Ethnogenesis Reconstruction in the Population of South Siberia and Tuva Using Data on N-M178 and O-M175 Polymorphisms

Ethnogenesis Reconstruction in the Population of South Siberia and Tuva Using Data on N-M178 and O-M175 Polymorphisms
Dmitry Adamov
1,*
 Georgy Ponomarev
1
Larissa Damba
1,2
Elvira Pocheshkhova
1,3
Elena Balanovska
1
Georgy Ponomarev
1
Larissa Damba
1,2
Elvira Pocheshkhova
1,3
Elena Balanovska
1
Received: 25 March 2026 Revised: 20 April 2026 Accepted: 29 May 2026 Published: 18 June 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Introduction
A bustling crossroads of ancient migration routes, South Siberia was successively a part of the Xiongnu Empire, the Göktürk, Uyhgur and Yenisei Kyrgyz Khaganates, the Genghisid Mongol Empire, and the Manchurian Qing Empire. Among the ethnic groups traditionally recognized as indigenous to South Siberia are Tofalars (the Irkutsk region), Tuvans and Tozhu Tuvans (Tuva), Shors (Mountainous Shoria), Khakass (Khakassia), Northern Altaians, including Kumandins, Tubalars, and Chelkans (Altai), Southern Altaians, including Altai-Kizhi, Telengits, and Teleuts (Altai). All of them are Turkic-speaking. The most populous group is the Tuvans (295,000), followed by Altaians (78,000), Khakass (61,000), Shors (11,000), and Tofalars (700).
Studies of ancient DNA, particularly those focusing on the Eurasian steppe, the Mongolian Plateau, and Siberia, have fundamentally transformed our understanding of population dynamics during the Bronze Age and the Early Iron Age. During the Bronze Age, large-scale population migrations, driven by the steppe pastoralists of the Yamnaya culture, spread as far as Siberia and Mongolia, bringing with them advanced livestock-rearing technologies and Indo-European languages. In the Late Bronze Age, populations associated with the Sintashta culture established the Andronovo cultural complex east of the Urals. These processes significantly increased the presence of western steppe genetic components as far as Lake Baikal and eastern Mongolia. In the Early Iron Age, Scythian-Saka populations dominated the Eurasian steppe. Later, the first nomadic Xiongnu empire was multi-ethnic and exhibited high genetic diversity. From this time onward, the main migratory flows of nomads occurred from east to west, accompanied by a gradual increase in the proportion of East Eurasian ancestry across the Eurasian steppe and adjacent territories [1].
Published studies of ancient DNA from the Tuva region mainly pertain to the Scythian-Siberian period (first millennium BCE) [2,3,4,5,6,7,8]. This is because the earliest archaeological artifacts of the Scythian world were found precisely in the territory of Tuva. Whole-genome studies were conducted on individual samples from elite burial complexes [2,7,8]. A significantly larger number of ancient samples were studied using Y-STR markers [3,4,5,6]. The genetic diversity of the Y-chromosome in the ancient population during this period was low, dominated by carriers of haplogroups R1a and Q [9].
Research into the distribution of Y-SNP and Y-STR markers in indigenous South Siberian populations has made it possible to describe the genetic diversity of the region [10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45] and reconstruct its genetic history. Of course, uniparental markers, such as Y-SNPs and Y-STRs, reflect only the paternal lineage. Ethnogenesis itself, however, is a multifactorial process influenced by culture, language, and social structure.
Peak frequencies (~60%) of haplogroup R1a-M198 are observed in Shors and Southern Altaians. Haplogroup Q-М242 is very common in Kets (~94%), Selkups (~66%), and Chelkans (~50%), which might suggest a Proto-Uralic and Paleo-Asian contribution to the ethnogenesis of South Siberia. Collectively, haplogroups N-M178 and N-P43 occur in about one-third of the Shor population, with N-M178 constituting a substantial 25%. By contrast, N-M178 is very rare (~1%) in the neighboring population of Northern Altaians, where N-P43 carriers make up about one-fifth of the gene pool. Haplogroup R1b-М73 is found only in Shors (~10%) and Northern Altaians (~15%); however, within the Northern Altaian population, it occurs in as many as ~50% of Kumandins. С-М217 branches have peak frequencies in Southern Altaians (~13%), possibly indicating a genetic influence of the Mongol expansion across the steppe, south of the Altai Mountains.
The most populous indigenous South Siberian ethnic group is the Tuvans, who live in close settlements, chiefly in the Republic of Tuva (~290,000 individuals), but also in Mongolia and China (~6000 Tuvans in total). Although their historical habitat suffered a division into military and administrative units at the time of the Qing Empire, their clan structure exists to this day. As shown by previous studies, a clan-based social organization effectively reflects the genetic structure of populations in various regions of North Eurasia (Central Asia, Ural, Siberia) and, considering its patrilineal succession, can be most accurately described with Y-chromosome markers [30,32,34,35,36,45]. In passports that the state began to issue for Tuvans after Tuva became a part of the USSR in 1944, their first names and clan names were transposed: clan names were treated as first names, while first names were treated as last names. Even so, it is still possible to reconstruct clan affiliations in the Tuvan population using genealogy analysis and evidence from the informants [45]. The main components of the Tuvan Y-gene pool are branches of the North Eurasian macrohaplogroup N-F963, the Paleo-Siberian macrohaplogroup Q-L56, and the Central Asian macrohaplogroup С-М217 [45]. In the Tuvan population, the most common haplogroups are N-P43 (22%), Q-M242 (21%), R1a-Z93 (15%), N-M178 (15%), C-M217(xM48, M407) (6%), C-M48 (6%), C-M407 (4%), and O-M175 (5%) [9].
As shown in previous publications, N-P43 lineages represent a Samoyedic contribution in Altaian, Nenets, Siberian Tatar, Khakass, Khanty, Chulym, and Shor populations [38,44], while the presence of haplogroup Q-M242 suggests a Yeniseian substrate [39,46]. The analysis of autosomal genomes [47] reveals a connection between Yeniseian speakers and populations of the Cis-Baikal Late Neolithic—Bronze Age (5100–3700 YBP). Branches of the C-M217 macrohaplogroup, which collectively make up 16% of the Tuvan gene pool, are descended from Central Asian steppe populations [41], testifying to their late Medieval migrations to West Mongolia and Tuva [41]. The possibility of R1a-Z93 inheritance from a Scythian-Siberian population was researched by [9]; the article also shows that one-third of ancient Tuvan Scythians were carriers of various Q lineages, of which only Q-YP1102 persists to this day.
Very little data is available on haplogroups N-M178 and O-M175 that collectively constitute ~20% of the Tuvan Y-chromosome gene pool. In the context of the Tuvan population, the history of O-M175 has not been studied at all, while N-M178, together with haplogroup N-P43, has been analyzed only as part of the North-Eurasian haplogroup N-F963 in South Siberian populations [45]. According to a hypothesis proposed in [42], the sources of two major N-M178 branches in the Tuvan population are different. N-B187(хB499) might be descended from an autochthonous population, but its connection to Samoyedic, Ugric, and other local ethnic groups is controversial, whereas N-B219(xB199) might be associated with the dispersal of Mongolians throughout Tuva.
This study aims at elucidating significant aspects of the ethnogenesis of indigenous populations inhabiting South Siberia and, more specifically, Tuva, using information about N-M178 and O-M175 polymorphisms.
2. Materials and Methods
Genealogical information about the indigenous populations of Tuva and their biological samples were collected during 2009–2023 field work supervised by Prof. Elena Balanovska. The locations of the collected samples on the map of Tuva are shown in Supplementary Figure S1. Written informed consent was obtained from every participant, and the collected samples were anonymized. The study was approved by the Ethics Committee of the Research Centre for Medical Genetics (Protocol No. 1 dated 29 June 2020). The dataset comprised a total of 2180 samples from the Biobank of North Eurasia [48] collected from unrelated male individuals whose ethnic affiliation was confirmed to a depth of three generations. N-M178 phylogeny was analyzed from 113 samples: Tuvan (n = 86), Tofalar (n = 13), Siberian Tatar (n = 3), Khakass (n = 1), and Mongol (n = 10). Only Tuvan samples (n = 24) were used to analyze the O-M175 phylogeny because this haplogroup is rare in other South Siberian populations.
DNA was extracted by phenol-chloroform extraction. For branch identification, N-M178 samples were genotyped for a broad set of SNPs using TaqMan custom array plates and a QuantStudio 12K Flex amplification system (Thermo Fisher Scientific, Waltham, MA, USA). The samples selected for O-M175 analysis were genotyped for the most common SNPs, including M119, P31, and M122. Some O-M175 branches were predicted from published Y-STR haplotypes. The results were compared to the modal haplotypes on the world Y-chromosome tree [49].
All samples were genotyped for 37 Y-STR markers using a Nanofor-05 genetic analyzer (Syntol LLC, Moscow, Russia): DYS389I, DYS389II, DYS390, DYS456, DYS19, DYS385a, DYS385b, DYS458, DYS437, DYS438, DYS448, GATA-H4, DYS391, DYS392, DYS393, DYS439, DYS635, DYS533, DYS504, DYS481, DYS525, DYS576, GATA-A10, DYS449, DYS445, DYS552, DYS537, DYS505, DYS460, DYF387S1a, DYF387S1b, DYS570, GGAAT1B07, DYS549, DYS643, DYS627, DYS518.
Phylogenetic networks were constructed using a median-joining algorithm [50] in Network v.10.2.0.0. The networks were visualized in Network Publisher v.2.1.2.5. The weight of each of 37 Y-STR markers was assumed to be 10, at ε = 0. Time to the most recent common ancestor (TMRCA) was estimated using the ASD method [51]. The average mutation rate for the 37-Y-STR haplotypes was assumed to be 0.0039 per locus per generation [52,53]. The average generation time was assumed to be 31.5 years [54].
Y-chromosome branches of the ancient samples from the literature were compared against the YFull database [49], haplotree.info (https://docs.google.com/spreadsheets/d/1xfeK8HvVjkCY7mKj3WEKAjap
AqltooWJMptY0nStKbo/edit?gid=1942507897#gid=1942507897, accessed on 26 December 2025), FamilyTreeDNA Discover™ (https://discover.familytreedna.com/, accessed on 13 January 2026) and TheYtree (https:// www.theytree.com/tree/, accessed on 13 January 2026).
3. Results
The phylogenetic networks in Figure 1 and Figure 2 show well-defined, distinct clusters of Y-STR haplotypes; the clusters appear well discriminated due to the high resolution of the applied 37-Y-STR marker panel. The clusters fully correspond to the branches of haplogroups N-M178 and O-M175 identified on the basis of their Y-SNP markers.
3.1. Haplogroup N-M178
The phylogenetic network for haplogroup N-M178 (Figure 1; 113 Tuvan, Tofalar, Siberian Tatar, Khakass, Mongol samples) shows 6 clusters of Y-STR haplotypes.
The most distant ethno-specific N-Y24317(xB499) cluster (~900 YBP, n = 32) is formed by Western and Central Tuvans (the Oorzhak, Kuzhuget, Dongak, Kuular, Mongush, and Ondar clans, Figure 1). According to [49], N-Y24317 branched off the N-M178 trunk at ~10,700 YBP and is marked by the B497 mutation [31].
The rest of the Tuvan samples are located on N-F4205 branches. The cluster of N-Y16319(xY16223) (~680 YBP, n = 23) haplotypes comprises Tofalars of the Irkutsk region (n = 12), Tozhu-Tuvans of Northwestern Tuva (n = 7, the Baraan, Choodu, Ak, Kol, and Maady clans), and Tuvans from the South and South-East of Tuva (n = 4, the Kyrgys, Todut, and Khertek clans). Unfortunately, Y-SNPs specific to N-Y16319(xY16223) remain unknown in the absence of sequenced whole genomes of its carriers, but on the YFull tree, two SNPs Y16319 and Y16223, appear on the same hierarchical level as F4205 [49]. This indicates that N-Y16319(xY16223) diverged earlier than other N-F4205 branches.
Another ethno-specific branch is N-Y16223(xZ35328, F2288), with 11 tandem repeats in the DYS385b locus (~660 YBP). It is common in Tuvans (n = 27) from Central (the Mongush, Khaazut, Khertek, Khovalyg clans, n = 12), Western (the Dongak, Mongush, Oorzhak, Irgit, Kuular clans, n = 8), and other parts of Tuva (the Kyrgys, Ak, Baraan, Irgit, Kol, Shynaa clans, n = 7). It also sometimes occurs in other ethnic groups: Tofalars (n = 1), Khakass (n = 1), Siberian Tatars (n = 3), and Mongolians (n = 4) from Western Mongolia (the Uriankhai and Miangat clans).
Other N-Z35328 and N-F2288 haplotypes in the phylogenetic network represent branches that are widespread in Turkic- and Mongolian-speaking populations and a few unidentified branches that we named N-Y16223(xZ35328, F2288) and DYS385 = 11,13 (in most branches, there are 10 tandem repeats in the DYS391 locus). N-Z35328 mostly occurs in Mongolian-speaking populations, reaching 82% in Trans-Baikal Khori Buryats. N-F2288 is a more “western” haplogroup: it has higher frequencies in Turkic-speaking peoples. All Turkic-Mongolian branches are represented in the Tuvan population, occurring in 15 carriers from the western and central parts of Tuva (the Dongak, Oorzhak, Sat, Tulush, Dolaan, Mongush, Saaya, Khovalyg clans) and in one Tozhu-Tuvan from the Ak clan.
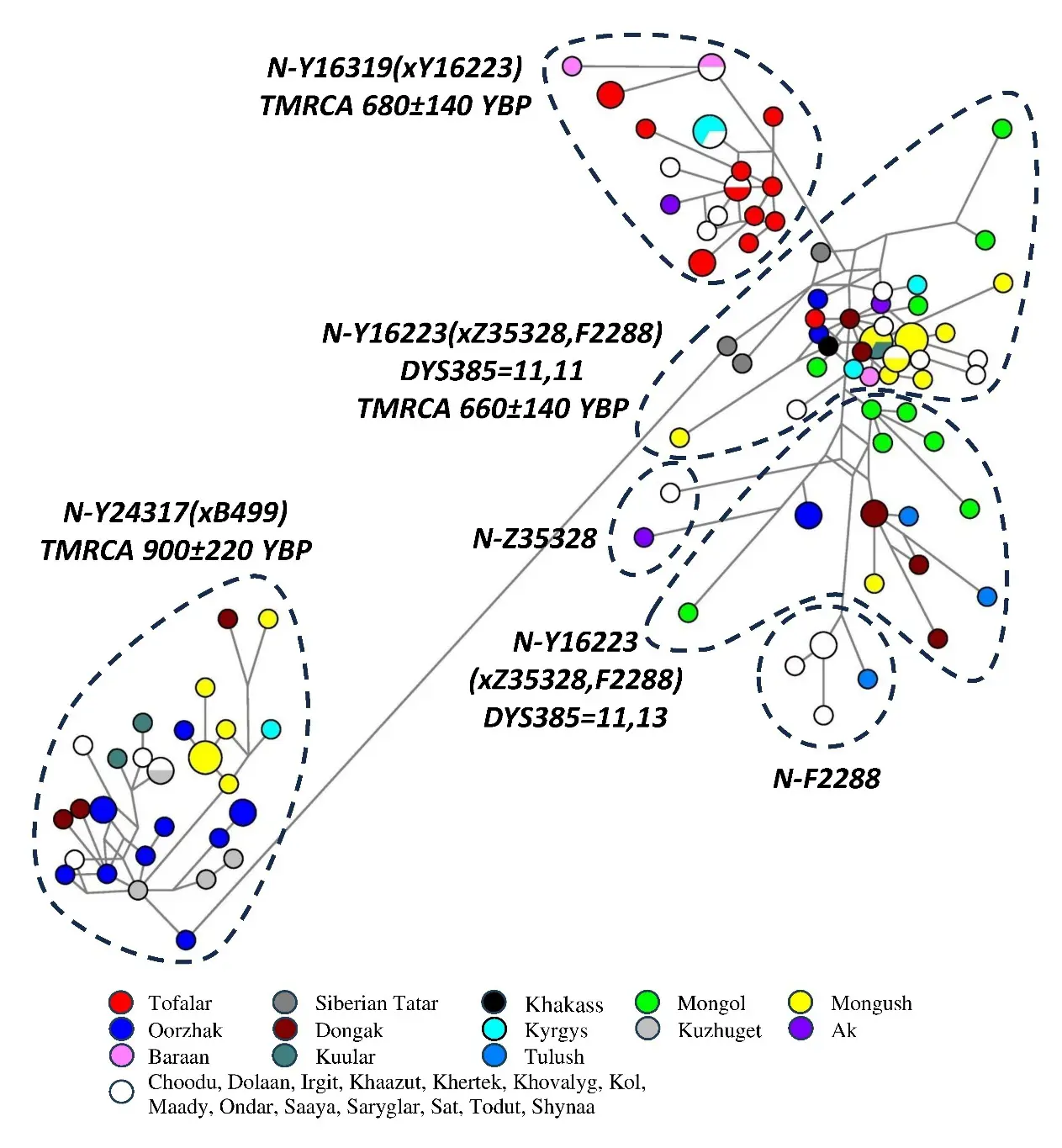
Figure 1. The median network of Y-STR haplotypes for haplogroup N-M178 (n = 113). Legend: Colored circles represent populations and clans.
Summing up, N-M178 branches can be mostly found in the western and central parts of Tuva, representing the Mongush, Oorzhak, Dongak, Kuzhuget, Kuular, Tulush, and some other clans (Table 1). The only exception is N-Y16319(xY16223), which occurs in the district of Tozhu, in the South-East and South of Tuva (the Baraan, Choodu, Ak, Kol, Maady, Kyrgys, Todut, Khertek clans) and in the Tofalars of the Irkutsk region.
Table 1. The frequency and distribution of N-M178 and O-M175 branches among the Tuvans.
|
Branches |
Frequency, % |
Distribution Features |
|---|---|---|
|
N-Y24317(xB499) |
5.6 |
Tuvan ethno-specific lineage. Western and central parts of Tuva. |
|
N-Y16319(xY16223) |
1.9 |
Tuvan and Tofalar ethno-specific lineage. Eastern and southern parts of Tuva. |
|
N-Y16223(xZ35328, F2288) DYS385 = 11,11 |
4.7 |
Tuvan ethno-specific lineage. Western and central parts of Tuva. |
|
N-Y16223(xZ35328, F2288) DYS385 = 11,13 |
1.6 |
Rare Turkic-Mongolian lineages. Western and central parts of Tuva. |
|
N-F2288 |
0.9 |
Turkic-Mongolian lineage. Rare in Tuva. |
|
N-Z35328 |
0.3 |
Predominantly Mongolian lineage. Rare in Tuva. |
|
O-M122-M134-M1706 |
1.9 |
One of the most common Han Chinese Y-lineages (Oα). |
|
O-M122-M134-FGC85750 |
1.0 |
One of the most common Han Chinese Y-lineages (Oβ). |
|
O-M119-F492 |
1.9 |
Widespread among the holders of the Chinese last name Ye. |
|
O-P31-Y24057 |
0.2 |
Common among Chinese, Japanese, Koreans, Southeast Asians. Rare in Tuva. |
3.2. Haplogroup O-M175
The branches O-M119, O-P31, and O2-M122 of haplogroup О-М175 also appear well discriminated in the phylogenetic network (Figure 2) constructed from 24 Tuvan samples on the basis of 37 Y-STR marker haplotypes.
Branch prediction from Y-STR haplotypes places all of our O-M122 samples on two large branches of subhaplogroup O-M134: O-M134-M1706 (~7000 YBP) and O-M134-FGC85750 (~8400 YBP) [49]. Their haplotype clusters in the phylogenetic network correspond to two of the three most common Han Chinese Y-lineages that originated in the Chinese Neolithic and are designated as Oα and Oβ [55].
Importantly, the haplotype clusters representing these two oldest and largest O-M122 branches are not dominated by any Tuvan clan: of 14 samples, the Kuzhuget and Kyrgys clans are represented by two individuals, and the rest 10 clans (Choodu, Khertek, Kol, Mongush, Ondar, Oorzhak, Oyun, Sartyyl, Saryglar, Shalyk) are represented by only one individual each. At the same time, O-M122 distribution has a clear geographical pattern: six samples come from the central part of Tuva, and another five from its southern and southeast regions.
By contrast, the compact O-M119-F492 cluster is undoubtedly young (~300 YBP), with a pronounced founder effect (Table 2, Figure 2). All carriers of haplotypes encompassed in this cluster reside in the southeast of Tuva: five samples represent the Kyrgys clan, and another four the Choodu, Irgit, Khertek, and Soyan clans. According to the literature, haplogroup O-M119 is one of the major lineages in southeastern Chinese nationalities [56], and its branch O-F492 is common in the holders of the Chinese last name Ye [57]. In our dataset, haplogroup O-P31 is represented by a single Tuvan sample from the southeast of Tuva (its clan affiliation is unknown).
Table 2. TMRCA of Tuvan Y-chromosome branches (YBP) estimated from Y-STR markers.
|
Clusters of Y-STR Haplotypes |
Number of Y-STR Loci Used in the Calculation |
TMRCA, YBP |
|---|---|---|
|
N-Y24317(xB499) |
36 (deletion in DYF389S1) |
900 ± 220 |
|
N-Y16319(xY16223) |
36 (deletion in DYF389S1) |
680 ± 140 |
|
N-Y16223(xZ35328, F2288), DYS385 = 11,11 |
36 (deletion in DYF389S1) |
660 ± 130 |
|
O-F492 |
35 (without DYF389S1) |
280 ± 160 |
Although macrohaplogroup O-M175 is represented by a variety of clans, its carriers are concentrated in the southeast of Tuva (the Tere-Khol and Erzin regions) and in the adjacent Central (Tandy) region.
The analysis of literature data shows that 3 of 24 samples have a match in steppe populations (Figure 2). A Tuvan haplotype from cluster O-FGC85750 belongs to branch O-FGC16885, which is a dominant branch in the Naiman Kazakh of the Middle Zhuz (Supplementary Table S1 [58]) and is also found in Altai Telengits (n = 2) and Kyrgyz (n = 1) (from our own dataset). The Tuvan Mongolian Ancestry 1 haplotype (Figure 2) is only two mutation steps away from the haplotypes of an Uriankhai Mongolians and two Derbets from Kalmykia (from our own dataset), which might point to its Oirat origin. It is also found in eight Altaians (n = 6 from our own data; n = 2 [28]).
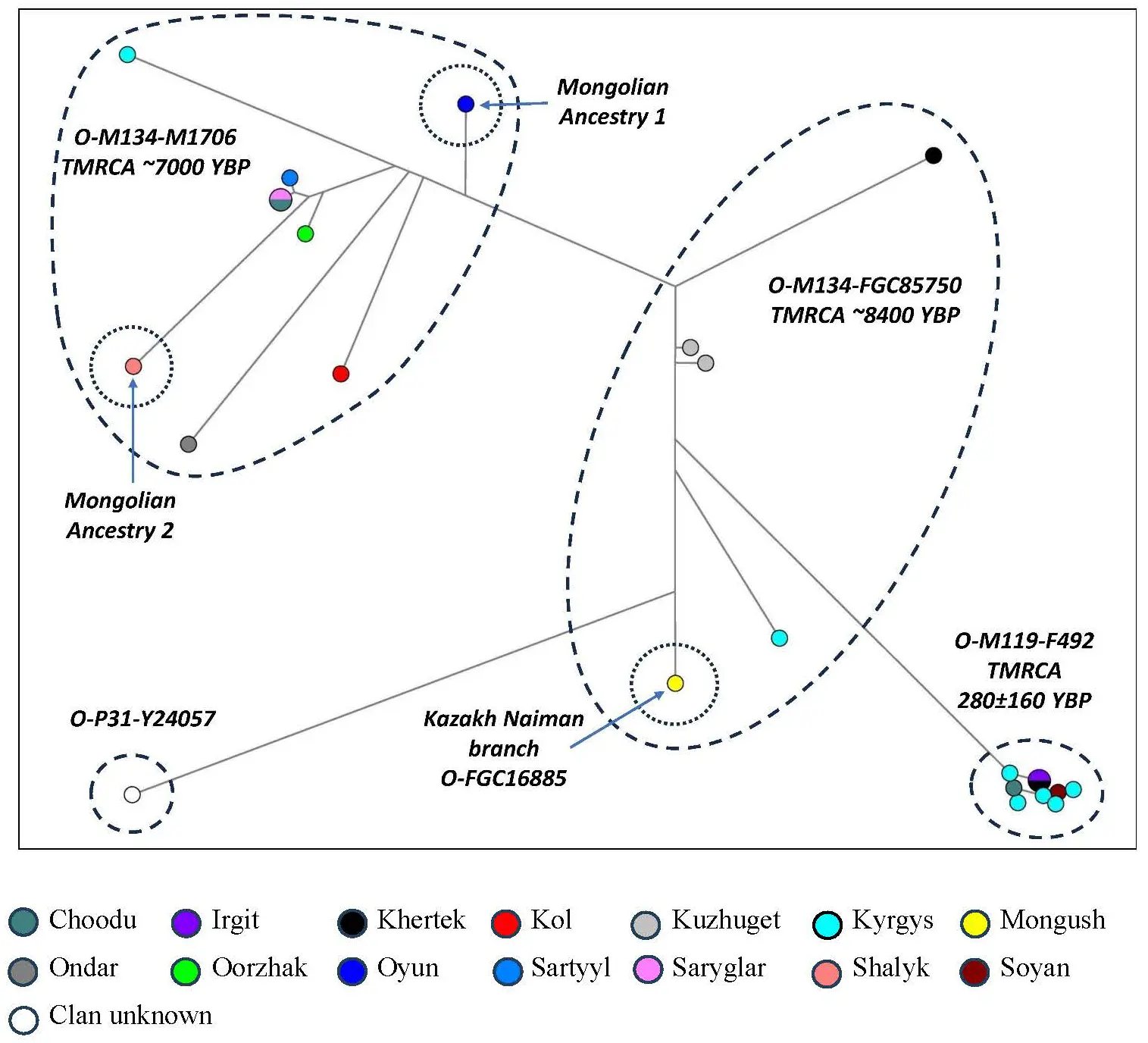
Figure 2. The median network of Y-STR haplotypes for haplogroup O-M175 (n = 24). Legend: Colored circles represent populations and clans.
The Mongolian Ancestry 2 haplotype is only one mutation step away from the haplotype of two Dariganga Mongolians from Eastern Mongolia and two mutation steps away from the haplotype of a Hotgoit Mongolians (Khovsgol Aimak) from our own dataset. The systematic search of the published databases has not revealed other matches with Turkic-Mongolian populations.
So far, only 10 ancient samples of macrohaplogroup O-M175 have been discovered in Mongolia and Russia (9 in Mongolia and 1 in the Primorsky Territory in Russia [37,59,60]. Most of them (6) are dated to the Middle Ages, but there is no connection between their ancient DNA and Tuvan haplotypes (Figure 2).
Thus, the origin of most O-M175 samples in the Tuvan gene pool might be associated with China and less so with Mongolia and Kazakhstan.
4. Discussion
The results of our analysis invite the reader to look at the genetic history of the region and, more specifically, at N-M178 and O-M175 in a broader historical context: from Bronze Age migrations to Mongol invasions.
During the Early Iron Age, the territory of Tuva was inhabited by populations associated with the Aldybel (8th–6th centuries BCE) and Uyuk-Sagly (5th–3rd centuries BCE) cultures. Their genomes differed from those of modern Tuva people in having a significant proportion of a steppe genetic component from the Middle and Late Bronze Age (Steppe_MLBA, [47]) with a predominance of West Eurasian traits. In contrast, modern Tuva people show a high proportion of various East Eurasian genetic components (China_YellowRiver_MN, China_AmurRiver_HG, etc. [47]). This proportion gradually increased, beginning in the Xiongnu era and continuing throughout the first millennium CE, and was accompanied by an increase in the frequency of Y-chromosome haplogroups N-M178, C-M217, O-M175, and D-M174, with a simultaneous decrease in the frequency of haplogroups R1a-M198 and Q-M242 [9].
About 4000 YBP, ancestors of Uralic-speaking N-M178 carriers began migrating from the territory of today’s Yakutia, spreading to Western Siberia and Northern Europe [47]. They migrated across the forest-steppe zone and left almost no trace in the populations who lived further to the south: the “North-European” branches of haplogroup N-M178 (N-B211, N-CTS10760, N-Z1936) are almost non-existent in the modern Tuvan, Khakass, Shor and Altai peoples. The literature describes two ancient samples of the then young (4200-4100 YBP) haplogroup N-L392 (N-CTS6967) discovered in the forest-steppe of the Krasnoyarsk territory (Supplementary Table S2) [47,61]. Perhaps, that forest-steppe migration is also the reason why the relict N-Z1979(xL392) lineage is still found in Teleuts [26,62,63].
In Northern and Southern Altaians, the frequency of haplogroup N-M178 does not exceed a 5% threshold for a polymorphism [27]. However, the frequency of its subhaplogroup N-Y24317, represented by N-Y24317(xB499), reaches 6% in the Tuvan population, and N-B499 occurs in about 20% of the Shor and Khakass populations. At the same time, this haplogroup is rarely found in their neighbors, and its origin is unknown. Among very few modern carriers of this haplogroup are the Kyrgyz of Tadzhikistan (from our own dataset), Nepalis [64], and Indians [49] (Supplementary Table S1). The oldest N-Y24317 carriers (Supplementary Table S2) are represented by four samples from Siberia (~8000–5000 YBP [47,65,66]), two samples from Northwest Mongolia (~3800 YBP [60] and ~3200 YBP [37]) and one sample from Xinjiang (~2700–2300 YBP [67]).This data is consistent with a hypothesis proposed by [42] that haplogroup N-Y24317 first spread from Tuva to Mountainous Shoria and then to Khakassia. Perhaps N-Y24317 carriers arrived in Tuva from Mongolia or Northwest China in the Bronze or Early Iron Age and survived among isolated mountainous populations. This means that the history of N-Y24317 in Tuva might have begun from Central Asian tribes of unknown origin, but not from Samoyedic or Ugric tribes. To support or refute this hypothesis, more data on ancient South Siberian and Central Asian DNA are needed.
In the Khakass and Shor populations, haplogroup N-M178 is represented by only one branch (N-B499). In Tuvans, however, two of its branches can be found: N-Y24317(xB499), which occurs at ~6%, and N-F4205 at ~9%. A lot of N-L708 carriers dispersed to all sides of the world at 7000–3000 YBP. They gave rise [68] to the European branches N-B211, N-CTS10760, and N-Z1936, the Northeastern branch (the Kyordyughen warrior branch) N-GG1002, the Chukchi-Koryak-Itelmen branch N-Z35350 and the Nanai branch N-B479. By contrast, the ancient ancestors of N-F4205 carriers remained in Transbaikalia and Mongolia (Supplementary Table S2); their descendants became steppe nomads. Today, N-F4205 branches are found primarily among Turkic- and Mongolian-speaking peoples. The oldest N-F4205 sample (~2000 YBP) was discovered at a burial site of late Xiongnu individuals in Buryatia [37]). Its haplotype was marked by the presence of the F4205 and F3271 mutations and the absence of the Y16221 and Y16220 mutations; Y16319 and Y16223 sites were uncalled. The second oldest N-F4205 sample (~1600 YBP) was found in Kazakhstan [59] and placed on branch N-Y16313. Another N-F4205 carrier was a Xianbei noble (~1600–1400 YBP) reported in [69]. Multiple N-F4205 samples from Medieval Hungarian and Austrian Avars [70,71,72,73] are located on various branches of haplogroup N-F4205, except for N-Y226011.
These findings can be explained by an initially rapid increase in the number of individuals carrying the F4205 allele around 2500 YBP [49] within some ancient population. As a hypothesis, this could have been the Xiongnu, or the Dingling (possible ancestor of the Tiele). The expansion of the Xiongnu across the East Eurasian steppe, their political and cultural influence led to the arrival in Tuva of new population groups who established the Ulug-Khem culture (2nd–1st centuries BCE [74]) in Tuva. Perhaps, carriers of the F4205 branches that are now ethno-specific to Tuvans (N-Y16319(xY16223), N-Y16223(xZ35328, F2288), DYS385 = 11,11 and DYS391 = 11) were among those newcomers. Given the current scarcity of diverse ancient DNA samples from the Xiongnu period in Tuva, this connection remains a hypothesis. Further research is needed to confirm it. The Ulug-Khem culture ceased to exist after the Xiongnu nomadic empire came apart, and its place was taken by the Kokel culture (2nd–4th centuries CE). During that period, the Tuvan population remained isolated from the rest of the world [75], which means that other F4205 branches (N-Y16223(xZ35328, F2288), DYS385 = 11,13, N-Z35328, N-F2288) entered Tuva much later in the Middle Ages and the Modern Era.
Table 2 shows TMRCA for the clusters within the constructed phylogenetic network estimated from Y-STR markers (Figure 1 and Figure 2). TMRCA estimates suggest that the most recent common ancestors of all studied branches lived in the second millennium CE. Therefore, it would be reasonable to hypothesize that these branches are so young because they arose during the Mongol invasions in the 13th–14th centuries CE.
At that time, the territory of modern Tuva was part of the Lingbei Province of the Mongolian Yuan Empire (1271–1368 CE). Founders of the local lineages (N-Y24317(xB499), N-Y16319(xY16223), N-Y16223(xZ35328, F2288), DYS385 = 11,11) produced a lot of male offspring; perhaps, their reproductive success was due to the large-scale administrative and demographic reforms implemented by the Empire.
In 1267, the Empire set up a yam system of 14 relay posts stretching all the way to the Chazadyr River, a tributary of the Barlyk river [76]. Before the Yuan general Tutuha launched his campaign against the Yenisei Kyrgyz in 1291, six relay posts that had probably been destroyed during the long Kaidu–Kublai war were rebuilt [76]. The last station on the route was downstream the estuary of the Barlyk river, on the right bank of the Khemchik, near a trading post operated by Central Asian merchants [77]. The Empire used the relay system to relocate the local population from the territories of modern Tuva and Khakassia to other provinces. Chinese, Mongols, and Uyghurs were moved in the opposite direction into the settlements of craftsmen and farmers. Such a policy was aimed at neutralizing any potential threat from the local population and creating a supply chain for the Mongolian army [77].
According to the Yuanshi chronicle, “in 1269 Liu Hao-Li was appointed to govern the Jilijisi and the Hanghen, as well as the regions of Qianzhou and Yilanzhou. It was then that storehouses and granaries were built here, relay posts were set up, and administrative institutions were established. Before that, the local tribes made cups and dishes from wood. They could not cast metal to make agricultural tools. Their knowledge of all good things came from the ruling Yuan dynasty. That is when craftsmen were sent there to teach pottery, metal casting and boat building to the local population of these two regions. This helped the local people a lot” [78].
The historical facts mentioned above explain the increased frequency of haplogroup O-M175 in Tuvans in comparison with other Siberian peoples. The Yuan region Yilanzhou spanned the territory of the modern Tandy district, where O-M122 samples were found (the Choodu, Kuzhuget, Ondar, Oyun, Sartyyl, Saryglar clans). The residence of the viceroy Liu Hao-Li was in Yilanzhou. This might imply that the majority of the samples within the phylogenetic network for O-M175 (Figure 2), including the O-F492 lineage, have a Han origin and are associated with Medieval Chinese craftsmen and farmers whom the Mongols forcibly resettled in the 13th century to what is now Tuva.
According to the Yuanshi chronicle, Kublai founded the city of Zhaozhou (1293) in the land of the Mongol prince Nayan after suppressing his uprising. Kublai ordered three tribes from Tuva to live there, including the Yuansuhan, Kanas and Jilijisi (Kyrgyz) [78]. The city lied in what is now the county of Zhaodong in the province of Heilongjiang [79]. We searched the literature on Northeast China for the Y-STR haplotypes of some lineages specific to modern Tuvans, including N-Y24317(xB499), N-Y16319(xY16223), and N-Y16223(xZ35328, F2288) at DYS385 = 11,11 [80]. The search returned no results, which is unsurprising for clusters whose age is comparable to that of Zhaozhou. At the same time, we found two Han samples of R1a-Y39884 and Q-YP1102 from Heilongjiang, the possible descendants of the Tuvans coerced to move from Tuva in 1293 (Supplementary Table S1). Carriers of R1a-Y39884 and Q-YP1102 lineages have inhabited Tuva since the Scythian-Siberian era [9].
5. Conclusions
Some N-M178 branches are exclusive to the Tuvans and are very rare outside of the areas inhabited by this ethnic group. The N-Y24317(xB499) branch may have reached Tuva from Mongolia or Northeast China during the Bronze or Early Iron Age. The origin of the ethno-specific Tuvan branches N-Y16319(xY16223) and N-Y16223(xZ35328, F2288), DYS385 = 11,11, correlates with the growth and expansion in Tuva of some external population approximately 2100 YBP. The Xiongnu, as well as the Dingling they conquered, can be considered as a hypothesis. The rest of the smaller branches (N-Z35328, N-F2288, and N-Y16223(xZ35328, F2288), DYS385 = 11,13) are not specific to Tuvans: they are common in Turkic-Mongolian populations, and their history in Tuva may be associated with the Medieval expansion of the Mongols.
The Turkic-Mongolian contribution of haplogroup O-M175 to the gene pool of Tuvans is insignificant. The possible ancestors of most O-M175 lineages might have been Hans forced to resettle at the time of the Yuan Empire.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/1068, Figure S1: Location of the clan groups from which samples were collected on a map of Tuva; Table S1: Modern Y-STR haplotypes from literature [28,58,64,80,81]; Table S2: Ancient Y-chromosomes [37,47,59,60,61,65,66,67,69].
Acknowledgments
The authors thank all the sample donors and the Biobank of Northern Eurasia for the access to DNA collections.
Author Contributions
Conceptualization, D.A. and E.B.; Methodology, D.A.; Validation, E.B. and E.P.; Formal Analysis, D.A.; Investigation, D.A. and G.P.; Resources, L.D. and G.P.; Data Curation, E.B. and G.P.; Writing—Original Draft Preparation, D.A.; Writing—Review & Editing, D.A., E.B. and E.P.; Visualization, D.A. and E.B.; Supervision, E.B.; Project Administration, E.B.; Funding Acquisition, E.B. All authors have read and approved the final manuscript.
Ethics Statement
The study was conducted according to the guidelines of the Declaration of Helsinki, and approved by the Ethics Committee of Research Centre for Medical Genetics (protocol No. 1 of 29 June 2020).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
All data used in the paper will be available to download from anyone with an approval from Medical Genetic Research Centre committee in accordance with Federal Law of Russian Federation 43-FZ of 20 February 2026.
Funding
The research was carried out within the state assignment of Ministry of Science and Higher Education of the Russian Federation for Research Centre for Medical Genetics.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Zhao X, Sun R, Li C, Cui Y. Genetic History of Ancient Xinjiang Revealed by Ancient DNA Study: A Hub of Eurasian Population Migration and Cultural Exchange. Nat. Anthropol. 2025, 3, 10010. DOI:10.70322/natanthropol.2025.10010 [Google Scholar]
-
Unterländer M, Palstra F, Lazaridis I, Pilipenko A, Hofmanová Z, Groß M, et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe. Nat. Commun. 2017, 8, 14615. DOI:10.1038/ncomms14615 [Google Scholar]
-
Mary L, Zvénigorosky V, Kovalev A, Gonzalez A, Fausser J-L, Jagorel F, et al. Genetic kinship and admixture in Iron Age Scytho-Siberians. Hum. Genet. 2019, 138, 411–423. DOI:10.1007/s00439-019-02002-y [Google Scholar]
-
Kilunovskaya ME, Alborova IE, Busova VS, Brown S, Lazarevskaya NA, Mustafin KK, et al. A Comprehensive Study of the Monuments of the End of 2nd Millennium—Beginning of 1st Millennium BC in the Valley of the River Eerbek (Central Tuva). Sib. Hist. Res. 2022, 4, 267–307. DOI:10.17223/2312461X/38/14 (In Russian) [Google Scholar]
-
Kilunovskaya ME, Alborova IA, Busova VS, Semenov MK, Uchaneva EN. Archaeological sites of the mid-1st millennium in the Urochishche Eki-Ottug (Central Tuva): From typology and chronology to anthropology and genetics. Archaeol News 2023, 40, 245–273. Available online: https://www.academia.edu/113376042/Archaeological_sites_of_the_mid_1st_millennium_in_the_Urochishche_Eki_Ottug_Central_Tuva_from_typology_and_chronology_to_anthropology_and_genetics_%D0%9F%D0%B0%D0%BC%D1%8F%D1%82%D0%BD%D0%B8%D0%BA%D0%B8_%D1%81%D0%B5%D1%80%D0%B5%D0%B4%D0%B8%D0%BD%D1%8B_I_%D1%82%D1%8B%D1%81%D1%8F%D1%87%D0%B5%D0%BB%D0%B5%D1%82%D0%B8%D1%8F_%D0%B4%D0%BE_%D0%BD_%D1%8D_%D0%B2_%D1%83%D1%80%D0%BE%D1%87%D0%B8%D1%89%D0%B5_%D0%AD%D0%BA%D0%B8_%D0%9E%D1%82%D1%82%D1%83%D0%B3_%D0%A6%D0%B5%D0%BD%D1%82%D1%80%D0%B0%D0%BB%D1%8C%D0%BD%D0%B0%D1%8F_%D0%A2%D1%83%D0%B2%D0%B0_%D0%BE%D1%82_%D1%82%D0%B8%D0%BF%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D0%B8_%D0%B8_%D1%85%D1%80%D0%BE%D0%BD%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D0%B8_%D0%BA_%D0%B0%D0%BD%D1%82%D1%80%D0%BE%D0%BF%D0%BE%D0%BB%D0%BE%D0%B3%D0%B8%D0%B8_%D0%B8_%D0%B3%D0%B5%D0%BD%D0%B5%D1%82%D0%B8%D0%BA%D0%B5 (accessed on 28 May 2026). (In Russian)
-
Kilunovskaya ME, Alborova IE, Anikeeva OV, Brown S, Busova VS, Kozhukhova EI, et al. Reconstructing Ethnocultural Processes in the Sausken Valley (Tuva) during the Later Half of the 1st Millennium BC. Ufa Archaeol. Her. 2024, 24, 263–295. DOI:10.31833/uav/2024.24.2.016 (In Russian) [Google Scholar]
-
McColl H, Kroonen GJ. Moreno-Mayar V, Seersholm FV, Scorrano G, Pinotti T, et al. Steppe Ancestry in western Eurasia and the spread of the Germanic Languages. bioRxiv 2024, preprint. DOI:10.1101/2024.03.13.584607 [Google Scholar]
-
Nedoluzhko A, Vergasova E, Sharko F, Agapitova N, Kharitonov D, Sukhanova X, et al. Ancient DNA analysis of elite nomadic warrior from Chinge-Tey I funerary commemorative complex in the “Valley of the Kings”, Tuva. BMC Genom. 2025, 26, 220. DOI:10.1186/s12864-025-11361-y [Google Scholar]
-
Adamov D, Ponomarev G, Damba L, Balanovska E. The Yeniseian and Andronovo substrates in the gene pools of the indigenous southern Siberian populations analyzed by Q and R1a Y-haplogroups. 2026, in press. [Google Scholar]
-
Karafet TM, Zegura SL, Posukh O, Osipova L, Bergen A, Long J, et al. Ancestral Asian sources of new world Y-chromosome founder haplotypes. Am. J. Hum. Genet. 1999, 64, 817–831. DOI:10.1086/302282 [Google Scholar]
-
Stepanov VA, Puzyrev VP. Haplotypes of two diallelic Y chromosome loci in the indigenous and migrant populations of Siberia. Genetika 2000, 36, 87–92. Available online: http://www.medgenetics.ru/UserFile/File/Doc/Evolution%20Doc/Stepanov-RJGenetica-2000-1(36)-76-80.pdf (accessed on 28 May 2026). (In Russian)
-
Stepanov VA, Puzyrev VP. Analysis of the allele frequencies of seven Y-chromosome microsatellite loci in three Tuvinian populations. Genetika 2000, 36, 241–248. Available online: http://www.medgenetics.ru/UserFile/File/Doc/Evolution%20Doc/Stepanov-Rus.J.Genetics-2000-36(2)-179-185-Y-(English).pdf (accessed on 28 May 2026). (In Russian)
-
Stepanov VA, Puzyrev VP. Y-chromosome microsatellite haplotypes demonstrate absense of subdivision and presence of several components in the tuvinian male gene pool. Genetika 2000, 36, 377–384. Available online: https://elibrary.ru/item.asp?id=13346384 (accessed on 28 May 2026). (In Russian)
-
Stepanov VA, Khitrinskaya IY, Puzyrev VP. Genetic differentiation of the Tuva population with respect to the Alu-insertions. Russ. J. Genet. 2001, 37, 453–459. DOI:10.1023/A:1016623030663 [Google Scholar]
-
Wells RS, Yuldasheva N, Ruzibakiev R, Underhill PA, Evseeva I, Blue-Smith J, et al. The Eurasian heartland: A continental per spective on Y-chromosome diversity. Proc. Natl. Acad. Sci. USA 2001, 98, 10244–10249. DOI:10.1073/pnas.171305098 [Google Scholar]
-
Karafet TM, Osipova LP, Gubina M, Posukh OL, Zegura SL, Hammer MF. High Levels of Y-Chromosome Differentiation among Native Siberian Populations and the Genetic Signature of a Boreal Hunter-Gatherer Way of Life. Hum. Biol. 2002, 74, 761–789. DOI:10.1353/hub.2003.0006 [Google Scholar]
-
Lell JT, Sukernik RI, Starikovskaya YB, Su B, Jin L, Schurr TG, et al. The dual origin and Siberian affinities of Native American Y chromosomes. Am. J. Hum. Genet. 2002, 70, 192–206. DOI:10.1086/338457 [Google Scholar]
-
Derenko MV, Malyarchuk BA, Denisova GA, Dorzhu CM, Ondar UN, Karamchakova ON, et al. Polymorphism of the Y-chromosome diallelic loci in ethnic groups of the Altai-Sayan Region. Russ. J. Genet. 2002, 38, 309–314. DOI:10.1023/A:1014863020171 (In Russian) [Google Scholar]
-
Zerjal T, Xue Y, Bertorelle G, Wells RS, Bao W, Zhu S, et al. The genetic legacy of the Mongols. Am. J. Hum. Genet. 2003, 72, 717–721. DOI:10.1086/367774 [Google Scholar]
-
Tambets K, Rootsi S, Kivisild T, Help H, Serk P, Loogvali EL, et al. The Western and Eastern Roots of the Saami—The Story of Genetic “Outliers” Told by Mitochondrial DNA and Y Chromosomes. Am. J. Hum. Genet. 2004, 74, 661–682. DOI:10.1086/383203 [Google Scholar]
-
Pakendorf B, Novgorodov IN, Osakovskij VL, Danilova AP, Protod’jakonov AP, Stoneking M. Investigating the effects of prehistoric migrations in Siberia: Genetic variation and the origins of Yakuts. Hum Genet 2006, 120, 334–353. DOI:10.1007/s00439-006-0213-2 [Google Scholar]
-
Derenko MV, Malyarchuk BA, Wozniak M, Dambuveva IK, Dorzhu CM, Luzina FA, et al. The diversity of Y-chromosome lineages in indigenous population of South Siberia. Dokl. Biol. Sci. 2006, 411, 466–470. DOI:10.1134/s0012496606060123 [Google Scholar]
-
Stepanov VA, Kharkov VN, Puzyrev VP. Evolution and phylogeography of human Y-chromosomal lineages. Vavilov J. Genet. Breed. 2006, 10, 57–73. Available online: https://vavilovj-icg.ru/download/06_Stepanov.pdf (accessed on 28 May 2026). (In Russian)
-
Derenko M, Malyarchuk B, Denisova G, Wozniak M, Grzybowski T, Dambueva I, et al. Y-chromosome haplogroup N dispersals from south Siberia to Europe. J. Hum. Genet. 2007, 52, 763–770. DOI:10.1007/s10038-007-0179-5 [Google Scholar]
-
Kharkov VN, Stepanov VA, Medvedeva OF, Spiridonova MG, Puzyrev VP, Voevoda NI, et al. Gene pool differences between Northern and Southern Altaians inferred from the data on Y-chromosomal haplogroups. Russ. J. Genet. 2007, 43, 551–562. DOI:10.1134/S1022795407050110 [Google Scholar]
-
Kharkov VN, Medvedeva OF, Luzina FA, Kolbasko AV, Gafarov NI, Puzyrev VP, et al. Comparative Characteristics of the Gene Pool of Teleuts Inferred from Y-Chromosomal Marker Data. Russ. J. Genet. 2009, 45, 994–1003. DOI:10.1134/S1022795409080158 [Google Scholar]
-
Balaganskaya OA, Lavryashina MB, Kuznetchova MA, Romanov AG, Dibirova KhD, Frolova SA, et al. Gene pool of the Altai ethnic groups (from Russia, Kazakhstan, and Mongolia) analyzed by the Y chromosomal markers. Mosc. Univ. Anthropol. Bull. 2011, 2, 25–36. Available online: https://cyberleninka.ru/article/n/geneticheskaya-struktura-po-markeram-y-hromosomy-narodov-altaya-rossii-kazahstana-mongolii (accessed on 28 May 2026). (In Russian)
-
Dulik MC, Zhadanov SI, Osipova LP, Askapuli A, Gau L, Gokcumen O, et al. Mitochondrial DNA and Y chromosome variation provides evidence for a recent common ancestry between Native Americans and Indigenous Altaians. Am. J. Hum. Genet. 2012, 90, 229–246. DOI:10.1016/j.ajhg.2011.12.014 [Google Scholar]
-
Kharkov VN, Khamina KV, Medvedeva OF, Simonova KV, Khitrinskaya IY, Stepanov VA. Gene-pool structure of Tuvans inferred from Y-chromosome mark er data. Russ. J. Genet. 2013, 49, 1236–1244. DOI:10.1134/S102279541312003X [Google Scholar]
-
Balanovsky O, Zhabagin M, Agdzhoyan A, Chukhryaeva M, Zaporozhchenko V, Utevska O, et al. Deep phylogenetic analysis of haplogroup G1 provides estimates of SNP and STR mutation rates on the human Y-chromosome and reveals migrations of Iranic speakers. PLoS ONE 2015, 10, e0122968. DOI:10.1371/journal.pone.0122968 [Google Scholar]
-
Ilumäe AM, Reidla M, Chukhryaeva M, Järve M, Post H, Karmin M, et al. Human Y chromosome haplogroup N: A non-trivial time-resolved phylogeography that cuts across language families. Am. J. Hum. Genet. 2016, 99, 163–173. DOI:10.1016/j.ajhg.2016.05.025 [Google Scholar]
-
Balanovska E, Yusupov Y, Shalyaho R, Stepanov G, Asilgujin R, Zhabagin M, et al. Genetic portraits of seven clans of North-Western Bashkirs: Contribution of the Finno-Ugric genetic component to the Bashkirian gene pool. Mosc. Univ. Anthropol. Bull. 2017, 3, 94–103. Available online: https://cyberleninka.ru/article/n/geneticheskie-portrety-semi-klanov-severo-zapadnyh-bashkir-vklad-finno-ugorskogo-komponenta-v-genofond-bashkir (accessed on 28 May 2026). (In Russian)
-
Damba LD, Balanovska EV, Zhabagin MK, Yusupov YM, Bogunov YV, Sabitov ZM, et al. Estimating the impact of Mongol expansion on gene pool of Tuvans. Vavilov J. Genet. Breed. 2018, 22, 611–619. DOI:10.18699/VJ18.402 (In Russian) [Google Scholar]
-
Balanovska EV, Damba LD, Agdzhoyan AT, Zhabagin MK, Olkova MV, Kagazezheva ZA, et al. The gene pool of hunter-reindeer herders of Southern Siberia: Tofalars and Todzhins. Mosc. Univ. Anthropol. Bull. 2019, 4, 67–80. DOI:10.32521/2074-8132.2019.4.067-080 (In Russian) [Google Scholar]
-
Damba LD, Balanovska EV, Agdzhoyan AT, Korotkova NA, Olkova MV, Utrivan SА, et al. Gene pool of three Eastern Tuvans clans according to Y-chromosome polymorphism. Mosc. Univ. Anthropol. Bull. 2019, 1, 74–85. Available online: https://cyberleninka.ru/article/n/genofond-treh-vostochnyh-rodov-tuvintsev-po-dannym-polimorfizma-y-hromosomy (accessed on 28 May 2026). (In Russian)
-
Zhabagin MK, Damba LD, Korotkova NA, Chernishenko DN, Utrivan SA, Pilev VY, et al. Analysis of Clan Structure of Tuvans by Y-Chromosome Markers. Russ. J. Genet. 2020, 56, 763–768. DOI:10.1134/S1022795420060137 [Google Scholar]
-
Jeong C, Wang K, Wilkin S, Taylor WTT, Miller BK, Bemmann JH, et al. A Dynamic 6000-Year Genetic History of Eurasia’s Eastern Steppe. Cell 2020, 183, 890–904.e29. DOI:10.1016/j.cell.2020.10.015 [Google Scholar]
-
Agdzhoyan AT, Damba LD, Zaporozhchenko VV, Balanovsky OP. In addressing the question about the Samoyedic substrate in the South Siberian populations: The phylogeography of Y-chromosome haplogroup N-L666. Mosc. Univ. Anthropol. Bull. 2021, 2, 75–86. DOI:10.32521/2074-8132.2021.2.075-086 (In Russian) [Google Scholar]
-
Agdzhoyan AT, Damba LD, Gurianov VM, Zaporozhchenko VV, Balanovsky OP. Phylogenetic Analysis of the South Siberian Haplogroup Q-YP1102 from Y-SNP and Y-STR-Data on Tuvinian and Surrounding Populations. Russ. J. Genet. 2021, 57, 1398–1407. DOI:10.1134/S1022795421120024 (In Russian) [Google Scholar]
-
Damba LD, Zaporozhchenko VV, Balanovsky OP, Balanovska EV. Phylogenetic analysis of Y-chromosomal haplogroup C2a1a2a2a2-SK1066 in the general pool of Tuvan general groups in the context of Central Asian populations. Vestn. Tuvan State Univ. Nat. Agric. Sci. 2021, 4, 6–17. DOI:10.24412/2077-5326-10.24411/2221-0458-2021-85-06-17 (In Russian) [Google Scholar]
-
Damba LD, Aiyzhy EV, Balanovsky OP, Markina NV, Zhabagin MK, Balanovskaya EV. The Central Asian component in the gene pool of the Tuvan tribal group Mongush: On the question of the Mongolian contribution to the ethnogenesis of the Tuvans. Mosc. Univ. Anthropol. Bull. 2022, 1, 77–85. DOI:10.32521/2074-8132.2022.1.077-085 (In Russian) [Google Scholar]
-
Stepanov VA, Kolesnikov NA, Valikhova LV, Zarubin AA, Khitrinskaya IY, Kharkov VN. Structure and origin of Tuvan gene pool according to autosome SNP and Y-chromosome haplogroups. Vavilov J. Genet. Breed. 2023, 27, 36–45. DOI:10.18699/VJGB-23-06 [Google Scholar]
-
Damba LD, Ponomarev GY, Pylev VY, Balanovska EV. The structure of the gene pool of the Southern tribal groups of Tuvans. Mosc. Univ. Anthropol. Bull. 2024, 3, 91–101. DOI:10.55959/MSU2074-8132-24-3-8 (In Russian). [Google Scholar]
-
Kharkov VN, Urmanchieva AY, Kolesnikov NA, Zarubin AA, Valikhova LV, Khitrinskaya IY, et al. Population genetics and comparative linguistics: Phylogeny and phylogeography of haplogroup N1a2b and the linguistic prehistory of the Samoyed peoples. J. Lang. Relatsh. 2024, 22, 109–130. DOI:10.31826/jlr-2024-221-207 (In Russian) [Google Scholar]
-
Balanovska EV, Damba LD, Adamov DS, Ponomarev GY, Potanina AY, Pocheshkhova EA. Diversity of gene pools in twelve Tuvan tribal groups (based on Y-chromosome haplogroup data). Mosc. Univ. Anthropol. Bull. 2025, 3, 61–71. DOI:10.55959/MSU2074-8132-25-3-5 (In Russian) [Google Scholar]
-
Kolesnikova SY, Kharkov VN, Valikhova LV. To a question of the Yeniseian ethnogenesis. Tomsk. State Univ. J. Hist. 2022, 79, 164–170. DOI:10.17223/19988613/79/20 (In Russian) [Google Scholar]
-
Zeng TC, Vyazov LA, Kim A, Flegontov P, Sirak K, Maier R, et al. Ancient DNA reveals the prehistory of the Uralic and Yeniseian peoples. Nature 2025, 644, 122–132. DOI:10.1038/s41586-025-09189-3 [Google Scholar]
-
Balanovska EV, Zhabagin MK, Agdzhoyan AT, Chukhryaeva MI, Markina NV, Balaganskaya OA, et al. Population biobanks: Organizational models and prospects of application in gene geography and personalized medicine. Russ. J. Genet. 2016, 52, 1227–1243. DOI:10.1134/S1022795416120024 [Google Scholar]
-
YFull YTree v13.07.00. 2025. Available online: https://www.yfull.com/tree/ (accessed on 26 December 2025).
-
Bandelt HJ, Forster P, Röhl A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. DOI:10.1093/oxfordjournals.molbev.a026036 [Google Scholar]
-
Zhivotovsky LA, Underhill PA, Cinnioğlu C, Kayser M, Morar B, Kivisild T, et al. The effective mutation rate at Y chromosome short tandem repeats, with application to human population-divergence time. Am. J. Hum. Genet. 2004, 74, 50–61. DOI:10.1086/380911 [Google Scholar]
-
Willuweit S, Roewer L. The new Y Chromosome Haplotype Reference Database. Forensic Sci. Int. Genet. 2015, 15, 43–48. DOI:10.1016/j.fsigen.2014.11.024 [Google Scholar]
-
Ballantyne KN, Goedbloed M, Fang R, Schaap O, Lao O, Wollstein A, et al. Mutability of Y-Chromosomal Microsatellites: Rates, Characteristics, Molecular Bases, and Forensic Implications. Am. J. Hum. Genet. 2010, 87, 341–153. DOI:10.1016/j.ajhg.2010.08.006 [Google Scholar]
-
Fenner J. Cross-cultural estimation of the human generation interval for use in genetics-based population divergence studies. Am. J. Phys. Anthropol. 2005, 128, 415–423. DOI:10.1002/ajpa.20188 [Google Scholar]
-
Yan S, Wang C-C, Zheng H-Z, Wang W, Qin Z-D, Wei L-H, et al. Y Chromosomes of 40% Chinese Descend from Three Neolithic Super-Grandfathers. PLoS ONE 2014, 9, e105691. DOI:10.1371/journal.pone.0105691 [Google Scholar]
-
Sun J, Li Y-X, Ma P-C, Yan S, Cheng H-Z, Fan Z-Q, et al. Shared paternal ancestry of Han, Tai-Kadai-speaking, and Austronesian-speaking populations as revealed by the high resolution phylogeny of O1a-M119 and distribution of its sub-lineages within China. Am. J. Phys. Anthropol. 2021, 174, 686–700. DOI:10.1002/ajpa.24240 [Google Scholar]
-
Zeng Z, Tian J, Jiang C, Ye W, Liu K, Li Y. Inferring the history of surname Ye based on Y chromosome high-resolution genotyping and sequencing data. J. Hum. Genet. 2019, 64, 703–709. DOI:10.1038/s10038-019-0616-2 [Google Scholar]
-
Ashirbekov Y, Nogay A, Abaildayev A, Zhunussova A, Sabitov Z, Zhabagin M. Genetic polymorphism of 27 Y-STR loci in Kazakh populations from Eastern Kazakhstan. Ann. Hum. Biol. 2023, 50, 48–51. DOI:10.1080/03014460.2023.2170465 [Google Scholar]
-
Damgaard PDB, Marchi N, Rasmussen S, Peyrot M, Renaud G, Korneliussen T, et al. 137 ancient human genomes from across the Eurasian steppes. Nature 2018, 557, 369–374. DOI:10.1038/s41586-018-0094-2 [Google Scholar]
-
Wang C-C, Yeh H-Y, Popov AN, Zhang H-Q, Matsumura H, Sirak K, et al. Genomic insights into the formation of human populations in East Asia. Nature 2021, 591, 413–419. DOI:10.1038/s41586-021-03336-2 [Google Scholar]
-
Kılınç GM, Kashuba N, Koptekin D, Bergfeldt N, Dönertaş HM, Rodríguez-Varela R, et al. Human population dynamics and Yersinia pestis in ancient northeast Asia. Sci. Adv. 2021, 7, eabc4587. DOI:10.1126/sciadv.abc4587 [Google Scholar]
-
Kharkov VN, Kolesnikov NA, Zarubin AA, Valikhova LV, Khitrinskaya IY, Voevoda MI, et al. Paleo-Siberian Substrate in the Gene Pool of Koryaks according to Data on Autosomal SNP Polymorphism and Y-Chromosome Haplogroups. Russ. J. Genet. 2024, 60, 787–796. DOI:10.1134/S1022795424700236 [Google Scholar]
-
Karafet TM, Osipova LP, Savina OV, Hallmark B, Hammer MF. Siberian genetic diversity reveals complex origins of the Samoyedic-speaking populations. Am. J. Hum. Biol. 2018, 30, e23194. DOI:10.1002/ajhb.23194 [Google Scholar]
-
Gayden T, Chennakrishnaiah S, La Salvia J, Jimenez S, Regueiro M, Maloney T, et al. Y-STR diversity in the Himalayas. Int. J. Legal Med. 2011, 125, 367–375. DOI:10.1007/s00414-010-0485-x [Google Scholar]
-
Damgaard PDB, Martiniano R, Kamm J, Moreno-Mayar JV, Kroonen G, Peyrot M, et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia. Science 2018, 360, eaar7711. DOI:10.1126/science.aar7711 [Google Scholar]
-
Sirak K, Fernandes D, Cheronet O, Harney E, Mah M, Mallick S, et al. Human auditory ossicles as an alternative optimal source of ancient DNA. Genome Res. 2020, 30, 427–436. DOI:10.1101/gr.260141.119 [Google Scholar]
-
Kumar V, Wang W, Zhang J, Wang Y, Ruan Q, Yu J, et al. Bronze and Iron Age population movements underlie Xinjiang population history. Science 2022, 376, 62–69. DOI:10.1126/science.abk1534 [Google Scholar]
-
Adamov D, Zhabagin M, Balanovska E. Modern descendants of Kyordyughen warrior (Yakutia, 4200 years before present) in populations of Far East. J. Hum. Genet. 2026, 71, 413–418. DOI:10.1038/s10038-026-01459-w [Google Scholar]
-
Zhang J, Zhang F, Qu Y, Suo M, Song G, Chen Y, et al. Archaeogenetic analysis sheds light on genomic substructure and kinship practices of Xianbei nobles from the Yihe Nur site. Archaeol. Anthropol. Sci. 2025, 17, 198. DOI:10.1007/s12520-025-02325-0 [Google Scholar]
-
Maróti Z, Neparáczki E, Schütz O, Maár K, Varga GIB, Kovács B, et al. The genetic origin of Huns, Avars, and conquering Hungarians. Curr. Biol. 2022, 32, 2858–2870.e7. DOI:10.1016/j.cub.2022.04.093 [Google Scholar]
-
Gnecchi-Ruscone GA, Szécsényi-Nagy A, Koncz I, Csiky G, Rácz Z, Rohrlach AB, et al. Ancient genomes reveal origin and rapid trans-Eurasian migration of 7th century Avar elites. Cell 2022, 185, 1402–1413.e21. DOI:10.1016/j.cell.2022.03.007 [Google Scholar]
-
Gnecchi-Ruscone GA, Rácz Z, Samu L, Szeniczey T, Faragó N, Knipper C, et al. Network of large pedigrees reveals social practices of Avar communities. Nature 2024, 629, 376–383. DOI:10.1038/s41586-024-07312-4 [Google Scholar]
-
Wang K, Tobias B, Pany-Kucera D, Berner M, Eggers S, Gnecchi-Ruscone GA, et al. Ancient DNA reveals reproductive barrier despite shared Avar-period culture. Nature 2025, 638, 1007–1014. DOI:10.1038/s41586-024-08418-5 [Google Scholar]
-
Kilunovskaya ME, Leus PM. Dating of burial grounds of the Xiongnu Era Ala-Tey 1 and Terezin in Tuva. Sayan-Altai Sci. Rev. 2021, 1, 79–91. DOI:10.52782/KRIL.2021.1.29.007 (In Russian) [Google Scholar]
-
Sadykov T, Caspari G, Blochin J, Lösch S, Kapinus Y, Milella M. The Kokel of Southern Siberia: New data on a post-Xiongnu material culture. PLoS ONE 2021, 16, e0254545. DOI:10.1371/journal.pone.0254545 [Google Scholar]
-
Khrapachevsky RP. “The Annals of Kublai Khan”, The Main Source on the History of the Reign of the First Emperor of the Yuan Dynasty; Center for the Study of Military and General History: Moscow, Russia, 2019; pp. 133, 576. [Google Scholar]
-
Kyzlasov LR. History of Tuva in the Middle Ages; Moscow State University Publishing House: Moscow, Russia, 1969; pp. 138, 169. (In Russian) [Google Scholar]
-
Kyrgyz-Turkish University “Manas”. Materials on the History of the Kyrgyz People and Kyrgyzstan (Excerpts from Chinese Sources from the 2nd Century BC to the 18th Century); Dzhusayev K, Ed.; Kyrgyz-Turkish University “Manas”: Bishkek, Kyrgyzstan, 2003; Volume 2; p. 55. [Google Scholar]
-
Jarkyn T. Research on the Kyrgyz of Heilongjiang (China). In Peoples and Cultures of the Sayan-Altai and Bordering Territories; Mainagasheva NS, Ed.; Publishing House of the Khakass Research Institute for Language, Literature, and History: Abakan, Russia, 2023; pp. 127–135. (In Russian) [Google Scholar]
-
Song W, Zhou S, Yu W, Fan Y, Liang X. Genetic analysis of 42 Y-STR loci in Han and Manchu populations from the three northeastern provinces in China. BMC Genom. 2023, 24, 578. DOI:10.1186/s12864-023-09636-3 [Google Scholar]
-
Malyarchuk B, Derenko M, Denisova G, Khoyt S, Woźniak M, Grzybowski T, et al. Y-chromosome diversity in the Kalmyks at the ethnical and tribal levels. J. Hum. Genet. 2013, 58, 804–811. DOI:10.1038/jhg.2013.108 [Google Scholar]