1. Introduction
Natural compounds produced by microorganisms have been used to treat bacterial infections for more than 50 years and have been involved in increasing of the human life expectancy [
1]. Originally, the term antibiotic was used to refer to natural products of microorganisms. Then, antimicrobial chemotherapeutic agents were synthesized by relying on the structure of natural products [
2]. Thus, the term antibiotics includes natural products of microorganisms, semisynthetic compounds derived from natural products, and drugs chemically inspired by the structure of natural products [
1]. Two types of antibiotics exist. Bacteriostatic antibiotics inhibit bacterial growth without killing the bacteria by targeting protein and folate synthesis. Bactericidal antibiotics kill bacteria by targeting the cell-wall, cytoplasmic membrane, and DNA replication [
3]. Since the accidental discovery of penicillin in 1928 by Alexander Fleming [
4] and its appearance on the drug market, in 1941, as an important chemotherapeutic agent [
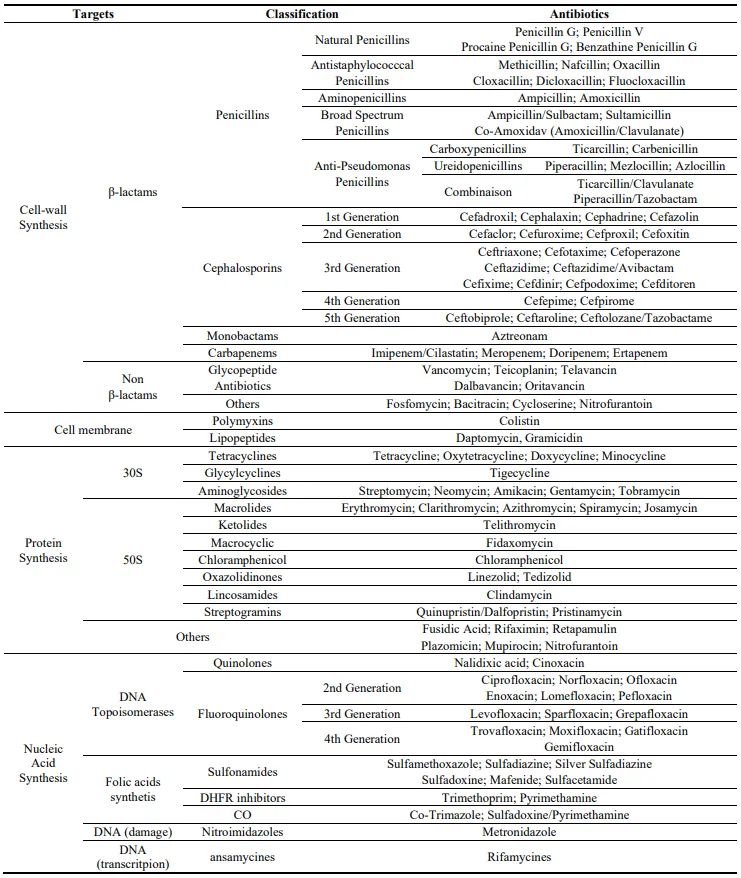
2], about 10–20 principal classes of antibiotics have been identified. These can be classified based on their mechanism of action, which targets components of bacteria such as cell-wall, cytoplasmic membrane, enzymes, folates and ribosomes.
Peptidoglycan is an essential component of the bacterial cell wall [
5] which is the target of β-lactam antibiotics. This class of antibiotics, including penicillins, cephalosporins, carbapenems, and monobactams, is most commonly used to treat bacterial infections [
6]. β-lactam antibiotics, known as inhibitors of the cell–wall synthesis of both Gram-negative and Gram-positive bacteria [
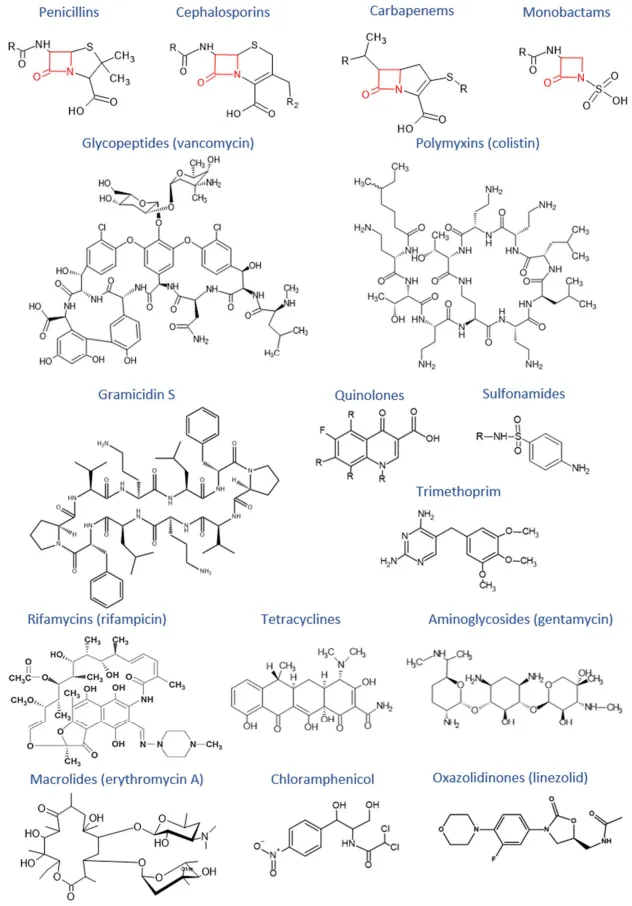
7], share a common 4-membered β-lactam ring containing an amide bond () [
8]. Their mechanism of action is based on covalent binding to penicillin-binding proteins (PBP) [
6], transpeptidase enzymes involved in the final step of peptidoglycan synthesis [
9,
10]. Similarly, glycopeptides, including vancomycin and teicoplanin, form the second class of antibiotics that inhibit the cell-wall synthesis of Gram-positive bacteria [
11,
12]. They are produced from
Actinomycetes bacteria and consist of cyclic or polycyclic heptapeptide cores that are generally glycosylated [
13].
The cytoplasmic membrane of Gram-positive and Gram-negative bacteria, located within the cell-wall, delimits the bacterial cytoplasm [
14] and is targeted by the antibiotics polymyxin [
15] and gramicidin [
16]. Polymyxins are natural antibiotics obtained by fermentation from different species of
Paenibacillus polymyxa [
14]. Five classes of polymyxins (A, B, C, D, E) are known, but only polymyxin B and E (colistin) are used clinically due to their lower nephrotoxicity [
15,
17]. Those medications are polycationic lipopeptides with 1200 Da consisting of a characteristic heptapeptide ring and a short prominent tripeptide bound to a fatty acid tail () [
18,
19]. The cationic moiety of these drugs makes them water soluble, while the hydrophobic part enables their incorporation into bacterial membranes readily [
20]. Although their mechanism of action is still poorly understood, it’s clear that polymyxins destabilize phospholipids and lipopolysaccharides (LPS) present in the outer membrane and stabilized by divalent cations as Ca
2+ and Mg
2+. In fact, the positively charged polymyxins interact with the negative charges localized on lipid A of LPS, generating displacement of cations from phosphate groups. As a result, destabilization and permeabilization of the outer membrane are observed. Then, polymyxins insert into the cytoplasmic membrane causing the cell lysis [
15,
21,
22]. Another class of antibiotics called gramicidin is involved in the disturbance of the cytoplasmic membrane. Discovered in 1942, Gramicidin S is an ionophore antibiotic produced by
Bacillus brevis [
23]. Its structure is based on a cyclodecapeptide comprised of two comparable pentapeptides with the head attached to the tail () [
24]. Gramicidin S acts on the cell membrane by disrupting the structure of the lipid bilayer and generating the appearance of pores or other localized anomalies [
25,
26]. However, Gramicidin S also shows hemolytic activity which limits its use as a topical antibiotic [
27].
The DNA replication is a process in which both identical DNA molecules are formed from an original DNA molecule [
28]. Quinolones inhibit DNA synthesis by targeting topoisomerase enzymes, more specifically DNA gyrase and topoisomerase IV [
29]. Emerged in the 1960’s for treating Gram-positive and Gram-negative bacterial infections, quinolones are synthetic antibiotics whose structure contains a bicyclic core () [
30]. Quinolones bind in a non-covalent manner to the enzyme-DNA cleavage complex, presenting a double strand break, and prevent the ligation of DNA fragments [
31]. In this way, DNA strands cannot assemble anymore, leading to inhibition of the replication process and consequent slow cell death. Moreover, quinolones can interfere with the positioning of the topoisomerase on DNA, making the strand-break free. If the strand-break remains unfixed, the chromosomal fragmentation and subsequent rapid cell death will occur. Furthermore, the presence of breaks and cleavage complexes in the DNA molecule can generate ROS (Reactive Oxygen Species). The accumulation of these toxic molecules can lead to more DNA breaks and eventually rapid cell death [
32,
33].
Folates are used by prokaryotic and eukaryotic cells for metabolic processes, including the synthesis of nucleic basis, particularly purine and thymine, and the synthesis of methionine, which are essential for DNA replication and transcription. Due to the presence of a pathway for folate synthesis in bacteria and its absence in mammalian cells, folate synthesis is an interesting target for antimicrobial agents. Sulfonamides are antibiotics composed of a sulfonamide group within their structure () and involved in the inhibition of folate biosynthesis. The molecular structure of sulfonamide is close to
p-aminobenzoic acid, which is one of the constituting cores of folate [
34], which may explain these mechanism of action. This class of antibiotics targets the last step of folate synthesis [
35]. Trimethoprim is a synthetic antibiotic used to treat various bacterial infections such as urinary tract ones [
36]. This antibiotic is described as an inhibitor of DHFR (dihydrofolate reductase), an enzyme involving in the last step of folate synthesis [
34]. Thus, trimethoprim blocks folate synthesis and, therefore, hinders nucleic acids synthesis in bacteria.
Transcription is the process by which mRNA is synthetized from DNA strand by RNA polymerase. This process includes three steps: Initiation, elongation, and termination [
37]. Some antibiotics are able to act on these stages of transcription. Rifamycins, as rifampicin, belonging to the family of ansamycin antibiotics, are naturally produced by the bacterial strain
Amycolatopsis mediterranei. They have a particular basket-like structure obtained by the junction of non-adjacent positions of the aromatic moiety via an aliphatic chain forming an ansa loop (). The aromatic side is composed of a napthalene, a naphtoquinolone, a benzene, or a benzoquinolone ring [
38,
39]. The antibacterial activity of rifamycin interlinks with its capacity to interfere with the initiation step by binding to the β-subunit of RNA polymerase. It leads to disruption of the binding of β-subunit to DNA. This prevents the elongation of mRNA, leading to cell death [
39].
Tetracyclines and aminoglycosides inhibit protein synthesis by linking with high affinity to the 30S ribosomal subunit. Other classes of antibiotics inhibit protein synthesis by linking to the 50S ribosomal subunit, such as macrolides, chloramphenicol, and oxazolidinones. Tetracyclines, discovered in the 1940s, present a chemical structure composed of linear merged tetracyclic cores containing a large number of substituents (). The mechanism of action of tetracyclines is based on strong link with the small ribosomal subunit and prevention of the link of the tRNA at the A site. Thus, the elongation step is inhibited leading to the stop of the polypeptide chain growth [
40]. Aminoglycosides, as gentamycin, are natural or semisynthetic antibiotics derived from
Actinomycetes. They are formed by a central structure of amino sugars linked by glycosidic bonds to a 2-deoxystreptamine () [
41]. They also link to the A site of the 30S ribosomal subunit. Thus, aminoglycosides induce a false codon leading to a false translation. Thereupon, a false amino acid is added to the polypeptide chain damaging the cell membrane and entailing the cellular death [
41]. Macrolides are constituted by a macrocyclic lactone core of 12 or more components with deoxy–sugars attached to the ring (). These antibiotics bind to the 50S ribosomal subunit conducing to the protein synthesis shutdown [
42,
43,
44]. Chloramphenicol is a natural antibiotic produced by
Streptomyces venezuelae [
45]. This drug is an organochloride compound formed by a nitrobenzene core linked to a dichloroacetamide group through the two carbon atoms substituted by two alcohol functions () [
46,
47]. Its mechanism of action is the same as that of macrolides. This antibiotic binds to the 50S ribosomal subunit and avoid the transpeptidation by the peptidyltransferase enzyme [
48]. Oxazolidinones, as linezolid, are antibiotics containing a 2-oxazolidone in their structure (). Oxazolidinones are used against Gram-positive bacteria by binding to P site of the 50S ribosomal subunit and inhibiting the initiation complex [
49]. Thus, the formation of the ribosome is prevented. However, if the ribosome is already formed, the link of oxazolidinones on the ribosome inhibits the translocation of the peptide chain from the A site to the P site during the peptide bond formation [
50,
51]. These data are compiled in .
. Antibiotic classes and their targets. Adapted from [
52].
. Structures of antibiotics.
2. Bacterial Resistance to Antibiotics
Unlike antiseptics used to cure microorganism infections, antibiotics are specific to bacteria and, as a result, they are only used to treat bacterial infections from both Gram-negative and Gram-positive bacteria [
52]. However, the previous decades have been marked by the emergence of bacterial antibiotic resistance due to the massive consumption of antibiotic exerting selective pressure on bacterial genome [
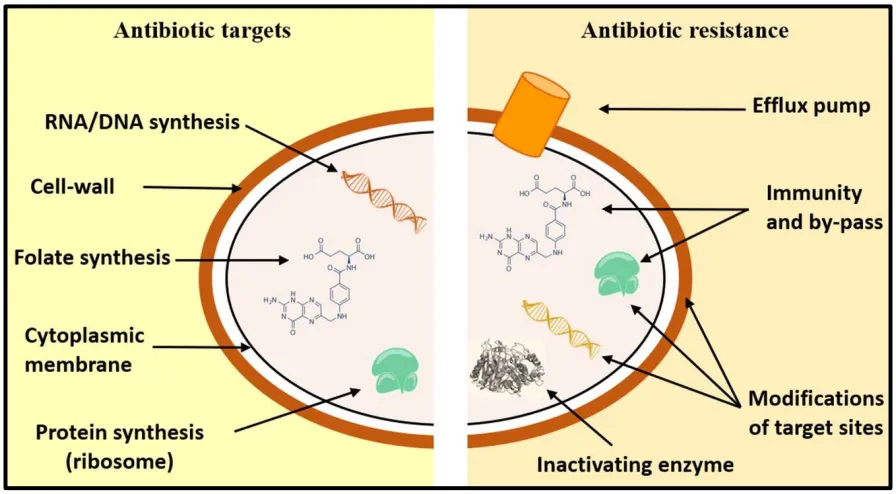
53]. This issue spearheads different kinds of resistance mechanisms in bacteria to escape the lethal effects of antimicrobial substances such as the modification of the antibiotic molecule, loss of antibiotic permeability and greater efflux, and modification of target sites ().
. Comparison of antibiotic targets and mechanisms of antibiotic resistance. Adapted from [
54].
The production of enzymes, able to introduce chemical modifications on the antibiotic, is a well-known mechanism in the antibiotic resistance acquisition in Gram-positive and Gram-negative bacteria. Most antibiotics concerned by this type of mechanism perform their activity by inhibiting the protein synthesis as chloramphenicol and aminoglycosides. Acetylation, phosphorylation, and adenylation are biochemical reactions most currently catalyzed by enzymes leading to an important steric hindrance decreasing the affinity of the drug for its target [
55].
The mechanism of antibiotic destruction involves, in particular, β-lactam antibiotics. β-lactamase enzymes are able to hydrolyze the β-lactam ring of antibiotics to destroy the amide bond and make the drug inactive [
55,
56,
57]. These hydrolytic enzymes are divided into two classes: Ser β-lactamases and metallo-β-lactamases. Ser β-lactamases form an enzyme-antibiotic intermediate quickly hydrolyzed, while metallo-β-lactamases use Zn
2+ ion to activate the β-lactam ring hydrolysis [
58,
59,
60].
Porins are membrane proteins capable of transporting antibiotic molecule inside of the Gram-negative bacterial cells by diffusion [
61]. However, bacteria developed many mechanisms to avoid antibiotics to achieve their cytoplasmic and periplasmic targets by decreasing the antibiotic absorption by porins. Therefore, mutations on genes coding for porins are responsible of a decrease of the outer membrane permeability to antibiotics. These mutations can lead to porin loss, change of the size or the conductance of the porin channel, or a decrease of the number of porins within the membrane. These modifications lead to the lowest diffusion of the antibiotic in the cell and thus to lower capacity to kill bacteria [
62,
63].
Discovered in the 1980s in
E. coli, efflux pumps are located and encased within the bacterial plasma membrane of both Gram-positive and Gram-negative bacteria. These bacterial complexes are able to recognize toxic molecules that have penetrated the bacterial cell–wall and reached the periplasm or the cytoplasm, and bring them out before they reach their targets [
64]. These systems affect a wide range of antibiotics including inhibitors of protein synthesis, quinolones, β-lactams, and polymyxins [
62].
A common strategy for bacteria to develop an antibiotic resistance is to avoid the antibiotic action by interfering with the target site. In this purpose, bacteria has developed many strategies, including the protection of targets and the modification of the target site that lead to a decrease of the affinity for the antibiotic [
65].
• The alteration of ribosomal subunits leads to drug resistance that affects the protein synthesis.
• The alteration of PBP is a favored mechanism for Gram-positive bacteria resistance. Mutations in PBP lead to a decrease of the β-lactam antibiotic affinity to PBP.
• The alteration of cell-wall precursors consists of a modification of D-Ala-D-Ala sequence to a D-Ala-lactate sequence. Glycopeptides cannot link to residues, resulting in a resistance development.
• The mutation of DNA gyrase and topoisomerase IV leads to a bacterial resistance to quinolones. The resistance mechanism implies modifications of two enzymes. It leads to a failed replication and, so, quinolones cannot bind with enzymes.
• Ribosomal protection mechanisms induce a bacterial resistance to tetracyclines.
• RNA polymerase modifications make bacteria resistant to rifampicin.
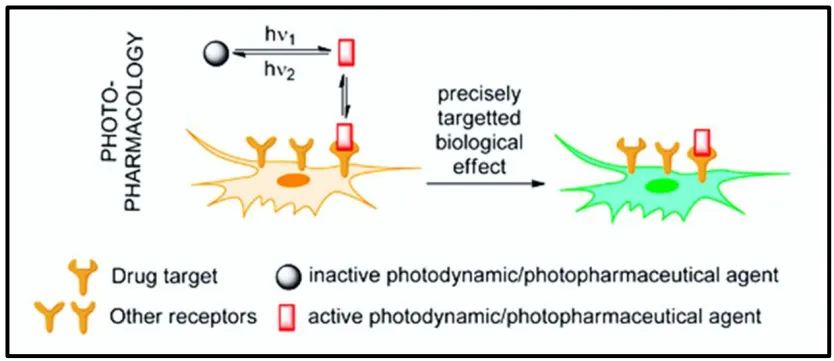
3. Useful Photopharmacology
The spread of bacterial antibioresistance is favored by selection pressure, related to the repeated exposure of environmental microbial populations to active principles released in the environment and via mechanisms of gene resistance transfer [
66]. To combat this problem, according to the “one health” concept whereby the human, animal and environmental health are closely related [
67], photopharmacology, derived from the combination of photochemistry and pharmacology, is a promising interdisciplinary approach [
68]. This technology is an effective way to photoregulate many essential biological processes [
69,
70,
71,
72], such as structure and function of nucleic acid [
73] and peptides [
74], DNA transcription and translation [
75], protein fold [
76], enzymatic activity [
77], protein–ligand interactions [
78], membrane transport [
79], and modulation of receptors [
57,
70,
71]. Photopharmacology is based on the use of light as an external stimulus to activate drug reversibly [
74,
75]. First, the medicine is irradiated at the wavelength of the incident light stemming from a first source, leading to the drug activation as well as a reversible modification of structure and properties of the compound [
80]. After the interaction with its target, the drug is irradiated at the wavelength of the incident light stemming from a second source making the drug inactive () [
57,
74,
79]. One of the most generally applied strategy for adding a light-responsive moiety to an antibiotic molecule is its functionalization with a molecular photoswitch [
81] which offers a high level of spatiotemporal resolution and a precise control of the light wavelength and intensity [
82].
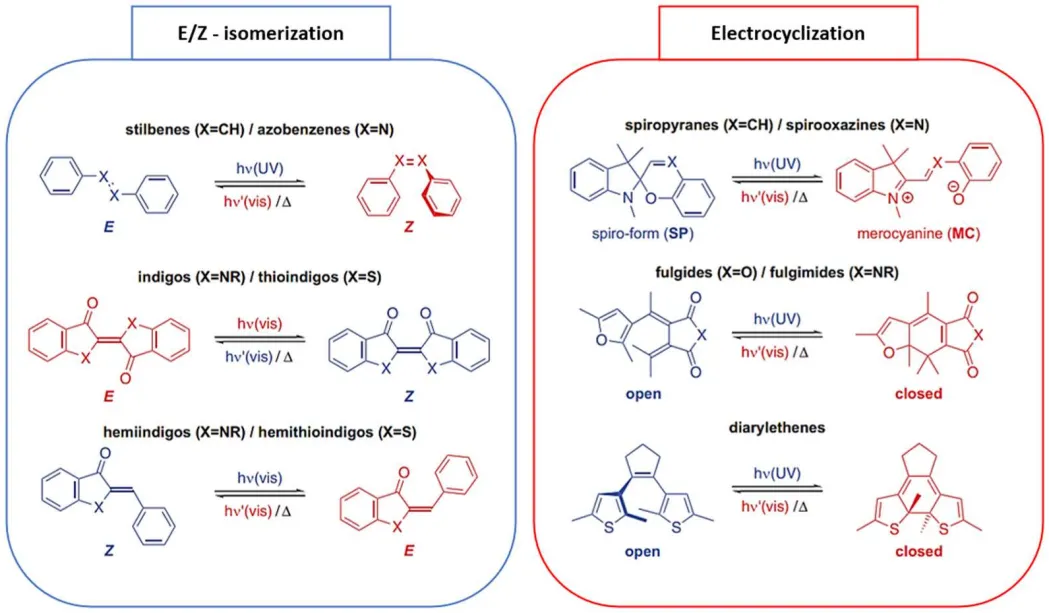
Recently, several classes of photochroms have been developed and their use on photopharmacology has been widely reported in literature [
67,
83,
84,
85,
86]. Thus, azobenzenes/stilbenes [
83,
84,
85], spiropyrans/spirooxazines [
86,
87,
88,
89,
90], hemiindigos/hemithioindigos [
91,
92], fulgides/fulgimides [
93,
94,
95], and diarylethenes [
96,
97] are the most used photochromic compounds for biomolecules functionalization (). These photoresponsive moieties can be divided into two groups according to their isomerization mode. Azobenzenes/stilbenes [
98,
99,
100,
101], indigos/thioindigos [
102,
103,
104], and hemiindigo/hemithioindigo [
104,
105,
106] isomerize via an E/Z isomerization mode (, blue box) [
107,
108,
109]. Spiropyrans/spirooxazines, fulgides/fulgimides and diarylethenes [
75,
107], isomerize via an electrocyclization reaction (, red box). As the result, a modification of the geometry of the photoswitch is observed. Moreover, the modification of certain photoswitches conformation as azobenzenes, spiropyrans and diarylethenes, leads to polarity and charges changes within the molecule [
110,
111].
. The concept of photopharmacology. The isomerization of the photopharmaceutical agent, induced by the light, leads to the activation of the drug which can interact with its target [
112].
. Molecular photoswitches based on E/Z-isomerization (left box) and electrocyclization (right box) [
113].
4. Coupling between Photoswitches and Antibiotics
Although the coupling between photoswitches and antibiotics is recent, it was used to fight bacterial antibiotic resistance. Incorporation of a photochromic group, such as azobenzenes, diarylethenes and spiropyrans, on a classical antibiotic molecule enables the design of photoresponsive antibiotics of which the antibacterial capacity can be regulated by a light irradiation [
81,
114,
115,
116]. Thus, several research teams studied this photopharmacologic approach. Among them, Feringa et al. demonstrated the efficiency of a modified-trimethoprim on bacterial growth inhibition in
E. coli [
117,
118]. They also worked on ciprofloxacin and norfloxacin on which they incorporated different photoswitches such as azobenzenes and spiropyrans [
119,
120] to evaluate the antibacterial activity of photoresponsive molecules. Furthermore, Fu et al. [
121,
122] also used azobenzenes and coupled them with a fluoroquinolone antibiotic via an amide bond. Moreover, Zhang et al. [
123] and Li et al. [
124] used dithienylethenes as photoswitches to couple them with a fluoroquinolone antibiotic whereas Babii et al. [
125] incorporated a diarylethene photoswitch with Gramicidin S antibiotic.
4.1. Coupling between Azobenzenes and Antibiotics
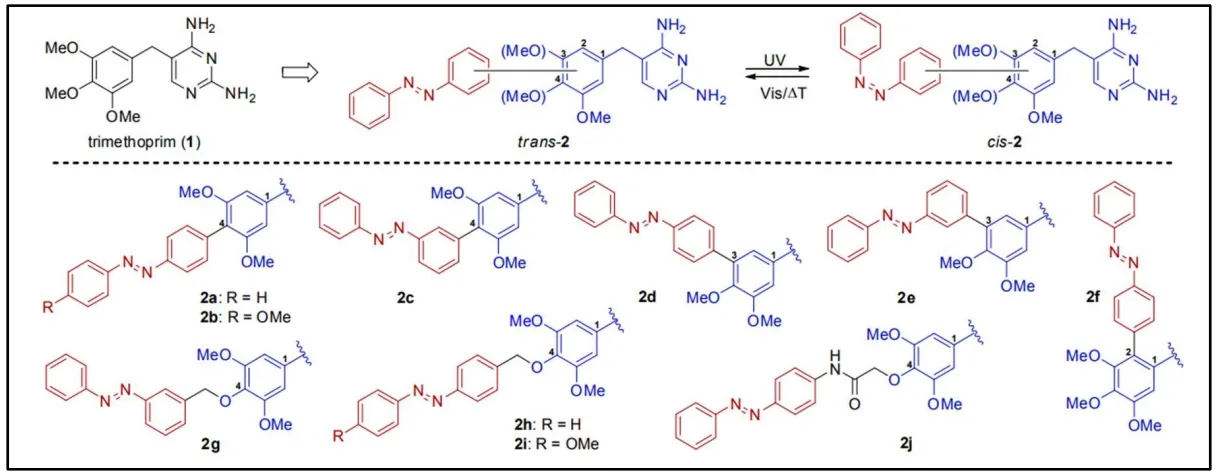
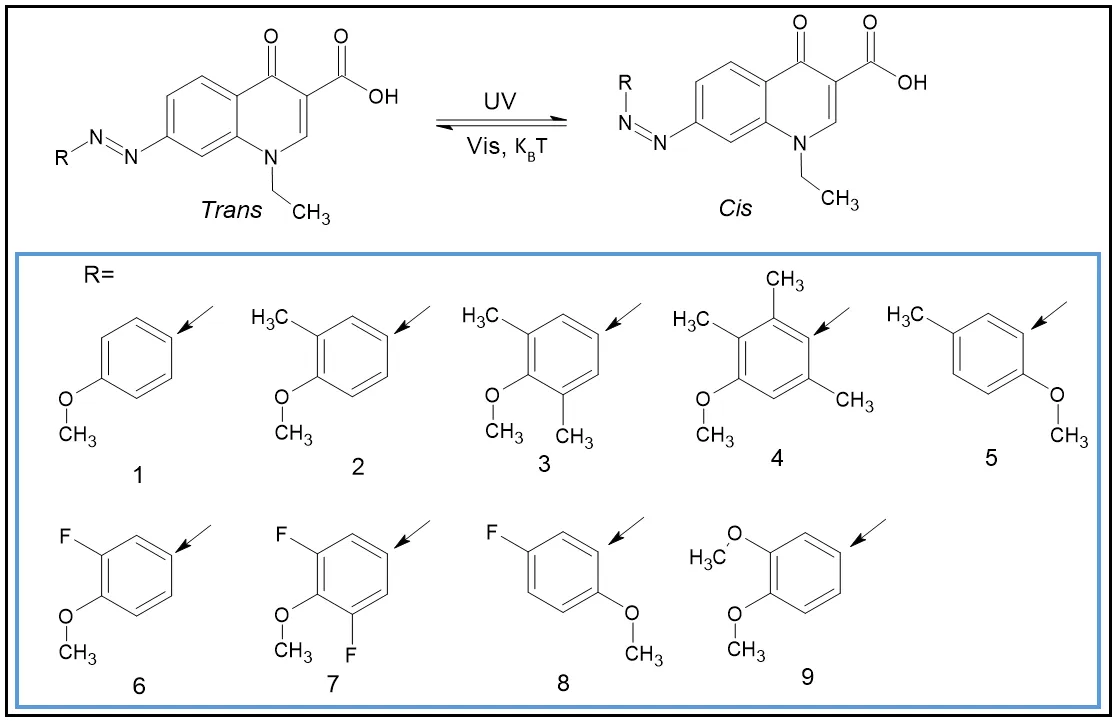
Trimethoprim is a synthetic antibiotic acting as a DHFR inhibitor to block the folates synthesis in bacteria. Wegener et al. [
117] synthetized photoresponsive analogues of trimethoprim to control their activity in a spatio-temporal manner by the light. In this way, they synthetized azobenzenes and incorporated them to the 2, 3 or 4 positions of the methoxyphenyl group of the trimethoprim by Suzuki cross-coupling (
2a–f), or by hydroxyl group alkylation at the 4 position via linkers (
2g–j). When the compound was irradiated with a UV light, trans-azobenzene isomerized to cis-azobenzene. Inversely, when the cis-azobenzene was exposed to visible light, or thermally, it returned to its trans conformer ().
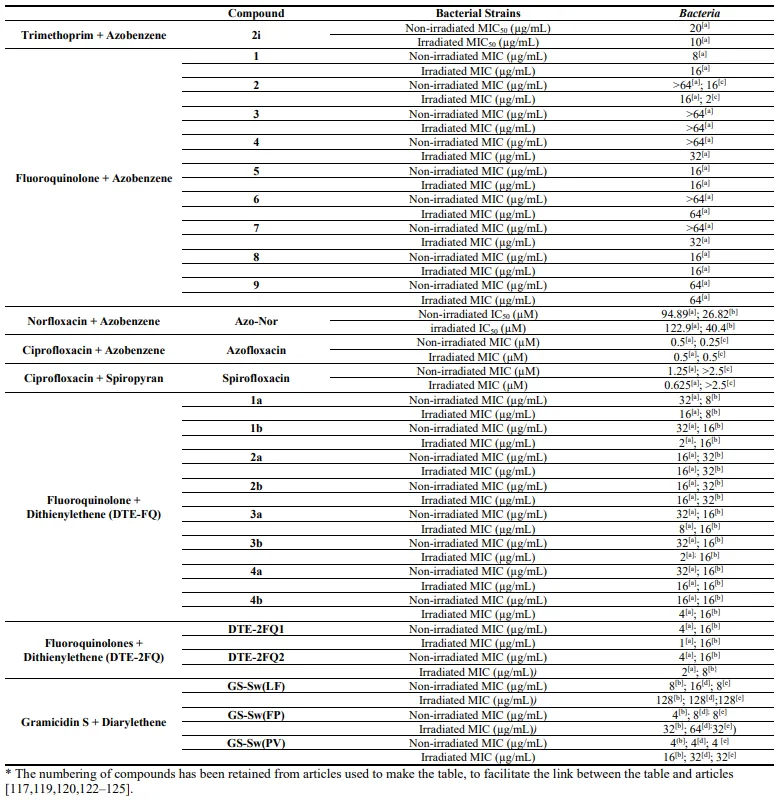
All compounds were tested on
E. coli before and after irradiation with UV light at 365 nm and the authors determined the minimal inhibitory concentration of molecules at which 50% of bacterial growth was inhibited (MIC
50). Among all compounds,
2f and
2j were mostly inactive up to a concentration of 20 µM before and after irradiation. However, other molecules were active on
E. coli with different degrees of efficacy. If, for most of compounds, the light irradiation did not improve the antibacterial activity, irradiated compound
2i showed a better antibacterial activity (MIC
50 = 10 µM) than the non-irradiated compound (MIC
50 = 20 µM) attesting that
trans-
cis isomerization increased the antibacterial activity. The attainment of compounds coming from the coupling of azobenzene photoswitches and antibiotics is the most commonly used coupling method. Feringa et al. widely worked on this subject. In a first publication [
112], they brought the proof of concept whereby the use of photoactivable antibiotics enabled the light-control of the bacterial growth. In this way, they obtained a photoactivable antibiotic by substituting the piperazine ring of a quinolone by an aryldiazo moiety bearing different substituents (). The addition of an aryldiazo moiety made the compound photoswitchable. Under a UV-light irradiation at 365 nm, the trans-isomer was converted into the
cis-isomer. Inversely, under a visible-light irradiation, the
cis-isomer was converted into the trans-isomer. The
cis-
trans conversion was also possible at room temperature ().
The efficiency of photoswitchable compounds
1–
9 was tested, before and after UV-light irradiation by determining their MIC on
E. coli strain. Among the compounds,
2,
4 and
7 showed an increase of antimicrobial activity after irradiation compared to the antimicrobial activity of non-irradiated compounds (). The compound
2 showed the higher discrepancy between the antibacterial activity before irradiation (MIC > 64 µg/mL) and the antibacterial activity after irradiation (MIC = 16 µg/mL). The compound
2 was, then, tested on
M. luteus. The antimicrobial activity after UV-light irradiation at 365 nm (MIC = 2 µg/mL) was 8 times higher than the antimicrobial activity before irradiation (MIC = 16 µg/mL). Thus, the UV-light irradiation on photoswitchable antimicrobial agents enabled the
trans→
cis isomerisation which led to an increase of molecules potency whereas the visible-light irradiation enabled the cis→trans conversion which led to a large decrease of the antibiotic activity.
. Structurally diverse set of UV light-responsive diaminopyrimidines [
117].
. MIC values of all compounds * stemming from the incorporation of a photoswitch with an antibiotic before and after irradiation. MIC values determined for
E. coli [a],
S. aureus [b],
M. luteus [c],
S. epidermidis [d],
S. xylosus [e] (values from [
117,
119,
120,
122,
123,
124,
125]).
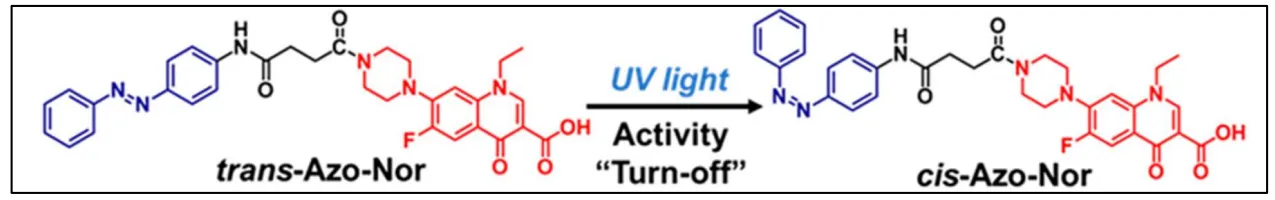
Fu et al. [
122] coupled a commercial fluoroquinolone, norfloxacin, with a photochromic azobenzene. Norfloxacin is formed by a piperazine ring with a free amino group that can be used to be linked to an azobenzene. Thus, they synthetized a photoswitchable antibacterial agent by conjugating azobenzene (blue) to the amino group of the piperazine ring of norfloxacin (red) via a linker (black) (). The incorporation of the molecular azobenzene photoswitch on the antibiotic enabled the drug activity control by the light. Indeed, under UV light irradiation, the
trans→
cis azobenzene isomerization occured whereas the
cis→
trans azobenzene isomerization happened under visible light irradiation. To evaluate the capability of the compound to control the antibacterial activity, it was tested on
E. coli and
S. aureus by determining the corresponding half maximal inhibitory concentration (IC
50). Contrary to the results of Feringa et al, the tests showed a large decrease of the antimicrobial activity after UV-light irradiation on both bacterial strains. In
E. coli, the antimicrobial activity was IC
50 = 94.89 µM before irradiation and IC
50 = 122.9 µM after irradiation. Similarly, in
S. aureus the antimicrobial activity before irradiation was IC
50 = 26,82 µM and IC
50 = 40.40 µM after irradiation. So, the antimicrobial effect can be turned-off under UV light.
The use of an amide linker to couple a fluoroquinolone with a photoswitch unit was used and conduced to coupled compounds. Thus, Velema et al. [
120] worked on the coupling between ciprofloxacin and either an azobenzene, or, a spiropyran photoswitch to compare both obtained compounds. To synthetize the compounds, carboxylic acid-modified spiropyran and azobenzene were used. The corresponding acyl chlorides were obtained and conjugated to the amine of the piperazine ring constituting ciprofloxacin. It led to compounds named spirofloxacin and azofloxacin (A) [
120]. Spirofloxacin and azofloxacin isomerized under UV light irradiation at 365 nm by two processes. Indeed, spirofloxacin was composed of a spiropyran photoswitch having an electrocyclization isomerization process. Under a UV light irradiation, the ring-closed isomer was converted into the open-ring isomer also called merocyanine. Under visible light irradiation or thermally, the open-ring isomer was converted into the ring-closed isomer (B). Furthermore, azofloxacin, was composed of an azobenzene photoswitch unit, having a
trans/
cis isomerization process. Indeed, under a UV light irradiation, the
trans→
cis azobenzene isomerization occured whereas under visible-light irradiation or thermally, the
cis→
trans azobenzene isomerization happened (C).
. Isomerization process and synthesis of photoswitchable compounds from quinolone. Adapted from [
119].
. Structure and isomerization process of the compound Azo-Nor [
122].
. Syntheses and isomerization processes of azofloxacin and spirofloxacin [
120].
After the synthesis, the researchers compared the antibacterial activity of spirofloxacin with the antibacterial activity of azofloxacin by determining the MIC of compounds on
E. coli and
M. luteus. They also tested the antibacterial activity of the ciprofloxacin alone to use it as control. Without irradiation, spirofloxacin showed a MIC of 1.25 µM on
E. coli, whereas, under UV light irradiation at 365 nm, the value of MIC was 0.625 µM. Moreover, there was no difference of activity before (MIC > 2.5 µM) and after (MIC > 2.5 µM) irradiation on
M. luteus. In the case of azofloxacin, the antibacterial activity was the same before and after UV light irradiation (MIC = 0.5 µM) on
E. coli. However, on
M. luteus, a significant difference between antibacterial activities before and after UV light irradiation was observed. Indeed, the non-irradiated azofloxacin showed a better antibacterial activity (MIC = 0.250 µM) compared to the antibacterial activity of the irradiated azofloxacin (MIC = 0.5 µM). Thus, the
trans-isomer of azofloxacin had a better antibacterial activity when it was non-irradiated. Nevertheless, it’s interesting to note that the antibacterial activity of azofloxacin was 50 times lower than the antibacterial activity of the control (MIC = 12 µM) on
M. luteus.
4.2. Coupling between Diarylethene and Antibiotics
The synthesis of new compounds composed of an antibiotic and a photochromic unit different than azobenzene were developed. Thus, Li et al. [
124] worked on the synthesis of a dithienylethene bearing a carboxylic acid group to use it as a photochrom and coupled it with ciprofloxacin or norfloxacin antibiotics. Indeed, the peptide coupling between the amine of the piperazine ring of fluoroquinolone and the carboxylic acid group of dithienylethene was realized, by using coupling agents HATU and DIPEA, to obtain products
1–
4 ().
. Synthesis route of switchable antibacterial agents
1–
4. Adapted from [
122].
The isomerization capacity of all compounds was tested at room temperature. It was found that dithienylethene compounds
1–
4 isomerized by an electrocyclization process. The irradiation by a UV-light at 254 nm of the open-ring isomer led to the ring closed isomer. Inversely, upon a visible light irradiation (ʎ > 402 nm), the ring-closed isomer underwent a cycloreversion reaction leading to the initial open-ring isomer.
The antibacterial properties of all compounds were tested before and after irradiation at 254 nm by determining their MIC on
E. coli and
S. aureus (). Thus, the photoresponsive antimicrobial molecules
1–
4 had a similar activity before the UV-light irradiation on
E. coli (MIC = 16–32 µg/mL) and
S. aureus (MIC = 8–32 µg/mL). Moreover, all closed-ring isomers did not show a difference of activity in comparison with opened-ring isomers on
S. aureus (MIC = 8–32 µg/mL). On
E. coli, all compounds, except
2a and
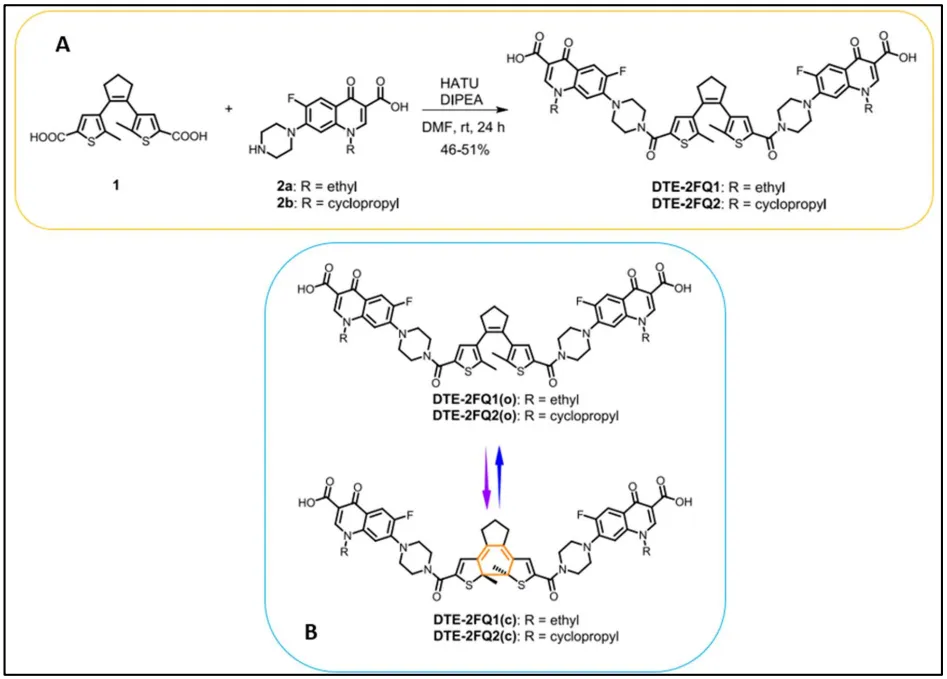
2b showed an increase of the antibacterial activity after the light irradiation (MIC = 2–16 µg/mL). The observed results confirmed that the light irradiation can modulate the antibacterial activity of compounds, but in a weakly efficient manner in comparison with native fluoroquinolone activity (MIC = 0.125 µg/mL). To increase the antimicrobial activity of the previous compound, Zhang et al. [
123] synthetized a molecule formed by a dithienylethene (DTE) linked to two fluoroquinolone units (FQ) via an amide bond. In this way, they designed
DTE-
2FQ1 and
DTE-
2FQ2 compounds by condensation reaction between carboxylic groups of dithienylethenes and amine of piperazine ring from norfloxacin and ciprofloxacin, respectively, by using coupling agents HATU and DIPEA (). These two compounds presented an electrocyclization process of isomerization. Indeed, under a UV-light irradiation at 254 nm, the open-ring isomer isomerized into the ring-closed isomer. Inversely, under a blue light irradiation at 460–470 nm, ring-closed isomer underwent a cycloinversion reaction which led to the initial open-ring isomer ().
. (
A) Synthesis of Diethienylethene-bridged Fluoroquinolones (DTE-2FQ); (
B) isomerization process of compounds
DTE-
2FQ1 and
DTE-
2FQ2 [
123].
Both compounds were tested on
E. coli and S. aureus before and after UV light irradiation by determining their MIC (). Before UV light irradiation,
DTE-
2FQ1 and
DTE-
2FQ2 had lower antimicrobial activities on
E. coli (MIC = 4 µg/mL) and S. aureus (MIC = 16-32 µg/mL) compared with ciprofloxacin and norfloxacin (MIC = 0.125 µg/mL). This observation was probably due to the DTE switch which limited the access to bacteria. However,
DTE-
2FQ1 and
DTE-
2FQ2 including two fluoroquinolone molecules showed an improvement of the antibacterial activity especially on
E. coli compared to the corresponding mono-fluoroquinolone molecules (
DTE-FQ1 and
DTE-FQ2) [
124]. Indeed, before UV light irradiation,
DTE-
2FQ1 (MIC = 4µg/mL) had a higher potency than its analogue
DTE-FQ1 (MIC = 32 µg/mL). In the same way,
DTE-
2FQ2 (MIC = 4 µg/mL) had a higher potency than its analogue
DTE-FQ2 (MIC = 8 µg/mL). Moreover, a difference of antimicrobial activity for both compounds on
E. coli was observed after UV light irradiation. However, the most interesting was the difference of activity of
DTE-
2FQ1 compound which showed a four times higher antimicrobial activity after irradiation (MIC = 1 µg/mL) compared to its activity before irradiation (MIC = 4 µg/mL). Nevertheless, on S. aureus, the difference of antimicrobial activity before and after irradiation was negligible. Finally, this work showed that the introduction of a DTE photoswitch enabled an increase of antimicrobial activity after the UV light irradiation of compounds. However, compounds activities remained low in comparison with the native antibiotics.
4.3. Coupling between Diarylethenes and Antibiotics
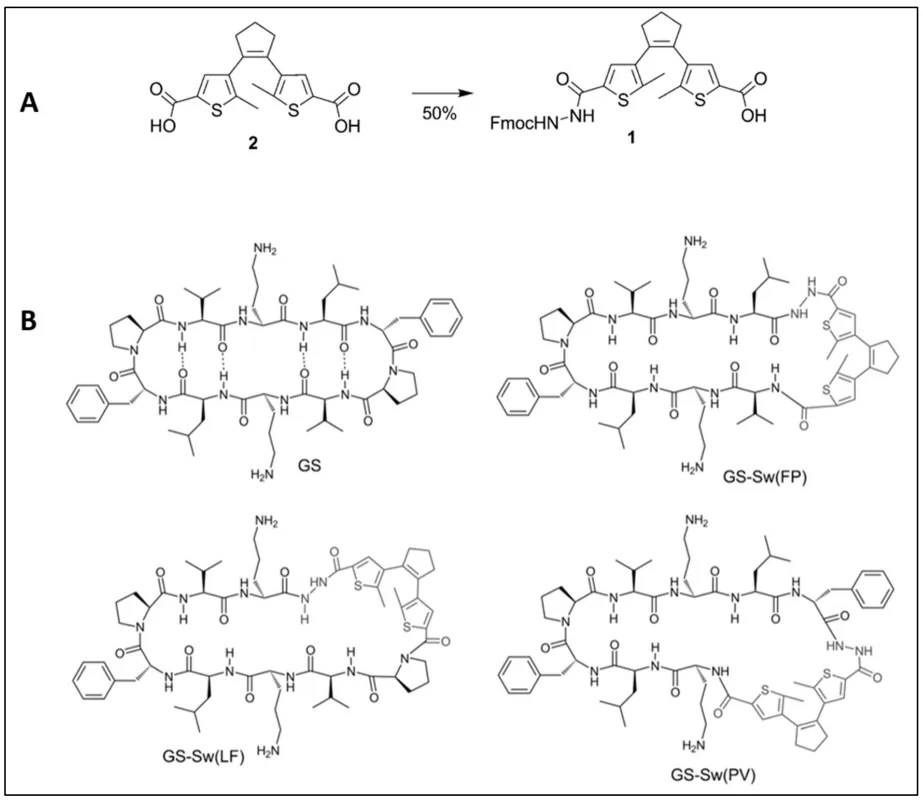
Babii et al. [
125] worked on the synthesis of Gramicidin S analogues containing a diarylethene unit to control its activity by the light. In this way, they firstly synthetized the photochromic molecule from a diarylethene carboxylic acid 2 by protecting one of both carboxylic acid group with Fmoc–NH–NH
2. Thus, they obtained a protected diarylethene-based amino acid analogue 1 in one step (A). Then, they selected three nonpolar dipeptide units of Gramicidin S (L
DF,
DFP, and PV) and replaced it with the photoswitchable diarylethene (Sw). Therefore, they obtained three different photoresponsive cyclic peptides (GS-Sw(LF), GS-Sw(FP), and GS-Sw(PV)) that mimic Gramicidin S (B). Moreover, the incorporation of a diarylethene photoswitch in Gramicidin S enabled the compounds isomerization. Diarylethenes showed an electrocyclization photoisomerization process which involved the transition between opened and closed photoswitches. Under UV light irradiation at 256 nm the compound isomerized into its ring-closed isomer from its open-ring isomer. Inversely, under visible light irradiation at 530 nm, the ring closed isomer underwent a cycloreversion and isomerized into the open-ring isomer.
. (
A) Synthesis of the photoswitching building block.Conditions: Fmoc–hydrazine, DIC, DIPEA, DMF, 8 h. Fmoc—9-fluorenylmethyloxycarbonyl, DIC—1,3-diisopropylcarbodiimide, DIPEA—
N,
N-diisopropylethylamine, DMF—
N,
N-dimethylformamide. (
B) Gramicidin S (GS) and its three analogues; the photoswitchable peptidomimetics GS-Sw(LF), GS-Sw(FP), and GS-Sw(PV) [
125].
The three Gramicidin S analogues were tested on different Gram-positive bacteria by determining their MIC. The results showed that all analogues exhibited antimicrobial activity at 256 nm without UV light irradiation and had similar antibacterial activity to Gramicidin S (). Under UV light irradiation, the compounds isomerized and the ring-closed isomer were less active than the open-ring isomer for all bacteria strains tested. Moreover, in some concentration ranges, the analogues were able to suppress the bacterial growth without light irradiation, while they were completely inactive under UV light irradiation.
5. Conclusions
Antibiotics are widely used to treat bacterial infections in animals and human. However, their excessive use is a cause for concern because of the development of high bacterial resistance due to the accumulation of antibiotics in the environment. Therefore, the use of photopharmacology seems to be a promising way to combat this problem. This approach, based on the coupling of photochromic compounds such as azobenzenes, spiropyrans, and diarylethenes, with antibiotics, was applied to determine the effects of light use on the antibacterial activity of the obtained compounds. For most of the evaluated compounds on E. coli, light exposure allowed an increase in their antibacterial activity (). However, the compounds derived from the coupling between norfloxacin and azobenzene showed a loss of antibacterial activity on E. coli and S. aureus throughout light irradiation. Similarly, Gramicidin S derivatives showed a decrease in antibacterial activity upon light irradiation on many Gram-positive bacteria (). Thus, the use of light is an interesting way to enhance or decrease the antibacterial activity of compounds. Consequently, light is an attractive alternative to control drug activity and thus to limit the antibiotic resistance in bacteria.
Acknowledgments
The authors would like to thank Université de Technologie de Compiègne (UTC), ESCOM Chimie and Junia for their funding, as well as the MSTD funding from UTC.
Author Contributions
Conceptualization, A.A. and E.L.; Methodology, A.A.; Validation, M.V., A.F. and M.B.; Writing—Original Draft Preparation, A.A.; Writing—Review & Editing, E.L.; Visualization, M.B. and M.V.; Supervision, A.F.; Project Administration, E.L. Funding Acquisition, E.L.
Funding
This research was funded by [MSTD] grant number [2021].
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Demain AL. Antibiotics: Natural products essential to human health.
Med. Res. Rev. 2009,
29, 821–842.
[Google Scholar]
-
2.
Waksman SA. What Is an Antibiotic or an Antibiotic Substance?
Mycologia 1947,
39, 565–569.
[Google Scholar]
-
3.
-
4.
Tan SY, Tatsumura Y. Alexander Fleming (1881–1955): Discoverer of penicillin.
Singapore Med. J. 2015,
56, 366–367.
[Google Scholar]
-
5.
Vollmer W, Blanot D, De Pedro MA. Peptidoglycan structure and architecture.
FEMS Microbiol. Rev. 2008,
32, 149–167.
[Google Scholar]
-
6.
Bush K, Bradford PA. β-Lactams and β-Lactamase Inhibitors: An Overview.
Cold Spring Harb. Perspect. Med. 2016,
6, a025247.
[Google Scholar]
-
7.
Sarkar P, Yarlagadda V, Ghosh C, Haldar J. A review on cell wall synthesis inhibitors with an emphasis on glycopeptide antibiotics.
MedChemComm 2017,
8, 516–533.
[Google Scholar]
-
8.
-
9.
Lima LM, da Silva BNM, Barbosa G, Barreiro EJ. β-lactam antibiotics: An overview from a medicinal chemistry perspective.
Eur. J. Med. Chem. 2020,
208, 112829.
[Google Scholar]
-
10.
Edoo Z, Arthur M, Hugonnet JE. Reversible inactivation of a peptidoglycan transpeptidase by a β-lactam antibiotic mediated by β-lactam-ring recyclization in the enzyme active site.
Sci. Rep. 2017,
7, 9136.
[Google Scholar]
-
11.
Wilhelm MP. Vancomycin.
Mayo Clin. Proc. 1991,
66, 1165–1170.
[Google Scholar]
-
12.
Shea KW, Cunha BA. Teicoplanin.
Med. Clin. N. Am. 1995,
79, 833–844.
[Google Scholar]
-
13.
Butler MS, Hansford KA, Blaskovich MAT, Halai R, Cooper MA. Glycopeptide antibiotics: Back to the future.
J. Antibiot. 2014,
67, 631–644.
[Google Scholar]
-
14.
Strahl H, Errington J. Bacterial Membranes: Structure, Domains, and Function.
Ann. Rev. Microbiol. 2017,
71, 519–538.
[Google Scholar]
-
15.
-
16.
Zerfas BL, Joo Y, Gao J. Gramicidin A Mutants with Antibiotic Activity against Both Gram-Positive and Gram-Negative Bacteria.
ChemMedChem 2016,
11, 629–636.
[Google Scholar]
-
17.
Satlin MJ, Jenkins SG. 151 - Polymyxins. In Infectious Diseases, 4th ed.; Elsevier: Oxford, UK, 2017; pp. 1285–1288.
-
18.
Shaheen M, Li J, Ross AC, Vederas JC, Jensen SE. Paenibacillus polymyxa PKB1 Produces Variants of Polymyxin B-Type Antibiotics.
Chem. Biol. 2011,
18, 1640–1648.
[Google Scholar]
-
19.
Trimble MJ, Mlynárčik P, Kolář M, Hancock REW. Polymyxin: Alternative Mechanisms of Action and Resistance.
Cold Spring Harb. Perspect. Med. 2016,
6, a025288.
[Google Scholar]
-
20.
Evans ME, Feola DJ, Rapp RP. Polymyxin B Sulfate and Colistin: Old Antibiotics for Emerging Multiresistant Gram-Negative Bacteria.
Ann. Pharmacother. 1999,
33, 960–967.
[Google Scholar]
-
21.
Ledger EVK, Sabnis A, Edwards AM. Polymyxin and lipopeptide antibiotics: membrane-targeting drugs of last resort.
Microbiology 2022,
168, 001136.
[Google Scholar]
-
22.
Yu Z, Qin W, Lin J, Fang S, Qiu J. Antibacterial Mechanisms of Polymyxin and Bacterial Resistance.
Biomed. Res. Int. 2015,
2015, 679109.
[Google Scholar]
-
23.
Burkhart BM, Gassman RM, Langs DA, Pangborn WA, Duax WL, Pletnev V. Gramicidin D conformation, dynamics and membrane ion transport.
Biopolymers 1999,
51, 129–144.
[Google Scholar]
-
24.
Duttagupta I, Ghosh KC, Sinha S. Synthetic Studies Toward Nonribosomal Peptides. In Studies in Natural Products Chemistry; Elsevier: Oxford, UK, 2016; pp. 29–64.
-
25.
Prenner EJ, Lewis RNAH, Kondejewski LH, Hodges RS, McElhaney RN. Differential scanning calorimetric study of the effect of the antimicrobial peptide gramicidin S on the thermotropic phase behavior of phosphatidylcholine, phosphatidylethanolamine and phosphatidylglycerol lipid bilayer membranes.
Biochim. Biophys. Acta Biomembr. 1999,
1417, 211–223.
[Google Scholar]
-
26.
Prenner EJ, Lewis RNAH, Neuman KC, Gruner SM, Kondejewski LH, Hodges RS, et al. Nonlamellar Phases Induced by the Interaction of Gramicidin S with Lipid Bilayers. A Possible Relationship to Membrane-Disrupting Activity.
Biochemistry 1997,
36, 7906–7916.
[Google Scholar]
-
27.
Ashrafuzzaman Md, Andersen OS, McElhaney RN. The antimicrobial peptide gramicidin S permeabilizes phospholipid bilayer membranes without forming discrete ion channels.
Biochim. Biophys. Acta Biomembr. 2008,
1778, 2814–2822.
[Google Scholar]
-
28.
-
29.
Anderson VE, Osheroff N. Type II Topoisomerases as Targets for Quinolone Antibacterials Turning Dr. Jekyll into Mr. Hyde.
Curr. Pharm. Des. 2001,
7, 337–353.
[Google Scholar]
-
30.
Pham TDM, Ziora ZM, Blaskovich MAT. Quinolone antibiotics.
MedChemComm 2019,
10, 1719–1739.
[Google Scholar]
-
31.
-
32.
-
33.
-
34.
Bermingham A, Derrick JP. The folic acid biosynthesis pathway in bacteria: evaluation of potential for antibacterial drug discovery.
Bioessays 2002,
24, 637–648.
[Google Scholar]
-
35.
Sköld O. Sulfonamide resistance: mechanisms and trends.
Drug Resist. Updates 2000,
3, 155–160.
[Google Scholar]
-
36.
Gleckman R, Blagg N, Joubert DW. Trimethoprim: Mechanisms of Action, Antimicrobial Activity, Bacterial Resistance, Pharmacokinetics, Adverse Reactions, and Therapeutic Indications.
Pharmacother. J. Hum. Pharmacol. Drug Ther. 1981,
1, 14–19.
[Google Scholar]
-
37.
Abril AG, Rama JLR, Sánchez-Pérez A, Villa TG. Prokaryotic sigma factors and their transcriptional counterparts in Archaea and Eukarya.
Appl. Microbiol. Biotechnol. 2020,
104, 4289–4302.
[Google Scholar]
-
38.
Floss HG, Yu TW. Rifamycin
—Mode of Action, Resistance, and Biosynthesis.
Chem Rev. 2005,
105, 621–632.
[Google Scholar]
-
39.
-
40.
Chopra I, Roberts M. Tetracycline Antibiotics: Mode of Action, Applications, Molecular Biology, and Epidemiology of Bacterial Resistance.
Microbiol. Mol. Biol. Rev. 2001,
65, 232–260.
[Google Scholar]
-
41.
Krause KM, Serio AW, Kane TR, Connolly LE. Aminoglycosides: An Overview.
Cold Spring Harb. Perspect. Med. 2016,
6, a027029.
[Google Scholar]
-
42.
-
43.
Kanoh S, Rubin BK. Mechanisms of Action and Clinical Application of Macrolides as Immunomodulatory Medications.
Clin. Microbiol. Rev. 2010,
23, 590–615.
[Google Scholar]
-
44.
-
45.
Fernández-Martínez LT, Borsetto C, Gomez-Escribano JP, Bibb MJ, Al-Bassam MM, Chandra G, et al. New Insights into Chloramphenicol Biosynthesis in Streptomyces venezuelae ATCC 10712.
Antimicrob. Agents Chemother. 2014,
58, 7441–7450.
[Google Scholar]
-
46.
-
47.
-
48.
-
49.
Barbachyn MR. The Oxazolidinones. In Antibacterials; Springer International Publishing: Cham, Switzerland, 2017; Topics in Medicinal Chemistry; Volume 26; pp. 97–121.
-
50.
Bozdogan B, Appelbaum PC. Oxazolidinones: activity, mode of action, and mechanism of resistance.
Int. J. Antimicrob. Agents 2004,
23, 113–119.
[Google Scholar]
-
51.
Dahshan H. Pharma Guide, Basic and Clinical Pharmacology; University Book Centre: Cairo, Egypt, 2015.
-
52.
Nankervis H, Thomas KS, Delamere FM, Barbarot S, Rogers NK, Williams HC. Antimicrobials Including Antibiotics, Antiseptics and Antifungal Agents. Scoping Systematic Review of Treatments for Eczema. NIHR Journals Library, 2016. Available online:
https://www.ncbi.nlm.nih.gov/books/NBK363143/ (accessed on 28 October 2022).
-
53.
Kolář M, Urbánek K, Látal T. Antibiotic selective pressure and development of bacterial resistance.
Int. J. Antimicrob. Agents 2001,
17, 357–363.
[Google Scholar]
-
54.
Lasemi E, Navi F, Lasemi R, Lasemi N, Lasemi E, Navi F, et al. Complications of Antibiotic Therapy and Introduction of Nanoantibiotics. In A Textbook of Advanced Oral and Maxillofacial Surgery Volume 3; IntechOpen: Rijeka, Croatia, 2016.
-
55.
Munita JM, Arias CA. Mechanisms of Antibiotic Resistance. Microbiol. Spectr. 2016, 4. doi:10.1128/microbiolspec.VMBF-0016‑2015.
-
56.
Pan X, He Y, Chen T, Chan KF, Zhao Y. Modified Penicillin Molecule with Carbapenem-Like Stereochemistry Specifically Inhibits Class C β-Lactamases.
Antimicrob. Agents Chemother. 2017,
61, e01288-17.
[Google Scholar]
-
57.
D’Costa V, Wright GD. Biochemical Logic of Antibiotic Inactivation and Modification. In Antimicrobial Drug Resistance: Mechanisms of Drug Resistance; Humana Press: Totowa, NJ, USA, 2009; pp. 81–95.
-
58.
Elufisan T, Oyedara O, Oluyide O. Updates on microbial resistance to drugs.
Afr. J. Microbiol. Res. 2012,
6, 4833–4844.
[Google Scholar]
-
59.
Salahuddin P, Kumar A, Khan AU. Structure, Function of Serine and Metallo-β-lactamases and their Inhibitors.
Curr. Protein Pept. Sci. 2018,
19, 130–144.
[Google Scholar]
-
60.
Bush K, Bradford PA. Interplay between β-lactamases and new β-lactamase inhibitors.
Nat. Rev. Microbiol. 2019,
17, 295–306.
[Google Scholar]
-
61.
Schulz GE. Bacterial porins: structure and function.
Curr. Opin. Cell Biol. 1993,
5, 701–707.
[Google Scholar]
-
62.
Fernández L, Hancock REW. Adaptive and Mutational Resistance: Role of Porins and Efflux Pumps in Drug Resistance.
Clin. Microbiol. Rev. 2012,
25, 661–681.
[Google Scholar]
-
63.
Pagès JM, James CE, Winterhalter M. The porin and the permeating antibiotic: a selective diffusion barrier in Gram-negative bacteria.
Nat. Rev. Microbiol. 2008,
6, 893–903.
[Google Scholar]
-
64.
Amaral L, Martins A, Spengler G, Molnar J. Efflux pumps of Gram-negative bacteria: what they do, how they do it, with what and how to deal with them.
Front. Pharmacol. 2014,
4, 168.
[Google Scholar]
-
65.
Kapoor G, Saigal S, Elongavan A. Action and resistance mechanisms of antibiotics: A guide for clinicians.
J. Anaesthesiol. Clin. Pharmacol. 2017,
33, 300–305.
[Google Scholar]
-
66.
Andersson DI, Hughes D. Selection and Transmission of Antibiotic-Resistant Bacteria. Microbiol. Spectr. 2017, 5. doi:10.1128/microbiolspec.mtbp-0013-2016.
-
67.
Mackenzie JS, Jeggo M. The One Health Approach—Why Is It So Important?
Tropic. Med. Infect. Dis. 2019,
4, 88.
[Google Scholar]
-
68.
Schulte AM, Kolarski D, Sundaram V, Srivastava A, Tama F, Feringa BL, et al. Light-Control over Casein Kinase 1δ Activity with Photopharmacology: A Clear Case for Arylazopyrazole-Based Inhibitors.
Int. J. Mol. Sci. 2022,
23, 5326.
[Google Scholar]
-
69.
Fregoni J, Granucci G, Coccia E, Persico M, Corni S. Manipulating azobenzene photoisomerization through strong light–molecule coupling.
Nat. Commun. 2018,
9, 4688.
[Google Scholar]
-
70.
Beharry AA, Woolley GA. Azobenzene photoswitches for biomolecules.
Chem. Soc. Rev. 2011,
40, 4422–4437.
[Google Scholar]
-
71.
Volarić J, Szymanski W, Simeth NA, Feringa BL. Molecular photoswitches in aqueous environments.
Chem. Soc. Rev. 2021,
50, 12377–12449.
[Google Scholar]
-
72.
Specht A, Bolze F, Omran Z, Nicoud J, Goeldner M. Photochemical tools to study dynamic biological processes.
HFSP J. 2009,
3, 255–264.
[Google Scholar]
-
73.
Brieke C, Heckel A. Spiropyran Photoswitches in the Context of DNA: Synthesis and Photochromic Properties.
Chem. Eur J. 2013,
19, 15726–15734.
[Google Scholar]
-
74.
Kirchner S, Leistner AL, Pianowski ZL. Photoswitchable Peptides and Proteins. In Molecular Photoswitches; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2022; pp. 987–1013.
-
75.
Jäschke A. Genetically encoded RNA photoswitches as tools for the control of gene expression.
FEBS Lett. 2012,
586, 2106–2111.
[Google Scholar]
-
76.
Ihalainen JA, Bredenbeck J, Pfister R, Helbing J, Chi L, van Stokkum IHM, et al. Folding and unfolding of a photoswitchable peptide from picoseconds to microseconds.
Proc. Natl. Acad. Sci. USA 2007,
104, 5383–5388.
[Google Scholar]
-
77.
Chai J, Zhao Y, Xu L, Li Q, Hu XY, Guo DS, et al. A Noncovalent Photoswitch for Photochemical Regulation of Enzymatic Activity.
Angew. Chem. Int. Ed. 2022,
61, e202116073.
[Google Scholar]
-
78.
Shimoboji T, Ding ZL, Stayton PS, Hoffman AS. Photoswitching of Ligand Association with a Photoresponsive Polymer−Protein Conjugate.
Bioconjug. Chem. 2002,
13, 915–919.
[Google Scholar]
-
79.
Adrian M, Nijenhuis W, Hoogstraaten RI, Willems J, Kapitein LC. A Phytochrome-Derived Photoswitch for Intracellular Transport.
ACS Synth. Biol. 2017,
6, 1248–1256.
[Google Scholar]
-
80.
Hüll K, Morstein J, Trauner D. In Vivo Photopharmacology.
Chem. Rev. 2018,
118, 10710–10747.
[Google Scholar]
-
81.
Szymański W, Beierle JM, Kistemaker HAV, Velema WA, Feringa BL. Reversible Photocontrol of Biological Systems by the Incorporation of Molecular Photoswitches.
Chem. Rev. 2013,
113, 6114–6178.
[Google Scholar]
-
82.
Bacchus W, Fussenegger M. The use of light for engineered control and reprogramming of cellular functions.
Curr. Opin. Biotechnol. 2012,
23, 695–702.
[Google Scholar]
-
83.
Majima T, Tojo S, Ishida A, Takamuku S. Cis-Trans Isomerization and Oxidation of Radical Cations of Stilbene Derivatives.
J. Org. Chem. 1996,
61, 7793–800.
[Google Scholar]
-
84.
Chen PC, Chieh YC. Azobenzene and stilbene: a computational study.
J. Mol. Struct. THEOCHEM 2003,
624, 191–200.
[Google Scholar]
-
85.
Drake HF, Day GS, Xiao Z, Zhou HC, Ryder MR. Light-induced switchable adsorption in azobenzene- and stilbene-based porous materials.
TRECHEM 2022,
4, 32–47.
[Google Scholar]
-
86.
Ali AA, Kharbash R, Kim Y. Chemo- and biosensing applications of spiropyran and its derivatives—A review.
Anal. Chim. Acta 2020,
1110, 199–223.
[Google Scholar]
-
87.
Lukyanov BS, Lukyanova MB. Spiropyrans: Synthesis, Properties, and Application. (Review).
Chem. Heterocycl. Compd. 2005,
41, 281–311.
[Google Scholar]
-
88.
Aldoshin SM. Spiropyrans: Structural Features and Photochemical Properties.
Mol. Cryst. Liquid Cryst. Sci. Technol. Sect. A Mol. Cryst. Liquid Cryst. 1994,
246, 207–214.
[Google Scholar]
-
89.
Kortekaas L, Browne WR. The evolution of spiropyran: fundamentals and progress of an extraordinarily versatile photochrome.
Chem. Soc. Rev. 2019,
48, 3406–3424.
[Google Scholar]
-
90.
Fagan A, Bartkowski M, Giordani S. Spiropyran-Based Drug Delivery Systems.
Front. Chem. 2021,
9, 720087.
[Google Scholar]
-
91.
Koeppe B, Römpp F. Reversible Spatial Control in Aqueous Media by Visible Light: A Thioindigo Photoswitch that is Soluble and Operates Efficiently in Water.
Chem. Eur. J. 2018,
24, 14382–14386.
[Google Scholar]
-
92.
Li D, Yang Y, Li C, Liu Y. Unveiling the mechanism of the promising two-dimensional photoswitch—Hemithioindigo. Spectrochim.
Acta Part A Mol. Biomol. Spectrosc. 2018,
200, 1–9.
[Google Scholar]
-
93.
Vlajić M, Unger W, Bruns J, Rueck-Braun K. Photoswitching of fulgimides in different environments on silicon surfaces.
Appl. Surface Sci. 2019,
465, 686–692.
[Google Scholar]
-
94.
Renth F, Temps F. Fulgides and Fulgimides. In Molecular Photoswitches; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2022; pp. 177–192.
-
95.
Lachmann D, Lahmy R, König B. Fulgimides as Light‐Activated Tools in Biological Investigations.
Eur. J. Org. Chem. 2019,
2019, 5018–5024.
[Google Scholar]
-
96.
Matsuda K, Irie M. Diarylethene as a photoswitching unit.
J. Photochem. Photobiol. C Photochem. Rev. 2004,
5, 169–182.
[Google Scholar]
-
97.
Liu R, Yang Y, Cui Q, Xu W, Peng R, Li L. A Diarylethene-Based Photoswitch and its Photomodulation of the Fluorescence of Conjugated Polymers.
Chem. Eur. J. 2018,
24, 17756–17766.
[Google Scholar]
-
98.
Nakatani K, Sato H, Fukuda R. A catalyzed E/Z isomerization mechanism of stilbene using para-benzoquinone as a triplet sensitizer.
Phys. Chem. Chem. Phys. 2022,
24, 1712–1721.
[Google Scholar]
-
99.
Wang C, Waters MDJ, Zhang P, Suchan J, Svoboda V, Luu TT, et al. Different timescales during ultrafast stilbene isomerization in the gas and liquid phases revealed using time-resolved photoelectron spectroscopy.
Nat Chem. 2022,
14, 1126–1132.
[Google Scholar]
-
100.
Waldeck DH. Photoisomerization dynamics of stilbenes in polar solvents.
J. Mol. Liquids. 1993,
57, 127–148.
[Google Scholar]
-
101.
Crecca CR, Roitberg AE. Theoretical Study of the Isomerization Mechanism of Azobenzene and Disubstituted Azobenzene Derivatives.
J. Phys. Chem. A 2006,
110, 8188–8203.
[Google Scholar]
-
102.
Farka D, Scharber M, Głowacki ED, Sariciftci NS. Reversible Photochemical Isomerization of N,N′-Di(t -butoxycarbonyl)indigos.
J. Phys. Chem. A 2015,
119, 3563–3568.
[Google Scholar]
-
103.
Lemieux RP. Photoswitching of ferroelectric liquid crystals using chiral thioindigo dopants: The development of a photochemical switch hitter.
Chem. Record. 2004,
3, 288–295.
[Google Scholar]
-
104.
Petermayer C, Dube H. Indigoid Photoswitches: Visible Light Responsive Molecular Tools.
Acc. Chem. Res. 2018,
51, 1153–1163.
[Google Scholar]
-
105.
Wiedbrauk S, Dube H. Hemithioindigo—an emerging photoswitch.
Tetrahedr. Lett. 2015,
56, 4266–4274.
[Google Scholar]
-
106.
Berdnikova DV. Visible-range hemi-indigo photoswitch: ON–OFF fluorescent binder for HIV-1 RNA.
Chem. Commun. 2019,
55, 8402–8405.
[Google Scholar]
-
107.
Bossi ML, Murgida DH, Aramendía PF. Photoisomerization of Azobenzenes and Spirocompounds in Nematic and in Twisted Nematic Liquid Crystals.
J. Phys. Chem. B 2006,
110, 13804–13811.
[Google Scholar]
-
108.
Crespi S, Simeth NA, König B. Heteroaryl azo dyes as molecular photoswitches.
Nat. Rev. Chem. 2019,
3, 133–146.
[Google Scholar]
-
109.
Göstl R, Senf A, Hecht S. Remote-controlling chemical reactions by light: Towards chemistry with high spatio-temporal resolution.
Chem. Soc. Rev. 2014,
43, 1982.
[Google Scholar]
-
110.
Stranius K, Börjesson K. Determining the Photoisomerization Quantum Yield of Photoswitchable Molecules in Solution and in the Solid State.
Sci. Rep. 2017,
7, 41145.
[Google Scholar]
-
111.
Imato K, Momota K, Kaneda N, Imae I, Ooyama Y. Photoswitchable Adhesives of Spiropyran Polymers.
Chem. Mater. 2022,
34, 8289–8296.
[Google Scholar]
-
112.
Velema WA, Szymanski W, Feringa BL. Photopharmacology: Beyond Proof of Principle.
J. Am. Chem. Soc. 2014,
136, 2178–2191.
[Google Scholar]
-
113.
Boelke J, Hecht S. Designing Molecular Photoswitches for Soft Materials Applications.
Adv. Opt. Mater. 2019,
7, 1900404.
[Google Scholar]
-
114.
Bonardi F, London G, Nouwen N, Feringa BL, Driessen AJM. Light-Induced Control of Protein Translocation by the SecYEG Complex.
Angew. Chem. Int. Ed. 2010,
49, 7234–7238.
[Google Scholar]
-
115.
Mayer G, Heckel A. Biologically Active Molecules with a “Light Switch”.
Angew. Chem. Int. Ed. 2006,
45, 4900–4921.
[Google Scholar]
-
116.
Koçer A, Walko M, Meijberg W, Feringa BL. A Light-Actuated Nanovalve Derived from a Channel Protein.
Science 2005,
309, 755–758.
[Google Scholar]
-
117.
Wegener M, Hansen MJ, Driessen AJM, Szymanski W, Feringa BL. Photocontrol of Antibacterial Activity: Shifting from UV to Red Light Activation.
J. Am. Chem. Soc. 2017,
139, 17979–17986.
[Google Scholar]
-
118.
Lauxen AI, Kobauri P, Wegener M, Hansen MJ, Galenkamp NS, Maglia G, et al. Mechanism of Resistance Development in
E. coli against TCAT, a Trimethoprim-Based Photoswitchable Antibiotic.
Pharmaceuticals 2021,
14, 392.
[Google Scholar]
-
119.
Velema WA, van der Berg JP, Hansen MJ, Szymanski W, Driessen AJM, Feringa BL. Optical control of antibacterial activity.
Nat. Chem. 2013,
5, 924–928.
[Google Scholar]
-
120.
Velema WA, Hansen MJ, Lerch MM, Driessen AJM, Szymanski W, Feringa BL. Ciprofloxacin–Photoswitch Conjugates: A Facile Strategy for Photopharmacology.
Bioconjug. Chem. 2015,
26, 2592–2597.
[Google Scholar]
-
121.
Fu X, Bai H, Qi R, Zhao H, Peng K, Lv F, et al. Optically-controlled supramolecular self-assembly of an antibiotic for antibacterial regulation.
Chem. Commun. 2019,
55, 14466–14469.
[Google Scholar]
-
122.
Fu X, Yu J, Dai N, Huang Y, Lv F, Liu L, et al. Optical Tuning of Antibacterial Activity of Photoresponsive Antibiotics.
ACS Appl. Bio Mater. 2020,
3, 4751–4755.
[Google Scholar]
-
123.
Zhang H, Qi Y, Zhao X, Li M, Wang R, Cheng H, et al. Dithienylethene-Bridged Fluoroquinolone Derivatives for Imaging-Guided Reversible Control of Antibacterial Activity.
J. Org. Chem. 2022,
87, 7446–7455.
[Google Scholar]
-
124.
Li Z, Wang Y, Li M, Zhang H, Guo H, Ya H, et al. Synthesis and properties of dithienylethene-functionalized switchable antibacterial agents.
Org. Biomol. Chem. 2018,
16, 6988–6997.
[Google Scholar]
-
125.
Babii O, Afonin S, Berditsch M, Reiβer S, Mykhailiuk PK, Kubyshkin VS, et al. Controlling Biological Activity with Light: Diarylethene-Containing Cyclic Peptidomimetics.
Angew. Chem. Int. Ed. 2014,
53, 3392–3395.
[Google Scholar]