1. Introduction
In boreal ecosystems, various mammals and birds rely on aquatic habitats for food. Some of these species remain active at northern latitudes through the winter months when most water bodies are ice-covered: they do not migrate or hibernate. Four semi-aquatic mammals—river otter (
Lontra canadensis), mink (
Neogale vison), beaver (
Castor canadensis) and muskrat (
Ondatra zibethicus)—are prominent examples. Relatively few boreal bird species that feed in aquatic ecosystems avoid migration. However, the American dipper (
Cinclus mexicanus) and some waterfowl (Anatidae) stay in unfrozen aquatic habitats at high latitudes through winter [
1,
2]. In addition, waterfowl migrating in spring (April-May) congregate at these areas of open water at the outflows of a wide range of sizes of large boreal lakes that are still mostly ice-covered [
1,
3].
The proportion of a boreal water body that remains unfrozen all winter and spring is clearly quite small because consistent cold results in extensive ice cover over the majority of most water bodies north of about 55° N Latitude [
4,
5]. Consequently, areas of open water are limited in availability and likely of high value as foraging habitats to the species that use them. These species are part of the aquatic food web, and here we are specifically concerned with semi-aquatic mammals and birds that access aquatic foods. However, reviews of winter limnology of northern lakes [
4,
5] provide very limited discussion of the incidence, dynamics, and ecology of long-lasting patches of open water in winter.
Portions of some boreal lakes stay open (unfrozen) for various reasons. Groundwater, at and below the water table, can percolate into lakes at the lake surface and in deeper water, sometimes maintaining open water along shorelines [
6]. Springs, where underground aquifers emerge as streams on the surface, can maintain open water downstream, often into lakes, until exposure to cold air brings water temperature low enough to freeze [
6]. However, most noticeable are open water patches at some lake inflows, narrows between lake basins, and especially at lake outflows.
Persistent and recurring patches of open water are frequently created and maintained by currents moving water into, through and out of the lakes. The current flows fairly directly under and close to the ice to the outflow, driven by drainage from upstream in the catchment [
6,
7,
8]. However, the incidence and persistence of patches of open water are also influenced by the considerable heat stored in benthic sediments and in the deeper lake water. Water temperature in winter generally increases with depth in northern latitude lakes [
4,
6]. This pattern emerges after autumn turnover when surface water that has cooled to 4 °C then sinks to the lake bottom because it is densest at that temperature. To a smaller extent, the pattern is maintained by heat stored during summer in lake sediments [
4,
6]. This thermal profile with depth is remarkably stable after freeze-up because covers of ice and snow provide excellent insulation from ambient temperatures [
4,
6]. Although the entire water column gradually cools with ongoing ambient air below freezing, the overall result is that substantial heat, stored in water above freezing point, is available in the body of the lake to warm the near freezing surface waters under the ice that is close to the outlet [
6,
7]. This warming happens partly through convection but also because deeper water can upwell in response to the downstream current at the outflow pulling water out of the lake [
4,
6]. Data from a large Yukon lake show that this upwelling can occur from depths as much as 15 m and distances of many hundreds of meters from the outlet, and results in water temperature at the outflow being at least 1 °C [
7].
We call these patches of open water “polynyas”, a term coined for persistent open water in ice-covered oceans, but also used in a freshwater context [
7]. It is suitable for freshwater because persistent near-surface currents plus upwelling of warmer water drive these recurring open patches in freshwater and seawater [
4,
6,
9,
10,
11], and because the word derivation does not depend on a marine association [
11]. However, it is noteworthy that recurring, strong wind plays a large part in the formation of many marine polynyas but does not play such a strong role in most freshwater settings [
6,
11]. Also, marine polynyas are much larger in surface area (c. 800 to 9000 km
2 in April [
10]) than even the freshwater ones on large boreal lakes in the northern boreal mountains (0.7 km
2 in January [
7]; 2 to 8 km
2 in April [
1]).
Our initial winter and spring observations of outflow polynyas on various lakes in Yukon Territory indicated that they are: very localized and seasonal ecosystems that recur annually; used frequently by semi-aquatic mammals and some birds; poorly studied with regard to their occurrence and the diversity of species using them throughout winter.
We investigated two questions. First, what are the frequencies and hydrological correlates of various types of open water, in particular outflow polynyas? We proposed three hypotheses: (i) that the mean size of lakes with outflow polynyas is significantly larger than the mean size of lakes without outflow polynyas, (ii) that outflow polynyas are significantly associated with higher stream orders, and (iii) that there is a consistent threshold in upstream catchment size at which outflow polynyas occur in the drainage.
Second, we asked what mammals and bird species visit such relatively small sites of open water from late autumn through to spring, well before the lakes lose all their ice, and what they do at these sites. We hypothesized that: (i) semi-aquatic mammals visit from late autumn through spring mainly for foraging because of the easy access to water, (ii) the few species of water birds known to overwinter in some areas of Yukon would also forage throughout the winter because they require ready access to water, and (iii) migratory waterfowl would visit in spring to forage because their long migrations to subarctic latitudes plus the selective advantage of nesting as early as possible demand nutrient intake at northern sites to maximize reproductive output [
12,
13].
2. Materials and Methods
2.1. Study Area
We undertook this research in the Southern Lakes Ecoregion of southern Yukon Territory, Canada; this ecoregion is part of Canada’s Boreal Cordillera ecozone [
14]. In this portion of the northern boreal mountains, Pleistocene glaciers covered most of the region but had mostly retreated by the Holocene. They left relatively wide valleys and rounded plateaux and ranges compared to the more deeply incised valleys and higher, still glaciated, peaks of the Coast Range closer to the Pacific Ocean. Bedrock is a diverse mix of metamorphic and igneous types, interspersed with sedimentary limestone and sandstone, and with volcanic intrusions. Surficial geology is dominated by glaciofluvial and glaciolacustrine deposits from the last Pleistocene glaciation, modified by erosional and depositional features of surface streams and rivers, as well as mass wasting of relatively uncommon bedrock exposures [
14]. In this ecoregion, ponds and lakes are moderately common compared to other parts of the Boreal Cordillera ecozone, and include some of the largest lakes in the western cordillera of North America, such as Teslin Lake (36,240 ha) and Lake Laberge (20,100 ha) [
15].
Lying in the rain shadow of the Coast Range but at a high latitude (c. 60.5° N Lat), this ecoregion has a subarctic continental climate. The mean annual precipitation is only 200 to 325 mm, and the mean annual temperature ranges from −1 °C to −4°C, warmed in winters and cooled in summers, compared to continental regimes to the north and east, by air masses from the Pacific Ocean [
16]. Nevertheless, winters are quite long (ice cover on small lakes from mid-October to mid-May) and cold (mean January daily temperatures from −21 °C to −25 °C) [
16]. Over the past 50 years, temperatures have warmed significantly, by over 2 °C, with the biggest increase in winter, and total annual precipitation has increased by 3%, especially in summer, but with substantial inter-annual variability [
17].
Our focal study area (c. 1945 km
2, elevation range 687 to 1546 m, centered at approximately 65°15′ N, −133°35′ W, ) includes the upper Squanga and the Teenah/Seaforth River drainages flowing into the Teslin River, along with the Little Atlin, Haunka, Snafu and Tarfu Creek drainages flowing into Atlin Lake. The latter set is within Agay Mene Territorial Park. All of these drainages are headwater tributaries to the Yukon River, including streams of orders one to four [
18], and lentic water bodies of a few to 1140 ha. We did not include the largest regional lakes (>5000 ha) because their outflow polynyas were already well documented [
1,
3]. We purposefully sampled complete drainages (i) holding lakes both with and without polynyas and (ii) with lakes of the small to medium sizes most useful to semi-aquatic mammals [
19,
20]. Lakes were often in sequence along drainages, and ranged from 687 to 1020 m in elevation.
. Map of focal study area showing (i) the locations of all systematically mapped lakes in the four drainages (Little Atlin, Squanga, Teenah and Snafu), and (ii) the locations of camera trapping sites (A, B and C as described in the text). Lakes with outflow polynyas are farther downstream, with higher stream order, in each drainage. The inset map shows the location of the focal study area (dark square) within Yukon Territory in northwestern North America.
Over most of the focal study area, including most valley floors with lakes that we sampled, the probability of permafrost is projected to be 0.1 (isolated patches) to 0.4 (sporadic), but occasionally reaching as high as 0.9 (continuous) on the crests of the highest mountains [
21].
The focal study area is mostly forested, with canopies of white spruce (
Picea glauca), lodgepole pine (
Pinus contorta), subalpine fir (
Abies lasiocarpa) and trembling aspen (
Populus tremuloides), and willows (
Salix spp.), especially near water [
22]. Open bedrock and alpine tundra cover some mountain ridges. Most lakes we sampled are in the Boreal Low Bioclimate Zone, below 950 m in elevation [
22].
2.2. Frequency and Correlates of Open Water in Winter
To determine the frequency of occurrence of various types of open water (“polynya”), we mapped open water (patches > 50 m
2) on all lakes (
>2 ha) along the main stems and tributaries of 4 headwater drainages (Little Atlin, Squanga, Teenah/Seaforth, Snafu) (a) systematically by helicopter on 10 December 2014, and (b) by repeated ground visits to these drainages in the course of winter field work on river otters from 2012 to 2020 (
). With the exception of many small ponds (<5 ha) that lacked above ground drainage, we visited all lakes at least twice during the study, and as many as five times per winter. To understand the persistence of open water during winter and among years, we visited 14 lakes with open water at least 3 times from December to April across at least 3 years.
We recognized three types of winter open water on lakes: inflows (mouths of tributary streams providing current into the lakes, or groundwater seeps along lakeshores); narrows (constrictions < 50 m wide in the width of the water body, concentrating the current through the lake); outflows (downstream gradient inducing current out of the lake plus upwelling near the outflow).
For outflow polynyas, the most common and persistent type of open water, we assessed lake size, stream order, and upstream catchment size as potential hydrological correlates. These analyses were based on ground visits in December to all lakes with above-ground drainage and larger than two hectares in the upstream portions of the four drainages we surveyed from the air. Our measure of lake size was surface area (quantified in ArcGIS Pro 3.4.0); bathymetry was lacking for these lakes. The stream order was based on the Horton-Strahler ordination system [
18], with the first order being placed at the headwaters. To quantify catchment size, we used the total length of all stream courses plus total length of all lakeshores (±500 m), as displayed in the 1:50,000 Canadian National Topographic Series (CanVec digital data) maps. We chose this distance-based measurement of all hydrological features upstream of a lake outflow as a proxy for an area-based measurement of catchment size because the available digital elevation model was relatively coarse scale (30 m resolution) and known to be error prone.
2.3. Mammal and Bird Species Diversity and Behaviors
To quantify the relative abundances of mammal and bird species, the timing of their visits, and their behaviors at patches of open water, we deployed automated cameras (camera traps) close to the shoreline of three open water sites during the approximately 6-month long period of ice cover on these smaller lakes (). In 2014–15 and 2016–17, we used Bushnell Trophy Cam (Model 119537CN) cameras, and we augmented these with Reconyx HC600 Hyperfire cameras in 2012–2013. Cameras were powered with lithium-ion batteries because of the better performance of this battery type in cold temperatures. To increase the sample size of open water sites potentially supporting the two species most strongly associated with open water in mid-winter (river otter, American dipper), we recorded evidence (sightings and snow tracks) of these species at our ground visits to the complete sample of lakes with outflow open water.
Of the three sites (, ), two were in the Tarfu drainage: Site A being the inflow polynya to un-gazetted Otter Lake (60.0710° N, 133.7625° W; 32.4 ha); site B being the outflow polynya from Otter Lake and continuous open water downstream for 250 m. One was in the Snafu drainage: Site C being the outflow polynya to Lower Snafu Lake (60.1532° N, 133.8022° W; 301.0 ha). We accessed sites on snowshoe or ski, travelling 3 to 8 km from road. Other people occasionally travelled to the same lakes, mainly by snowmobile, but we did not capture images of people on our cameras.
.
Summary of sampling effort using camera traps, including winters, time periods, and numbers of photos recorded at each site under continuous and periodic sampling. Site codes correspond to sites described in the text.
Sampling
Regime |
Site
Code |
Winters |
Time Periods
(Day/Month) |
Number of
Cameras |
Number of
Photos |
Photo
Mode |
| Continuous |
A |
2016–17 |
13/10–30/04 |
1 |
8088 |
Still |
|
B |
2016–17 |
03/11–30/04 |
5 |
24,799 |
Still |
|
C |
2014–15 |
11/10–30/04 |
1 |
576 |
Video |
| Periodic |
A |
2013 |
08/01–21/04 |
2 |
554 |
Still |
|
A |
2014–15 |
13/10–14/01 |
3 |
3289 |
Video |
|
|
|
& 30/03–30/04 |
|
|
|
|
B |
2014–15 |
06/10–14/01 |
3 |
2540 |
Video |
|
|
|
& 20/02–28/02 |
|
|
|
|
|
|
& 30/03–30/04 |
|
|
|
|
C |
2014–15 |
10/10–13/02 |
3 |
1608 |
Video |
|
|
|
& 20/04–30/04 |
|
|
|
Our primary approach to sampling was “continuous” deployment of the same set of cameras at the same sites from near freeze-up in mid or late October (some variation between sites) through the winter until the end of April. We chose this end-point, a little prior to break-up on these lakes (mid-May), because polynyas had increased in size, so the camera traps could no longer capture the full extent of open water. Migratory waterfowl (swans, geese, and ducks (Anatidae)) visitation to the outflow polynyas on the very large lakes in this region tapers off by the end of April as the birds either continue their migrations or disperse to smaller water bodies locally for the breeding season [
1,
3,
23]. Our sampling may have missed some species diversity and abundance in May. We augmented this “continuous” sampling with “periodic” camera trapping during portions of some winters; this sampling had temporal breaks in the record (). We present the results of the “periodic” camera trapping in the Supplementary Material.
Cameras were set to be triggered by motion (infrared) and record an image every hour on the hour. We wanted to increase the likelihood of capturing images of species that spend considerable time underwater and those that might be quite far from the camera. Because animals generally need to be within about 30 m of the camera to trigger it, we aimed and placed cameras quite close to particular features (e.g., spillways on beaver dams, upstream ice edge at lake outlet, latrine sites used by river otters) that animals were most likely to occupy if they were present at the open water. Each camera also had a field of view beyond the particular feature of interest, often capturing the presence of individuals other than the ones that triggered the camera. We oriented these fields of view to cover the entire open water area, so up to 5 cameras were used at one site with fairly extensive open water. Most photos were stills (2 photos per trigger), but in 2014–15, we set cameras to record 20-s videos.
We measured relative abundance as the number of individuals recorded on photos per day for each species (count per day). One author (ML) analysed and classified all photos to avoid interpersonal bias. Photos were not always independent, however, because one or more individuals of a species could repeatedly trigger the camera within one day. To remove much of this potential repeat counting within a day, we considered photos that repeatedly recorded individuals of the same species, with less than a two-hour interval between consecutive photos, to be showing the same individuals. This applied to photos from all cameras recording at the same site, unless photos from different cameras captured the species in different portions of the site at exactly the same time. Counts per day were then the sum of the maximum number of “individuals recorded” during all of the time periods that were separated by at least two hours within the same day. We had to apply this procedure mainly for waterfowl in spring, because they are nearly continuously visible on the water surface. Even when applying this rule, our counts per day may be inflated, especially for waterfowl, because the same individual birds may appear within more than one of the time periods separated by two hours. However, we consider the two-hour rule as a reasonable compromise to deal with our field observations of considerable daily turnover of waterfowl within sites in spring, and our goal to quantify general patterns of seasonal use of these sites rather than absolute counts.
For each still photo, we classified the observed behaviour using the following categories: Forage—search or eat; Excrete; Travel—fly; Travel—swim; Travel—walk; Rest and Sleep; Play; Groom—allo or auto; Vigilance; Smell and scent mark; Display—mating; Interspecific interaction; Investigate camera; Aversion to camera. Some behaviors, such as otters searching for food, would be very difficult to capture in above-water photos, so the photos may not give a complete view of how a species used the site. Some behavior categories were not entirely independent. For example, animals can travel while foraging, so many instances of “Travel-swim” may also have been “Forage-search”, especially for waterfowl. In videos, the primary behavior we classified was the one lasting the longest in the 20-second sequence.
3. Results
3.1. Frequency and Correlates of Open Water in Winter
During the helicopter flight along main-stem drainages, 26 of 42 surveyed lakes had at least one area of open water. However, such open water was very localized, covering only 0.11% of the total surface area of all lakes surveyed. Regarding lakes with some open water, outflow polynyas were the most common and occurred on all 26 (100%), followed by open water at inflows on 14 (54%) and at narrows on 5 (19%). One inflow polynya was definitely fed from a karst resurgent stream, whereas the great majority were caused by the current of inflowing tributaries from upstream lakes or from first-order streams whose sources of water were more likely in winter to be diffuse inputs of groundwater. A large proportion (12 or 86%) of these inflow polynyas were fed by open water along the entire length of the streams from the outflow of the nearest upstream lake; these inflow polynyas were directly related to upstream outflows. We could not systematically map the incidence of open water originating from groundwater along lakeshores: from the air, they were too indistinct, and on the ground, many froze over and became covered in snow as winter progressed. Early to mid-December was a suitable time of year to map polynyas, with at least six weeks having passed since freezing of virtually the entire surface area of this size range of lakes.
Of the 26 outflows, open water occurred both on the level surface of the lake itself and also in 100% of the streams draining the lakes downstream. The length of open water downstream ranged from 50 m to 3500 m (mean = 683 m (SD = 775 m), median = 425 m), flowing all the way to become an inflow to the next lake downstream in 12 (46%) instances. Fifteen (58%) of these 26 outflow polynyas included an existing intact or partly washed-out beaver dam, with open water both above and below the dams. Overall, open water flowing out of the lake in a stream is a prominent component of each outflow polynya (a). However, its extent tended to shorten or become intermittent in the colder periods of January through March.
Open water was remarkably persistent throughout individual winters and recurred at the same places across many winters on the 14 lakes we regularly visited. All 14 outflow polynyas persisted all winter, all 6 inflow polynyas persisted all winter, and one of two polynyas at the narrows persisted all winter. All of these open water sites recurred at the same sites in the minimum of three winters with sampling, and many for at least 8 consecutive winters. Polynya size often contracted with some ice cover during periods of deeper cold (<−25 °C), leading to complete ice cover of some polynyas in narrows. However, this ice cover generally disappeared in warmer weather. With daytime temperatures regularly above freezing in April and May, all areas of open water recurred and gradually expanded in size.
. (<b>a</b>) Aerial view of the dark open water of an outflow polynya (lake on left) and open water in stream feeding an inflow polynya (lake on right) in March (Credit: Gerry Whitley); (<b>b</b>) River otters at open water in stream from an outflow polynya, camera trapping site B in March (Credit: Maria Leung).
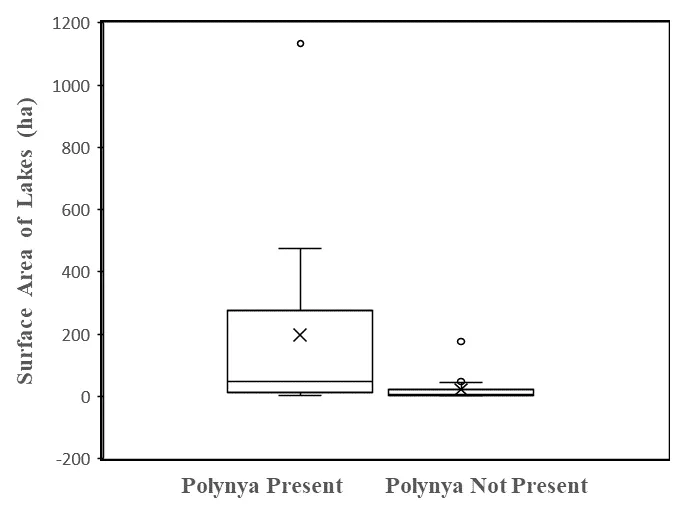
Regarding the systematic survey of all lakes in the upstream portions of four drainages, 22 (37%) of 59 lakes had an outflow polynya. Importantly, most of these lakes were arranged in sequences within each drainage and are not fully independent sample units. However, we treat them as such in the following statistical analyses. We found support for our first hypothesis: the mean size of lakes with outflow polynyas (197.8 ha, range 3.5 to 1138.4 ha) was indeed significantly larger than the mean size of lakes without such open water (22.7 ha, range 2.0 to 175.6 ha) (t = 3.88, df = 57,
p < 0.001(one-tailed)) (). However, some lakes with outflow polynyas were relatively small, and were found quite far downstream in the drainage sequence, suggesting that size alone was not driving the occurrence of open water ().
. Surface areas (ha) of lakes with an outflow polynya are larger than those without an outflow polynya. The boxes in these plots are bounded by first and third quartiles, with mean values (X) and median values (horizontal lines). Whiskers display maximum and minimum values without outliers. Outliers are open circles with values greater or less than 1.5 times the span between first and third quartiles.
We found support for our second hypothesis that outflow polynyas were significantly associated with higher stream orders of the outflow streams, given an expectation that polynyas would be distributed among stream orders in similar proportions to the relative incidence of those orders in our full sample of 59 lakes (Lakes with outflow polynyas: Chi-square goodness of fit = 22.2,
df = 3,
p < 0.001; Lakes without outflow polynyas: Chi-square goodness of fit = 13.2,
df = 3,
p < 0.001). Thirty-five of the 37 lakes without an outflow polynya had outflow stream orders of 1 or 2; the other two were stream order 3. Sixteen of 22 lakes with an outflow polynya had outflow stream orders of 3 or 4, and none had stream order 1.
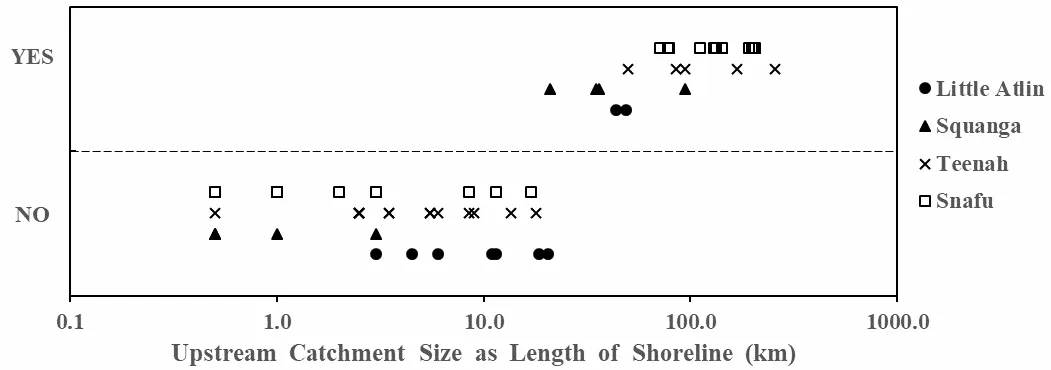
We found a fairly consistent threshold in upstream catchment size at which outflow polynyas begin to occur in a drainage, supporting our third hypothesis. The threshold between lakes with and without an outflow polynya occurred at an upstream catchment size of 10 to 20 km in Squanga drainage, and 30 to 40 km in other drainages (
). The smallest catchment size supporting an outflow polynya had 21 km of upstream shoreline. There was no size range of upstream catchment shoreline length that fell in both categories, with and without outflow polynya (
). Taken together, these results indicate that the further downstream along the principal stream in a drainage the higher the likelihood of an outflow polynya.
. Upstream catchment sizes, estimated as upstream shoreline lengths (km), for lakes with an outflow polynya (“YES”) are consistently higher than for lakes without an outflow polynya (“NO”). Catchment size (x-axis) is displayed on a logarithmic scale for better graphical separation of the smaller lakes. Symbols indicate individual lakes in each of the four drainages sampled.
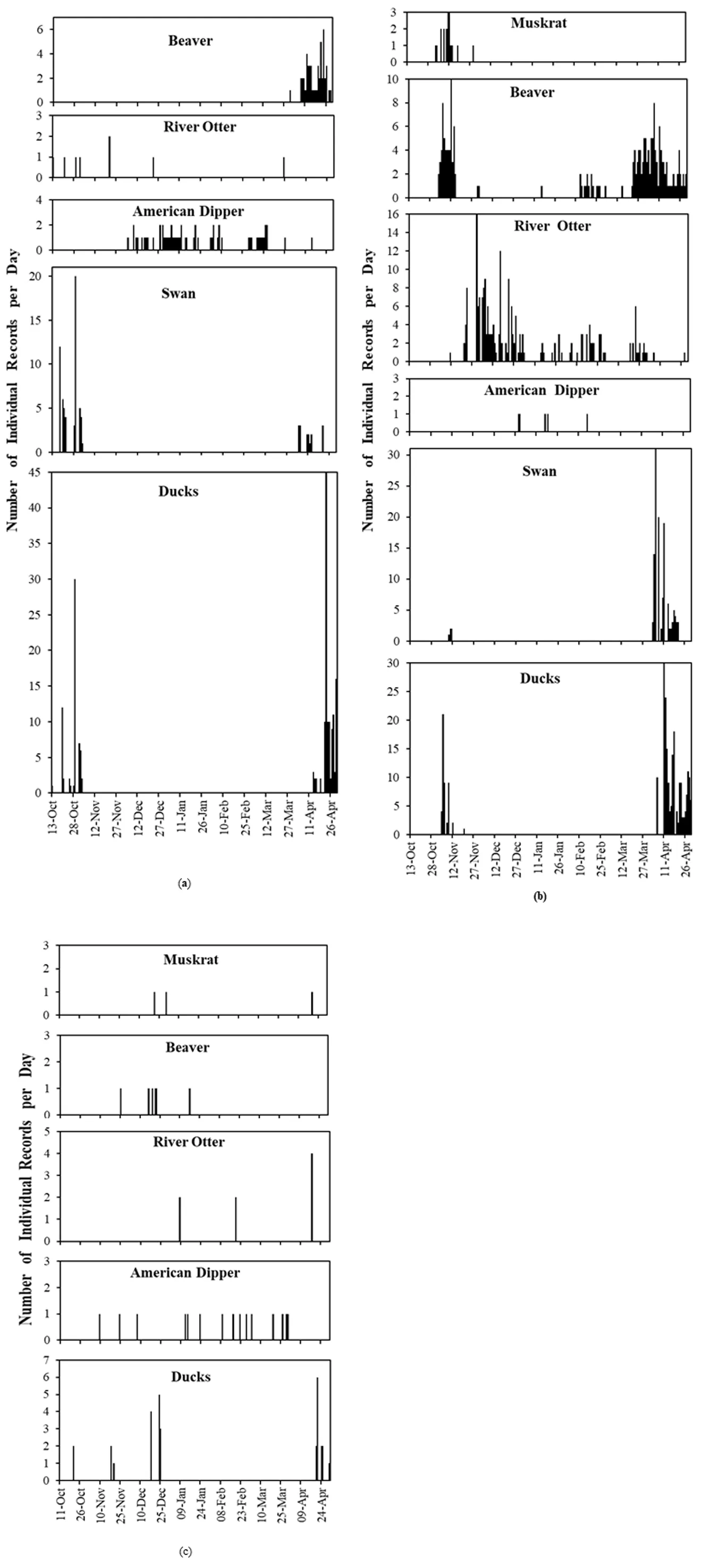
Regarding semi-aquatic mammals, the continuous () and periodic (Figure S1) camera trapping revealed that only some semi-aquatic mammals used open water sites from freeze-up right through to start of break-up in late April. River otters were most prominent and consistent through the seasons, with beavers also prominent but using polynyas less in the middle of winter. Muskrats and mink were rarely recorded.
. Histograms of relative abundance (number of individual records per day) of semi-aquatic mammals, American dipper, swans, and ducks daily through the winter at: (<b>a</b>) site A (2016–17); (<b>b</b>) site B (2016–17); (<b>c</b>) site C (2014–15). “Swans” were most often Trumpeter Swans (<i>Cygnus buccinator</i>), and “Ducks” include various species listed in the text. See Methods for how “Individual Records” were calculated from photos.
River otters were recorded throughout winter, but this species used sites in the middle of winter (mid-November to the end of March) much more than other species. Photos captured them alone or in groups as large as five (b). Their presence at a site was often separated by substantial time with no recorded activity, suggesting that they moved away from the site(s) quite often. Otters exhibited a wide diversity of behaviors, but we could not observe underwater foraging with our cameras. Foraging included otters eating fish on the ice edge (). This may have been under-estimated, because, based on our observations at other sites, otters sometimes take freshly-caught fish back under the ice, perhaps to shoreline air cavities under ice or bank burrows (). We mostly recorded otters swimming in the open water, or above-ground at their communal latrines and established trails where they groomed, investigated scent, scent-marked, and played ().
During our repeated visits over at least 5 winters, we had clear evidence of otters using all 28 outflow polynyas we sampled.
The closest active beaver lodge to each site was 425 m at site A, 150 m at site B, and 275 m at site C. Beavers were frequently recorded, especially at the closest polynya to an active lodge that may have been directly accessed under the ice. They were mostly recorded as individuals but occasionally in groups of 2 or 3. Records of foraging were based on beavers eating vegetation on the shoreline, including ice edge, and of beavers carrying recently cut shrub branches (). Most occurred in late winter (February through April), with a few in November. Most often beavers were recorded swimming or walking through the field of view. Building territorial scent piles and marking them, especially in late winter (late March and April), were fairly common beaver activities ().
Although rarely recorded by cameras ( and ), muskrats and American mink occasionally used open water for foraging. Our observations in all seasons confirm this relative rarity.
Regarding overwintering aquatic birds, only American dippers were regularly present throughout the entire winter, especially from November through March ( and Figure S1). They were recorded by cameras at all sites, and always as individuals in the continuous record. They were first recorded no earlier than 9 November and often considerably later, were present through the heart of winter (December through March), and were last recorded on 15 April (). Cameras most often recorded them foraging (), as inferred from their perching and posture at the water’s edge (associated with diving into the water), standing in shallow water, and occasionally with prey in their bills.
During our repeated visits over at least 5 winters, we observed American dippers using 17 of 28 outflow polynyas. They may have used a greater proportion because individual birds could have used more than one polynya along a stretch of stream, and could have been present without being active and observable.
Of the few species of ducks and swans known to overwinter regionally, we only occasionally recorded common merganser (
Mergus merganser) and mallard (
Anas platyrhynchos) ducks from November to January on the relatively small polynyas we sampled (Site C,
). They were foraging, perhaps on small fish which, in some photos, were breaking through the surface of fairly deep water above a beaver dam, which is typical of the time of year and behavior of least cisco (
Coregonus sardinella) when spawning [
24].
Regarding migratory aquatic birds, the data support our hypothesis of substantial use of polynyas in spring (starting late March), when lakes were still frozen but polynyas were expanding due to melt of ice at their edges, and also in late autumn (mid-October to as late as mid-December) when lakes were already mostly frozen (
and
Figure S1). The vast majority of swans were trumpeter swans (
Cygnus buccinator), and we plot them separately from other waterfowl for comparison to the data from other studies of outlet polynyas on larger lakes in the region. We did not record them after mid-November; the earliest spring recording was 23 March. Swans were often recorded individually but in groups as large as twelve (Mean group size 2.5; Modal group size 1). Cygnets were often present in early winter. Swans were most often foraging, as inferred from food in the bill or posture with head underwater, or they were swimming, which might have included searching for forage (
). They also came onto the ice edge where they rested, slept, and groomed. Swans likely choose polynyas based on the depths of water that would allow them to reach aquatic macrophytes growing in the lake sediments.
The “ducks” (Anatidae) totalled 520 records, 18.5% of which we could not identify to species because they were too far from the camera or in poor lighting. In order of decreasing relative abundance, their proportional contribution to the total records was mallard (62.1%), bufflehead (
Bucephala albeola) (7.1%), common merganser (5.4%), Barrow’s goldeneye (
Bucephala islandica) (3.4%), green-winged teal (
Anas carolinensis) (1.2%), ring-necked duck (
Aythya collaris) (1.2%), common goldeneye (
Bucephala clangula) (1.0%), and lesser scaup (
Aythya affinis) (0.2%). During our sampling period, our cameras did not record any grebes (Podicipedidae) or loons (Gaviidae), although some species from these families regularly used lakes associated with our sites in summer.
Collectively, these ducks, like swans, had generally left open water areas in autumn by mid-November, and first appeared in spring in the last week of March or early April, a little after the arrival of swans. Like swans, ducks were most often foraging, as inferred from their head underwater in dabblers, or swimming. Swimming might have included searching for food, especially for diving ducks that could not be recorded underwater (
). Ducks came onto the ice edge to rest and sometimes were clearly vigilant regarding activity elsewhere.
A considerable diversity of other bird and mammal species visited the sites, being recorded in the continuous camera trapping record (
). Of the 5 species of birds, the great horned owl was generally perched in trees overlooking open water, and likely hunting. We found one muskrat freshly killed by a great-horned owl at site C in 2014–15, but the action was not captured on camera. The bald eagle (
Haliaeetus leucocephalus) was likely hunting for fish or ducks. The three species of Corvidae were foraging by scavenging at locations where river otters had eaten fish on the ice and where beavers had marked scent piles. Of the 11 species of mammals, only the American mink and the moose (
Alces alces) were foraging in the open water. In their autumn and spring visits, moose were in the water and likely had foraged for aquatic macrophytes. The other species (notably Canada lynx (
Lynx canadensis) and American marten (
Martes americana)) appeared attracted to the edges of open water because of scent marking, especially by beavers and river otters. At other visits, the various mammalian carnivores may have been prospecting for potential prey, such as waterfowl. Some records may have been incidental travel.
.
Primary behaviors of the semi-aquatic mammals and aquatic birds from the continuous photo sampling, quantified as percentage of all Records of the species. “Records” refers to the number of individuals recorded on photos per day summed across all days, where photos repeatedly showing an individual of the same species, with less than a two-hour interval between consecutive photos, were judged to be showing the same individual (i.e., one record).
|
|
|
Percent of Species’ Records by Behavior Category |
|
|
|
Forage |
Travel |
Rest |
Play |
Groom |
Vigilant |
Mark |
Other |
| Site |
Species * |
Records |
|
Fly |
Swim |
Walk |
|
|
|
|
|
|
| A |
Beaver |
52 |
2 |
|
82 |
12 |
|
|
|
|
4 |
|
|
Otter |
8 |
|
|
38 |
50 |
|
|
12 |
|
|
|
|
Dipper |
58 |
98 |
2 |
|
|
|
|
|
|
|
|
|
Swan |
76 |
78 |
|
17 |
|
2 |
|
3 |
|
|
|
|
Ducks |
207 |
38 |
|
45 |
2 |
15 |
|
|
|
|
|
| B |
Beaver |
193 |
12 |
|
71 |
12 |
|
|
|
1 |
5 |
|
|
Muskrat |
18 |
|
|
100 |
|
|
|
|
|
|
|
|
Otter |
210 |
|
|
6 |
49 |
21 |
7 |
3 |
6 |
8 |
|
|
Dipper |
5 |
80 |
|
|
|
|
|
|
20 |
|
|
|
Swan |
132 |
44 |
1 |
46 |
|
5 |
|
1 |
|
|
|
|
Ducks |
265 |
18 |
1 |
80 |
|
1 |
|
|
|
|
|
| C |
Beaver |
6 |
|
|
17 |
83 |
|
|
|
|
|
|
|
Muskrat |
3 |
|
|
33 |
67 |
|
|
|
|
|
|
|
Otter |
8 |
75 |
|
|
25 |
|
|
|
|
|
|
|
Dipper |
14 |
86 |
|
|
|
|
|
|
14 |
|
|
|
Ducks |
41 |
49 |
|
27 |
5 |
|
|
|
14 |
|
5 |
.
Number of independent sightings (records) by month for the less frequently observed species and the dominant behaviour they exhibited in the photos.
|
|
Month |
Dominant |
| Species |
Site |
Oct |
Nov |
Dec |
Jan |
Feb |
Mar |
Apr |
Behavior |
Bald eagle
(Haliaeetus leucocephalus)
|
B |
|
|
|
|
|
1 |
|
Foraging |
| C |
1 |
1 |
|
|
|
|
2 |
Foraging |
| Black-billed magpie (Pica hudsonia) |
B |
|
|
|
|
|
4 |
1 |
Foraging |
| Common raven (Corvus corax) |
C |
|
|
|
3 |
|
|
1 |
Foraging |
Great horned owl
(Bubo virginianus)
|
A |
|
|
|
|
|
1 |
|
Foraging |
| B |
|
|
|
|
|
|
3 |
Foraging |
| Canada jay (Perisoreus canadensis) |
B |
|
|
|
|
|
8 |
12 |
Foraging |
| Black bear (Ursus americanus) |
C |
|
|
|
|
|
|
2 |
Travel–walking |
| Red fox (Vulpes vulpes) |
B |
|
|
1 |
|
|
1 |
|
Travel–walking |
| Coyote (Canis latrans) |
B |
|
|
1 |
3 |
1 |
|
|
Travel–walking |
| C |
|
1 |
|
|
|
|
1 |
Travel–walking |
| Grey wolf (Canis lupus) |
B |
|
|
|
1 |
1 |
|
|
Travel–walking |
| Canada lynx (Lynx canadensis) |
A |
|
|
|
|
|
1 |
|
Travel–walking |
| B |
|
1 |
3 |
8 |
2 |
2 |
|
Travel–walking |
| C |
|
|
|
|
|
|
1 |
Travel–walking |
| American mink (Neogale vison) |
B |
|
|
|
|
|
|
2 |
Travel–walking or Foraging |
| American marten (Martes americana) |
B |
|
|
|
1 |
|
|
|
Smelling site |
| Snowshoe hare (Lepus americanus) |
B |
|
1 |
|
|
|
|
|
Travel–walking |
| American red squirrel (Tamiasciurus hudsonicus) |
B |
|
|
|
|
4 |
|
|
Travel–walking |
| Mule deer (Odocoileus hemionus) |
A |
|
1 |
|
|
|
|
|
Travel–walking |
| Moose (Alces alces) |
A |
4 |
1 |
|
2 |
|
|
6 |
Travel–walking |
| B |
|
|
|
|
|
|
4 |
Travel–walking |
4. Discussion
4.1. Frequency and Correlates of Open Water in Winter
Freshwater polynyas are recurring and predictable, though only seasonal, winter habitat features on numerous small to medium-sized lakes in the mountainous region of the northern boreal biome. This is the first detailed documentation of their occurrence on lakes other than the very large lakes (>10,000 ha) that have dominated hydrological studies in this region [
7]. As with the largest lakes, these polynyas on smaller lakes predominantly occur in three hydrological settings: lake inflows, major constrictions or narrows in the body of a lake, and lake outflows. They also occur as seeps of groundwater along lake margins, though these are very small and more ephemeral during the course of the winter. Outflow polynyas are the most common and persistent, and also the largest because they consistently keep water open in streams draining the lakes.
Persistent open water during at least six months of ice-cover depends on an ongoing supply of water draining into ice-covered lakes from upstream for the duration of winter. Inflow currents push water through the lakes, under the ice, feeding the outflow and drawing warmer water from depth towards the outflow polynyas [
7]. We show that headwater lakes, on first and second-order streams with limited upstream catchment, lack persistent open water after freeze-up, so likely lack enough inflow to push water to the outflow. There is a clear threshold of upstream catchment size above which outflow polynyas, at least, are predictable, and regularly occur on main valley lakes with stream order of 3 or 4. As catchment size increases, stream and groundwater inflows increase, enhancing the likelihood of enough current to keep outflows ice-free. We hypothesize that mountainous settings, such as our study area, enhance the likelihood of outflow polynyas because ground water is readily stored during summer and autumn in headwater drainages at higher elevations than the main valley bottoms. In this scenario, the higher elevation provides a hydraulic head, perhaps in a set of perched water tables (unconfined aquifers), to induce downslope flow and feed water through the drainage [
25] long after the ground surface is snow-covered and often frozen. By contrast, we expect relatively flat regions with similar seasonal patterns to support fewer lakes with persistent open water in headwater drainages. For example, there were no outflow polynyas on lakes, other than one with stream order 5, during field studies of the North American river otter set in headwater drainages in the non-mountainous Boreal Plains ecozone of Alberta (55°30′ N Lat), with a duration of winter ice cover similar to the current study [
26].
The clear association of outflow polynyas with higher stream order and larger catchments means that most are at relatively low elevations. Consequently, conservation of the fauna relying on the polynyas during winter and spring depends on protecting an adequate collection of lower elevation lakes, focusing on those that provide the best foraging opportunities for the birds and mammals in question. Conservation also means removing the risk that human activity, notably noise and movement, near polynyas will alienate these key habitats from the dependent species.
We do not expect that a threshold of upstream drainage size identified to support outflow polynyas in one particular drainage will necessarily be characteristic of all drainages in a region. Various factors influencing groundwater storage, distribution, and flow regime will differ between drainages. For instance, the karst bedrock geology in parts of our study area may have enhanced underground water storage in summer, making it more available for the duration of the following winter.
Permafrost also plays a major role in groundwater hydrology at northern latitudes [
27]. With continuous permafrost, groundwater storage and flow are confined mostly to the active layer (ground thawing in summer) above the permafrost, so groundwater is (i) more likely to drain above the permafrost into streams during summer than be stored in deep aquifers, or (ii) will become immobile when the active layer freezes in autumn and early winter [
27]. We expect that, in regions with continuous permafrost, such as mountainous regions to the north of our study area [
21], liquid ground water would be less available, and lakes less likely to have winter polynyas, than in regions with discontinuous or no permafrost, such as our study area and much of northern British Columbia. The overheating climate is acting as an experiment in subarctic regions to illustrate this pattern. The active layer is deepening because the upper layers of permafrost are melting, and sometimes not re-freezing in winter. This provides more space for liquid groundwater storage and flow on top of, or through, the remaining permafrost, even in winter, and leads to increases in flows in groundwater-charged streams in subarctic regions [
28].
Given projected increases in precipitation and winter temperatures in most northern boreal portions of North America [
29], we expect continued occurrence of winter polynyas on lakes in our study area and other mountainous boreal regions with little or no permafrost, including much of southern Yukon and northern British Columbia.
4.2. Mammal and Bird Species Diversity and Behaviors
Outflow polynyas, and the flowing open water they produced downstream, provided American dippers and river otters with reliable access to water for foraging throughout the six to seven months of ice cover on lakes and notably in the heart of winter (November through February) when other semi-aquatic mammals and birds rarely visited the open water. These stretches of second to fourth-order stream can be excellent foraging habitat for dippers. The birds feed on aquatic invertebrates and small fish, which they catch on the stream bed and in the water column [
30,
31,
32]. The water must be largely ice free and clear for the birds to readily search the stream bed while perched on shore, and complete their diving and swimming attacks. Relatively shallow streams without ice cover are a very useful winter habitat for dippers.
American dippers are confined to mountainous regions of western North America, where they can be resident year-round on stretches of stream or river, but also seasonal migrants moving to higher elevation streams after ice melt for nesting and summer seasons [
33,
34]. We do not know the life history of those we observed, but infer that many were seasonal migrants because camera traps did not record them in October. During the summer we rarely observed them at the streams that were open in winter. The open water streams we studied were short, and, after being used regularly by even one dipper through a winter, may not be able to sustain a nesting pair with offspring unless the contiguous summer habitat were much more extensive than the length of open water in winter. We suggest that the relatively high incidence of outflow polynyas and open water in downstream reaches, associated with small to medium-sized lakes in mountainous compared to non-mountainous boreal regions, is a major determinant of the distribution of the American dipper in the boreal portions of its range. Mountainous relief, with perched water tables, seems to provide the necessary ground and surface water flow regime to power the through-lake currents and associated upwelling necessary to maintain polynyas. In flatter terrain, flow would be significantly reduced, especially in groundwater. Outflow polynyas and associated open streams likely comprise the majority of the winter habitat for dippers in the northern boreal mountains where shallow streams are otherwise frozen over [
2], unless kept open by inflow from springs and groundwater [
35]. These key winter habitats need conservation because of their small size and irreplaceability for American dippers.
Open water in winter is probably less essential for river otters than dippers but still highly valuable. River otters can overwinter in boreal regions with virtually no open water as long as they can access water via den structures (mostly old beaver lodges and bank burrows) that have both above-ground and underwater entrances [
26]. Otters need access to water to catch fish, their primary prey in winter [
36], and most fish species, especially in size classes large enough as useful otter food, reside in lakes rather than streams during winter in headwater reaches of the Yukon River drainage [
24,
37]. Frequent, though small, patches of open water at lake outflows and inflows provide otters with substantial foraging opportunities. They visited and often used all the open water patches we sampled, using them as centers for a wide variety of activities. They likely swam considerable distances from polynyas under ice to reach underwater entrances to dens (beaver lodges, bank burrows, air cavities under subsided ice) and expand the lake volumes they searched for fish.
The photo record showed that otters stayed at polynyas intermittently, but not continuously through the winter. This is best explained by denning availability, and by fish availability through time. Otters require an above-ground, somewhat insulated, den with ready access to the open water. Beaver lodges are ideal, and burrows, also often dug by beavers in shoreline banks, are also useful [
26]. These were not always available within under-ice swimming distance of a polynya. Otters appeared to use some polynyas only for short periods during travel between various polynyas. In one instance, we found an otter using a den dug out of a snowdrift, and lined with dead grass, immediately beside a polynya. Regarding fish availability, otters’ foraging likely reduces fish abundance quite quickly within the small portion of a lake accessible from the limited open water, as fish are captured or induced to escape. Otters would respond to this declining availability by moving to another polynya where fish are more abundant in the portion of the lake available for foraging. They could return to a previously used polynya after enough time has passed for the fish to lose their aversion and return to the pelagic and benthic zones close to the polynya.
The question of how fish respond to persistent open water in winter was outside our scope, but it is of high interest to understand these seasonal ecosystems [
38] and their use by piscivorous mammals and birds. We discuss two issues fish face: oxygen availability for all species and early winter spawning habitat for a few species. The dissolved oxygen content of water in boreal lakes with long periods of ice cover generally declines as winter progresses, sometimes leading to oxygen starvation and winter-kill of fish [
4,
39]. Although we did not observe winter-kill in any of the drainages in this study, we are aware of a shallow lake in the same ecoregion where this has occurred in some winters and where fish congregate in small patches of water kept open by groundwater seeps. Northern boreal streams with open water have somewhat higher oxygen concentrations than areas under ice because of some re-aeration of water [
40]. So, it seems possible that inflow polynyas fed by open water streams with a gradient sufficient for aeration will have somewhat elevated oxygen concentrations compared to the main body of the lake. This relatively high oxygen concentration may attract fishes to polynyas, establishing interesting trophic relationships in the fully aquatic portion of the food web, and also raising the value of these sites for foraging otters and piscivorous waterfowl. Regarding spawning habitat, the open water streams that were part of four of our sampled outflow polynyas were either known spawning sites for a morph or ecotype of the lake whitefish (
Coregonus clupeaformis) [
41] or gathering places for large groups of this species moving in streams during spawning season. These polynyas attracted substantial numbers of otters during fish spawning.
Contrary to our hypothesis, polynyas were not heavily used by other semi-aquatic mammals, especially in mid-winter. The beavers’ reduced activity in mid-winter probably indicates energy conservation by avoiding colder air temperatures and moving less from their lodges. Muskrats may have been limited by insufficient foods (notably aquatic macrophytes) in the vicinity of polynyas. Mink may be limited regionally by the competition from the prominent river otter population, and by the scarcity of muskrats, a key prey species.
In late autumn and spring, waterfowl foraged and rested at all polynyas we sampled, but the birds rarely visited these relatively small polynyas in winter, possible because of insufficient food to sustain them for many months. Some trumpeter swans and ducks (notably mallards and goldeneye) overwinter at the large polynya draining Teslin Lake (c. 45 km northeast of our continuous camera trapping). Even small polynyas can be valuable over shorter periods for the foods they provide: aquatic macrophytes (notably
Potomageton spp.) for swans and dabbling ducks; zooplankton, invertebrates and small fish for diving ducks. Ducks are still present after late October and have not joined the peak migration, but they might try to spend winter in the region.
In the spring, waterfowl visited our small polynyas during April, coincident with the main pulse of migration observed on the large regional lakes, which can vary significantly because of weather variations among years [
1,
3]. Our data cannot be readily converted to absolute counts, but give a similar picture of waterfowl abundance as the direct counts in early May 1983 on polynyas ranging in size from 0.1 to 3.0 ha [
3], similar to those we surveyed. These aerial surveys in 1983 were in other headwater drainages to the Yukon River, and associated with Teslin Lake, and found a mean of 63 individuals (100% ducks) at 8 lake outflows, and a mean of 22 individuals (99% ducks) at 20 lake inflow polynyas [
3]. The migratory grebes and loons might have been absent in spring before May because they need more extensive open water with their tendency to be pelagic foragers, also often requiring long stretches of open water to take off. Consequently, their spring migration is delayed with respect to ducks and swans.
Female waterfowl in particular have higher energy needs in spring as they have to produce eggs and incubate them. Their time in the vicinity of nesting sites but before egg-laying may be crucial for augmenting the stock of nutrients they bring with them from wintering grounds to proceed with reproduction in northern latitudes, especially given the benefits of early laying to reproductive success [
12,
13]. Polynyas are likely highly valuable to these waterfowl in spring, either as stop-over sites on the migration path, or resting and foraging sites close to the nesting destinations. In late May, the relative abundances of apparent nesting pairs of the most abundant ducks (Anatidae) on the drainages we sampled (including with our continuous camera record), in decreasing order were mallard (17.2%), scaup (17.0%), American wigeon (12.2%), Barrow’s goldeneye (12.2%), bufflehead (11.9%), green-winged teal (7.2%), ring-necked duck (5.6%) and common goldeneye (5.2%), based on aerial counts [
42]. This set of species and their relative abundances was quite similar to those we recorded at polynyas in April, supporting the likely value of the small polynyas we studied as early spring foraging sites for locally nesting waterfowl. The ecological value of the big polynyas on major regional lakes has been recognized, and some have been designated as protected areas [
23,
43]. Here we show that there are many smaller polynyas providing early season, highly dispersed, foraging sites several weeks before the onset of egg-laying. Historically, that occurred from mid-May to mid-June in these ducks [
2], but, with a warming climate, it is likely advancing as the timing of ice melt advances in spring [
17] and waterfowl respond by migrating earlier [
44]. These small polynyas are localized, but predictable, sites also worthy of special attention from a waterfowl conservation perspective.
5. Conclusions
Freshwater polynyas are recurring and predictable, though only seasonal, winter habitat features on numerous small to medium-sized lakes in mountainous regions of the northern boreal biome where permafrost is absent, sporadic, or discontinuous. During at least six months of ice-cover, such persistent open water depends on an ongoing supply of water draining into ice-covered lakes from upstream. We show that headwater lakes, on first and second-order streams with limited upstream catchment, lack persistent open water after freeze-up. There is a clear threshold of upstream catchment size above which outflow polynyas, at least, are predictable, and regularly occur on valley-bottom lakes with stream order of 3 or 4. Mountainous relief may be a necessary condition for the relatively high incidence of polynyas at northern boreal latitudes.
River otter and American dipper regularly forage at polynyas throughout winter, with dippers likely relying on them for survival. In spring, migratory waterfowl (Anatidae: swans and ducks) forage and rest at polynyas, likely gaining significant nutritional value for spring reproduction from these spatially-limited ecosystems.
Without previous winter-long study, these temporary and spatially-limited ecosystems on a wide diversity of northern boreal lakes have not been well mapped and appreciated. Based on our new findings, we urge greater attention to their conservation, to protect these habitats for semi-aquatic mammals and birds, and to acquire better knowledge of their aquatic ecology. Future work needs to more completely quantify the incidence and hydrological correlates of winter open water in other boreal regions (especially non-mountainous). It needs to quantify the spatial buffers and timing windows around polynyas within which human activity must be constrained to ensure undisturbed use of these high value and sometimes irreplaceable habitats by semi-aquatic mammals and water birds.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/665, Figure S1: Histograms of relative abundance (number of individual records per day) of semi-aquatic mammals, American dipper, swans, and ducks as recorded by periodically operating camera traps at: (a) site A (2013); (b) site A (2014–15); (c) site B (2014–15); (d) augmented site C (2014–2015). “Swans” were most often Trumpeter Swans (Cygnus buccinator), and “Ducks” includes various species listed in the text. See Methods for how “Individual Records” were calculated from photos. Portions of graphs blocked out in grey are time periods without camera trapping.
Acknowledgments
We thank the Carcross/Tagish First Nation and the Teslin Tlingit Council for the opportunity to work in their traditional territories. We thank the administration staff at Wildlife Conservation Society Canada for assistance in acquiring funding and for their management of financial and logistical processes.
Author Contributions
Conceptualization, D.G.R. and M.C.Y.L.; Methodology, D.G.R. and M.C.Y.L.; Validation, D.G.R. and M.C.Y.L.; Formal Analysis, D.G.R., M.C.Y.L. and P.K.; Investigation, D.G.R., M.C.Y.L. R.R. and P.K.; Resources, D.G.R., M.C.Y.L. and R.R.; Data Curation, D.G.R., M.C.Y.L. and P.K.; Writing—Original Draft Preparation, D.G.R.; Writing—Review & Editing, D.G.R., M.C.Y.L., R.R. and P.K.; Visualization, D.G.R. and M.C.Y.L.; Project Administration, D.G.R.; Funding Acquisition, D.G.R.
Ethics Statement
Ethical review and approval were waived for this study, because camera trapping with automated remote cameras was deemed non-invasive and non-interventionary to the mammals and birds being recorded, and because the jurisdiction of Yukon only requires wildlife research permits for projects involving live-capture and handling of animals.
Informed Consent Statement
Not applicable.
Data Availability Statement
An archive of images captured and data analysed in this study is available as part of the Wildlife Conservation Society Canada’s data storage portal: https://show.pics.io/don-reid/open-water-in-winter (accessed 5 September 2025).
Funding
This research was funded by The Weston Family Foundation and by the T-Gear Foundation, with grants to Wildlife Conservation Society Canada. The APC was funded by the journal.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Mossop DH. Studies of Waterfowl Staging Areas: Winter 1976; Game Branch, Yukon Environment: Whitehorse, YT, Canada, 1976. Available online: https://emrlibrary.gov.yk.ca/environment/studies-of-waterfowl-staging-areas-winter-1976.pdf (accessed on 4 September 2025).
-
2.
Sinclair PH, Nixon WA, Eckert CD, Hughes N. Birds of the Yukon Territory, 1st ed.; UBC Press: Vancouver, BC, Canada, 2003.
-
3.
Johnson WG, McEwen CA. Spring Staging of Waterbirds at Early Open Water Sites in the Yukon River Basin; Canadian Wildlife Service: Whitehorse, YT, Canada, 1983.
-
4.
Kirillin G, Leppäranta M, Terzhevik A, Granin N, Bernhardt J, Engelhardt C, et al. Physics of seasonally ice-covered lakes: A review.
Aquat. Sci. 2012,
74, 659–682.
[Google Scholar]
-
5.
Leppäranta M. Freezing of Lakes and the Evolution of Their Ice Cover; Springer-Verlag: Berlin/Heidelberg, Germany, 2015. doi:10.1007/978-3-642-29081-7.
-
6.
Larsen P, Ahston G, Gosink J, Marcotte N, Müller A, Osterkamp TE. Thermal regime of lakes and rivers. In River and Lake Ice Engineering, 1st ed.; Ashton GD, Ed.; Water Resources Publications: Littleton, CO, USA, 1986; pp. 203–260.
-
7.
Carmack EC, Wiegand RC, Marles EM, Alford ME, Chamberlain VA. Physical Limnology of an Ice-Covered Lake with through-Flow: Lake Laberge, Yukon Territory; Paper No. 35; National Hydrology Research Institute: Saskatoon, SK, Canada, 1988.
-
8.
Bengtsson L. Mixing in ice-covered lakes.
Hydrobiologia 1996,
322, 91–97.
[Google Scholar]
-
9.
Dunbar MJ. Physical causes and biological significance of polynyas and other open water in sea ice. In Polynyas in the Canadian Arctic; Stirling I, Cleator H, Eds.; Occasional Paper Number 45; Canadian Wildlife Service: Ottawa, ON, Canada. 1981; pp. 29–43.
-
10.
Smith SD, Muench RD, Pease CH. Polynyas and leads: An overview of physical processes and environment.
J. Geophys. Res. C: Oceans 1990,
95, 9461–9479.
[Google Scholar]
-
11.
Williams WJ, Carmack EC, Ingram RG. Physical oceanography of polynyas.
Elsevier Oceanogr. Ser. 2007,
74, 55–85.
[Google Scholar]
-
12.
Drent RH, Fox AD, Stahl J. Travelling to breed.
J. Ornithol. 2006,
147, 122–134. doi:10.1007/s10336-006-0066-4.
[Google Scholar]
-
13.
Janke AK, Anteau MJ, Markl N, Stafford JD. Is income breeding an appropriate construct for waterfowl?
J. Ornithol. 2015,
156, 755–762.
[Google Scholar]
-
14.
Yukon Ecoregions Working Group. Yukon southern lakes ecoregion. In Ecoregions of the Yukon Territory: Biophysical Properties of Yukon Landscapes; Smith CAS, Meikle JC, Roots CF, Eds.; PARC Technical Bulletin No. 04-01; Agriculture and AgriFood Canada: Summerland, BC, Canada, 2004; pp. 35–38.
-
15.
Potié J, Reid D. Conservation of Lakeshore Zones in the Northern Boreal Mountains: State of Knowledge, and Principles and Guidelines for Planning and Management; Conservation Report no. 14; Wildlife Conservation Society Canada: Toronto, ON, Canada, 2021. doi:10.19121/2021.Report.39964.
-
16.
Wahl H. Climate; Smith CAS, Meikle JC, Roots CF, Eds.; PARC Technical Bulletin No. 04-01; Agriculture and AgriFood Canada: Summerland, BC, Canada, 2004; pp. 19–23.
-
17.
Perrin A, Jolkowski D. Yukon Climate Change Indicators and Key Findings 2022; Yukon University Research Centre: Whitehorse, YT, Canada, 2022; pp. 1–126. Available online: https://www.yukonu.ca/sites/default/files/inline-files/Indicators2022_FinalReport_Oct2022.pdf (accessed on 5 September 2025).
-
18.
Strahler AN. Quantitative analysis of watershed geomorphology.
Trans. Am. Geophys. Union 1957,
38, 913–920.
[Google Scholar]
-
19.
Baker BW, Hill EP. Beaver. In Wild Mammals of North America: Biology, Management and Conservation, 2nd ed.; Feldhamer GA, Thompson BC, Chapman JA, Eds.; The John Hopkins University Press: Baltimore, MD, USA, 2003; pp. 288–310.
-
20.
Melquist WE, Polechla PJ, Toweill D. River otter. In Wild Mammals of North America: Biology, Management and Conservation, 2nd ed.; Feldhamer GA, Thompson BC, Chapman JA, Eds.; The John Hopkins University Press: Baltimore, MD, USA, 2003; pp. 708–734.
-
21.
Bonnaventure PP, Lewkowicz AG, Kremer M, Sawada MC. A permafrost probability model for the southern Yukon and northern British Columbia, Canada.
Permafr. Periglac. Process. 2012,
23, 52–68. doi:10.1002/ppp.1733.
[Google Scholar]
-
22.
McKenna K, Flynn N. The Boreal Low zone of Yukon. In Southern Lakes Boreal Low Subzone (BOLsl): A Field Guide to Ecosite Identification, 1st ed.; Department of Environment, Government of Yukon: Whitehorse, YT, Canada, 2017; Chapter 5, pp. 1-39–1-45. Available online: https://yukon.ca/sites/default/files/env/env-field-guide-ecosite-identification.pdf (accessed on 5 September 2025).
-
23.
Southern Lakes Wildlife Coordinating Committee. Regional Assessment of Wildlife in the Yukon Southern Lakes Region: Species Status Assessment; Environment Yukon: Whitehorse, YT, Canada, 2012; Volume 2, pp. 1–168.
-
24.
McPhail JD. The Freshwater Fishes of British Columbia, 1st ed.; University of Alberta Press: Edmonton, AB, Canada, 2007.
-
25.
Şen Z. Practical and Applied Hydrogeology, 1st ed.; Elsevier: Amsterdam, The Netherlands, 2014.
-
26.
Reid DG, Code TE, Reid ACH, Herrero SM. Spacing, movements, and habitat selection of the river otter in boreal Alberta.
Can. J. Zool. 1994,
72, 1314–1324.
[Google Scholar]
-
27.
Woo M-K. Permafrost Hydrology, 1st ed.; Springer: Berlin/Heidelberg, Germany, 2012.
-
28.
Lamontagne-Hallé P, McKenzie JM, Kurylyk BL, Zipper SC. Changing groundwater discharge dynamics in permafrost regions.
Environ. Res. Lett. 2018,
13, 084017.
[Google Scholar]
-
29.
IPCC. Climate Change 2022: Impacts, Adaptation, and Vulnerability. Contribution of Working Group II to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Pörtner H-O, Roberts DC, Tignor M, Poloczanska EC, Mintenbeck K, Alegría A, et al., Eds.; Cambridge University Press: Cambridge, UK, 2022; pp. 1–3056. doi:10.1017/9781009325844.
-
30.
Thut RN. Feeding habits of the Dipper in southwestern Washington.
Condor 1970,
72, 234–235.
[Google Scholar]
-
31.
Ealey DM. Aspects of the Ecology and Behaviour of a Breeding Population of Dippers (Cinclus mexicanus) in Southern Alberta. Master’s thesis, University of Alberta, Edmonton, AB, Canada, 1977.
-
32.
Young DA, Kraft LF. Dippers catching fish during winter in Yukon Territory.
West. Birds 1981,
12, 141–142.
[Google Scholar]
-
33.
Morrissey CA, Bendell-Young LI, Elliott JE. Seasonal trends in population density, distribution, and movement of American dippers within a watershed of southwestern British Columbia, Canada.
Condor 2004,
106, 815–825.
[Google Scholar]
-
34.
Gillis EA, Green DJ, Middleton HA, Morrissey CA. Life history correlates of alternative migratory strategies in American Dippers.
Ecology 2008,
89, 1687–1695.
[Google Scholar]
-
35.
Gallagher CP, Lea EV. Overwintering habitat of American Dipper,
Cinclus mexicanus, observed in an Arctic groundwater spring feeding on Dolly Varden,
Salvelinus malma.
Arctic 2019,
72, 82–87.
[Google Scholar]
-
36.
Reid DG, Code TE, Reid ACH, Herrero SM. Food habits of the river otter in a boreal ecosystem.
Can. J. Zool. 1994,
72, 1306–1313.
[Google Scholar]
-
37.
Roberge M, Hume JMB, Minns CK, Slaney T. Life history characteristics of freshwater fishes occurring in British Columbia and the Yukon, with major emphasis on stream habitat characteristics.
Can. Manuscr. Rep. Fish. Aquat. Sci. 2002,
2611, 1–248.
[Google Scholar]
-
38.
Salonen K, Leppäranta M, Viljanen M, Gulati R. Perspectives in winter limnology: closing the annual cycle of freezing lakes.
Aquat. Ecol. 2009,
43, 609–616.
[Google Scholar]
-
39.
Greenbank J. Limnological conditions in ice-covered lakes, especially as related to winter-kill of fish.
Ecol. Monogr. 1945,
15, 343–392.
[Google Scholar]
-
40.
Schreier H, Erlebach W, Albright L. Variations in water quality during winter in two Yukon rivers with emphasis on dissolved oxygen concentration.
Water Res. 1980,
14, 1345–1351.
[Google Scholar]
-
41.
Bodaly RA, Clayton JW, Lindsey CC, Vuorinen J. Evolution of lake whitefish (
Coregonus clupeaformis) in North America during the Pleistocene: Genetic differentiation between sympatric populations.
Can. J. Fish. Aquat. Sci. 1992,
49, 769–779.
[Google Scholar]
-
42.
Kenyon JK, Spiewak EK. Waterbird Use of Agay Mene and Kusawa Territorial Parks, 2007; Ducks Unlimited Canada: Whitehorse, YT, Canada, 2008; pp. 1–20.
-
43.
Cooke HA. Securing a Wild Future: Planning for Landscape-Scale Conservation in Yukon’s Boreal Mountains; Conservation Report no. 9; Wildlife Conservation Society Canada: Toronto, ON, Canada, 2017.
-
44.
Pöysä H. Tracking ice phenology by migratory waterbirds: settling phenology and breeding success of species with divergent population trends.
J. Avian Biol. 2019,
50. doi:10.1111/jav.02327.
[Google Scholar]