1. Introduction
The molecular mechanisms that regulate progressive tissue fibrosis remain poorly understood. Although the causes of pulmonary fibrosis remain largely unknown, it is believed that progressive pulmonary fibrosis is an epithelial-fibroblastic disorder that results from numerous microinjuries to the alveolar epithelia that lead to excessive fibroblast activation, deposition of extracellular matrix, and eventually loss of normal lung architecture [
1]. Lung fibrosis is regulated by a variety of processes including TGFbeta signaling [
2,
3], metabolism [
4,
5,
6], non-coding RNAs [
7,
8,
9], epigenetic changes [
10], as well as aging [
11].
In the lung, type 2 alveolar epithelial cells (AEC2s) function as adult stem cells [
12]. Our previous work showed that there is a loss of AEC2s [
13,
14] and a failure of AEC2 renewal in the lungs of idiopathic pulmonary fibrosis (IPF) patients [
13]. We and others have showed that AEC2 renewal is regulated by several pathways [
13,
15,
16,
17,
18,
19] and metabolic reprogramming [
6,
20]. Furthermore, AEC2 stem cell activity is also regulated by the AEC2 niche [
18,
21,
22,
23], including basal-like cells [
24], fibroblasts [
18,
25,
26], endothelial cells [
20,
27,
28], macrophages [
17,
29], and T cells [
30], through various mechanisms such as extracellular vesicle trafficking [
18], cytokine release [
18,
24], or cell–cell interactions.
Beta-arrestins are classically known to participate in agonist-mediated G-protein-coupled receptor (GPCR) signaling through receptor desensitization and internalization [
31,
32,
33], playing a wide range of roles in development, cancer, asthma [
34,
35], and tissue fibrosis [
36]. We previously reported that beta-arrestin 1 and 2 deficient mice exhibit decreased mortality, decreased matrix deposition, and increased lung function in a model of bleomycin-induced lung fibrosis [
37], and serve as the key regulators of fibroblast invasion [
37]. However, the role of beta-arrestins in AEC2 regeneration is unclear.
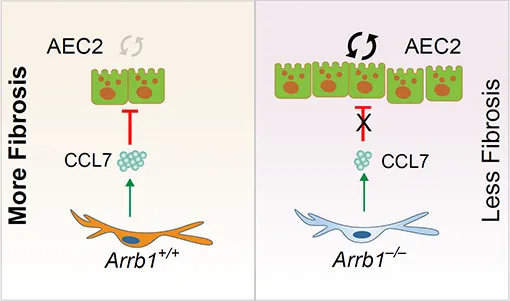
In this study, we investigated the role of Arrestin beta 1 (ARRB1) [
38] in AEC2 renewal and in lung fibrosis. We investigated the role of ARRB1 in regulating AEC2 regeneration using conventional deletion as well as cell type-specific deletion of ARRB1 in mice with a lung fibrosis mouse model. We found that targeted deletion of ARRB1 in fibroblasts promoted AEC2 regeneration and ameliorated lung fibrosis.
2. Materials And Methods
2.1. Mice
Arrb1−/− mice (Strain #: 011131, full name B6.129X1(Cg)-
Arrb1tm1Jse/J) [
39] and wild type (WT) C57BL/6J were purchased from The Jackson Laboratory.
Arrb1−/− mice were crossbred onto C57BL/6J background for more than five generations.
Arrb1flox/flox mice [
37] were from R. Lefkowitz of Duke University. Col1a2-Cre mice were from Dr. P. Angel of German Cancer Research Center (Das Deutsche Krebsforschungszentrum, DKFZ) [
40]. SPC-Cre mice were described previously [
41]. All mice were housed in a pathogen-free facility at Cedars-Sinai Medical Center. All animal experiments were approved by the Institutional Animal Care and Use Committee at Cedars-Sinai Medical Center (IACUC008529).
2.2. Bleomycin-induce Lung Injury and Fibrosis in Mice
Bleomycin-induce lung injury and fibrosis in age matched 8- to 12-week mice was described previously [
13]. Under anesthesia, the trachea was surgically exposed. 1.25 U/kg bleomycin (Hospira, Lake Forest, IL, USA) in 25 mL PBS was instilled into the mouse trachea with a 25-G needle inserted between the cartilaginous rings of the trachea. On day 0, day 7, and day 14, mice were sacrificed and lung tissues were collected for experiments.
2.3. Mouse Lung Tissue Digestion
Mouse lung tissue digestion was described previously [
6,
12,
13,
42]. In brief, mouse lungs were first digested with 4 U/mL Elastase in DMEM/F12 for 15 min. Then tissues were chopped into small pieces, and digested further with 100 mg/mL DNase I in DMEM/F12 for 15 min. Digestion was terminated with DMEM/F12 medium. Single cells were filtered through 100 mm filter, spined down, and resuspend with 700 mL HBSS
+ buffer for downstream experiments.
2.4. Flow Cytometry
Flow cytometrical analysis was described previously [
6,
13]. Mouse lung single cell suspension was stained with fixable viability dye, EPCAM, CD45, CD31, CD34, PDGFRa
+, Sca-1 and CD24. AEC2s were gated as live EPCAM
+CD45
−CD31
−CD34
−Sca-1
−CD24
− cells, PDGFRa
+ fibroblasts were gated as live EPCAM
−CD45
−CD31
−CD34
− PDGFRa
+ cells. Primary antibodies Percp-Cy5.5-EPCAM (clone G8.8, catalog# 118220), APC-Cy7-CD45 (clone 30-F11, catalog# 103116), APC-Cy7-CD31 (clone MEC13.3, catalog# 102534), APC-Cy7-CD34 (clone HM34, catalog# 128622), FITC-CD24 (clone M1/69, catalog# 101806), PE-Cy7-Sca-1 (clone D7, catalog# 108114) and APC-PDGFRa
+ (clone APA5, catalog# 135908) were from BioLegend (San Diego, CA, USA). Fixable viability dye (catalog# 65-0865-14) was from Thermo Fisher (Waltham, MA, USA). Stained cells were sorted on Aria III (BD Immunocytometry Systems; Franklin Lakes, NJ, USA) or analyzed on Fortessa (BD Immunocytometry Systems) and the data were analyzed with FlowJo software.
2.5. Cell Cycle Analysis
Surface marker-stained single cell suspension was fixed with Foxp3/Transcription Factor Staining Buffer Set (catalog# 00-5523-00, Thermo Fisher) for 1 h, then stained with conjugated APC-Ki-67 antibody (clone 16A8, catalog 652406, BioLegend) for another hour, then cells were washed, resuspended, and analyzed on Fortessa and the data were analyzed with FlowJo software.
2.6. RNA Isolation and Analysis
RNA isolation and analysis was described previously [
6,
24]. Briefly, 500 mL Trizol reagent (catalog# 15596026, Thermo Fisher) was added to freshly isolated PDGFRa
+ fibroblasts, and incubated at room temperature for 5 min. 100 mL chloroform solution was added and incubated for 3 min. 250 mL isopropanol was then added and incubated for another 10 min. The mixture was then centrifuged for 10 min at 13,500 rpm, and supernatant was discarded, pellets were washed with 500 mL of 75% ethanol, followed by centrifuging for 5 min at 7500
g at 4 °C. After removal of the supernatant, RNA was air dried for 10 min, resuspend in 30 mL RNase-free water. Total RNAs were sent for RNA-Seq and reverse-transcribed into cDNA for real-time PCR.
2.7. Immunofluorescence
Mouse lung tissues were fixed with 10% formalin solution and embedded in paraffin. The tissues were sectioned, and the slices were blocked with 5% goat serum, stained with primary antibodies against pro-SPC (catalog# ab170699, Abcam; Boston, MA, USA) overnight at 4 °C. On the next day, the samples were washed with PBS+0.01% Tween 20 buffer and stained with secondary antibody. Results were visualized with Zeiss LSM 780 confocal microscope (Carl Zeiss AG, Oberkochen, Germany).
2.8. Hydroxyproline Assay
Hydroxyproline assay to determine collagen contents in mouse lungs was described previously [
6,
24]. In brief, lung tissues were heat-dried, minced, and hydrolyzed overnight. The other day, pH was adjusted and samples were diluted with PBS. Then, samples were incubated with Chloramine T solution, perchloric acid and P-DMAB solution. Finally, samples were read under 557 nm wavelength. The ability of the assay to completely hydrolyze and recover hydroxyproline from collagen was confirmed using samples containing known amounts of purified collagen.
2.9. 3D Organoid Cultures of AEC2s
AEC2 organoid assays were described previously [
6,
12,
13,
24,
42]. Col1a2-Cre;
Arrb1fl/fl and
Arrb1fl/fl mice were treated with 1.25 U/kg bleomycin. 14 days after treatment, 1 × 105 fresh isolated lung PDGFRa
+ fibroblasts were mixed with 3000 flow-sorted WT AEC2 cells, resuspended in 100 μL matrigel/medium (1:1) mixture (catalog# 356252, Corning Inc.; Corning, NY, USA) and plated into 24-well transwell insert, and cultured for another 14 days. For colony-formation assay with CCL7, WT AEC2 cells were pre-treated with 10 ng/mL recombinant CCL7 protein (catalog# 250-08, Peprotech; Cranbury, NJ, USA) at 37 °C for 30 min, then mixed with mlg2908 fibroblasts. 400 μL medium was added to the lower chamber of the insert with a half medium change every other day. In the treatment group, 10 ng/mL CCL7 was also added to lower chamber medium. 14 days after culturing, colonies were counted and colony-formation efficiency was calculated.
2.10. Statistics
Results were shown as mean ± SD, statistical difference was calculated with Prism 8 (GraphPad Software, San Diego, CA, USA). Student’s two-tailed t test was used for two-group comparisons. Results were considered statistically significant at
p < 0.05.
2.11. Data and Material Availability
The data used in this paper can be accessed via GSE47460 [
43] and GSE48455 [
44]. Raw data of RNA-seq is available upon request. Further information and requests for resources and reagents should be directed to D. Jiang (Dianhua.Jiang@cshs.org).
3. Results
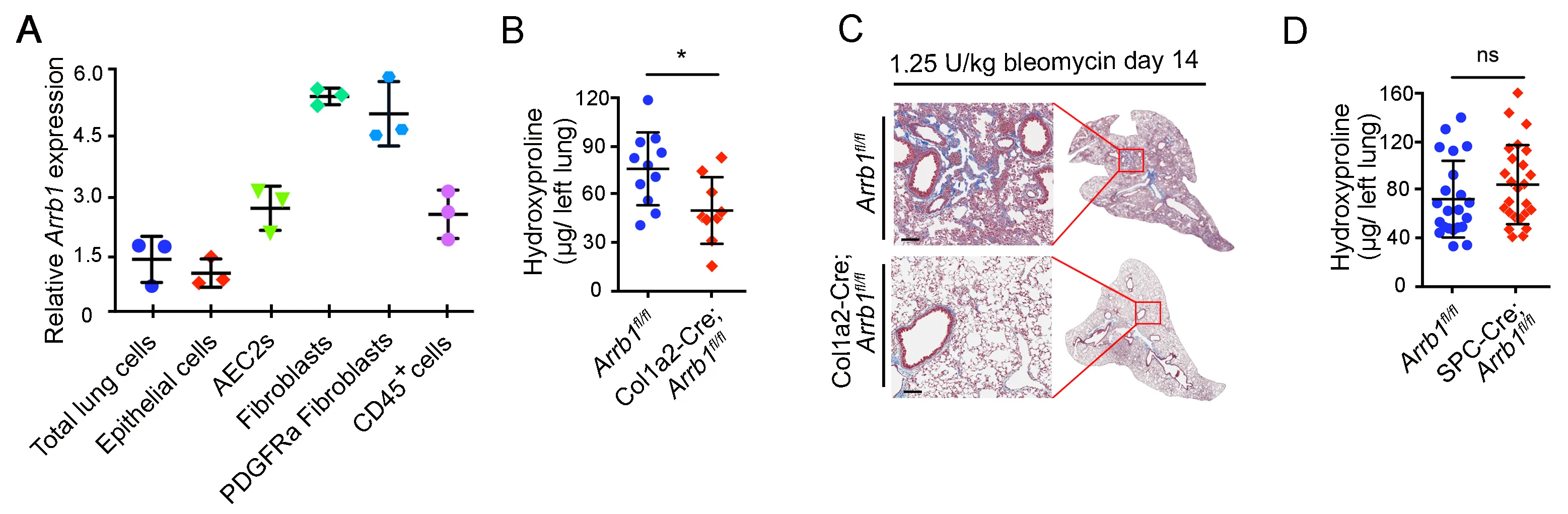
3.1. Arrb1 Deficiency in Fibroblasts Protects Mice from Lung Fibrosis
Our previous data showed that mice with conventional deletion of
Arrb1 were protected from bleomycin-induced injury and fibrosis [
37]. To study the cell type responsible for this protective role, we analyzed
Arrb1 expression in different cell types in the lung. Results showed that
Arrb1 transcripts were highly expressed in lung fibroblasts, especially PDGFRa
+ fibroblasts (A). We further generated a mouse strain with lung fibroblast-specific deficiency of
Arrb1. When challenged with bleomycin, mice with lung fibroblast-specific deficiency of
Arrb1 showed to a decrease in hydroxyproline content in the lungs compared to littermate control mice (B), and reduced lung fibrosis was confirmed with Trichrome staining (C). In contrast, deletion of Arrb1 in SPC
+ AEC2 compartment did not affect fibrosis when challenged with bleomycin (D). These data demonstrate a detrimental role of ARRB1 in fibroblasts in bleomycin-induced lung fibrosis.
Figure 1. Arrb1 deficiency in fibroblasts protects mice from lung fibrosis. (<b>A</b>) Expression of <em>Arrb1</em> in different cell types from mouse lungs was determined. Cells were isolated via FACS, and total RNAs were extracted. <em>Arrb1</em> expression was determined with RT-PCR (<em>n</em> = 3). (<b>B</b>) Hydroxyproline assays of mouse left lungs 14 days after 1.25 U/kg bleomycin treatment (<em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 11, Col1a2-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> =9, *<em>p</em> < 0.05). (<b>C</b>) Trichrome staining of mouse lungs 14 days after 1.25 U/kg bleomycin injury. (<b>D</b>) Hydroxyproline assays of mouse left lungs 14 days after 1.25 U/kg bleomycin treatment (<em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 22, SPC-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> =25, *<em>p</em> < 0.05).
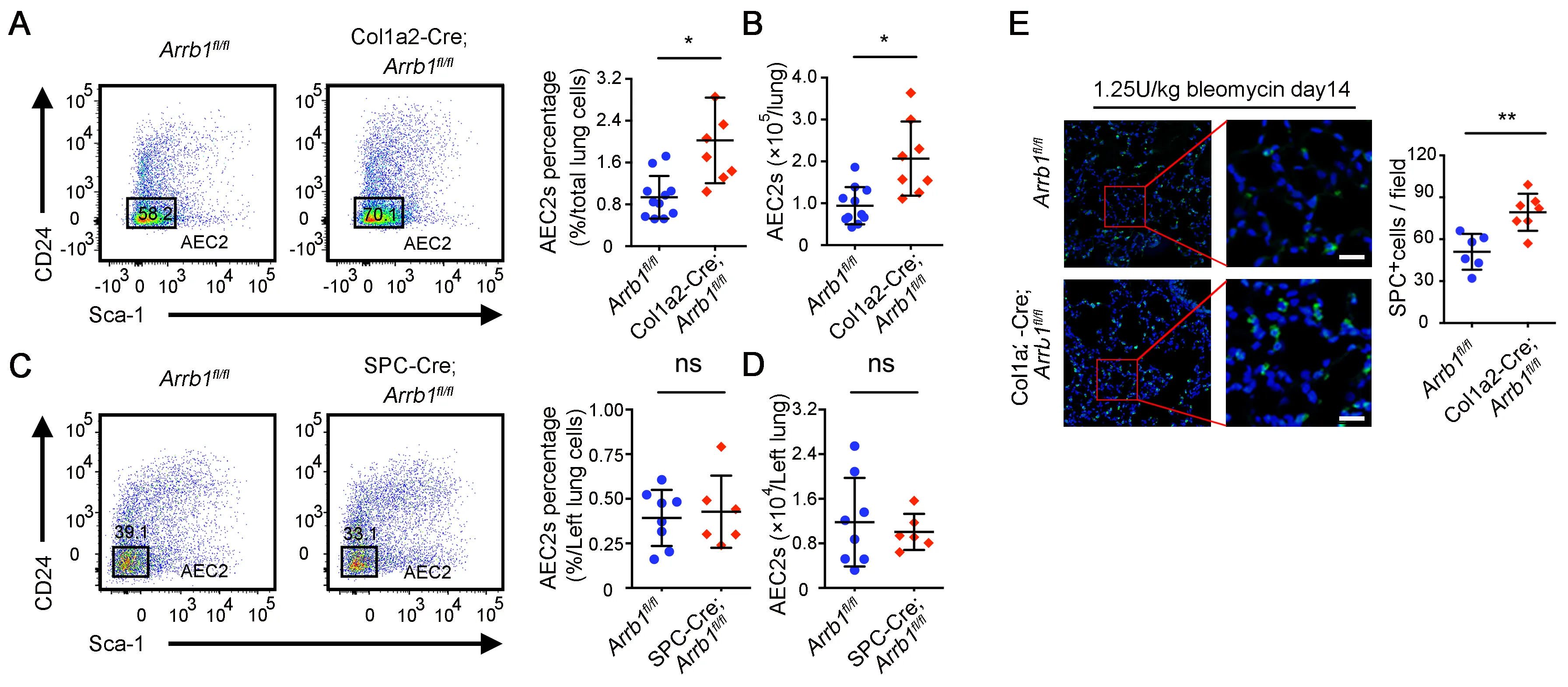
The mounting evidence indicates that the stem cell niche plays a crucial role in governing the regeneration of stem cells [
45]. We recently showed that injured fibroblasts were less supportive role of AEC2 renewal [
18]. We thus investigated the potential impact of
Arrb1 deletion in fibroblasts on the regeneration of AEC2. We found that mouse lungs with fibroblast-specific
Arrb1 deficiency have a higher percentage (A) and number (B) of AEC2 cells after bleomycin treatment. In contrast, deletion of
Arrb1 in SPC
+ AEC2 compartment did not affect the percentage and number of AEC2 cells after bleomycin treatment (C,D). These data indicated that deficiency of
Arrb1 in fibroblasts may promote AEC2 regeneration after injury. The results were further confirmed with immunofluorescence staining of pro-SPC (E). These data showed that
Arrb1 deficiency in fibroblasts leads to enhanced recovery of AEC2 cells after lung injury.
. <em>Arrb1</em> deficiency in fibroblasts leads to increased AEC2 recovery in injured mouse lung. (<b>A</b>–<b>D</b>) 14 days after 1.25 U/kg bleomycin treatment, AEC2 cells were analyzed as live CD45<sup>−</sup>CD31<sup>−</sup>CD34<sup>−</sup>Epcam<sup>+</sup>CD24<sup>−</sup>Sca-1<sup>−</sup> cells with flow cytometry. Percentage (<b>A</b>, <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 11, Col1a2-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 7; *<em>p</em> < 0.05) and total number (<b>B</b>, <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 11, Col1a2-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 8; *<em>p</em> < 0.05) of AEC2 cells in Col1a2-Cre;<em>Arrb1</em><sup>fl/fl</sup> mouse lungs as well as percentage (<b>C</b>, <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 8, SPC-Cre;<em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 6; ns, not significant) and total number (<b>D</b>, <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 8, SPC-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 6; ns, not significant) of AEC2 cells in SPC-Cre;<em>Arrb1</em><sup>fl/fl</sup> mouse lungs were calculated according to flow results. (<b>E</b>) Immunofluorescence staining of SPC<sup>+</sup> cells from mouse lungs 14 days after 1.25 U/kg bleomycin treatment (<em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 7, Col1a2-Cre; <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 8; **<em>p</em> < 0.01).
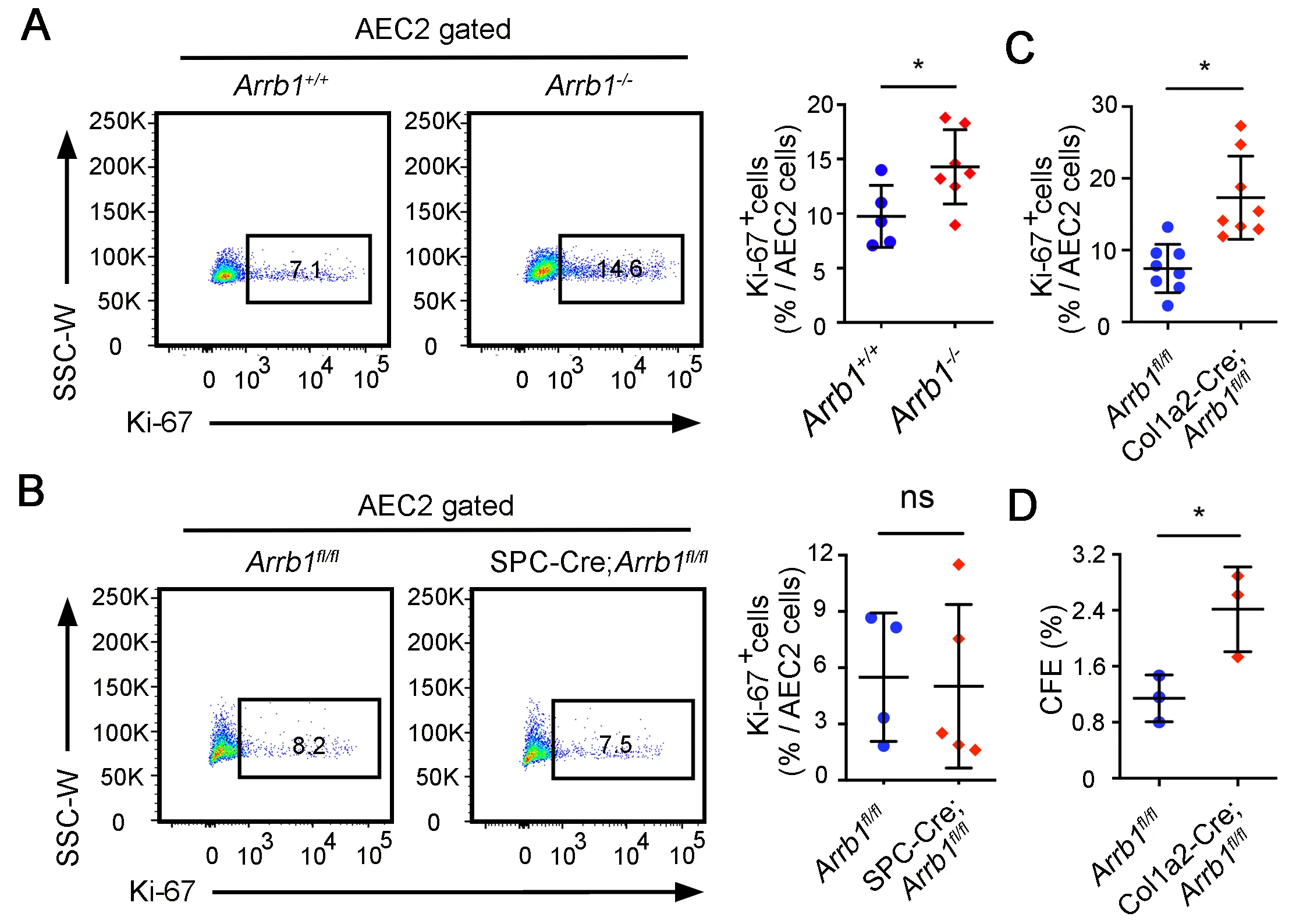
AEC2 cells are adult stem cells in the lung, and regenerate the alveolar epithelial cells after injury [
12,
13]. To further demonstrate that ARRB1 affects AEC2 regeneration, we first assessed AEC2 proliferation in
Arrb1−/− total knockout mice after bleomycin injury. The results showed that 7 days after bleomycin, there are more Ki-67
+ AEC2 cells in
Arrb1−/− mice compared to control mice (A). However, the increase of Ki-67
+ AEC2 cells was not observed in the lungs of mice with
Arrb1 deletion specifically in AEC2s (SPC-Cre;
Arrb1fl/fl mice) (B), indicating that ARRB1 promotes AEC2 proliferation via fibroblasts rather than AEC2s themselves. Indeed, when we deleted Arrb1 in Col1a2
+ fibroblasts, an increase of Ki-67
+ AEC2 cells was observed (C). To further validate that
Arrb1−/− fibroblasts promote AEC2 proliferation, WT AEC2 cells were cocultured with
Arrb1-deficient PDGFRa
+ fibroblasts. Results showed that
Arrb1 deficiency in fibroblasts significantly increased colony-formation ability of AEC2 cells (D), confirming that ARRB1 from fibroblasts affects AEC2 cell proliferation and renewal.
. <em>Arrb1</em>-deficient fibroblasts promote AEC2 proliferation and renewal. (<b>A</b>–<b>C</b>) All mice were subjected to 1.25 U/kg bleomycin and mouse lungs were harvested 14 days after injury. AEC2 cells were gated as live CD45<sup>−</sup>CD31<sup>−</sup>CD34<sup>−</sup>Epcam<sup>+</sup>CD24<sup>−</sup>Sca-1<sup>−</sup>, percentage of Ki-67<sup>+</sup> cells were calculated. Ki-67<sup>+</sup> AEC2 cells were analyzed by FACS in <em>Arrb1</em><sup>+/+</sup> and <em>Arrb1</em><sup>−/−</sup> mice (<b>A</b>, <em>Arrb1</em><sup>+/+</sup> <em>n</em> = 5, <em>Arrb1</em><sup>−/−</sup> <em>n</em> = 7; *<em>p</em> < 0.05), <em>Arrb1</em><sup>fl/fl</sup> and SPC-Cre;<em>Arrb1</em><sup>fl/fl</sup> mice (<b>B</b>, <em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 7, SPC-Cre;<em>Arrb1</em><sup>fl/fl</sup> <em>n</em> = 6; ns, not significant), <em>Arrb1</em><sup>fl/fl</sup> and Col1a2-Cre; <em>Arrb1</em><sup>fl/fl</sup> mice (<b>C</b>, <em>n</em> = 8; *<em>p</em> < 0.05). (<b>D</b>) AEC2 cells from wild type C57BL/6 mice were cocultured with freshly isolated PDGFRa<sup>+</sup> fibroblasts (CD45<sup>−</sup>CD31<sup>−</sup>CD34<sup>−</sup>EPCAM<sup>−</sup>PDGFRa<sup>+</sup>) from <em>Arrb1</em><sup>fl/fl</sup> and Col1a2-Cre;<em>Arrb1</em><sup>fl/fl</sup> mice lungs 7 days after 1.25 U/kg bleomycin treatment. 14 days after culture, colony-formation efficiency (CFE) was calculated (<em>n</em> = 3, *<em>p</em> < 0.05).
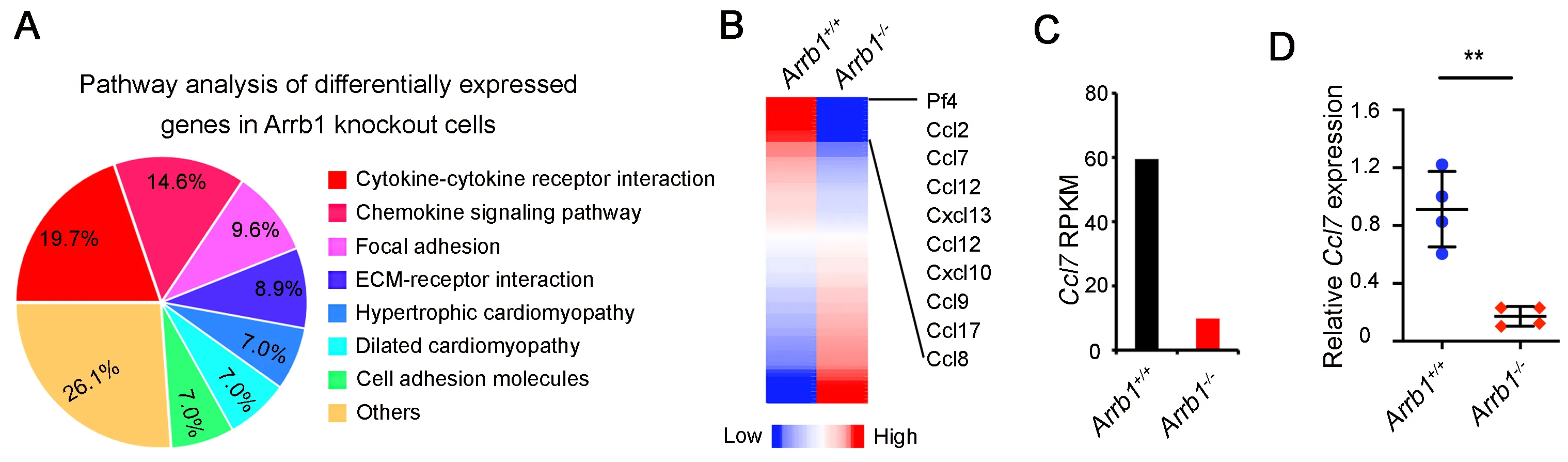
Next, we sought to determine how
Arrb1-deficent fibroblasts promote AEC2 proliferation. We isolated PDGFRa
+ fibroblasts from
Arrb1+/+ and
Arrb1−/− mice, and performed RNA sequencing (RNA-Seq). Results showed that differentially expressed genes were enriched in cytokine-cytokine receptor interactions and chemokine signaling pathways (A). Cytokines and chemokines analysis showed that
Ccl7 was among the top differential expressed genes in Arrb1 deficiency (B). RPKM results also confirmed that
Ccl7 was downregulated in
Arrb1-deficient fibroblasts (C). We confirmed these results by RT-PCR analysis, showing that Ccl7 expression was decreased in
Arrb1-deficient fibroblasts (D). Thus, these data demonstrated that
Arrb1 deficiency downregulates
Ccl7 expression in fibroblasts.
. Arrb1 promotes Ccl7 expression in fibroblasts. (<b>A</b>) <em>Arrb1</em><sup>+/+</sup> and <em>Arrb1</em><sup>−/−</sup> mice were subjected to 1.25 U/kg bleomycin injury, PDGFRa<sup>+</sup> fibroblasts were isolated with FACS 7 days after treatment. RNA-seq was performed with total RNAs isolated from these cells. Pathway analysis was conducted with Ingenuity Pathway Analysis (IPA) on the differentially expressed genes (DEGs). (<b>B</b>) Differentially expressed cytokines and chemokines were extracted. C–C chemokine genes are indicated. (<b>C</b>) RPKM of <em>Ccl7</em> in PDGFRa<sup>+</sup> fibroblasts from <em>Arrb1</em><sup>+/+</sup> and <em>Arrb1</em><sup>−/−</sup> lungs in RNA-Seq analysis. (<b>D</b>) <em>Ccl7</em> expression in PDGFRa<sup>+</sup> fibroblasts was analyzed with RT-PCR (<em>n</em> = 4; **<em>p</em> < 0.01).
Our data show that mice with
Arrb1 deficiency in fibroblasts were protected from bleomycin-induced lung fibrosis,
Arrb1-deficient fibroblasts promoted AEC2 proliferation, and
Arrb1 deficiency in fibroblasts led to decreased Ccl7 expression. CCL7 (C–C Motif Chemokine Ligand 7, aka Monocyte Chemoattractant Protein 3, MCP3) is a member of the C–C subfamily of chemokines [
46], which plays a role in macrophage recruitment during inflammation [
47] and in tumor metastasis [
46].
Ccl7 is among several upregulated inflammatory genes identified during early injury [
44].
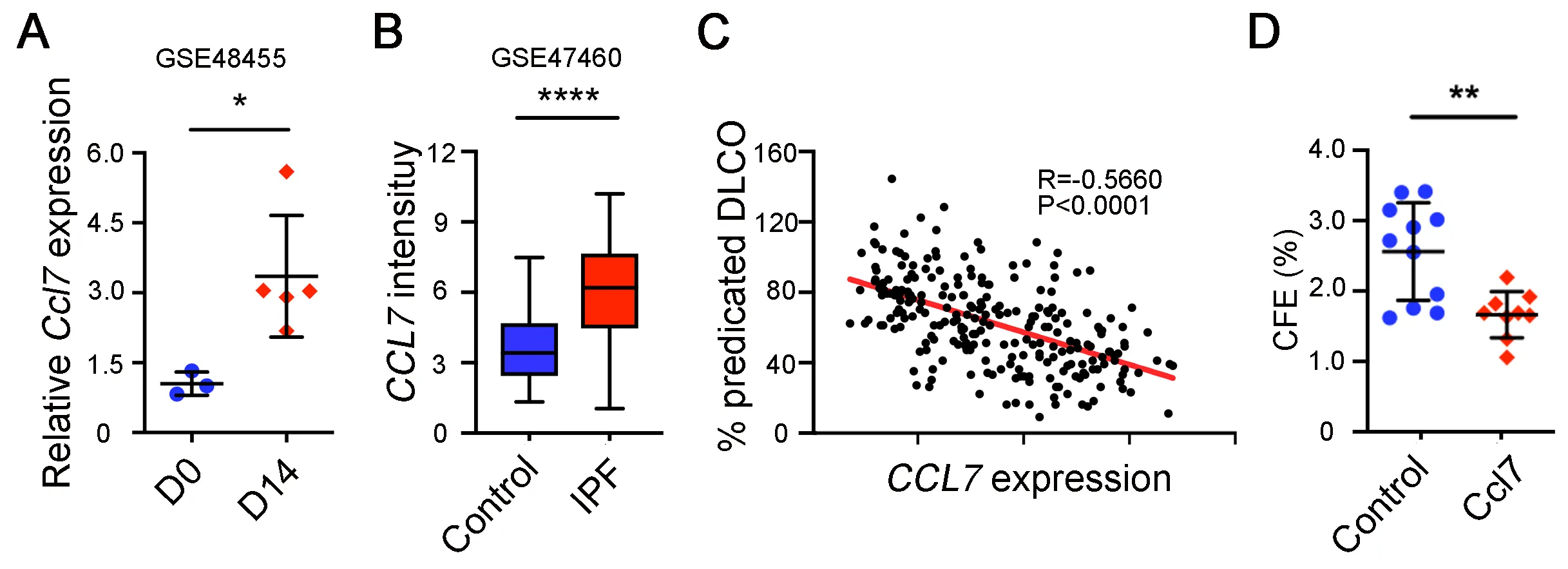
Next, we determined whether CCL7 directly affects AEC2 renewal. Analysis of rat lung injury data [
44] showed that Ccl7 expression was increased after bleomycin treatment (A). Importantly, CCL7 expression was also found to be increased in lungs of IPF patients compared to healthy lungs (B). Furthermore, CCL7 expression is correlated with reduced lung function shown as the negative correlation between CCL7 expression and the percentage of predicted diffusion capacity for carbon monoxide (DLCO) (C). In addition, 3D organoid experiments showed that adding CCL7 in organoid assays significantly decreased AEC2 renewal capacity (D), confirming that CCL7 inhibits AEC2 regeneration.
. CCL7 inhibits AEC2 renewal. (<b>A</b>) Ccl7 expression on day 0 and day 14 rat lungs after bleomycin treatment in GSE48455 dataset was analyzed (Day 0 <em>n</em> = 3, Day 14 <em>n</em> = 5, *<em>p</em> < 0.05). (<b>B</b>) <em>CCL7</em> expression in IPF patients and healthy controls in GSE47460 dataset was analyzed (Control <em>n</em> = 108, IPF <em>n</em> = 160, ****p < 0.0001). (<b>C</b>) Pearson correlation of <em>CCL7</em> expression and the percentage of predicted diffusion capacity for carbon monoxide (DLCO) in GSE47460 dataset was determined (<em>n</em> = 242, <em>R</em> = −0.5660, <em>p</em> < 0.0001). (<b>D</b>) Colony formation efficiency of AEC2 from wild type C57BL/6 mice treated with 10 ng/mL CCL7 (Control <em>n</em> = 11, CCL7 <em>n</em> = 9; **p < 0.01).
In summary, these results demonstrate that fibroblast ARRB1 mediates AEC2 regeneration, possibly by releasing chemokine CCL7, which inhibits AEC2 regeneration, leading to fibrosis in the lung.
4. Discussion
AEC2s are adult stem cells in the lung [
12]. We and others have demonstrated a loss of AEC2s and a failure of AEC2 renewal in IPF lungs [
13,
14]. We previously reported that beta-arrestins are the key regulators of fibroblast invasion, and beta-arrestin 1 and 2 deficient mice exhibit decreased lung fibrosis [
37]. However, the role of beta-arrestins in AEC2 regeneration is unclear. In this study, we investigated the role of ARRB1 in AEC2 renewal and in lung fibrosis and found that
Arrb1 deficiency in fibroblasts enhanced AEC2 regeneration and protected mice from lung fibrosis.
AEC2 stem cell regenerative capacity is regulated by the AEC2 stem cell niche [
18,
21,
22,
23]. We recently showed that basal-like cells may produce WNT ligands, mediating AEC2 renewal [
24]. Several studies have showed that lipofibroblasts and PDGFRa
+ fibroblasts regulate alveolar progenitor cell activities [
18,
25,
26]. Other cell types such as endothelial cells [
20,
27,
28], macrophages [
17,
29], and T cells [
30] regulate alveolar progenitor cell activities. The current study is consistent with these studies and makes the novel observation that fibroblast ARRB1 is crucial for alveolar progenitor cell regeneration. Beta-Arrestins are known to act as a scaffold to regulate GPCR signaling through receptor desensitization and internalization [
31,
32,
33]. Further studies are needed to determine how ARRB1 interacts with specific GPCRs in fibroblasts for this effector function.
Chemokine CCL7 (MCP3) is a member of the C–C subfamily [
46]. It has been shown to play a role in macrophage recruitment during inflammation [
47]. Both lung macrophages and myofibroblasts secrete CCL7 into the blood [
48]. We found that CCL7 is increased in IPF and negatively correlated with lung function, consistent with a previous report showing an increase in CCL7 in fibroblasts from patients with usual interstitial pneumonia [
49], in serum of patients with systemic sclerosis [
50], and a recent preprint showing a significant increase of CCL7 in IPF plasma compared to that of matched controls [
48]. We showed that CCL7 is also increased after bleomycin injury in mice, consistent with a previous report that CCL7 and CCL2 (MCP1) are increased in the BALF of Sftpc BRICHOS mutant mice [
51]. CCL7 may also be a feature of aging [
52]. Inhibition of CCL7 receptor CCR2 enhanced aged muscle regeneration and functional recovery after skeletal muscle injury [
52].
CCL7 is known for its role in macrophage recruitment, mediating immune response [
46]. It is reported that CCL7 promotes activation of the TGF-beta signaling pathways leading to increased type I collagen production [
53]. We previously reported that beta-arrestins are key regulators of fibroblast invasion [
37]. It is unclear if CCL7 has a role in fibroblast invasion. In this study, we showed that fibroblast-derived CCL7 participates in the AEC2 stem cell niche and inhibits AEC2 renewal. We previously showed that CXC chemokines such as CXCL1 [
18] and cytokines such as IL-6 promote AEC2 renewal [
13,
18]. CCL7 binds CCR2 [
54], CCR1, and CCR3, which are typical GPCRs. Future studies are needed to determine which and how CCL7 receptor participates in the AEC2 niche. At the cellular level, CCL7 may bind to CCR2 on macrophages, the latter may negatively regulate the AEC2 niche. Macrophages can influence lung stem cells in a pneumonectomy model in mice [
29]. Since CCR2 expression level on AEC2 cells is notably scant, it is unlikely that CCL7 inhibits AEC2 regeneration through CCL7–CCR2 interactions on AEC2s. Future studies are warranted to determine if the effect of CCL7 on AEC2 renewal is direct or indirect.
In conclusion, we demonstrate that fibroblast ARRB1 impedes AEC2 regeneration, possibly by releasing chemokine CCL7, leading to fibrosis in the lung.
Acknowledgments
The authors thank the members of our laboratory for support and helpful discussion during the study. We thank Ning Yu, and Jo Suda from Cedars-Sinai Medical Center Flow Core for help with FACS sorting.
Author Contributions
J.L., P.W.N., and D.J. conceived the study. G.H., Y.G. designed, performed experiments, and analyzed data. V.K., F.T., and N.L. performed experiments. X.L. performed data analysis and interpretion. G.H., J.L., P.W.N., and D.J. wrote the paper.
Ethics Statement
The mouse studies were approved by the Institutional Animal Care and Use Committee at Cedars-Sinai Medical Center (IACUC008529).
Informed Consent Statement
Not applicable.
Funding
This work was supported in part by National Institutes of Health grants P01 HL108793, R01 AG078655, and R01 HL122068.
Declaration of competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Noble PW, Barkauskas CE, Jiang D. Pulmonary fibrosis: patterns and perpetrators.
J. Clin. Invest. 2012,
122, 2756–2762.
[Google Scholar]
-
2.
Massague J, Sheppard D. Massague J, Sheppard D. TGF-beta signaling in health and
disease.
Cell 2023,
186, 4007–4037.
[Google Scholar]
-
3.
Taleb SJ, Ye Q, Baoyinna B, Dedad M, Pisini D, Parinandi NL, et al. Molecular Regulation of Transforming Growth Factor-beta1-induced Thioredoxin-interacting Protein Ubiquitination and Proteasomal Degradation in Lung Fibroblasts: Implication in Pulmonary Fibrosis.
J. Respir. Biol. Transl. Med. 2024,
1, 10002.
[Google Scholar]
-
4.
Xie N, Tan Z, Banerjee S, Cui H, Ge J, Liu RM, et al. Glycolytic Reprogramming in Myofibroblast Differentiation and Lung Fibrosis.
Am. J. Respir. Crit. Care. Med. 2015,
192, 1462–1474.
[Google Scholar]
-
5.
Bueno M, Lai YC, Romero Y, Brands J, St Croix CM, Kamga C, et al. PINK1 deficiency impairs mitochondrial homeostasis and promotes lung fibrosis.
J. Clin. Invest. 2015,
125, 521–538.
[Google Scholar]
-
6.
Liang J, Huang G, Liu X, Taghavifar F, Liu N, Wang Y, et al. The ZIP8/SIRT1 axis regulates alveolar progenitor cell renewal in aging and idiopathic pulmonary fibrosis.
J. Clin. Invest. 2022,
132, e157338.
[Google Scholar]
-
7.
Xie T, Liang J, Geng Y, Liu N, Kurkciyan A, Kulur V, et al. MicroRNA-29c Prevents Pulmonary Fibrosis by Regulating Epithelial Cell Renewal and Apoptosis.
Am. J. Respir. Cell. Mol. Biol. 2017,
57, 721–732.
[Google Scholar]
-
8.
Ge L, Habiel DM, Hansbro PM, Kim RY, Gharib SA, Edelman JD, et al. miR-323a-3p regulates lung fibrosis by targeting multiple profibrotic pathways.
JCI Insight 2016,
1, e90301.
[Google Scholar]
-
9.
Liu G, Friggeri A, Yang Y, Milosevic J, Ding Q, Thannickal VJ, et al. miR-21 mediates fibrogenic activation of pulmonary fibroblasts and lung fibrosis.
J. Exp. Med. 2010,
207, 1589–1597.
[Google Scholar]
-
10.
Huan C, Yang T, Liang J, Xie T, Cheng L, Liu N, et al. Methylation-mediated BMPER expression in fibroblast activation in vitro and lung fibrosis in mice in vivo.
Sci. Rep. 2015,
5, 14910.
[Google Scholar]
-
11.
Liang J, Huang G, Liu X, Liu N, Taghavifar F, Dai K, et al. Reciprocal interactions between alveolar progenitor dysfunction and aging promote lung fibrosis.
Elife 2023,
12, e85415.
[Google Scholar]
-
12.
Barkauskas CE, Cronce MJ, Rackley CR, Bowie EJ, Keene DR, Stripp BR, et al. Type 2 alveolar cells are stem cells in adult lung.
J. Clin. Invest. 2013,
123, 3025–3036.
[Google Scholar]
-
13.
Liang J, Zhang Y, Xie T, Liu N, Chen H, Geng Y, et al. Hyaluronan and TLR4 promote surfactant-protein-C-positive alveolar progenitor cell renewal and prevent severe pulmonary fibrosis in mice.
Nat. Med. 2016,
22, 1285–1293.
[Google Scholar]
-
14.
Morse C, Tabib T, Sembrat J, Buschur KL, Bittar HT, Valenzi E, et al. Proliferating SPP1/MERTK-expressing macrophages in idiopathic pulmonary fibrosis.
Eur. Respir. J. 2019,
54, 1802441.
[Google Scholar]
-
15.
Chen Q, Suresh Kumar V, Finn J, Jiang D, Liang J, Zhao YY, et al. CD44(high) alveolar type II cells show stem cell properties during steady-state alveolar homeostasis.
Am. J. Physiol. Lung Cell. Mol. Physiol. 2017,
313, L41–l51.
[Google Scholar]
-
16.
Glisinski KM, Schlobohm AJ, Paramore SV, Birukova A, Moseley MA, Foster MW, et al. Interleukin-13 disrupts type 2 pneumocyte stem cell activity.
JCI Insight 2020,
5, e131232.
[Google Scholar]
-
17.
Choi J, Park JE, Tsagkogeorga G, Yanagita M, Koo BK, Han N, et al. Inflammatory Signals Induce AT2 Cell-Derived Damage-Associated Transient Progenitors that Mediate Alveolar Regeneration.
Cell Stem Cell 2020,
27, 366–382.
[Google Scholar]
-
18.
Xie T, Kulur V, Liu N, Deng N, Wang Y, Rowan SC, et al. Mesenchymal growth hormone receptor deficiency leads to failure of alveolar progenitor cell function and severe pulmonary fibrosis.
Sci. Adv. 2021,
7, eabg6005.
[Google Scholar]
-
19.
Liu X, Zhang X, Yao C, Liang J, Noble PW, Jiang D. Transcriptomics Analysis Identifies the Decline in the AT2 Stem Cell Niches in Aged Human Lungs.
Am. J. Respir. Cell. Mol. Biol. 2024,
70, DOI: 10.1165/rcmb.2023-0363OC.
[Google Scholar]
-
20.
Chen Q, Rehman J, Chan M, Fu P, Dudek SM, Natarajan V, et al. Angiocrine Sphingosine-1-Phosphate Activation of S1PR2-YAP Signaling Axis in Alveolar Type II Cells Is Essential for Lung Repair.
Cell Rep. 2020,
31, 107828.
[Google Scholar]
-
21.
Donne ML, Lechner AJ, Rock JR. Evidence for lung epithelial stem cell niches.
BMC Dev. Biol. 2015,
15, 32.
[Google Scholar]
-
22.
Basil MC, Katzen J, Engler AE, Guo M, Herriges MJ, Kathiriya JJ, et al. The Cellular and Physiological Basis for Lung Repair and Regeneration: Past, Present, and Future.
Cell Stem Cell 2020,
26, 482–502.
[Google Scholar]
-
23.
Juul NH, Stockman CA, Desai TJ. Niche Cells and Signals that Regulate Lung Alveolar Stem Cells In Vivo.
Cold Spring Harb. Perspect. Biol. 2020,
12, a035717.
[Google Scholar]
-
24.
Huang G, Liang J, Huang K, Liu X, Taghavifar F, Yao C, et al. Basal Cell-derived WNT7A Promotes Fibrogenesis at the Fibrotic Niche in Idiopathic Pulmonary Fibrosis.
Am. J. Respir. Cell. Mol. Biol. 2023,
68, 302–313.
[Google Scholar]
-
25.
Zysman M, Baptista BR, Essari LA, Taghizadeh S, Thibault de Menonville C, Giffard C, et al. Targeting p16(INK4a) Promotes Lipofibroblasts and Alveolar Regeneration after Early-Life Injury.
Am. J. Respir. Crit. Care. Med. 2020,
202, 1088–1104.
[Google Scholar]
-
26.
Gao F, Li C, Danopoulos S, Al Alam D, Peinado N, Webster S, et al. Hedgehog-responsive PDGFRa(+) fibroblasts maintain a unique pool of alveolar epithelial progenitor cells during alveologenesis.
Cell Rep. 2022,
39, 110608.
[Google Scholar]
-
27.
Rafii S, Cao Z, Lis R, Siempos, II, Chavez D, Shido K, et al. Platelet-derived SDF-1 primes the pulmonary capillary vascular niche to drive lung alveolar regeneration.
Nat. Cell. Biol. 2015,
17, 123–136.
[Google Scholar]
-
28.
Li K, Wu Q, Sun X, Geng Y, Leng D, Li H, et al. Tsp1 promotes alveolar stem cell proliferation and its down-regulation relates to lung inflammation in intralobar pulmonary sequestration.
Oncotarget 2017,
8, 64867–64877.
[Google Scholar]
-
29.
Lechner AJ, Driver IH, Lee J, Conroy CM, Nagle A, Locksley RM, et al. Recruited Monocytes and Type 2 Immunity Promote Lung Regeneration following Pneumonectomy.
Cell Stem Cell 2017,
21, 120–134.
[Google Scholar]
-
30.
Kaiser KA, Loffredo LF, Santos-Alexis KL, Ringham OR, Arpaia N. Regulation of the alveolar regenerative niche by amphiregulin-producing regulatory T cells.
J. Exp. Med. 2023,
220, e20221462.
[Google Scholar]
-
31.
Luttrell LM, Lefkowitz RJ. The role of beta-arrestins in the termination and transduction of G-protein-coupled receptor signals.
J. Cell Sci. 2002,
115, 455–465.
[Google Scholar]
-
32.
Ma L, Pei G. Beta-arrestin signaling and regulation of transcription.
J. Cell Sci. 2007,
120, 213–218.
[Google Scholar]
-
33.
Pydi SP, Barella LF, Zhu L, Meister J, Rossi M, Wess J. beta-Arrestins as Important Regulators of Glucose and Energy Homeostasis.
Annu. Rev. Physiol. 2022,
84, 17–40.
[Google Scholar]
-
34.
Walker JK, Fong AM, Lawson BL, Savov JD, Patel DD, Schwartz DA, et al. Beta-arrestin-2 regulates the development of allergic asthma.
J. Clin. Invest. 2003,
112, 566–574.
[Google Scholar]
-
35.
Hollingsworth JW, Theriot BS, Li Z, Lawson BL, Sunday M, Schwartz DA, et al. Both hematopoietic-derived and non-hematopoietic-derived beta-arrestin-2 regulates murine allergic airway disease.
Am. J. Respir. Cell. Mol. Biol. 2010,
43, 269–275.
[Google Scholar]
-
36.
Jiang D, Xie T, Liang J, Noble PW. beta-Arrestins in the immune system.
Prog. Mol. Biol. Transl. Sci. 2013,
118, 359–393.
[Google Scholar]
-
37.
Lovgren AK, Kovacs JJ, Xie T, Potts EN, Li Y, Foster WM, et al. beta-arrestin deficiency protects against pulmonary fibrosis in mice and prevents fibroblast invasion of extracellular matrix.
Sci. Transl. Med. 2011,
3, 74ra23.
[Google Scholar]
-
38.
Parruti G, Peracchia F, Sallese M, Ambrosini G, Masini M, Rotilio D, et al. Molecular analysis of human beta-arrestin-1: cloning, tissue distribution, and regulation of expression. Identification of two isoforms generated by alternative splicing.
J. Biol. Chem. 1993,
268, 9753–9761.
[Google Scholar]
-
39.
Conner DA, Mathier MA, Mortensen RM, Christe M, Vatner SF, Seidman CE, et al. beta-Arrestin1 knockout mice appear normal but demonstrate altered cardiac responses to beta-adrenergic stimulation.
Circ. Res. 1997,
81, 1021–1026.
[Google Scholar]
-
40.
Florin L, Alter H, Grone HJ, Szabowski A, Schutz G, Angel P. Cre recombinase-mediated gene targeting of mesenchymal cells.
Genesis 2004,
38, 139–144.
[Google Scholar]
-
41.
Eblaghie MC, Reedy M, Oliver T, Mishina Y, Hogan BL. Evidence that autocrine signaling through Bmpr1a regulates the proliferation, survival and morphogenetic behavior of distal lung epithelial cells.
Dev. Biol. 2006,
291, 67–82.
[Google Scholar]
-
42.
Liang J, Liu N, Liu X, Mena JM, Xie T, Geng Y, et al. Mitogen-activated Protein Kinase-activated Protein Kinase 2 Inhibition Attenuates Fibroblast Invasion and Severe Lung Fibrosis.
Am. J. Respir. Cell. Mol. Biol. 2019,
60, 41–48.
[Google Scholar]
-
43.
Kusko RL, Brothers JF, 2nd, Tedrow J, Pandit K, Huleihel L, Perdomo C, et al. Integrated Genomics Reveals Convergent Transcriptomic Networks Underlying Chronic Obstructive Pulmonary Disease and Idiopathic Pulmonary Fibrosis.
Am. J. Respir. Crit. Care. Med. 2016,
194, 948–960.
[Google Scholar]
-
44.
Bauer Y, Tedrow J, de Bernard S, Birker-Robaczewska M, Gibson KF, Guardela BJ, et al. A novel genomic signature with translational significance for human idiopathic pulmonary fibrosis.
Am. J. Respir. Cell. Mol. Biol. 2015,
52, 217–231.
[Google Scholar]
-
45.
Wu H, Tang N. Stem cells in pulmonary alveolar regeneration.
Development 2021,
148, dev193458.
[Google Scholar]
-
46.
Opdenakker G, Froyen G, Fiten P, Proost P, Van Damme J. Human monocyte chemotactic protein-3 (MCP-3): molecular cloning of the cDNA and comparison with other chemokines. Biochem. Biophys.
Res. Commun. 1993,
191, 535–542.
[Google Scholar]
-
47.
Mercer PF, Williams AE, Scotton CJ, Jose RJ, Sulikowski M, Moffatt JD, et al. Proteinase-activated receptor-1, CCL2, and CCL7 regulate acute neutrophilic lung inflammation.
Am. J. Respir. Cell. Mol. Biol. 2014,
50, 144–157.
[Google Scholar]
-
48.
Unterman A, Zhao AY, Neumark N, Schupp JC, Ahangari F, Cosme C, Jr., et al. Single-cell profiling reveals immune aberrations in progressive idiopathic pulmonary fibrosis.
medRxiv. 2023,
, doi: 10.1101/2023.1104.1129.23289296.
[Google Scholar]
-
49.
Choi ES, Jakubzick C, Carpenter KJ, Kunkel SL, Evanoff H, Martinez FJ, et al. Enhanced monocyte chemoattractant protein-3/CC chemokine ligand-7 in usual interstitial pneumonia.
Am. J. Respir. Crit. Care. Med. 2004,
170, 508–515.
[Google Scholar]
-
50.
Yanaba K, Komura K, Kodera M, Matsushita T, Hasegawa M, Takehara K, et al. Serum levels of monocyte chemotactic protein-3/CCL7 are raised in patients with systemic sclerosis: association with extent of skin sclerosis and severity of pulmonary fibrosis.
Ann. Rheum. Dis. 2006,
65, 124–126.
[Google Scholar]
-
51.
Katzen J, Wagner BD, Venosa A, Kopp M, Tomer Y, Russo SJ, et al. An SFTPC BRICHOS mutant links epithelial ER stress and spontaneous lung fibrosis.
JCI Insight 2019,
4, e126125.
[Google Scholar]
-
52.
Blanc RS, Kallenbach JG, Bachman JF, Mitchell A, Paris ND, Chakkalakal JV. Inhibition of inflammatory CCR2 signaling promotes aged muscle regeneration and strength recovery after injury.
Nat. Commun. 2020,
11, 4167.
[Google Scholar]
-
53.
Ong VH, Carulli MT, Xu S, Khan K, Lindahl G, Abraham DJ, et al. Cross-talk between MCP-3 and TGFbeta promotes fibroblast collagen biosynthesis.
Exp. Cell Res. 2009,
315, 151–161.
[Google Scholar]
-
54.
Szymczak WA, Deepe GS, Jr. The CCL7-CCL2-CCR2 axis regulates IL-4 production in lungs and fungal immunity.
J. Immunol. 2009,
183, 1964–1974.
[Google Scholar]
 Paul W. Noble
1,*
Dianhua Jiang
1,4,*
Paul W. Noble
1,*
Dianhua Jiang
1,4,*