1. Introduction
Fibrosis, or fibrotic scarring, is a pathological process that if left unchecked can result in tissue remodeling and the formation of a permanent scar. Its mechanism is very similar to wound healing, however in contrast, wound healing is the controlled deposition of collagens. Once the wound is closed collagen is no longer secreted in abundance. Fibrosis can manifest in a single organ or can be systemic affecting many organs and tissues. Numerous different insults can contribute to the development of fibrosis. In many cases the causative agent is unknown, however, in some cases, fibrosis can be attributed to pathogens [
1,
2,
3], environmental factors [
4,
5,
6], genetics [
7,
8], autoimmunity [
9,
10,
11], or cancer [
12,
13]. Furthermore, the risk of developing a fibrotic disorder significantly increases with increasing age [
14,
15]. Depending on the organ affected, there can be significant morbidity, and this can lead to death.
Much has been learned in recent years about various mechanisms that drive fibrosis, and this has largely centered on the myofibroblast. Myofibroblasts have heterogeneous origins (Reviewed in [
16,
17,
18]), however, the most commonly described process is the differentiation of fibroblasts into myofibroblasts. During fibroblast to myofibroblast differentiation, there are numerous phenotypic changes to the cell. The endoplasmic reticulum (ER) becomes expanded and there is increased production of proteins, especially those belonging to the extracellular matrix family. The expanded ER is partly due to the role of ER stress in these cells [
19], but it is also because there is increased folding and processing of procollagen and other large extracellular matrix proteins requiring export out of the ER. Under ER stress, the spliced form of XBP1 (X-box binding protein-1) enhances the transcription of various genes, allowing the ER to better cope with elevated levels of unfolded proteins. The spliced form of XBP1 also directly promotes ER/Golgi expansion and functions to control the secretory phenotype of the cell [
20,
21,
22,
23]. XBP1 also helps to increase the number and size of ER exit sites [
24], induces a subset of proteins involved in vesicular trafficking and transport such as TANGO1 [
25], and promotes Coat Protein Complex II (COPII) gene expression [
26]. Overall, this helps the ER to deal with the elevated folding and processing of proteins by enhancing their export out of the ER.
2. Components of the COPII Vesicle
Approximately one-third of the proteins synthesized in the ER traffic to the Golgi. Many of these proteins are destined for secretion, or they reside within the various organelles in the cell. Proteins accumulate at ER exit sites for export [
27]. These regions are devoid of ribosomes but are rich with COPII coat proteins and have between 2 to 6 stable COPII buds [
28]. ER exit sites function as departure gates for proteins, pushing them out to other regions in the cell. Thus, the formation of the COPII vesicle for protein export is crucial to this process. The COPII vesicle includes two proteinaceous layers. These layers can deform the ER membrane to accommodate the protein cargo. The inner layer contains the Sar1-GTPase and the heterodimer Sec23/Sec24 lattice, while the outer layer comprises of a the rod-shaped heterotetramer made of Sec13/Sec31 proteins. These vesicles are assembled and disassembled as needed [
29].
To start the formation of a COPII vesicle, the inner coat of the vesicle begins to assemble when Sar1-GDP is activated by the guanine nucleotide exchange factor Sec12. Sec12 loads GTP onto Sar1 causing it to bury its amphipathic helix into the outer ER membrane [
30]. Sar1-GTP then engages Sec23/Sec24, which starts forming the inner coat of the vesicle. Sec23 directly interacts with Sar1. However, Sec23 also engages with Sec13/Sec31, and this causes the formation of the outer cage. In this complex, the role of Sec24 is the recruitment of various receptors for the binding of a variety of cargo. Recent studies show that the outer cage is necessary for the stabilization of the inner membrane [
31]. Once the vesicle is large enough (~60–70 nm in size), it becomes unstable and the GTPase activity of Sec23 then hydrolyzes Sar1-GTP. This destabilizes the vesicle, and it buds off holding the cargo.
While overall this process is efficient for the export of smaller proteins, eukaryotic cells have evolved additional mechanisms for the export of much larger proteins such as collagens. TANGO1 (Transport ANd Golgi Organization protein 1, also called MIA3 (Melanoma Inhibitory Activity protein 3), and its paralogs in concert with other proteins have evolved to capture large proteins and expand the vesicle to hold that captured protein. The role of these proteins in the export of collagen is discussed below.
3. The Function of the TANGO1 Family of Proteins at ER Exit Sites
The synthesis of procollagen has been extensively studied. However, extraordinarily little is known about the export of procollagen from the ER in the context of fibrosis. The export of large proteins such as collagen is crucial under basal conditions and increased export is especially important during wound healing. Once the procollagen is folded, it must exit the ER in COPII vesicles and travel to the Golgi for further modifications. Here there is a size paradox as many of the large extracellular proteins are too big to fit into regular-sized COPII vesicles. The export of procollagen VII from the ER was first elucidated in
Drosophila and was able to shed some light on this paradox [
32]. These first fly studies found that TANGO1 and cTAGE5 (Cutaneous T-cell lymphoma-Associated antiGen 5) interacted together to create a mega-COPII transport carrier large enough to hold procollagen. More recently, other proteins have been shown to interact with TANGO1 to delay the closure of the vesicle allowing for the accommodation of procollagen.
TANGO1 is a resident protein found in ER exit sites. It is coded on chromosome 1 and has two well-studied functional isoforms, TANGO1-Long and TANGO1-Short (), although there are several additional splice variants with minor alterations (see Genbank). Whether these other variants have a functional role or affect protein export is currently unknown.
Saito et al. [
32], showed that TANGO1 helps to orchestrate the loading of procollagen into growing vesicles. Furthermore, other studies by Saito et al. [
33], demonstrate that TANGO1 also functions to organize the ER exit sites and this is a necessary step in protein export. There is controversy, however, as to how procollagen moves between the ER and Golgi. Several investigators have reported enlarged COPII vesicles are responsible while others suggest tubular-saccular structures are involved [
32,
34,
35]. Irrespective of the mode of export out of the ER, TANGO1 plays a crucial role in this process.
. Topology of TANGO1 (Long and Short), TALI, and cTAGE5. (
A) TANGO1 isoforms. (
B) The TANGO1 paralog and its isoforms. The TANGO1 isoforms and paralogs embed themselves in the ER membrane using a transmembrane helix. Except for cTAGE5, there is an additional helix that partially embeds into the inner leaflet of the ER membrane to help stabilize their interaction when recruiting ERGIC membranes and procollagen. On the cytoplasmic side of the ER membrane, the proline-rich domain (PRD) binds to Sec23 and Sec16, which helps to start the formation of the COPII vesicle. The coiled-coil (CC)1 domain recruits the NBAS/RINT1/ZW10 (NRZ) tethering complex which drives the recruitment of ERGIC-53 membranes. The CC2 domain interacts with other members of the TANGO1 family; cTAGE5, TALI, and TANGO1-Short. On the lumenal side of the ER, the SH3-like domain binds HSP47 which then recruits procollagen into the complex. Figure adapted from Raote et al., 2018 [
36]. Created in BioRender.com.
TANGO1-Long is a large protein consisting of 1907 amino acids. Approximately one-third of the protein is found on the cytoplasmic side of the ER. It has a transmembrane helical domain with an additional helical membrane loop that partially anchors itself into the leaflet of the ER membrane (). On the lumenal side of the ER, TANGO1-Long has a SH3-like domain [
37] that binds HSP47. It is the HSP47 that recruits procollagen into the growing vesicle. HSP47 is a critical collagen-specific ER chaperone that is upregulated in parallel with COL1A1 expression [
38,
39]. Studies show that the SH3 domain of TANGO1 can barely recognize and bind procollagen [
40], while it is the HSP47 that strongly binds to the native type I, II, III, IV, V, VI, and X procollagens [
39] and as a result recruits them into the growing vesicle.
TANGO1-Long interacts directly with cTAGE5 [
34,
40] and Sec12 [
40]. The interaction with Sec12 helps to delay the closure of the vesicle. TANGO1-Long also has a binding site that can recruit TANGO1-S [
36] and its paralog isoform TALI [
41]. In keeping with the recruitment of procollagen into the growing vesicle, TANGO1-Long also recruits Sec23, allowing for the envelope to increase in size. Sec23 also co-localizes with procollagen at these sites. TANGO1-Long can also homodimerize with itself [
36,
42].
3.2. TANGO1-Short
The isoform, TANGO1-Short is 785 amino acids in length and also embeds into the ER membrane. TANGO1-Short contains the cytoplasmic domains of TANGO1-Long but lacks the HSP47 binding domain and therefore cannot bind directly to procollagen. However, TANGO1-Short is involved in procollagen export [
40]. It is thought that TANGO1-Short helps to recruit the ERGIC membranes with the role of aiding in the expansion of the vesicles, allowing for procollagen to be incorporated [
34,
43]. TANGO1-Short competes with Sec31 for binding to Sec23. Because of this competition, there is a delay in the fission of the vesicle. TANGO1-Short has been shown to form a protein complex with cTAGE5 and Sec12 [
40]. Elegant studies by Maeda et al. showed that TANGO1-Short can form homotrimers [
40].
When both TANGO1-Long and TANGO1-Short are knocked down in cells, there is dissociation of Sec31 from the ER exit site [
40] with subsequent destabilization of the exit site [
43]. Furthermore, the depletion of both isoforms led to the decreased expression of cTAGE5 and the loss of Sec12 from the exit site [
40].
3.3. TALI and cTAGE5
TANGO1 has a paralog located on chromosome 14. The TANGO1 duplication results in the protein TANGO1-like protein, which also has two isoforms (). The larger isoform TALI is located in the liver and was found to be crucial in the export of apolipoprotein B [
41]. Apolipoprotein B has a molecular weight of 540 kDa and is likely to be one of the largest proteins to ever be made by any cell. TALI was also found to be expressed in tissues devoid of apolipoprotein synthesis such as the lung, testes, small intestine, colon, pancreas, kidney, and prostate [
41]. This suggests that TALI may be involved in the export of other large proteins. Since TALI has a HSP47 binding domain we speculate that it could also be involved in the export of collagen although to date there is no evidence for this. The short isoform of TALI, now called cTAGE5, is ubiquitously expressed and is also involved in procollagen export. The export of apolipoprotein B requires both TANGO1 and TALI, and the export of procollagen requires TANGO1 and cTAGE5.
cTAGE5 lacks a lumenal domain and therefore cannot bind directly to cargo, however, cTAGE5 interacts with TANGO1 via its CC2 domain and interacts with Sec12 via its CC1 domain. It is this binding of Sec12 and the increased recruitment of the inner COPII-coat proteins Sec23/24, concordant with the increased recruitment of outer-coat proteins Sec13/31 that helps to create larger vesicles that accommodate procollagen. cTAGE5 is the central (core) protein in cargo export. It functions as a cargo receptor via its engagement with TANGO1 (Long and Short) and it acts as a scaffolding protein for the recruitment of Sec12 [
44]. The function of cTAGE5 was determined by the integration of point mutations in the protein where cTAGE5 failed to bind to Sec12, and as a result, collagen could not be exported [
44].
Maeda et al. [
40] reported that cTAGE5 together with Sec12 formed distinct complexes with TANGO1-Long and TANGO1-Short. Together these complexes were found to take part in the export of procollagen. The depletion of both TANGO1-Long and -Short reduced the expression of cTAGE5 without affecting the expression of Sec12. However, it was noted that Sec12 was no longer localized into the ER exit site when cTAGE5 was depleted [
40]. Both TANGO1-Long/cTAGE5/Sec12 and TANGO1-Short/cTAGE5/Sec12 were found to be stable complexes within the ER exit sites. In the absence of TANGO1-Long, TANGO1-Short is sufficient to interact with cTAGE5 and Sec12 for the export of procollagen [
40].
4. The Interaction of Other Proteins with TANGO1 for Procollagen Export
There are numerous other proteins directly or indirectly involved in the export of procollagen or other larger bulky proteins. They primarily function by delaying the closure of the vesicle so that these large proteins can be incorporated before they bud off.
4.1. COPII Proteins
Sec23 has two isoforms (Sec23A and Sec23B) and mutations in Sec23 have been associated with various diseases [
45,
46,
47]. Mutations in Sec23A at amino acid 382 resulted in a nonsynonymous change from phenylalanine to leucine that caused craniolenticulosutural dysplasia and defects in procollagen export [
46,
47]. This observation was recapitulated in Sec23-KO mice, which exhibited embryonic lethality due to impaired collagen secretion [
48].
Sec24 has four isoforms that can stably interact with both of the Sec23 isoforms. They have a hierarchical preference for procollagens. Lu et al. [
49] showed that procollagen interacts with multiple Sec24 isoforms, but Sec24A and Sec24D played a major role in the export of procollagen. Sec24B and Sec24C can also export procollagen but to a lesser degree [
49].
To date, there are no isoforms of Sec13, however, mutations within the Sec13 gene caused defects in protein secretion. Zebrafish depleted of Sec13 had developmental impairment of retinal and gut tissues associated with a defect in procollagen export [
50,
51]. In mammals, a Sec13-KO is embryonic lethal [
52].
There are two isoforms of Sec31, Sec31A and Sec31B. Sec31A is generally expressed more than Sec31B [
53], but there is differential expression of Sec31A and Sec31B in various tissues. Sec31A is highly expressed in the ovary [
53] and Sec31B is highly expressed in the kidney [
53]. Mutations in Sec31A result in skeletal abnormalities [
54] suggesting a direct role in procollagen export.
Overall, the coupling of Sec23 to Sec24 and Sec13 to Sec31 is crucial for the export of procollagen. Sec13 stabilizes Sec31 [
55]. In a study using zebrafish the depletion of Sec13/Sec31 halted the formation of the COPII vesicle causing distension of the ER [
55] and retention of procollagen, while smaller cargoes were unaffected.
TANGO1 directly binds to only one of the COPII proteins. This binding would help to feed the collagen molecule directly into the growing vesicle. It directly binds to the inner layer protein Sec23, and then via this engagement Sec23 binds to Sec24. Sec13/Sec31 are recruited by Sec24. The interaction between TANGO1 and Sec23 occurs via the gelsolin-like domain of Sec23 and the repeated phosphoprotein phosphatase motifs in the TANGO1 cytosolic region [
34]. Structural abnormalities and mutations in COPII proteins reported above would result in an abnormal COPII coat that cannot contain large macromolecules like collagen. This would likely explain the phenotypic abnormalities seen in mice and fish.
4.2. KLHL12 and CUL3
Kelch Like Family Member-12 (KLHL12), is a Bric-a-brac, Tram-track, and Broad-complex (BTB)–domain protein also involved in the export of large cargo molecules such as procollagen. Cullin-3 (CUL3)-RING ubiquitin ligase engages KLHL12 causing the monoubiquitylation of Sec31 [
56]. This monoubiquination delays the closure of the vesicle, allowing for more procollagen to be incorporated. Studies have also shown the neddylation of CUL3 to be important in the process of delaying vesicle closure [
57], whereas others have shown it not to be important [
58]. The mild over-expression of KLHL12 leads to the formation of large COPII vesicles [
56], suggesting a direct role for KLHL12/CUL3 in procollagen export. For CUL3 and KLHL12 to function properly, two calcium-binding proteins are required. These are PEF1 (Penta-EF-Hand Domain Containing 1 or peflin) and ALG2 (Alpha-1,3/1,6-mannosyltransferase) and aid in CUL3/KLHL12 recognition of Sec31. PEF1 and ALG2 dimerize to form a target-specific adaptor that causes a transient rise in cytosolic calcium levels. The increased calcium levels result in a persistence of Sec31 ubiquitylation [
59,
60], which in turn helps to trigger the formation of larger COPII vesicles. Calcium is a very important signaling molecule, especially in collagen synthesis [
61]. Overall, this calcium-dependent regulation of CUL3/KLHL12 integrates collagen synthesis with secretion by interacting with the outer leaflet of the COPII vesicle which is composed of Sec13/Sec31 [
62]. With TANGO1 anchored in the ER membrane, KLHL12/CUL3 functions to help regulate the size of the COPII transport vesicle [
56].
4.3. Sedlin
Sedlin is a TRAPPC2 protein (TRAnsport Protein Particle C2) that aids in the targeting or fusion of ER-to-Golgi transport vesicles. Sedlin interacts directly with TANGO1 and regulates the efficient cycling of Sar1. If Sedlin is knocked down, this causes the premature closure of the vesicle leading to less incorporation of collagen and smaller vesicles [
62,
63].
4.4. Sar1
Sar1 is bound into the procollagen export machinery via its engagement with Sec12 which is directly bound by TANGO1 [
64]. Sar1 has two isoforms, Sar1A and Sar1B, which share about 90% sequence identity [
65]. Sar1 isoforms have a unique amphipathic amino-terminal helix but have to be activated by Sec12 to stably penetrate the bilayer to start the remodeling and deformation of the ER membrane [
31]. Studies suggest that the expression of Sar1A and Sar1B are important for vesicle size and each isoform delays the closure of the vesicle at different rates. Sar1A is required for the formation of vesicles and controls membrane deformation so that vesicles can bud off when they reach the correct size. Sar1A engages with the GTPase of Sec31 more strongly than Sar1B does and therefore likely creates smaller vesicles due to the increased GTPase activity of Sec31. Therefore, for smaller cargoes, the faster kinetics mediated by Sar1A would allow for the export of these proteins. However, Sar1B binds to Sec23 more strongly than does Sar1A [
66]. Sar1B is needed for the export of chylomicrons and patients with mutations in Sar1B have disorders in fat malabsorption [
65,
67]. In zebrafish, deficiency in Sar1B causes deficits in extracellular matrix collagen and the intracellular accumulation of procollagen [
68]. Overall, these observations suggest that Sar1B is also an important protein that helps to regulate the size of the COPII vesicle for the incorporation of larger-sized proteins.
4.5. Sec12
TANGO1 also interacts with Sec12 in the ER exit site and this interaction also helps to delay the closure of the vesicle causing the expansion of the COPII vesicle [
69]. Along with the procollagen, TANGO1 and Sec12 are exported out of the ER in the enlarged COPII vesicles and then recruited back to the ER via COPI-mediated retrograde transport [
69]. HSP47 facilitates this process of retrieval of TANGO1 back to the ER. Sec12 also interacts with cTAGE5 [
64] which then interacts with TANGO1 [
70]. However, another study suggested that TANGO1 was retained in the ER [
71].
4.6. Sec16
During the cell cycle, protein export from the ER is impeded [
72], however, until now it was not fully understood as to how this happened. Maeda et al. recently discovered the role of TANGO1 and Sec16 in this process. During mitosis, ER exit sites are disassembled [
73]. It has been shown that Sec16 is an organizer of the ER exit sites, but it is considered to be a peripheral membrane protein. Therefore, it must be recruited to the ER exit site for it to function as a scaffolding protein with other COPII components; while TANGO1-Long and TANGO1-Short span the ER membrane at ER exit sites. TANGO1-Long also interacts with Sec16 [
32,
70]. Maeda et al. demonstrated that TANGO1 recruits Sec16 into the ER exit site, where they interact with the organization of the site [
74]. Intriguingly, during the cell cycle, TANGO1 is hyperphosphorylated and this reduces its affinity for Sec16. It is believed that this loss in Sec16/TANGO1 interaction causes the disassembly of the ER exit site [
75].
5. TANGO1 and Fibrosis
Overall, the export of procollagen from the ER is poorly characterized in mammals and under-studied in fibrosis and other collagenopathies. The first study to report an increase and role for TANGO1 in any fibrotic disease was published by Maiers et al., [
25] who elegantly showed that TANGO1 was elevated in hepatic stellate cells leading to fibrosis. They further went on to prove that the depletion of TANGO1 blocked type I collagen (COL1A1) secretion, but in their experiments the export of other matrix proteins was unaffected. Depletion of TANGO1 led to the retention of procollagen in the ER. This caused the unfolded protein response and apoptosis. They further went on to find that the unfolded protein response caused the upregulation of TANGO1 by TGFβ, and this was mediated by X-box binding protein 1 (XBP1) [
25].
Our laboratories then studied TANGO1 in systemic sclerosis [
76]. Systemic sclerosis (SSc) is an autoimmune fibrotic disorder that primarily affects women. In the fibrotic lesions, there are persistently activated myofibroblasts. These activated fibroblasts drive the excessive secretion and deposition of collagen and extracellular matrix proteins in the skin, vasculature, and internal organs [
77,
78,
79,
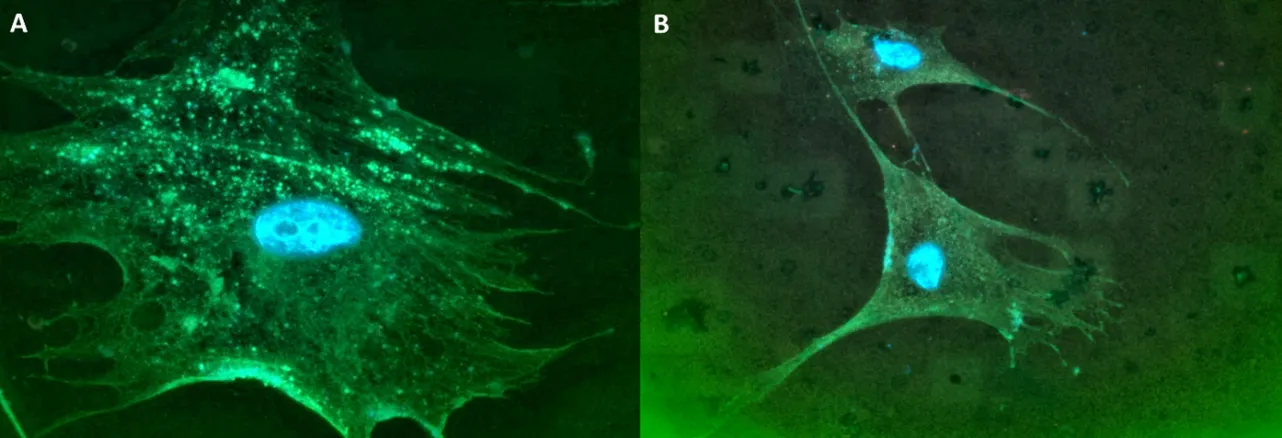
80]. Myofibroblasts have the increased ability to secrete abundant amounts of protein and produce more collagens and other ECM proteins compared to quiescent fibroblasts. Therefore, to meet this increased secretory demand, myofibroblasts must depend on the ER to process and export the augmented protein load. We stained myofibroblasts derived from fibrotic lesions for TANGO1 and found there were increased positive punctates in fibroblasts isolated from patients with SSc ().
Given that there is increased procollagen export from the ER during fibrosis, we reasoned that there also had to be elevated COPII proteins. Using Sec13 as a marker for COPII proteins, we found that SSc fibroblasts also had more distinct punctates of Sec13 compared to normal fibroblasts ().
We previously showed elevated TANGO1 in lung biopsies isolated from patients with SSc [
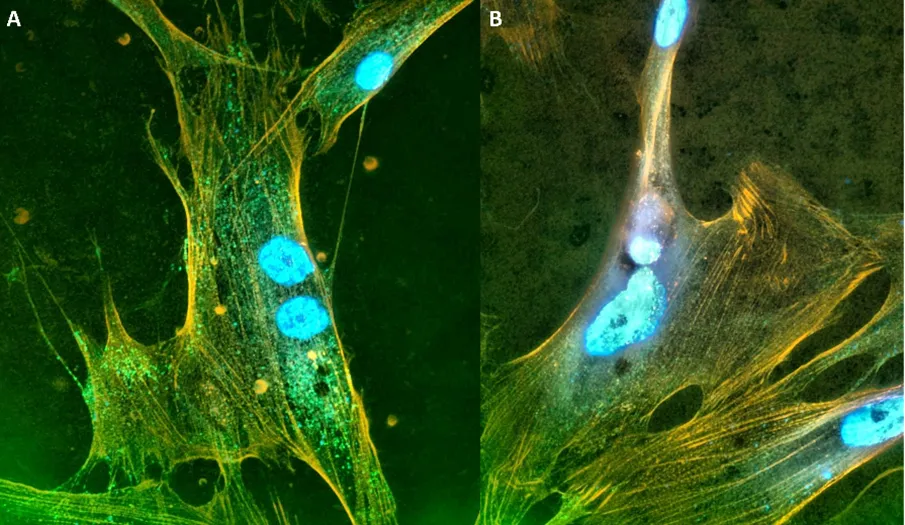
76]. In the neointima of vessels from SSc patients, we saw that TANGO1 was co-localization with α-smooth muscle actin, which is suggestive of increased TANGO1 in vascular myofibroblasts (). We especially noted this co-localization occurred in the neointima of occluded vessels (A). In control samples, TANGO1 was not found in the vessels while α-smooth muscle actin was abundantly present (B).
We further went on to show that TANGO1-Long and TANGO1-Short are significantly elevated in SSc fibroblasts [
76] and that their expression is dependent on the activity of caspase-1. This strongly suggests the role of the inflammasome in TANGO1 expression. We further show that inhibition of the TGF-β receptor lowered both collagen and TANGO1-Long and -Short. The inhibition of the IL-1 receptor lowered TANGO1-Long and TANGO1-Short but caused the retention of collagen in the cells. In our study, we speculated that the lowered collagen expression mediated by blocking of the TGF-β receptor took pressure off the ER for the export of collagen and therefore TANGO1 expression went down. Whereas inhibition of the IL-1 receptor decreased TANGO1 expression but did not affect COL1A1 expression and therefore collagen was retained in the cell. We also found that inhibition of TANGO1 with the caspase-1 inhibitor or blockade of the TGF-β or IL-1 receptors reduced the total amounts of secreted proteins by the SSc fibroblasts suggesting that TANGO1 is also involved in the export of other proteins, not just collagen. We observed that the high molecular weight proteins were more affected than low molecular weight proteins [
76]. However, we did not test to determine if the high molecular weight proteins were more likely to be retained within the cell or if these proteins also went down with the blockade of the receptors.
. TANGO1 punctates in fibroblasts derived from SSc lesions. (A) Fibroblasts isolated from the lung from SSc or (B) normal lung tissues were stained for TANGO1 (green) and stained for f-actin (orange) using phalloidin. The nuclei were stained blue with DaPI. 1000× magnification.
. SSc fibroblasts have enlarged Sec13 punctates. SSc (A) and normal (B) fibroblasts were stained for Sec13 (green). Note the increased positive green punctates in the enlarged SSc fibroblast, compared to the normal fibroblast. 1000× magnification.
. Increased expression of TANGO1 with α-smooth muscle actin in SSc vessels. (
A) Lung tissue was isolated from a patient with SSc undergoing lung transplants or (
B) from a normal individual who had died of unrelated pulmonary events and was dual stained for TANGO1 (red) and α-smooth muscle actin (green). Only vessels are shown. Note the increased expression of TANGO1 (red) corresponding with α-smooth muscle actin (green) in the neointima of a vessel derived from a patient with SSc (
A), while no TANGO1 is present in the α-smooth muscle actin stained vessel derived from normal lung tissue in (
B) [
76].
6. The Loss of Functional TANGO1 Correlates with Disease Phenotypes
Mutations in the TANGO1 gene have been associated with various diseases. In a study on clinically diagnosed Ehler’s-Danlos syndrome who were negative for the classic mutations in connective tissue genes, one hundred patients were tested for mutations in TANGO1 [
81]. The authors found 5 variations in the
TANGO1 gene, 4 were missense mutations and 1 was a frameshift mutation. All mutations were determined to be pathogenic. All the genetic alterations found were heterozygous and absent in the control group. The patient with the truncated form of TANGO1 had skeletal abnormalities and this was associated with scoliosis, osteopenia, brachydactyly and clinodactyly, dentinogenesis imperfecta, and mild intellectual disability. This individual also had a substantial reduction in COL1A1 secretion [
81]. Another study found a homozygous synonymous substitution in the
TANGO1 gene leading to exon 8 skipping and the formation of a truncated TANGO1 protein. This truncated protein also affected COL1A1 secretion. This mutation was found in a consanguineous family [
82]. Affected individuals in this family had dentinogenesis imperfecta delayed eruption of the permanent teeth, numerous skeletal abnormalities, and mild intellectual disability [
82]. In another consanguineous family from India, the fetus presented with early lethality and an almost complete absence of bone formation. Whole-exome sequencing was performed and showed a novel homozygous frameshift variant of the
TANGO1 gene. This caused a premature termination codon and the complete absence of the TANGO1 protein [
83].
The loss of a functional TANGO1 has been recapitulated in zebrafish and mice and confirms many of the findings seen in human diseases. When TANGO1 was depleted in zebrafish, the fish were significantly shorter in length with craniofacial defects. They did not survive to adulthood [
84]. Mice lacking the
TANGO1 gene also showed chondrodysplasia and dwarfing of fetal pups. There was a shortening of the snout and limbs, compounded by tissue fragility. The complete absence of an ossified skeleton led to death [
85]. Intriguingly, a new syndrome was reported in Cane Corso dogs with hereditary dental pathology that demonstrated a homozygote splice mutation that caused exon skipping in the
TANGO1 gene. The authors propose that there was some residual TANGO1 protein activity that allowed for the export of some collagen molecules and therefore the pathology exhibited in the canines was not as severe as that seen in complete knockout animals [
86].
7. Targeting TANGO1
Targeting TANGO1 as a means to control fibrosis is an attractive option, however, there are potential caveats to this approach. We speculate that the direct targeting and blockade of TANGO1 could result in adverse events or toxicities when collagen cannot be exported, and the ER exit sites are disassembled. Although TANGO1 (Long and Short) is needed for the export of collagen it also has a functional role in the export of other proteins [
76]. These will also be significantly affected. For example, TANGO1 is required for the export of apolipoprotein, and a certain amount of this protein is needed for normal mammalian health. However, more studies are warranted on the TANGO1 isoforms and paralog isoforms, certainly in the context of fibrosis, other collagenopathies, and cancer [
87,
88] to give a better understanding of the function of these proteins in these types of diseases. Currently, there are no TANGO1 inhibitors on the market or in clinical trials. The careful targeting of the interaction between TANGO1 and procollagen, possibly via the inhibition of the HSP47 binding domain, or with other export factors that regulate vesicle size may prove to be an effective therapeutic modality to treat a spectrum of fibrotic diseases that otherwise are difficult to control.
. Interaction of TANGO1 with other export factors. TANGO1 binds/interacts with multiple proteins that delay the closure of the vesicle allowing for the accommodation of procollagen so that it can be exported out of the ER.
8. Conclusions
Over the last decade, studies have shown the importance of TANGO1 in regulating the export of collagen out of the ER. Overall, TANGO1 interacts with other export factors to delay the closure of the vesicle so that it can accommodate large proteins such as procollagen (). Taken together, there is an elaborate interaction between TANGO1 and its co-factors that establish enlarged vesicles or tubules [
89,
90] for collagen export. Further evidence suggests that TANGO1 could be involved in the export of other cargoes as depletion or knockdown of this protein affected the secretion of other extracellular matrix proteins, while smaller cargoes were largely unaffected [
76,
91]. More studies on this protein are needed for us to better understand the role of TANGO1 in health and disease.
Author Contributions
Conceptualization, C.M.A.; Methodology, L.M.C.; Formal Analysis, C.M.A. and L.M.C.; Data Curation, C.M.A. and L.M.C.; Writing – Original Draft Preparation, C.M.A.; Writing – Review & Editing, C.M.A. and L.M.C.; Funding Acquisition, C.M.A.
Funding
This research was funded by the National Institutes of Health, grant number R21-AR-073947 to C.M.A., and the National Scleroderma Foundation to C.M.A.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Declaration of Competing Interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
References
-
1.
Wootton SC, Kim DS, Kondoh Y, Chen E, Lee JS, Song JW, et al. Viral infection in acute exacerbation of idiopathic pulmonary fibrosis.
Am. J. Respir. Crit. Care Med. 2011,
183, 1698–1702.
[Google Scholar]
-
2.
Collazos J, Carton JA, Asensi V. Immunological status does not influence hepatitis c virus or liver fibrosis in HIV-hepatitis C virus-coinfected patients.
AIDS Res. Hum. Retroviruses 2011,
27, 383–389.
[Google Scholar]
-
3.
Olson PD, Kuechenmeister LJ, Anderson KL, Daily S, Beenken KE, Roux CM, et al. Small molecule inhibitors of Staphylococcus aureus RnpA alter cellular mRNA turnover, exhibit antimicrobial activity, and attenuate pathogenesis.
PLoS Pathog 2011,
7, e1001287.
[Google Scholar]
-
4.
Yarnold J, Brotons MC. Pathogenetic mechanisms in radiation fibrosis.
Radiother. Oncol. 2010,
97, 149–161.
[Google Scholar]
-
5.
Meltzer EB, Noble PW. Idiopathic pulmonary fibrosis.
Orphanet. J. Rare Dis. 2008,
3, 8.
[Google Scholar]
-
6.
Daba MH, El-Tahir KE, Al-Arifi MN, Gubara OA. Drug-induced pulmonary fibrosis.
Saudi Med. J. 2004,
25, 700–706.
[Google Scholar]
-
7.
Tirelli C, Pesenti C, Miozzo M, Mondoni M, Fontana L, Centanni S. The Genetic and Epigenetic Footprint in Idiopathic Pulmonary Fibrosis and Familial Pulmonary Fibrosis: A State-of-the-Art Review.
Diagnostics 2022,
12, 3107.
[Google Scholar]
-
8.
Mercier S, Küry S, Barbarot S. Hereditary Fibrosing Poikiloderma with Tendon Contractures, Myopathy, and Pulmonary Fibrosis. In GeneReviews®; University of Washington: Seattle, WA, USA, 1993.
-
9.
Sisto M, Ribatti D, Lisi S. Sjögren’s Syndrome-Related Organs Fibrosis: Hypotheses and Realities.
J. Clin. Med. 2022,
11, e51319.
[Google Scholar]
-
10.
Diesler R, Cottin V. Pulmonary fibrosis associated with rheumatoid arthritis: from pathophysiology to treatment strategies.
Expert Rev. Respir. Med. 2022,
16, 541–553.
[Google Scholar]
-
11.
Case AH. Clinical Overview of Progressive Fibrotic Interstitial Lung Disease.
Front. Med. 2022,
9, 858339.
[Google Scholar]
-
12.
Liang C, Shi S, Meng Q, Liang D, Ji S, Zhang B, et al. Complex roles of the stroma in the intrinsic resistance to gemcitabine in pancreatic cancer: where we are and where we are going.
Exp. Mol. Med. 2017,
49, e406.
[Google Scholar]
-
13.
Blažević A, Hofland J, Hofland LJ, Feelders RA, de Herder WW. Small intestinal neuroendocrine tumours and fibrosis: an entangled conundrum.
Endocrine-Related Cancer 2018,
25, r115–r130.
[Google Scholar]
-
14.
Cupit-Link MC, Kirkland JL, Ness KK, Armstrong GT, Tchkonia T, LeBrasseur NK, et al. Biology of premature ageing in survivors of cancer.
ESMO Open 2017,
2, e000250.
[Google Scholar]
-
15.
Meng X, Wang H, Song X, Clifton AC, Xiao J. The potential role of senescence in limiting fibrosis caused by aging.
J. Cell Physiol. 2020,
235, 4046–4059.
[Google Scholar]
-
16.
Romano E, Rosa I, Fioretto BS, Matucci-Cerinic M, Manetti M. New Insights into Profibrotic Myofibroblast Formation in Systemic Sclerosis: When the Vascular Wall Becomes the Enemy.
Life 2021,
11, 610.
[Google Scholar]
-
17.
Tai Y, Woods EL, Dally J, Kong D, Steadman R, Moseley R, et al. Myofibroblasts: Function, Formation, and Scope of Molecular Therapies for Skin Fibrosis.
Biomolecules 2021,
11, 1095.
[Google Scholar]
-
18.
Romano E, Rosa I, Fioretto BS, Matucci-Cerinic M, Manetti M. The Role of Pro-fibrotic Myofibroblasts in Systemic Sclerosis: From Origin to Therapeutic Targeting.
Curr. Mol. Med. 2022,
22, 209–239.
[Google Scholar]
-
19.
Baek HA, Kim do S, Park HS, Jang KY, Kang MJ, Lee DG, et al. Involvement of endoplasmic reticulum stress in myofibroblastic differentiation of lung fibroblasts.
Am. J. Respir. Cell Mol. Biol. 2012,
46, 731–739.
[Google Scholar]
-
20.
Calfon M, Zeng H, Urano F, Till JH, Hubbard SR, Harding HP, et al. IRE1 couples endoplasmic reticulum load to secretory capacity by processing the XBP-1 mRNA.
Nature 2002,
415, 92–96.
[Google Scholar]
-
21.
Moore BD, Jin RU, Lo H, Jung M, Wang H, Battle MA, et al. Transcriptional Regulation of X-Box-binding Protein One (XBP1) by Hepatocyte Nuclear Factor 4alpha (HNF4Alpha) Is Vital to Beta-cell Function.
J. Biol. Chem. 2016,
291, 6146–6157.
[Google Scholar]
-
22.
Huh WJ, Esen E, Geahlen JH, Bredemeyer AJ, Lee AH, Shi G, et al. XBP1 controls maturation of gastric zymogenic cells by induction of MIST1 and expansion of the rough endoplasmic reticulum.
Gastroenterology 2010,
139, 2038–2049.
[Google Scholar]
-
23.
Lee AH, Chu GC, Iwakoshi NN, Glimcher LH. XBP-1 is required for biogenesis of cellular secretory machinery of exocrine glands.
Embo J. 2005,
24, 4368–4380.
[Google Scholar]
-
24.
Farhan H, Weiss M, Tani K, Kaufman RJ, Hauri HP. Adaptation of endoplasmic reticulum exit sites to acute and chronic increases in cargo load.
Embo J. 2008,
27, 2043–2054.
[Google Scholar]
-
25.
Maiers JL, Kostallari E, Mushref M, deAssuncao TM, Li H, Jalan-Sakrikar N, et al. The unfolded protein response mediates fibrogenesis and collagen I secretion through regulating TANGO1 in mice.
Hepatology 2017,
65, 983–998.
[Google Scholar]
-
26.
Acosta-Alvear D, Zhou Y, Blais A, Tsikitis M, Lents NH, Arias C, et al. XBP1 controls diverse cell type- and condition-specific transcriptional regulatory networks.
Mol. Cell 2007,
27, 53–66.
[Google Scholar]
-
27.
Mironov AA, Mironov AA Jr, Beznoussenko GV, Trucco A, Lupetti P, Smith JD, et al. ER-to-Golgi carriers arise through direct en bloc protrusion and multistage maturation of specialized ER exit domains.
Dev. Cell 2003,
5, 583–594.
[Google Scholar]
-
28.
Bannykh SI, Rowe T, Balch WE. The organization of endoplasmic reticulum export complexes.
J. Cell Biol. 1996,
135, 19–35.
[Google Scholar]
-
29.
Sato K. COPII coat assembly and selective export from the endoplasmic reticulum.
J. Biochem. 2004,
136, 755–760.
[Google Scholar]
-
30.
Lee MC, Orci L, Hamamoto S, Futai E, Ravazzola M, Schekman R. Sar1p N-terminal helix initiates membrane curvature and completes the fission of a COPII vesicle.
Cell 2005,
122, 605–617.
[Google Scholar]
-
31.
Hutchings J, Stancheva V, Miller EA, Zanetti G. Subtomogram averaging of COPII assemblies reveals how coat organization dictates membrane shape.
Nat. Commun. 2018,
9, 4154.
[Google Scholar]
-
32.
Saito K, Chen M, Bard F, Chen S, Zhou H, Woodley D, et al. TANGO1 facilitates cargo loading at endoplasmic reticulum exit sites.
Cell 2009,
136, 891–902.
[Google Scholar]
-
33.
Saito K, Maeda M. Not just a cargo receptor for large cargoes; an emerging role of TANGO1 as an organizer of ER exit sites.
J. Biochem. 2019,
166, 115–119.
[Google Scholar]
-
34.
Ma W, Goldberg J. TANGO1/cTAGE5 receptor as a polyvalent template for assembly of large COPII coats.
Proc. Natl. Acad. Sci. USA 2016,
113, 10061–10066.
[Google Scholar]
-
35.
Bonfanti L, Mironov AA Jr, Martínez-Menárguez JA, Martella O, Fusella A, Baldassarre M, et al. Procollagen traverses the Golgi stack without leaving the lumen of cisternae: evidence for cisternal maturation.
Cell 1998,
95, 993–1003.
[Google Scholar]
-
36.
Raote I, Ortega-Bellido M, Santos AJ, Foresti O, Zhang C, Garcia-Parajo MF, et al. TANGO1 builds a machine for collagen export by recruiting and spatially organizing COPII, tethers and membranes.
eLife 2018,
7, e32723.
[Google Scholar]
-
37.
Arnolds O, Stoll R. A new fold in TANGO1 evolved from SH3 domains for the export of bulky cargos. BioRxiv 2022, doi:10.1101/2022.02.02.478833.
-
38.
Ferreira LR, Norris K, Smith T, Hebert C, Sauk JJ. Association of Hsp47, Grp78, and Grp94 with procollagen supports the successive or coupled action of molecular chaperones.
J. Cell Biochem. 1994,
56, 518–526.
[Google Scholar]
-
39.
Omari S, Makareeva E, Gorrell L, Jarnik M, Lippincott-Schwartz J, Leikin S. Mechanisms of procollagen and HSP47 sorting during ER-to-Golgi trafficking.
Matrix Biol. 2020,
93, 79–94.
[Google Scholar]
-
40.
Maeda M, Saito K, Katada T. Distinct isoform-specific complexes of TANGO1 cooperatively facilitate collagen secretion from the endoplasmic reticulum.
Mol. Biol. Cell 2016,
27, 2688–2696.
[Google Scholar]
-
41.
Santos AJ, Nogueira C, Ortega-Bellido M, Malhotra V. TANGO1 and Mia2/cTAGE5 (TALI) cooperate to export bulky pre-chylomicrons/VLDLs from the endoplasmic reticulum.
J. Cell Biol. 2016,
213, 343–354.
[Google Scholar]
-
42.
Reynolds HM, Zhang L, Tran DT, Ten Hagen KG. Tango1 coordinates the formation of endoplasmic reticulum/Golgi docking sites to mediate secretory granule formation.
J. Biol. Chem. 2019,
294, 19498–19510.
[Google Scholar]
-
43.
Raote I, Ortega Bellido M, Pirozzi M, Zhang C, Melville D, Parashuraman S, et al. TANGO1 assembles into rings around COPII coats at ER exit sites.
J. Cell Biol. 2017,
216, 901–909.
[Google Scholar]
-
44.
Tanabe T, Maeda M, Saito K, Katada T. Dual function of cTAGE5 in collagen export from the endoplasmic reticulum.
Mol. Biol. Cell 2016,
27, 2008–2013.
[Google Scholar]
-
45.
King R, Lin Z, Balbin-Cuesta G, Myers G, Friedman A, Zhu G, et al. SEC23A rescues SEC23B-deficient congenital dyserythropoietic anemia type II.
Sci. Adv. 2021,
7, eabj5293.
[Google Scholar]
-
46.
Boyadjiev SA, Fromme JC, Ben J, Chong SS, Nauta C, Hur DJ, et al. Cranio-lenticulo-sutural dysplasia is caused by a SEC23A mutation leading to abnormal endoplasmic-reticulum-to-Golgi trafficking.
Nat. Genet. 2006,
38, 1192–1197.
[Google Scholar]
-
47.
Boyadjiev SA, Kim SD, Hata A, Haldeman-Englert C, Zackai EH, Naydenov C, et al. Cranio-lenticulo-sutural dysplasia associated with defects in collagen secretion.
Clin. Genet. 2011,
80, 169–176.
[Google Scholar]
-
48.
Zhu M, Tao J, Vasievich MP, Wei W, Zhu G, Khoriaty RN, et al. Neural tube opening and abnormal extraembryonic membrane development in SEC23A deficient mice.
Sci. Rep. 2015,
5, 15471.
[Google Scholar]
-
49.
Lu CL, Ortmeier S, Brudvig J, Moretti T, Cain J, Boyadjiev SA, et al. Collagen has a unique SEC24 preference for efficient export from the endoplasmic reticulum.
Traffic 2022,
23, 81–93.
[Google Scholar]
-
50.
Schmidt K, Cavodeassi F, Feng Y, Stephens DJ. Early stages of retinal development depend on Sec13 function.
Biol. Open 2013,
2, 256–266.
[Google Scholar]
-
51.
Niu X, Gao C, Jan Lo L, Luo Y, Meng C, Hong J, et al. Sec13 safeguards the integrity of the endoplasmic reticulum and organogenesis of the digestive system in zebrafish.
Dev. Biol. 2012,
367, 197–207.
[Google Scholar]
-
52.
Moreira TG, Zhang L, Shaulov L, Harel A, Kuss SK, Williams J, et al. Sec13 Regulates Expression of Specific Immune Factors Involved in Inflammation In Vivo.
Sci. Rep. 2015,
5, 17655.
[Google Scholar]
-
53.
Krupp M, Marquardt JU, Sahin U, Galle PR, Castle J, Teufel A. RNA-Seq Atlas--a reference database for gene expression profiling in normal tissue by next-generation sequencing.
Bioinformatics 2012,
28, 1184–1185.
[Google Scholar]
-
54.
Halperin D, Kadir R, Perez Y, Drabkin M, Yogev Y, Wormser O, et al. SEC31A mutation affects ER homeostasis, causing a neurological syndrome.
J. Med. Genet. 2019,
56, 139–148.
[Google Scholar]
-
55.
Townley AK, Feng Y, Schmidt K, Carter DA, Porter R, Verkade P, et al. Efficient coupling of Sec23-Sec24 to Sec13-Sec31 drives COPII-dependent collagen secretion and is essential for normal craniofacial development.
J. Cell Sci. 2008,
121, 3025–3034.
[Google Scholar]
-
56.
Jin L, Pahuja KB, Wickliffe KE, Gorur A, Baumgartel C, Schekman R, et al. Ubiquitin-dependent regulation of COPII coat size and function.
Nature 2012,
482, 495–500.
[Google Scholar]
-
57.
Asmar AJ, Beck DB, Werner A. Control of craniofacial and brain development by Cullin3-RING ubiquitin ligases: Lessons from human disease genetics.
Exp. Cell Res. 2020,
396, 112300.
[Google Scholar]
-
58.
Moretti T, Kim K, Tuladhar A, Kim J. KLHL12 can form large COPII structures in the absence of CUL3 neddylation.
Mol. Biol. Cell 2023,
34, br4.
[Google Scholar]
-
59.
McGourty CA, Akopian D, Walsh C, Gorur A, Werner A, Schekman R, et al. Regulation of the CUL3 Ubiquitin Ligase by a Calcium-Dependent Co-adaptor.
Cell 2016,
167, 525–538.e514.
[Google Scholar]
-
60.
Akopian D, McGourty CA, Rapé M. Co-adaptor driven assembly of a CUL3 E3 ligase complex.
Mol. Cell 2022,
82, 585–597.e511.
[Google Scholar]
-
61.
Follonier Castella L, Gabbiani G, McCulloch CA, Hinz B. Regulation of myofibroblast activities: calcium pulls some strings behind the scene.
Exp. Cell Res. 2010,
316, 2390–2401.
[Google Scholar]
-
62.
Malhotra V, Erlmann P. The pathway of collagen secretion.
Ann. Rev. Cell Dev. Biol. 2015,
31, 109–124.
[Google Scholar]
-
63.
Venditti R, Scanu T, Santoro M, Di Tullio G, Spaar A, Gaibisso R, et al. Sedlin controls the ER export of procollagen by regulating the Sar1 cycle.
Science 2012,
337, 1668–1672.
[Google Scholar]
-
64.
Saito K, Yamashiro K, Shimazu N, Tanabe T, Kontani K, Katada T. Concentration of Sec12 at ER exit sites via interaction with cTAGE5 is required for collagen export.
J. Cell Biol. 2014,
206, 751–762.
[Google Scholar]
-
65.
Jones B, Jones EL, Bonney SA, Patel HN, Mensenkamp AR, Eichenbaum-Voline S, et al. Mutations in a Sar1 GTPase of COPII vesicles are associated with lipid absorption disorders.
Nat. Genet. 2003,
34, 29–31.
[Google Scholar]
-
66.
Melville DB, Studer S, Schekman R. Small sequence variations between two mammalian paralogs of the small GTPase SAR1 underlie functional differences in coat protein complex II assembly.
J. Biol. Chem. 2020,
295, 8401–8412.
[Google Scholar]
-
67.
Sané A, Ahmarani L, Delvin E, Auclair N, Spahis S, Levy E. SAR1B GTPase is necessary to protect intestinal cells from disorders of lipid homeostasis, oxidative stress, and inflammation.
J. Lipid Res. 2019,
60, 1755–1764.
[Google Scholar]
-
68.
Levic DS, Minkel JR, Wang WD, Rybski WM, Melville DB, Knapik EW. Animal model of Sar1b deficiency presents lipid absorption deficits similar to Anderson disease.
J. Mol. Med. 2015,
93, 165–176.
[Google Scholar]
-
69.
Yuan L, Kenny SJ, Hemmati J, Xu K, Schekman R. TANGO1 and SEC12 are co-packaged with procollagen I to facilitate the generation of large COPII carriers.
Proc. Natl. Acad. Sci. USA 2018,
115, e12255–e12264.
[Google Scholar]
-
70.
Saito K, Yamashiro K, Ichikawa Y, Erlmann P, Kontani K, Malhotra V, et al. cTAGE5 mediates collagen secretion through interaction with TANGO1 at endoplasmic reticulum exit sites.
Mol. Biol. Cell 2011,
22, 2301–2308.
[Google Scholar]
-
71.
Santos AJ, Raote I, Scarpa M, Brouwers N, Malhotra V. TANGO1 recruits ERGIC membranes to the endoplasmic reticulum for procollagen export.
eLife 2015,
4, e10982.
[Google Scholar]
-
72.
Kreiner T, Moore HP. Membrane traffic between secretory compartments is differentially affected during mitosis.
Cell Regul. 1990,
1, 415–424.
[Google Scholar]
-
73.
Hughes H, Stephens DJ. Sec16A defines the site for vesicle budding from the endoplasmic reticulum on exit from mitosis.
J. Cell Sci. 2010,
123, 4032–4038.
[Google Scholar]
-
74.
Maeda M, Katada T, Saito K. TANGO1 recruits Sec16 to coordinately organize ER exit sites for efficient secretion.
J. Cell Biol. 2017,
216, 1731–1743.
[Google Scholar]
-
75.
Maeda M, Komatsu Y, Saito K. Mitotic ER Exit Site Disassembly and Reassembly Are Regulated by the Phosphorylation Status of TANGO1.
Dev. Cell 2020,
55, 237–250.
[Google Scholar]
-
76.
Connolly LM, McFalls CM, McMahon IG, Bhat AM, Artlett CM. Caspase-1 Enhances TANGO1 Expression to Promote Procollagen Export from the Endoplasmic Reticulum in Systemic Sclerosis Contributing to Fibrosis.
Arthritis Rheum. 2023,
75, 1831–1841.
[Google Scholar]
-
77.
Varga J, Abraham D. Systemic sclerosis: a prototypic multisystem fibrotic disorder.
J. Clin. Invest. 2007,
117, 557–567.
[Google Scholar]
-
78.
Abraham DJ, Eckes B, Rajkumar V, Krieg T. New developments in fibroblast and myofibroblast biology: implications for fibrosis and scleroderma.
Curr. Rheum. Rep. 2007,
9, 136–143.
[Google Scholar]
-
79.
Tomasek JJ, Gabbiani G, Hinz B, Chaponnier C, Brown RA. Myofibroblasts and mechano-regulation of connective tissue remodelling.
Nat. Rev. 2002,
3, 349–363.
[Google Scholar]
-
80.
Kirk TZ, Mark ME, Chua CC, Chua BH, Mayes MD. Myofibroblasts from scleroderma skin synthesize elevated levels of collagen and tissue inhibitor of metalloproteinase (TIMP-1) with two forms of TIMP-1.
J. Biol. Chem. 1995,
270, 3423–3428.
[Google Scholar]
-
81.
Junkiert-Czarnecka A, Pilarska-Deltow M, Bąk A, Heise M, Haus O. A novel mutation in collagen transport protein, MIA3 gene, detected in a patient with clinical symptoms of Ehlers-Danlos hypermobile syndrome.
Adv. Clin. Exp. Med. 2023,
32, 777–781.
[Google Scholar]
-
82.
Lekszas C, Foresti O, Raote I, Liedtke D, König EM, Nanda I, et al. Biallelic TANGO1 mutations cause a novel syndromal disease due to hampered cellular collagen secretion.
eLife 2020,
9, e51319.
[Google Scholar]
-
83.
Guillemyn B, Nampoothiri S, Syx D, Malfait F, Symoens S. Loss of TANGO1 Leads to Absence of Bone Mineralization.
JBMR Plus 2021,
5, e10451.
[Google Scholar]
-
84.
Clark EM, Link BA. Complementary and divergent functions of zebrafish Tango1 and Ctage5 in tissue development and homeostasis.
Mol. Biol. Cell 2021,
32, 391–401.
[Google Scholar]
-
85.
Wilson DG, Phamluong K, Li L, Sun M, Cao TC, Liu PS, et al. Global defects in collagen secretion in a Mia3/TANGO1 knockout mouse.
J. Cell Biol. 2011,
193, 935–951.
[Google Scholar]
-
86.
Christen M, Booij-Vrieling H, Oksa-Minalto J, de Vries C, Kehl A, Jagannathan V, et al. MIA3 Splice Defect in Cane Corso Dogs with Dental-Skeletal-Retinal Anomaly (DSRA).
Genes 2021,
12, 1497.
[Google Scholar]
-
87.
Wanbiao Z, Jing M, Shi Z, Chen T, Zhao X, Li H. MIA3 promotes the degradation of GSH (glutathione) by binding to CHAC1, thereby promoting the progression of hepatocellular carcinoma. Mol. Cell Biochem. 2023, doi:10.1007/s11010-023-04850-9.
-
88.
Man J, Zhou W, Zuo S, Zhao X, Wang Q, Ma H, et al. TANGO1 interacts with NRTN to promote hepatocellular carcinoma progression by regulating the PI3K/AKT/mTOR signaling pathway.
Biochem. Pharmacol. 2023,
213, 115615.
[Google Scholar]
-
89.
Liu M, Feng Z, Ke H, Liu Y, Sun T, Dai J, et al. Tango1 spatially organizes ER exit sites to control ER export.
J. Cell Biol. 2017,
216, 1035–1049.
[Google Scholar]
-
90.
Yang K, Liu M, Feng Z, Rojas M, Zhou L, Ke H, et al. ER exit sites in Drosophila display abundant ER-Golgi vesicles and pearled tubes but no megacarriers.
Cell Rep. 2021,
36, 109707.
[Google Scholar]
-
91.
McCaughey J, Stevenson NL, Mantell JM, Neal CR, Paterson A, Heesom K, et al. A general role for TANGO1, encoded by MIA3, in secretory pathway organization and function.
J. Cell Sci. 2021,
134, jcs259075.
[Google Scholar]