Morphological and Genome Characterization of Alternaria alternata Causing Blueberry (Vaccinium corymbosum L.) Leaf Spot in Peru

Morphological and Genome Characterization of Alternaria alternata Causing Blueberry (Vaccinium corymbosum L.) Leaf Spot in Peru
Ricardo Velasquez 1,† Valentina Osorio 2,† Ana Maria Leiva 2 Juan Manuel Pardo 2 Alejandra Gil-Ordóñez 2 Ida Bartolini 3 Wilmer J. Cuellar 2,*
Received: 26 January 2026 Revised: 10 April 2026 Accepted: 09 May 2026 Published: 20 May 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Introduction
Blueberries (Vaccinium corymbosum L.), a perennial crop native to North America and part of the Ericaceae family, are highly valued for their nutritional properties and global economic importance. The increasing global demand for blueberries has driven expansion in Peru’s production. Although commercial production began in 2007, the industry experienced remarkable growth following a national campaign launched in 2012, expanding from just 50 hectares to over 18,000 hectares in 2022. That year, blueberries had become Peru’s top agro-export crop, generating $1.435 billion in revenue and accounting for approximately 14% of the nation’s total agro-export earnings [1]. Today, Peru ranks as the world’s third-largest blueberry producer, following the United States (U.S.) and Canada, and is the leading producer in Latin America. The country exports to major markets, including the U.S., Europe, and China [2,3].
Nevertheless, this crop is highly susceptible to fungal pathogens that cause severe economic losses. Among them, Alternaria spp. are a broad-host-range pathogen responsible for yield reductions of 20% to 80% widely [4], and postharvest losses of up to 46% in China. Economic losses in the blueberry industry due to Alternaria spp. infections are estimated to be between $230 million and $1 billion globally [5]. These substantial financial impacts underscore the critical importance of early detection and timely implementation of management to contain the spread of the disease and lower the associated costs of disease management at such scales.
Globally, the most frequently reported Alternaria species affecting blueberries are A. alternata and A. tenuissima, which have been associated with leaf spot in major producing countries such as the U.S., China, South Korea, and Australia [6,7,8,9]. Additionally, these species have been linked to fruit rot in blueberries in the U.S. and Argentina [10,11]. These are polycyclic pathogens that persist in soil and plant debris through various survival structures. Under favorable environmental conditions, conidia initiate infections through natural openings and wounds [12,13]. However, despite this extensive global documentation of Alternaria species affecting blueberries, no formal reports have documented these pathogens in blueberry crops in Peru. Although Alternaria spp. have been reported affecting other crops in Peru, such as citrus and tomato [14,15], this knowledge gap represents a phytosanitary concern that limits effective disease management.
Therefore, the objective of this study was to isolate, identify, and characterize the causal agent of leaf spot symptoms observed in blueberry plants in Peru, integrating morphological, molecular, and biological approaches (including fulfillment of Koch’s postulates). By scientifically documenting its identity, this work provides a robust foundation for future epidemiological studies and targeted disease management strategies.
2. Materials and Methods
2.1. Sample Collection and Pathogen Isolation
In 2022, leaf spots were first observed on V. corymbosum cv. Biloxi in a field in northern Lima (–11.51478, –77.23869). Plants were evaluated for symptoms by examining lesion characteristics, including color, shape, pattern distribution, and presence of fungal signs. Disease incidence, calculated as the percentage of affected leaves per plant, was assessed using a stratified random sampling of 30 plants across three homogeneous field strata (10 plants per stratum) [16], covering a total area of 1800 m2. Fruit rot incidence was evaluated by collecting 9250 fruits from the same 30 plants across three harvest periods and expressing it as the percentage of symptomatic fruits. For pathogen isolation, five plants showing moderate symptom severity were selected to avoid advanced lesions potentially colonized by secondary microorganisms. From these plants, ten symptomatic leaves were collected from the middle canopy, as this location represents the active disease front, ensuring that tissues retain cellular integrity and nutrients necessary for pathogen metabolism, unlike lower leaves already compromised by advanced senescence or upper leaves that may not yet have been colonized [17]. Small tissue fragments (5 mm diameter) were excised from the margins of necrotic lesions using a sterile scalpel.
Samples were surface disinfected by sequential immersion in 1% NaClO for 1 min, followed immediately by 70% ethanol for 1 min, and then rinsed three times with sterile distilled water. All procedures were conducted under aseptic conditions without interruption. Disinfected tissues were placed on potato dextrose agar (PDA) and incubated at 25 °C under continuous darkness. Fungal growth was observed after three days, and single-spore purification was subsequently performed to obtain pure cultures.
2.2. Morphological Characterization
A total of five fungal isolates were obtained from symptomatic tissues, all showing consistent macroscopic and microscopic characteristics typical of Alternaria alternata. Due to this uniformity, one representative isolate was selected for detailed characterization. Macroscopic characteristics were evaluated on PDA, including colony color (surface and reverse), texture, margin type, and mycelial appearance. For microscopic characterization, cultures were grown on potato carrot agar (PCA), a medium known to promote sporulation in Alternaria spp., and incubated at 25 °C under an 8 h light/16 h dark photoperiod for 7 days. Conidia were examined using a light microscope (BA210, Motic, Xiamen, China) at 40× magnification, and measurements were obtained from 50 randomly selected conidia following the criteria described by Simmons (2007) [18].
2.3. Genome Characterization
A single representative isolate obtained by single-spore purification was selected for genomic analysis and designated as Alternaria alternata isolate Per1. For DNA extraction, isolate Per1 was grown on PDA medium at 25 °C under continuous darkness for 2–4 weeks. Approximately 0.03 g of fresh mycelium was scraped from agar plates and processed according to the protocol described by Gil-Ordóñez et al. (2024) [19]. DNA integrity was confirmed by gel electrophoresis, and concentration was determined using Qubit™ dsDNA HS Assay Kit (Invitrogen, Life Technologies, Carlsbad, CA, USA). Whole-genome sequencing was performed using Oxford Nanopore Technology (FLO-MIN114 R10 chemistry, LSK114 kit). Genome assembly was conducted using Flye v2.9 [20] with default parameters, specifying only the input read (--nano-raw) and computational settings. Assembly and sequencing coverage metrics were calculated using BBMap v38.18 [21] and Qualimap v2.2.2 [22], respectively. Genome completeness and quality were assessed with BUSCO v5.2.2 using the eukaryota_odb10 (8 January 2024) and pleosporales_odb10 (8 January 2024) datasets asreferences [23]. Subsequently, in silico prediction of coding regions was performed using AUGUSTUS, previously trained with the genome of A. alternata (GenBank: GCA_036971565). Orthologous gene clusters were inferred using OrthoVenn3 (https://orthovenn3.bioinfotoolkits.net/home, accessed on 20 January 2025) and compared with the reference genomes of A. alternata (GenBank: GCA_036971565), A. arborescens (GenBank: GCA_004154835), and A. tenuissima (GenBank: GCA_041430775) using the OrthoMCL algorithm (e-value = 1 × 10−10), with all other parameters set to default. Phylogenetic relationships were inferred based on conserved single-copy orthologs using a maximum likelihood approach implemented within the OrthoVenn3 pipeline [24].
2.4. Pathogenicity Test
Pathogenicity tests were conducted on whole blueberry plants grown in pots under screen-house conditions to fulfill Koch’s postulates [8,9,25]. The experiment followed a completely randomized design using five healthy six-month-old blueberry plants (biological replicates), with six leaves per plant (technical replicates), totaling 30 leaves per treatment. The experiment was conducted twice. Inoculations were done using a conidial suspension (3.0 × 105 conidia mL−1) of A. alternata obtained from a 10-day-old single-spore culture grown on PDA, suspended in sterile distilled water containing 0.025% Tween 20. Before inoculation, four wounds per leaf were made using a sterile needle to ensure consistent infection. The conidial suspension was applied using sterile brushes to cover the entire leaf surface, while control leaves received sterile water containing 0.025% Tween 20. All leaves remained attached to living plants throughout the experiment. Plants were maintained at >70% relative humidity and 26 ± 2 °C for 14 days. Disease severity was evaluated at 0, 7, and 14 days post-inoculation (DPI) using paired measurements from five leaves per plant, with one additional leaf per plant reserved for re-isolation and documentation. Necrotic leaf area was quantified through manual analysis using the R package pliman [26]. Data were analyzed descriptively using means and standard deviations.
2.5. Re-Isolation and Validation
Following pathogenicity testing, re-isolation was attempted from both inoculated symptomatic tissues and mock-inoculated leaves using the same isolation procedure described in Section 2.1. Re-isolated organisms were subjected to morphological characterization following the same methodology described in Section 2.2, to confirm identity with the original isolate. Rapid molecular validation was conducted by amplification of the internal transcribed spacer (ITS) regions of nuclear ribosomal DNA using the primers ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) [27]. PCR reactions were performed in 25 μL volumes containing GoTaq® Green Master Mix (12.5 μL, 1× final concentration), upstream and downstream primers (0.5 μL each, 10 μM), DNA template (3 μL, 20 ng), and nuclease-free water to a final volume. Thermal cycling conditions included initial denaturation at 94 °C for 4 min, followed by 35 cycles of 94 °C for 30 s, 55 °C for 30 s, and 72 °C for 1 min, with final extension at 72 °C for 10 min. PCR products were sequenced using the Sanger method at Macrogen Co., Ltd. (Daejeon, Korea). Additionally, we carried out whole-genome sequencing and assembly of the re-isolated pathogen as previously described for multilocus validation (including regions gapdh, rpb2, tef1-α, and Alt-a1). Sequences were compared with those from the original isolate using BLASTn analysis (e-value < 1 × 10−25).
3. Results and Discussion
3.1. Field Symptoms and Disease Incidence
Leaf spot symptoms were characterized by brown, circular or irregular necrotic lesions with sunken, dark-brown centers and concentric rings, often surrounded by yellow halos and small black dots (Figure 1a). Lesions initially appeared as small spots (approximately 3–7 mm in diameter), predominantly on the abaxial leaf surface, and became evident on both surfaces as the disease progressed. Fruit rot symptoms were also observed in the field, characterized by flattening of mature fruits with a characteristic dry appearance, later covered by a mass of mycelium. Damage assessment at the field level revealed that 4.80% of fruits were affected during the first production cycle and 1.28% in the last production cycle of the year. Additionally, leaf spot incidence was recorded at 4.8%, a level comparable to previous reports of leaf spot incidence in South Korea [8].
3.2. Morphological Characterization
Five isolates were obtained that showed consistent morphology, with simple conidiophore architecture and short conidia chains (up to seven conidia) characteristic of A. alternata. Fungal colonies exhibited a dark olive color on the surface and a dark gray color on the reverse. The colonies had white filamentous margins, a circular form, and a velvety texture. They produced loose, aerial hyphae on the culture surfaces, with a cottony mycelial appearance (Figure 1b), consistent with the characteristics reported in other studies [28,29]. Single suberect or erect pale brown conidiophores were observed, ranging in size from 23.7 to 66.6 μm (mean: 36.5 ± 1.7 μm) and producing up to seven chained conidia (Figure 1c). Each conidium measured from 20.2 to 43.7 μm in length, with a mean of 28.6 μm ± 4.7 μm (n = 50), and 8.2 to 16.9 μm in width, with a mean of 11.7 μm ± 1.9 μm (n = 50). They contained three to six transverse septa and up to two longitudinal septa. Conidia were olivaceous or dull green-brown in color, ellipsoid to ovoid, often featuring a short conical or cylindrical beak (Figure 1d). The first conidium produced on a conidiophore was also narrowly elliptical, with three transverse septa.

Figure 1. Disease symptoms on V. corymbosum caused by A. alternata and its cultural and morphological characteristics. (a) Necrotic lesions on blueberry leaves taken from the field; (b) Colony morphology after 10 days of growth on PDA medium; (c) Conidiophores and conidia after 7 days of growth on PCA medium; (d) Microscopic view showing the diverse shapes of macroconidia (40×).
Based on these key features, the isolate was identified as A. alternata, distinct from closely related species in the Alternaria complex, according to Simmons’ taxonomic key [18]. Key distinguishing features included conidial dimensions consistent with the ellipsoid form, simple conidiophore architecture with short chains (up to 7 conidia) typical of Group J species rather than the longer unbranched chains (10–25+ conidia) characteristic of A. tenuissima (Group H), and typical beak morphology and septation patterns. The observed morphological traits are consistent with previous reports describing conidia with 3 to 6 transverse septa and dimensions ranging from 16 to 42 × 7 to 16 μm [7]. Likewise, other authors have described conidia as obclavate to elliptical [30]. However, differences were observed when comparing conidial dimensions with isolates from California, which showed average measurements of 22.04 μm in length and 8.7 μm in width [11]. Such variation may be influenced by geographic factors or local adaptation, emphasizing the importance of regional studies for understanding pathogen variability and its possible implications in disease management.
A. alternata has been confirmed as the causal agent of leaf spots on blueberries in South Korea and southern Poland [8,31]. Similarly, in California, 61.5% of Alternaria isolates from decayed blueberries were identified as A. alternata [11]. However, other studies have reported A. tenuissima as responsible for leaf spots in Western Australia and Argentina [9,32]. These results underscore the complexity of Alternaria spp. in different regions and highlight the need for accurate identification to develop precise, pathogen-targeted disease management strategies.
In other crops, Alternaria alternata has been reported causing brown spots on citrus crops across several Peruvian provinces [14]. Given that citrus crops are well-established in Peru, particularly in regions such as Lima [33], they could serve as a potential reservoir for A. alternata populations affecting blueberries. The widespread occurrence of A. alternata across Latin America, including citrus in Argentina, alfalfa in Brazil, quinoa in Colombia, hazelnut in Chile, broccoli in Ecuador, and weeds in Uruguay, underscores its adaptability to different environmental conditions and hosts [34,35,36,37,38,39]. The presence of many of these crops in Peru may have facilitated the expansion and adaptation of A. alternata to the more recently introduced blueberries.
3.3. Genome Characterization
DNA had high integrity and yielded 90.4 ng/µL before sequencing. Whole-genome sequencing yielded approximately 6 Gb of estimated bases (506,720 reads, average read length: 11.8 kb, N50 raw reads: 20.04 kb, mean quality score: Q10), and resulted in an assembled genome of 34.2 Mb with a coverage of ~166×, comprising 17 contigs (N50: 435,246 bp, L50: 18, largest contig: 6,834,098 bp) and a GC content of 49.93%. BUSCO analysis revealed 98.8% completeness (98.8% complete, 0.8% fragmented) and 95.3% completeness (94.8% complete, 0.5% fragmented) based on eukaryota_odb10 and pleosporales_odb10 databases, respectively. The genome size of A. alternata Per1 (34.2 Mb) and GC content fall within the expected range for this species, which has been reported to vary between 30–35 Mb and ~50%, respectively, across isolates from different host plants using various sequencing technologies, including Nanopore [6,40]. Protein-based gene cluster analysis was performed using the entire predicted proteome (all protein-coding genes) derived from the assembled genome of isolate Per1 (12,169 protein-coding genes). Orthologous gene cluster analysis identified 11,720 clusters and showed strong clustering with reference genomes of A. alternata (Figure 2).
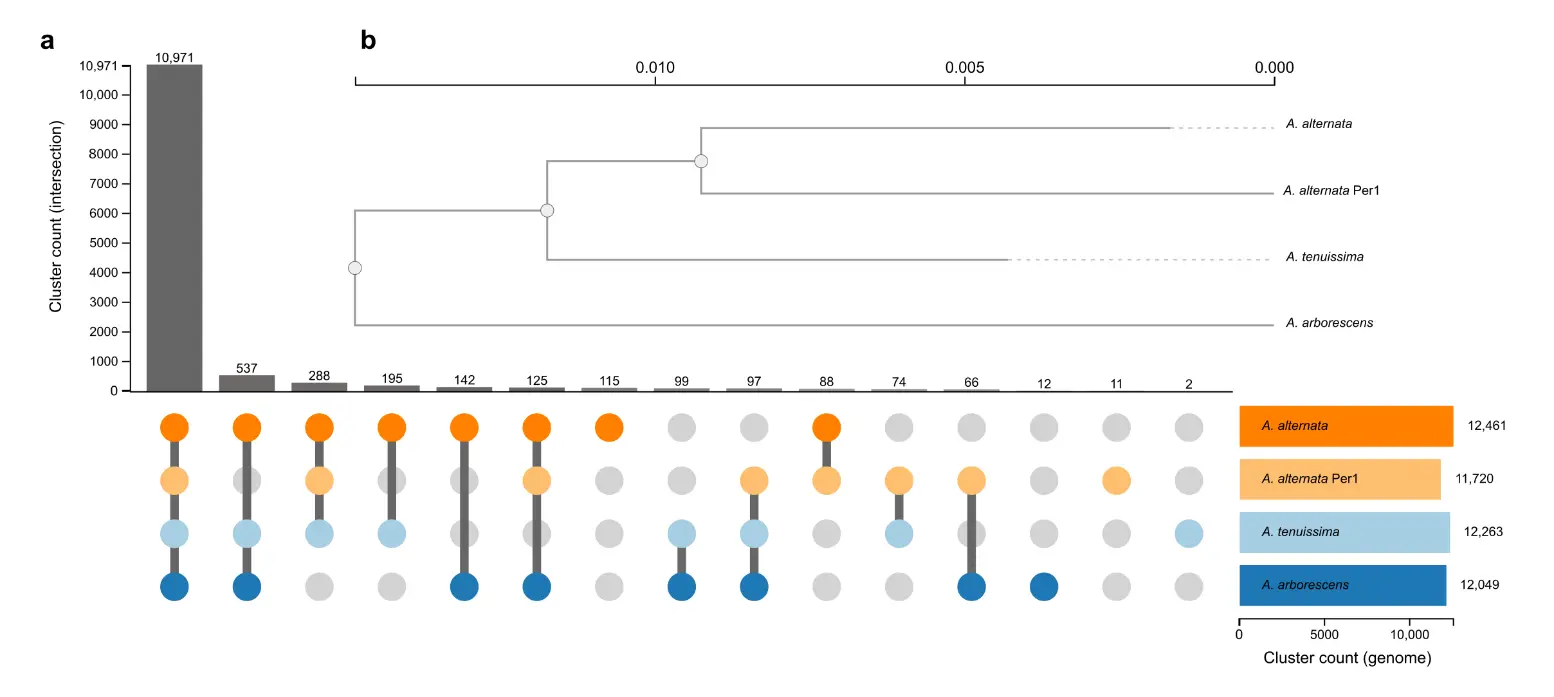
Figure 2. Phylogenetic relationships and orthologous cluster analysis of an Alternaria isolate from Peruvian blueberries. (a) Diagram showing distinct and shared orthologous clusters among A. alternata Per1 (orange dots) and the reference genomes of A. alternata, A. tenuissima, and A. arborescens available in the OrthoVenn3 web service. The bar graph on the left displays the count of orthologous clusters. (b) Maximum likelihood phylogenetic tree illustrating evolutionary relationships and genetic distances based on single-copy conserved genes. The tree was constructed using the JTT+CAT evolutionary model. This analysis provides insights into the genetic relationships between the Peruvian Alternaria isolate and other reference genomes.
3.4. Pathogenicity Test
Symptoms began to appear on inoculated leaves seven days post-inoculation, and by day 14, all inoculated leaves had developed necrotic spots identical to those observed in the field. In contrast, control plants remained asymptomatic (Figure 3). These results are consistent with previous studies reporting symptom onset between 7 and 8 days after inoculation [27,41]. Similarly, leaf spot symptoms observed after 15 days were comparable to those occurring under natural conditions [10]. Disease severity increased over time, with the affected leaf area ranging from 9.35 ± 0.71% at 7 days to 25.61 ± 6.2% at 14 days post-inoculation (Figure 3).

Figure 3. Results of the pathogenicity test showing A. alternata causing leaf spots 14 days post-inoculation. (a) No symptoms on a control leaf at 14 days post-inoculation; (b) Necrotic symptoms on a leaf 7 days post-inoculation; (c) Necrotic symptoms on a leaf 14 days post-inoculation. All leaves shown were attached to living, whole plants at the time of inoculation and symptom development.
The pathogen was successfully re-isolated from 86.7% of symptomatic tissues, confirming pathogenicity and fulfilling Koch’s postulates. Fusarium spp. were occasionally recovered as contaminants; however, contamination was controlled through colony purification and morphological differentiation, as Fusarium spp. colonies exhibited white coloration distinct from the characteristic olive-green appearance of A. alternata isolates.
Morphological and cultural comparisons confirmed the re-isolated organism as A. alternata, consistent with the inoculated pathogen (Figure 4). Comprehensive molecular validation included multilocus sequence analysis (extracted from whole-genome data) of ITS, gapdh, rpb2, tef1-α, Alt-a 1 locus, which confirmed the identity of the re-isolated fungus with that of the inoculated isolate (sequences for all loci shared 100% identity; GenBank nros: PV069777, PZ316321-PZ316324). These results fully satisfy Koch’s postulates through both morphological and robust molecular confirmation.

Figure 4. Morphological confirmation of re-isolated A. alternata. (a) Colony morphology after 10 days of growth on PDA medium; (b) Conidiophores and conidia after 7 days of growth on PCA medium; (c) Microscopic view of conidia (40×).
4. Conclusions
Alternaria alternata has been identified as the causal agent of leaf spot symptoms in blueberries in Peru. The combined use of morphological, pathogenicity, and whole-genome analyses enabled precise identification of the pathogen and confirmed its pathogenicity. These results establish a foundation for targeted disease management and inform the selection of specific control strategies.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this manuscript, the authors used Grammarly in order to improve the English language and writing clarity. After using this tool, the authors reviewed and edited the content as needed and take full responsibility for the content of the published article.
Acknowledgments
We extend our gratitude to David Pavel Casanova Núñez Melgar of INIA–Peru; Jesús Iván Chumpitaz Fernández at the Peruvian Institute of Agroexporters; and Jacobo Trujillo, Technical Support at INIA EEA Donoso–Huaral and the National Agrarian Health Service (SENASA), for their valuable assistance in molecular laboratory work, with special thanks to Arturo Olortegui Livia. We also acknowledge the technical support provided by Viviana Domínguez and Ronal Montes from CIAT.
Author Contributions
Conceptualization, R.V., J.M.P. and W.J.C.; Biological test, R.V., V.O. and J.M.P.; Formal Analysis, R.V., V.O., A.M.L. and A.G.-O.; Validation, R.V. and V.O.; Data Curation, V.O., A.M.L. and A.G.-O.; Writing—Original Draft Preparation, All authors; Writing—Review & Editing, W.J.C.; Supervision, J.M.P. and W.J.C.; Project Administration, R.V., I.B. and W.J.C.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
The genome assembly and associated data of isolate Per1 and re-isolate have been deposited in NCBI under accession JBMHEI000000000, BioProject PRJNA1189477, and BioSample SAMN44971619 and SAMN57370065 and SAMN57370065.
Funding
This research was funded by the Ministry of Agriculture of Peru (MINAGRI) through the Secretaría Técnica de Cooperación del CGIAR (STC-CGIAR).
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
- Bell MG, Zimmerer KS, Tinoco OSR. Following the water uphill? The spread of blueberry cultivation to the mountains of Áncash, Peru. J. Lat. Am. Geogr. 2023, 22, 156–166. DOI:10.1353/lag.2023.a899558 [Google Scholar]
- FAO. FAOSTAT: Crops and Livestock Products. Available online: https://www.fao.org/faostat/en/#data/QCL/visualize (accessed on 10 January 2026).
- Montes Ninaquispe JC, Arbulú Ballesteros MA, Cruz Salinas LE, García Juárez HD, Farfán Chilicaus GC, Martel Acosta R, et al. A strategy for the sustainability of Peru’s blueberry exports: Diversification and competitiveness. Sustainability 2024, 16, 6606. DOI:10.3390/su16156606 [Google Scholar]
- Deka B, Upamanya GK, Baruah RMB. Eco-friendly management of Alternaria blight disease: A review. Asian J. Adv. Agric. Res. 2024, 24, 34–43. DOI:10.9734/ajaar/2024/v24i10554 [Google Scholar]
- Cong K, Cheng J, Li T, Wu C, Fan G, Li X, et al. Alternaria in berry: Effects on its quality, pathogenic mechanisms, and potential control strategies. Plant Pathol. 2024, 73, 1317–1333. DOI:10.1111/ppa.13916 [Google Scholar]
- Gai Y, Ma H, Chen Y, Li L, Cao Y, Wang M, et al. Chromosome-scale genome sequence of Alternaria alternata causing citrus brown spot. Mol. Plant-Microbe Interact. 2021, 34, 726–732. DOI:10.1094/MPMI-10-20-0278-SC [Google Scholar]
- Luan YS, Feng L, Xia XY, An LJ. First report of Alternaria tenuissima on blueberry in China. Plant Dis. 2007, 91, 464. DOI:10.1094/PDIS-91-4-0464A [Google Scholar]
- Kwon JH, Kang DW, Cheon MG, Kim J. First report of Alternaria leaf spot on highbush blueberry in South Korea. Plant Dis. 2014, 98, 1434. DOI:10.1094/PDIS-04-14-0344-PDN [Google Scholar]
- You MP, Lanoiselet V, Wang CP, Barbetti MJ. First report of Alternaria tenuissima on blueberry in Western Australia. Plant Dis. 2014, 98, 423. DOI:10.1094/PDIS-07-13-0737-PDN [Google Scholar]
- Greco M, Patriarca A, Terminiello L, Pose G. Toxigenic Alternaria species from Argentinean blueberries. Int. J. Food Microbiol. 2012, 154, 187–191. DOI:10.1016/j.ijfoodmicro.2012.01.004 [Google Scholar]
- Zhu XQ, Xiao CL. Characterization of Alternaria species associated with fruit rot of blueberry in California. Phytopathology 2015, 105, 1555–1567. DOI:10.1094/PHYTO-05-15-0122-R [Google Scholar]
- Neugebauer KA, Mattupalli C, Hu M, Oliver JE, VanderWeide J, Lu Y, et al. Managing fruit rot diseases of blueberry. Front. Plant Sci. 2024, 15, 1428769. DOI:10.3389/fpls.2024.1428769 [Google Scholar]
- Salotti I, Giorni P, Battilani P. Biology, ecology, and epidemiology of Alternaria species affecting tomato. Front. Plant Sci. 2024, 15, 1430965. DOI:10.3389/fpls.2024.1430965 [Google Scholar]
- Marín JE, Fernández HS, Peres NAR, Andrew M, Peever TL, Timmer LW. First report of citrus brown spot in Peru. Plant Dis. 2007, 90, 686. DOI:10.1094/PD-90-0686C [Google Scholar]
- Schmey T, Auxier B, Krebs S, Patneedi SK, Ahmad F, Habig M, et al. Evolutionary divergence in sympatric populations of the fungal pathogen Alternaria alternata across wild tomato hosts. bioRxiv 2025, 2025-10. DOI:10.1101/2025.10.01.678058 [Google Scholar]
- Delp BR, Stowell LJ, Marois JJ. Evaluation of field sampling techniques for estimation of disease incidence. Phytopathology 1986, 76, 1299–1305. DOI:10.1094/Phyto-76-1299 [Google Scholar]
- Dai YF, Wu XM, Wang HC, Li WH, Cai LT, Li JX, et al. Spatio-temporal variation in the phyllospheric microbial biodiversity of alternaria alternata-infected tobacco foliage. Front. Microbiol. 2022, 13, 920109. DOI:10.3389/fmicb.2022.920109 [Google Scholar]
- Simmons EG. Alternaria: An Identification Manual; CBS Fungal Biodiversity Centre, Ed.; ASM Press: Washington, DC, USA, 2007. [Google Scholar]
- Gil-Ordóñez A, Pardo JM, Sheat S, Xaiyavong K, Leiva AM, Arinaitwe W, et al. Isolation, genome analysis, and tissue localization of Ceratobasidium theobromae. Sci. Rep. 2024, 14, 18139. DOI:10.1038/s41598-024-69061-8 [Google Scholar]
- Kolmogorov M, Yuan J, Lin Y, Pevzner PA. Assembly of long, error-prone reads using repeat graphs. Nat. Biotechnol. 2019, 37, 540–546. DOI:10.1038/s41587-019-0072-8 [Google Scholar]
- Bushnell B. BBMap: A Fast, Accurate, Splice-Aware Aligner. 2014. Available online: https://escholarship.org/uc/item/1h3515gn (accessed on 4 February 2025).
- García-Alcalde F, Okonechnikov K, Carbonell J, Cruz LM, Götz S, Tarazona S, et al. Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 2012, 28, 2678–2679. DOI:10.1093/bioinformatics/bts503 [Google Scholar]
- Simão FA, Waterhouse RM, Ioannidis P, Kriventseva EV, Zdobnov EM. BUSCO: Assessing genome assembly completeness. Bioinformatics 2015, 31, 3210–3212. DOI:10.1093/bioinformatics/btv351 [Google Scholar]
- Sun J, Lu F, Luo Y, Bie L, Xu L, Wang Y. OrthoVenn3: Orthologous data visualization platform. Nucleic Acids Res. 2023, 51, W397–W403. DOI:10.1093/nar/gkad313 [Google Scholar]
- Haq IU, Ijaz S, Khan IA. A journey from Koch’s postulates to molecular systems biology. In Phytomycology and Molecular Biology of Plant–Pathogen Interactions; CRC Press: Boca Raton, FL, USA, 2022; pp. 1–25. [Google Scholar]
- Olivoto T. Lights, camera, pliman! An R package for plant image analysis. Methods Ecol. Evol. 2022, 13, 789–798. DOI:10.1111/2041-210X.13803 [Google Scholar]
- Glass NL, Donaldson GC. Development of primer sets for conserved genes in filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. DOI:10.1128/aem.61.4.1323-1330.1995 [Google Scholar]
- Hu LJ, Li Y, Xuan JP, Gu YH, Li DL, Mo ZH, et al. A new leaf spot disease caused by Alternaria alternata on Atractylodes lancea. Plant Dis. 2024, 108, 3658. DOI:10.1094/PDIS-07-24-1349-PDN [Google Scholar]
- Liu Z, Zhang R, Sun D, Tuerdi M, Zhu T, Chen X. Alternaria alternata causing brown spot on walnut in China. Plant Dis. 2023, 107, 2224. DOI:10.1094/PDIS-07-22-1536-PDN [Google Scholar]
- Olmez S, Mutlu N, Kaba A. First report of Alternaria alternata on cotton in Türkiye. Plant Dis. 2023, 107, 3296. DOI:10.1094/PDIS-04-23-0724-PDN [Google Scholar]
- Nadziakiewicz M, Kurzawińska H, Mazur S, Tekielska D. Alternaria alternata—The main causal agent of disease symptoms in juniper, rose, yew and highbush blueberry in nurseries in southern Poland. Folia Hortic. 2018, 30, 15–25. DOI:10.2478/fhort-2018-0002 [Google Scholar]
- Wright ER, Rivera MC, Esperón J, Cheheid A, Rodríguez Codazzi A. Alternaria diseases of blueberry in Argentina. Plant Dis. 2004, 88, 1383. DOI:10.1094/PDIS.2004.88.12.1383B [Google Scholar]
- Ruiz Camacho W, Julca Otiniano A. Comportamiento del cultivo de naranja (Citrus sinemis) variedad Valencia en dos zonas agroecológicas de la provincia de Chanchamayo, Junín, Perú. Idesia 2022, 40, 89–94. DOI:10.4067/S0718-34292022000300089 [Google Scholar]
- Durán P, Barra PJ, Mora MLL, Morina F, Viscardi S, Meriño-Gergichevich C. First report of fungal complex causing grey necrosis of hazelnut in Chile. New Dis. Rep. 2020, 42, 7. DOI:10.5197/j.2044-0588.2020.042.007 [Google Scholar]
- Fonseca-Guerra IR, Beltrán Pineda ME, Benavides Rozo ME. Characterization of Alternaria alternata and Alternaria scrophulariae brown spot in Colombian quinoa. J. Fungi 2023, 9, 947. DOI:10.3390/jof9090947 [Google Scholar]
- Kaspary TE, García MA, Stewart S, Casaroto G, Ramos RF, Bellé C. First report of Alternaria alternata causing leaf spot on Rumex crispus in Uruguay. Plant Dis. 2019, 103, 2139. DOI:10.1094/PDIS-03-19-0590-PDN [Google Scholar]
- Miguel DA, Mariana RA, José AM, Gerarda BPS, Raquel SC, José ARS, et al. First report in Southern Brazil of Alternaria alternata causing Alternaria leaf spot in alfalfa (Medicago sativa). Afr. J. Agric. Res. 2015, 10, 491–493. DOI:10.5897/AJAR2014.9205 [Google Scholar]
- Peres NAR, Agostini JP, Timmer LW. Outbreaks of citrus brown spot in Brazil and Argentina. Plant Dis. 2003, 87, 750. DOI:10.1094/PDIS.2003.87.6.750C [Google Scholar]
- Ramirez-Villacis DX, Barriga-Medina N, Llerena-Llerena S, Pazmino-Guevara C, Leon-Reyes A. First report of Alternaria alternata on broccoli in Ecuador. Plant Dis. 2023, 107, 2866. DOI:10.1094/PDIS-03-23-0598-PDN [Google Scholar]
- Dettman JR, Eggertson Q. Phylogenomic analyses of Alternaria section Alternaria. Mycologia 2021, 113, 1218–1232. DOI:10.1080/00275514.2021.1950456 [Google Scholar]
- Shahid AA, Iftikhar S, Nawaz K, Anwar W, Ali M. First report of Alternaria alternata causing brown leaf spot of potato in Pakistan. J. Plant Pathol. 2017, 99, 300. Available online: https://www.cabidigitallibrary.org/doi/full/10.5555/20183088575 (accessed on 4 February 2025).