1. Introduction
Plastics have been applied in the fields of clothing, packaging, pharmaceutical compounds, and building, owing to the performance of light-weight, inexpensive, strong, durable, and flexibility [
1,
2]. In 2022, the global production of plastics amounted to 400.3 million tons, in which 90% derived from fossil fuels [
3]. Given that the majority of plastics were disposable, the accumulation of vast plastic waste has resulted in severe environmental issues. For example, the global life-cycle greenhouse gas emissions from plastics were growing rapidly with an average annual growth rate of 4%. Additionally, the persistence of a substantial amount of plastic waste in the marine environment can cause severe damage to oceanic ecosystems and human health [
4,
5].
Four main strategies, which were used for plastic waste disposal around the world, included landfill, incineration, regenerative granulation, and pyrolysis [
6]. Most plastic waste was degraded and processed through these strategies, but they also had some apparent drawbacks. Landfill carried the risk of soil leaching and compaction, potentially leading to groundwater contamination. The process of plastic waste through waste treatment systems can lead to the accumulation of organic pollutants, which had various adverse effects on human health [
7,
8]. The improper disposal of plastic waste may result in the loss of its economic value such as traditional plastics recycling process, which often involved energy-intensive treatment steps. In addition, the environmental pollution resulted in additional costs for environmental remediation and treatment [
9]. In chemical recycling, pyrolysis and solvent decomposition routes can effectively convert polymers back into monomers and oligomers. However, the low cost of virgin monomers derived from fossil carbon-based feedstocks often rendered chemical recycling commercially unfeasible [
10]. Biodegradation and sustainable conversion methods for plastic waste were gaining more attention due to their potential to address the shortcomings of traditional disposal methods. Biodegradation of plastic waste can be achieved through enzymatic degradation or microbial degradation. In comparison to chemical recycling, enzymatic degradation proved to be a gentler and more energy-efficient process, with reduced associated environmental pollution. Additionally, the utilization of biological techniques, such as enzyme immobilization, can facilitate the reuse of enzymes and reduce costs. Furthermore, the conversion of waste into high-value compounds enabled the reuse and the recycle of resources, reducing the necessity for virgin resources.
Biodegradation referred to the process in which microorganisms in the environment can convert materials into natural substances [
11]. Biodegradable plastics, such as polylactic acid (PLA), poly(butylene succinate) (PBS), poly(butylene adipate terephthalate) (PBAT) and polyhydroxyalkanoates (PHAs) [
12], had chemical structures that allowed them to be broken down by microorganisms. Microorganisms mainly broke down these plastics into one-carbon substances such as carbon dioxide and methane [
13]. Non-biodegradable synthetic plastics are those that cannot be degraded significantly by microbial cells in the natural environment within a reasonable time. These plastics were usually made from petroleum-based raw materials, including polyethylene terephthalate (PET), polyethylene (PE), polypropylene (PP), polyvinyl chloride (PVC), and polystyrene (PS) [
14]. Even so, they can be broken down into corresponding monomers or small molecules through enzymatic degradation. Some degradation products had natural value, such as adipic acid (AA) [
15]. Some monomers can be converted into value-added compounds [
16]. For instance, monomer ethylene glycol (EG) can be converted into glycolic acid (GA) [
17]. The economic value of plastic waste was low in its original state, due to the costs associated with its disposal. Conversely, the value was significantly enhanced through the conversion of plastic waste into high-value chemicals. The process of high-value conversion of plastics can be divided into three stages: collection and handling of plastics, degradation and conversion of plastics, and separation of high-value products. Among them, the biodegradation and conversion of plastics was technically feasible. Several molecular manipulation tools can be used to modify microorganisms to enhance their biodegradation efficiency and optimize conversion pathways to meet the needs of industrial production. In addition, in the stage of collection and handling of plastics, the biodegradation process was a relatively low-energy process, with reduced energy costs compared to the physical method of recovery. The value of the target product had a significant influence on the economic viability of the conversion process. For example, the bioconversion of PET to adipic acid was potentially economically beneficial. The bioconversion has been technically achieved in
E. coli with the conversion rate of 79%. Furthermore, the recycling rate of PET bottles in China has exceeded 94%, which provided an adequate raw material base and reduced the cost of raw materials for the high-value conversion of PET.
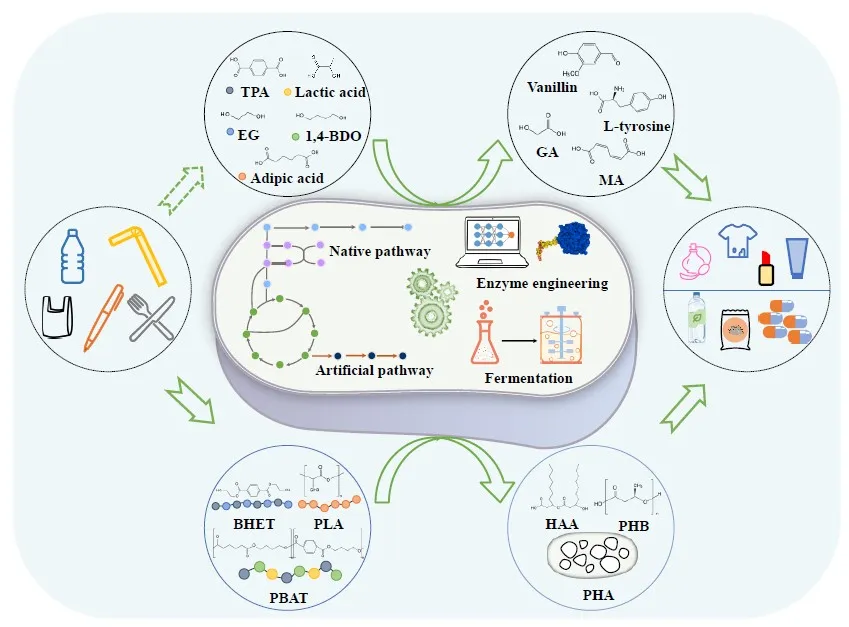
So far, many enzymes and microorganisms which can degrade plastics, have been reported. Based on this, this review summarized the current biodegradation and conversion of plastic waste and analyzes the current problems in converting plastics into value-added compounds. As shown in , we classified the high-value conversion of plastics into direct conversion of monomers and continuous conversion. Finally, these value-added compounds can be used to manufacture cosmetics, clothes and bioplastic products.
Figure 1. Bioconversion of the degradation products from plastics into value-added chemicals.
2. Biological Depolymerization Methods on Plastic
2.1. Depolymerization of Biodegradable Plastic
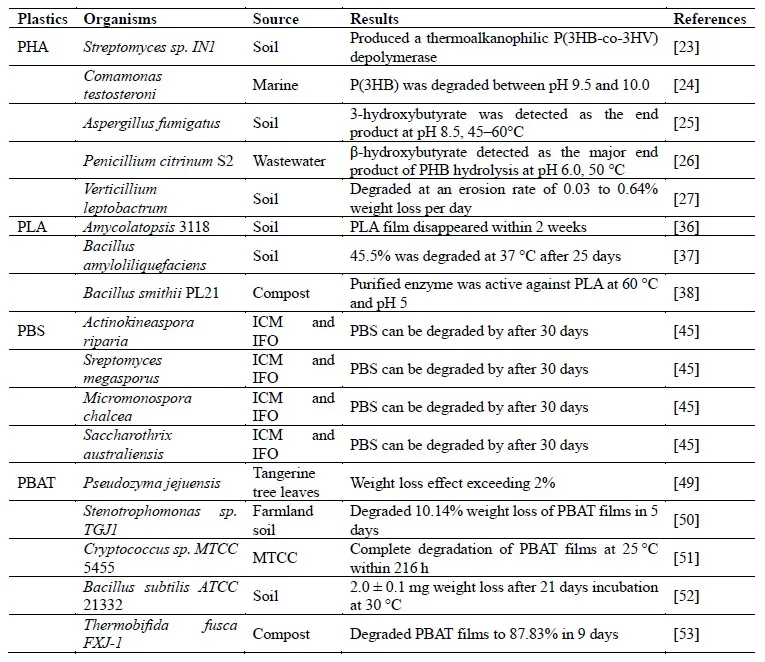
Biodegradable plastics, which included PLA, PBS, PBAT and PHAs, were the plastics that can be degraded by microorganisms to generate CO
2, CH
4, and microbial biomass [
18]. Some microorganisms that degraded biodegradable plastic were listed in . The biodegradability made these plastics significantly reduced their impact on ecosystems.
2.1.1. PHA
PHAs were biodegradable microbial polyesters that can be produced by microorganisms [
19]. The superiority of PHAs compared with other plastics was the complete biodegradability under aerobic and anaerobic conditions [
20]. Various microorganisms, which were capable of degrading PHAs, have been discovered in environments such as soil, compost, and ocean () [
21]. Among these degrading microorganisms, the main bacteria were
Pseudomonas lemoigne [
22],
Streptomyces sp. IN1 [
23],
Comamonas testosteroni [
24], and the main fungi were
Aspergillus fumigatus [
25],
Penicillium citrinum S2 [
26], and
Verticillium leptobactrum [
27]. Generally, the natural environments were conducive to the degradation of PHA in aerobic conditions. Besides, there were still some microorganisms that can degrade PHA under anaerobic conditions, such as
Ilyobacter delafieldii sp. nov,
Cloacamonales,
Thermotogales and
Clostridium [
28,
29,
30,
31].
Ilyobacter delafieldii sp. nov was the first anaerobic bacterium that can degrade exogenous PHB into 3-hydroxybutyrate.
Clostridium that isolated from sludge can degrade PHB powder in a pH-controlled bioreactor system. The anaerobic degradation of PHB can be achieved within 3 days.
The enzymatic degradation of PHA can be categorized into intracellular depolymerases and extracellular depolymerases. Intracellular depolymerases broke down PHA chains into small molecules within the PHA-producing strains, and these molecules can be utilized as carbon and energy source for the growth of microorganism [
32]. Extracellular depolymerases, which acted on PHA outside of cells, have been identified in various bacteria and fungi. These enzymes can encounter PHA biomaterials and initiate the process of depolymerization [
33]. Recently, Zhou et al. investigated that
Schlegelella thermodepolymerans (
S. thermodepolymerans) may possess extracellular depolymerases, and
S. thermodepolymerans showed a faster extracellular PHA degradation speed at 50 °C compared with mesophilic conditions [
28,
34]. The presence of these enzymes provided the possibility for the degradation of PHA under extreme conditions. However, the complete degradation of PHA to the monomers remained a challenge. For example, the use of base catalysts for the degradation of poly(3-hydroxybutyrate) (P3HB) only produced the non-original monomer transcrotonic acid, which affected the repolymerization of bioplastic P3HB [
35].
2.1.2. PLA& PBS
PLA was a biodegradable plastic with a wide range of applications in packaging materials, agriculture and pharmaceuticals. According to PlasticDB (http://plasticdb.org), over 150 strains, 6 uncultured bacteria, and 27 PLA-depolymerases were capable of degrading PLA, blends of PLA, or copolymers of lactic acid. These strains contained bacteria like
Amycolatopsis 3118 [
36],
Bacillus amyloliliquefaciens [
37],
Bacillus smithii PL21 [
38], and as well as fungi
Cryptococcus magnus [
39] and
Penicillium, etc [
40]. The commercial product Proteinase K was a hydrolase which was capable of effectively degrading PLA [
41,
42]. Additionally, a fungal-like keratinase (CLE1) which was expressed in
Saccharomyces cerevisiae, produced a crude supernatant that efficiently hydrolyzed different types of PLA materials, releasing up to 9.44 g/L lactic acid from 10 g/L PLA film [
43]. PBS was a biodegradable plastic which can be synthesized through polycondensation reaction between 1,4-butanediol (1,4-BDO) and succinic acid. Microorganisms that can degrade PBS, contained
Actinokineaspora riparia,
Sreptomyces megasporus,
Micromonospora chalcea,
Saccharothrix australiensis, and
Excellospora viridilutea, etc [
44,
45]. However, microbial degradation was often slow and required strict environmental condition. Currently, only a few enzymes had the ability of degrading PBS, and their efficiency were generally low. Low-efficiency enzymes prolonged the degradation process, which resulted in extended production cycles and decreased production efficiency. This further increased costs, including energy, labor, and equipment usage costs [
46].
2.1.3. PBAT
PBAT, which was synthesized by polymerizing a mixture of AA, 1,4-BDO, and terephthalic acid (TPA) [
47], has gained significant attention due to its desirable physical properties. One of its key characteristics was high flexibility but relatively low strength compared with other bioplastics [
48]. Several microorganisms that can degrade PBAT contained
Pseudozyma jejuensis,
Stenotrophomonas sp. TGJ1,
Cryptococcus sp. MTCC 5455,
Bacillus subtilis ATCC 21332, and
Thermobifida fusca FXJ-1 [
49,
50,
51,
52,
53]. Biodegradable polyesters were typically broken down by enzymes such as esterases, lipases, proteases and cutinases. Among these hydrolases, lipases and esterases showed low efficiency due to the narrow substrate-binding tunnel, and the lid region that covered the active center. In contrast, cutinases harbored broad substrate-binding cavities and lid-free active site, which appeared to be the most promising PBAT hydrolase [
54,
55]. For example, the double mutation strategy was applied to modify cutinase that derived from
Thermobifida fusca, generating a mutant with a more flexible substrate-binding pocket and high decomposition rate [
56]. Moreover, cutinase variant TfCut-DM Q132Y can completely degrade PBAT film to TPA in 24 h at 60 ℃, which represented the most efficient and energy-saving choice for enzyme-mediated PBAT decomposition [
57]. In addition, biosensors were applied to the high-throughput screening of hydrolases. Shin et al. constructed an AA biosensor based on PcaR transcription factor. The optimized sensor showed high sensitivity (low detection concentration < 10 μM) and dynamic range (~50-fold). In particular, the novel AA sensors can contribute to the discovery and evolution of effective biocatalysts [
58]. However, the degradation of PBAT in the actual application process was not ideal. When degrading PBAT mulch that used in agriculture, the degradation process was usually affected by light, temperature, pH and mechanical forces [
47]. Therefore, the development of enzymes or bacteria with improved stability and degradation efficiency of PBAT was urgently needed. These stable enzymes or bacteria can maintain activity under different environmental conditions and effectively break down PBAT.
. Microorganisms that degrade biodegradable plastic.
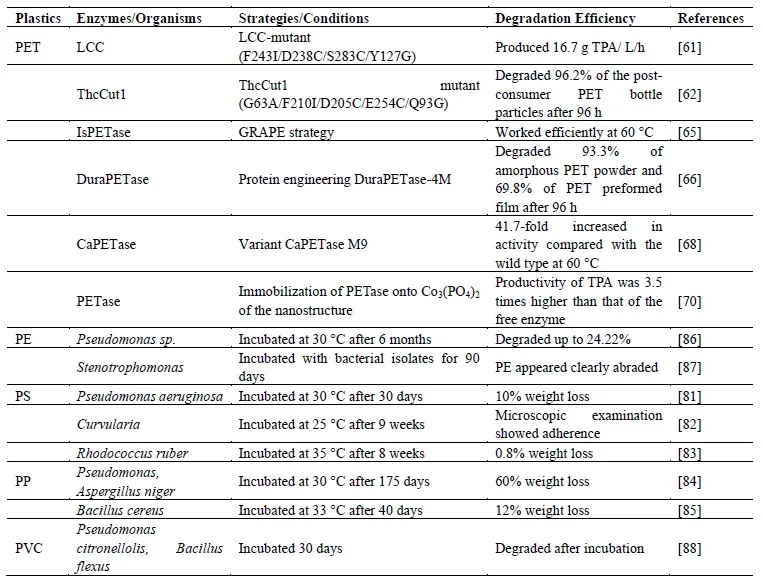
2.2.1. PET
Many methods can be used to degrade PET into monomers TPA and EG, such as hydrolysis, methanolysis, glycolysis, aminolysis, and ammonolysis [
59]. Enzymatic hydrolysis offered a milder and more environmentally friendly approach to degrading PET compared with traditional chemical methods. Carboxylic ester hydrolases (EC 3.1.1) such as lipases, esterases, cutinases, and PETases, played a crucial role in the PET hydrolysis [
60]. We listed enzymes that degraded PET in .
Tournier et al. utilized computer-aided enzyme engineering to modify leaf-branch compost cutinase (LCC) derived from leaf-cutinase branch compost metagenome. The LCC-variant (F243I/D238C/S283C/Y127G) has been designed to enhance its depolymerization specific activity, and the introduction of disulfide bonds at the calcium-binding site contributed to thermostability. The LCC-variant catalyzed PET to TPA with a mean productivity of 16.7 g
TPA·l
−1·h
−1 [
61]. Cutinase 1 (ThcCut1) derived from Thermobifida cellulosilytica exhibited higher activity with ester bonds. The ThcCut1 mutant (G63A/F210I/D205C/E254C/Q93G) can degrade 96.2% of the post-consumer PET bottle particles after 96 h [
62].
A two-enzyme system of PET hydrolase (PETase) and MHET hydrolase (MHETase) derived from
Ideonella sakaiensis (
I. sakaiensis), was employed to depolymerize PET to TPA and EG. The system was biochemically identified and characterized to degrade PET in vitro [
63,
64]. Recently, Cui et al. improved the robustness of a IsPETase (PETase derived from
I. sakaiensis) through the GRAPE strategy (greedy accumulated strategy for protein engineering), the IsPETase mutant (DuraPETase) possessed high catalytic efficiency and worked efficiently at 60 °C for more than 24 h [
65]. Further, Liu et al. designed DuraPETase through protein engineering. The constructed DuraPETase-4M can degrade 93.3% of amorphous PET powder and 69.8% of PET preformed film after 96 h. These degradation rates were 3.2 and 5.4 times higher than DuraPETase, respectively [
66]. Directed evolution was conducted on PETase using a novel high-throughput fluorescence screening assay, which utilized a novel substrate bis(2-hydroxyethyl)2-hydroxyterephthalate (BHET-OH). The generated DepoPETase enabled complete depolymerization of untreated PET wastes at mild temperature [
67]. The novel PET hydrolase CaPETase, which combined the advantages of PETase and LCC, exhibited high catalytic activity and high thermal stability at ambient temperature. After rational protein engineering, the variant CaPETase M9 showed a remarkable 41.7-fold increase in activity compared with the wild type at 60 °C [
68]. A hydrophobic cell surface display (HCSD) system was constructed as a whole-cell catalyst. The truncated outer membrane hybrid protein FadL exposed PETase and hydrophobic protein HFBII to the surface of
E. coli and exhibited efficient PET accessibility and degradation, with a higher degradation rate than free PETase [
69]. Immobilization of enzymes was also an important method to improve enzyme activity. Immobilization of PETase onto Co
3(PO
4)
2 of the nanostructure resulted in an effective increase in enzyme loading, with long reaction times showing that the productivity of TPA was 3.5 times higher than that of the free enzyme [
70]. The mining and directed evolution of PET and Phthalate esters (PAEs) hydrolases were slow due to the lack of fast and efficient detection methods. Li et al. established PA and TPA cell sensors by directional evolution transcription factor XylS, and used this sensor to aid the directed evolution of PAE hydrolase GoEst15. The degradation efficiency of dibutyl phthalate and 4-nitrophenylbutyrate in the resulting mutant was increased by 2 and 2.5 times compared with the wild type, respectively [
71].
However, PET hydrolases exhibited significant degradation effects primarily on PET with low crystallinity, typically around 10%. If the crystallinity of PET exceeded 20%, high-temperature pretreatment was necessary. This process not only consumed a considerable amount of energy but also increased processing costs [
72]. PET was typically composed of a complex structure comprising tightly packed parallel chains in the crystalline regions and disordered chains in the amorphous regions [
73]. PET molecules with higher crystallinity were more tightly arranged, with smaller gaps between the molecules, making it more difficult for enzyme molecules to penetrate the structure of the PET molecule. Consequently, the rate of degradation was lower. Additionally, with increased crystallinity, the flexibility and mobility of the polymer chains were constrained, reducing their susceptibility to enzymes. Furthermore, the repetitive units of terephthalate present in PET also limited its mobility and reduced its degradation efficiency [
74].
2.2.2. PE, PP, PS, PVC
Plastics with carbon-carbon backbone such as PE, PP, PS, PVC, were difficult to be degraded using microbes, because C-C bonds were resistant to microbial attack and biodegradation. Some Bacterial species and fungi can depolymerize PE after some forms of pretreatment [
75]. Over 20 bacterial genera can degrade PE such as
Pseudomonas,
Ralstonia,
Stenotrophomonas,
Klebsiella and
Acinetobactor [
76]. Fungi were thought to be more efficient than bacteria for the degradation of PE because they can attach to the hydrophobic surface of the polymers [
77]. Fungal laccases and peroxidases, generally used by fungi to degrade lignin, showed high efficiency in degrading PE and PVC. However, the complex biodegradation mechanism of PE was not fully understood yet. Peixoto et al. found a novel role of nitrogen metabolism in the breakdown and oxidation of PE, which was associated with biological production of NOx in three PE-degrading strains of
Comamonas,
Delftia, and
Stenotrophomonas [
78]. The biodegradation of PE and PP was challenging due to the lack of unsaturated double-bonds in their structure. Consequently, the abiotic pretreatment before degradation was typically required. Some pretreatment methods, such as ultraviolet radiation (UV), heating, and chemical agent can enhance the degradation efficiency [
79,
80]. The biodegradation of PS by several microbes such as
Pseudomonas aeruginosa,
Curvularia, and
Rhodococcus ruber etc, has been demonstrated [
81,
82,
83]. The number of published studies on the degradation of polypropylene by various microorganisms was limited. Among the microorganisms that have been observed to degrade PP were
Pseudomonas and
Vibrio, as well as the fungus
Aspergillus niger [
84,
85].
. Enzymes and microorganisms that degrade non-biodegradable plastic.
3. Direct Conversion of Monomers into Value-Added Chemicals
The high-value conversion of plastic waste has been widely studied, with particular emphasis on the transformation of plastic monomers. For example, TPA was one of the monomers of PET, which can be converted into value-added compounds such as vanillin. Additionally, some monomers derived from plastic degradation, such as 1,4-BDO and succinic acid, were natural value-added compounds with significant applications.
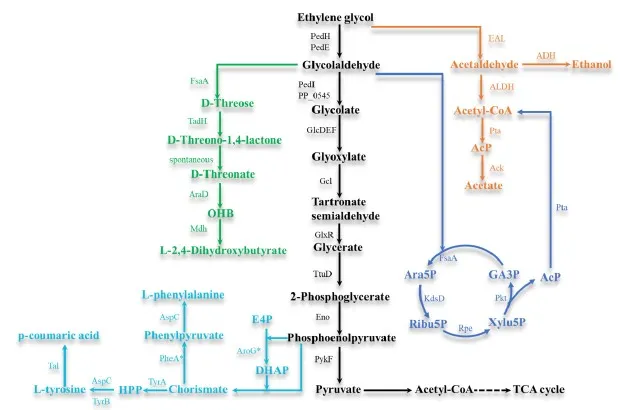
3.1 EG
EG was widely used as antifreeze agent and chemical intermediate, especially as critical precursor in the production of PET [
89,
90]. The catabolic pathways of EG to value-added chemicals have been shown in . Acetic acid bacteria (AAB), such as
Acetobacter,
Gluconobacter,
Gluconacetobacter and
Komagataeibacter genera, possessed a unique metabolism that allows them to oxidize several carbohydrates, alcohols and polyols into carboxylic acids, aldehydes or ketones [
91]. EG can be converted into GA by several AAB especially
Gluconobacter oxydans [
92].
Yarrowia lipolytica (
Y. lipolytica) may also share an incomplete EG oxidation pathway, because 429.5 mM GA can be obtained in bioreactors from EG [
93]. EG was also naturally utilized by other bacteria like
Pseudomonas aeruginosa. EG can be converted into glycolaldehyde and GA through two functionally redundant periplasmic Quino proteins PedE and PedH [
94,
95]. Franden et al. clarified the metabolic pathway of EG in
Pseudomonas putida KT2440 (
P. putida KT2440). The additional overexpression of the glycolate oxidase (
glcDEF) operon overcame the toxicity of glycolaldehyde, and the engineered strain can utilize 31 g/L EG to produce medium chain length PHA (mcl-PHA) with a productivity of 0.06 g/g [
96].
Wild type
Escherichia coli (
E. coli) cannot utilize EG as sole carbon for growth because the propanediol oxidoreductase (FucO) that utilized EG was oxygen sensitive [
97]. Lu et al. discovered a FucO mutant (Fuco
∗) with I7L and L8V, which possessed improved oxygen tolerance and can be used to consume EG [
98,
99]. Fuco
∗ and glycolaldehyde dehydrogenase (aldA) have been co-expressed in
E. coli to produce GA. Pandit et al. integrated modeling with experimental investigations and identified oxygen concentration as a crucial metabolic regulator. Finally, a GA titer of 10.4 g/L was attained after 112 h in a fed-batch bioreactor [
17]. L-tyrosine was one of the three aromatic amino acids involved in protein biosynthesis [
100]. Panda et al. engineered
E. coli to transform EG into L-tyrosine. The engineered strain produced 2 g/L L-tyrosine from 10 g/L EG through the improvement of EG assimilation and the overcoming of L-tyrosine feedback resistance, and this strain outperformed glucose under the same conditions. Otherwise, a similar quantity of L-tyrosine can be obtained when using EG derived PET waste to replace commercial EG [
101]. Frazão et al. extended the range of glycolaldehyde-dependent pathways by demonstrating production of 2,4-dihydroxybutyric acid (DHB). The nonnatural pathway involved several enzymatic steps in the conversion of glycolaldehyde to DHB. The sequential action of the D-threose aldolase, D-threose dehydrogenase, D-threono-1,4-lactonase, D-threonate dehydratase and 2-oxo-4-hydroxybutyrate (OHB) reductase enzyme was employed. Indeed, incorporating an EG oxidation step into the nonnatural pathway enabled the conversion of EG into DHB [
102].
Apart from the value-added chemicals mentioned above, EG also can be converted into important platform chemicals and green fuels such as ethanol. Acetyl-CoA was an important metabolic intermediate that can be used to regulate intracellular growth and generate a variety of chemicals such as 3-hydroxypropionic acid, terpenoids and fatty acids [
103,
104,
105]. The carbon utilization efficiency of glycolaldehyde was found to be low in the native strains, serving a precursor function for producing acetyl-CoA. Recently, some new acetyl-CoA biosynthetic pathways have been created to increase carbon utilization efficiency [
106]. A synthetic acetyl-CoA (SACA) pathway was constructed by repurposing glycolaldehyde synthase and acetyl-phosphate synthase, which was verified by
13C-labeled metabolites. Meanwhile, SACA pathway was further verified by cell growth using glycolaldehyde, formaldehyde and methanol as supplemental carbon source. The SACA pathway was proved to be the shortest, ATP-independent, carbon-conserving and oxygen-insensitive pathway for acetyl-CoA biosynthesis [
107]. The Ara5P-dependent glycolaldehyde assimilation pathway (GAA) was proposed for the conversion of EG into acetyl-CoA without carbon loss. GAA pathway has been verified in vitro and vivo [
108,
109]. Acetate and ethanol were important products of the acetyl-CoA metabolic pathway.
I. sakaiensis was capable of growth on PET as the major carbon and energy source. The consumption of 25 mM EG resulted in the production of 3.3 mM ethanol and 21.8 mM acetate after 17 and 22 days, respectively. [
110,
111].
Figure 2. Catabolic pathways of EG to value-added chemicals. OHB:2-oxo-4-hydroxybutyrate; Ara5P: d-arabinose 5-phosphate; Ribu5P: d-ribulose 5-phosphate; Xylu5P: d-xylulose 5-phosphate; GA3P: glyceraldehyde 3-phosphate; E4P: erythrose-4-phosphate; DHAP: 3-deoxy-D-arabino-heptulosonate 7-phosphate; HPP: 4-hydroxyphenylpyruvate. PedE/PedH: PQQ-dependent alcolhol dehydrogenase; PedI: peroxisomal 3-ketoacyl-COA thiolase; GlcDEF: glycolate dehydrogenase; GlxR: tatronate semialdehyde reductase; Gcl: glyoxylate carboligase; TtuD: hydroxypyruvate reductase; Eno: Enolase; pykF: pyruvate kinase; FsaA: D-threose aldolase; TaDH:D-threose dehydrogenase; AraD: D-threonate dehydratase; Mdh: OHB reductase; FsaA: arabinose 5-phosphate aldolase; KdsD: D-arabinose 5-phosphate isomerase; Rpe: D-ribulose 5-phosphate 3-epimerase; Pkt: heterologous phosphoketolase; Pta: phosphate acetyltransferase; AroG∗: DAHP synthase D146N; TyrA: chorismate mutase/prephenate dehydrogenase; Tal: tyrosine ammonia lyase; EAL: ethanolamine ammonia lyase; ADH: alcohol dehydrogenase; ALDH: aldehyde dehydrogenase; Ack: acetate kinase. PheA∗: Bifunctional chorismate mutase/prephenate dehydratase T326P.
Currently, TPA was primarily obtained through the oxidation of p-xylene using either chemical or microbial methods [
112,
113]. TPA has been widely utilized in various fields, such as polyester fibers, feed additives and pharmaceuticals. Besides, TPA can also be converted into value-added compounds. Several strains, including
Comamonas sp. E6,
I. sakaiensis,
P. umsongensis GO16, and
R. jostii RHA1.13 [
114], have been verified in metabolizing TPA.
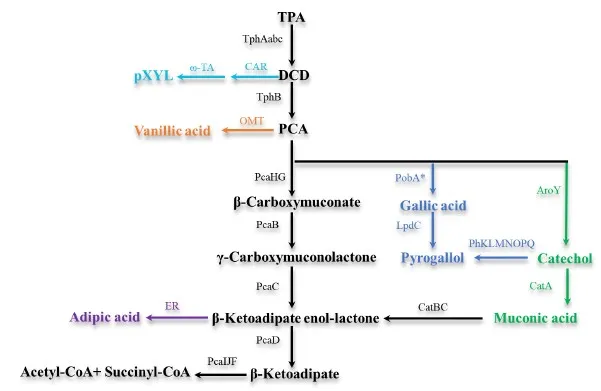
TPA can be converted by the engineered microorganisms into five different aromatics or aromatic-derived compounds, including gallic acid, pyrogallol, catechol, muconic acid (MA), and vanillic acid (). These five products were produced using protocatechuic acid as the principal intermediate, achieving relatively high molar conversion yields ranging from 32.7% to 92.5% [
115]. Vanillin was a widely used additive in the food industry [
116]. Sadler et al. developed a novel pathway in engineered
E. coli for the direct conversion of TPA into vanillin using whole-cell catalysts. The pathway involved several enzymes including terephthalate 1,2-dioxygenase, dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylic acid dehydrogenase, carboxylic acid reductase and catechol O-methyltransferase. After optimizing the reaction conditions, the maximum yield of vanillin reached 119 mg/L with a conversion rate of 79% [
117]. In addition, four cleavable vanillin-based polyols were prepared from vanillin, 2,5-furandicarboxylic acid and TPA, followed by the cross-linking with two different isocyanates. The resulting polyurethane samples exhibited reasonable mechanical strength, thermostability, solvent resistance and outstanding degradability [
118].
Bayer et al. proposed a new strategy for the conversion of TPA into various diamines. Carboxylic acid reductase (CAR) catalyzed TPA to produce terephthalaldehyde (TAL). TAL was catalyzed through a chemical reductive amination cascade in a one-pot process to produce 1,4-bis-(aminomethyl) benzene and benzylamine [
119]. Additionally, para-xylylenediamine (pXYL), which was used as a building block for polymeric materials, can be obtained of 69 ± 1% yield when combining CAR from
Segniliparus rotundus (srCAR) and ω-transaminase from
Chromobacterium violaceum (cvTA) [
120]. Valenzuela-Ortega et al. developed the first one-pot bio-upcycling of PET plastic waste into AA in
E. coli. The rate-limiting enoate reductase (BcER) was interfaced with hydrogen gas and a biocompatible Pd catalyst to synthesize AA. The product conversion is high (79%, 115 mg/L) under ambient conditions in 24 h [
15].
Figure 3. Catabolic pathways of TPA to value-added chemicals. TPA: Terephthalic acid; DCD: 1,6-dihydroxycylohexa-2,4-diene-dicarboxylate. TphAabc: TPA 1,2-dioxygenase; TphB: 1,2-dihydroxy-3,5-cyclohexadiene-1,4-dicarboxylate dehydrogenase; PcaHG: protocatechuate 3,4-dioxygenase; PcaB: 3-carboxy-cis,cis-muconate cycloisomerase; PcaC: γ-carboxy-muconolactone decarboxylase; PcaD: β-ketoadipate enol-lactone; PcaIJF: 3-oxoadipate CoA-transferase; PobA∗: p-hydroxybenzoate hydroxylase T294A/Y385F; LpdC: Gallic acid decarboxylase; AroY: PCA decarboxylase; PhKLMNOPQ: phenol hydroxylase; CatA: catechol 1,2-dioxygenase; CatBC: Muconate cycloisomerase 1/Muconolactone Delta-isomerase; ER: Enoate reductase; ω-TA: ω-transaminase; CAR carboxylic acid reductase; OMT:O-methyltransferase.
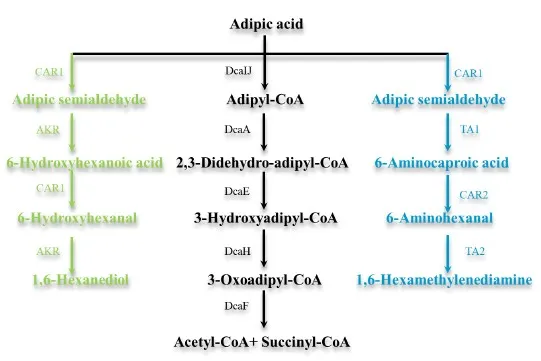
AA was a dicarboxylic acid with a wide range of applications, especially used for the production of PBAT and nylon 6-6 fibers, microbial degradation of AA has been characterized in
Acinetobacter baylyi [
121,
122,
123]. Fedorchuk et al. reported the biocatalytic conversion of AA into 6-aminocaproic acid (6-ACA) and 1,6-hexamethylenediamine (HMD), the key precursors for nylon synthesis (). The cascades of reduction/amination reactions involved carboxylic acid reductases (CARs) and transaminases (TAs). To increase the cascade activity for the transformation of 6-ACA to HMD, three CARs mutant with enhanced activity were designed by protein engineering. For the transformation of AA to HMD (via 6-ACA), the wild type CAR was combined with the variant and two different TAs, resulting in up to 30% conversion to HMD and 70% to 6-ACA [
124]. Besides, new CARs were created using accurate protein structure prediction-based virtual screening method, resulting in twice conversion in the enzymatic cascade synthesis of HMD [
125]. Additionally,
P. putida was engineered to produce PHA when utilizing AA as a carbon source. The initial uptake and activation steps for dicarboxylates were conducted, and adaptive laboratory evolution was employed to enable the growth of
P. putida on AA as the sole carbon source. In a nitrogen-limited mineral medium, the engineered strain produced 25.3 ± 4.2% PHA, representing a yield of 9.2% (g/g of carbon) [
126].
Figure 4. Catabolic pathways of AA to value-added chemicals. DcaIJ: CoA transferase subunits; DcaA: acyl-CoA dehydrogenase; DcaE: enoyl-CoA hydratase; DcaH: hydroxyl-CoA dehydrogenase; DcaF: thiolase; CAR1: carboxylic acid reductase; CAR2: engineered CAR; AKR: aldo-keto reductases; TA1/2: ω-transaminase.
Lactic acid was an important building block especially for the production of PLA [
127,
128]. Three methods were known for the synthesis of polymer PLA (Mw > 10,000), including direct condensation polymerization, azeotropic dehydration condensation, and lactide ring-opening polymerization [
129]. Lactic acid was also used in the production of some organic acids. The strain
Megasphaera elsdenii has been described to consume lactic acid and convert it into n-caproate. In this process, lactic acid was converted to acetyl-CoA, which then entered the reverse β-oxidation cycle. A maximum n-caproate productivity of 6.9 g COD/L-day was observed in 17 days (COD represents chemical oxygen demand), representing the highest sustained lactate to n-caproate conversion rate [
130]. Propionate was a valuable platform chemical with versatile applications such as food preservatives.
Clostridium saccharoperbutylacetonicum has been engineered to produce propionate by implementing the acrylate pathway from
Anaerobiospirillum succiniciproducens and introducing a D-lactate dehydrogenase enzyme. This modification allowed the bacterium to metabolize sugars to produce propionate as an end product [
131].
3.5 Succinic acid and 1,4-BDO
Succinic acid has been recognized as one of the most promising bio-based platform chemicals. In 2017, the market of bio-succinic acid was estimated at 175.7 million US dollars. The market is expected to grow at an annual growth rate of 20%, reaching 900 million US dollars by 2026 [
132]. Cui et al. engineered the strictly aerobic yeast
Y. lipolytica for efficient succinic acid production without pH control, the engineered strain produced 111.9 g/L succinic acid with a yield of 0.79 g/g glucose within 62 h [
133]. Succinic acid can be utilized as a precursor for industrially valuable commodity chemicals including 1,4-BDO, 5-Aminolevulinic acid (5-ALA), gamma-butyrolactone, tetrahydrofuran, and as a monomer for manufacturing various polymers [
134].
1,4-BDO was involved in the synthesis of PBS and PBAT as a monomer. In 2015, the global market size of 1,4-BDO was valued at 6.19 billion dollars, which is expected to reach 12.6 billion dollars by 2025 [
135]. It was known that 1,4-BDO can be produced in
E. coli by introducing an artificial pathway involving six exogenous genes. In this pathway, succinate and succinyl-CoA were important precursors to synthesize 1,4-BDO [
136]. Metabolism of 1,4-BDO has been reported in
P. putida KT2440 and
Y. lipolytica. Adaptive Laboratory Evolution enabled strain
P. putida KT2440 to grow using 1,4-BDO as the sole carbon source.1,4-BDO was oxidized to 4-hydroxybutyrate (4-HBA), which can be further metabolized to support growth by
P. putida KT2440. [
93,
137].
4. Continuous Conversion of Plastic into Value-Added Chemicals
The transformation of monomers after plastic degradation partially alleviated the issue of plastic waste. However, the conversion may require an additional carbon source to support growth. Moreover, the separation and purification of plastic monomers from their hydrolysis products are time-consuming and resource-intensive, rendering the entire process unsustainable. Continuous conversion from plastic to value-added compounds demonstrates significant advantages compared with monomers conversion. Continuous conversion eliminated the purification process of intermediates and the need for obtaining purified monomers. Additionally, direct utilization of hydrolysis products also enhanced the efficiency of conversion.
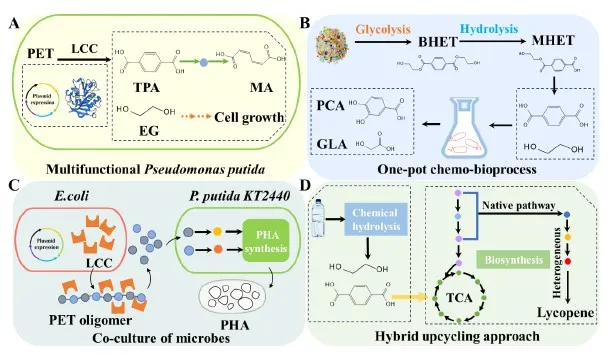
As shown in A, Liu et al. proposed a multifunctional
P. putida KT2440 for converting PET into MA, a precursor for the production of AA or TPA [
138]. The engineered strain was constructed to simultaneously secrete PET hydrolase LCC and synthesize MA. The hydrolysis product TPA served as a precursor for MA production, while EG supported cell growth. After optimizing the process, the yield reached 0.50 g MA/g PET [
139]. Diao et al. developed a hybrid strategy for converting PET into lycopene (D). PET was converted to TPA and EG using alkaline hydrolysis. Then,
Rhodococcus jostii strain PET (RPET) was able to directly use both TPA and EG as sole carbon sources to produce 1300 μg/L of lycopene, which provided beneficial effects to cellular functions in the human body [
140,
141].
In addition to using a single strain for the conversion of PET, the hybrid upcycling approach greatly increased the rate of PET degradation and conversion efficiency. Kim et al. developed a one-pot chemo-bioprocess, which integrated PET chemical glycolysis, enzymatic hydrolysis, and bioconversion (B). The pivotal strategy involved employing the biocompatible catalyst betaine during glycolysis. Finally, protocatechuic acid and GA were obtained [
142]. Besides,
Bacillus subtilis esterase (Bs2Est) catalyzed TPA to catechol via a whole-cell biotransformation. The catechol solution produced from TPA can be directly used for functional coating on various substrates after simple cell removal from the culture medium without further purification and water-evaporation [
143]. Tiso et al. utilized evolved
P. umsongensis GO16 KS3, which was capable of growth with the PET hydrolysis solution, to produce hydroxyalkanoyloxy alkanoates (HAAs) at a yield of 35 mg/L. HAAs showed surface-active properties and can be polymerized into bio-based poly(amide urethane) (bio-PU) [
144,
145]. Other than PET, PLA also can be converted into value-added compounds in a one-pot process. PLA can be converted into alanine by a simple ammonia solution treatment using a Ru/TiO2 catalyst. The process had a 77% yield of alanine at 140 °C [
146]. Jiao et al. demonstrated a one-pot catalytic process for the direct cracking of PLA into acrylic acid. This process employed an acid catalyst in conjunction with an ionic liquid, Bu4PBr, which facilitated the conversion of PLA to acrylic acid. The acrylic acid was then vaporized directly from the reactor and collected in a collector when PLA was completely converted [
147].
The co-culture of microorganisms has emerged as a more promising approach to enhance biosynthetic efficiency. Compared with monoculture, microbial co-culture can reduce the metabolic burden and meet the expression requirements of different genes. Additionally, multiple strains can provide varied cellular environments and confer significant benefits in utilizing mixed substrates. Liu et al. developed a one-step biological process for converting PET oligomers to PHA by the co-cultivation of
E. coli and
P. putida KT2440. The system produced PHA utilizing PET as substrate.
E. coli BL21 (DE3) was used to express and secrete the PET hydrolase (C). The genetically modified strain
P. putida was used to activate the metabolic pathway of TPA and EG while simultaneously overexpressing the PHA synthetic pathway [
148]. Liu et al. proposed a co-culture system to directly hydrolyze bis(2-hydroxyethyl) terephthalate (BHET) to generate the PHB in one fermentation step. The system contained a
Y. lipolytica Po1f harboring PETase from
I. sakaiensis and a TPA-degrading
Pseudomonas stutzeri containing the PHB synthesis pathway [
149,
150]. Bao et al. presented a synthetic microbial consortium that involved two
Pseudomonas putida strains, specializing in TPA and EG utilization respectively. Compared with a single strain for TPA and EG co-consumption under various substrate conditions, the microbial consortium has reduced catabolic cross-interactions between the pathways for a faster rate of substrate assimilation. The engineered system can be further optimized to design and integrate metabolic pathways that synthesized mcl-PHA and MA [
151].
The continuous conversion approach enhanced the efficiency of high-value transformation of plastics. In comparison to monomer conversion, it eliminated the need for monomer separation and purification steps. Moreover, the combination of multiple methods ensured an efficient connection between degradation and conversion, reducing the loss of intermediate products.
Figure 5. Four methods for continuous conversion of plastic into value-added chemicals.
5. From Plastic to Bioplastic
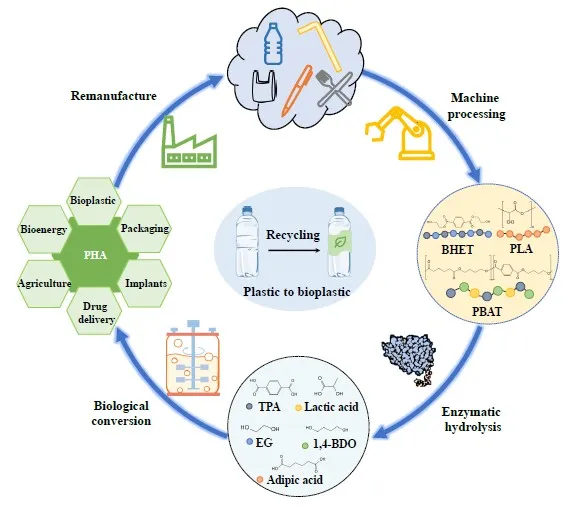
Bioplastics referred to the plastics of bio-based origin, or the plastics with biodegradable property, they have been applied in packaging, consumer goods, textiles, and biomedical devices. Total production of fossil plastics currently exceeded 380 million tons per year, while global production of 100% bio-based polymers was approximately 2 million tons per year. The global market share of bioplastics was expected to remain low at 2%, mainly driven by upcoming market regulations as well as increased consumer demand for sustainable products. If bioplastics received subsidies and political support, global growth could reach 10–20% [
152]. The bioplastics market was expected to grow at an annual rate of 7.1% until 2026 and the revenue from bioplastics was expected to reach $4.4 billion by 2026 [
153]. The growing interest of some major organizations and chemical companies in developing and producing sustainable plastics expanded the bioplastics market in the future [
154]. As shown in , transforming plastic waste into bioplastics through biological methods contributed to a more sustainable method on plastic production and usage. Initially, plastic waste was crushed into polymers through machine processing. The polymers were then biodegraded by enzymes into synthetic monomers, which can be synthesized into bioplastics such as PHA through polymerization. PHA can be reused in food packaging, clothing industry and other fields after processing. This process demonstrated the potential for the reuse of plastic waste.
The conversion of PET into bioplastic PHA was currently being investigated to address plastic pollution and promote sustainable plastic production.
P. umsongensis GO16 can use TPA as the sole carbon source for the synthesis of both mcl-PHA and short chain length PHA (scl-PHA). Tiso et al. engineered
P. umsongensis GO16 by adaptive laboratory evolution, which allowed
P. umsongensis GO16 to use EG as sole carbon source and TPA for PHA synthesis, the PHA concentration peaked at 0.15 g/L [
145,
155]. Dou et al. proposed a hybrid upcycling approach to converting mixed PET/PLA. They used a biocompatible ionic liquid (IL) to chemically depolymerize plastic and then converted the depolymerized stream via biological upgrading. More than 95% of mixed PET/PLA was depolymerized into the respective monomers, which then served as the sole carbon source for the growth of
Pseudomonas putida, enabling the conversion of the depolymerized plastics into PHA [
156]. Many non-biodegradable plastics (PE, PP, PS) can also be transformed into bioplastics PHA. Plastics were depolymerized and converted into small monomers, which were utilized for the conversion to PHA via fermentation process [
157].
Pseudomonas aeruginosa PAO-1 accumulated PHA with almost 25% of the cell dry weight as mcl-PHA when supplied with the PE pyrolysis wax in the presence of rhamnolipids. This was the first time that PE was employed as an initial substance for the manufacturing of a biodegradable polymer [
158]. Johnston et al. investigated the molecular structure of the PHA that produced via
Cupriavidus necator H16.
Cupriavidus necator H16 consumed oxidized PP waste to produce PHA tert-polymer, the ESI-MS/MS analysis confirmed the result [
159].
Some research indicated that the products of plastic degradation can be utilized for plastic repolymerization. Li et al. synthesized 554 mg virgin PET directly from the chemical-enzymatic degradation PET solution and completed a closed-loop PET recycling. The entire cycle from degradation to polymerization can be completed within a few days [
160]. Youngpreda et al. conducted the re-polymerization of PLA. The protease produced by
Actinomadura keratinilytica strain T16-1 was used to degrade PLA, and the degradable products lactic acid were re-polymerized by using commercial lipase as a catalyst under a nitrogen atmosphere for 6 h [
161]. In addition, circular biological recycling of PHB has been reported. Chemical or enzymatic degradation of PHB can produce crotonic acid (CA), 3-hydoxybutyrate methyl ester (3HMB) and 3-hydroxybutyrate (3HB).
Paraburkholderia sacchari can utilize 3HB to produce new PHB. Sequential addition of 3HB led to 6.5 g/L total dry weight and 2.5 g/L PHB with 0.26 g/g yield, which was slightly higher than the respective values achieved on glucose [
162].
Figure 6. The cycle from plastic to bioplastic.
6. Conclusion and Future Perspectives
The increasing waste and impact caused by plastics have become significant concerns. Converting plastic waste into value-added compounds can greatly alleviate the current accumulation of plastic waste. The design and modification of hydrolase play a crucial role in the degradation of plastics and further conversion process. Currently, the methods for modifying hydrolases involve rational protein design, directed evolution, and random mutagenesis [
163]. New artificial intelligence techniques such as machine learning and deep learning are becoming increasingly important in this field. Machine learning can effectively analyze large datasets and hold great promise for accelerating the development of new enzymes. It has been widely applied in enzyme engineering to alleviate the burden of high-throughput experiments [
164]. Some machine learning methods have been employed to improve enzyme stability, catalytic activity, and thermostability [
165]. Moreover, in the high-value conversion process, the construction of new synthetic pathways also requires machine learning for the exploration of novel enzymes. Some new methods which can convert degradation products have been studied. Electrochemical upcycling has recently emerged as a new alternative, which enables precise control of chemical reactions. Electrocatalytic treatments involve chemical reactions that facilitate electron transfer through the electrode/electrolyte interface driven by electrical energy [
166].
Tenebrio molitor's abundant gut microbiota makes it a promising solution for plastic degradation and the biological retrieval of PHA [
167].
The continuous conversion of plastics into value-added compounds avoids the depletion of intermediate degradation products, thereby enhancing conversion efficiency. Currently, the most effective methods involve the combination of chemical and biological approaches or co-cultivation techniques. The combination of chemical and biological approaches involves the modification of degradative enzymes and design of conversion process. Biosensors can be used to optimize the combination of chemical and biological approaches. On the one hand, biosensors have been designed to identify hydrolysis products, thereby screening efficient hydrolase and optimizing the biodegradation [
71]. On the other hand, biosensors can be used to control metabolic pathways and reconstruct the conversion circuits [
168]. Coupling the sensing elements of biosensors with downstream responsive gene circuits enables the dynamic control of plastic conversion. Besides, the co-culture of microorganisms provides an optimal catalytic environment for continuous conversion of plastics, but the incompatibility between different species may break the balance between biodegradation and bioconversion. Computational modelling can be used to balance degradation and conversion process by designing metabolic network. In addition, several technologies, including genome engineering, macro-proteomics, and macro-metabolomics contribute to the co-culture of microorganisms. In summary, the high value conversion of plastics will be further improved through the development of a range of tools and the combination of technologies.
Acknowledgments
The authors thank the Hebei Natural Science Foundation (grant no. B2023105008), the National Natural Science Foundation of China (grant no. 22278033), and the Biological & Medical Engineering Core Facilities (Beijing Institute of Technology).
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Funding
This research received no external funding.
Declaration of competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Geyer R, Jambeck JR, Law KL. Production, use, and fate of all plastics ever made.
Sci. Adv. 2017,
3, e1700782.
[Google Scholar]
-
2.
Andrady AL, Neal MA. Applications and societal benefits of plastics.
Philosoph. Trans. Royal Soc. B Bio. Sci. 2009,
364, 1977–1984.
[Google Scholar]
-
3.
-
4.
Wright SL, Kelly FJ. Plastic and Human Health: A Micro Issue?
Environ. Sci. Tec. 2017,
51, 6634–6647.
[Google Scholar]
-
5.
Tamoor M, Samak NA, Yang M, Xing J. The Cradle-to-Cradle Life Cycle Assessment of Polyethylene terephthalate: Environmental Perspective.
Molecules 2022,
27, 1599.
[Google Scholar]
-
6.
Jatoi AS, Ahmed J, Mubarak NM, Shaikh MS, Kumar S, Ahmed MI, et al. A Comprehensive Review on Handling of Plastic Waste For Energy Generation—Current Status and Future Challenges.
Water Air Soil Pollu. 2023,
234, 773.
[Google Scholar]
-
7.
Huang Q, Chen G, Wang Y, Chen S, Xu L, Wang R. Modelling the global impact of China’s ban on plastic waste imports.
Res. Conser. Recycl. 2020,
154, 104607.
[Google Scholar]
-
8.
Jung YS, Sampath V, Prunicki M, Aguilera J, Allen H, LaBeaud D, et al. Characterization and regulation of microplastic pollution for protecting planetary and human health.
Environ. Pollu. 2022,
315, 120442.
[Google Scholar]
-
9.
Banu JR, Sharmila VG. A systematic review on plastic waste conversion for a circular economy: recent trends and emerging technologies.
Catal. Sci. Tec. 2023,
13, 2291–2302.
[Google Scholar]
-
10.
Weckhuysen BM. Creating value from plastic waste.
Science 2020,
370, 400–401.
[Google Scholar]
-
11.
Lackner M, Mukherjee A, Koller M. What Are “Bioplastics”? Defining Renewability, Biosynthesis, Biodegradability, and Biocompatibility.
Polymers 2023,
15, 4695.
[Google Scholar]
-
12.
Gupta PK, Gahtori R, Govarthanan K, Sharma V, Pappuru S, Pandit S, et al. Recent trends in biodegradable polyester nanomaterials for cancer therapy.
Mater. Sci. Eng. C 2021,
127, 112198.
[Google Scholar]
-
13.
Huneault MA. Special Issue on Biobased Polymers.
Inter. Polym. Proc. 2007,
22, 384–384.
[Google Scholar]
-
14.
Song X, Wu X, Song X, Shi C, Zhang Z. Sorption and desorption of petroleum hydrocarbons on biodegradable and nondegradable microplastics.
Chemosphere 2021,
273, 128553.
[Google Scholar]
-
15.
Valenzuela-Ortega M, Suitor JT, White MFM, Hinchcliffe T, Wallace S. Microbial Upcycling of Waste PET to Adipic Acid.
ACS Central Sci. 2023,
9, 2057–2063.
[Google Scholar]
-
16.
Tamoor M, Samak NA, Jia Y, Mushtaq MU, Sher H, Bibi M, et al. Potential Use of Microbial Enzymes for the Conversion of Plastic Waste Into Value-Added Products: A Viable Solution.
Front. Microb. 2021,
12, 777727.
[Google Scholar]
-
17.
Pandit AV, Harrison E, Mahadevan R. Engineering Escherichia coli for the utilization of ethylene glycol.
Micro.Cell Fact. 2021,
20, 22.
[Google Scholar]
-
18.
Flury M, Narayan R. Biodegradable plastic as an integral part of the solution to plastic waste pollution of the environment.
Curr.Op. Green Sust. Chem. 2021,
30, 100490.
[Google Scholar]
-
19.
Mannina G, Presti D, Montiel-Jarillo G, Carrera J, Suárez-Ojeda ME. Recovery of polyhydroxyalkanoates (PHAs) from wastewater: A review.
Biore. Tech. 2020,
297, 122478.
[Google Scholar]
-
20.
Choi SY, Lee Y, Yu HE, Cho IJ, Kang M, Lee SY. Sustainable production and degradation of plastics using microbes.
Nat. Micro. 2023,
8, 2253–2276.
[Google Scholar]
-
21.
Zhou W, Bergsma S, Colpa DI, Euverink G-JW, Krooneman J. Polyhydroxyalkanoates (PHAs) synthesis and degradation by microbes and applications towards a circular economy.
J. Env. Manag. 2023,
341, 118033.
[Google Scholar]
-
22.
Focarete ML, Ceccorulli G, Scandola M, Kowalczuk M. Further Evidence of Crystallinity-Induced Biodegradation of Synthetic Atactic Poly(3-hydroxybutyrate) by PHB-Depolymerase A from Pseudomonas lemoignei. Blends of Atactic Poly(3-hydroxybutyrate) with Crystalline Polyesters.
Macromolecules 1998,
31, 8485–8492.
[Google Scholar]
-
23.
Allen AD, Anderson WA, Ayorinde F, Eribo BE. Isolation and characterization of an extracellular thermoalkanophilic P(3HB-co-3HV) depolymerase from Streptomyces sp. IN1.
Inter. Bio. Biodeg. 2011,
65, 777–785.
[Google Scholar]
-
24.
Mukai K, Yamada K, Doi Y. Enzymatic degradation of poly(hydroxyalkanoates) by a marine bacterium.
Pol. Deg. Stab. 1993,
41, 85–91.
[Google Scholar]
-
25.
Iyer S, Shah R, Sharma A, Jendrossek D, Desai A. Purification of Aspergillus fumigatus (Pdf1) Poly(β-hydroxybutyrate) (PHB) Depolymerase Using a New, Single-Step Substrate Affinity Chromatography Method: Characterization of the PHB Depolymerase Exhibiting Novel Self-Aggregation Behavior.
J. Poly. Env. 2000,
8, 197–203.
[Google Scholar]
-
26.
Shivakumar S, Jagadish SJ, Zatakia H, Dutta J. Purification, Characterization and Kinetic Studies of a Novel Poly(β) Hydroxybutyrate (PHB) Depolymerase PhaZPenfrom Penicillium citrinum S2.
App. Biochem. Biotec. 2011,
164, 1225–1236.
[Google Scholar]
-
27.
Mergaert J, Anderson C, Wouters A, Swings J. Microbial degradation of poly(3-hydroxybutyrate) and poly(3-hydroxybutyrate-co-3-hydroxyvalerate) in compost.
J. Env. Poly. Degrada. 1994,
2, 177–183.
[Google Scholar]
-
28.
Meereboer KW, Misra M, Mohanty AK. Review of recent advances in the biodegradability of polyhydroxyalkanoate (PHA) bioplastics and their composites.
Green Chem. 2020,
22, 5519–5558.
[Google Scholar]
-
29.
Janssen PH, Harfoot CG. Ilyobacter delafieldii sp. nov., a metabolically restricted anaerobic bacterium fermenting PHB.
Arch. Micro. 1990,
154, 253–259.
[Google Scholar]
-
30.
Abou-Zeid D-M, Müller R-J, Deckwer W-D. Biodegradation of Aliphatic Homopolyesters and Aliphatic−Aromatic Copolyesters by Anaerobic Microorganisms.
Biomacromolecules 2004,
5, 1687–1697.
[Google Scholar]
-
31.
Wang S, Lydon KA, White EM, Grubbs JB, III, Lipp EK, Locklin J, et al. Biodegradation of Poly(3-hydroxybutyrate-co-3-hydroxyhexanoate) Plastic under Anaerobic Sludge and Aerobic Seawater Conditions: Gas Evolution and Microbial Diversity.
Envir. Sci. Tech. 2018,
52, 5700–5709.
[Google Scholar]
-
32.
Boey JY, Mohamad L, Khok YS, Tay GS, Baidurah S. A Review of the Applications and Biodegradation of Polyhydroxyalkanoates and Poly(lactic acid) and Its Composites.
Polymer 2021,
13, 1544.
[Google Scholar]
-
33.
Martínez V, de Santos PG, García-Hidalgo J, Hormigo D, Prieto MA, Arroyo M, et al. Novel extracellular medium-chain-length polyhydroxyalkanoate depolymerase from Streptomyces exfoliatus K10 DSMZ 41693: a promising biocatalyst for the efficient degradation of natural and functionalized mcl-PHAs.
App. Micro. Biotech. 2015,
99, 9605–9615.
[Google Scholar]
-
34.
Zhou W, Colpa DI, Permentier H, Offringa RA, Rohrbach L, Euverink G-JW, et al. Insight into polyhydroxyalkanoate (PHA) production from xylose and extracellular PHA degradation by a thermophilic Schlegelella thermodepolymerans.
Res. Con. Recyc. 2023,
194, 107006.
[Google Scholar]
-
35.
Westlie AH, Quinn EC, Parker CR, Chen EYX. Synthetic biodegradable polyhydroxyalkanoates (PHAs): Recent advances and future challenges.
Prog. Poly. Sci. 2022,
134, 101608.
[Google Scholar]
-
36.
Ikura Y, Kudo T. Isolation of a microorganism capable of degrading poly-(L-lactide).
J. Gen. App. Micro. 1999,
45, 247–251.
[Google Scholar]
-
37.
Urbanek AK, Rymowicz W, Strzelecki MC, Kociuba W, Franczak Ł, Mirończuk AM. Isolation and characterization of Arctic microorganisms decomposing bioplastics.
AMB Exp. 2017,
7, 148.
[Google Scholar]
-
38.
Sakai K, Kawano H, Iwami A, Nakamura M, Moriguchi M. Isolation of a thermophilic poly-l-lactide degrading bacterium from compost and its enzymatic characterization.
J. Biosci. Bioeng. 2001,
92, 298–300.
[Google Scholar]
-
39.
Suzuki K, Sakamoto H, Shinozaki Y, Tabata J, Watanabe T, Mochizuki A, et al. Affinity purification and characterization of a biodegradable plastic-degrading enzyme from a yeast isolated from the larval midgut of a stag beetle, Aegus laevicollis.
App. Micro. Biotech. 2013,
97, 7679–7688.
[Google Scholar]
-
40.
Tournier V, Duquesne S, Guillamot F, Cramail H, Taton D, Marty A, et al. Enzymes’ Power for Plastics Degradation.
Chem. Rev. 2023,
123, 5612–5701.
[Google Scholar]
-
41.
Lee SH, Kim IY, Song WS. Biodegradation of polylactic acid (PLA) fibers using different enzymes.
Macro. Res. 2014,
22, 657–663.
[Google Scholar]
-
42.
Cui L, Wang X, Szarka G, Hegyesi N, Wang Y, Sui X, et al. Quantitative analysis of factors determining the enzymatic degradation of poly(lactic acid).
Inter. J. Biolog. Macro. 2022,
209, 1703–1709.
[Google Scholar]
-
43.
Myburgh MW, Favaro L, van Zyl WH, Viljoen-Bloom M. Engineered yeast for the efficient hydrolysis of polylactic acid.
Biores. Tech. 2023,
378, 129008.
[Google Scholar]
-
44.
Rajgond V, Mohite A, More N, More A. Biodegradable polyester-polybutylene succinate (PBS): a review.
Poly. Bull. 2023, ,
81, 5703-5752.
[Google Scholar]
-
45.
Tokiwa Y, Jarerat A. Microbial degradation of aliphatic polyesters.
Macro. Sym. 2003,
201, 283–290.
[Google Scholar]
-
46.
Kim SH, Cho JY, Cho DH, Jung HJ, Kim BC, Bhatia SK, et al. Acceleration of Polybutylene Succinate Biodegradation by Terribacillus sp. JY49 Isolated from a Marine Environment.
Polymers 2022,
14, 3978.
[Google Scholar]
-
47.
kanwal A, Zhang M, Sharaf F. Synergistic degradation of PBAT poly (butylene adipate-co-terephthalate) co-polyesters using a bacterial co-culture system.
Poly. Bull. 2024,
81, 2741–2755.
[Google Scholar]
-
48.
Roy S, Ghosh T, Zhang W, Rhim J-W. Recent progress in PBAT-based films and food packaging applications: A mini-review.
Food Chem. 2024,
437, 137822.
[Google Scholar]
-
49.
Jang Y, Kim M, Kim Y, Yu J, Kim S-K, Han J, et al. Enhancing biodegradation of PBAT through bio-stimulation using Pseudozyma jejuensis for effective plastic waste reduction.
Chemosphere 2023,
340, 139867.
[Google Scholar]
-
50.
Jia H, Zhang M, Weng Y, Zhao Y, Li C, Kanwal A. Degradation of poly(butylene adipate-co-terephthalate) by Stenotrophomonas sp. YCJ1 isolated from farmland soil.
J. Envir. Sci. 2021,
103, 50–58.
[Google Scholar]
-
51.
Aarthy M, Puhazhselvan P, Aparna R, George AS, Gowthaman MK, Ayyadurai N, et al. Growth associated degradation of aliphatic-aromatic copolyesters by Cryptococcus sp. MTCC 5455.
Poly. Degrad. Stab. 2018,
152, 20–28.
[Google Scholar]
-
52.
Trinh Tan F, Cooper DG, Marić M, Nicell JA. Biodegradation of a synthetic co-polyester by aerobic mesophilic microorganisms.
Poly. Degrad. Stab. 2008,
93, 1479–1485.
[Google Scholar]
-
53.
Jia X, Zhao K, Zhao J, Lin C, Zhang H, Chen L, et al. Degradation of poly(butylene adipate-co-terephthalate) films by Thermobifida fusca FXJ-1 isolated from compost.
J. Hazard. Mater. 2023,
441, 129958.
[Google Scholar]
-
54.
Yang Y, Ko T-P, Liu L, Li J, Huang C-H, Chen J, et al. Roles of tryptophan residue and disulfide bond in the variable lid region of oxidized polyvinyl alcohol hydrolase.
Biochem. Biophy. Res. Com. 2014,
452, 509–514.
[Google Scholar]
-
55.
Yang Y, Ko T-P, Liu L, Li J, Huang C-H, Chan H-C, et al. Structural Insights into Enzymatic Degradation of Oxidized Polyvinyl Alcohol.
ChemBioChem 2014,
15, 1882–1886.
[Google Scholar]
-
56.
Yang Y, Min J, Xue T, Jiang P, Liu X, Peng R, et al. Complete bio-degradation of poly(butylene adipate-co-terephthalate) via engineered cutinases.
Nat. Com. 2023,
14, 1645.
[Google Scholar]
-
57.
Yang Y, Cheng S, Zheng Y, Xue T, Huang J-W, Zhang L, et al. Remodeling the polymer-binding cavity to improve the efficacy of PBAT-degrading enzyme.
J. Hazard. Mater. 2024,
464, 132965.
[Google Scholar]
-
58.
Shin S-M, Jha RK, Dale T. Tackling the Catch-22 Situation of Optimizing a Sensor and a Transporter System in a Whole-Cell Microbial Biosensor Design for an Anthropogenic Small Molecule.
ACS Syn. Bio. 2022,
11, 3996–4008.
[Google Scholar]
-
59.
Shojaei B, Abtahi M, Najafi M. Chemical recycling of PET: A stepping-stone toward sustainability.
Poly. Adv. Technol. 2020,
31, 2912–2938.
[Google Scholar]
-
60.
Danso D, Schmeisser C, Chow J, Zimmermann W, Wei R, Leggewie C, et al. New Insights into the Function and Global Distribution of Polyethylene Terephthalate (PET)-Degrading Bacteria and Enzymes in Marine and Terrestrial Metagenomes.
App. Env. Micro. 2018,
84, e02773–17.
[Google Scholar]
-
61.
Tournier V, Topham CM, Gilles A, David B, Folgoas C, Moya-Leclair E, et al. An engineered PET depolymerase to break down and recycle plastic bottles.
Nature 2020,
580, 216–219.
[Google Scholar]
-
62.
Zhang Z, Huang S, Cai D, Shao C, Zhang C, Zhou J, et al. Depolymerization of post-consumer PET bottles with engineered cutinase 1 from Thermobifida cellulosilytica.
Green Chem. 2022,
24, 5998–6007.
[Google Scholar]
-
63.
Yoshida S, Hiraga K, Takehana T, Taniguchi I, Yamaji H, Maeda Y, et al. A bacterium that degrades and assimilates poly(ethylene terephthalate).
Science 2016,
351, 1196–1199.
[Google Scholar]
-
64.
Knott BC, Erickson E, Allen MD, Gado JE, Graham R, Kearns FL, et al. Characterization and engineering of a two-enzyme system for plastics depolymerization.
Proc. Nat. Aca. Sci. 2020,
117, 25476–25485.
[Google Scholar]
-
65.
Cui Y, Chen Y, Liu X, Dong S, Tian Ye, Qiao Y, et al. Computational Redesign of a PETase for Plastic Biodegradation under Ambient Condition by the GRAPE Strategy.
ACS Catal. 2021,
11, 1340–1350.
[Google Scholar]
-
66.
Liu Y, Liu Z, Guo Z, Yan T, Jin C, Wu J. Enhancement of the degradation capacity of IsPETase for PET plastic degradation by protein engineering.
Sci. Total Env. 2022,
834, 154947.
[Google Scholar]
-
67.
Shi L, Liu P, Tan Z, Zhao W, Gao J, Gu Q, et al. Complete Depolymerization of PET Wastes by an Evolved PET Hydrolase from Directed Evolution.
Angewandte Chem. Inter. Ed. 2023,
62, e202218390.
[Google Scholar]
-
68.
Hong H, Ki D, Seo H, Park J, Jang J, Kim K-J. Discovery and rational engineering of PET hydrolase with both mesophilic and thermophilic PET hydrolase properties.
Nat. Com. 2023,
14, 4556.
[Google Scholar]
-
69.
Jia Y, Samak NA, Hao X, Chen Z, Wen Q, Xing J. Hydrophobic cell surface display system of PETase as a sustainable biocatalyst for PET degradation.
Front. Microb. 2022,
13, 1005480.
[Google Scholar]
-
70.
Jia Y, Samak NA, Hao X, Chen Z, Yang G, Zhao X, et al. Nano-immobilization of PETase enzyme for enhanced polyethylene terephthalate biodegradation.
Biochem. Eng. J. 2021,
176, 108205.
[Google Scholar]
-
71.
Li J, Nina MRH, Zhang X, Bai Y. Engineering Transcription Factor XylS for Sensing Phthalic Acid and Terephthalic Acid: An Application for Enzyme Evolution.
ACS Syn. Bio. 2022,
11, 1106–1113.
[Google Scholar]
-
72.
Wei R, Breite D, Song C, Gräsing D, Ploss T, Hille P, et al. Biocatalytic Degradation Efficiency of Postconsumer Polyethylene Terephthalate Packaging Determined by Their Polymer Microstructures.
Adv. Sci. 2019,
6, 1900491.
[Google Scholar]
-
73.
Schubert SW, Thomsen TB, Clausen KS, Malmendal A, Hunt CJ, Borch K, et al. Relationships of crystallinity and reaction rates for enzymatic degradation of poly (ethylene terephthalate), PET.
ChemSusChem 2024,
, e202301752.
[Google Scholar]
-
74.
Carr CM, Clarke DJ, Dobson ADW. Microbial Polyethylene Terephthalate Hydrolases: Current and Future Perspectives.
Front. Microb. 2020,
11, 571265.
[Google Scholar]
-
75.
Mohanan N, Montazer Z, Sharma PK, Levin DB. Microbial and Enzymatic Degradation of Synthetic Plastics.
Front. Microb. 2020,
11, 580709.
[Google Scholar]
-
76.
Ghatge S, Yang Y, Ahn J-H, Hur H-G. Biodegradation of polyethylene: a brief review.
App. Biolog. Chem. 2020,
63, 27.
[Google Scholar]
-
77.
Seo M-J, Yun S-D, Kim H-W, Yeom S-J. Polyethylene-biodegrading Microbes and Their Future Directions.
Biotech. Bioproc. Eng. 2023,
28, 977–989.
[Google Scholar]
-
78.
Peixoto J, Vizzotto C, Ramos A, Alves G, Steindorff A, Krüger R. The role of nitrogen metabolism on polyethylene biodegradation.
J. Hazard. Mater. 2022,
432, 128682.
[Google Scholar]
-
79.
Arkatkar A, Juwarkar AA, Bhaduri S, Uppara PV, Doble M. Growth of Pseudomonas and Bacillus biofilms on pretreated polypropylene surface.
Inter. Biodet. Biodeg. 2010,
64, 530–536.
[Google Scholar]
-
80.
Taghavi N, Udugama IA, Zhuang W-Q, Baroutian S. Challenges in biodegradation of non-degradable thermoplastic waste: From environmental impact to operational readiness.
Biotech. Adv. 2021,
49, 107731.
[Google Scholar]
-
81.
Shimpi N, Borane M, Mishra S, Kadam M. Biodegradation of polystyrene (PS)-poly(lactic acid) (PLA) nanocomposites using Pseudomonas aeruginosa.
Macro. Res. 2012,
20, 181–187.
[Google Scholar]
-
82.
Motta O, Proto A, De Carlo F, De Caro F, Santoro E, Brunetti L, et al. Utilization of chemically oxidized polystyrene as co-substrate by filamentous fungi.
Inter. J. Hyg. Env. Heal. 2009,
212, 61–66.
[Google Scholar]
-
83.
Mor R, Sivan A. Biofilm formation and partial biodegradation of polystyrene by the actinomycete Rhodococcus ruber.
Biodegradation 2008,
19, 851–858.
[Google Scholar]
-
84.
Auta HS, Emenike CU, Fauziah SH. Screening of Bacillus strains isolated from mangrove ecosystems in Peninsular Malaysia for microplastic degradation.
Env. Pollu. 2017,
231, 1552–1559.
[Google Scholar]
-
85.
Cacciari I, Quatrini P, Zirletta G, Mincione E, Vinciguerra V, Lupattelli P, et al. Isotactic polypropylene biodegradation by a microbial community: physicochemical characterization of metabolites produced.
App. Env. Micro. 1993,
59, 3695–3700.
[Google Scholar]
-
86.
Patinge KP, Zodpe SN. Screening of Low Density Polyethylene Degrading Microflora From Garbage Soil.
Inter. J. Sci. Res. Sci. Tech. 2020,
7, 110–116.
[Google Scholar]
-
87.
Peixoto J, Silva LP, Krüger RH. Brazilian Cerrado soil reveals an untapped microbial potential for unpretreated polyethylene biodegradation.
J. Hazard. Mater. 2017,
324, 634–644.
[Google Scholar]
-
88.
Giacomucci L, Raddadi N, Soccio M, Lotti N, Fava F. Polyvinyl chloride biodegradation by Pseudomonas citronellolis and Bacillus flexus.
New Biotech. 2019,
52, 35–41.
[Google Scholar]
-
89.
Lu X, Yao Y, Yang Y, Zhang Z, Gu J, Mojovic L, et al. Ethylene glycol and glycolic acid production by wild-type Escherichia coli.
Biotech. App. Biochem. 2021,
68, 744–755.
[Google Scholar]
-
90.
Marchesan AN, Oncken MP, Maciel Filho R, Wolf Maciel MR. A roadmap for renewable C2–C3 glycols production: a process engineering approach.
Green Chem. 2019,
21, 5168–5194.
[Google Scholar]
-
91.
Román-Camacho JJ, Santos-Dueñas IM, García-García I, Moreno-García J, García-Martínez T, Mauricio JC. Metaproteomics of microbiota involved in submerged culture production of alcohol wine vinegar: A first approach.
Inter. J. Food Micro. 2020,
333, 108797.
[Google Scholar]
-
92.
Hua X, Zhou X, Xu Y. Improving techno-economics of bioproduct glycolic acid by successive recycled-cell catalysis of ethylene glycol with Gluconobacter oxydans.
Bioproc. Biosys. Eng. 2018,
41, 1555–1559.
[Google Scholar]
-
93.
Carniel A, Santos AG, Chinelatto Júnior LS, Castro AM, Coelho MAZ. Biotransformation of ethylene glycol to glycolic acid by Yarrowia lipolytica: A route for poly(ethylene terephthalate) (PET) upcycling.
Biotech. J. 2023,
18, 2200521.
[Google Scholar]
-
94.
Mückschel B, Simon O, Klebensberger J, Graf N, Rosche B, Altenbuchner J, et al. Ethylene Glycol Metabolism by Pseudomonas putida.
App. Env. Micro. 2012,
78, 8531–8539.
[Google Scholar]
-
95.
Wehrmann M, Billard P, Martin-Meriadec A, Zegeye A, Klebensberger J. Functional Role of Lanthanides in Enzymatic Activity and Transcriptional Regulation of Pyrroloquinoline Quinone-Dependent Alcohol Dehydrogenases in Pseudomonas putida KT2440.
mBio 2017,
8, 1128.
[Google Scholar]
-
96.
Franden MA, Jayakody LN, Li W-J, Wagner NJ, Cleveland NS, Michener WE, et al. Engineering Pseudomonas putida KT2440 for efficient ethylene glycol utilization.
Metab. Eng. 2018,
48, 197–207.
[Google Scholar]
-
97.
Aziz RK, Monk JM, Andrews KA, Nhan J, Khaw VL, Wong H, et al. The aldehyde dehydrogenase, AldA, is essential for L-1, 2-propanediol utilization in laboratory-evolved Escherichia coli.
Microb. Res. 2017,
194, 47–52.
[Google Scholar]
-
98.
Zheng H, Wang X, Yomano Lorraine P, Geddes Ryan D, Shanmugam Keelnatham T, Ingram Lonnie O. Improving Escherichia coli FucO for Furfural Tolerance by Saturation Mutagenesis of Individual Amino Acid Positions.
App. Env. Micro. 2013,
79, 3202–3208.
[Google Scholar]
-
99.
Lu Z, Cabiscol E, Obradors N, Tamarit J, Ros J, Aguilar J, et al. Evolution of an Escherichia coli Protein with Increased Resistance to Oxidative Stress *.
J. Biolog. Chem. 1998,
273, 8308–8316.
[Google Scholar]
-
100.
Kurpejović E, Burgardt A, Bastem GM, Junker N, Wendisch VF, Sariyar Akbulut B. Metabolic engineering of Corynebacterium glutamicum for l-tyrosine production from glucose and xylose.
J. Biotech. 2023,
363, 8–16.
[Google Scholar]
-
101.
Panda S, Zhou JFJ, Feigis M, Harrison E, Ma X, Fung Kin Yuen V, et al. Engineering Escherichia coli to produce aromatic chemicals from ethylene glycol.
Metab. Eng. 2023,
79, 38–48.
[Google Scholar]
-
102.
Frazão CJR, Wagner N, Rabe K, Walther T. Construction of a synthetic metabolic pathway for biosynthesis of 2, 4-dihydroxybutyric acid from ethylene glycol.
Nat. Com. 2023,
14, 1931.
[Google Scholar]
-
103.
Qin N, Li L, Ji X, Li X, Zhang Y, Larsson C, et al. Rewiring Central Carbon Metabolism Ensures Increased Provision of Acetyl-CoA and NADPH Required for 3-OH-Propionic Acid Production.
ACS Syn. Bio. 2020,
9, 3236–3244.
[Google Scholar]
-
104.
Zhang Q, Zeng W, Xu S, Zhou J. Metabolism and strategies for enhanced supply of acetyl-CoA in Saccharomyces cerevisiae.
Biores. Tech. 2021,
342, 125978.
[Google Scholar]
-
105.
Pietrocola F, Galluzzi L, Bravo-San Pedro José M, Madeo F, Kroemer G. Acetyl Coenzyme A: A Central Metabolite and Second Messenger.
Cell Metab. 2015,
21 , 805–821.
[Google Scholar]
-
106.
Nie M, Wang J, Zhang K. Engineering a Novel Acetyl-CoA Pathway for Efficient Biosynthesis of Acetyl-CoA-Derived Compounds.
ACS Syn. Bio. 2024,
13, 358–369.
[Google Scholar]
-
107.
Lu X, Liu Y, Yang Y, Wang S, Wang Q, Wang X, et al. Constructing a synthetic pathway for acetyl-coenzyme A from one-carbon through enzyme design.
Nat. Com. 2019,
10, 1378.
[Google Scholar]
-
108.
Yang X, Yuan Q, Luo H, Li F, Mao Y, Zhao X, et al. Systematic design and in vitro validation of novel one-carbon assimilation pathways.
Metab. Eng. 2019,
56, 142–153.
[Google Scholar]
-
109.
Wagner N, Bade F, Straube E, Rabe K, Frazão CJR, Walther T. In vivo implementation of a synthetic metabolic pathway for the carbon-conserving conversion of glycolaldehyde to acetyl-CoA.
Front. Bioeng. Biotech. 2023,
11, 1125544.
[Google Scholar]
-
110.
Xue T, Liu K, Chen D, Yuan X, Fang J, Yan H, et al. Improved bioethanol production using CRISPR/Cas9 to disrupt the ADH2 gene in Saccharomyces cerevisiae.
World J. Microb. Biotech. 2018,
34, 154.
[Google Scholar]
-
111.
Kalathil S, Miller M, Reisner E. Microbial Fermentation of Polyethylene Terephthalate (PET) Plastic Waste for the Production of Chemicals or Electricity**.
Angewandte Chem. Inter. Ed. 2022,
61, e202211057.
[Google Scholar]
-
112.
Luo ZW, Lee SY. Biotransformation of p-xylene into terephthalic acid by engineered Escherichia coli.
Nat. Com. 2017,
8, 15689.
[Google Scholar]
-
113.
Luo ZW, Choi KR, Lee SY. Improved terephthalic acid production from p-xylene using metabolically engineered Pseudomonas putida.
Metab. Eng. 2023,
76, 75–86.
[Google Scholar]
-
114.
Pardo I, Jha RK, Bermel RE, Bratti F, Gaddis M, McIntyre E, et al. Gene amplification, laboratory evolution, and biosensor screening reveal MucK as a terephthalic acid transporter in Acinetobacter baylyi ADP1.
Metab. Eng. 2020,
62, 260–274.
[Google Scholar]
-
115.
Kim HT, Kim JK, Cha HG, Kang MJ, Lee HS, Khang TU, et al. Biological Valorization of Poly(ethylene terephthalate) Monomers for Upcycling Waste PET.
ACS Sust. Chem. Eng. 2019,
7, 19396–19406.
[Google Scholar]
-
116.
Wang Q, Wu X, Lu X, He Y, Ma B, Xu Y. Efficient Biosynthesis of Vanillin from Isoeugenol by Recombinant Isoeugenol Monooxygenase from Pseudomonas nitroreducens Jin1.
App. Biochem. Biotech. 2021,
193, 1116–1128.
[Google Scholar]
-
117.
Sadler JC and Wallace S. Microbial synthesis of vanillin from waste poly(ethylene terephthalate).
Green Chem. 2021,
23, 4665–4672.
[Google Scholar]
-
118.
Luo H, Hua X, Liu W, Xu Y, Cao F, Nikafshar S, et al. Toward Utilization of Lignin-Derivatized Monomers: Development of Degradable Vanillin-Based Polyurethane Materials.
ACS App. Polym. Mater. 2023,
5, 6061–6068.
[Google Scholar]
-
119.
Bayer T, Pfaff L, Branson Y, Becker A, Wu S, Bornscheuer UT, et al. Biosensor and chemo-enzymatic one-pot cascade applications to detect and transform PET-derived terephthalic acid in living cells.
iScience 2022,
25, 104326.
[Google Scholar]
-
120.
Gopal MR, Dickey RM, Butler ND, Talley MR, Nakamura DT, Mohapatra A, et al. Reductive Enzyme Cascades for Valorization of Polyethylene Terephthalate Deconstruction Products.
ACS Catal. 2023,
13, 4778–4789.
[Google Scholar]
-
121.
Zhao M, Huang D, Zhang X, Koffas MAG, Zhou J, Deng Y. Metabolic engineering of Escherichia coli for producing adipic acid through the reverse adipate-degradation pathway.
Metab. Eng. 2018,
47, 254–262.
[Google Scholar]
-
122.
Jiang B, Wang Y, Peng Z, Lim KH, Wang Q, Shi S, et al. Synthesis of Poly(butylene adipate terephthalate)-co-poly(glycolic acid) with Enhanced Degradability in Water.
Macromolecules 2023,
56, 9207–9217.
[Google Scholar]
-
123.
Parke D, Garcia MA, Ornston LN. Cloning and Genetic Characterization of dca Genes Required for β-Oxidation of Straight-Chain Dicarboxylic Acids in Acinetobacter sp. Strain ADP1.
App. Env. Micro. 2001,
67, 4817–4827.
[Google Scholar]
-
124.
Fedorchuk TP, Khusnutdinova AN, Evdokimova E, Flick R, Di Leo R, Stogios P, et al. One-Pot Biocatalytic Transformation of Adipic Acid to 6-Aminocaproic Acid and 1, 6-Hexamethylenediamine Using Carboxylic Acid Reductases and Transaminases.
J. Am. Chem. Soc. 2020,
142, 1038–1048.
[Google Scholar]
-
125.
Shi K, Li J-M, Zhang Z-J, Chen Q, Xu J-H, Yu H-L. Virtual screening of carboxylic acid reductases for biocatalytic synthesis of 6-aminocaproic acid and 1, 6-hexamethylenediamine.
Biotech. Bioeng. 2023,
120, 1773–1783.
[Google Scholar]
-
126.
Ackermann YS, Li W-J, Op de Hipt L, Niehoff P-J, Casey W, Polen T, et al. Engineering adipic acid metabolism in Pseudomonas putida.
Metab. Eng. 2021,
67, 29–40.
[Google Scholar]
-
127.
Singhvi MS, Zinjarde SS, Gokhale DV. Polylactic acid: synthesis and biomedical applications.
J. App. Microb. 2019,
127, 1612–1626.
[Google Scholar]
-
128.
Paswan M, Adhikary S, Salama HH, Rusu AV, Zuorro A, Dholakiya BZ, et al. Microbial Synthesis of Lactic Acid from Cotton Stalk for Polylactic Acid Production.
Microorganisms 2023,
11, 1931.
[Google Scholar]
-
129.
Li G, Zhao M, Xu F, Yang B, Li X, Meng X, et al. Synthesis and Biological Application of Polylactic Acid.
Molecules 2020,
25 , 5023.
[Google Scholar]
-
130.
Kucek LA, Nguyen M, Angenent LT. Conversion of l-lactate into n-caproate by a continuously fed reactor microbiome.
Water Res. 2016,
93, 163–171.
[Google Scholar]
-
131.
Baur T, Wentzel A, Dürre P. Production of propionate using metabolically engineered strains of Clostridium saccharoperbutylacetonicum.
App. Micro. Biotech. 2022,
106, 7547–7562.
[Google Scholar]
-
132.
Li C, Ong KL, Cui Z, Sang Z, Li X, Patria RD, et al. Promising advancement in fermentative succinic acid production by yeast hosts.
J. Hazard. Mater. 2021,
401, 123414.
[Google Scholar]
-
133.
Cui Z, Zhong Y, Sun Z, Jiang Z, Deng J, Wang Q, et al. Reconfiguration of the reductive TCA cycle enables high-level succinic acid production by Yarrowia lipolytica.
Nat. Com. 2023,
14, 8480.
[Google Scholar]
-
134.
Cui Z, Zhu Z, Zhang J, Jiang Z, Liu Y, Wang Q, et al. Efficient 5-aminolevulinic acid production through reconstructing the metabolic pathway in SDH-deficient Yarrowia lipolytica.
Biochem. Eng. J. 2021,
174, 108125.
[Google Scholar]
-
135.
Cheng J, Li J, Zheng L. Achievements and Perspectives in 1, 4-Butanediol Production from Engineered Microorganisms.
J. Agr. Food Chem. 2021,
69, 10480–10485.
[Google Scholar]
-
136.
Wu M-Y, Sung L-Y, Li H, Huang C-H, Hu Y-C. Combining CRISPR and CRISPRi Systems for Metabolic Engineering of E. coli and 1, 4-BDO Biosynthesis.
ACS Syn. Bio. 2017,
6, 2350–2361.
[Google Scholar]
-
137.
Li W-J, Narancic T, Kenny ST, Niehoff P-J, O’Connor K, Blank LM, et al. Unraveling 1, 4-Butanediol Metabolism in Pseudomonas putida KT2440.
Front. Microb. 2020,
11, 382.
[Google Scholar]
-
138.
Fujiwara R, Noda S, Tanaka T, Kondo A. Muconic Acid Production Using Gene-Level Fusion Proteins in Escherichia coli.
ACS Syn. Bio. 2018,
7, 2698–2705.
[Google Scholar]
-
139.
Liu P, Zheng Y, Yuan Y, Zhang T, Li Q, Liang Q, et al. Valorization of Polyethylene Terephthalate to Muconic Acid by Engineering Pseudomonas Putida.
Inter. J. Mol. Sci. 2022,
23, 10997.
[Google Scholar]
-
140.
Watcharawipas A, Sansatchanon K, Phithakrotchanakoon C, Tanapongpipat S, Runguphan W, Kocharin K. Novel carotenogenic gene combinations from red yeasts enhanced lycopene and beta-carotene production in Saccharomyces cerevisiae from the low-cost substrate sucrose.
FEMS Yeast Res. 2021,
21, foab062.
[Google Scholar]
-
141.
Diao J, Hu Y, Tian Y, Carr R, Moon TS. Upcycling of poly(ethylene terephthalate) to produce high-value bio-products.
Cell Rep. 2023,
42, 111908.
[Google Scholar]
-
142.
Kim DH, Han DO, In Shim K, Kim JK, Pelton JG, Ryu MH, et al. One-Pot Chemo-bioprocess of PET Depolymerization and Recycling Enabled by a Biocompatible Catalyst, Betaine.
ACS Catal. 2021,
11, 3996–4008.
[Google Scholar]
-
143.
Kim HT, Hee Ryu M, Jung YJ, Lim S, Song HM, Park J, et al. Chemo-Biological Upcycling of Poly(ethylene terephthalate) to Multifunctional Coating Materials.
ChemSusChem 2021,
14, 4251–4259.
[Google Scholar]
-
144.
Welsing G, Wolter B,
Hintzen HMT, Tiso T, Blank LM. Chapter Eighteen - Upcycling of hydrolyzed PET by microbial conversion
to a fatty acid derivative. Methods In Enzymology; Elsevier: Gainesville,
FL, USA, 2021; pp. 391–421
-
145.
Tiso T, Narancic T, Wei R, Pollet E, Beagan N, Schröder K, et al. Towards bio-upcycling of polyethylene terephthalate.
Metab. Eng. 2021,
66, 167–178.
[Google Scholar]
-
146.
Tian S, Jiao Y, Gao Z, Xu Y, Fu L, Fu H, et al. Catalytic Amination of Polylactic Acid to Alanine.
J. Am. Chem. Soc. 2021,
143, 16358–16363.
[Google Scholar]
-
147.
Jiao Y, Wang M, Ma D. Catalytic Cracking of Polylactic Acid to Acrylic Acid.
Chin. J. Chem. 2023,
41, 2071–2076.
[Google Scholar]
-
148.
Liu P, Zheng Y, Yuan Y, Han Y, Su T, Qi Q. Upcycling of PET oligomers from chemical recycling processes to PHA by microbial co-cultivation.
Waste Manag. 2023,
172, 51–59.
[Google Scholar]
-
149.
Briassoulis D, Tserotas P, Athanasoulia I-G. Alternative optimization routes for improving the performance of poly(3-hydroxybutyrate) (PHB) based plastics.
J. Clean. Prod. 2021,
318, 128555.
[Google Scholar]
-
150.
Liu P, Zhang T, Zheng Y, Li Q, Su T, Qi Q. Potential one-step strategy for PET degradation and PHB biosynthesis through co-cultivation of two engineered microorganisms.
Eng. Microb. 2021,
1, 100003.
[Google Scholar]
-
151.
Bao T, Qian Y, Xin Y, Collins JJ, Lu T. Engineering microbial division of labor for plastic upcycling.
Nat. Com. 2023,
14, 5712.
[Google Scholar]
-
152.
Rosenboom J-G, Langer R, Traverso G. Bioplastics for a circular economy.
Nat. Rev. Mater. 2022,
7, 117–137.
[Google Scholar]
-
153.
Bioplastics Market to Touch
$4.4 bn by 2026. Focus Catal. 2020, 2020, 2. Available online:
http://www.chemicalweekly.com (accessed on 20 March 2020)
-
154.
Kumar R, Sadeghi K, Jang J, Seo J. Mechanical, chemical, and bio-recycling of biodegradable plastics: A review.
Sci. Total Env. 2023,
882 , 163446.
[Google Scholar]
-
155.
Narancic T, Salvador M, Hughes GM, Beagan N, Abdulmutalib U, Kenny ST, et al. Genome analysis of the metabolically versatile Pseudomonas umsongensis GO16: the genetic basis for PET monomer upcycling into polyhydroxyalkanoates.
Microb. Biotech. 2021,
14, 2463–2480.
[Google Scholar]
-
156.
Dou C, Choudhary H, Wang Z, Baral NR, Mohan M, Aguilar RA, et al. A hybrid chemical-biological approach can upcycle mixed plastic waste with reduced cost and carbon footprint.
One Earth 2023,
6, 1576–1590.
[Google Scholar]
-
157.
Abu-Thabit NY, Pérez-Rivero C, Uwaezuoke OJ, Ngwuluka NC. From waste to wealth: upcycling of plastic and lignocellulosic wastes to PHAs.
J. Chem. Tech. Biotech. 2022,
97, 3217–3240.
[Google Scholar]
-
158.
Guzik MW, Kenny ST, Duane GF, Casey E, Woods T, Babu RP, et al. Conversion of post consumer polyethylene to the biodegradable polymer polyhydroxyalkanoate.
App. Micro. Biotech. 2014,
98, 4223–4232.
[Google Scholar]
-
159.
Johnston B, Radecka I, Chiellini E, Barsi D, Ilieva VI, Sikorska W, et al. Mass Spectrometry Reveals Molecular Structure of Polyhydroxyalkanoates Attained by Bioconversion of Oxidized Polypropylene Waste Fragments.
Polymers 2019,
11, 1580.
[Google Scholar]
-
160.
Li A, Sheng Y, Cui H, Wang M, Wu L, Song Y, et al. Discovery and mechanism-guided engineering of BHET hydrolases for improved PET recycling and upcycling.
Nat. Com. 2023,
14, 4169.
[Google Scholar]
-
161.
Youngpreda A, Panyachanakul T, Kitpreechavanich V, Sirisansaneeyakul S, Suksamrarn S, Tokuyama S, et al. Optimization of Poly(dl-Lactic Acid) Degradation and Evaluation of Biological Re-polymerization.
J. Polym. Env. 2017,
25, 1131–1139.
[Google Scholar]
-
162.
Zoghbi L, Argeiti C, Skliros D, Flemetakis E, Koutinas A, Pateraki C, et al. Circular PHB production via Paraburkholderia sacchari cultures using degradation monomers from PHB-based post-consumer bioplastics as carbon sources.
Biochem. Eng. J. 2023,
191, 108808.
[Google Scholar]
-
163.
Samak NA, Jia Y, Sharshar MM, Mu T, Yang M, Peh S, et al. Recent advances in biocatalysts engineering for polyethylene terephthalate plastic waste green recycling.
Env. Inter. 2020,
145, 106144.
[Google Scholar]
-
164.
Ma R, Li Y, Zhang M, Xu F. Methanol tolerance upgrading of Proteus mirabilis lipase by machine learning-assisted directed evolution.
Sys. Microb. Bio. 2023,
3, 427–439.
[Google Scholar]
-
165.
Lu H, Diaz DJ, Czarnecki NJ, Zhu C, Kim W, Shroff R, et al. Machine learning-aided engineering of hydrolases for PET depolymerization.
Nature 2022,
604, 662–667.
[Google Scholar]
-
166.
Kim S, Kong D, Zheng X, Park JH. Upcycling plastic wastes into value-added products via electrocatalysis and photoelectrocatalysis.
J. Energy Chem. 2024,
91, 522–541.
[Google Scholar]
-
167.
Sangiorgio P, Verardi A, Dimatteo S, Spagnoletta A, Moliterni S, Errico S. Tenebrio molitorin the circular economy: a novel approach for plastic valorisation and PHA biological recovery.
Envir. Sci. Pollu. Res. 2021,
28, 52689–52701.
[Google Scholar]
-
168.
Hong K-Q, Zhang J, Jin B, Chen T, Wang Z-W. Development and characterization of a glycine biosensor system for fine-tuned metabolic regulation in Escherichia coli.
Microb. Cell Fact. 2022,
21, 56.
[Google Scholar]