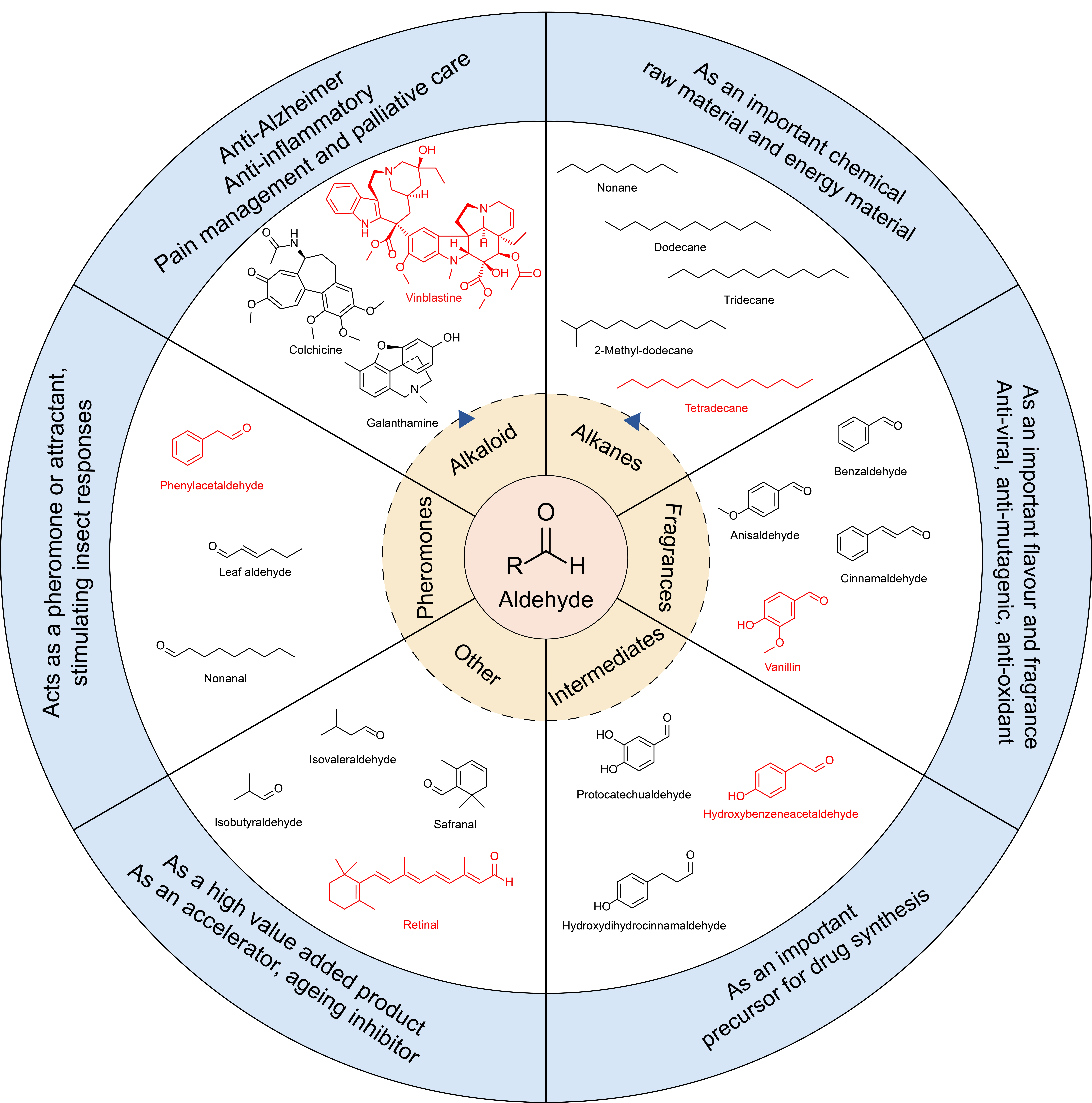
In light of recent strides achieved in the fields of genome engineering and synthetic biology, the paradigm of
de novo aldehyde biosynthesis in microorganisms emerges as a compelling alternative avenue when juxtaposed against traditional modalities, such as the customary physical (e.g., aldehyde extraction from botanical sources [
21]) or chemical routes. Nonetheless, most microorganisms are unable to accumulate aldehydes naturally because aldehydes are readily converted to the corresponding alcohols or acids by the endogenous NADH-dependent alcohol dehydrogenase (ADH) family, NADPH-dependent aldo-keto reductase (AKR) family, or aldehyde dehydrogenase (ALDH) family. Therefore, the pivotal interventions necessitate the targeted elimination of ADHs, AKRs, and ALDHs, simultaneous with the orchestrated implementation of metabolic pathways dedicated to channeling the carbon fluxes in the direction of aldehyde biosynthesis. To facilitate a structured discussion, this review offers a preliminary classification of biogenic aldehydes into two primary groups based on molecular structure: aliphatic and aromatic aldehydes. A comprehensive compilation of representative examples of
de novo aldehyde biosynthesis, as detailed in the subsequent sections, is concisely summarized in for ease of reference.
. Microbial synthesis of aliphatic aldehydes and their derivatives.
Aliphatic aldehydes, produced through fermentation or fatty acid synthesis, have been strategically employed to synthesize alcohols [
37,
38] as well as alkanes [
22,
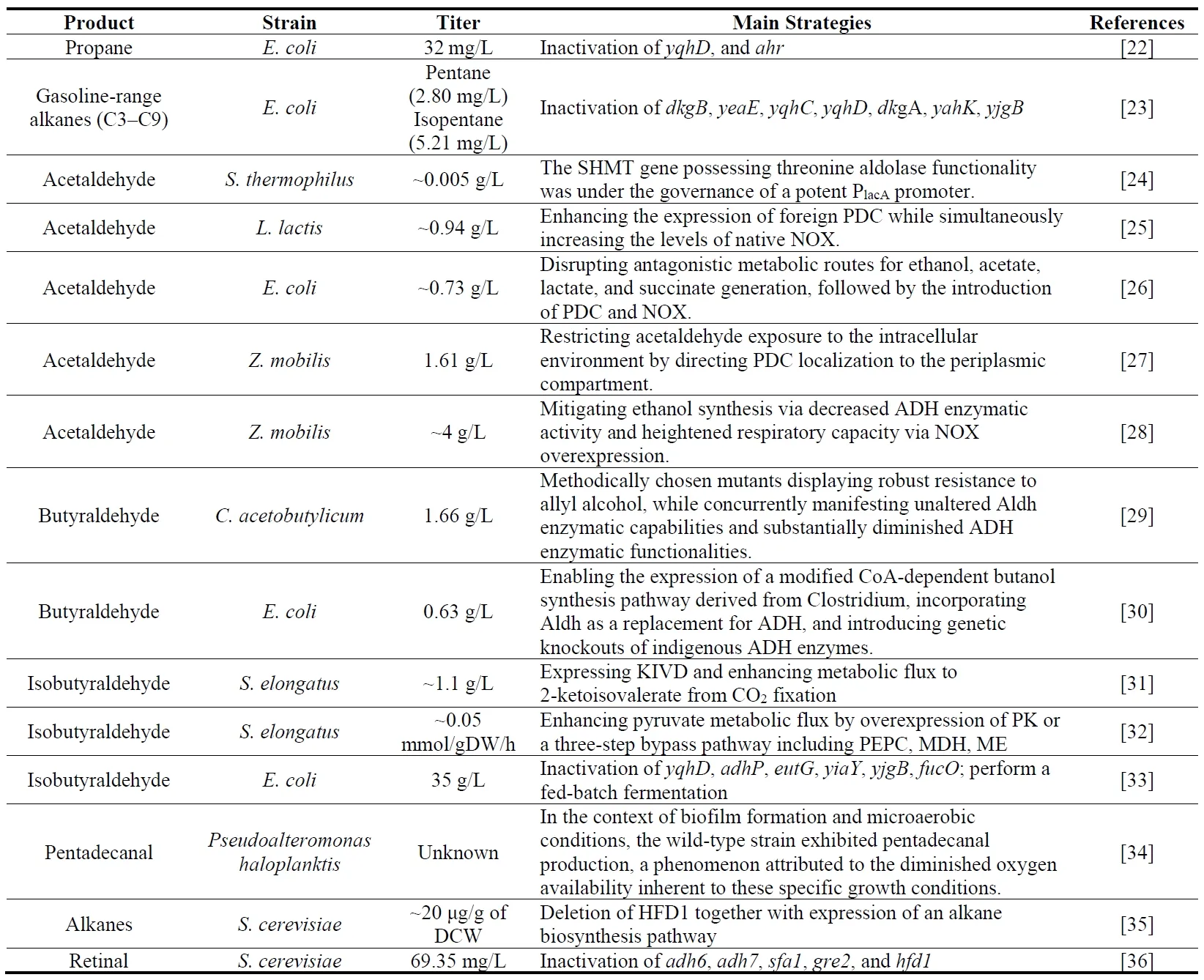
23], thereby catering to their utilities as both fuels and foundational chemical compounds. As aliphatic aldehydes are classified based on carbon chain length, they are categorized into short-chain (C ≤ 5), medium-chain (C6–C12), and long-chain (C > 12) fatty aldehydes. Extensive research has demonstrated the capacity of diverse engineered microorganisms, such as
Escherichia coli,
Pseudomonas aeruginosa, and
Clostridium acetobutylicum, to synthesize these aldehydes, as summarized in . Notably, the microbial synthesis of short-chain fatty aldehydes has reached a comparatively advanced stage, with numerous studies reporting gram-scale product yields. Acetaldehyde, in particular, functions as an essential food additive and flavoring agent, and acts as a precursor for producing various commercial chemicals, including acetic anhydride, acetic acid, and butadiene.In 2002, Chaves et al. [
24] reported that overexpressing the
glyA gene (encoding serine hydroxymethyltransferase) showed an increase in threonine aldolase activity and acetaldehyde (~0.005 g/L) and folic acid formation when the engineered
Streptococcus thermophilus were grown in LM17 medium. In 2005, efficient glucose to acetaldehyde (~0.94 g/L) conversion was achieved in
Lactococcus lactis by nisibiotic-controlled overexpression of
Zymomonas mobilis Pdc and
L. lactis Nox [
25]. In 2018, Balagurunathan et al. [
26] introduced Pdc from
Z. mobilis and NADH oxidase (Nox) from
L. lactis into engineered
E. coli. They knocked out genes related to competitive metabolic pathways (
ΔadhE ΔldhA ΔfrdC ΔackA-pta), resulting in the production of acetaldehyde with a titer of 0.73 g/L from glucose. In 2019, Balodite et al. [
27] overexpressed pyruvate decarboxylase (Pdc) from the cytosol into the periplasm, culminating in the synthesis of acetaldehyde at 1.66 g/L with glucose as the carbon source. Kallenieks et al. [
28] used glucose as a carbon source to modify the motile fermentative bacterium, producing 4 g/L acetaldehyde by reducing the activity of ADHs and enhancing respiratory capacity (overexpression of respiratory NADH dehydrogenase (NDH) or mutations in other components metabolized by respiration).
For the synthesis of butyraldehyde, in 1987, Rogers et al. [
29] discovered that a spontaneous mutant of
C. acetobutylicum NRRL B643 could produce 1.66 g/L butyraldehyde from glucose. In 2017, Ku et al. [
30] overexpressed Coenzyme A-acylating ALDH from
Clostridium beijerinckii in
E. coli while knocking out the endogenous ethanol dehydrogenases YjgB, FucO, EutG, YbbO, AdhP, GldA, YahK, and YghA to synthesize 0.63 g/L butyraldehyde from glucose. Isobutyraldehyde, an isomer of butyraldehyde, was considered a promising diesel or jet fuel component. In 2009, Atsumi et al. [
31] introduced the isobutyraldehyde synthesis pathway (ketoacid decarboxylase gene
kivd from
L. lactis,
alsS from
Bacillus subtilis,
ilvC and
ilvD from
E. coli) into
Synechococcus elongatus PCC7942. They overexpressed ribulose 1,5-diphosphocarboxylase/oxygenase (Rubisco) to improve productivity. The modified engineering strain can directly produce 1.1 g/L isobutyraldehyde from CO
2. Jazmin et al. [
32] adjusted the carbon flux of the pyruvate kinase (PK) reaction step and its bypass pathway based on the study of engineering strains and improved the ability of engineering strains to produce isobutyraldehyde using CO
2 in small-scale experiments (0.05 g mmol/gDW/h). In
E. coli, Rodriguez and Atsumi [
33] identified and deleted 8 natural isobutyraldehyde reductases in the strain JCL260 that produces isobutanol. Combined with an in-situ product removal (ISPR) system, they achieved isobutyraldehyde production capacity of 35 g/L, which is the highest reported yield to date. However, due to incomplete knockout of the intracellularly related reductase, the strain still produced quite a lot of isobutanol after fermentation. In 2014, Rodriguez and Atsumi [
39] reported that the simultaneous inactivation of 13 genes (
adhE,
yqhD,
adhP,
eutG,
yiaY,
ahr,
betA,
fucO,
yahK,
dkgA,
gldA,
ybbO, and
yghA) in
E. coli strain AL1728 led to a reduction of 90–99% in the strain’s endogenous aldehyde reductase activity for a variety of aliphatic aldehyde (C2 to C12).
In comparison, fewer studies have focused on the production of medium-chain and long-chain fatty aldehydes using microorganisms. Santiago-Gómez et al. [
40] achieved a yield of 600 mg/L hexanal by expressing hydrogen peroxide lyase (HPL) in green pepper fruit using
Yarrowia lipolytica. Šalić et al. [
41] employed NAD
+ dependent ADH from
Saccharomyces cerevisiae in a microreactor system to facilitate the oxidation of hexanol to hexanal. Akhtar et al. [
37] demonstrated that carboxylic acid reductase (CAR) from
Mycobacterium marinum is capable of converting various fatty acids (C6–C18) into their corresponding aldehydes. In 2020, Zhu et al. [
42] realized an efficient synthesis of medium-chain fatty acids in
S. cerevisiae employing multidimensional engineering techniques. Further introduction of CAR could potentially facilitate efficient synthesis of medium-chain fatty aldehydes.
2.2. Microbial Synthesis of Aromatic Aldehydes
Aromatic aldehydes serve as essential precursors in the synthesis of fragrances, pharmaceuticals, agrochemicals, and polymers [
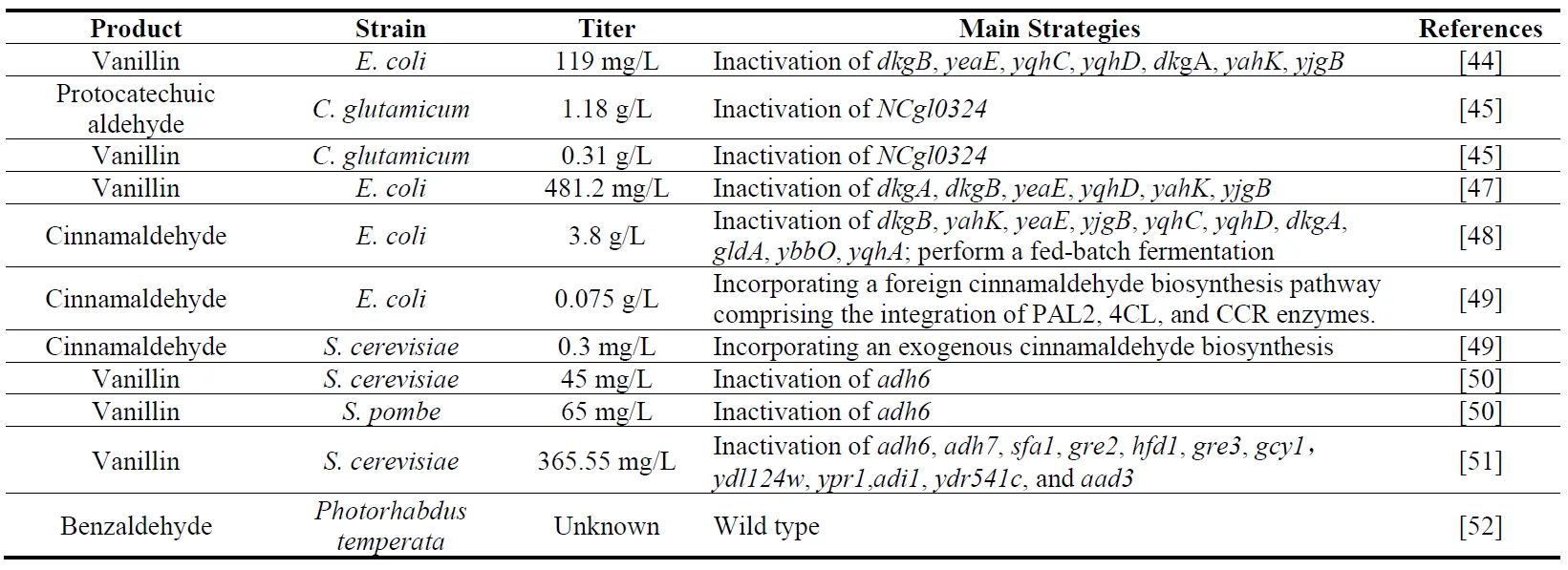
43]. Owing to their high-value applications and substantial market demand, there has been a significant surge in commercial interest for microbial aldehyde synthesis in recent years. A compilation of recent advancements in the microbial accumulation of aromatic aldehydes is presented in . Among these, vanillin and benzaldehyde are particularly prominent, holding substantial market share in the flavor and fragrance industry. In 2014, Kunjapur et al. [
44] demonstrated that knockout of ADHs (YqhD, YahK, and YjgB) and AKRs (DkgB, YeaE, DkgA) in
E. coli (RARE strain) had a significant effect on improving the microbial accumulation of aldehydes, and achieved the accumulation of benzaldehyde and vanillin under specific conditions. After introducing the vanillin synthesis pathway, 119 ± 3 mg/L vanillin can be synthesized from glucose (more than a 55-fold increase over the wild-type strain). In 2022, Kim et al. [
45] found that a single knockout of
NCgl0324 in
Corynebacterium glutamicum substantially enhanced the production of protocatechualdehyde (1.18 g/L) and vanillin (0.31 g/L). In 2023, Butler et al. [
46] executed an extensive genetic manipulation study, systematically inactivating a suite of oxidoreductases (AldB, PuuC, BetB, PatD, FeaB, GabD, AldA, Sad, AstD, and PutA) within RARE strains. This strategic intervention led to the creation of ROAR strains, which, employed as whole-cell biocatalysts, exhibited a markedly improved capacity for accumulating eight aromatic aldehydes: piperonaldehyde, naphthaldehyde, anisaldehyde,
m-hydroxybenzaldehyde, butyraldehyde, vanillin, benzaldehyde, and furfural. Particularly noteworthy, the retention rates of these aldehydes, assessed 4 hours following their introduction, revealed that six maintained levels above 50%. In 2024, Wu et al. [
47] synthesized 481.2 mg/L vanillin in
E. coli by optimizing the protocatechuic acid pathway, deleting three endogenous AKRs (encoded by DkgA, DkgB and YeaE) and three ADHs (YqhD, YahK and YjgB). Cinnamaldehyde, derived from the oil of cinnamon bark, is extensively utilized as a flavoring agent in a broad range of food and beverage products due to its substantial impact on enhancing taste and aroma. In 2023, Bang et al. [
48] knocked out oxidoreductases (DkgA, DkgB, YahK, YeaE, YbbO, YjgB, YqhD, YqhC, GldA, and YqhA) in
E. coli to achieve a higher level accumulation of cinnamaldehyde (3.8 g/L). However,
E. coli and
C. glutamicum are unsuitable for food-related fermentation applications, which severely limits their utility and potential uses.
. Microbial synthesis of simple aromatic aldehydes.
Compared to
E. coli, the aldehyde accumulation platform in
S. cerevisiae has been relatively slow to be studied in recent years. presents an exemplary
de novo vanillin biosynthesis from glucose via 3-dehydroshikimate in engineered yeasts,
S. cerevisiae and
Schizosaccharomyces pombe, facilitated by the introduction of four pivotal enzymes: 3-dehydroshikimate dehydratase, CAR, phosphopantetheinyl transferase, and catechol O-methyltransferase (COMT) [
50]. However, endogenous enzyme activity posed a limitation on vanillin accumulation. Inactivation of ADH6 among 29 candidate ADHs, aryl-ADHs, and related aldehyde reductases (ALDRs) led to a 50% reduction in vanillin conversion to vanillyl alcohol, overcoming this obstacle [
50]. More recently, our group deleted a total of 12 genes comprising Adh6,
Adh7,
Sfa1,
Gre2,
Hfd1,
Gre3,
Gcy1,
Ydl124w,
Ypr1,
Ari1,
Ydr541c, and
Aad3 in
S. cerevisiae, and the resulting minimal aromatic aldehyde reduction (MARE) yeast platform allowed improved accumulation of vanillin [
51]. This collective effort of engineering
S. cerevisiae produced an impressive 365.55 mg/L vanillin from glucose. This concerted genetic intervention yielded marked reductions in alcohol formation, concomitantly fostering the augmentation of vanillin accumulation, thereby reflecting a salient exemplification of the approach.
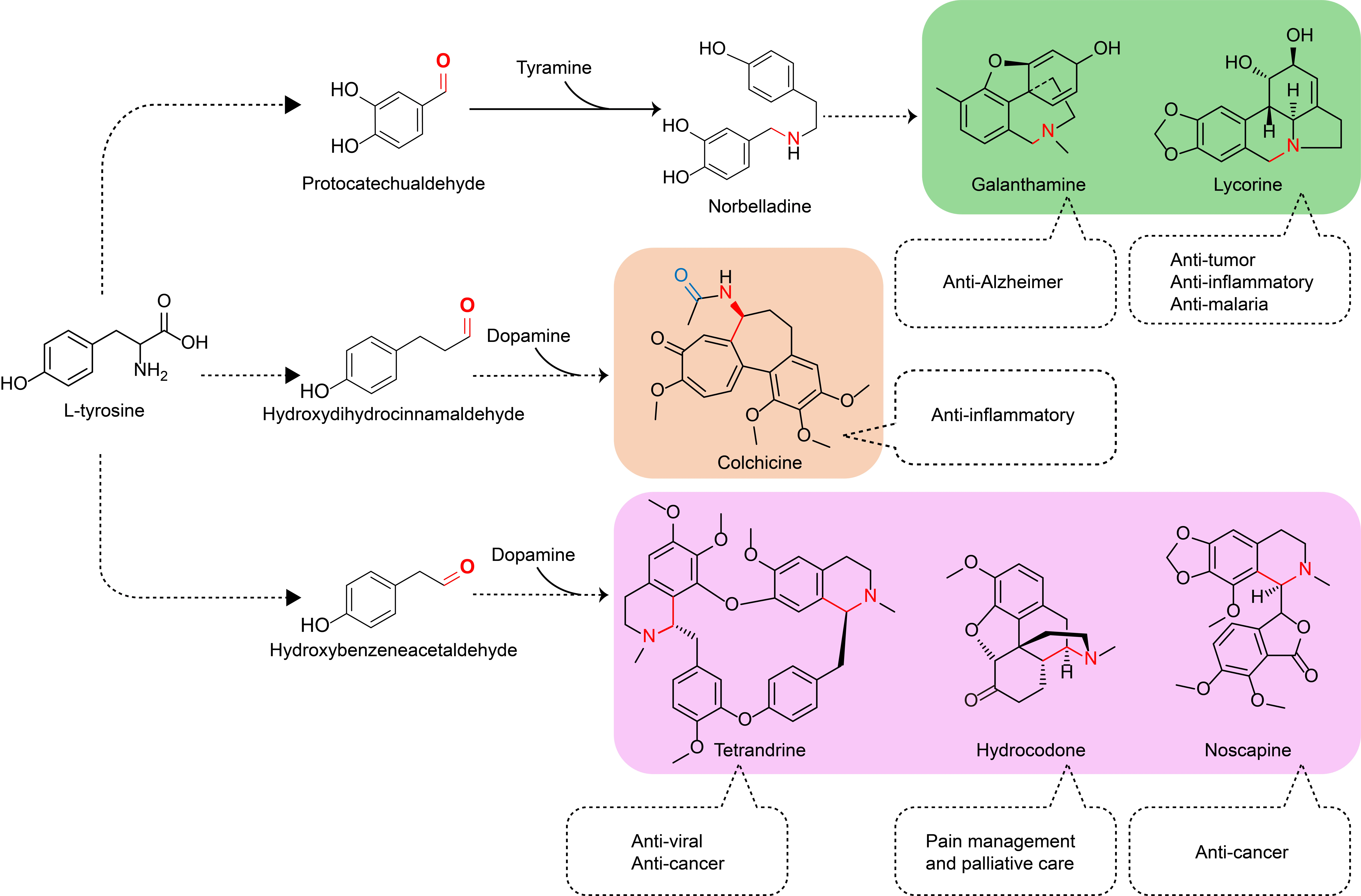
2.3. Microbial Synthesis of Aromatic Aldehyde Derived Complex Alkaloids
The alkaloids are the largest class of natural products and include several of the most important human medicines [
53]. Hence, an increasing number of studies are embracing biomass-derived aromatic aldehydes as feedstocks to expedite the biosynthesis of alkaloids (). (
S)-Reticuline is a crucial intermediate for the production of alkaloids, which are applied in the manufacture of pharmaceuticals, green biological pesticides, and food additives [
54]. The biosynthetic pathway of (
S)-reticuline in plants starts with dopamine and 4-hydroxyphenylacetaldehyde (4-HPAA). 4-HPAA condenses with dopamine to form (
S)-reticuline through enzyme-mediated Pictet-Spengler reaction, followed by 6OMT-catalyzed methylation of a 6-position hydroxyl group to form (
S)-reticuline [
55]. In 2020, Martin et al. [
1] continued to implement >20 successive strain modifications to the yeast shikimate, Ehrlich, and L-tyrosine metabolic pathway based on the previous studies. The refinement of the shikimate pathway involved the amplification of branch acid synthase, prenylate dehydrogenase, and phenylpyruvate decarboxylase within the manganic acid pathway. Furthermore, the targeted elimination of genes responsible for seven related reductases and oxidases in the Ehrlich pathway was performed, effectively impeding the reduction and oxidative functions of 4-HPAA. Through feedback regulation of enzyme inhibition and optimization of the fermentation process, the yield of
de novo synthesized (
S)-reticuline by yeast increased 57,000-fold to 4.6 g/L [
1]. The research results indicate that microbial synthesis of (
S)-reticuline has reached the industrial production level of BIAs, highlighting the great potential of microbial systems for producing plant secondary metabolites. . A range of alkaloids, including berberine, scopolamine, tropine, vinblastine, and catharanthine, has been successfully demonstrated to be synthesized
de novo in
S. cerevisiae and
Pichia pastoris (). Considering that yeast cells are more friendly to host diverse plant-derived metabolic pathways, it is like to explore further the MARE yeast platform [
51] to synthesize other more complex natural alkaloids such as galanthamine ().
. Schematic representation of the microbial system used to convert aldehydes into value-added alkaloids.
. Microbial synthesis of plant derived alkaloids.
As depicted in , a diverse array of aldehydes is utilized in the spice, pharmaceutical, and food industries. Given their potential for direct or indirect human exposure, the designation of “naturally derived” through microbial biosynthesis could expand the application spectrum of these compounds. To date, numerous aldehydes have been successfully produced via engineered microbial platforms, leveraging the capabilities of microbial chassis. With the ongoing refinement of microbial metabolic engineering, strain development, and evolutionary techniques, the
de novo biosynthesis of aldehydes that are rare or possess intricate structures has become feasible.
Retinal and its derivatives have been used in food, cosmetics, pharmaceuticals, nutritional supplements, and animal feed additives [
62,
63,
64]. In 2022, we first reported the microbial production of retinal as the sole product [
36]. Prior to this, there were reports of microbial synthesis of retinal, which resulted in a mixture of retinoids (including retinal, retinol, retinyl acetate, etc.) [
65,
66,
67]. In our study [
36], the co-inactivation of four ADHs (Adh6, Adh7, Sfa1, Gre2) and one ALDH (Hfd1) in
S. cerevisiae enhanced the yeast’s capacity to accumulate retinal. By integrating
crtE/YB/I genes from
Xanthophyllomyces dendrorhous and β-carotene 15,15′-monooxygenase (BCMO) from the marine bacterium 66A03 into the yeast genome, we optimized β-carotene synthesis, yielding 69.13 ± 5.82 mg/L retinal with >99% purity in shake-flask cultures.
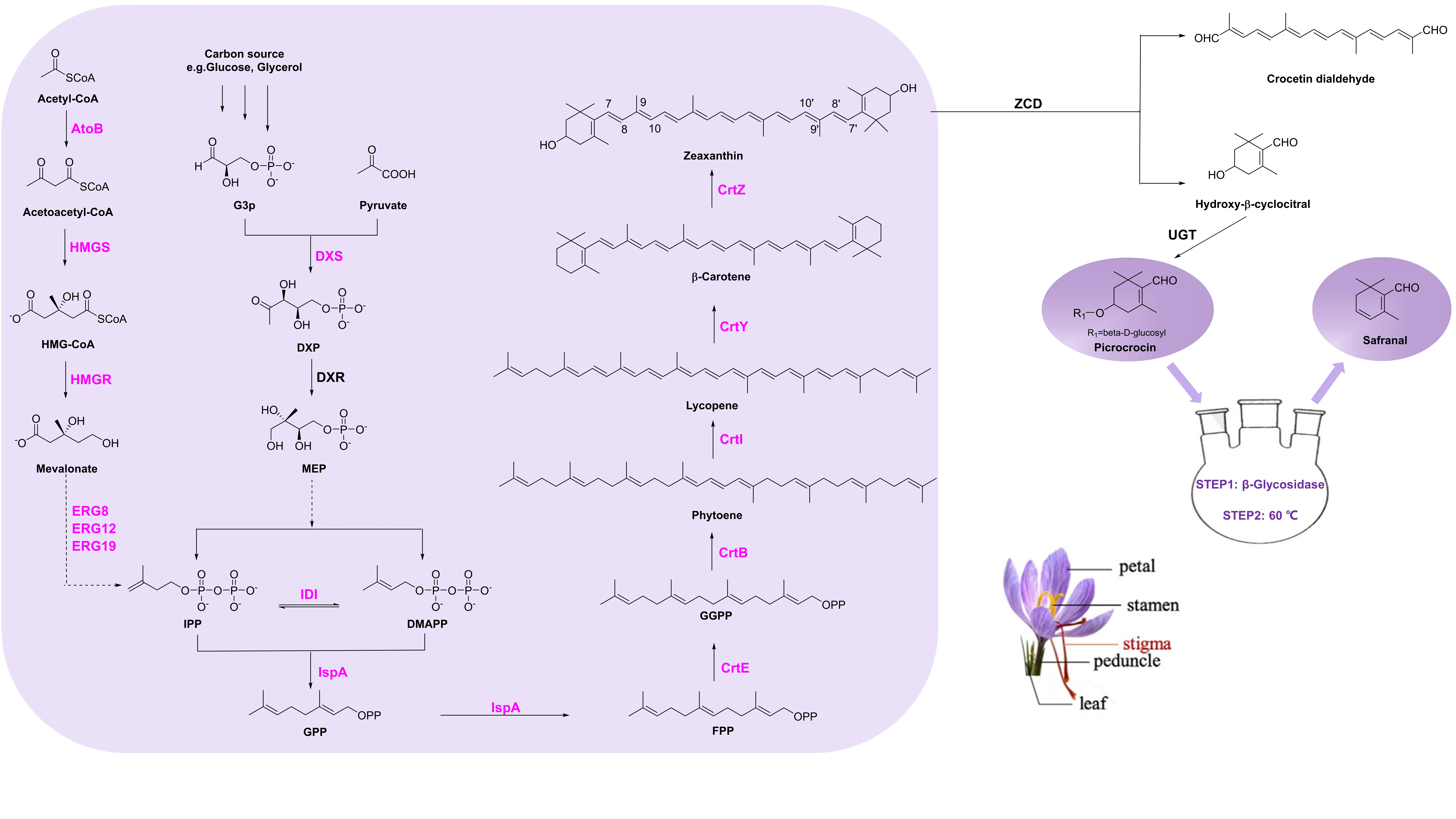
Plants constitute a vital reservoir for natural aldehyde compounds. A case in point is safranal, the principal volatile component responsible for the distinct aroma of saffron. Beyond its characteristic spicy fragrance, safranal has also been recognized for its diverse pharmacological activities, including potential therapeutic benefits for anxiety, depression, and other neurological disorders [
68]. Here, we predicted the possible synthesis modes of saffron aldehyde in microbial platforms (). Zeaxanthin is the key compound in the synthesis pathways of safranal. In 2016, Shen et al. [
69] introduced the dynamically regulated mevalonate (MVA) pathway from
S. cerevisiae into
E. coli, achieving a zeaxanthin titer of 722.46 mg/L via batch fermentation with feed supplementation. In 2023, Zhang et al. [
70] performed metabolic engineering on
Y. lipolytica, enabling the engineered strain to produce up to 775.3 mg/L of zeaxanthin in YPD shake flasks. The production of zeaxanthin, an intermediate in the safrole biosynthesis pathway, has been validated and optimized across diverse microbial hosts. Furthermore, prior studies have identified carotenoid cleavage dioxygenase 2 (
CsCCD2) [
71] for its role in catalyzing the conversion of zeaxanthin to hydroxy-β-cyclocitral and
CsUGT709G1 [
72] for catalyzing the formation of picrocrocin from hydroxy-β-cyclocitral in
Crocus sativus. Integrating
CsCCD2 and
CsUGT709G1 into the zeaxanthin-producing microbial platform could facilitate the microbial synthesis of picrocrocin. Picrocrocin, the precursor of the volatile safranal, could be extracted from saffron stigmas and biotransformed to 4-hydroxysafranal with complete conversion (>99%) using β-glucosidase from
Alicyclobacillus acidophilus, followed by a thermic treatment at 60 ℃. A one-pot 2-step chemo-enzymatic transformation is established to produce a marketable natural safranal [
73]. To date, there are few higher plant genes to construct microbial cell factories for safranal accumulation. Thus, the introduction of higher plant genes into microorganisms to increase safranal preparation will be a significant trial. The results provide important insights into the engineering of safranal producer. Reconstitution of the safranal biosynthetic pathway in
E. coli RARE and the yeast MARE platform might eventually give a sustainable production of safranal from economical fermentation processes.
. Proposed synthetic pathway of plant natural product of safranal. HMG-CoA, 3-hydroxy-3-methylglutaryl-CoA; DXP, 1-deoxyxylulose-5-phosphate; MEP, 2-C-methyl-D-erythritol 4-phosphate; IPP, isopentenyl pyrophosphate; DMAPP, dimethylallyl diphosphate; FPP, farnesyl pyrophosphate; GGPP, geranylgeranyl pyrophosphate; AtoB, acetoacetyl-CoA thiolase; HMGS, HMG-CoA synthase; HMGR, HMG-CoA reductase; DXS, 1-deoxy-D-xylulose 5-phosphate synthase; DXR, DXP reductoismerase; IDI, IPP isomerase; IspA, GPP/FPP synthase; CrtE, GGPP synthase; CrtB, phytoene synthase; CrtI, phytoene desaturase; CrtY, lycopene cyclase; CrtZ, β-carotene hydroxylase; ZCD, zeaxanthin cleavage dioxygenase; UGT, UDPG-glucosyltransferase.
Improving aldehyde production in microbial cell factories typically involves modulating carbon flux in metabolic pathways, inactivating endogenous enzymes that rapidly convert aldehydes into alcohols or acids, and mitigating the toxicity of aldehydes to cells. Regulating carbon flux through metabolic pathways is a standard strategy for enhancing microbial cell factories, primarily by steering metabolic carbon flux towards product synthesis pathways to augment yield. In this section, we will not delve into carbon flux regulation strategies but rather concentrate on the prevalent issues of endogenous transformation and cytotoxicity associated with aldehydes as target products.
3.1. Minimizing Endogenous Consumption of Aldehydes
Because of the presence of endogenous ADHs, AKRs, ALDRs, and ALDHs, most microorganisms cannot naturally accumulate aldehydes [
19,
20]. These endogenous enzymes pose significant barriers to the enhanced production of aldehydes in engineered microorganisms. As previously described, many microbial platforms for aldehyde production employ the knockout of endogenous oxidoreductases to varying extents to increase aldehyde yields. For different target aldehyde products, selectively screening and knocking out specific oxidoreductases can facilitate the accumulation of the desired aldehyde. However, the inactivation of these enzymes may disrupt the endogenous metabolic regulation of the cells, potentially impacting their growth and diminishing their industrial applicability. For instance, in a study on the production of isobutyraldehyde using
E. coli, the deletion of the gene
fucO encoding endogenous isobutyraldehyde reductase resulted in higher levels of isobutyraldehyde accumulation. Still, the deletion of this gene led to a significant reduction in 24-h growth of the strain [
33]. Therefore, screening for endogenous enzymes that can accumulate aldehydes without affecting growth after knockout is crucial for subsequent industrial applications. In synthetic biology,
E. coli and
S. cerevisiae are the two most commonly utilized host microorganisms. Currently, several modified platforms of
E. coli and
S. cerevisiae have been reported to successfully accumulate aldehydes after the knockout of various endogenous oxidoreductases, with growth rates comparable to those of wild-type strains. For instance, the engineered strains of
E. coli AL1827 [
39],
E. coli RARE [
44],
E. coli ROAR [
46], and
S. cerevisiae MARE [
51] are capable of accumulating a range of fatty aldehydes (C2–C12) and aromatic aldehydes, with a normal growth state.
3.2. Mitigating the Cytotoxicity of Aldehydes
Microbial production of aldehydes presents an enticing alternative to the extraction of aldehydes from botanical sources or their chemical synthesis. Nonetheless, the inherent toxicity of aldehydes to host microorganisms stands as a principal barrier to their widespread industrial utilization [
74,
75]. Within microbial cells, aldehydes can inflict damage on the functionality of macromolecules, including DNA, RNA, proteins, and lipids, which results in the disruption of cellular processes and ultimately manifests as cytotoxicity [
76,
77,
78,
79,
80,
81]. To realize the large-scale application of microbial-derived aldehydes, addressing the issue of aldehyde-induced cytotoxicity is imperative. This challenge necessitates the development of strategies to mitigate or detoxify the impact of aldehydes on cellular health, ensuring the viability and productivity of the microbial cell factories.
3.2.1.
In Situ Product Recovery (ISPR) Techniques
The integration of ISPR in the microbial production of aldehydes, which are often toxic to the host organisms, emerges as a highly promising strategy. This process facilitates the continuous elimination of the target aldehydes from the culture medium, thereby averting the accumulation of toxic levels that could impede cell growth and metabolism [
30,
31,
33,
48]. On one hand, ISPR can provide a stimulus for the further formation of the product by reducing the concentration of the product. On the other hand, it enhances productivity by circumventing the growth inhibition that would otherwise be caused by the product [
82].
For instance, Ku et al. [
30] employed oil alcohol as an in situ extractant to remove butyraldehyde from the fermentation broth during its synthesis with engineered
E. coli. This approach led to a threefold increase in the yield of butyraldehyde. Similarly, in a study focused on the production of isobutyraldehyde in
E. coli, a gas stripping system was implemented to remove the product
in situ, leading to a significant enhancement in the production titer, reaching up to 35g/L [
33]. In another investigation aimed at producing cinnamaldehyde with
E. coli, Bang et al. opted for the in situ isolation of the product from tributyl ester, which yielded a product titer of up to 3.8 g/L [
48].
The effectiveness of ISPR technology employed in microbial cell factories hinges on the judicious selection of extraction methods and the choice of solvents [
82]. The selected method must not only demonstrate high extraction efficiency for the target aldehyde but also maintain high biocompatibility with the production host.The efficacy of the ISPR process is highly dependent on the physical properties of the target aldehyde, such as its boiling point and solubility, which dictate the optimal conditions for product removal and cell growth. Therefore, a tailored approach based on these properties is essential for the successful implementation of ISPR in the large-scale production of aldehydes by microbial cell factories.
3.2.2. Alternative Processing Strategies
Efflux pumps Snq2p and Pdr5p have been reported to potentially facilitate the efflux of long-chain aldehydes, thereby enhancing cellular tolerance to these compounds [
83]. Considering the substrate specificity of these pumps, strategies such as directed evolution or structure-based enzyme engineering could be effectively employed to develop aldehyde-specific efflux pumps [
84,
85]. Notably, several efflux pumps, including multidrug-resistant nodulation pumps like MexEF-OprN, have been identified for their role in improving bacterial tolerance to aldehydes [
78,
86,
87]. The introduction of an external discharge pump system may significantly improve the tolerance of microbial cell factories to aldehydes, offering a promising avenue for strain engineering in synthetic biology.
The employment of resting whole-cell biocatalysts for the production of toxic products, such as aldehydes, presents a viable alternative. These biocatalysts are initially amassed through high-density cell cultivation, followed by concentration and resuspension under buffered conditions, and subsequently, the addition of substrates to initiate product synthesis. Recombinant cells utilized as whole-cell biocatalysts can be in the form of resting cells or freeze-dried cells, which do not necessitate further growth, thus making the synthesis process less susceptible to cytotoxic effects of the products [
88,
89]. Whole-cell catalysis has been effectively used to produce certain cytotoxic compounds. When combined with
de novo synthesis, it enables the generation of toxic products from cost-effective carbon sources like glucose or glycerol [
90,
91]. This approach may offer a feasible solution for mitigating the cytotoxicity associated with aldehydes.
The acquisition of microbial chassis cells with heightened aldehyde tolerance through directed evolution may effectively mitigate the cytotoxic effects of aldehydes. Adaptive laboratory evolution (ALE) permits the accumulation of mutations that enhance cellular resistance [
92]. This method has been extensively applied to improve tolerance to various stressful environments, including the presence of toxic products. Industrially relevant microbial cell factories have been successfully tailored by integrating ALE with rational strain engineering strategies [
93,
94,
95]. ALE enables further optimization of engineered genomes and fine-tuning of translation and transcription mechanisms to achieve desired phenotypes.Recently, Wang et al. [
96] developed an innovative random mutation system based on functional TadA variants, which can accelerate genomic evolution. This system applied to the ALE of Cd
2+ and kanamycin resistance, enhanced
E. coli’s short-term tolerance to heavy metals and antibiotics. Similarly, this system could possibly be adapted to improve cellular tolerance to aldehydes. By applying ALE to refine and evolve the adaptability and metabolic traits of chassis cells in high-aldehyde environments, there is a possibility of developing a stable and heritable microbial platform with aldehyde tolerance.
Conceptualization, J.Y.; Investigation, Y.C. (Yueyang Chen), J.F., Q.M. and Y.C. (Yongming Choe); Writing—Original Draft Preparation, Y.C. (Yueyang Chen), J.F. and Q.M.; Writing—Review & Editing, Y.C. (Yueyang Chen) and H.C.; Visualization, Q.M. and Y.C. (Yongming Choe); Project Administration, Y.Z. and J.Y.; Funding Acquisition, Z.X. and J.Y.
Not applicable.
Not applicable.
This work was supported by the National Key Research and Development Program of China (2023YFA0915800), the National Natural Science Foundation of China (grant no. 32270087), Xiamen University (grant no. 20720240120) and ZhenSheng Biotech.
The authors declared no competing interest.
 Zheyong Xue
3,*
Zheyong Xue
3,*