The Diverse Landscape of Cancer Genomics in Africa

The Diverse Landscape of Cancer Genomics in Africa
Karla Christina Sousa Silva
1
 Chiara Lea Manon Foret
1
Martha Wium
1
Maria Stella Zerbini
2
Stefano Cacciatore
3
Lara Donaldson
4
Luiz Fernando Zerbini
1,*
Chiara Lea Manon Foret
1
Martha Wium
1
Maria Stella Zerbini
2
Stefano Cacciatore
3
Lara Donaldson
4
Luiz Fernando Zerbini
1,*
Received: 21 February 2026 Revised: 17 April 2026 Accepted: 24 April 2026 Published: 14 May 2026

© 2026 The authors. This is an open access article under the Creative Commons Attribution 4.0 International License (https://creativecommons.org/licenses/by/4.0/).
1. Introduction
Cancer burden is unevenly distributed across global populations, with individuals of African ancestry experiencing disproportionately higher mortality rates, earlier age of onset, and a greater prevalence of aggressive disease phenotypes [1,2]. While structural determinants such as socio-economic inequality, barriers to healthcare access, and delayed diagnosis undeniably shape cancer outcomes, accumulating evidence suggests that genetic ancestry makeup also plays a substantive role in cancer susceptibility, progression, and therapeutic response [3,4,5]. The contribution of African genetic diversity to cancer risk remains insufficiently characterised [6], notwithstanding that Africa harbours the greatest human genetic diversity worldwide. The average African genome contains approximately 25% more genetic variation than genomes from non-African populations and reflects exceptionally complex population structure [7,8,9].
In this context, African Indigenous populations, whose genetic lineages have contributed substantially to the ancestry of contemporary African populations across the world, remain markedly underrepresented in cancer genomics and population health research [10,11]. This underrepresentation limits understanding of ancestry-specific disease risk and constrains the generalisability of findings derived largely from non-African or urbanised African cohorts. As a result, the biological relevance of African genetic diversity for cancer risk and tumour behaviour remains insufficiently characterized and analysed.
Indigenous peoples are defined by the United Nations as populations with historical continuity to pre-colonial or pre-settlement societies; cultural, linguistic, and social distinctiveness from dominant populations; and sustained relationships with ancestral territories and natural resources [12]. In this review, the term “Indigenous” is used for populations that align with the United Nations descriptive framing cited above or are described as such in the source literature; where this is unclear, we use the specific community’s name or ethnolinguistic designation and avoid assigning Indigenous status. The general geographic distribution of the Indigenous and ethnolinguistic populations of Africa, as discussed in this narrative review, is shown in Figure 1.
An estimated 476 million Indigenous people live in over 90 countries of the world, including Africa, collectively constituting about 6.2% of the world’s population [13]. In contrast to global estimates, comprehensive population statistics for Indigenous people in Africa are limited, with available data largely restricted to country or group-specific estimates [14]. Within Africa, Indigenous populations encompass diverse hunter-gatherer, pastoralist, and small-scale agrarian societies whose demographic histories often predate major population expansions, including Bantu-speaking migrations [15,16,17]. These populations retain genetic lineages that reflect some of the earliest divergences in human evolutionary history and continue to shape the genetic architecture not only of modern African populations but also of the Homo sapiens species as a whole [15,18,19]. Genomic reconstructions of African population history demonstrate that Indigenous groups are related through deep ancestral lineages that diverged tens of thousands of years ago, with population structure broadly aligning with geographic regions across North, West, Central, East, and Southern Africa [20,21]. Subsequent migration events, such as pastoralist expansions and later agricultural dispersals, facilitated varying degrees of genetic exchange among groups, while many Indigenous populations maintained relative genetic distinctiveness due to geographic, ecological, or cultural continuity [21,22,23]. Contemporary African populations therefore represent complex mosaics of Indigenous and later-arriving ancestries, with important implications for patterns of disease susceptibility and health outcomes. Studying the genetic makeup of African Indigenous populations is therefore critical. These populations provide unique opportunities to identify genetic variants that may influence cancer susceptibility in contemporary African populations, uncover population-specific risk factors for both common and rare diseases, and elucidate gene-environment interactions shaped by long-standing subsistence practices [18,24,25].
Beyond population-specific insights, inclusion of Indigenous populations has broader translational implications. Because African populations harbor the greatest human genetic diversity and retain genetic diversity that reflects early human population divergence [7,8,9], discoveries made in these groups can strengthen the foundational architecture of disease gene discovery and phenotypes. This dual benefit enables both the development of tailored precision medicine strategies for specific communities and the generation of genomic insights that improve risk prediction and therapeutic approaches across multiple populations [26,27,28].
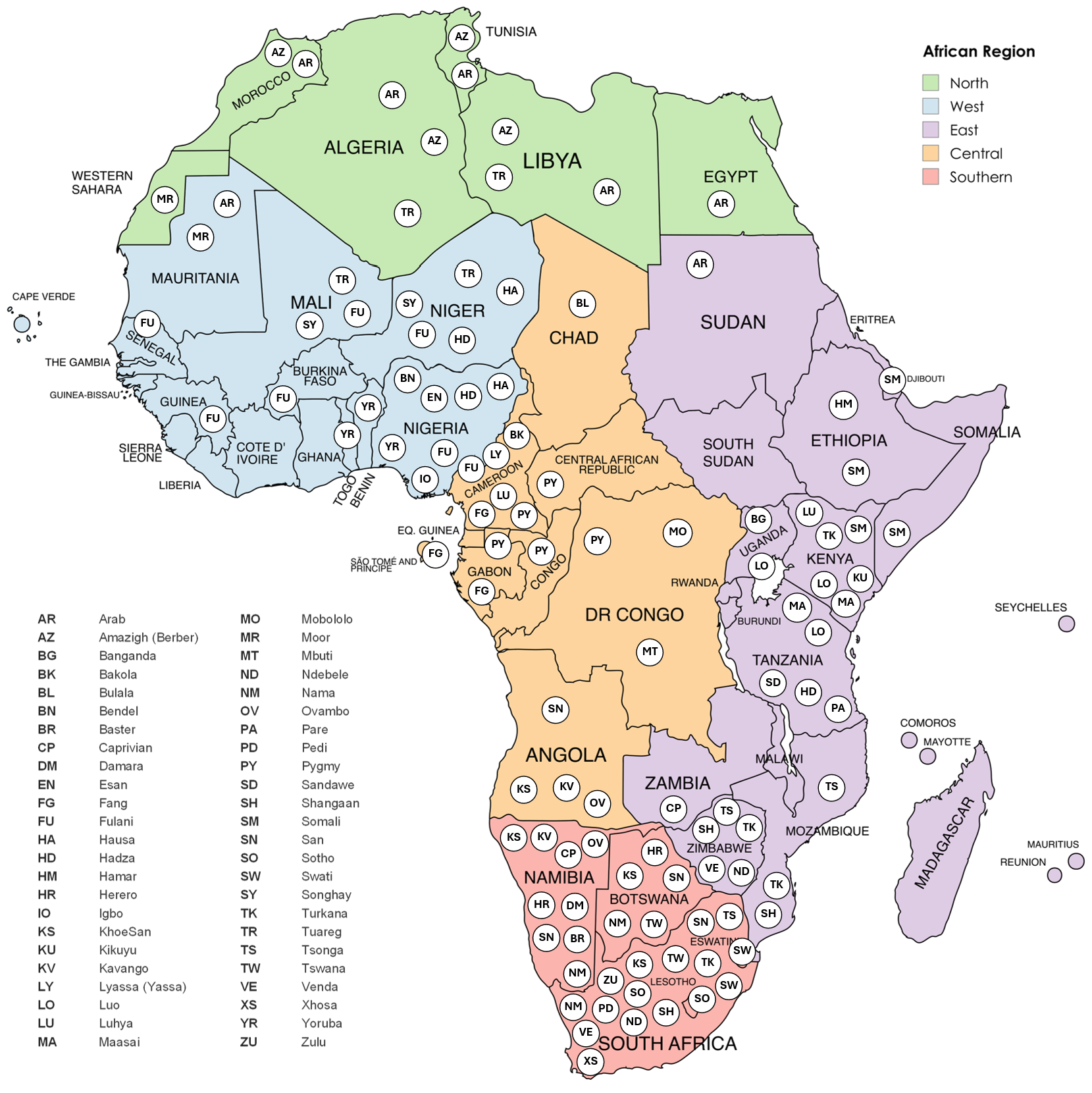
Figure 1. Map of selected African population groups discussed in the review. This map illustrates the spatial distribution of selected population groups across the African continent, overlaid on the five African regions defined by the United Nations (North, West, East, Central, and Southern Africa). Each circle marks the country in which a population group is situated, identified by standardized abbreviations (see legend). The map of Africa was created with MapChart.
By integrating anthropological, evolutionary, and biomedical perspectives, this narrative literature review situates health disparities among African Indigenous populations within the broader context of African population history and contemporary cancer burden. It examines regional patterns of African Indigenous and non-Indigenous populations’ distribution and associated health disparities across Africa, focusing on the six most prevalent types of cancer on the continent according to Global Cancer Observatory (GLOBOCAN) [2], as illustrated in Figure 2 and Table 1. Understanding the genomic landscape through a more inclusive and population-oriented approach is essential for developing health solutions that are more effective, context-appropriate, and aligned with the realities of the continent, ultimately contributing to greater health equity.
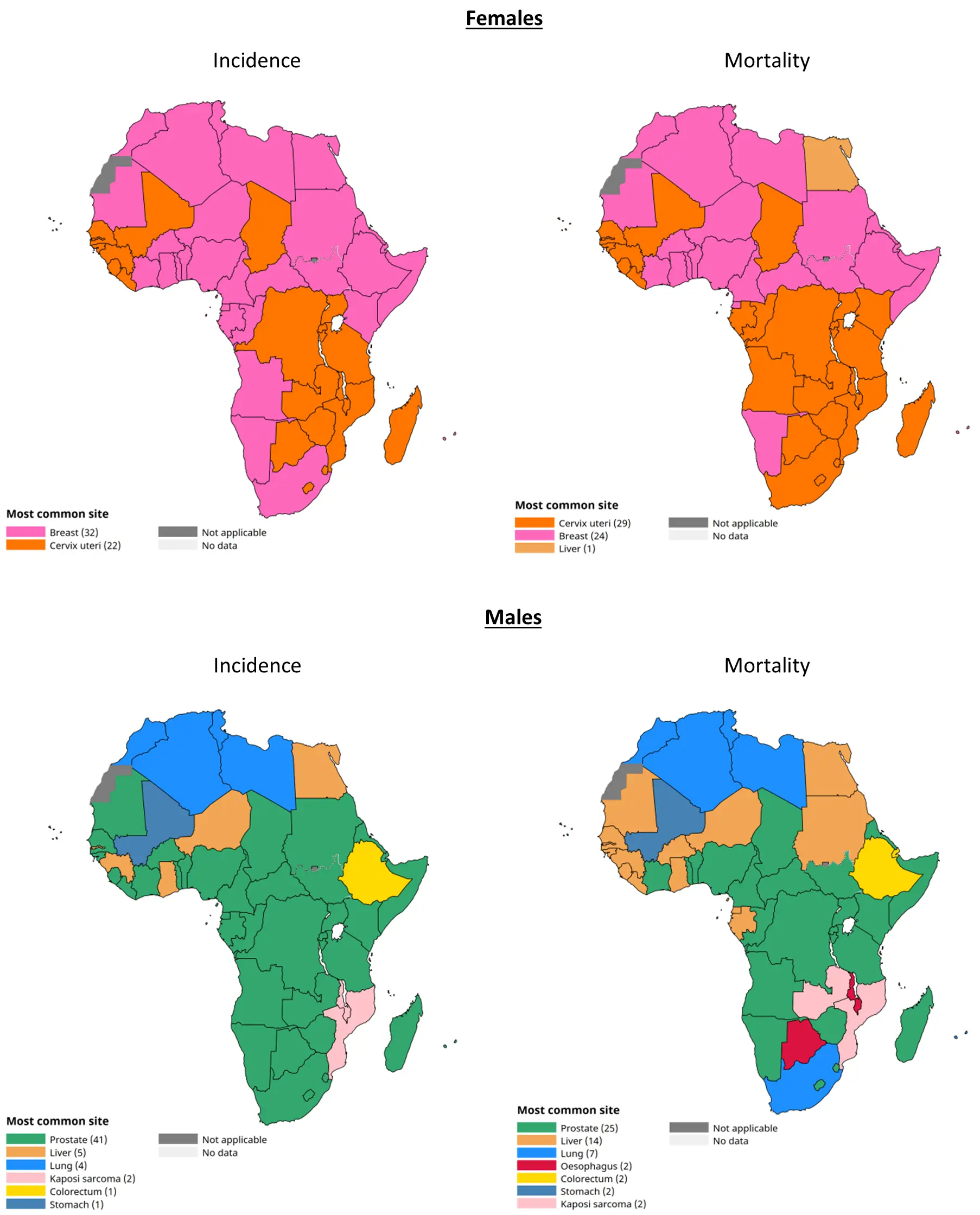
Table 1. Incidence and mortality rates for the most common cancers in Africa in males and females in 2022, according to GLOBOCAN [2].
|
Central Africa |
East Africa |
North Africa |
Southern Africa |
West Africa |
Africa |
World |
||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
Incidence (ASR) |
Mortality (ASR) |
|
|
Breast |
||||||||||||||
|
Female |
30.0 |
16.8 |
31.9 |
17.6 |
53.2 |
19.0 |
46.2 |
16.6 |
41.6 |
22.3 |
40.5 |
19.2 |
46.8 |
12.7 |
|
Cervical |
||||||||||||||
|
Female |
31.1 |
22.9 |
40.4 |
28.9 |
6.5 |
3.8 |
34.9 |
20.4 |
26.7 |
16.3 |
26.4 |
17.6 |
14.1 |
7.1 |
|
Prostate |
||||||||||||||
|
Male |
44.2 |
27.2 |
28.0 |
16.8 |
16.1 |
6.9 |
59.9 |
29.7 |
36.9 |
23.5 |
30.3 |
17.3 |
29.4 |
7.3 |
|
Liver |
||||||||||||||
|
Male |
9.8 |
9.5 |
5.9 |
5.7 |
18.7 |
18.0 |
6.1 |
5.7 |
9.9 |
9.6 |
11.1 |
10.7 |
12.7 |
10.9 |
|
Female |
5.0 |
4.9 |
3.9 |
3.8 |
10.1 |
9.6 |
3.7 |
3.4 |
6.0 |
5.9 |
6.2 |
6.0 |
4.8 |
4.1 |
|
Lung |
||||||||||||||
|
Male |
3.2 |
3.0 |
3.9 |
3.7 |
20.6 |
18.6 |
25.7 |
23.7 |
2.6 |
2.5 |
9.9 |
9.1 |
32.1 |
24.8 |
|
Female |
1.6 |
1.5 |
2.7 |
2.5 |
3.8 |
3.4 |
10.4 |
9.8 |
1.6 |
1.5 |
3.2 |
3.0 |
16.2 |
9.8 |
|
Colorectal |
||||||||||||||
|
Male |
5.9 |
4.3 |
8.5 |
6.1 |
11.0 |
6.2 |
16.1 |
10.2 |
7.4 |
5.6 |
9.1 |
6.1 |
21.9 |
9.9 |
|
Female |
5.2 |
3.8 |
7.2 |
5.2 |
9.6 |
5.3 |
10.7 |
7.2 |
5.8 |
4.4 |
7.5 |
5.1 |
15.2 |
6.5 |
ASR: age-standardized incidence per 100,000 people. Source: International Agency for Research on Cancer (IARC), GLOBOCAN, [accessed on 18 February 2026].
2. Prostate Cancer
Numerous publications show that, in African men, prostate cancer (PCa) is the most devastating cancer in terms of incidence and mortality [29,30,31]. PCa diagnosis typically involves a Prostate-Specific Antigen (PSA) blood test and a digital rectal examination (DRE), often conducted for men over 50 years of age. PSA is a serum biomarker produced by the prostate gland and, in healthy men, is ≤4 ng/mL, although it lacks specificity as elevated levels can indicate non-cancerous conditions or infections. Well-established non-modifiable risk factors that are thought to contribute to PCa development include: advanced age (diagnosis in men aged <40 years is rare, with incidence steadily rising thereafter), positive family history, genetic predisposition (e.g., BRCA1, ERCC6, ARHGAP21, and ADAMTSL3 mutations [32]), and ethnicity, with the latter two being somewhat interlinked [33]. It is of great concern that PCa mortality disproportionately affects African men despite having a similar, or in some cases lower, incidence to European men. This disparity is characterised either by earlier age at diagnosis, presentation of advanced-stage disease, a lack of access to appropriate healthcare, and genetic susceptibility or hereditary predisposition [34].
As of 2022, Northern Africa has the lowest age-standardised rate (ASR) of PCa incidence and mortality in Africa (Table 1). In Morocco, however, the burden of PCa is the highest in the region. It ranks as the most frequently diagnosed cancer (23.6 per 100,000) and the second leading cause of cancer-related deaths in men (10.8 per 100,000) [2,35]. Although prostatic adenocarcinoma histology is predominant at presentation, the prevalence of advanced disease in the population (29.2% of cases) is considered lower than that in other African regions and affects mainly older individuals [36]. Therefore, the high rate of mortality has been attributed to delayed diagnosis, absence of a PCa screening program, smoking, and socioeconomic status rather than clinical or genomic characteristics [36]. Genetic studies have found that North Africa is somewhat unique in that there exists minimal differentiation between the Arab and Indigenous Amazigh populations in the region, due to the high degree of inter-population admixture and their common Neolithic Berber ancestry [37,38]. Indeed, a small study of a familial PCa cohort from Morocco suggests that there are seemingly no discernible differences in PCa presentation for the Amazigh and the remainder of the population. At presentation, both the Amazigh and their counterparts were on average 70 years of age, had similar median PSA levels (45 ng/mL and 34 ng/mL, respectively), and had a high proportion classified as having intermediate disease (Gleason score ≤ 3+4) [39]. However, since this study was limited to familial PCa and concerned a cohort of just 30 participants, these findings may not be broadly representative. Interestingly, this homogeneity of PCa presentation amongst Indigenous and non-Indigenous peoples was mirrored in Sudan, whose population faces both the lowest incidence (11.9 per 100,000 men) and the second lowest mortality rates (6.1 per 100,000) of the disease in the region [2]. PCa burden and aggressiveness were relatively similar amongst the various ethnolinguistic groups in Sudan, including Niger-Congo, Nilo-Saharan, and Afro-Asiatic speaking populations. The majority (85.4%) of patients presented at the age of 70 and above with advanced (stage III and IV) disease [34,40,41]. The Afro-Asiatic-speaking population presented with a lower median PSA level of 52 ng/mL compared to the overall Sudanese population median of 100 ng/mL, but this was not significantly different [34,40]. One study, however, found that the incidence of perineural invasion in prostatic tissue was elevated among Afro-Asiatic patients, which tends to be more prevalent in advanced-stage tumors, and it is often associated with unfavourable outcomes [34].
PCa is the most common cancer in Eastern African men, with incidence and mortality ASRs of 28.0 and 16.8 per 100,000, respectively (Table 1). At present, there are no epidemiological studies that describe the burden or presentation of PCa in East African Indigenous populations. However, a few genomics studies have identified PCa susceptibility loci in selected East African populations. One such study was performed using genomics data from two Kenyan populations: the Luhya and Maasai [42]. This study found that Maasai had positive selection for a deleterious missense variant, rs773663, in the gene BAZ2A, the overexpression of which has been associated with PCa recurrence [43]. A second genomics study sought to investigate the contribution of susceptibility loci to PCa disparities across African populations. They found that several East African populations had an intermediate PCa risk in comparison to that of non-African populations [44]. In particular, the Pare and Luhya of Kenya, as well as the Hadza of Tanzania and the Hamar of Ethiopia, had the highest predicted risk of PCa, likely due to the predominance of the known PCa susceptibility loci rs9623117 (TNRC6B gene, responsible for mRNA degradation mediated by miRNA) and rs7584330 (MLPH gene, part of the androgen receptor binding site) [44,45,46].
This same genomics study found that the genetic risk of PCa is elevated for West African populations [44]. Like in Eastern Africa, PCa is the leading cause of cancer incidence and mortality in the region, with respective ASRs of 33.1 and 20.2 per 100,000 (Table 1). The vast majority of PCa epidemiological data in Western Africa comes from Nigeria. Nigeria bears the heaviest burden of the disease among the sixteen countries that comprise Western Africa. In fact, known PCa-associated SNPs (rs9623117 and rs11672691) with the highest West African risk scores are particularly concentrated in the Esan and Yoruba populations of Nigeria [44]. Nigerian men are typically diagnosed between the ages of 55 and 71 years [47]. Advanced or metastatic disease (Gleason scores ≥ 4+3) at presentation is common, and PSA levels > 100 ng/mL are correlated with a higher burden of disease [47]. Interestingly, although the PSA levels at diagnosis amongst Igbo men were far lower than those of the general population (1.20–33.9 ng/mL and 0.5–760 ng/mg, respectively), they tended to present earlier with similar stages of PCa [47,48]. For the Igbo, advanced disease (Gleason score ≥ 4+3) was most common at presentation and was accompanied by chronic prostatitis in one third of all cases [48]. Advanced or late-stage disease in relatively young men usually suggests that there may be biological/environmental predisposition or heritable factors at play. This may also be the case with other Nigerian ethnic population groups, as the incidence in Hausa and Yoruba men was found to be the highest in the 50–60 year age group compared to that of 60–69 years for Nigeria overall [47,48,49]. However, since no studies have been reported on the clinical features of these groups, this is purely conjecture.
Southern Africa experiences the most aggressive burden of PCa in Africa, where it ranks as having the highest incidence and mortality of the disease. The ASR of incidence and mortality is nearly four times that of Northern Africa (Table 1) [2]. The ethnolinguistic landscape of the region comprises two major branches: the Indigenous KhoeSan and various Bantu-speaking peoples. For example, Southern Africans represent a unique ancestry of predominantly Bantu-speaking groups (including the Ndebele, Pedi, Shangaan, Sotho, Swati, Tswana, Tsonga, Venda, Xhosa, and Zulu), with contributing KhoeSan heritage [29]. In Namibia, the Tswana, Herero, and Baster (descended from the Cape Coloured and KhoeSan of South Africa) are more likely to develop PCa than all other groups [50], and the Nguni, Tsonga, Venda, and KhoeSan of South Africa were more likely than men of European descent to be diagnosed with PCa [29]. By way of explanation, studies have found that Southern African ancestry is linked to high-risk PCa (Gleason score ≥ 8) and that this association is directly influenced by an increase in the mean KhoeSan genetic contribution. Indeed, four chromosomal regions were identified as high-risk loci within ancient KhoeSan genomes—2p11.2, 3p14, 8q23, and 22q13.2—of which variants of 2p11.2 and 22q13.2 had previously been associated with PCa in Japanese, Australian, and various European populations [51]. This suggests that modern humans’ earliest ancestors may already have carried genomic signatures for high-risk PCa. A similar genomics study in Bantu-speaking populations in South Africa found that they carried genetic variants significantly associated with clinical characteristics of aggressive PCa [52]. This may partly provide an explanation as to why the Bantu-speaking Nguni, Tsonga, and Venda men from South Africa are at a higher risk of PCa than all other groups, with one third being diagnosed with advanced PCa (Gleason score ≥ 8) at presentation [29,52].
Despite the high incidence of PCa in the Central African region, the second highest in Africa (Table 1), data regarding Indigenous populations have proven scarce. The Fulani from Cameroon have the highest median genetic risk scores compared to all other Eastern, Western, or Non-African (i.e., Asian, European, and South American) population groups investigated, closely followed by other groups such as the Lemande, Bakola, Lyassa, and Mobololo of Cameroon, and the Bulala of Chad [44]. In Gabon, a study investigating the relationship between Angiotensin-Converting Enzyme (ACE) gene insertion/deletion polymorphism and disease onset found that one-third of individuals with PCa carry the ACE deletion polymorphism [53]. The ACE enzyme is synthesized by the prostate, and the double deletion genotype of this gene has been previously associated with an advanced stage of PCa [54]. This deletion polymorphism was significantly more frequent among the Baka (Pygmy; 0.862) and the Fang (Bantu-speaking; 0.687) [54].
The PCa burden and biology among African populations, including several Indigenous populations, reflect the continent’s evolutionary history and complex demographic structure. While in some regions clinical homogeneity is observed between Indigenous and non-Indigenous groups, genomic analyses increasingly reveal enrichment of distinct susceptibility loci within particular ancestral backgrounds. Importantly, several of the populations discussed in this review present with advanced-stage PCa, yet the relative contributions of inherited susceptibility, long-standing gene–environment interactions, and structural inequities in healthcare access remain insufficiently addressed.
3. Breast Cancer
Breast cancer (BrCa) is the most common female malignancy in Africa, being the cancer with the first or second highest incidence and/or mortality in most African countries, despite having lower incidence rates than in the rest of the world (Table 1). Relative to BrCa in women of European ancestry, the disease in Indigenous women in Africa is typically characterized by a younger age, an advanced stage, and a higher grade at presentation [55]. Although this is certainly exacerbated by factors such as cultural beliefs, socio-economic status, the lack of screening programs, and deficiencies in healthcare infrastructure, genetic susceptibility and heritable factors also play a role in BrCa development and prognosis [56,57,58]. BrCa is molecularly classified by oestrogen (ER), progesterone (PR), and human epidermal growth factor-2 (HER2) receptor status, as they inform not only BrCa aetiology and incidence trends, but are also a major determinant of treatment options, disease outcomes, and survival [54]. This is often further delineated into four molecular subtypes based on the receptor’s expression: luminal A (Lum A; ER+ and/or PR+, HER2−), luminal B (Lum B; ER+, PR+ and/or HER2+), HER2-enriched (HER2+), and triple-negative breast cancer (TNBC), which lacks expression of all three receptors [59]. The Lum A and Lum B molecular subtypes are associated with a better prognosis and long-term survival estimated at 80–85% for 5 years, while the TNBC, characterized by aggressiveness and higher resistance to therapies, has the highest mortality rate of 40% within a 5-year diagnostic period [58]. The luminal subtypes of BrCa, particularly those with ER+ expression, account for 45–65% of BrCa cases worldwide [59].
Although dominated by the luminal subtypes of BrCa, which typically correspond to better prognosis, Northern Africa has the highest incidence and the second highest mortality rates of the disease in Africa (Table 1). And yet, studies relating to the burden of disease in the region’s Indigenous populations remain scarce and lack clinical or genetic etiological data. At present, only the Indigenous Amazigh people of Morocco have been included in BrCa epidemiological studies. Although the composition of these studies’ cohorts closely reflected that of the rest of the Moroccan population, with approximately 30% identifying as Amazigh, they could not provide any conclusion about the distribution or genetic contribution of BrCa in the Indigenous Amazigh people [60,61,62].
Within Eastern Africa, the majority of studies on BrCa distribution and genetic contributions among Indigenous groups come from Kenya. After Ethiopia, Kenya faces the highest incidence of BrCa in the region, and it is the leading cause of cancer-related deaths in Kenyan women (ASR mortality of 19.6 per 100,000) [2]. Kenya’s population is often divided into three distinct ethnolinguistic and cultural groups: the Bantu-speakers (e.g., the Kikuyu and Luhya), the Nilotes (e.g., the Luo and the Indigenous Massai and Turkana), and the Cushites (e.g., the Indigenous Somali) population groups. When BrCa characteristics among these three groups were compared, the Nilotes tended to present with larger tumours, likely directly correlated with their slightly higher proportion of HER2-enriched tumours than the Bantus [63]. In contrast, the Cushites tended to present at a markedly earlier age (36 years) than both the other two groups and the broader Kenyan population (median of 48 years) [63,64]. Later, a study investigating population-specific variation in microRNA (miRNA) associated with cancer risk found the tumor suppressor miRNA hsa-miR-202, which contains a T-allele polymorphism with a protective effect on BrCa mortality due to increased expression of the mature miRNA, had a varied frequency amongst selected East African population groups. Interestingly, this T-allele occurred at lower frequency in the nonhunter-gatherer Maasai and Luhya populations of Kenya (0.08%) compared to the hunter-gatherer Hadza and Sandawe populations of Tanzania (43%) [65]. Although there are no studies directly comparing the rates of incidence and mortality amongst these Indigenous peoples, the overall BrCa mortality rate in Tanzania is almost half that of Kenya despite the predominance of the more aggressive triple-negative subtype [58]. This is in part due to the relatively high frequency of the protective hsa-miR-202 T-allele in people of Tanzania, but this cannot be determined based solely on this one study [65]. It does, however, warrant further investigation of hsa-miR-202 allele polymorphisms in a broader comparative study across the region or even the continent, as they may serve as prognostic markers of BrCa and prove a valuable target for precision chemotherapeutics.
Despite having the second highest incidence of BrCa in Africa, Southern Africa has the second lowest mortality rate of any region (Table 1). A meta-analysis of BrCa prognostic factors showed that the age at diagnosis tended to be younger across most of Africa compared to Southern Africa, likely because the highly admixed population in the region may exhibit genetic risk profiles closer to those observed in European populations and because the relatively high frequency of luminal BrCa subtypes [56,58]. Indeed, in Namibia BrCa was the most common cancer in the European-descent (44.7%) and admixed (38.3%) populations, followed by women from Nama (28.7–35.7%), Baster (31.4%), Damara (29.5%), Herero (26.5%), Ovambo (24.5%), Kavango (21.5%), Tswana (17.6%) and Caprivian (11.8%) [50]. In South Africa, European-descent and admixed women also had the highest ASR of mortality compared to any other group [66]. However, this does not mean that the Bantu-speaking populations of the region do not also have a heavy burden of disease. In South Africa, 82.6% of Bantu-speaking women referred to the clinic were diagnosed with early-onset TNBC before the age of 50 years, indicating that pre-menopausal Bantu women are generally at a higher risk of hereditary BrCa than other populations in South Africa [67]. BRCA1 and BRCA2 are tumour suppressor genes involved in DNA damage repair; mutations in these genes are associated with an increased risk of BrCa [68]. Surprisingly, only 4.2% of Bantu-speaking patients tested positive for BRCA1, despite displaying BRCA1-like characteristics [67]. However, previous studies have also demonstrated a low incidence of pathogenic BRCA1 variants in Bantu-speaking South African populations, such as the Xhosa (contributing just 7.1% variants), compared to other groups, suggesting that there may be other population-specific pathogenic variants that are influencing the development of BrCa in the region that have yet to be discovered [69].
The highest BrCa mortality and poorest survival rate of any African (and, in fact, world) region is found in Western Africa (ASR of 22.3 per 100,000), despite having similar incidence rates to those of other African regions (Table 1). West African ancestry has been correlated with a higher risk of developing the TNBC subtype and shorter disease-free survival [64,70,71,72]. West African women typically present with early-stage invasive BrCa with a median age at diagnosis of 44 years [71]. A limited case study of three Yoruba women with bilateral BrCa found that all patients presented at an early age with invasive ductal carcinoma, the most common type of histologically diagnosed BrCa in Nigeria [73]. All three patients were later confirmed to have TNBC, the most common subtype in both Nigeria and the broader region [58,71]. Indeed, high proportions of BRCA1 (7.4%) and BRCA2 (4.3%) variants have been identified among Yoruba BrCa patients, but this was not significantly different from that of non-Yoruba Nigerian women [74]. Additionally, the hsa-miR-202 T-allele with a known protective effect on BrCa mortality occurred at a lower frequency in the Yoruba (0.08%) than African population groups of other regions, such as the Hadza and Sandawe, suggesting that there may be differences in the potential applicability of prognostic markers amongst the Indigenous people and broader population of Nigeria [65]. Similarly, in Niger, younger age (<45 years) and late diagnosis were also reported as common features of Djerma-Sonrai and Haussa Indigenous women with BrCa [75]. Although the majority (89.4%) of women had localised tumours at diagnosis, survival was found to be low at just 13% after 12 months [75].
Of all African regions, Central Africa bears the lowest incidence and mortality of BrCa, perhaps because the less aggressive luminal subtypes predominate the encompassing countries (Table 1). However, much like its highly burdened counterpart, Northern Africa, very little is known about the burden of BrCa in the region’s Indigenous populations, as only two genomics studies to date have been performed for this purpose. One such study found several pathogenic variants in known BrCa susceptibility genes (BRCA1, BRCA2, CDH1, CHEK2, PALB2, TP53, ATM, and BARD1) among the Indigenous Baganda women of Uganda and the Bantu-speaking and Indigenous Pygmy population groups of Cameroon [76]. Notably, two miRNAs previously associated with BrCa were discovered in the Pygmy people: hsa-miR-4640, which has roles in tumorigenesis and metastasis, and hsa-miR-202, which had a low frequency of the protective T-allele in the population [65]. Since Central Africa has the lowest rates of incidence and mortality in Africa, it seems pertinent to investigate whether there are protective factors at play or if BrCa is simply underreported in the region.
4. Liver Cancer (Hepatocellular Carcinoma)
Hepatocellular carcinoma (HCC) is the most common primary malignancy of the liver [77]. Africa and Southeast Asia together account for more than 80% of the global incidence and prevalence, and HCC ranks as the sixth most frequently diagnosed cancer and the third leading cause of cancer-related mortality worldwide [2]. The development of HCC is typically driven by chronic liver injury characterized by persistent inflammation, progressive fibrosis, and dysregulated hepatocyte generation. These pathological processes promote cirrhosis and create a pro-tumorigenic microenvironment in which accumulated genetic and epigenetic alterations ultimately result in malignant transformation [77]. The main risk factors for the development of HCC include non-alcoholic fatty liver disease (NAFLD) [78], hepatitis infections (hepatitis B or hepatitis C), aflatoxin exposure, and iron overload to a lesser extent [79]. In Sub-Saharan Africa (SSA), approximately 80% of HCC cases are attributable to chronic hepatitis B virus (HBV) infection [80]. Due to dietary changes, NAFLD is increasingly being reported as an emerging and potentially growing contributor to HCC in SSA [81]. HBV contributes to hepatocarcinogenesis through both indirect and direct mechanisms: chronic infection induces ongoing hepatic inflammation, oxidative stress, and cycles of hepatocyte death and regeneration that promote fibrosis and cirrhosis, while integration of HBV DNA into the host genome can disrupt tumour suppressor genes or activate oncogenes [82,83]. Additionally, the viral HBx protein has been shown to interfere with p53 function, cell cycle regulation, and DNA repair pathways, thereby facilitating malignant transformation [84]. Due to dietary changes, particularly the increasing shift towards a Westernised diet characterised by high intake of processed foods, saturated fats, and refined sugars, NAFLD is increasingly being reported as an emerging and potentially growing contributor to HCC in SSA [81]. Ethnic differences in HCC susceptibility and clinical outcomes have been extensively investigated between Africans (including African descendants, e.g., African Americans) and Europeans [85], but comparable analysis considering the population plurality amongst different African populations is scarce.
In Mali, in the northern region of Africa, there’s a contrast between the southern and northern Saharan zones. Southern Malian groups near Bamako face a multiplicative interaction of HBV and high aflatoxin exposure in HCC, while northern Indigenous groups in Timbuktu, including Tuareg, Moor, and Songhay, lack markers of aflatoxin exposure but have an 11% endemic HBV carriage rate [86]. The molecular characterization reveals that 86.7% of HBV carriers of these groups in Timbuktu harbour truncated forms of the viral HBx gene in cell-free DNA, a molecular signature of viral integration and high-risk progression of HCC [86]. These findings indicate that HCC risk factors are not evenly distributed across Mali, but vary according to climatic, socioeconomic, and anthropological conditions, which may influence the patterns of chronic liver disease and HCC development.
Further exploring risk factors implicated in the development of HCC, an aflatoxin-induced mutation at codon 249 of the TP53 gene (p.Arg249Ser), an oncogenic hotspot mutation, has been reported in HCC patients from Nigeria, Gambia, Senegal, and Cameroon [87,88,89,90]. Aflatoxin B1 carcinogenicity depends on its hepatic bioactivation to the mutagenic AFB1-exo-8,9-epoxide, a process partly mediated by the polymorphic enzyme CYP3A5, a cytochrome P450 enzyme involved in the metabolism of xenobiotics. Certain CYP3A5 variants are associated with increased formation of this mutagenic metabolite, particularly in individuals with high CYP3A5 expression, thereby potentially increasing HCC susceptibility. Notably, a study reports moderate or high expression levels of CYP3A5 in Gambian individuals, slightly more than reported in African American liver biopsies. This difference is probably due to the European ancestry admixture in African Americans, since European ancestry individuals express low levels of CYP3A5, although the main expressing haplotype is present at similar frequencies in Gambians and African Americans [91]. This highlights how the admixed populations do not completely reflect the genetic makeup of African populations.
An analysis of the frequency of five HCC-related SNPs (rs1800562, rs2596542, rs2267716, rs9275572, and rs17401966) across 53 human populations found that the Mbuti and San Indigenous populations had the highest calculated risk score of HCC development [92]. These variations have established links to HCC susceptibility: rs1800562 is associated with hereditary hemochromatosis (iron overload); rs17401966 and rs9275572 have been linked to HBV- or HCV-related HCC risk; rs2596542 lies in the promoter region of the MICA gene and is associated with increased HCC susceptibility; and rs2267716, located in the CRHR2 gene, has been associated with HBV-related HCC in Chinese populations [92].
Most studies on HCC in Africa have primarily focused on the interplay between viral risk factors and how genomic integration events affect human genome integrity. In contrast, investigations exploring biomarkers such as miRNAs are emerging. In South Africa, a study has described 91 deregulated miRNAs in the serum of HCC HBV associated in African patients (the individuals self-identified their ethnicity, individuals of mixed ancestry were not included) that can be used as diagnostic biomarkers [93]. However, research exploring the mechanistic aspects of HCC in Africa remains unexplored.
Collectively, these findings underscore that HCC risk across Africa is not uniform, but is shaped by complex interactions between viral exposure, environmental carcinogens, metabolic transitions, and deeply rooted ancestral genomic variation. Importantly, Indigenous African populations remain markedly underrepresented in molecular, genomic, and biomarker-driven studies of HCC.
5. Cervical Cancer
Cervical cancer ranks as the second most common malignancy and the second leading cause of cancer-related mortality among women in Africa (Table 1). The burden of cervical cancer in the African population [94] is closely linked to the high prevalence of persistent infection with the oncogenic HPV, especially high-risk genotypes such as HPV-16 and HPV-18, in Western and Southern Africa, respectively [95,96]. Interestingly, HPV has coevolved with ancestral Africans for at least 500,000 years; the HPV-16 lineage is believed to have infected ancestral human populations long before modern humans emerged from Africa [97]. The development of cervical cancer is the result of persistent infection of the HPV virus, followed by the viral genome integration into the host genome, leading to the expression of E6 and E7 oncogenes. These oncogenes disturb mechanisms of DNA repair, apoptosis, and cell cycle progression checkpoints. This process transforms normal cervical epithelial cells into pre-cancerous cells and, if not detected on time, ultimately into invasive carcinoma [98].
It is important to highlight that, besides risk factors such as HPV, in SSA, approximately 60–75% of women who develop cervical cancer live in rural areas, where access to medical facilities is poorest [99]. This often leads to late-stage diagnosis and higher mortality rates compared to urban populations [100].
Cultural habits may contribute to the increase in the chances of infections by HPV. The Maasai people from Kenya and Tanzania face a substantial obstacle to controlling cervical cancer due to the lack of both quantitative and qualitative healthcare data, which hinders effective surveillance and methods of control [101]. Besides this, there are societal and cultural pressures such as polygynous family structures (a practice common among Indigenous groups), wife-sharing, and pressure to conceive that may contribute to the spread of HPV infection [101,102]. Other social factors, such as misinformation, serve as drivers of vaccine hesitancy [103]. On the other hand, there are isolated cases of success, such as Rwanda, which implemented a school-based approach and achieved HPV vaccine coverage of 80–90% in its routine cohorts [104]. However, data on the long-term impact of this program on HPV prevalence and cervical cancer incidence are not yet available.
HPV vaccination is the main target for primary prevention, with the potential to avert up to 90% of cervical cancer cases [101,105,106]. Despite the substantial burden of cervical cancer among African women (Table 1), research has largely concentrated on epidemiological patterns and viral aetiology. In contrast, cancer genomic studies on cervical cancer in African populations are markedly underrepresented, particularly among Indigenous groups, with most of the efforts focused on raising awareness about the importance of HPV vaccination across the continent.
6. Colorectal Cancer
Colorectal cancer (CRC) is the fifth most common malignancy and the fifth leading cause of cancer-related mortality in the African population. It poses a major public health challenge due to its rising incidence, with an overall ASR of 9.1 and 7.5 per 100,000 person-years for males and females, respectively (Table 1). The burden of CRC varies across the continent; Northern and Southern Africa generally report higher age-standardized incidence rates compared to the rest of the regions [107]. A quarter of affected individuals are younger than 40 years of age at presentation, and many CRC cases are already metastatic or in an advanced state [30,108]. Potential risk factors include tobacco smoking, consumption of red meat, alcohol drinking, low dietary fibre, and sedentary lifestyle; and these factors are thought to be on the incline in Indigenous African populations due to urbanisation [109,110]. However, there is also a heritable and genetic component to the disease, such as Lynch syndrome (DNA mismatch repair (MMR) disorder, which substantially increases lifetime risk of colorectal cancer), but the contribution of this is understudied in African populations [30].
As previously mentioned, Northern Africa has one of the highest burdens of CRC among African regions, with ASRs for incidence and mortality of 10.3 and 5.7, respectively [2]. Although North African countries have a similar population structure (consisting mainly of the Arab and Amazigh), they exhibit diversity in both the disease’s distribution and genetic susceptibility factors. For example, Morocco and Tunisia both have similar rates of CRC incidence, but the mortality rates in Morocco are slightly higher [2]. Population comparisons have since revealed significant differences between the Indigenous Moroccan Amazigh and Tunisians in the occurrence of CYP3A4 and CYP3A5 variants—specifically the two-variant haplotype CYP3A4*1B/CYP3A5*3, which has been linked to more severe forms of CRC (although the latter is more likely associated with increased resistance to chemotherapeutic treatment) [111]. This haplotype was found at considerably higher frequencies in the Indigenous Amazigh of Morocco (18.4%) than that of the general Tunisian population (8.4%), and higher still than that observed in African American and European men (5.3% and 0.6%, respectively) [111]. This may potentially explain mortality variance seen in the region, as the population of Amazigh peoples is relatively higher in heavily burdened countries such as Morocco and Algeria, as compared to countries with a lower proportion of Amazigh, like Egypt (mortality ASR of 6.7, 9.4, and 3.4 per 100,000, respectively) [2].
In Eastern Africa, CRC has a high ratio of mortality as compared to its relatively low incidence (7.8 vs. 5.6 ASR per 100,000, respectively; Table 1) as it presents at an advanced stage in the affected population [112]. In Uganda, where the CRC mortality is similar to that of the rest of the region, the presentation of many of the Indigenous Baganda patients was below 40 years compared to the mean of 54 years for the general population, and they had both a higher predominance and number of adenomas in the colon [112]. Notably, the risk of having multiple adenomas (indicative of aggressive malignancy) was higher in Baganda patients who had a familial history of colon or rectal cancer, pointing to a potential heritable risk factor for advanced disease in this population [112]. However, to date, no genomic discovery studies have been conducted in this population to ascertain this.
Within Southern Africa, South Africa has one of the highest rates of CRC-related mortality in Africa (ASR of 8.9 per 100,000), second only to Libya, despite having one of the lowest rates of incidence. Generally, the Bantu-speaking population has more aggressive disease and a poorer prognosis than the European-descent population. The mean age at presentation is 56.8 in South Africa, but the median age among the Bantu patients was significantly lower than the median age among European-descent patients (55 vs. 63) [113]. This is despite the fact that left-sided colon cancer (which typically bears a better prognosis than right-side colon cancer) was more common in Bantu-speaking (21.2%) patients compared to European-descent (15.2%) patients [114], and was specifically linked to the BAT25 and BAT26 microsatellite markers [114]. These markers are commonly used to detect Microsatellite Instability high (MSI-H) CRC tumours, as they can cause frame-shift mutations in tumor-suppressor genes when shortened by DNA MMR deficiencies, thus leading to a worse prognosis [115]. They therefore appear also to be a negative prognostic indicator in the Bantu-speaking population of South Africa [114]. Notably, only germline variation of these markers was found in Bantu-speaking patients, suggesting that these individuals may have a stronger hereditary basis than in other South African populations [114]. A few recent studies have also linked South African Bantu ethnicity with more frequent MSI-H CRC, and the development of early onset CRC in these patients was associated with Lynch syndrome and MMR gene variants [116,117]. In comparison to that of the colon, there was little variation in the presentation of rectal cancer, including high-grade disease and malignant signet ring cell morphology. However, there was a significant difference in stage at presentation. More advanced stage disease (i.e., stage III and stage IV) was diagnosed among Bantu (81%) patients, who were also more likely to have metastasis, compared to European-descent patients (60.7%). However, this is influenced by differences in cultural health-seeking behaviours and access to appropriate facilities [113]. Likely due to their late stage at presentation, overall Bantu patients had a significantly reduced median survival of 20.9 months compared to that of 32.2 months in European-descent patients [113].
Interestingly, Western Africans displayed similar clinical characteristics to that of Southern Africans, despite having a lower overall incidence and mortality rate (Table 1). Genetically, Nigerians have a significantly higher rate of MSI-H tumours and a lower frequency of Wnt pathway and APC mutations compared to North American cohorts, suggesting alternative drivers for the disease in the population [118]. When focusing just on the Indigenous and ethnic populations from Nigeria, 73% of overall recorded hospital cases were Yoruba, 13.5% were Igbo, and 12.9% were Bendel, while Hausa, Cross-river, and Rivers made up the remaining 1.5%, 1.8% and 0.5%, respectively [119]. Over the last 40 years, reports from studies in Ibadan (majority Yoruba) have shown a steady increase in case numbers [119]. Certainly, the Yoruba appear to have more aggressive disease than other Indigenous Nigerian groups, as indicated both in their relatively high incidence and the presentation of CRC. A study found that the mean age at presentation in the Yoruba was 54 years, and a large proportion of patients were just 30 years of age at the time of diagnosis (6.9%) [113]. Right-sided CRC were most common in the Yoruba (65.8%) [113]. They were also more likely to present with high-grade and/or advanced disease stages (82.4%). The median survival among the Nigerian cohort was 21.9 months [113]. The cancer profile in Niger is similar to that of Nigeria. Most CRC in Niger were adenocarcinomas, and 8.6% of individuals died during the 17 year study period of the Niger Cancer Registry [120]. The average age at diagnosis was 47 years, with the highest rates for women and men in the range of 40–69 years and 35–69 years, respectively [120]. CRC risk also varied among various Indigenous groups in Niger. The Djerma Sonrai were more likely to develop CRC (55%), followed by Hausa (29.5%) and Touareg (6.6%) [120]. As the culture, diet, and environment in each population are drastically different and can affect CRC risk, it is not entirely surprising that the incidence varies among the different Indigenous populations. Much like the CRC studies in Nigeria, the sample sizes of the various ethnic and Indigenous groups in Niger were small and geographically limited, perhaps by difficulties in accessing healthcare or even just the small size of the Indigenous population itself. This speaks to the need for large-scale regional or country-wide comparative studies of CRC that are equitable and includes Indigenous populations that are not heavily influenced by the Western lifestyle/environment, as it may obscure population-specific susceptibility factors.
7. Lung Cancer
Lung cancer is the sixth most common cancer and the sixth leading cancer-related cause of death in Africa [2]. The occurrence of lung cancer differs throughout the African continent; Northern and Southern regions display higher lung cancer incidence compared to the other regions of the continent (Table 1). Although SSA comprises one of the regions with higher occurrences of lung cancer, it still lacks epidemiological and genomic data; however, efforts are being made to cover this gap [121].
Lung cancer arises from the epithelial cells of the respiratory tract; it encompasses two principal histopathological categories: non-small cell lung cancer (NSCLC) and small-cell lung cancer (SCLC). The pathogenesis of lung cancer reflects a complex interplay of genetic alterations and environmental exposures, with tobacco smoking remaining the dominant risk factor globally. However, exposure to environmental carcinogens (such as air pollution and occupational agents) and inherited susceptibility also contribute to disease development [122,123].
The epidermal growth factor receptor (EGFR) is a transmembrane protein that is highly expressed in more than 60% of NSCLC, making it an important target for lung cancer therapeutics (e.g., tyrosine kinase inhibitors) [124]. In North Africa, the prevalence of EGFR mutations ranges between 21% and 24.3%, consistently higher than the reported for European populations (approximately 15%), but significantly lower than Asian populations (approximately 50%) [125]. Within Moroccan cohorts, exon 19 deletions are the most frequent EGFR alteration, which may predict a favourable response to targeted tyrosine kinase inhibitors [125]. Other significant drivers in Morocco include TP53 and KRAS mutations as well as ALK fusions [126]. Notably, compared with the Cancer Atlas Genome (TCGA) database, considerable differences were observed in the prevalence of EGFR (Moroccan cohort 24.3% vs. 14.9% in TCGA), KRAS (Moroccan cohort 22.9% vs. 25.9%), and TP53 (Moroccan cohort 34.3% vs. 50.9%). In Tunisia, the EGFR mutation rate is lower, though ALK positive expression was found to be relatively high (9.09%) [127] compared to the Moroccan and Egyptian cohorts. In Egypt, the prevalence of EGFR mutations is approximately 17.5% [128].
Overall, lung cancer in Africa demonstrates significant regional heterogeneity in incidence, histopathology, and genomic profiles, with Northern and Southern regions exhibiting higher disease burden. Molecular analyses highlight population-specific differences in key driver mutations, such as EGFR, TP53, KRAS, and ALK, with important implications for targeted therapies and clinical outcomes, as they underpin precision medicine approaches in lung cancer. Despite these insights, Indigenous populations remain markedly underrepresented in both epidemiological and genomic studies, limiting our understanding of lung cancer risk factors, mutational landscapes, and therapeutic responsiveness within these communities.
8. Future Perspectives
The main limitation emerging from the evidence reviewed is that the genomic diversity of African Indigenous populations remains largely unexplored, despite its clear relevance for understanding the biological basis of non-communicable diseases. Many cancers discussed in this review show population-specific patterns of incidence, aggressiveness, and age of onset. However, the scarcity of ancestry-resolved genomic data from Indigenous populations limits our ability to identify risk and protective variants, reconstruct disease-relevant evolutionary histories, and determine how deeply rooted genetic lineages contribute to cancer susceptibility in contemporary African populations [1]. Firstly, more coordinated efforts are needed to collect health data from Indigenous populations. This knowledge gap can hinder the development of precision medicine in African contexts. The incomplete and uneven characterization of cancer genomics in African Indigenous populations points to the need for ancestry-aware analytic frameworks to translate genomic advances into equitable cancer care [1]. Most existing genomic reference datasets, risk models, and therapeutic targets have been derived from non-African populations, reducing their predictive accuracy and clinical utility for Africans, particularly those with substantial Indigenous ancestry [1,129]. As cancer genomics advances, applying ancestry-informed precision frameworks will be critical [129]. Without systematic inclusion of Indigenous genomes, precision oncology efforts risk perpetuating health disparities by applying tools that inadequately capture African genetic architecture and tumour biology.
Looking forward, integrating whole-genome sequencing, tumour profiling, and detailed clinical data from these populations can uncover ancestry-specific drivers of cancer and therefore improve prediction of treatment response and toxicity. However, advancing understanding in this area requires more than the collection of genetic samples; it is also essential to elucidate the mechanistic pathways through which genetic variation, in conjunction with environmental, lifestyle, and socio-cultural factors, contributes to differences in cancer incidence, progression, mortality, and therapeutic outcomes. Such efforts have the potential to benefit not only Indigenous groups themselves but also the broader African population and the global community, whose genomes are shaped by varying degrees of Indigenous ancestry. Achieving these goals would likely require a fundamental shift in research infrastructure, including the increased establishment and strengthening of African-owned biobanks, sequencing facilities, and technical expertise. Locally controlled biobanks can help ensure ethical stewardship of samples, respect for Indigenous data sovereignty, long-term sustainability, and equitable benefit-sharing [130]. Additionally, the use of specimens in these biobanks should be prioritized in studies pertaining to conditions of importance within African communities [130]. African-led biobanking initiatives would enable large-scale, longitudinal studies that link genomic, environmental, and clinical data, while reducing dependence on external repositories that often limit African access and control [131]. Strengthening cross-institutional networks and engagement across Africa will be critical to fully leverage these African-owned biobanks and advance impactful genomic research [131].
In this context, practical approaches such as implementing consent processes and frameworks that explicitly recognize participants’ rights over their data and biospecimens, alongside locally led institutional review boards and community advisory structures, are essential to ensure contextually appropriate oversight. Importantly, to enable scientific progress that aligns with the sovereignty of Indigenous and local populations, these frameworks must prioritize community ownership, equitable benefit-sharing, and culturally grounded governance. The feasibility of such approaches is demonstrated by initiatives such as the Human Heredity and Health in Africa Consortium [132], which have successfully implemented Africa-led governance, ethical data-sharing, and community engagement frameworks. With adequate support, these models show that sustainable, locally governed biobanking is achievable, underscoring that initiatives in cancer genomics and related biobanking efforts are both feasible and urgently needed.
Raising awareness of the necessity for such improvements to the current research infrastructure is therefore central to translating Indigenous genomic research into lasting scientific and public health impact in all of the diverse communities across the continent. Ultimately, such efforts are aligned with the broader goal of ensuring equitable healthcare access and outcomes across the continent, contributing directly to the achievement of The United Nation’s Sustainable Development Goal 3 (SDG3) on good health and well-being.
Statement of the Use of Generative AI and AI-Assisted Technologies in the Writing Process
During the preparation of this work, the authors used ChatGPT 5.2 to improve readability and language. After using this tool/service, the authors reviewed and edited the content as needed and take full responsibility for the content of the publication.
Acknowledgments
We extend our gratitude to the body of work that we cited in this paper and their invaluable contribution towards health equity in African populations.
Author Contributions
Conceptualization: K.C.S.S., C.L.M.F., S.C. and L.F.Z.; Investigation: K.C.S.S., C.L.M.F., M.W. and M.S.Z.; Writing—Original Draft Preparation: K.C.S.S., C.L.M.F., M.W. and M.S.Z.; Writing—Review & Editing: K.C.S.S., C.L.M.F., M.W., M.S.Z., S.C., L.D. and L.F.Z.; Visualization: C.L.M.F.; Supervision: S.C., L.D. and L.F.Z.; Project administration: L.F.Z.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable to this article as no datasets were generated or analyzed during the current study.
Funding
This research was funded by International Centre for Genetic Engineering and Biotechnology.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
Thomas CE, Peters U. Genomic Landscape of Cancer in Racially and Ethnically Diverse Populations. Nat. Rev. Genet. 2025, 26, 336–349. DOI:10.1038/s41576-024-00796-w [Google Scholar]
-
Bray F, Laversanne M, Sung H, Ferlay J, Siegel RL, Soerjomataram I, et al. Global Cancer Statistics 2022: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2024, 74, 229–263. DOI:10.3322/caac.21834 [Google Scholar]
-
Freedman JA, Al Abo M, Allen TA, Piwarski SA, Wegermann K, Patierno SR. Biological Aspects of Cancer Health Disparities. Annu. Rev. Med. 2021, 72, 229–241. DOI:10.1146/annurev-med-070119-120305 [Google Scholar]
-
Ngwa W, Addai BW, Adewole I, Ainsworth V, Alaro J, Alatise OI, et al. Cancer in Sub-Saharan Africa: A Lancet Oncology Commission. Lancet Oncol. 2022, 23, e251–e312. DOI:10.1016/S1470-2045(21)00720-8 [Google Scholar]
-
Joko-Fru WY, Bardot A, Bukirwa P, Amidou S, N’da G, Woldetsadik E, et al. Cancer Survival in Sub-Saharan Africa (SURVCAN-3): A Population-Based Study. Lancet Glob. Health 2024, 12, e947–e959. DOI:10.1016/S2214-109X(24)00130-X [Google Scholar]
-
Fatumo S, Chikowore T, Choudhury A, Ayub M, Martin AR, Kuchenbäcker K. Diversity in Genomic Studies: A Roadmap to Address the Imbalance. Nat. Med. 2022, 28, 243–250. DOI:10.1038/s41591-021-01672-4 [Google Scholar]
-
Pfennig A, Petersen LN, Kachambwa P, Lachance J. Evolutionary Genetics and Admixture in African Populations. Genome Biol. Evol. 2023, 15, evad054. DOI:10.1093/gbe/evad054 [Google Scholar]
-
Jakobsson M, Scholz SW, Scheet P, Gibbs JR, VanLiere JM, Fung H-C, et al. Genotype, Haplotype and Copy-Number Variation in Worldwide Human Populations. Nature 2008, 451, 998–1003. DOI:10.1038/nature06742 [Google Scholar]
-
Tishkoff SA, Reed FA, Friedlaender FR, Ehret C, Ranciaro A, Froment A, et al. The Genetic Structure and History of Africans and African Americans. Science 2009, 324, 1035–1044. DOI:10.1126/science.1172257 [Google Scholar]
-
Segelov E, Garvey G. Cancer and Indigenous Populations: Time to End the Disparity. JCO Glob. Oncol. 2020, 6, JGO-19. DOI:10.1200/JGO.19.00379 [Google Scholar]
-
D’Angelo CS, Hermes A, McMaster CR, Prichep E, Richer É, van der Westhuizen FH, et al. Barriers and Considerations for Diagnosing Rare Diseases in Indigenous Populations. Front. Pediatr. 2020, 8, 579924. DOI:10.3389/fped.2020.579924 [Google Scholar]
-
United Nations Department of Economic and Social Affairs. State of the World’s Indigenous Peoples; Vereinte Nationen, Ständiges Forum für Indigene Angelegenheiten, Ed.; United Nations Department of Economic and Social Affairs: New York, NY, USA, 2009; ISBN 978-92-1-130283-7. [Google Scholar]
-
Go AE, Guinaran RC, Dee EC. Closing the Cancer Care Gap: Cancer Disparities among Indigenous Peoples Globally. Lancet Reg. Health-West. Pac. 2025, 57, 101543. DOI:10.1016/j.lanwpc.2025.101543 [Google Scholar]
-
IWGIA. Indigenous World, 37th ed.; IWGIA: Copenhagen, Denmark, 2023; ISBN 978-87-93961-61-6. [Google Scholar]
-
Schlebusch CM, Malmström H, Günther T, Sjödin P, Coutinho A, Edlund H, et al. Southern African Ancient Genomes Estimate Modern Human Divergence to 350,000 to 260,000 Years Ago. Science 2017, 358, 652–655. DOI:10.1126/science.aao6266 [Google Scholar]
-
Ehret C. Bantu Expansions: Re-Envisioning a Central Problem of Early African History. Int. J. Afr. Hist. Stud. 2001, 34, 5–41. DOI:10.2307/3097285 [Google Scholar]
-
Skoglund P, Thompson JC, Prendergast ME, Mittnik A, Sirak K, Hajdinjak M, et al. Reconstructing Prehistoric African Population Structure. Cell 2017, 171, 59–71.e21. DOI:10.1016/j.cell.2017.08.049 [Google Scholar]
-
Rotimi CN, Bentley AR, Doumatey AP, Chen G, Shriner D, Adeyemo A. The Genomic Landscape of African Populations in Health and Disease. Hum. Mol. Genet. 2017, 26, R225–R236. DOI:10.1093/hmg/ddx253 [Google Scholar]
-
Fan S, Kelly DE, Beltrame MH, Hansen MEB, Mallick S, Ranciaro A, et al. African Evolutionary History Inferred from Whole Genome Sequence Data of 44 Indigenous African Populations. Genome Biol. 2019, 20, 82. DOI:10.1186/s13059-019-1679-2 [Google Scholar]
-
Prendergast ME, Sawchuk EA, Sirak KA. Genetics and the African Past. In Oxford Research Encyclopedia of African History; Oxford University: Oxford, UK, 2022; ISBN 978-0-19-027773-4. [Google Scholar]
-
Schlebusch CM, Sjödin P, Breton G, Günther T, Naidoo T, Hollfelder N, et al. Khoe-San Genomes Reveal Unique Variation and Confirm the Deepest Population Divergence in Homo Sapiens. Mol. Biol. Evol. 2020, 37, 2944–2954. DOI:10.1093/molbev/msaa140 [Google Scholar]
-
Fan S, Spence JP, Feng Y, Hansen MEB, Terhorst J, Beltrame MH, et al. Whole-Genome Sequencing Reveals a Complex African Population Demographic History and Signatures of Local Adaptation. Cell 2023, 186, 923–939.e14. DOI:10.1016/j.cell.2023.01.042 [Google Scholar]
-
Sirak KA, Sawchuk EA, Prendergast ME. Ancient Human DNA and African Population History. In Oxford Research Encyclopedia of Anthropology; Oxford University Press: Oxford, UK, 2022; ISBN 978-0-19-085458-4. [Google Scholar]
-
Corpas M, Pius M, Poburennaya M, Guio H, Dwek M, Nagaraj S, et al. Bridging Genomics’ Greatest Challenge: The Diversity Gap. Cell Genom. 2025, 5, 100724. DOI:10.1016/j.xgen.2024.100724 [Google Scholar]
-
Nguyen ADK, Meehan K, Redfern AD, Brown A, Robinson M, Papertalk L, et al. The Impact of Genetics and the Environment on Cancer Risk in Indigenous Australians: A Narrative Review. Lancet Reg. Health-West. Pac. 2025, 61, 101627. DOI:10.1016/j.lanwpc.2025.101627 [Google Scholar]
-
Sirugo G, Williams SM, Tishkoff SA. The Missing Diversity in Human Genetic Studies. Cell 2019, 177, 26–31. DOI:10.1016/j.cell.2019.02.048 [Google Scholar]
-
Martin AR, Kanai M, Kamatani Y, Okada Y, Neale BM, Daly MJ. Clinical Use of Current Polygenic Risk Scores May Exacerbate Health Disparities. Nat. Genet. 2019, 51, 584–591. DOI:10.1038/s41588-019-0379-x [Google Scholar]
-
Caron NR, Chongo M, Hudson M, Arbour L, Wasserman WW, Robertson S, et al. Indigenous Genomic Databases: Pragmatic Considerations and Cultural Contexts. Front. Public Health 2020, 8, 111. DOI:10.3389/fpubh.2020.00111 [Google Scholar]
-
Gheybi K, Mmekwa N, Lebelo MT, Patrick SM, Campbell R, Nenzhelele M, et al. Linking African Ancestral Substructure to Prostate Cancer Health Disparities. Sci. Rep. 2023, 13, 20909. DOI:10.1038/s41598-023-47993-x [Google Scholar]
-
Hamdi Y, Abdeljaoued-Tej I, Zatchi AA, Abdelhak S, Boubaker S, Brown JS, et al. Cancer in Africa: The Untold Story. Front. Oncol. 2021, 11, 650117. DOI:10.3389/fonc.2021.650117 [Google Scholar]
-
Rebbeck TR. Prostate Cancer Genetics: Variation by Race, Ethnicity, and Geography. Semin. Radiat. Oncol. 2017, 27, 3–10. DOI:10.1016/j.semradonc.2016.08.002 [Google Scholar]
-
Temilola DO, Wium M, Paccez J, Salukazana AS, Rotimi SO, Otu HH, et al. Detection of Cancer-Associated Gene Mutations in Urinary Cell-Free DNA among Prostate Cancer Patients in South Africa. Genes 2023, 14, 1884. DOI:10.3390/genes14101884 [Google Scholar]
-
Berenguer CV, Pereira F, Câmara JS, Pereira JAM. Underlying Features of Prostate Cancer—Statistics, Risk Factors, and Emerging Methods for Its Diagnosis. Curr. Oncol. 2023, 30, 2300–2321. DOI:10.3390/curroncol30020178 [Google Scholar]
-
Ahmed D, Mohammed EAA, Ahmed MEM, Abdalla YMO, Hadad I, Elimam AA, et al. Epidemiologic Study on Prostate Cancer in Sudanese Men across African Ethnic Groups. Sci. Rep. 2024, 14, 29646. DOI:10.1038/s41598-024-77475-7 [Google Scholar]
-
Safiri S, Shamekh A, Hassanzadeh K, Fazlollahi A, Sullman MJM, Raeisi M, et al. The Burden of Prostate Cancer in the North Africa and Middle East Region from 1990 to 2021. Sci. Rep. 2025, 15, 1853. DOI:10.1038/s41598-024-83840-3 [Google Scholar]
-
Samtal C, Bouguenouch L, Ismaili N, El Baldi M, El Makhzen B, El Rhazi K, et al. Update on Prostate Cancer Epidemiology in Morocco. Afr. J. Urol. 2024, 30, 22. DOI:10.1186/s12301-024-00419-0 [Google Scholar]
-
Arauna LR, Comas D. Genetic Heterogeneity between Berbers and Arabs. In Encyclopedia of Life Sciences; Wiley: Hoboken, NJ, USA, 2017; pp. 1–7; ISBN 978-0-470-01617-6. [Google Scholar]
-
Arauna LR, Mendoza-Revilla J, Mas-Sandoval A, Izaabel H, Bekada A, Benhamamouch S, et al. Recent Historical Migrations Have Shaped the Gene Pool of Arabs and Berbers in North Africa. Mol. Biol. Evol. 2016, 34, 318–329. DOI:10.1093/molbev/msw218 [Google Scholar]
-
Salmi F, Maachi F, Tazzite A, Aboutaib R, Fekkak J, Azeddoug H, et al. Next-Generation Sequencing of BRCA1 and BRCA2 Genes in Moroccan Prostate Cancer Patients with Positive Family History. PLoS ONE 2021, 16, e0254101. DOI:10.1371/journal.pone.0254101 [Google Scholar]
-
Taha SM, Weng H-Y, Mohammed MEI, Osman YM, N’dri N, Mohammed SI, et al. Prostate Cancer Clinical Characteristics and Outcomes in Central Sudan. Ecancermedicalscience 2020, 14, 1116. DOI:10.3332/ecancer.2020.1116 [Google Scholar]
-
Elamin A, Ibrahim ME, Abuidris D, Mohamed KEH, Mohammed SI. Part I: Cancer in Sudan—Burden, Distribution, and Trends Breast, Gynecological, and Prostate Cancers. Cancer Med. 2015, 4, 447–456. DOI:10.1002/cam4.378 [Google Scholar]
-
Genome-Wide Analysis of Luhya and Maasai Genomes Reveals Signatures of Selection for Fertility, Prostate Cancer, Prostatitis and Vein Disease—MedCrave Online. Available online: https://medcraveonline.com/JCPCR/genome-wide-analysis-of-luhya-and-maasai-genomes-reveals-signatures-of-selection-for-fertility-prostate-cancer-prostatitis-and-vein-disease.html (accessed on 3 February 2026).
-
Gu L, Frommel SC, Oakes CC, Simon R, Grupp K, Gerig CY, et al. BAZ2A (TIP5) Is Involved in Epigenetic Alterations in Prostate Cancer and Its Overexpression Predicts Disease Recurrence. Nat. Genet. 2015, 47, 22–30. DOI:10.1038/ng.3165 [Google Scholar]
-
Lachance J, Berens AJ, Hansen MEB, Teng AK, Tishkoff SA, Rebbeck TR. Genetic Hitchhiking and Population Bottlenecks Contribute to Prostate Cancer Disparities in Men of African Descent. Cancer Res. 2018, 78, 2432–2443. DOI:10.1158/0008-5472.CAN-17-1550 [Google Scholar]
-
Sun J, Zheng SL, Wiklund F, Isaacs SD, Li G, Wiley KE, et al. Sequence Variants at 22q13 Are Associated with Prostate Cancer Risk. Cancer Res. 2009, 69, 10–15. DOI:10.1158/0008-5472.CAN-08-3464 [Google Scholar]
-
Bu H, Narisu N, Schlick B, Rainer J, Manke T, Schäfer G, et al. Putative Prostate Cancer Risk SNP in an Androgen Receptor-Binding Site of the Melanophilin Gene Illustrates Enrichment of Risk SNPs in Androgen Receptor Target Sites. Human Mutat. 2016, 37, 52–64. DOI:10.1002/humu.22909 [Google Scholar]
-
Iheanacho CO, Enechukwu OH. Epidemiology of Prostate Cancer in Nigeria: A Mixed Methods Systematic Review. Cancer Causes Control 2025, 36, 1–12. DOI:10.1007/s10552-024-01917-w [Google Scholar]
-
Ugwumba FO, Okoh AD, Echetabu KN, Udeh EI, Nnabugwu II. Prostate Cancer Detected by Screening in a Semi Urban Community in Southeast Nigeria: Correlations and Associations between Anthropometric Measurements and Prostate-specific Antigen. Niger. J. Surg. 2017, 23, 33–36. DOI:10.4103/1117-6806.199967 [Google Scholar]
-
Erhabor O, Hussaini AM, Abdulwahab-Ahmed A, Retsky M, Erhabor T. Some Oxidative Stress Biomarkers among Patients with Prostate Cancer in Sokoto, North Western Nigeria. Open J. Blood Dis. 2022, 12, 60–78. DOI:10.4236/ojbd.2022.123007 [Google Scholar]
-
Koegelenberg MR, Hansen MR-J. Namibia National Cancer Registry (NNCR) Cancer Incidences in Namibia 2010–2014. Available online: https://afcrn.org/images/M_images/attachments/125/Cancer%20in%20Namibia%202010-2014.pdf (accessed on 5 February 2026).
-
Petersen DC, Jaratlerdsiri W, van Wyk A, Chan EKF, Fernandez P, Lyons RJ, et al. African KhoeSan Ancestry Linked to High-Risk Prostate Cancer. BMC Med. Genom. 2019, 12, 82. DOI:10.1186/s12920-019-0537-0 [Google Scholar]
-
Tindall EA, Bornman MR, van Zyl S, Segone AM, Monare LR, Venter PA, et al. Addressing the Contribution of Previously Described Genetic and Epidemiological Risk Factors Associated with Increased Prostate Cancer Risk and Aggressive Disease within Men from South Africa. BMC Urol. 2013, 13, 74. DOI:10.1186/1471-2490-13-74 [Google Scholar]
-
Ollomo B, Mouélé LY, Bivigou-Mboumba B, Ondo BM, Lendoye E, Mezui J, et al. D Allele and DD Genotype of I/D Polymorphism in The ACE Gene in Patients with Hypertension, Stroke and Cancer Prostate in Libreville: A Concern Given the High Frequencies of These Signatures in Gabonese Population. J. Proteom. Genom. Res. 2019, 2, 3–12. DOI:10.14302/issn.2326-0793.jpgr-19-3114 [Google Scholar]
-
Medeiros R, Vasconcelos A, Costa S, Pinto D, Lobo F, Morais A, et al. Linkage of Angiotensin I-converting Enzyme Gene Insertion/Deletion Polymorphism to the Progression of Human Prostate Cancer. J. Pathol. 2004, 202, 330–335. DOI:10.1002/path.1529 [Google Scholar]
-
Eng A, McCormack V, dos-Santos-Silva I. Receptor-Defined Subtypes of Breast Cancer in Indigenous Populations in Africa: A Systematic Review and Meta-Analysis. PLoS Med. 2014, 11, e1001720. DOI:10.1371/journal.pmed.1001720 [Google Scholar]
-
Parenté A, Gnangnon FHR, Kinsou D, Chkeir M, Adou C, Robin Sacca H, et al. Prognostic Factors of Breast Cancer and Survival in Sub-Saharan Africa: A Systematic Review and Meta-Analysis. Bull. du Cancer 2025, 112, 1024–1034. DOI:10.1016/j.bulcan.2025.05.009 [Google Scholar]
-
Martini R, Delpe P, Chu TR, Arora K, Lord B, Verma A, et al. African Ancestry–Associated Gene Expression Profiles in Triple-Negative Breast Cancer Underlie Altered Tumor Biology and Clinical Outcome in Women of African Descent. Cancer Discov. 2022, 12, 2530–2551. DOI:10.1158/2159-8290.CD-22-0138 [Google Scholar]
-
Onyia AF, Nana TA, Adewale EA, Adebesin AO, Adegboye BE, Paimo OK, et al. Breast Cancer Phenotypes in Africa: A Scoping Review and Meta-Analysis. JCO Glob. Oncol. 2023, 9, e2300135. DOI:10.1200/GO.23.00135 [Google Scholar]
-
Orrantia-Borunda E, Anchondo-Nuñez P, Acuña-Aguilar LE, Gómez-Valles FO, Ramírez-Valdespino CA. Subtypes of Breast Cancer. In Breast Cancer; Mayrovitz HN, Ed.; Exon Publications: Brisbane, Australia, 2022; ISBN 978-0-6453320-3-2. [Google Scholar]
-
Elkhalloufi F. Access to Healthcare among Moroccan Breast Cancer Patients: Experience of the National Institute of Oncology in Rabat. J. Clin. Case Rep. Med. Images Health Sci. 2023, 3. DOI:10.55920/JCRMHS.2023.03.001129 [Google Scholar]
-
Bakkach J, Mansouri M, Derkaoui T, Loudiyi A, Fihri M, Hassani S, et al. Clinicopathologic and Prognostic Features of Breast Cancer in Young Women: A Series from North of Morocco. BMC Women’s Health 2017, 17, 106. DOI:10.1186/s12905-017-0456-1 [Google Scholar]
-
Derkaoui T, Bakkach J, Mansouri M, Loudiyi A, Fihri M, Alaoui FZ, et al. Triple Negative Breast Cancer in North of Morocco: Clinicopathologic and Prognostic Features. BMC Women’s Health 2016, 16, 68. DOI:10.1186/s12905-016-0346-y [Google Scholar]
-
Sayed S, Moloo Z, Wasike R, Bird P, Oigara R, Njoroge FW, et al. Ethnicity and Breast Cancer Characteristics in Kenya. Breast Cancer Res. Treat. 2018, 167, 425–437. DOI:10.1007/s10549-017-4511-2 [Google Scholar]
-
Popli P, Gutterman EM, Omene C, Ganesan S, Mills D, Marlink R. Receptor-Defined Breast Cancer in Five East African Countries and Its Implications for Treatment: Systematic Review and Meta-Analysis. JCO Glob. Oncol. 2021, 7, 289–301. DOI:10.1200/GO.20.00398 [Google Scholar]
-
Rawlings-Goss RA, Campbell MC, Tishkoff SA. Global Population-Specific Variation in miRNA Associated with Cancer Risk and Clinical Biomarkers. BMC Med. Genom. 2014, 7, 53. DOI:10.1186/1755-8794-7-53 [Google Scholar]
-
Statistics South Africa. Cancer in South Africa, 2008–2019 [Report. No. 03-08-00]; Statistics South Africa: Pretoria, South Africa, 2023. [Google Scholar]
-
Makhetha M, Chabilal N, Aldous C. Recent Trends in Hereditary Breast Cancer Incidence by Race and Age in KwaZulu-Natal, South Africa: An 11-Year Single-Centre Retrospective Study (2011–2021). Int. J. Cancer 2024, 154, 65–70. DOI:10.1002/ijc.34687 [Google Scholar]
-
Petrucelli N, Daly MB, Pal T. BRCA1- and BRCA2-Associated Hereditary Breast and Ovarian Cancer. In GeneReviews®; Adam MP, Bick S, Mirzaa GM, Pagon RA, Wallace SE, Amemiya A, Eds.; University of Washington: Seattle, WA, USA, 1993. [Google Scholar]
-
Francies FZ, Wainstein T, De Leeneer K, Cairns A, Murdoch M, Nietz S, et al. BRCA1, BRCA2 and PALB2 Mutations and CHEK2 c.1100delC in Different South African Ethnic Groups Diagnosed with Premenopausal and/or Triple Negative Breast Cancer. BMC Cancer 2015, 15, 912. DOI:10.1186/s12885-015-1913-6 [Google Scholar]
-
Biancolella M, Ouédraogo NLM, Zongo N, Zohoncon TM, Testa B, Rizzacasa B, et al. Breast cancer in West Africa: molecular analysis of BRCA genes in early-onset breast cancer patients in Burkina Faso. Hum Genomics. 2021, 15, 65. DOI:10.1186/s40246-021-00365-w [Google Scholar]
-
Reid S, Fan R, Venton L, Weidner A, Tezak A, Roberson ML, et al. West African Genetic Ancestry and Breast Cancer Outcomes Among Black Women. JAMA Netw. Open 2024, 7, e2449798. DOI:10.1001/jamanetworkopen.2024.49798 [Google Scholar]
-
Nwagu GC, Bhattarai S, Swahn M, Ahmed S, Aneja R. Prevalence and Mortality of Triple-Negative Breast Cancer in West Africa: Biologic and Sociocultural Factors. JCO Glob. Oncol. 2021, 7, 1129–1140. DOI:10.1200/GO.21.00082 [Google Scholar]
-
Olubunmi Obajimi M, Adeniji-Sofoluwe AT, Adeoye AO, Obajimi GO, Ajani MA, Adejumo PO, et al. Bilateral Breast Cancer among Three Yoruba Women in a Nigerian Teaching Hospital. BJR Case Rep. 2015, 1, 20150156. DOI:10.1259/bjrcr.20150156 [Google Scholar]
-
Fackenthal JD, Zhang J, Zhang B, Zheng Y, Hagos F, Burrill DR, et al. High Prevalence of BRCA1 and BRCA2 Mutations in Unselected Nigerian Breast Cancer Patients. Int. J. Cancer 2012, 131, 1114–1123. DOI:10.1002/ijc.27326 [Google Scholar]
-
Aissami A, Van Hal G, Nouhou H, Abari MM. What Can 27 Years of Niger Cancer Registry Tell Us about Breast Cancer Epidemiology? Incidence of Breast Cancer from 1992 to 2018 in Niger. J. Public Health Epidemiol. 2021, 13, 293–302. Available online: https://repository.uantwerpen.be/link/irua/200747 (accessed on 5 February 2026).
-
Adedokun B, Zheng Y, Ndom P, Gakwaya A, Makumbi T, Zhou AY, et al. Prevalence of Inherited Mutations in Breast Cancer Predisposition Genes among Women in Uganda and Cameroon. Cancer Epidemiol. Biomark. Prev. 2020, 29, 359–367. DOI:10.1158/1055-9965.EPI-19-0506 [Google Scholar]
-
Villanueva A. Hepatocellular Carcinoma. N. Engl. J. Med. 2019, 380, 1450–1462. DOI:10.1056/NEJMra1713263 [Google Scholar]
-
Sharma N, Singh L, Sharma A, Kumar A, Mahajan D. NAFLD-Associated Hepatocellular Carcinoma (HCC)—A Compelling Case for Repositioning of Existing mTORc1 Inhibitors. Pharmacol. Res. 2024, 208, 107375. DOI:10.1016/j.phrs.2024.107375 [Google Scholar]
-
Kedar Mukthinuthalapati VVP, Sewram V, Ndlovu N, Kimani S, Abdelaziz AO, Chiao EY, et al. Hepatocellular Carcinoma in Sub-Saharan Africa. JCO Glob. Oncol. 2021, 7, 756–766. DOI:10.1200/GO.20.00425 [Google Scholar]
-
Spearman CW, Afihene M, Ally R, Apica B, Awuku Y, Cunha L, et al. Hepatitis B in Sub-Saharan Africa: Strategies to Achieve the 2030 Elimination Targets. Lancet Gastroenterol. Hepatol. 2017, 2, 900–909. DOI:10.1016/S2468-1253(17)30295-9 [Google Scholar]
-
Spearman CW, Afihene M, Betiku O, Bobat B, Cunha L, Kassianides C, et al. Epidemiology, Risk Factors, Social Determinants of Health, and Current Management for Non-Alcoholic Fatty Liver Disease in Sub-Saharan Africa. Lancet Gastroenterol. Hepatol. 2021, 6, 1036–1046. DOI:10.1016/S2468-1253(21)00275-2 [Google Scholar]
-
Zhang M, Chen H, Liu H, Tang H. The Impact of Integrated Hepatitis B Virus DNA on Oncogenesis and Antiviral Therapy. Biomark. Res. 2024, 12, 84. DOI:10.1186/s40364-024-00611-y [Google Scholar]
-
Levrero M, Zucman-Rossi J. Mechanisms of HBV-Induced Hepatocellular Carcinoma. J. Hepatol. 2016, 64, S84–S101. DOI:10.1016/j.jhep.2016.02.021 [Google Scholar]
-
Shen X, Huang H, Sheng J, Tang X. HBV Reprograms the Tumor Microenvironment in Hepatocellular Carcinoma: Mechanisms and Therapeutic Implications. Clin. Exp. Med. 2026, 26, 148. DOI:10.1007/s10238-025-01851-4 [Google Scholar]
-
Wagle NS, Park S, Washburn D, Ohsfeldt RL, Rich NE, Singal AG, et al. Racial, Ethnic, and Socioeconomic Disparities in Curative Treatment Receipt and Survival in Hepatocellular Carcinoma. Hepatol. Commun. 2022, 6, 1186–1197. DOI:10.1002/hep4.1863 [Google Scholar]
-
Lawrence P, Chabane M, Abrouk L, Thiesson A, Berthé D, Diarra AB, et al. First Molecular Characterization of Chronic Hepatitis B Carriers in Timbuktu, Mali. Diagnostics 2023, 13, 375. DOI:10.3390/diagnostics13030375 [Google Scholar]
-
Coursaget P, Depril N, Chabaud M, Nandi R, Mayelo V, LeCann P, et al. High Prevalence of Mutations at Codon 249 of the P53 Gene in Hepatocellular Carcinomas from Senegal. Br. J. Cancer 1993, 67, 1395–1397. DOI:10.1038/bjc.1993.258 [Google Scholar]
-
Igetei R, Otegbayo JA, Ndububa DA, Lesi OA, Anumudu CI, Hainaut P, et al. Detection of P53 Codon 249 Mutation in Nigerian Patients with Hepatocellular Carcinoma Using a Novel Evaluation of Cell-Free DNA. Ann. Hepatol. 2008, 7, 339–344. DOI:10.1016/S1665-2681(19)31834-4 [Google Scholar]
-
Kirk GD, Camus-Randon A-M, Mendy M, Goedert JJ, Merle P, Trépo C, et al. Ser-249 P53 Mutations in Plasma DNA of Patients with Hepatocellular Carcinoma from The Gambia. JNCI J. Natl. Cancer Inst. 2000, 92, 148–153. DOI:10.1093/jnci/92.2.148 [Google Scholar]
-
Marchio A, Amougou Atsama M, Béré A, Komas N-P, Noah Noah D, Atangana PJA, et al. Droplet Digital PCR Detects High Rate of TP53 R249S Mutants in Cell-Free DNA of Middle African Patients with Hepatocellular Carcinoma. Clin. Exp. Med. 2018, 18, 421–431. DOI:10.1007/s10238-018-0502-9 [Google Scholar]
-
Wojnowski L, Turner PC, Pedersen B, Hustert E, Brockmöller J, Mendy M, et al. Increased Levels of Aflatoxin-Albumin Adducts Are Associated with CYP3A5 Polymorphisms in The Gambia, West Africa. Pharmacogenet. Genom. 2004, 14, 691–700. Available online: https://journals.lww.com/jpharmacogenetics/fulltext/2004/10000/Increased_levels_of_aflatoxin_albumin_adducts_are.7.aspx (accessed on 18 February 2026).
-
Ngamruengphong S, Patel T. Molecular Evolution of Genetic Susceptibility to Hepatocellular Carcinoma. Dig. Dis. Sci. 2014, 59, 986–991. DOI:10.1007/s10620-013-2984-3 [Google Scholar]
-
Sartorius K, Sartorius B, Winkler C, Chuturgoon A, Shen T-W, Zhao Y, et al. Serum microRNA Profiles and Pathways in Hepatitis B-Associated Hepatocellular Carcinoma: A South African Study. Int. J. Mol. Sci. 2024, 25, 975. DOI:10.3390/ijms25020975 [Google Scholar]
-
Bruni L, Diaz M, Castellsagué X, Ferrer E, Bosch FX, De Sanjosé S. Cervical Human Papillomavirus Prevalence in 5 Continents: Meta-Analysis of 1 Million Women with Normal Cytological Findings. J. Infect. Dis. 2010, 202, 1789–1799. DOI:10.1086/657321 [Google Scholar]
-
Magagi LH, Lagare A, Bowo-Ngandji A, Hassane F, Maiga AZ, Issa M, et al. Prevalence and Genotype Distribution of Human Papillomavirus (HPV) Infections among West African Populations: A Systematic Review and Meta-Analysis. Infect. Agents Cancer 2025, 20, 62. DOI:10.1186/s13027-025-00659-x [Google Scholar]
-
Maswanganye CK, Mkhize PP, Matume ND. Mapping the HPV Landscape in South African Women: A Systematic Review and Meta-Analysis of Viral Genotypes, Microbiota, and Immune Signals. Viruses 2024, 16, 1893. DOI:10.3390/v16121893 [Google Scholar]
-
Rifkin RF, Potgieter M, Ramond J, Cowan DA. Ancient Oncogenesis, Infection and Human Evolution. Evol. Appl. 2017, 10, 949–964. DOI:10.1111/eva.12497 [Google Scholar]
-
Rosendo-Chalma P, Antonio-Véjar V, Ortiz Tejedor JG, Ortiz Segarra J, Vega Crespo B, Bigoni-Ordóñez GD. The Hallmarks of Cervical Cancer: Molecular Mechanisms Induced by Human Papillomavirus. Biology 2024, 13, 77. DOI:10.3390/biology13020077 [Google Scholar]
-
Anorlu RI. Cervical Cancer: The Sub-Saharan African Perspective. Reprod. Health Matters 2008, 16, 41–49. DOI:10.1016/S0968-8080(08)32415-X [Google Scholar]
-
Ba DM, Ssentongo P, Musa J, Agbese E, Diakite B, Traore CB, et al. Prevalence and Determinants of Cervical Cancer Screening in Five Sub-Saharan African Countries: A Population-Based Study. Cancer Epidemiol. 2021, 72, 101930. DOI:10.1016/j.canep.2021.101930 [Google Scholar]
-
Picozzi K, Oliver CM. Barriers and Facilitators of Sexually Transmitted Infection Control in Maasai Communities: A Qualitative Systematic Review. Available online: https://era.ed.ac.uk/handle/1842/39472 (accessed on 18 February 2026).
-
Watson-Jones D, Mugo N, Lees S, Mathai M, Vusha S, Ndirangu G, et al. Access and Attitudes to HPV Vaccination amongst Hard-To-Reach Populations in Kenya. PLoS ONE 2015, 10, e0123701. DOI:10.1371/journal.pone.0123701 [Google Scholar]
-
Haddison EC, Engoung DB, Bodo CB, Njie VM. Overcoming HPV Vaccine Hesitancy: Insights from a Successful School-Based Vaccination Campaign in the Saa Health District of Cameroon. BMC Infect. Dis. 2025, 25, 465. DOI:10.1186/s12879-025-10864-z [Google Scholar]
-
Sayinzoga F, Tenet V, Heideman DAM, Sibomana H, Umulisa M-C, Franceschi S, et al. Human Papillomavirus Vaccine Effect against Human Papillomavirus Infection in Rwanda: Evidence from Repeated Cross-Sectional Cervical-Cell-Based Surveys. Lancet Glob. Health 2023, 11, e1096–e1104. DOI:10.1016/S2214-109X(23)00193-6 [Google Scholar]
-
Adeyanju GC, Essoh T-A, Sidibe AR, Kyesi F, Aina M. Human Papillomavirus Vaccination Acceleration and Introduction in Sub-Saharan Africa: A Multi-Country Cohort Analysis. Vaccines 2024, 12, 489. DOI:10.3390/vaccines12050489 [Google Scholar]
-
Amani A, Bloem P, Kobayashi E, Atuhaire B, Wiysonge CS, Impouma B. Scaling HPV vaccination in Africa to eliminate cervical cancer by 2030. Lancet Glob. Health 2025, 13, 12, e2006–e2008. DOI:10.1016/S2214-109X(25)00349-3 [Google Scholar]
-
Arhin N, Ssentongo P, Taylor M, Olecki EJ, Pameijer C, Shen C, et al. Age-Standardised Incidence Rate and Epidemiology of Colorectal Cancer in Africa: A Systematic Review and Meta-Analysis. BMJ Open 2022, 12, e052376. DOI:10.1136/bmjopen-2021-052376 [Google Scholar]
-
Yildiz S, Chambuso R, Rebello G, Ramesar R. High Burden of Variants of Uncertain Significance in Early-Onset Colorectal Cancer among Indigenous African Patients: A Call for Global Research Equity in Cancer Genetics. Mol. Biol. Rep. 2025, 52, 684. DOI:10.1007/s11033-025-10750-6 [Google Scholar]
-
Tazinkeng NN, Pearlstein EF, Manda-Mapalo M, Adekunle AD, Monteiro JFG, Sawyer K, et al. Incidence and Risk Factors for Colorectal Cancer in Africa: A Systematic Review and Meta-Analysis. BMC Gastroenterol. 2024, 24, 303. DOI:10.1186/s12876-024-03385-7 [Google Scholar]
-
Moen L, Liu B, Bukirwa P, Chingonzoh T, Chokunonga E, Finesse A, et al. Trends in the Incidence of Colorectal Cancer in Sub-Saharan Africa: A Population-Based Registry Study. Int. J. Cancer 2024, 155, 675–682. DOI:10.1002/ijc.34942 [Google Scholar]
-
Novillo A, Romero-Lorca A, Gaibar M, Bahri R, Harich N, Sánchez-Cuenca D, et al. Genetic Diversity of CYP3A4 and CYP3A5 Polymorphisms in North African Populations from Morocco and Tunisia. Int. J. Biol. Markers 2015, 30, 148–151. DOI:10.5301/jbm.5000118 [Google Scholar]
-
Wismayer R, Kiwanuka J, Wabinga H, Odida M. Risk Factors for Colorectal Adenocarcinoma in an Indigenous Population in East Africa. Cancer Manag. Res. 2022, 14, 2657–2669. DOI:10.2147/CMAR.S381479 [Google Scholar]
-
Alatise OI, Knapp GC, Bebington B, Ayodeji P, Dare A, Constable J, et al. Racial Differences in the Phenotype of Colorectal Cancer: A Prospective Comparison Between Nigeria and South Africa. World J. Surg. 2022, 46, 47–53. DOI:10.1007/s00268-021-06304-z [Google Scholar]
-
McCabe M, Penny C, Magangane P, Mirza S, Perner Y. Left-Sided Colorectal Cancer Distinct in Indigenous African Patients Compared to Other Ethnic Groups in South Africa. BMC Cancer 2022, 22, 1089. DOI:10.1186/s12885-022-10185-3 [Google Scholar]
-
Losso GM, Moraes RDS, Gentili AC, Messias-Reason IT. Microsatellite Instability—MSI Markers (BAT26, BAT25, D2S123, D5S346, D17S250) in Rectal Cancer. Arq. Bras. Cir. Dig. 2012, 25, 240–244. DOI:10.1590/s0102-67202012000400006 [Google Scholar]
-
Yildiz S, Musarurwa TN, Algar U, Chambuso R, Rebello G, Goldberg PA, et al. Genetic Insights: High Germline Variant Rate in an Indigenous African Cohort with Early-Onset Colorectal Cancer. Front. Oncol. 2023, 13, 1253867. DOI:10.3389/fonc.2023.1253867 [Google Scholar]
-
McCabe M, Perner Y, Magobo R, Magangane P, Mirza S, Penny C. Microsatellite Instability Assessment in Black South African Colorectal Cancer Patients Reveal an Increased Incidence of Suspected Lynch Syndrome. Sci. Rep. 2019, 9, 15019. DOI:10.1038/s41598-019-51316-4 [Google Scholar]
-
Alatise OI, Knapp GC, Sharma A, Chatila WK, Arowolo OA, Olasehinde O, et al. Molecular and Phenotypic Profiling of Colorectal Cancer Patients in West Africa Reveals Biological Insights. Nat. Commun. 2021, 12, 6821. DOI:10.1038/s41467-021-27106-w [Google Scholar]
-
Irabor D. Ethnic Differences in Colon and Rectal Cancer Incidence in Nigeria: A Case of Dietary Determinants? Ann. Niger. Med. 2012, 6, 71. DOI:10.4103/0331-3131.108123 [Google Scholar]
-
Garba SM, Hami H, Zaki HM, Soulaymani A, Nouhou H, Quyou A. P0190Colorectal Cancer in Niger: An Analysis of the Niger Cancer Registry Data. Eur. J. Cancer 2014, 50, e63. DOI:10.1016/j.ejca.2014.03.234 [Google Scholar]
-
Ghazali N, Brown C, Pugh T, Hung R, Okonta K, Saleh M, et al. Liquid Biopsies of Lung Cancer and Next Generation Sequencing at Sub-Saharan Africa Sites (LUNGS@AFRICA). J. Liq. Biopsy 2025, 9, 100373. DOI:10.1016/j.jlb.2025.100373 [Google Scholar]
-
Rojiani MV, Rojiani AM. Non-Small Cell Lung Cancer—Tumor Biology. Cancers 2024, 16, 716. DOI:10.3390/cancers16040716 [Google Scholar]
-
Leiter A, Veluswamy RR, Wisnivesky JP. The Global Burden of Lung Cancer: Current Status and Future Trends. Nat. Rev. Clin. Oncol. 2023, 20, 624–639. DOI:10.1038/s41571-023-00798-3 [Google Scholar]
-
Da Cunha Santos G, Shepherd FA, Tsao MS. EGFR Mutations and Lung Cancer. Annu. Rev. Pathol. Mech. Dis. 2011, 6, 49–69. DOI:10.1146/annurev-pathol-011110-130206 [Google Scholar]
-
Errihani H, Inrhaoun H, Boukir A, Kettani F, Gamra L, Mestari A, et al. Frequency and Type of Epidermal Growth Factor Receptor Mutations in Moroccan Patients with Lung Adenocarcinoma. J. Thorac. Oncol. 2013, 8, 1212–1214. DOI:10.1097/JTO.0b013e31829f6b4a [Google Scholar]
-
El Zaitouni S, Laraqui A, Ghaouti M, Benzekri A, Kettani F, Boustany Y, et al. Genetic Profiling of Non-Small Cell Lung Cancer in Moroccan Patients by Targeted Next-Generation Sequencing. Technol. Cancer Res. Treat. 2024, 23, 15330338241288907. DOI:10.1177/15330338241288907 [Google Scholar]
-
Arfaoui Toumi A, Blel A, Aloui R, Zaibi H, Ksentinini M, Boudaya MS, et al. Assessment of EGFR Mutation Status in Tunisian Patients with Pulmonary Adenocarcinoma. Curr. Res. Transl. Med. 2018, 66, 65–70. DOI:10.1016/j.retram.2018.02.004 [Google Scholar]
-
Boustany Y, Laraqui A, El Rhaffouli H, Bajjou T, El Mchichi B, El Anaz H, et al. Prevalence and Patterns of EGFR Mutations in Non-Small Cell Lung Cancer in the Middle East and North Africa. Cancer Control 2022, 29, 10732748221129464. DOI:10.1177/10732748221129464 [Google Scholar]
-
Holt CL, Hussain A, Wachbroit R, Scott J. Precision Medicine Across the Cancer Continuum: Implementation and Implications for Cancer Disparities. JCO Precis. Oncol. 2017, 1, 1–5. DOI:10.1200/PO.17.00102 [Google Scholar]
-
Yakubu A, Munung NS, De Vries J. How Should Biobanking Be Governed in Low-Resource Settings? AMA J. Ethics 2020, 22, E156–E163. DOI:10.1001/amajethics.2020.156 [Google Scholar]
-
Ofori-Acquah SF, Nuno M, Osae-Larbi JA, Tavares K, Osei-Tutu A, Barkers Ansah DA, et al. The West African Genetic Medicine Center as a Model to Address the Challenge of Genetic Disorders in Africa. Nat. Genet. 2026, 58, 467–471. DOI:10.1038/s41588-026-02509-x [Google Scholar]
-
Joubert BR, Berhane K, Chevrier J, Collman G, Eskenazi B, Fobil J, et al. Integrating Environmental Health and Genomics Research in Africa: Challenges and Opportunities Identified during a Human Heredity and Health in Africa (H3Africa) Consortium Workshop. AAS Open Res. 2019, 2, 159. DOI:10.12688/aasopenres.12983.1 [Google Scholar]