang X, Luo H, Wang Y, Wang Y, Tu T, et al. Exploiting heterologous and endogenous CRISPR-Cas systems for genome editing in the probiotic
Clostridium butyricum.
Biotechnol. Bioeng. 2021,
118, 2448–2459.
[Google Scholar]
-
78.
Walker JE, Lanahan AA, Zheng T, Toruno C, Lynd LR, Cameron JC, et al. Development of both type I-B and type II CRISPR/Cas genome editing systems in the cellulolytic bacterium
Clostridium thermocellum.
Metab. Eng. Commun. 2020,
10, e00116.
[Google Scholar]
-
79.
Bruder MR, Pyne ME, Moo-Young M, Chung DA, Chou CP. Extending CRISPR-Cas9 Technology from Genome Editing to Transcriptional Engineering in the Genus
Clostridium.
Appl. Environ Microbiol. 2016,
82, 6109–6119.
[Google Scholar]
-
80.
Li Q, Chen J, Minton NP, Zhang Y, Wen Z, Liu J, et al. CRISPR-based genome editing and expression control systems in
Clostridium acetobutylicum and
Clostridium beijerinckii.
Biotechnol. J. 2016,
11, 961–972.
[Google Scholar]
-
81.
Woolston BM, Emerson DF, Currie DH, Stephanopoulos G. Rediverting carbon flux in
Clostridium ljungdahlii using CRISPR interference (CRISPRi).
Metab. Eng. 2018,
48, 243–253.
[Google Scholar]
-
82.
Shen A. A Xylose-Inducible Expression System and a CRISPR Interference Plasmid for Targeted Knockdown of Gene Expression in
Clostridioides difficile.
J. Bacteriol. 2019,
201, e00711-18.
[Google Scholar]
-
83.
Ganguly J, Martin-Pascual M, van Kranenburg R. CRISPR interference (CRISPRi) as transcriptional repression tool for
Hungateiclostridium thermocellum DSM 1313.
Microb. Biotechnol. 2020,
13, 339–349.
[Google Scholar]
-
84.
Maier LK, Stachler AE, Brendel J, Stoll B, Fischer S, Haas KA, et al. The nuts and bolts of the Haloferax CRISPR-Cas system I-B.
RNA Biol. 2019,
16, 469–480.
[Google Scholar]
-
85.
Pickar-Oliver A, Black JB, Lewis MM, Mutchnick KJ, Klann TS, Gilcrest KA, et al. Targeted transcriptional modulation with type I CRISPR-Cas systems in human cells.
Nat. Biotechnol. 2019,
37, 1493–1501.
[Google Scholar]
-
86.
Young JK, Gasior SL, Jones S, Wang L, Navarro P, Vickroy B, et al. The repurposing of type I-E CRISPR-Cascade for gene activation in plants.
Commun. Biol. 2019,
2, 383.
[Google Scholar]
-
87.
Beri D, York WS, Lynd LR, Pena MJ, Herring CD. Development of a thermophilic coculture for corn fiber conversion to ethanol.
Nat. Commun. 2020,
11, 1937.
[Google Scholar]
-
88.
Zhang J, Hu L, Zhang H, He ZG. Cyclic di-GMP triggers the hypoxic adaptation of
Mycobacterium bovis through a metabolic switching regulator ArgR.
Environ. Microbiol. 2022,
24, 4382–4400.
[Google Scholar]
-
89.
Charlier D, Bervoets I. Regulation of arginine biosynthesis, catabolism and transport in
Escherichia coli.
Amino Acids 2019,
51, 1103–1127.
[Google Scholar]
-
90.
Qu C, Chen L, Li Y, Fu H, Wang J. The redox-sensing transcriptional repressor Rex is important for regulating the products distribution in
Thermoanaerobacterium aotearoense SCUT27.
Appl. Microbiol. Biotechnol. 2020,
104, 5605–5617.
[Google Scholar]
-
91.
Jiang Y, Liu J, Dong W, Zhang W, Fang Y, Ma J, et al. The Draft Genome Sequence of Thermophilic
Thermoanaerobacterium thermosaccharolyticum M5 Capable of Directly Producing Butanol from Hemicellulose.
Curr. Microbiol. 2018,
75, 620–623.
[Google Scholar]
-
92.
Yang X, Zhu M, Huang X, Lin CS, Wang J, Li S. Valorisation of mixed bakery waste in non-sterilized fermentation for L-lactic acid production by an evolved
Thermoanaerobacterium sp. strain.
Bioresour. Technol. 2015,
198, 47–54.
[Google Scholar]
-
93.
Iodice P, Senatore A, Langella G, Amoresano A. Advantages of ethanol-gasoline blends as fuel substitute for last generation Si engines.
Environ. Progress Sustain. Energy 2017,
36, 1173–1179.
[Google Scholar]
-
94.
Toor M, Kumar SS, Malyan SK, Bishnoi NR, Mathimani T, Rajendran K, Pugazhendhi A. An overview on bioethanol production from lignocellulosic feedstocks.
Chemosphere 2020,
242, 125080.
[Google Scholar]
-
95.
Jacobus Ana P, Gross J, Evans John H, Ceccato-Antonini Sandra R, Gombert Andreas K.
Saccharomyces cerevisiae strains used industrially for bioethanol production.
Essays Biochem. 2021,
65, 147–161.
[Google Scholar]
-
96.
Zhang L, Tang Y, Guo ZP, Ding ZY, Shi GY. Improving the ethanol yield by reducing glycerol formation using cofactor regulation in
Saccharomyces cerevisiae.
Biotechnol. Lett. 2011,
33, 1375–1380.
[Google Scholar]
-
97.
Yan X, Wang X, Yang Y, Wang Z, Zhang H, Li Y, et al. Cysteine supplementation enhanced inhibitor tolerance of
Zymomonas mobilis for economic lignocellulosic bioethanol production.
Bioresour. Technol. 2022,
349, 126878.
[Google Scholar]
-
98.
Todhanakasem T, Wu B, Simeon S. Perspectives and new directions for bioprocess optimization using
Zymomonas mobilis in the ethanol production.
World J. Microbiol. Biotechnol. 2020,
36, 112.
[Google Scholar]
-
99.
Koppolu V, Vasigala VK. Role of
Escherichia coli in Biofuel Production.
Microbiol. Insights 2016,
9, 29–35.
[Google Scholar]
-
100.
Cai Y, Lai C, Li S, Liang Z, Zhu M, Liang S, et al. Disruption of lactate dehydrogenase through homologous recombination to improve bioethanol production in
Thermoanaerobacterium aotearoense.
Enzyme Microb. Technol. 2011,
48, 155–161.
[Google Scholar]
-
101.
Fu HX, Luo S, Dai KQ, Qu CY, Wang JF. Engineering
Thermoanaerobacterium aotearoense SCUT27/Delta ldh with pyruvate formate lyase-activating protein (PflA) knockout for enhanced ethanol tolerance and production.
Process Biochem. 2021,
106, 184–190.
[Google Scholar]
-
102.
Olson DG, Sparling R, Lynd LR. Ethanol production by engineered thermophiles.
Curr. Opin. Biotechnol. 2015,
33, 130–141.
[Google Scholar]
-
103.
Cui J, Olson DG, Lynd LR. Characterization of the
Clostridium thermocellum AdhE, NfnAB, ferredoxin and Pfor proteins for their ability to support high titer ethanol production in
Thermoanaerobacterium saccharolyticum.
Metab. Eng. 2019,
51, 32–42.
[Google Scholar]
-
104.
Hitschler L, Kuntz M, Langschied F, Basen M.
Thermoanaerobacter species differ in their potential to reduce organic acids to their corresponding alcohols.
Appl. Microbiol. Biotechnol. 2018,
102, 8465–8476.
[Google Scholar]
-
105.
Scully SM, Orlygsson J. Biotransformation of Carboxylic Acids to Alcohols: Characterization of
Thermoanaerobacter Strain AK152 and 1-Propanol Production via Propionate Reduction.
Microorganisms 2020,
8, 945.
[Google Scholar]
-
106.
Borchers A, Pieler T. Programming pluripotent precursor cells derived from Xenopus embryos to generate specific tissues and organs.
Genes 2010,
1, 413–426.
[Google Scholar]
-
107.
Lo J, Zheng T, Hon S, Olson DG, Lynd LR. The bifunctional alcohol and aldehyde dehydrogenase gene, adhE, is necessary for ethanol production in
Clostridium thermocellum and
Thermoanaerobacterium saccharolyticum.
J. Bacteriol. 2015,
197, 1386–1393.
[Google Scholar]
-
108.
Zheng T, Olson DG, Tian L, Bomble YJ, Himmel ME, Lo J, et al. Cofactor Specificity of the Bifunctional Alcohol and Aldehyde Dehydrogenase (AdhE) in Wild-Type and Mutant
Clostridium thermocellum and
Thermoanaerobacterium saccharolyticum.
J. Bacteriol. 2015,
197, 2610–2619.
[Google Scholar]
-
109.
Desai SG, Guerinot ML, Lynd LR. Cloning of L-lactate dehydrogenase and elimination of lactic acid production via gene knockout in
Thermoanaerobacterium saccharolyticum JW/SL-YS485.
Appl. Microbiol. Biotechnol. 2004,
65, 600–605.
[Google Scholar]
-
110.
Shaw AJ, Hogsett DA, Lynd LR. Identification of the [FeFe]-hydrogenase responsible for hydrogen generation in
Thermoanaerobacterium saccharolyticum and demonstration of increased ethanol yield via hydrogenase knockout.
J. Bacteriol. 2009,
191, 6457–6464.
[Google Scholar]
-
111.
Eminoğlu A, Murphy SJ-L, Maloney M, Lanahan A, Giannone RJ, Hettich RL, et al. Deletion of the hfsB gene increases ethanol production in
Thermoanaerobacterium saccharolyticum and several other thermophilic anaerobic bacteria.
Biotechnol. Biofuels 2017,
10, 282.
[Google Scholar]
-
112.
Fu H, Yang X, Qu C, Li Y, Wang J. Enhanced ethanol production from lignocellulosic hydrolysates by inhibiting the hydrogen synthesis in
Thermoanaerobacterium aotearoense SCUT27(Δldh).
J. Chem. Technol. Biotechnol. 2019,
94, 3305–3314.
[Google Scholar]
-
113.
Tian L, Lo J, Shao X, Zheng T, Olson DG, Lynd LR. Ferredoxin:NAD+ Oxidoreductase of
Thermoanaerobacterium saccharolyticum and Its Role in Ethanol Formation.
Appl. Environ Microbiol. 2016,
82, 7134–7141.
[Google Scholar]
-
114.
Lo J, Zheng T, Olson DG, Ruppertsberger N, Tripathi SA, Tian L, et al. Deletion of nfnAB in
Thermoanaerobacterium saccharolyticum and Its Effect on Metabolism.
J. Bacteriol. 2015,
197, 2920–2929.
[Google Scholar]
-
115.
Beri D, Olson DG, Holwerda EK, Lynd LR. Nicotinamide cofactor ratios in engineered strains of
Clostridium thermocellum and
Thermoanaerobacterium saccharolyticum.
FEMS Microbiol. Lett. 2016,
363, fnw091.
[Google Scholar]
-
116.
Lovitt RW, Shen GJ, Zeikus JG. Ethanol production by thermophilic bacteria: Biochemical basis for ethanol and hydrogen tolerance in
Clostridium thermohydrosulfuricum.
J. Bacteriol. 1988,
170, 2809–2815.
[Google Scholar]
-
117.
Shaw AJ, Miller BB, Rogers SR, Kenealy WR, Meola A, Bhandiwad A, et al. Anaerobic detoxification of acetic acid in a thermophilic ethanologen.
Biotechnol. Biofuels 2015,
8, 75.
[Google Scholar]
-
118.
Yanase H, Miyawaki H, Sakurai M, Kawakami A, Matsumoto M, Haga K, et al. Ethanol production from wood hydrolysate using genetically engineered
Zymomonas mobilis.
Appl. Microbiol. Biotechnol. 2012,
94, 1667–1678.
[Google Scholar]
-
119.
Haque S, Singh R, Pal DB, Faidah H, Ashgar SS, Areeshi MY, et al. Thermophilic biohydrogen production strategy using agro industrial wastes: Current update, challenges, and sustainable solutions.
Chemosphere 2022,
307, 136120.
[Google Scholar]
-
120.
Ubando AT, Chen WH, Hurt DA, Conversion A, Rajendran S, Lin SL. Biohydrogen in a circular bioeconomy: A critical review.
Bioresour. Technol. 2022,
366, 128168.
[Google Scholar]
-
121.
Soboh B, Linder D, Hedderich R. A multisubunit membrane-bound [NiFe] hydrogenase and an NADH-dependent Fe-only hydrogenase in the fermenting bacterium
Thermoanaerobacter tengcongensis.
Microbiology 2004,
150, 2451–2463.
[Google Scholar]
-
122.
Carere CR, Rydzak T, Verbeke TJ, Cicek N, Levin DB, Sparling R. Linking genome content to biofuel production yields: a meta-analysis of major catabolic pathways among select H2 and ethanol-producing bacteria.
BMC Microbiol. 2012,
12, 295.
[Google Scholar]
-
123.
Li P, Zhu M. A consolidated bio-processing of ethanol from cassava pulp accompanied by hydrogen production.
Bioresour. Technol. 2011,
102, 10471–10479.
[Google Scholar]
-
124.
Cheng J, Zhu M. A novel anaerobic co-culture system for bio-hydrogen production from sugarcane bagasse.
Bioresour. Technol. 2013,
144, 623–631.
[Google Scholar]
-
125.
Jiang Y, Guo D, Lu J, Dürre P, Dong W, Yan W, et al. Consolidated bioprocessing of butanol production from xylan by a thermophilic and butanologenic
Thermoanaerobacterium sp. M5.
Biotechnol. Biofuels 2018,
11, 89.
[Google Scholar]
-
126.
Bhandiwad A, Shaw AJ, Guss A, Guseva A, Bahl H, Lynd LR. Metabolic engineering of
Thermoanaerobacterium saccharolyticum for n-butanol production.
Metab. Eng. 2014,
21, 17–25.
[Google Scholar]
-
127.
Liu Y, Yu P, Song X, Qu Y. Hydrogen production from cellulose by co-culture of
Clostridium thermocellum JN4 and
Thermoanaerobacterium thermosaccharolyticum GD17.
Int. J. Hydrog. Energy 2008,
33, 2927–2933.
[Google Scholar]
-
128.
Kremer F, Blank LM, Jones PR, Akhtar MK. A Comparison of the Microbial Production and Combustion Characteristics of Three Alcohol Biofuels: Ethanol, 1-Butanol, and 1-Octanol.
Front. Bioeng. Biotechnol. 2015,
3, 112.
[Google Scholar]
-
129.
Jiang Y, Lv Y, Wu R, Lu J, Dong W, Zhou J, et al. Consolidated bioprocessing performance of a two-species microbial consortium for butanol production from lignocellulosic biomass.
Biotechnol. Bioeng. 2020,
117, 2985–2995.
[Google Scholar]
-
130.
Morvan C, Folgosa F, Kint N, Teixeira M, Martin-Verstraete I. Responses of Clostridia to oxygen: From detoxification to adaptive strategies.
Environ. Microbiol. 2021,
23, 4112–4125.
[Google Scholar]
-
131.
Qu C, Dai K, Fu H, Wang J. Enhanced ethanol production from lignocellulosic hydrolysates by
Thermoanaerobacterium aotearoense SCUT27/ΔargR1864 with improved lignocellulose-derived inhibitors tolerance.
Renew. Energy 2021,
173, 652–661.
[Google Scholar]
-
132.
Lynd LR, Baskaran S, Casten S. Salt accumulation resulting from base added for pH control, and not ethanol, limits growth of
Thermoanaerobacterium thermosaccharolyticum HG-8 at elevated feed xylose concentrations in continuous culture.
Biotechnol. Progress 2001,
17, 118–125.
[Google Scholar]
-
133.
Atalah J, Caceres-Moreno P, Espina G, Blamey JM. Thermophiles and the applications of their enzymes as new biocatalysts.
Bioresour. Technol. 2019,
280, 478–488.
[Google Scholar]
-
134.
Wang F, Wang M, Zhao Q, Niu K, Liu S, He D, et al. Exploring the Relationship between
Clostridium thermocellum JN4 and
Thermoanaerobacterium thermosaccharolyticum GD17.
Front. Microbiol. 2019,
10, 2035.
[Google Scholar]
-
135.
Cheng J, Yu Y, Zhu M. Enhanced biodegradation of sugarcane bagasse by
Clostridium thermocellum with surfactant addition.
Green Chem. 2014,
16, 2689–2695.
[Google Scholar]
-
136.
Lynd LR, Guss AM, Himmel ME, Beri D, Herring C, Holwerda EK, et al. Advances in Consolidated Bioprocessing Using Clostridium thermocellum and Thermoanaerobacter saccharolyticum. In Industrial Biotechnology; Wiley-VCH: Weinheim, Germany, 2017; pp. 365–394.