Simulating Repetition Suppression and Enhancement in Infancy: An Interactive Approach

Simulating Repetition Suppression and Enhancement in Infancy: An Interactive Approach
Yang Ji *
Received: 05 September 2025 Revised: 10 November 2025 Accepted: 29 June 2026 Published: 14 July 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
Repetition suppression, a decrease in neural response to repeated stimuli, is considered “one of the most robust experience-related cortical dynamics” [1]. This phenomenon has been observed consistently across different sensory domains. It has been documented both through single-cell recordings in animal studies and brain imaging studies in humans. Interestingly, this phenomenon may manifest differently in infants. While a similar reduction in neural response has been observed when infants are exposed to repeated single syllable (3-month-old) [2], picture (6-month-old) [3], or action sequence (9-month-old) [4], repetition can lead to an enhancement of neural response in other instances. At the trial-by-trial level, neural responses often show enhancement during the initial learning phase of a task, followed by a gradual suppression as familiarity increases. This trend has been observed in studies involving visual categories (6-month-old) [5], syntactic/lexical speech patterns (neonates) [6,7], or pseudo-word (20-month-old) [8].
To reconcile the seemingly opposing phenomena of repetition suppression and enhancement, one possible explanation is perceptual integration efficiency, which is influenced by the interconnected factors of task complexity and individual differences in efficiency. Infants typically exhibit lower efficiency when performing more complex tasks. For example, Bouchon et al. [6] show that a more complex task elicits repetition enhancement, while a less complex task produces repetition suppression. Alternatively, for individuals with rapid perceptual integration, an objectively complex task may appear relatively easier than for those with lower efficiency. For example, Torkildsen et al. [8] demonstrate that high-ability individuals exhibit more rapid repetition suppression, while low-ability individuals show repetition enhancement with a reluctance to suppression. Therefore, whether repetition suppression or enhancement is observed may depend on infants’ subjective task experience (i.e., whether perceptual integration has been completed) [9]. However, further research is needed to elucidate the underlying mechanisms across the spectrum of repetition suppression and enhancement, particularly by integrating both phenomena into a unified computational framework [10].
To provide a unified view of repetition suppression and enhancement, this simulation study aims to offer a prediction-based approach that incorporates both bottom-up perceptual integration and top-down expectation. Furthermore, the prediction-based framework is situated in an environment where task-related processing competes with off-task novelty-seeking for shared temporal resources. Note that the term “expectation” in this paper does not equate with conscious awareness but merely refers to the graded predictive certitude about an outcome, whether implicit or explicit. Before introducing this prediction-based approach, however, the paper first discusses neuronal models typically used to simulate repetition suppression. While this study does not delve into the intricate details of neuronal models, it underscores the implication that such models may point toward a more integrated framework aimed at understanding infants’ experience-related neural dynamics.
1. Neuronal Models
Neuronal models examine how repeated presentations of a stimulus alter the response of neuronal populations at the perceptual level [1]. These models primarily focus on the response magnitude or latency of a neuronal population following repeated stimulation. One specific area of investigation within fatigue and sharpening models is how repetition affects stimulus-specific neural tuning curves. Fatigue and sharpening models differentiate themselves by proposing which one of these tuning curve parameters, peak value or precision, is primarily influenced by repeated stimulus presentation. The fatigue model proposes that repeated stimulation weakens the response of a neuronal population. This fatigue manifests as a decrease in the overall firing rate of the neurons [11,12]. To better capture the influence of stimulus-specific adaptation, the local-scaling model refines the fatigue model by considering the mismatch between the peak frequency of the neuronal population and the external stimulus frequency [11]. Fatigue occurs faster when the peak response aligns with the stimulus, and slower when they differ. On the other hand, the sharpening model contrasts the fatigue/local-scaling models by proposing that repetition suppression arises from increased neural precision, instead of generalized fatigue [11]; see also [12]. As a specific neuronal population repeatedly encounters a stimulus, its response becomes more refined, focusing on a narrow range of frequencies defined by its tuning curve. This focused response leads to a suppression for stimulus that fall outside of this preferred range, ultimately reducing the overall number of neurons responding to the repeated stimulus.
However, while neuronal models often focus exclusively on perceptual neuronal firing patterns, Vogels [13] suggests that stimulus-specific repetition effects cannot be solely explained by bottom-up perception. Repeated exposure to a stimulus also influences how we interpret it based on learning experiences. This suggests that top-down inputs, shaped by such predictions, are essential for understanding experience-related dynamics [1,11,14]. For example, Alink et al. [11] acknowledge that their local-scaling model cannot reproduce repetition effects without incorporating a β parameter. This parameter quantifies the rate at which adaptation changes in relation to the discrepancy between neural preference and stimulus property, suggesting a mechanism similar to prediction error that moderates the learning rate. On the other hand, neuronal sharpening is typically explained by a top-down predictive framework [14]. Following the sharpening view, we will explore later how specific learning mechanisms, driven by prediction error, can help discriminate between informative and less informative perceptual cues.
In contrast to models that focus on response magnitude, the facilitation model examines the temporal dimension of neural responses [1]. Information accumulates more rapidly for repeated stimuli, resulting in shorter latencies and attenuated neural responses. According to the accumulation model, information accumulation is primarily a stochastic process [15]. However, it is also plausible that information accumulation is guided by an implicit predictive process contributing to outcome anticipation [16]. From a prediction-based perspective, a reduction in prediction error (the discrepancy between the expected and actual stimulus) accelerates information flow through the system. Later, we will explore how response latency to an expected outcome may be influenced by its corresponding prediction error.
Taken together, fatigue/local-scaling and sharpening models suggest that some form of top-down influence, potentially related to a prediction-based learning mechanism, is required to explain local perceptual-level neural responses. Moreover, neural responses accumulated through perceptual integration may be further subject to latency-related neural attenuation.
2. An Interactive Model
The purpose of this article is to integrate previously introduced concepts, such as sharpening and facilitation, to demonstrate their interconnectedness within a prediction-based framework. Consider the typical experience of an infant interacting with their environment: they begin to form expectations when encountering a repeated stimulus or when a specific stimulus reliably predicts an outcome. Throughout this process, neural responses at both the bottom-up perceptual and top-down expectational levels gradually change in relation to their learning experience. Furthermore, these neural responses are not only tied to the target task but also to the infant’s explore-exploit behaviors within a broader ecological context. For instance, a sleeping infant’s state differs significantly from that of a wakeful infant, whose attention is constantly captured by both on-task and off-task activities in their surroundings. In the following sections, we describe how combining a discriminative learning mechanism with a latency function can integrate both sharpening and facilitation perspectives. Moreover, we show how a resource availability perspective can model the additional role of off-task activities.
2.1. Prediction-Based Learning Mechanism
In this article, the prediction-based framework is derived from the error-driven learning (EDL) mechanism. The standard EDL equation adapts the Rescorla-Wagner learning equation [17], which has successfully explained many Pavlovian conditioning phenomena [18]. Recently, EDL has been repurposed to simulate higher-level human cognition (see [19]; for an accessible introduction to the equation, also see [20]). For instance, in the context of early language acquisition, while EDL may not be optimally tuned for specific task performance, it excels at generalizing to new tasks following parameter optimization based on a single task (for a large-scale model comparison study, see [21]).
Conceptually, EDL is consistent with predictive coding (compare [22]) and is therefore primarily related to the sharpening perspective. Nevertheless, unlike predictive coding, EDL does not model graded perceptual hierarchies (e.g., auditory [23,24], visual [25]) or graded stimulus-specific adaptation [23,26]. Instead, the EDL architecture consists of only two layers—a cue layer representing perception and an outcome layer representing expectation—with a learning equation that updates cue-outcome associations. Additional assumptions within EDL, such as discriminative learning and common prediction error, give rise to critical properties like cue competition and outcome competition (to be described shortly).
Conveniently, EDL is supervised and atemporal. It is supervised because the modeler predefines the concurrence between perceived cues and expected outcomes during the learning phase. It is atemporal because the model updates cue-outcome associations on a trial-by-trial basis without considering directionality (e.g., cue-to-outcome perceptual integration followed by outcome-to-cue expectation) or continuous event-related epochs. This framework serves as an abstraction of the dynamic bidirectional processes involved, allowing the model to highlight the core learning mechanism and make trial-by-trial learning dynamics clearly interpretable [19]. Nevertheless, while not elaborated upon here, EDL can also be integrated into dynamic processing-based cognitive models [27]. Notably, EDL corresponds to basal ganglia function, which is characterized by excitatory and inhibitory pathways [28]. This evolutionarily ancient structure has been considered a learning hub for various higher-level cognitive functions (for a review, see [29,30]), a role reconfirmed by data-driven Granger causality analyses [31,32]. To illustrate the core learning mechanisms and properties of the two-layer EDL network, we provide schematic drawings framed within the context of the simulated empirical studies. More details on the learning equations for EDL can be found in the Supplemental Materials (Equations (S1)–(S3)).
Consider the repeated syllable “ba” from Nakano et al. [2], illustrated in the top panel of Figure 1. Here, the identical stimulus is conceptualized simultaneously as both the cue (representing bottom-up perception) and the outcome (representing top-down expectation). On the initial trial, both the cue “ba” (represented by the blue circles) and the outcome “ba” (represented by the black circle) are present, establishing the cue as a unique predictor of a unique outcome. Since no prior connection exists at this first presentation, the initial association weight is zero (indicated by the dotted line), rendering the initial prediction error maximal (visualized by the full blue bar labeled P.E. on the right). This maximal prediction error drives a substantial initial update to the associative weight between this specific cue-outcome pair. Consequently, across subsequent trials, the EDL network progressively strengthens this cue-outcome association (indicated by the bold connection line). As these learned associations accumulate to help predict the outcome, the resulting prediction error in subsequent trials is reduced (visualized by the diminishing blue bar), leading to less substantial additional updates to the associative weight. Alternatively, in the surprise condition illustrated in the bottom panel of Figure 1, a novel syllable, “pa” is perceived (indicated by the purple circles) and becomes expected (indicated by the red circle). Since there is initially no learned association between this new cue-outcome pair, the prediction error returns to its maximal level.
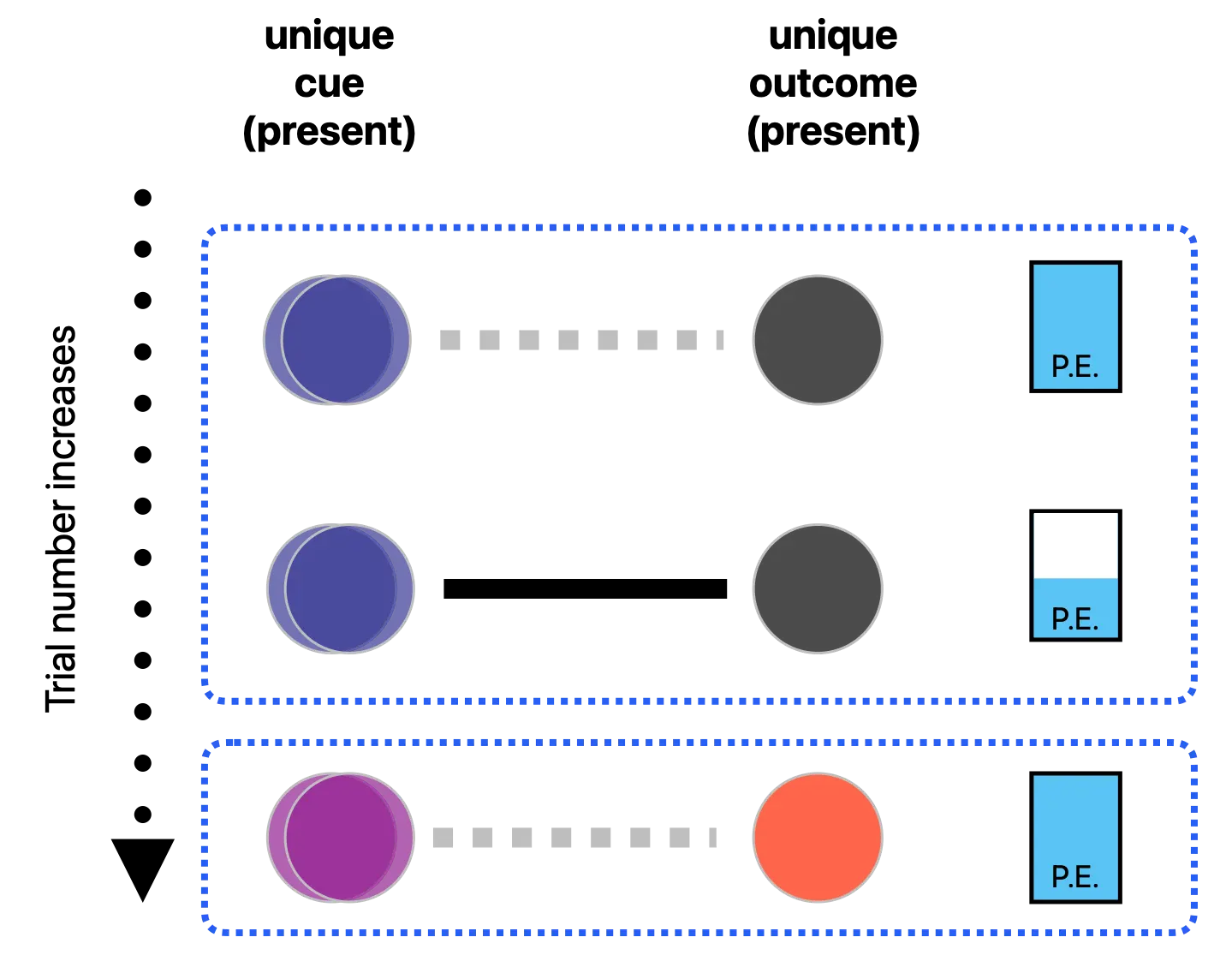
Figure 1. (Top panel): Updating of associative weights between a unique cue-outcome pair, with a concurrent reduction in prediction error (P.E. bar) across trials. (Bottom panel): The surprise condition, where a novel single syllable triggers the learning of a new association between a separate cue-outcome pair.
Another key property of the EDL model is the common prediction error. In the EDL model, this prediction error is common to a specific outcome rather than to individual cue-outcome associations. This limits the total associative weights that can be accumulated for any given outcome. For example, pre-existing associative weights will automatically reduce the learning rate of subsequent cue-outcome associations. In the extreme case where the common prediction error reaches zero, the network can no longer update or accommodate any additional cue-outcome associations for that outcome (i.e., blocking).
Since the cue-outcome pairs in both the repetition and surprise conditions of Nakano et al. [2] are unique, there is no discriminative learning where cues become differentially associated with an outcome. Instead, discriminative learning emerges when multiple cues—both unique and shared—are present to predict an outcome. For instance, in Emberson et al. [33], a squeak sound is paired with one specific smiley movement, and a rattle sound is paired with another. In such a scenario, the discriminative learning mechanism of the EDL network is engaged. As illustrated in the top panel of Figure 2, the currently present cue (the rattle sound, represented by the blue circles) uniquely predicts the active outcome (the corresponding smiley movement, represented by the solid black circle; the association indicated by the bold black connection line). Simultaneously, this same present cue acquires a negative association (indicated by the bold red connection line) with the absent outcome (the alternative smiley movement, represented by the faded black circle). Conversely, the absent cue (the squeak sound, represented by faded black circles) maintains a positive association with its alternative outcome (faded black circle), while developing a negative association with the currently present outcome (solid black circle).
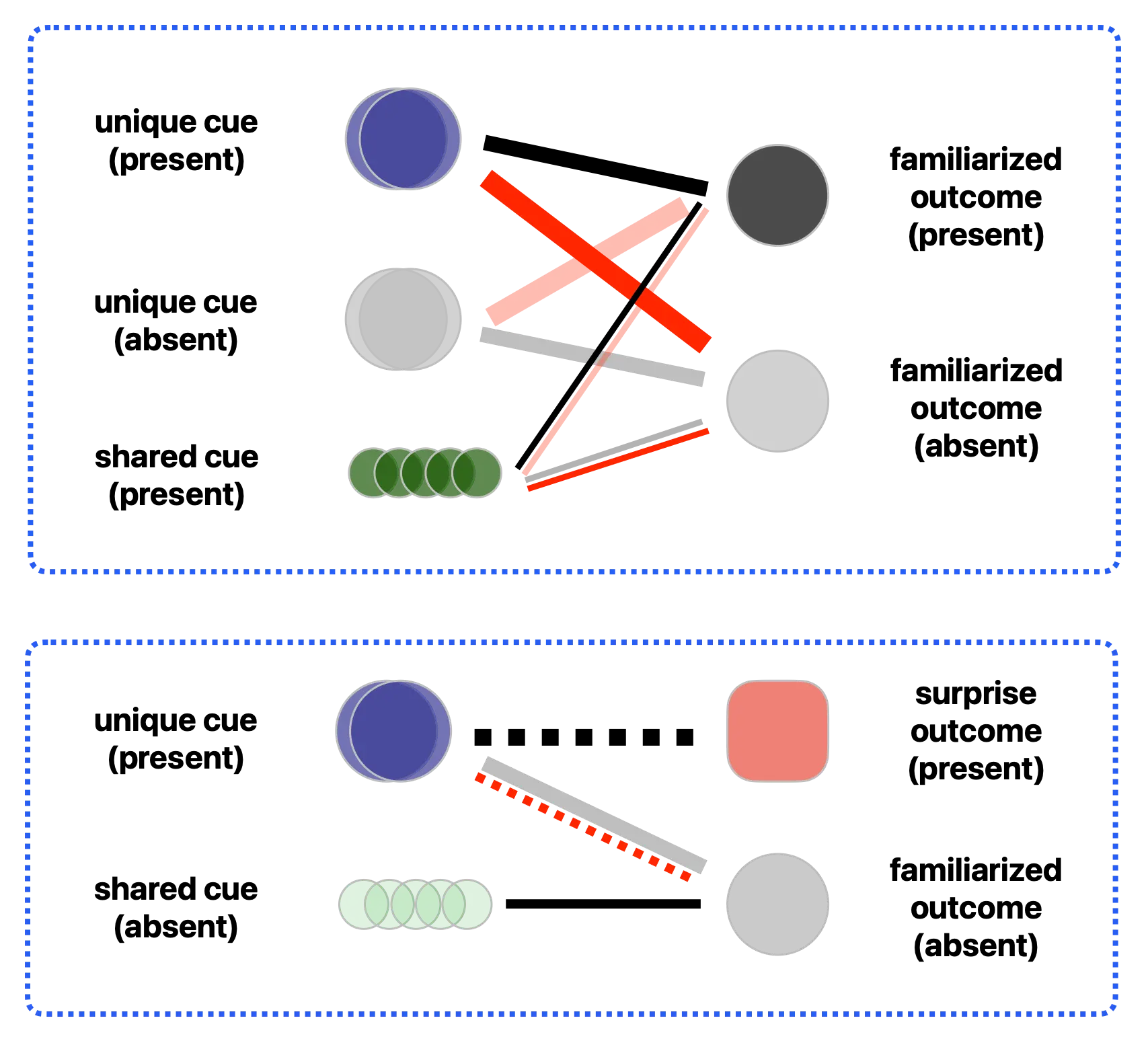
Figure 2. (Top panel): Discriminative learning of associative weights between unique and shared cues and outcomes. (Bottom panel): The surprise condition, where the unique cue signals a novel outcome.
In such a multi-cue scenario, there is also a shared cue (the smiley image, represented by the green circles). This shared cue develops a positive association (indicated by the thin black connection line) with the present outcome (solid black circle) and a negative association (indicated by the thin red connection line) with the absent outcome (faded black circle). However, in scenarios where the current outcome is absent, and the alternative outcome is present, the application of these positive and negative updates is reversed (as indicated by the thin faded connection lines). Consequently, the shared cue’s associative weights with both outcomes effectively cancel each other out over time. Through this mechanism, the shared cue is rendered less informative than the unique cues when predicting a specific outcome.
Furthermore, in this multi-cue scenario, the learned associative weights exhibit the property of outcome competition. As illustrated in the top panel of Figure 2, given the unique cue of the rattle sound and the shared cue of the smiley image, these concurrent predictors collectively drive the inference of one specific smiley movement over the alternative. In addition, discriminative learning results in cue competition. The unique cue progressively emerges as a more informative predictor of its corresponding outcome than the shared cue, whose associative strength is discounted. Note that this cue competition property depends on the common prediction error constraint outlined earlier; it cannot emerge if the prediction error is unconstrained at the outcome level, as occurs in typical associative learning frameworks.
In the surprise condition of Emberson et al. [33], shown in the bottom panel of Figure 2, the unique cue (the rattle sound, represented by the blue circles) is present, but the expected outcome (the smiley movement, represented by the red square) is suddenly replaced by a novel, unexpected outcome (the absence of movement, represented by the alternative red square). Since this switch causes the prediction error to return to its maximum value, the unique cue rapidly updates its weights—downgrading its association with the original outcome (indicated by the red dotted line) while developing a positive association with the new outcome (indicated by the black dotted line). Conversely, the shared cue (the smiley image, represented by the faded green circles) is absent during this surprise event. In this scenario, we assume that the pre-existing cue-outcome association that predicts the expected smiley movement remains embedded in the network’s learned matrix.
In real-time processing, the presentation of the unique cue (the rattle sound) would trigger a top-down expectation of the corresponding outcome (the internal inference of smiley movement) before that expectation is violated by the novel outcome (the absence of movement). Thus, the internal representation of the shared cue during this temporal window continues to be driven by top-down feedback from the expected outcome, despite the shared cue’s physical absence in the environment (indicated by the black connection line, see Simulation 2).
2.2. Latency Attenuation of Neural Response
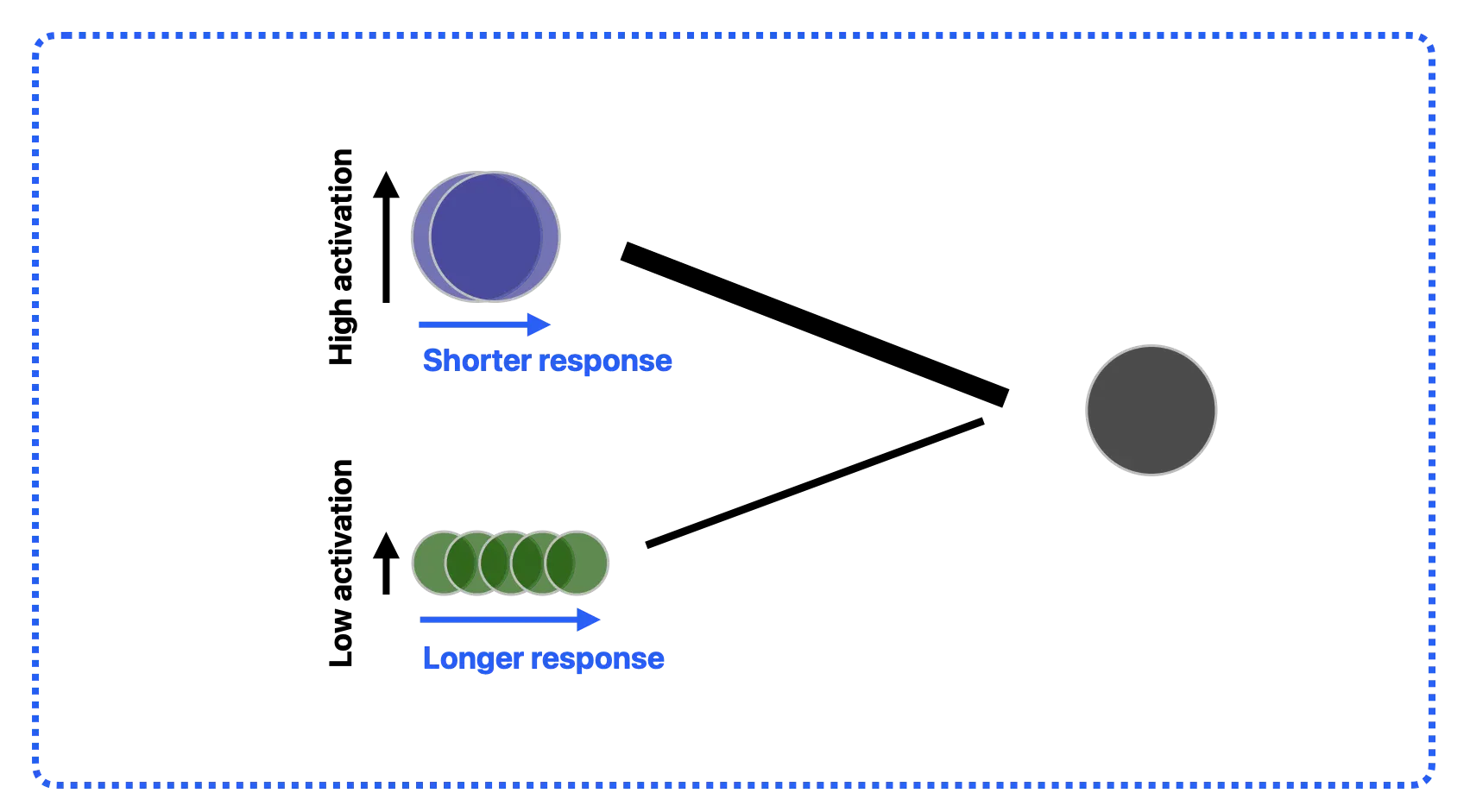
So far, we have explored how the EDL network differentiates the informativeness of various cues when predicting an outcome. These learned associative weights can then be used to calculate the activation levels of specific cues and outcomes [19]. This activation is calculated based on the associative weights for all relevant cues and outcomes that are currently present. For instance, in the multi-cue scenario depicted in the top panel of Figure 2, the activation of the unique cue is basically the cue-outcome association. The EDL approach is thus consistent with the sharpening view, where the decrease in prediction error is related to an increase in stimulus-specific activation [14]. However, directly applying the activation function of EDL cannot uniformly explain repetition suppression and enhancement; since both cue and outcome activations gradually increase as associative weights are strengthened, this would always lead to repetition enhancement.
To model both repetition enhancement and suppression, we also need to consider the facilitation view, where information accumulation leads to reduced response latency. Facilitation is usually modeled based on accumulation models. In these models, a faster accumulation rate of information (i.e., drift rate) is directly linked to shorter response latency [15]. Generally, this drift rate can be considered relating to the efficiency of perceptual integration (e.g., see [9]). The learning of simple stimulus typically exhibits higher drift rates than that of complex stimulus. Within a prediction-based framework, the stochastic drift rate can be explained in terms of a learning rate driven by prediction error (e.g., see [34]). As described above, the strengthening of associative weights in EDL can be directly used to calculate activation. Based on a parameter recovery study, van der Velde et al. [35] demonstrated that response latency in accumulation models is closely linked to these activation values. Furthermore, Brasoveanu and Dotlačil [36] showed that the latency exponent (i.e., f), which directly moderates activation, is crucial for accurately estimating latency. Additional details on the latency function, and how these equations together infer neural responses at perceptual and expectation-related areas (latency-attenuated explanation), can be found in the Supplemental Materials (Equations (S4)–(S6)).
When accounting for facilitation-based latency changes, it becomes clear that the neural response depends not only on the initial activation value, but also on the duration for which that response is sustained. In this sense, the neural response is modulated by at least two distinct variables. The first is the learning rate [35]. For example, predicting an outcome is learned more rapidly in a single-cue scenario than in a multi-cue scenario, leading to a faster attenuation of neural responses in the single-cue condition (see Figure 3). In adults, attenuation is accelerated when tasks feature longer (less demanding), rather than shorter (more demanding), inter-stimulus intervals [37,38,39]. This learning rate variable is specifically related to how fast cue-outcome associations can be acquired, and it is therefore relevant to the activation of cues and outcomes. While the learning rate can be higher when approaching a simpler task (e.g., single- vs. multi-cue; longer vs. shorter ISI), it can also be influenced by individual capacity (e.g., higher vs. lower performers).
The second variable is the rate of latency change (see [36,40]) which can be modulated by systemic states such as sleep and clinical pathology. For instance, neural responses to stimuli in infants decay more rapidly during sleep than during wakefulness (as summarized in the section “Differences in sensory activation between awake and sleep states” in Taga et al. [41]). Similarly, atypical populations, such as individuals with dyslexia, demonstrate altered, faster-decaying neural responses [38]. This type of latency change is closely related to how long an individual can sustain attention toward a presented stimulus. During wakefulness, neural responses are sustained throughout the duration of the stimulus. In contrast, during sleep or when pathology alters the processing of specific stimuli, neural responses decay quickly after stimulus onset.
It is important to note that an identical magnitude of neural responses can stem from fundamentally different mechanisms. For instance, a diminished response may reflect efficient processing in wakeful infants or healthy adults (driven by learning rate), or it may be caused by a rapid latency decay in sleeping infants or clinical groups (driven by rate of latency change; for an illustration, see [38]).
2.3. Wakeful Environmental Interaction
During wakefulness, although infants are able to sustain their attention to task-specific stimuli better compared to the sleep state, their attention can be modulated by additional factors, such as novel activities within the same environment. This is well documented in terms of explore-exploit behavior [10,42] where infants start by exploring their environment (e.g., the task-specific stimuli) and gradually disengage from the exploited activities to explore novel activities. Similarly, the preferential looking paradigm has revealed that infants initially show an attentional bias towards novel tasks, gradually disengaging as familiarity increases and shifting their focus to novel activities [43]. Further supporting this link, Snyder and Keil [44] demonstrated a correlation between infants’ behavioral shifts toward novel activities and neural changes in the task-specific gamma band, interpreting wake-state repetition suppression as neural attenuation due to the reorientation of attention to competing activities.
Such explore-exploit behavior can be modeled based on the resource availability perspective [45]. To illustrate the additional influence of novelty-seeking behavior, we continue the cue-activation examples from Figure 3. In Figure 4, the constrained time (indicated along the horizontal axis in the blue box) is defined as the cognitive resource. We have already mentioned that the unique cue (the rattle sound, represented by blue circles) acquires higher activation compared to the shared cue (the smiley, represented by green circles; see Figure 3). Thus, the response latency for the unique cue is shorter than that of the shared cue. This means that the unique cue (relative to the shared cue) allows more cognitive resources to be shared with alternative processes (brown squares). These other processes can be driven by off-task processes, such as occasional mind-wandering or overt novelty seeking, depending on the salience and learning rate of the novel activities. In this paper, the off-task occupation of cognitive resources is considered competitive to on-task processes (calculated as on-task neural responses subtracted from off-task neural responses). Consequently, a shorter response latency, compared to a longer one, results in more response attenuation due to competing off-task processing (novelty-attenuated explanation).
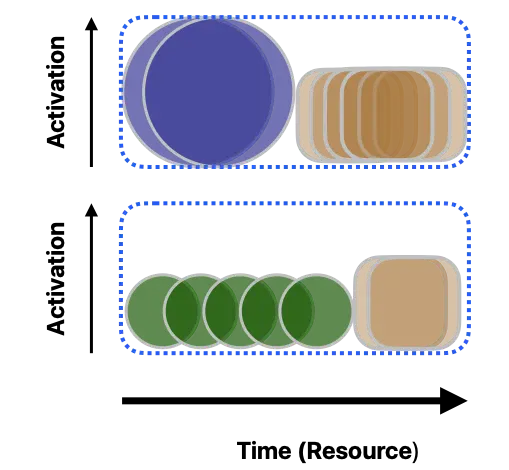
2.4. The Current Study
This study broadens the understanding of experience-dependent neural dynamics by considering (1) the interaction between bottom-up perception and top-down expectation, and (2) the interplay between on-task and off-task processing during wakefulness. The conceptual frameworks for the latency-attenuated model (Model 1) and the latency + novelty-attenuated model (Model 2) are illustrated above, with detailed equations provided in the Supplemental Materials. In the following, we present simulation studies targeting two sets of fNIRS findings that reveal differential experience-dependent neural dynamics [2,46,47] which have also been illustrated above. This simulation study seeks to model both trial-by-trial neural response changes and surprise-induced neural responses across perceptual and frontal regions.
3. Simulation 1: Single-Cue Scenario
This simulation study adapts a task from Nakano et al. [2] involving 3-month-old infants passively listening to a repeated syllable during sleep. The fNIRS recorded dynamic neural responses in the temporal and frontal areas, which are assumed in this paper to be relevant to cue perception and outcome expectation. For this simulation, the stimulus pattern is simplified as the repeated co-occurrence of a unique cue U1 and a unique outcome O1. This invariant U1-O1 pairing is presented for 100 time steps to simulate the repeated syllable presentation. This study also included a surprise condition in which the unique syllable was replaced with another syllable. In this simulation, the surprise is modeled by replacing the U1-O1 list with a novel U2-O2 co-occurrence from the 51st to the 55th time step, representing an abrupt switch to a new syllable. The presentation of the surprise stimulus follows the block design of the original study. Both repetition and surprise conditions were simulated, with each condition based on 50 individual model runs. Note also that in error-driven learning, it does not matter whether the surprise stimulus is arranged together within a block or placed sparsely as oddball signals (i.e., the same y-values would occur). This is because for the unique cue-outcome combinations (U1-O1 or U2-O2), the presenting cues (U1/2) are monotonously updated with their corresponding outcome (O1/2) without interfering with shared cues.
Model 1: Latency-Attenuated Only
The latency-attenuated Models 1a and 1b exhibited repetition enhancement and suppression, respectively, during single-cue stimulus repetition. Correspondingly, their surprise-induced responses were opposite: Model 1a produced suppression, whereas Model 1b produced enhancement effects (see Figure 5). Taken together, the simulation results of Model 1b align with Nakano et al. [2], whereas Model 1a produced the opposite pattern. These results suggest that latency attenuation in a single-cue scenario is not driven by a rapid learning rate (as illustrated by Model 1a with a default f value), but rather by an increased rate of latency changes (i.e., a higher f value of the corresponding parameter). As noted in the Introduction, a higher rate of neural attenuation in this context is thus better explained by the sleep state [41].
While the current simulation study primarily focuses on trial-by-trial fNIRS findings, it is worth noting that repetition suppression and corresponding surprise-induced enhancement have also been observed in electrophysiological studies. Although the atemporal trial-level granularity of the current model precludes the simulation of waveform results or the interpretation of event-related components, we focus primarily on components related to the well-studied early prediction-related component (N1/P1-related waveform dependent on maturation) and an additive component related to prediction error (difference waveform of MMR). In line with the imaging results of Nakano et al. [2], we do not need to reiterate the ample evidence for the gradual reduction in response amplitude to the standard stimulus and the abrupt increase in amplitude of the deviant surprise-induced response among infants (see [48]). Note that most of these studies are conducted with sleeping infants.
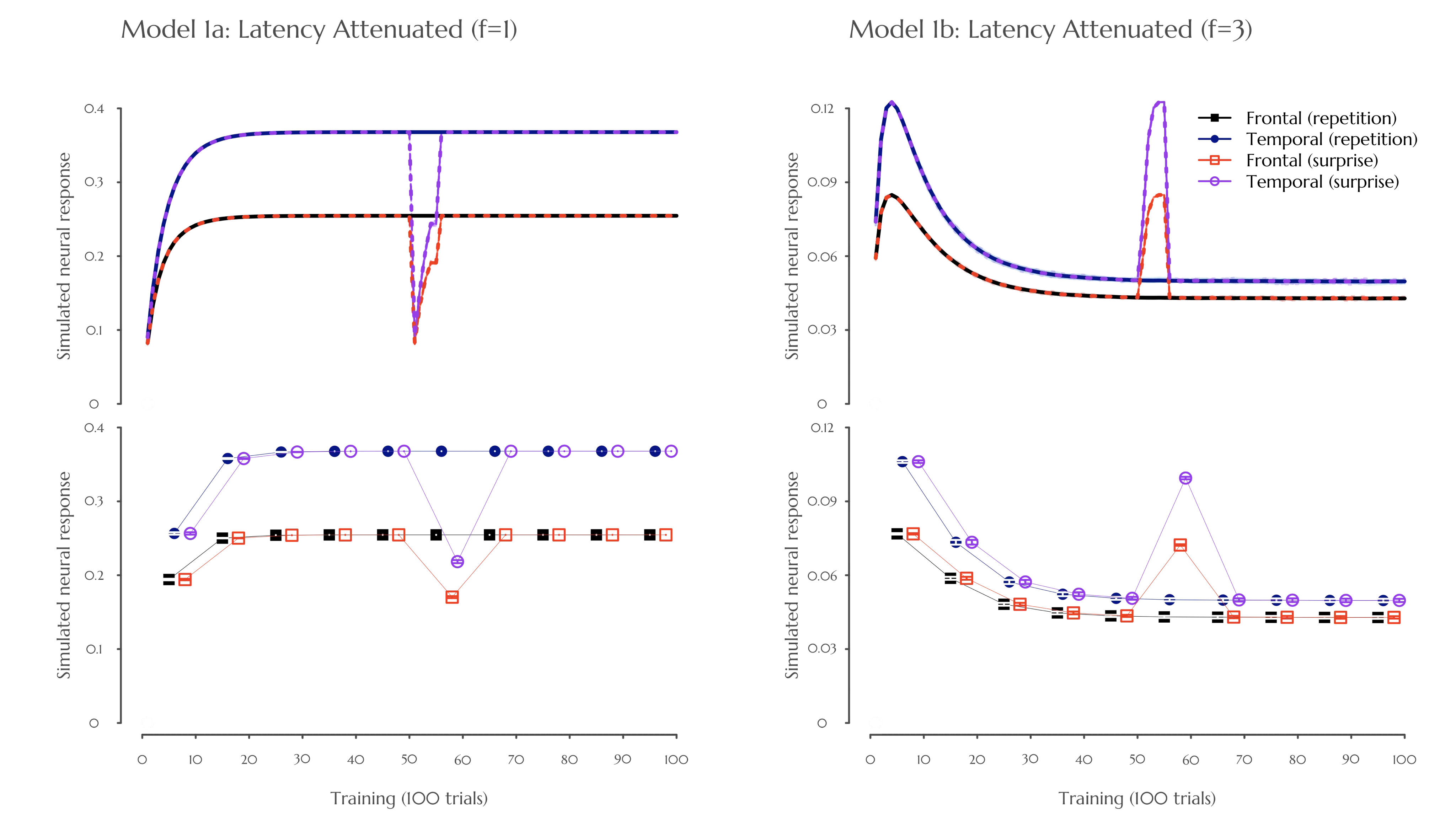
Figure 5. Simulated neural responses to the repetition of a single-cue stimulus (adapted from Nakano et al. [2]). Models 1a and 1b represent the same model with default (f = 1) and higher (f = 3) rate of latency attenuation, respectively. The top panel displays continuous changes in neural response over 100 trials. The bottom panel presents the same results binned into 10 blocks. The x-axis represents time steps (top) or blocks (bottom), and the y-axis represents the simulated neural response. In the top panel, solid lines indicate the repeated condition, whereas dashed lines represent the surprise condition. In the bottom panel, the 95% confidence intervals are indicated based on the error bars (negligible or null within the size of the points). Colors correspond to neural response regions associated with each condition: frontal lobe (black) and temporal lobe (blue) for the repetition condition, and frontal lobe (red) and temporal lobe (purple) for the surprise condition. Surprise trials occurred at time steps 51 to 55. Results are averaged across 50 independent model runs for both the repetition and surprise conditions.
4. Simulation 2: Multi-Cue Scenario
This simulation study adapts a multi-domain task from Kersey and Emberson [47] involving 6-month-old infants. In this task, wakeful infants actively discriminate between two sounds (rattle or squeak) that predict the corresponding visual movement of a red smiley (top or bottom entrance) before it exits the screen. FNIRS measures neural responses in various brain regions. In this paper, temporal responses are associated with perceiving unique auditory cues (rattle or squeak), while occipital responses are linked to processing the shared visual appearance (red smiley). Moreover, frontal lobe response is thought to reflect the perceptual integration of these auditory and visual cues, thereby forming outcome expectations about the smiley’s movement. For the simulation, the stimulus pattern is simplified as repeated pairings of unique sound cues U1/U2 with a shared smiley S corresponding to different outcomes O1/O2. These U1S-O1 and U2S-O2 pairings are randomly presented for 100 time steps to simulate the repeated multi-domain presentation. This study by Emberson et al. [46], which shared the same experiment data with Kersey and Emberson [47], also included a surprise condition. In this condition, the smiley movement was omitted. This simulation’s surprise is modeled by introducing a novel pairing U1-O3 at the 51st and 52nd time steps. Here, the shared visual cue S is absent, and the new outcome O3 represents the lack of visual movement. Both repetition and surprise conditions were simulated, with each condition based on 50 individual model runs.
Note that while the updating of cue-outcome associations during the two surprise trials relies on the co-occurrence of U1-O3, neural responses regarding S are calculated based on the expectation of prepotent outcome O1. We have already mentioned this predefinition of the S-O1 association in the Introduction (see Figure 2, bottom panel), which differs from the typical EDL equation, where such an absent association is neglected. The reason for this is that the EDL is restricted to predefined cue-outcome co-occurrences at the trial-by-trial level, without being able to account for the millisecond-level details of dynamic processing. In a processing-based model that integrates EDL [27], the unique cue U1 (rattle sound) would continue to drive the retrieval of the prepotent outcome O1 (smiley movement), which then infers the shared cue S (smiley image) from the top-down, thereby preserving the S-O1 association. Only when the expected outcome is violated (replaced by the blank screen O3) does the model recognize a change in context and begin learning the U1-O3 association.
4.1. Statistical Analyses
In Kersey and Emberson [47], a mixed-effects regression with a linear regressor revealed significant block-by-block linear trends in temporal and frontal responses. Although the linear trend was not significant for occipital responses, a follow-up t-test between blocks 1 and 5 showed a significant increase in occipital response, along with enhanced temporal and frontal responses. Furthermore, a mixed-effects regression incorporating both linear and quadratic regressors revealed significant linear and quadratic block-by-block trends for temporal, occipital, and frontal responses. The linear trend remained positive, consistent with the first regression analysis. Controlling for the linear trend, the negative coefficient on the quadratic term further confirmed an inverted U-shaped pattern, as suggested by the authors, indicating initial repetition enhancement followed by suppression.
To validate simulation results against experience-related neural dynamics (excluding surprise-induced effects), this study adapts the mixed-effects regression approach used by Kersey and Emberson [47] to analyze simulated trial-by-trial responses in the temporal, occipital, and frontal areas.
| ```latex\begin{aligned}&\text{lmerTest::lmer}( \\&\quad\quad\quad\,\,\,\,\,\;\text{scale}(\text{y}) \sim \text{poly}(\text{x}, \{\text{degree}\}) + (1 \mid \text{model run}), \\&\qquad\qquad\qquad\qquad\quad\quad\quad\quad\quad\quad\quad\text{data} = \{\text{model}\}\{\text{cortex}\} \\& \qquad\qquad\qquad\qquad\qquad\qquad\qquad\quad)\end{aligned}``` |
Following Kersey and Emberson [47], the analyses fit trial-by-trial responses to both linear and quadratic regressions. For these analyses, the same lmerTest package and identical regression structures are followed. The poly() function was applied across both linear and quadratic regressions. The only distinction between linear/quadratic was the degree: 1 for the linear regressor and 2 for the combined linear and quadratic regressors. Moreover, because the magnitudes of simulated neural responses were arbitrary, these values were standardized with the scale() function before inclusion in the regression analyses. Standardization of regressors and responses was performed separately for model-specific results in a given cortical area (e.g., Model2a-occipital), without pooling data across distinct models or cortical areas. In these analyses, the quadratic regression would further evaluate whether neural dynamics follow a U-shaped pattern, in addition to the linear regression. This is particularly useful for determining whether neural responses may exhibit initial enhancement followed by suppression, as reported by Kersey and Emberson [47]. Moreover, we can directly compare the linear and quadratic models to identify the best fit for the simulation data.
4.2. Model 2: Latency + Novelty-Attenuated
Regression analyses of Model 2a show positive linear trends (trial) in trial-by-trial neural responses and additional negative quadratic trends (trial squared) after accounting for the linear effect, across all cortical areas (see Table 1, Figure 6). Quadratic regression models outperformed linear models in all cortical areas (deviance of linear regression subtracted from quadratic regression, i.e., $${\chi }^{2}$$: temporal = 1457.6, occipital = 1605.5, frontal = 1323.8; all p < 0.001). Subsequent block-wise analyses revealed repetition enhancement followed by suppression for frontal and temporal neural responses (frontal: 5th block 95% CI = 7.25 $$×\, {10}^{-2}$$ to 7.34$$\,×\, {10}^{-2}$$, 10th block 95% CI = 7.21$$\,× \,{10}^{-2}$$ to 7.24$$\,×\,{ 10}^{-2}$$; temporal: 5th block 95% CI = 10.15 $$×\, {10}^{-2}$$ to 10.34$$\,×\,{ 10}^{-2}$$, 10th block 95% CI = 9.88$$\,×\, {10}^{-2}$$ to 9.97$$\,×\,{ 10}^{-2}$$), but no evidence of suppression for occipital responses (5th block 95% CI = 9.55$$\,×\, {10}^{-2}$$ to 9.86$$\,×\, {10}^{-2}$$, 10th block 95% CI = 9.65$$\,×\, {10}^{-2}$$ to 9.77$$\,×\,{ 10}^{-2}$$, see Figure 7). Furthermore, this attenuation occurs more rapidly for an informative unique cue (temporal) and overall expectation (frontal), as compared to a less informative shared cue (occipital). The results of Model 2a are therefore consistent with Kersey and Emberson [47]. Last but not least, the surprise-induced suppression of temporal responses, combined with the absence of changes in occipital responses (see Figure 7), is also consistent with Emberson et al. [46]. In contrast, the contribution of novelty-seeking, along with the rapid attenuation rate of Model 2b, produces results (see Table 1, Figure 6 and Figure 7) that are at odds with the empirical findings.
Table 1. Mixed-effects regressions were conducted on trial-by-trial responses simulated by Models 2a and 2b, separately for each cortical location. Statistical test results presented above the midline are based solely on a linear (trial) regressor, while those below incorporate both linear (trial) and quadratic (trial squared) regressors. All p values < 0.001.
|
Model 2a |
Model 2b |
||||
|---|---|---|---|---|---|
|
beta |
t |
beta |
t |
||
|
temporal |
x |
0.20 |
20.4 |
−0.53 |
81.4 |
|
occipital |
x |
0.32 |
35.8 |
−0.35 |
39.5 |
|
frontal |
x |
0.25 |
26.8 |
−0.56 |
91.7 |
|
temporal |
x |
0.20 |
23.7 |
−0.53 |
138.9 |
|
x2 |
−0.34 |
41.6 |
0.37 |
97.2 |
|
|
occipital |
x |
0.32 |
42.3 |
−0.35 |
48.1 |
|
x2 |
−0.33 |
44.4 |
0.35 |
49.0 |
|
|
frontal |
x |
0.25 |
30.7 |
−0.56 |
159.0 |
|
x2 |
−0.32 |
39.4 |
0.35 |
100.0 |
|
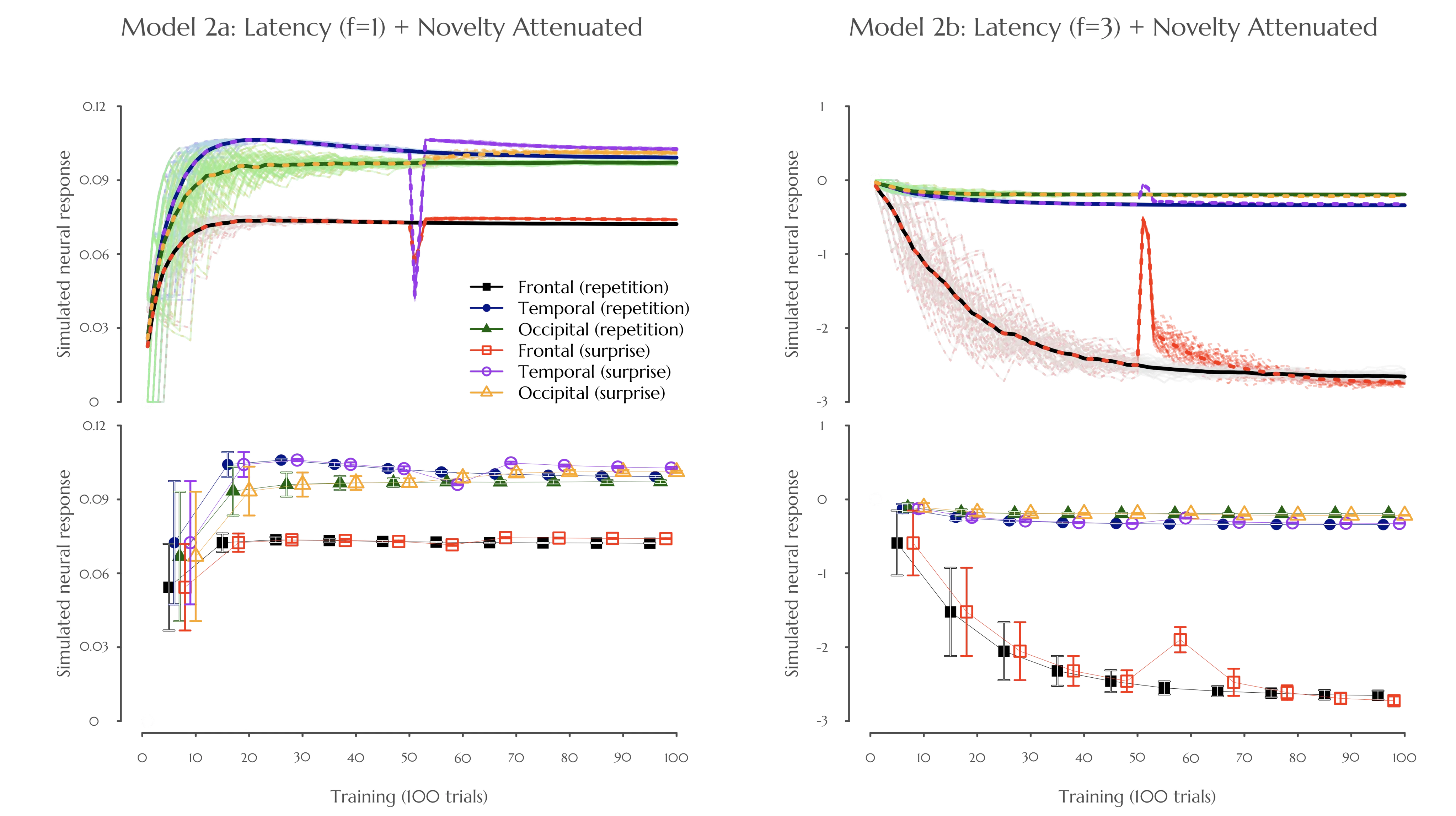
Figure 6. Simulated neural responses to the repetition of a multi-cue stimulus (adapted from [46,47]). Models 2a and 2b represent the same model with novelty attenuation, in addition to the respective default (f = 1) and higher (f = 3) latency attenuations. The top panel displays continuous changes in neural response over 100 trials. The bottom panel presents the same results binned into 10 blocks. The x-axis represents time steps (top) or blocks (bottom), and the y-axis represents the simulated neural response. In the top panel, solid lines indicate the repeated condition, whereas dashed lines represent the surprise condition. In the bottom panel, the 95% confidence intervals are indicated based on the error bars. Colors correspond to neural response regions associated with each condition: frontal lobe (black), temporal lobe (blue), and occipital lobe (green) for the repetition condition, and frontal lobe (red), temporal lobe (purple), and occipital lobe (orange) for the surprise condition. Surprise trials occurred at time steps 51 and 52. Results are averaged across 50 independent model runs for both the repetition and surprise conditions.
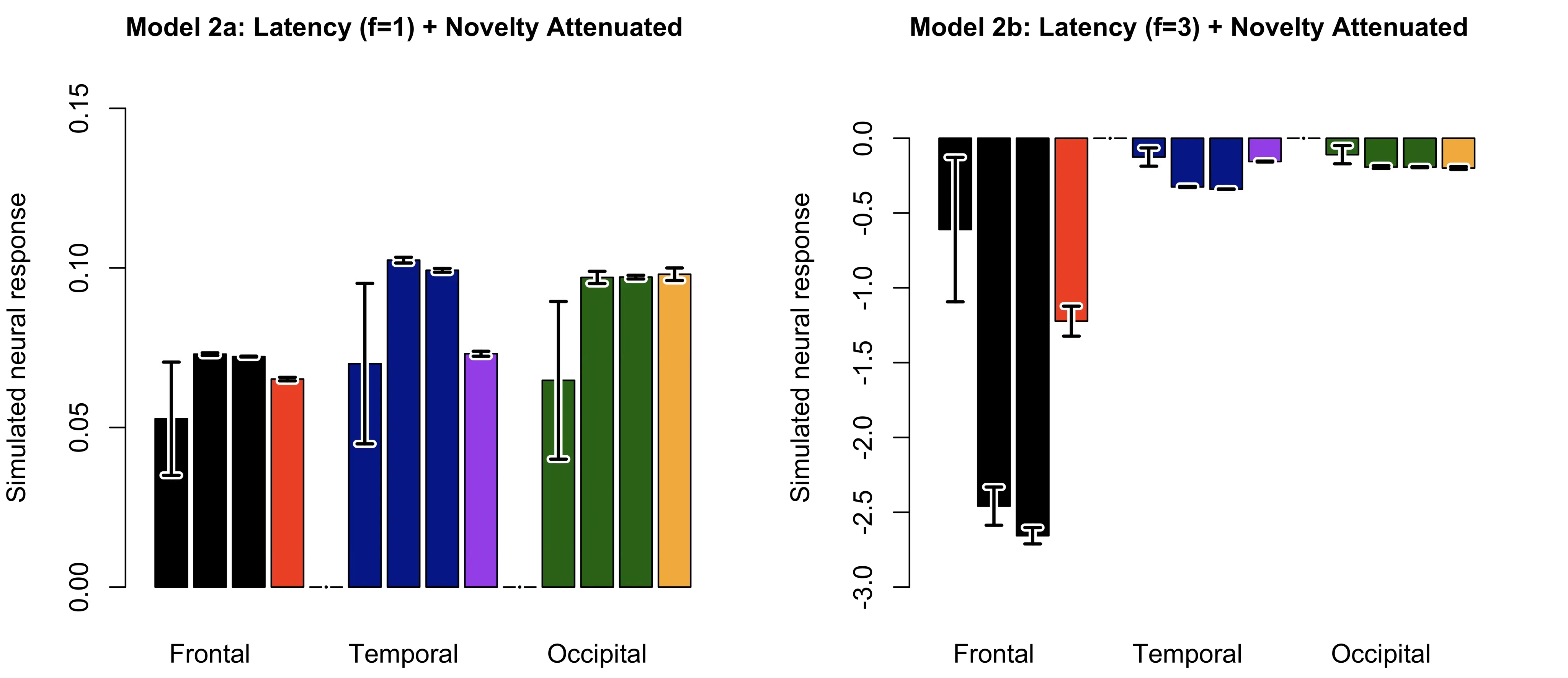
Figure 7. To better illustrate the overall experience-related neural dynamics, bar plots are provided for each model, showing neural responses at the 1st, 5th, and 10th blocks (each averaged across 10 repetitions), followed by the surprise-induced block (averaged across 2 repetitions) from left to right. The error bars indicate the 95% confidence intervals. Color coding for repeated and surprise stimulus for each cortical area is consistent with Figure 6.
5. General Discussion
This simulation study captures infants’ experience-related neural dynamics based on a model capable of dynamic, multi-domain learning through prediction-based EDL mechanisms. The model further emphasizes environmental interactions that promote ongoing exploration and exploitation, as characterized by wakeful infants. The simulation study aims more concretely to model the fNIRS findings [2,46,47] and to provide a comprehensive overview of experience-related dynamics in infants. Following streamlined discussions of the simulated results in relation to detailed empirical findings, the general discussion section will explore how the current simulation study relates to the broader topics of prediction-based learning, subjective perceptual integration, and wakeful environmental interaction.
5.1. Prediction-Based Unification of Experience-Related Dynamics
This simulation study hypothesized that a more rapid learning rate (i.e., perceptual integration efficiency) would lead to higher latency attenuation, which produces repetition suppression. As mentioned, faster perceptual integration results from the quicker occupation of a common prediction error toward predicting the current outcome expectation. Nevertheless, while the learning rate (or integration efficiency) is more rapid for the simpler single-cue versus multi-cue scenario, it alone does not lead to more rapid attenuation of neural responses from our simulation results. For instance, neural attenuation can happen if a higher rate of latency change is applied (e.g., f = 3 vs. the default f = 1; for more detail of this parameter, see [36,40]). This higher rate of latency change can be related to the faster attenuation rate when modeling a single-cue scenario during sleep (Model 1b, Figure 5), or a pathological attenuation rate when modeling wakeful multi-cue scenario processing (Model 2a, Figure 6; more discussion shortly). Alternatively, neural attenuation can originate from wakeful novelty-seeking behavior. In this case, single-cue (Model 2a, Figure S1 from Supplemental Materials) as compared to multi-cue (Model 2a, Figure 6) scenarios do lead to more rapid suppression of initial repetition enhancement. This study, therefore, explains how repetition suppression and enhancement may happen at different levels of perceptual integration efficiency [9] during wakeful processing alone, which can be relatively influenced by task complexity [6] and/or individual differences [8].
The simulation study also provides an interpretation of the contrasting surprise-induced effects. Surprise occurs when current perception no longer matches expectation. For novel stimuli, surprise leads to reduced neural response and high prediction error. When a single-cue stimulus is repeatedly presented (Model 1b, Figure 5), learning a novel stimulus from scratch elicits a larger surprise-induced enhancement compared to the attenuated repeated response, leading to an enhanced surprise-induced effect. Alternatively, in the case of repetition enhancement of a multi-cue stimulus (Models 2a, Figure 6), the suppressed surprise-induced effect was due to the learned outcome (smiley movement) being replaced by a novel outcome (smiley absence), leading to a decrease in acquired expectation. In other words, the acquired cue-outcome association (unique sound-movement inference) now undergoes an abrupt negative update when the previously unique cue (rattle sound) is paired with an alternative outcome (smiley absence) from afresh. The corresponding surprise-induced effects underlying both repetition suppression and enhancement can thus be interpreted based on the same EDL mechanism.
Taken together, the prediction-based model presented in this study extends previous work on perceptual neural dynamics (e.g., fatigue or sharpening, see [1]) by incorporating the interactive influence of bottom-up perception and top-down expectation to explain experience-related neural activity across distinct cortical areas. Moreover, perceptual integration efficiency, modeled by the learning rate, further modulates response latency. This offers an alternative to the stochastic accumulator (e.g., facilitation; see [1]) by providing an interpretive EDL mechanism (i.e., a mechanism for updating toward a reduction in the mismatch between perception and expectation). More concretely, the current simulation study is capable of simulating several fNIRS findings related to repetition suppression and enhancement, as well as corresponding surprise-induced effects, within both single- and multi-domain paradigms.
5.2. Discriminative Learning and Subjective Perceptual Integration
In terms of multi-domain integration, there is an additional distinction between the contributions of unique and shared cues, even though both cues are present during the task. As mentioned, informative, unique cues are positively updated toward multi-domain outcomes, while less informative shared cues are additionally subject to negative updates that cancel out their own associative weights. In the simulation of neural dynamics regarding the repetition of a multi-cue stimulus, the unique cue (sound: temporal area) exhibits more rapid neural enhancement compared to the shared cue (smiley: occipital area; see Model 2a, Figure 6). Note, though, that due to the arbitrary nature of response magnitudes, empirically quantifying absolute differences in neural responses across domain-specific cortical areas is challenging.
Furthermore, surprise-induced neural responses within domain-specific cortical areas elucidate the subjective contributions of unique and shared cues. As previously discussed, the simulation results are consistent with the counterintuitive finding that a present sound and absent smiley movement outcome generate a suppressed surprise-induced response for the unique cue (sound: temporal area) compared to the shared cue (smiley: occipital area), even though the smiley was now absent [46]. Anticipating the presence of the smiley’s usual movement, the perceptual response to the now physically absent smiley is preserved. In contrast, the unique cue that reliably infers the movement outcome—despite being physically present—leads to a suppressed response when paired with a novel outcome.
The above findings thus point to higher-level expectations. As revealed by empirical findings and simulated results, expectations are more subjective than what is actually presented in the environment. After prolonged exposure to a repeated multi-cue stimulus, infants infer environmental patterns, leading to greater responses to informative unique cues compared to less informative shared cues, despite both being objectively presented. When the inferred outcome expectation is challenged by a novel alternative, the perception of informative cues would demonstrate a more abrupt subjective change in neural response when the objective cue remains unchanged. These altogether demonstrate the subjective and dynamic nature of expectation formation and readaption.
5.3. Wakeful Environmental Interaction and Response Attenuation
Moreover, infants are not isolated processors but are actively situated within their environment, where their attention can freely orient toward task-related stimuli or alternative activities. To capture this, the simulation study adopts a resource availability perspective [45] to model the competition between ongoing engagement with the primary task and reorientation toward competing, novel off-task activities. Such novelty-seeking may become an increasingly crucial factor contributing to the suppression of neural responses during wakefulness [44].
By incorporating slower latency attenuation and considering novelty competition, the “wakeful” model (see Model 2a, Figure 6) simulated the multi-cue neural dynamics of repetition enhancement followed by repetition suppression [47], along with surprise-induced effects [46]. Similarly, the same model (see Model 2a, Figure S1 in Supplemental Materials) also simulated fNIRS findings of single-cue repetition suppression detected during wakefulness [49]. Taken together, these results suggest that interpreting experience-related neural dynamics, especially those observed in wakeful infants, may require considering environmental factors.
5.4. Pathological Neural Responses
Our simulation studies generally revealed that repetition suppression is related to simple task processing (such as single-cue scenarios), whereas repetition enhancement is linked to more complex task performance (such as multi-cue scenarios). However, contrary to these general trends, we also demonstrate that simulated repetition suppression may occur during multi-cue scenarios when the latency exponent is excessively large (f = 3, multi-cue scenario, Model 2b). In such cases, the attenuation of neural activity may not be driven by rapid information accumulation, but rather stems from a pathological origin, where wakeful processing collapses into an unreactive state. These simulation results align with the empirical findings of [38]. In their study, a simple frequency discrimination task was administered to both dyslexic individuals and healthy controls, with task demand manipulated by varying the length of the inter-stimulus intervals (ISIs). Crucially, their results show that the magnitude of the neural response does not directly correspond to psychopathology; for example, an identical response magnitude was observed in healthy controls performing an easier task (longer ISIs) and dyslexics performing a demanding task (shorter ISIs). In light of our model, we cannot differentiate a pathological state from a healthy one by simply looking at the magnitude of neural responses. While both groups display identical, diminished neural responses, the underlying mechanisms driving them fundamentally differ. For healthy controls, the reduction may reflect efficient perceptual integration (reflected by higher acquired activation, A; see [35]). Conversely, for dyslexic individuals, the exact same change in magnitude may be driven by a pathological, rapid overall latency attenuation (reflected by higher latency exponent, f; see [36,40]). Therefore, relying solely on the final response magnitude masks these opposing internal dynamics. Therefore, pathological neural dynamics are better characterized when mapped across varying levels of task demand, as demonstrated by Jaffe-Dax et al. [38].
5.5. Limitations
Although we are able to model both repetition suppression and enhancement based on a unified, prediction-based integrated modeling approach, several limitations of our study should be noted. Firstly, the models are not empirically fitted; therefore, the simulation findings are best viewed as a proof of concept rather than a predictive model. Moreover, while the current application of the two-layer EDL highlights the prediction-based mechanism, it does so by only looking at trial-by-trial changes without considering dynamic processing in millisecond detail. This leads to some aspects of the model (e.g., the surprising condition of a multi-cue scenario) being predefined rather than openly inferred through interaction with the task. Last but not least, our model is a high-level modeling approach that looks only at the long-range connections between cortical areas; this does not allow us to model the graded nature of neural responses happening at each of the cortical areas.
6. Conclusions
This study presents a computational model simulating key fNIRS findings related to repetition suppression and enhancement in infants. By incorporating prediction-based, error-driven learning, while considering environmental interactions, the model offers a novel perspective on distinct experience-related neural dynamics at the trial-by-trial level. The results emphasize the dynamic and subjective nature of expectation formation and its influence on perceptual responses, as well as the role of attentional orientation toward different activities in the environment of wakeful infants.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/1112, Figure S1: Simulated neural responses to the repetition of a single-cue stimulus (adapted from [2]); Figure S2: Simulated neural responses to the repetition of a multi-cue stimulus (adapted from [46,47]); Figure S3: To better illustrate the overall experience-related neural dynamics, bar plots are provided for each model, showing neural responses at the 1st, 5th, and 10th blocks (each averaged across 10 repetitions), followed by the surpriseinduced block (averaged across 2 repetitions) from left to right; Figure S4: Simulations regarding [2] where novelty attenuation is moderated; Figure S5: Simulations regarding [47] where novelty attenuation is moderated; Table S1: Mixed-effects regressions were conducted on trial-by-trial responses simulated by Models 1a and 1b, separately for each cortical location.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the author used Gemini for the sole purpose of proofreading, correcting grammar, and improving clarity. After using this tool/service, the author reviewed and edited the content as needed and takes full responsibility for the content of the published article.
Acknowledgments
I would like to express my gratitude to Niels Taatgen and Jacolien van Rij. Their modeling approaches and expertise provided the essential insights that inspired this work. I also wish to thank Lauren Emberson for her encouragement and perspective on modeling repetition-related effects regarding multimodal stimuli.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The main simulation code is publicly available at: https://osf.io/8p7kc/files/osfstorage.
Funding
This research received no external funding.
Declaration of Competing Interest
The author declares that he has no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Grill-Spector K, Henson R, Martin A. Repetition and the brain: Neural models of stimulus-specific effects. Trends Cogn. Sci. 2006, 10, 14–23. DOI:10.1016/j.tics.2005.11.006 [Google Scholar]
- Nakano T, Watanabe H, Homae F, Taga G. Prefrontal Cortical Involvement in Young Infants’ Analysis of Novelty. Cereb. Cortex 2009, 19, 455–463. DOI:10.1093/cercor/bhn096 [Google Scholar]
- Snyder KA, Garza J, Zolot L, Kresse A. Electrophysiological Signals of Familiarity and Recency in the Infant Brain. Infancy 2010, 15, 487–516. DOI:10.1111/j.1532-7078.2009.00021.x [Google Scholar]
- Bauer PJ, Wiebe SA, Carver LJ, Waters JM, Nelson CA. Developments in Long-Term Explicit Memory Late in the First Year of Life. Psychol. Sci. 2003, 14, 629–635. DOI:10.1046/j.0956-7976.2003.psci_1476.x [Google Scholar]
- Grossmann T, Gliga T, Johnson MH, Mareschal D. The Neural Basis of Perceptual Category Learning in Human Infants. J. Cogn. Neurosci. 2009, 21, 2276–2286. DOI:10.1162/jocn.2009.21188 [Google Scholar]
- Bouchon C, Nazzi T, Gervain J. Hemispheric Asymmetries in Repetition Enhancement and Suppression Effects in the Newborn Brain. PLoS ONE 2015, 10, e0140160. DOI:10.1371/journal.pone.0140160 [Google Scholar]
- Gervain J, Macagno F, Cogoi S, Peña M, Mehler J. The neonate brain detects speech structure. Proc. Natl. Acad. Sci. USA 2008, 105, 14222–14227. DOI:10.1073/pnas.0806530105 [Google Scholar]
- Torkildsen JVK, Friis Hansen H, Svangstu JM, Smith L, Simonsen HG, Moen I, et al. Brain dynamics of word familiarization in 20-month-olds: Effects of productive vocabulary size. Brain Lang. 2009, 108, 73–88. DOI:10.1016/j.bandl.2008.09.005 [Google Scholar]
- Dehaene-Lambertz G, Hertz-Pannier L, Dubois J, Mériaux S, Roche A, Sigman M, et al. Functional organization of perisylvian activation during presentation of sentences in preverbal infants. Proc. Natl. Acad. Sci. USA 2006, 103, 14240–14245. DOI:10.1073/pnas.0606302103 [Google Scholar]
- Nordt M, Hoehl S, Weigelt S. The use of repetition suppression paradigms in developmental cognitive neuroscience. Cortex 2016, 80, 61–75. DOI:10.1016/j.cortex.2016.04.002 [Google Scholar]
- Alink A, Abdulrahman H, Henson RN. Forward models demonstrate that repetition suppression is best modelled by local neural scaling. Nat. Commun. 2018, 9, 3854. DOI:10.1038/s41467-018-05957-0 [Google Scholar]
- Weiner KS, Sayres R, Vinberg J, Grill-Spector K. fMRI-Adaptation and Category Selectivity in Human Ventral Temporal Cortex: Regional Differences Across Time Scales. J. Neurophysiol. 2010, 103, 3349–3365. DOI:10.1152/jn.01108.2009 [Google Scholar]
- Vogels R. Sources of adaptation of inferior temporal cortical responses. Cortex 2016, 80, 185–195. DOI:10.1016/j.cortex.2015.08.024 [Google Scholar]
- Kok P, Jehee JFM, de Lange FP. Less Is More: Expectation Sharpens Representations in the Primary Visual Cortex. Neuron 2012, 75, 265–270. DOI:10.1016/j.neuron.2012.04.034 [Google Scholar]
- Ratcliff R, Smith PL. A Comparison of Sequential Sampling Models for Two-Choice Reaction Time. Psychol. Rev. 2004, 111, 333–367. DOI:10.1037/0033-295x.111.2.333 [Google Scholar]
- Friston K. A theory of cortical responses. Phil. Trans. R. Soc. B 2005, 360, 815–836. DOI:10.1098/rstb.2005.1622 [Google Scholar]
- Rescorla RA, Wagner AR. A theory of pavlovian conditioning: Variations in the effectiveness of reinforcement and non-reinforcement. Class. Cond. Curr. Res. Theory 1972, 2, 64–69. [Google Scholar]
- Miller RR, Barnet RC, Grahame NJ. Assessment of the Rescorla-Wagner model. Psychol. Bull. 1995, 117, 363–386. DOI:10.1037//0033-2909.117.3.363 [Google Scholar]
- Hoppe DB, Hendriks P, Ramscar M, van Rij J. An exploration of error-driven learning in simple two-layer networks from a discriminative learning perspective. Behav. Res. 2022, 54, 2221–2251. DOI:10.3758/s13428-021-01711-5 [Google Scholar]
- Dehaene S. How We Learn: The New Science of Education and the Brain; Penguin: London, UK, 2021. [Google Scholar]
- Kachergis G, Frank MC. A large-scale comparison of cross-situational word learning models. PsyArXiv 2021, preprints. DOI:10.31234/osf.io/4um9k [Google Scholar]
- Spratling MW. A review of predictive coding algorithms. Brain Cogn. 2017, 112, 92–97. DOI:10.1016/j.bandc.2015.11.003 [Google Scholar]
- Font-Alaminos M, Ribas-Prats T, Gorina-Careta N, Escera C. Emergence of prediction error along the human auditory hierarchy. Hear. Res. 2021, 399, 107954. DOI:10.1016/j.heares.2020.107954 [Google Scholar]
- Parras GG, Nieto-Diego J, Carbajal GV, Valdés-Baizabal C, Escera C, Malmierca MS. Neurons along the auditory pathway exhibit a hierarchical organization of prediction error. Nat. Commun. 2017, 8, 2148. DOI:10.1038/s41467-017-02038-6 [Google Scholar]
- Rao RPN, Ballard DH. Predictive coding in the visual cortex: A functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 1999, 2, 79–87. DOI:10.1038/4580 [Google Scholar]
- Recasens M, Leung S, Grimm S, Nowak R, Escera C. Repetition suppression and repetition enhancement underlie auditory memory-trace formation in the human brain: An MEG study. NeuroImage 2015, 108, 75–86. DOI:10.1016/j.neuroimage.2014.12.031 [Google Scholar]
- Ji Y, van Rij J, Taatgen N. Skill acquisition from a bottom-up perspective. In Proceedings of the 23rd International Conference on Cognitive Modelling; Sibert C, Hough AR, Eds.; Applied Cognitive Science Lab: University Park, PA, USA, 2025; pp. 82–88. [Google Scholar]
- Parent A, Hazrati LN. Functional anatomy of the basal ganglia. I. The cortico-basal ganglia-thalamo-cortical loop. Brain Res. Rev. 1995, 20, 91–127. DOI:10.1016/0165-0173(94)00007-c [Google Scholar]
- Helie S, Chakravarthy S, Moustafa AA. Exploring the cognitive and motor functions of the basal ganglia: An integrative review of computational cognitive neuroscience models. Front. Comput. Neurosci. 2013, 7, 174. DOI:10.3389/fncom.2013.00174 [Google Scholar]
- Kotseruba I, Tsotsos JK. 40 years of cognitive architectures: Core cognitive abilities and practical applications. Artif. Intell. Rev. 2020, 53, 17–94. DOI:10.1007/s10462-018-9646-y [Google Scholar]
- Hake HS, Sibert C, Stocco A. Inferring a Cognitive Architecture from Multitask Neuroimaging Data: A Data‐Driven Test of the Common Model of Cognition Using Granger Causality. Top. Cogn. Sci. 2022, 14, 845–859. DOI:10.1111/tops.12623 [Google Scholar]
- Stocco A, Sibert C, Steine-Hanson Z, Koh N, Laird JE, Lebiere CJ, et al. Analysis of the human connectome data supports the notion of a “Common Model of Cognition” for human and human-like intelligence across domains. NeuroImage 2021, 235, 118035. DOI:10.1016/j.neuroimage.2021.118035 [Google Scholar]
- Emberson LL, Cannon G, Palmeri H, Richards JE, Aslin RN. Using fNIRS to examine occipital and temporal responses to stimulus repetition in young infants: Evidence of selective frontal cortex involvement. Dev. Cogn. Neurosci. 2017, 23, 26–38. DOI:10.1016/j.dcn.2016.11.002 [Google Scholar]
- Luzardo A, Alonso E, Mondragón E. A Rescorla-Wagner drift-diffusion model of conditioning and timing. PLoS Comput. Biol. 2017, 13, e1005796. DOI:10.1371/journal.pcbi.1005796 [Google Scholar]
- van der Velde M, Sense F, Borst JP, van Maanen L, van Rijn H. Capturing Dynamic Performance in a Cognitive Model: Estimating ACT‐R Memory Parameters With the Linear Ballistic Accumulator. Top. Cogn. Sci. 2022, 14, 889–903. DOI:10.1111/tops.12614 [Google Scholar]
- Brasoveanu A, Dotlačil J. An extensible framework for mechanistic processing models: From representational syntax-semantics theories to quantitative model comparison. In Proceedings of the 16th International Conference on Cognitive Modeling (ICCM 2018), Madison, WI, USA, 21–24 July 2018. [Google Scholar]
- Heinemann LV, Kaiser J, Altmann CF. Auditory repetition enhancement at short interstimulus intervals for frequency-modulated tones. Brain Res. 2011, 1411, 65–75. DOI:10.1016/j.brainres.2011.07.009 [Google Scholar]
- Jaffe-Dax S, Frenkel O, Ahissar M. Dyslexics’ faster decay of implicit memory for sounds and words is manifested in their shorter neural adaptation. eLife 2017, 6, e20557. DOI:10.7554/elife.20557 [Google Scholar]
- Wang AL, Mouraux A, Liang M, Iannetti GD. The Enhancement of the N1 Wave Elicited by Sensory Stimuli Presented at Very Short Inter-Stimulus Intervals Is a General Feature across Sensory Systems. PLoS ONE 2008, 3, e3929. DOI:10.1371/journal.pone.0003929 [Google Scholar]
- Brasoveanu A, Dotlačil J. Computational Cognitive Modeling and Linguistic Theory; Springer: Berlin/Heidelberg, Germany, 2020. DOI:10.1007/978-3-030-31572-6 [Google Scholar]
- Taga G, Watanabe H, Homae F. Developmental changes in cortical sensory processing during wakefulness and sleep. NeuroImage 2018, 178, 519–530. DOI:10.1016/j.neuroimage.2018.05.075 [Google Scholar]
- Chun MM. Babies and Brains: Habituation in Infant Cognition and Functional Neuroimaging. Front. Hum. Neurosci. 2008, 2, 333. DOI:10.3389/neuro.09.016.2008 [Google Scholar]
- Hunter MA, Ames EW, Koopman R. Effects of stimulus complexity and familiarization time on infant preferences for novel and familiar stimuli. Dev. Psychol. 1983, 19, 338–352. DOI:10.1037/0012-1649.19.3.338 [Google Scholar]
- Snyder KA, Keil A. Repetition Suppression of Induced Gamma Activity Predicts Enhanced Orienting toward a Novel Stimulus in 6-month-old Infants. J. Cogn. Neurosci. 2008, 20, 2137–2152. DOI:10.1162/jocn.2008.20149 [Google Scholar]
- Taatgen NA, van Vugt MK, Daamen J, Katidioti I, Huijser S, Borst JP. The resource-availability model of distraction and mind-wandering. Cogn. Syst. Res. 2021, 68, 84–104. DOI:10.1016/j.cogsys.2021.03.001 [Google Scholar]
- Emberson LL, Richards JE, Aslin RN. Top-down modulation in the infant brain: Learning-induced expectations rapidly affect the sensory cortex at 6 months. Proc. Natl. Acad. Sci. USA 2015, 112, 9585–9590. DOI:10.1073/pnas.1510343112 [Google Scholar]
- Kersey AJ, Emberson LL. Tracing trajectories of audio‐visual learning in the infant brain. Dev. Sci. 2017, 20, e12480. DOI:10.1111/desc.12480 [Google Scholar]
- Näätänen R, Kujala T, Light G. The Mismatch Negativity: A Window to the Brain; Oxford University Press: Oxford, UK, 2019. DOI:10.1093/oso/9780198705079.001.0001 [Google Scholar]
- Emberson LL, Boldin AM, Robertson CE, Cannon G, Aslin RN. Expectation affects neural repetition suppression in infancy. Dev. Cogn. Neurosci. 2019, 37, 100597. DOI:10.1016/j.dcn.2018.11.001 [Google Scholar]