1. Introduction
Microplastics (MPs) refer to plastic particles, fibers, and films with a particle size less than 5 mm, mainly from plastic film covering, followed by sludge utilization, organic fertilizer application, wastewater irrigation, and atmospheric deposition [
1]. Low-density polyethylene (LDPE) is extensively utilized as a plastic film in farmland due to its cost-effectiveness, ease of processing, and high durability. It represents the most common type of MPs found in soil [
2,
3]. However, LDPE’s remarkable chemical stability, resulting from its high molecular weight and hydrophobic nature, contributes to severe ecological issues due to its non-biodegradability [
4].
Recent research has demonstrated that LDPE MPs in soil can alter microbial diversity and enzyme activity [
5]. Microbiota will colonize the surface of MPs, forming a biofilm commonly referred to as the plastisphere. Additionally, MPs can directly interact with soil microorganisms, contributing to the development of the soil plastisphere [
6]. The soil plastisphere typically exhibits lower microbial diversity than its surrounding environment [
7]. However, certain species of bacteria from the Vibrionaceae and Pseudomonadaceae families are abundant on the surface of plastic debris, but are relatively scarce in the surrounding environment [
8]. The effects of LDPE-MPs on soil enzyme activity are complex, as they can both promote and inhibit enzyme activity [
9]. The effects of LDPE-MPs on crop biomass varied with concentration, species, and particle size. For example, studies have shown that HDPE promotes maize growth by increasing aboveground and root biomass production [
10]. However, other studies have found that LDPE has significant negative effects on stem diameter, height, leaf area, and root-shoot ratio in plants [
11]. Therefore, our understanding of the relationship between these microplastics and their impacts on physical, chemical, and biological properties remains limited, impeding our ability to effectively address issues related to MP pollution in agricultural ecosystems.
Studies have shown that biochar can significantly improve the fertility and microbial diversity of LDPE-MPs contaminated soils [
12], but the production cost of this material is high, and the process is complicated. In contrast, wood chips, as a commonly utilized form of biomass material, exhibit distinct advantages in soil improvement. They are rich in organic matter and contain a variety of essential nutrients, including C, N, P, and K [
13]. Wood chips as a modifier can enhance soil structure, soil permeability, and water retention, decrease the mobility and bioavailability of toxic metals, reduce pathogenic microorganisms, improve crop disease resistance, promote plant growth rate and health, and meet the requirements of environmental friendliness and sustainable development [
14,
15]. An efficient bio-based cross-flow capture material (bioCap) has been explored [
16]. The flow capture of MPs through polyphenol-mediated interfacial interactions on wood chips removes a variety of MPs (ranging from approximately 100 nm to 22 μm in size) with any surface charge (cations, neutrals, and anions) and various chemical components from water [
17]. However, little is known about how the addition of wood chips to soil affects soil physical and chemical properties and plant growth under microplastic pollution.
Therefore, this study was designed to investigate the effects of wood chips on soil physicochemical properties, plant growth, and soil microbial communities in farmland contaminated with LDPE-MPs. Specifically, the objectives were: (i) to examine the impact of LDPE-MPs on soil physicochemical properties and plant growth in conventional farmland; (ii) to analyze the influence of LDPE-MPs on soil bacterial and fungal diversity, structure, composition, function, co-occurrence network; and (iii) to assess the potential of wood chips to alleviate LDPE-induced changes in soil health and crop performance.
2. Materials and Methods
2.1. Study Site
This experiment was conducted at the Yellow River Floodplain Ecosystems Research Station (34°53′42.03″ N, 113°26′19.77″ E), Henan Province, China. The research platform is a typical intensive management farmland in the North China Plain, approximately 7 km from the research station. In the area, maize and wheat have been cropped in rotation for a long time, and there is no history of plastic film mulching in the experimental plots. The climatic conditions in this area are characterized by a warm-temperate, continental monsoon climate, with an average annual temperature of 14.8 °C, an average annual rainfall of 608.8 mm, and a frost-free period of 233 days.
2.2. Experimental Design
The LDPE mulch film purchased from HuaXin Plastic Products Co., Ltd. (Jining, China) was crushed (5 mm × 5 mm) to form LDPE-MPs. They were washed five times with sterile ultrapure water, dried in a fume hood at 40 °C, and stored at 4 °C before use. Pomegranate branches and peach branches around the site were collected and crushed into wood chips about 2 cm in size using a branch crusher, mixed evenly, and dried. The basic chemical properties of wood chips are as follows: the total carbon content is 46.89 ± 3.61 g/kg; the total nitrogen content is 0.68 ± 0.07 g/kg. After the wheat harvest (June 2022), the experiment was arranged in the plot, and the nutrients of the field soil are shown in
Table S1.
The experiment was divided into four treatments: control (C), wood chips (W), LDPE (N), and wood chips + LDPE (WN). Each treatment was replicated four times, with all plots randomly arranged, totaling 16 plots, each with an area of 48 m
2 (4 m × 12 m). The north-south interval between the quadrats is 0.4 m, and the east-west interval is 3 m. To approximate the amount of microplastics from the film covering the ground, LDPE microplastics were added based on the product of the weight of one square of the LDPE film and the area of the quadrat,
i.e., 200 g of LDPE microplastics per quadrat (41.67 kg/ha). The amount of wood chips added was calculated similarly,
i.e., 52.8 kg per quadrat (11,000 kg/ha). Common fertilization management (urea) and watering measures were used in the experimental field.
2.3. Sample Collection and Processing
The crop rhizosphere soil was collected during the maize vigorous growing period (MVG, August 2022) and the wheat vigorous growing period (WVG, April 2023). The experiment lasted for approximately 10 months. Eight random sampling points per plot were selected to excavate the entire root systems of crops, removing the loosely attached soil particles, and collecting the rhizosphere soil (1–2 mm from the root). The soil samples from the same plot were fully mixed, placed in a box with ice packs, and immediately transported to the laboratory. 16 mixed soil samples (4 treatments × 4 replicates) were collected each time, and 32 were collected twice.
After the collected soil samples were brought back to the laboratory, the sand and plant roots in the soil were removed by a 2 mm sieve, and then the soil was divided into four parts. One part was stored in an ultra-low temperature refrigerator at −80 °C for DNA extraction of soil microorganisms. One part was stored in a refrigerator at −20 °C to determine soil enzyme activity; one part was stored in a refrigerator at 4 °C, and the determination of fresh soil indexes, such as soil available nitrogen and water content, was completed within 7 days. The last part was air-dried naturally, ground, and passed through a 0.149 mm sieve to determine soil pH, total carbon, total nitrogen, and available phosphorus.
Wheat plants were collected during the wheat mature season (June 2023), and three points were selected in each plot to dig up multiple crops to remove the roots and bring them back to the laboratory for drying at 105 °C for 30 min, followed by drying to constant weight at 65 °C.
2.4. Soil and Plant Measurement
Soil pH (soil-water mass ratio of 1:2.5) was determined using a pH device (PB 10, Sartorius, Göttingen, Germany). Soil total carbon (TC) and total nitrogen (TN) were measured using a Vario Macro cube (Elementar, Langenselbold, Germany). Soil ammonium (NH
4+-N) and nitrate (NO
3−-N) were measured using an automatic chemical analyzer (Smartchem 200, AMS, Novara, Italy). The sodium bicarbonate extraction-molybdenum antimony anti-colorimetric-ultraviolet method determined the available phosphorus (AP) in soil. The soil enzyme activities were determined by 96-well microplate fluorescence analysis: β-N-acetylglucosaminidase (NAG), β-xylosidase (BX), β-glucosidase (BG), α-glucosidase (αG), cellobiase (BCE), and acid phosphatase (AP). The activity was measured by a microplate reader (Synergy H4, BioTek, Winooski, VT, USA) with excitation and emission wavelengths at 365 nm and 450 nm, respectively.
Ten wheat plants were randomly selected from each plot to measure the plant height, panicle length, and stem diameter of wheat with a tape measure. The panicle weight and the whole plant weight were measured. The number of grains per panicle, the number of filled grains, and the thousand seed weight were counted.
2.5. Soil DNA Extraction, Amplification, and Sequencing
The rhizosphere soil microbial community in wheat and maize growing seasons was studied. According to the manufacturer’s instructions, the OMEGA Soil DNA Kit (Omega Bio-Tek, Norcross, GA, USA) was used to extract DNA from 0.50 g of soil. The V3-V4 region of the bacterial 16S rRNA gene and the fungal ITS2 sequence were amplified by an ABI GeneAmp 9700 polymerase chain reaction thermocycler (ABI, CA, Houston, TX, USA). The bacterial primers were 338F (5′-
ACTCCTACGGGAGGCAGCA-3′) and 806R (5′-
GGACTACHVGGGTWTCTAAT-3′). The fungal primers were ITS5F (5′-
GCATCGATGAAGAACGCAGC-3′) and IT1R (5′-
TCCTCCGCTTATTGATATGC-3′). PCR was carried out according to the following protocol: initial denaturation at 95 °C for 3 min, followed by 27 cycles of denaturation at 95 °C for 30 s, annealing at 55 °C for 30 s, and extension at 72 °C for 45 s. Finally, it was denatured at 72 °C for 10 min and ended at 10 °C. The PCR mixture contained 5× FastPfu buffer (4 μL), 2.50 mM dNTP (2 μL), 5 μM forward and reverse primers (0.80 μL), FastPfu polymerase (0.4 μL), and BSA (0.20 μL). Template DNA (10 ng) was supplemented with ddH
2O up to 20 μL. The amplicons were extracted from 2% agarose gel and purified using the AxyPrep DNA gel extraction kit (Axygen Biosciences, Union City, CA, USA). The purified amplicons were subjected to equimolar and paired terminal sequencing on the Illumina Miseq platform of Shanghai Parsono Biotechnology Co., Ltd., Shanghai, China.
After truncating the barcode and primer sequences, FLASH (v. 1.2.11) was used to splice the double-ended read length of each sample. The DADA2 module in the QIIME2 software was used to denoise, and the initial augmented subsequence variants (ASVs) were obtained. The bacterial 16S rRNA gene was annotated using the Silva database (Release 132, http://www.arb-silva.de, accessed on 6 April 2025) [
17]. Annotating the fungal ITS sequence from the UNITE database (Release 8.0, https://unite.ut.ee/, accessed on 6 April 2025) [
18].
2.6. Statistical Analysis
Shapiro-Wilk and Bartlett tests assessed the normal distribution and homogeneity of variance. A three-factor analysis of variance (ANOVA) was conducted using R software (v. 4.2.3) to evaluate the soil’s physical and chemical properties and its bacterial α-diversity, while a two-way ANOVA was employed to analyze the crop growth indices. Canoco5 was used for RDA graphics. Lastly, random forest analysis was applied to determine the bacterial species that contributed most to the treatments. Tukey’s HSD was used to test the differences between the treatments, and the statistical significance was set to
p < 0.05.
The alpha diversity indices, including Chao1 richness, Pielou’s evenness, Shannon diversity, and Observed_species, were calculated for each sample in QIIME2. Principal component analysis (PCoA, 999 permutations based on the Bray-Curtis dissimilarity matrix) was used to visually display the changes in soil bacterial community structure between different treatments. Permutation multivariate analysis of variance (PERMANOVA, permutation = 999) based on Bray-Curtis distance was used to evaluate the significance of principal component analysis (PCoA). The Spearman correlation coefficient was used to draw a correlation heatmap to evaluate the relationship between soil microbial taxa’s top 20 dominant genera and environmental parameters. Random forest regression (RandomForest) was used to analyze the importance of field rhizosphere classification variables and differences in taxa abundance.
The FAPROTAX method was used to predict the function of bacteria, and the ‘
FUNGuildR’ package was used to predict the function of fungi. A Spearman correlation matrix was constructed for ASVs with relative abundance > 0.01% was created using the ‘
WGCNA’ package (v. 1.72–5) in the R software (v. 4.2.3), and a significant correlation (r > 0.6 and
p < 0.01) was selected to construct a co-occurrence network of microbial communities. Gephi software (version 0.10.2) was used to visualize the correlation of the results, and the ‘
igraph’ package (v. 1.5.1) was used to calculate the topological properties of the network. Origin 2018 software was used to draw histograms and relative abundance stacked histograms.
3. Results
3.1. Soil Properties and Enzyme Activities
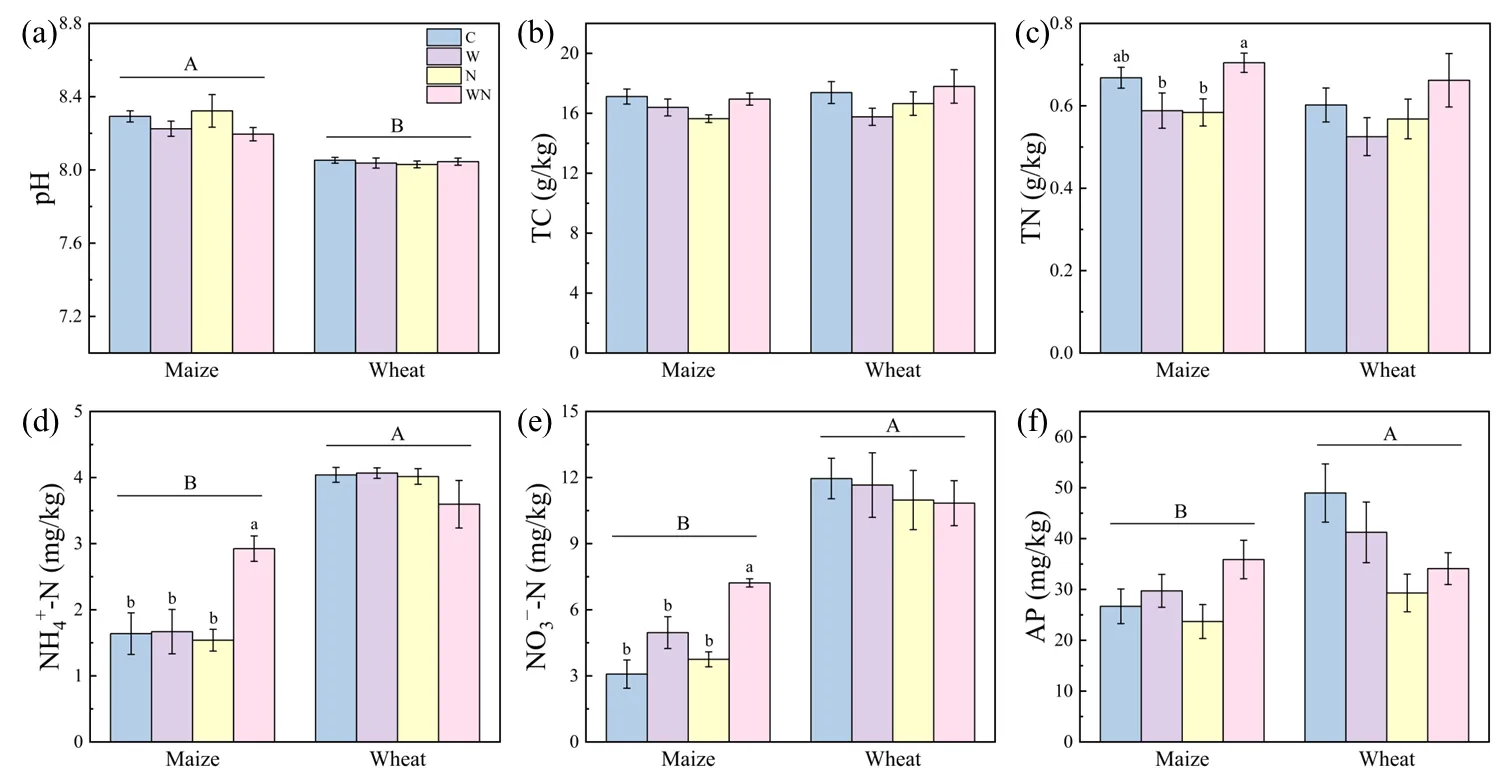
LDPE treatment reduced soil TC content in both periods by 8.63% and 5.63% respectively, while adding wood chips increased soil TC by 5.45% and 9.98% respectively (
b). Compared with LDPE treatment, wood chips + LDPE treatment significantly increased TN content by 20.65% in the MVG period (
c,
p < 0.05). Compared with the control, LDPE reduced NH
4+-N content in both periods (
d). During the MVG period, compared with the control, wood chips + LDPE treatment significantly increased NH
4+-N content by 89.96% (
d,
p < 0.05). Compared with the control, LDPE increased NO
3--N content by 21.80% in the MVG period, while wood chips + LDPE treatment significantly increased it by 134.45% (
e,
p < 0.05). During both the MVG and WVG period, LDPE reduced the AP content by 11.18% and 40.36% respectively, compared with the control, while wood chips increased the AP content by 51.49% and 16.36% respectively (
f).
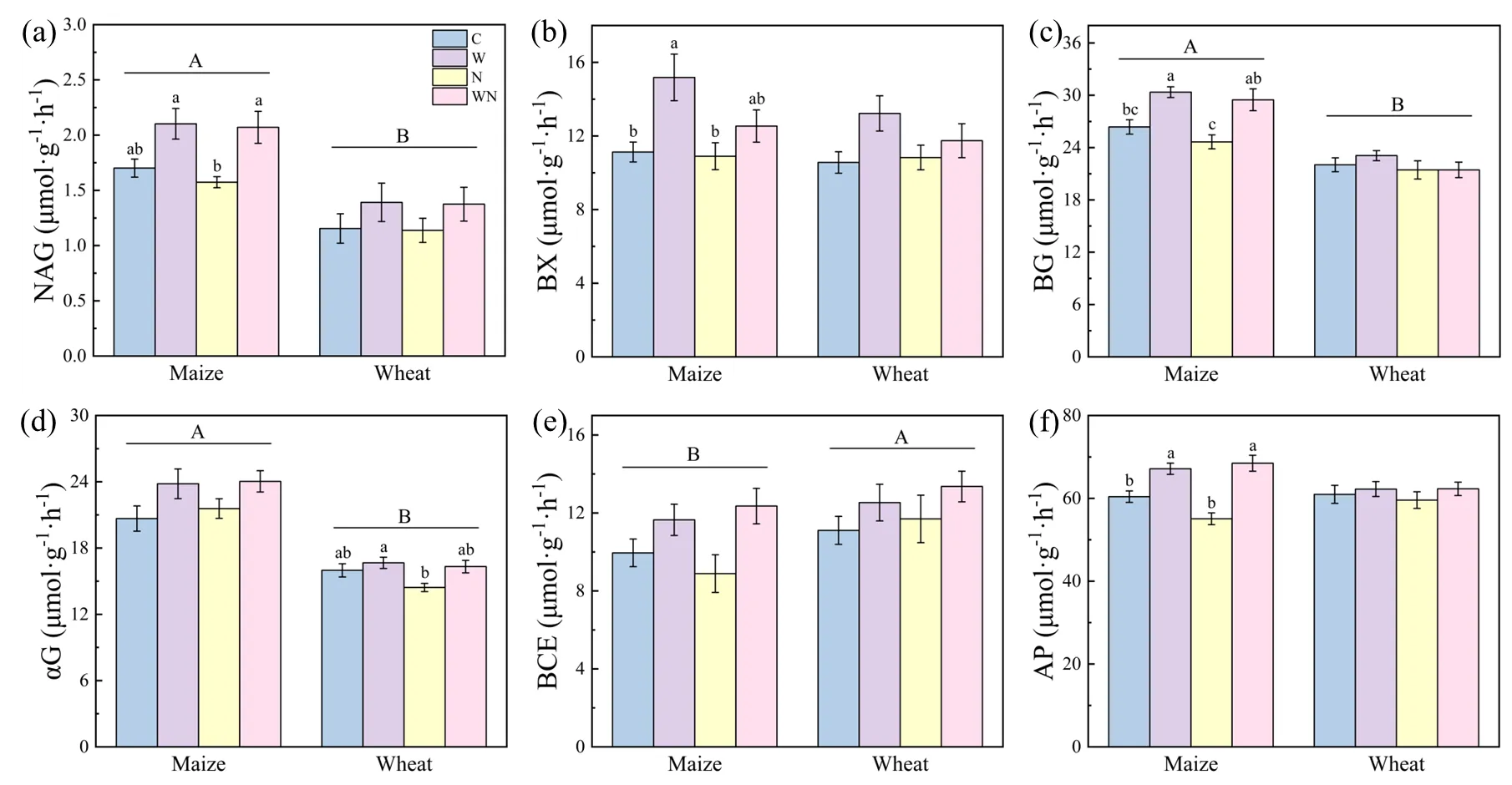
Wood chips (W) had a significant effect on soil C-related enzyme activities (BX, BG, αG, and BCE), N-related enzyme activities (NAG), and P-related enzyme activities (AP), which were significantly increased (
,
p < 0.05). Time (T) had a significant effect on soil NAG, BG, αG, and BCE enzyme activities. NAG, BG, and αG activities were significantly higher in the MVG period than those in the WVG period (
,
p < 0.05). In the MVG period, LDPE significantly reduced NAG activity by 7.49% compared with the control (
a,
p < 0.05), and it significantly reduced BX activity by 28.19% compared with the wood chips treatment (
b,
p < 0.05). In the WVG period, compared with the wood chips treatment, LDPE reduced αG activity by 13.40% (
d). LDPE treatment after adding wood chips increased αG activity in both periods (
d). LDPE treatment reduced BCE activity by 10.68% compared with the control during the MVG period (
e). In the MVG period, compared with LDPE treatment, the AP activity of wood chips and wood chips + LDPE treatments increased significantly by 21.94% and 24.36%, respectively. LDPE decreased AP activity, while wood chips increased activity by 24.36% and 4.57% in the MVG and WVG periods, respectively (
f,
p > 0.05).
.
Three-way ANOVA testing the effects of wood chips (W), LDPE (N) and time (T) on soil chemical properties and enzyme activities.
|
W |
N |
T |
W∗N |
N∗T |
W∗T |
W∗N∗T |
| pH |
0.051 |
0.932 |
<0.001 |
0.314 |
0.932 |
0.211 |
1.000 |
| TC |
0.958 |
0.845 |
0.433 |
0.017 |
0.249 |
0.581 |
0.700 |
| TN |
0.633 |
0.272 |
0.132 |
0.005 |
0.560 |
0.847 |
0.808 |
| NH4+-N |
0.135 |
0.330 |
<0.001 |
0.183 |
0.020 |
0.012 |
0.012 |
| NO3−-N |
0.074 |
0.671 |
<0.001 |
0.515 |
0.085 |
0.037 |
0.591 |
| AP |
0.307 |
0.057 |
0.004 |
0.079 |
0.018 |
0.136 |
0.776 |
| E-NAG |
<0.001 |
0.602 |
<0.001 |
0.795 |
0.732 |
0.257 |
0.796 |
| E-BX |
<0.001 |
0.102 |
0.171 |
0.095 |
0.497 |
0.387 |
0.784 |
| E-BG |
<0.001 |
0.063 |
<0.001 |
0.931 |
0.885 |
0.004 |
0.453 |
| E-αG |
0.003 |
0.755 |
<0.001 |
0.831 |
0.226 |
0.223 |
0.441 |
| E-BCE |
0.003 |
0.680 |
0.030 |
0.436 |
0.490 |
0.421 |
0.551 |
| E-AP |
<0.001 |
0.286 |
0.232 |
0.111 |
0.594 |
0.003 |
0.299 |
. Soil chemical properties under each treatment (C, control, W, wood chips; N, LDPE; WN: wood chips + LDPE) during vigorous growing season of maize and wheat. (<strong>a</strong>) soil pH; (<strong>b</strong>) soil total carbon (TC); (<strong>c</strong>) soil total nitrogen (TN); (<strong>d</strong>) soil ammonium nitrogen (NH<sub>4</sub><sup>+</sup>-N); (<strong>e</strong>) soil nitrate nitrogen (NO<sub>3</sub><sup>−</sup>-N); (<strong>f</strong>) soil available phosphorus (AP). Different letters indicate significant differences at <em>p </em>< 0.05 (mean ± SE., <em>n</em> = 4).
. Soil enzyme activity under each treatment (C, control, W, wood chips; N, LDPE; WN: wood chips + LDPE) during vigorous growing season of maize and wheat. (<strong>a</strong>) β-N-acetylglucosaminidase (NAG); (<strong>b</strong>) β-xylosidase (BX); (<strong>c</strong>)β-glucosidase (BG); (<strong>d</strong>) α-glucosidase (αG); (<strong>e</strong>) cellobiase (BCE); (<strong>f</strong>) acid phosphatase (AP). Different letters indicate significant differences at <em>p</em> < 0.05 (mean ± SE., <em>n</em> = 4).
The main effect of LDPE had significant effects on plant height and whole plant weight (
,
p < 0.05). The main effect of wood chips (W) and the interaction between wood chips and LDPE (W∗N) had significant effects on panicle length and panicle number (
,
p < 0.05). The main effect of wood chips (W) also had significant effects on panicle weight and kernel number (
,
p < 0.05). Compared with the control, LDPE treatment reduced the plant height, panicle length, filled grain number, and thousand seed weight of wheat by 5.72%, 4.31%, 3.73%, and 0.13% respectively. While the adverse effects of LDPE on wheat growth were alleviated by adding wood chips.
.
Two-factor ANOVA of the effects of wood chips (W) and LDPE (N) on wheat growth and the effects of different treatments on wheat growth.
| Wheat Growth Parameter |
Plant Height (cm) |
Stem Diameter (mm) |
Panicle Length (cm) |
Panicle Weight (g) |
Whole Plant Weight (g) |
Kernels per Spike |
Filled Grain Number |
Thousand Seed Weight (g) |
| W |
0.296 |
0.702 |
0.020 |
<0.001 |
0.070 |
<0.001 |
0.002 |
0.052 |
| N |
0.011 |
0.702 |
0.981 |
0.154 |
0.025 |
0.505 |
0.525 |
0.399 |
| W∗N |
0.207 |
0.898 |
0.041 |
0.862 |
0.482 |
0.018 |
0.627 |
0.387 |
| C |
59.575 ± 0.886 a |
4.525 ± 0.107 a |
10.270 ± 0.192 ab |
2.476 ± 0.111 b |
3.645 ± 0.151 b |
58.225 ± 1.882 b |
50.300 ± 1.646 ab |
36.660 ± 1.509 a |
| W |
59.380 ± 0.599 ab |
4.500 ± 0.101 a |
10.330 ± 0.238 ab |
2.861 ± 0.084 ab |
4.037 ± 0.106 ab |
69.125 ± 1.964 a |
54.725 ± 1.576 a |
39.295 ± 0.710 a |
| N |
56.165 ± 1.043 b |
4.575 ± 0.101 a |
9.828 ± 0.203 b |
2.620 ± 0.139 b |
4.104 ± 0.194 ab |
61.175 ± 1.420 b |
48.425 ± 1.829 b |
36.611 ± 3.805 a |
| WN |
58.230 ± 0.972 ab |
4.525 ± 0.080 a |
10.763 ± 0.212 a |
3.045 ± 0.117 a |
4.277 ± 0.156 a |
63.900 ± 1.485 ab |
54.475 ± 1.607 ab |
43.004 ± 0.479 a |
3.3. Soil Microbial Community Diversity, Composition, and Structure
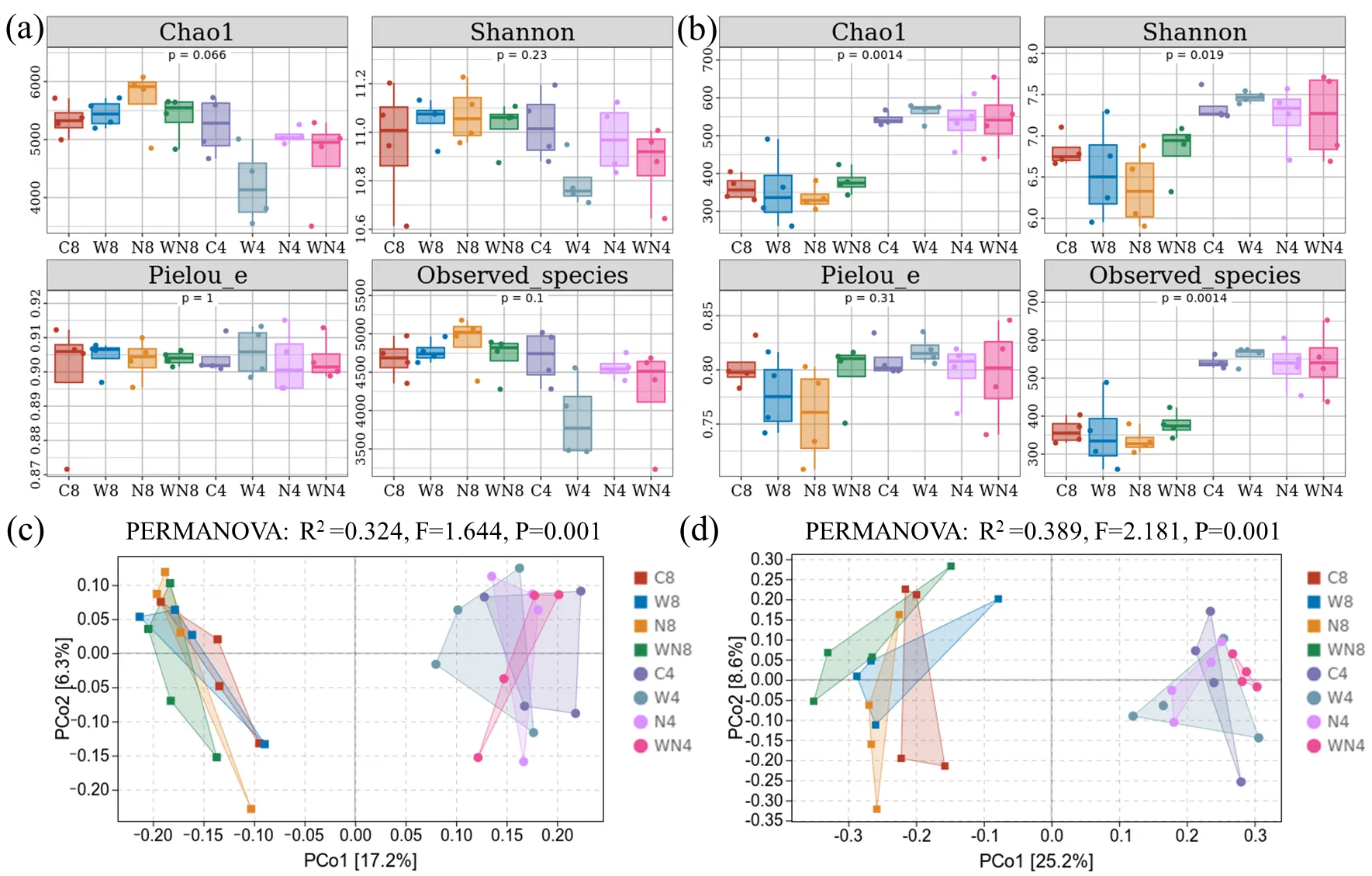
Two-way analysis of variance showed that time (T) had a significant effect on bacterial and fungal Chao1, Shannon, and Observed_species (
,
p < 0.05). The addition of wood chips did not increase the bacterial α diversity index (
a,
p > 0.05). The fungal Chao1, Shannon, Pielou_e, and Observed_species indexes under LDPE treatment decreased by 7.26%, 6.71%, 5.54% and 7.28%, respectively, compared with the control. But these adverse effects were mitigated in the WVG period, and the addition of wood chips increased the fungal α diversity index (
b,
p > 0.05).
.
Three-way ANOVA testing effects of wood chips (W), LDPE (N) and time (T) on alpha diversity of soil bacterial and fungal communities.
|
|
W |
N |
T |
W∗N |
N∗T |
W∗T |
W∗N∗T |
| Bacteria |
Chao1 |
0.031 |
0.411 |
<0.001 |
0.730 |
0.993 |
0.089 |
0.147 |
| Shannon |
0.182 |
0.580 |
0.045 |
0.963 |
0.761 |
0.084 |
0.206 |
| Pielou_e |
0.510 |
0.957 |
0.653 |
0.635 |
0.543 |
0.784 |
0.739 |
| Observed_species |
0.032 |
0.505 |
0.005 |
0.725 |
0.929 |
0.072 |
0.167 |
| Fungi |
Chao1 |
0.451 |
0.739 |
<0.001 |
0.647 |
0.795 |
0.875 |
0.452 |
| Shannon |
0.549 |
0.332 |
<0.001 |
0.278 |
0.790 |
0.865 |
0.135 |
| Pielou_e |
0.642 |
0.220 |
0.058 |
0.232 |
0.892 |
0.895 |
0.107 |
| Observed_species |
0.433 |
0.746 |
<0.001 |
0.627 |
0.791 |
0.898 |
0.454 |
. Soil (<strong>a</strong>) bacterial α diversity, (<strong>b</strong>) fungal α diversity, (<strong>c</strong>) bacterial β diversity and (<strong>d</strong>) fungal β diversity under different treatments (C: control; W: wood chips; N: LDPE; WN: wood chips + LDPE. 8: vigorous season of maize growth; 4: vigorous season of wheat growth).
The contribution rate of the first two principal components to bacterial community variation was 23.5% (
c). The first two principal components explained 33.8% of the observed fungal community variance (
d). There were significant differences in bacterial and fungal communities between the MVG period and the WVG period (
Table S2,
c,d,
p < 0.05). In the MVG period, the fungal communities between the control and LDPE treatments were significantly separated (
Table S2,
c,
p < 0.05). In the WVG period, there were significant differences in bacterial communities between the control and wood chips treatments, and between the control and LDPE treatment (
Table S2,
c,
p < 0.05). There were significant dissimilarities in fungal communities between wood chips and wood chips + LDPE treatments, LDPE treatment, and wood chips + LDPE treatment (
Table S2,
d,
p < 0.05).
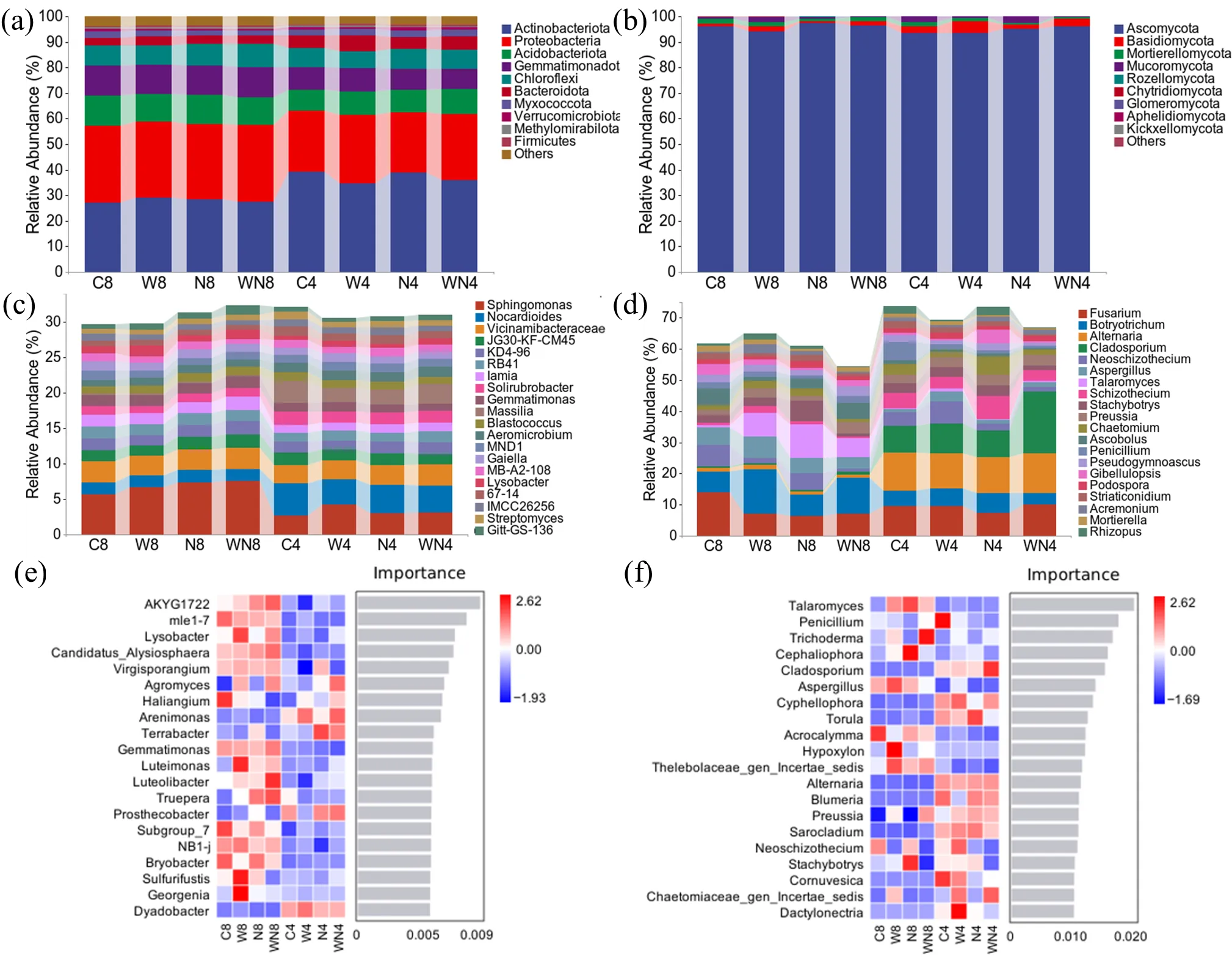
The dominant phyla of bacteria in our study were Actinobacteria, Proteobacteria, Acidobacteriota, and Gemmatimonadota (relative abundance > 10%,
a). The dominant phylum of fungi was Ascomycota (
b), which increased with the LDPE treatment. At the genus level, the dominant bacteria were
Sphingomonas,
Nocardioides, and
Vicinamibacteraceae (relative abundance > 2%,
c).
Fusarium was the dominant fungal genus in the soil (relative abundance > 2%,
d), which was inhibited by the LDPE addition.
It can be seen from
Figure 4e that LDPE treatment enriched
Virgisporangium at the WVG stage and
Truepera at the MVG stage.
Sulfurifustis and
Georgeni of the WVG stage were enriched by wood chip treatment. The relative abundance of
Agromyces increased under the treatment of wood chips + LDPE in the WVG and MVG periods.
From
f, we can also analyze that the relative abundance of
Talaromyces only increased under the treatment of wood chips, LDPE, and wood chips + LDPE in the MVG period. The relative abundance of
Trichoderma increased under the treatment of wood chips and wood chips + LDPE in the MVG period.
Cephaliophora was enriched under LDPE treatment during the MVG period. The relative abundance of
Cladosporium,
Cyphellophora,
Torula,
Alternaria,
Blumeria, and
Sarocladium decreased in the MVG period. It increased in the WVG period, while the relative abundance of
Aspergillus and
Thelebolaceae_gen_Incertae_sedis was opposite.
Acrocalymma was enriched under LDPE and wood chips + LDPE treatments in the MVG period. The relative abundance of
Hypoxylon increased under the treatment of wood chips in the MVG period.
. The species composition of the top 10 phyla of soil (<strong>a</strong>) bacteria (<strong>b</strong>) fungi and the top 20 genera of soil (<strong>c</strong>) bacteria (<strong>d</strong>) fungi under different treatments. Random forest analysis indicates species with significant differences between (<strong>e</strong>) bacterial and (<strong>f</strong>) fungal species groups. (C: control; W: wood chips; N: LDPE; WN: wood chips + LDPE. 8: vigorous season of maize growth; 4: vigorous season of wheat growth).
Chemoheterotrophs and aerobic chemoheterotrophy are the main bacterial functional groups (relative abundance > 10%,
a). Under LDPE treatment, the relative abundance of aromatic compound degradation and cellulose decomposition functional groups decreased compared to the control, except for a 57.80% increase in cellulose decomposition during the MVG period. Wood chips addition alleviated the inhibition of cellulose decomposition during the WVG period. In the MVG and WVG periods, LDPE treatment reduced the relative abundance of functional groups related to nitrate reduction, chitinolysis, nitrogen respiration, and nitrate respiration, while wood chips + LDPE treatment increased these groups during the WVG period. The relative abundance of animal parasites or symbionts and human-associated functional groups decreased by 11.25% and 12.37% respectively, under wood chips treatment in the MVG period, but increased by 13.04% and 11.97% respectively, in the WVG period compared to the control. LDPE treatment consistently reduced these groups in both periods.
Pathotroph-Saprotroph-Symbiotroph and Pathotroph-Saprotroph are the main functional groups of fungi (relative abundance > 10%,
b). Pathotroph-Saprotroph-Symbiotroph groups increased under LDPE and wood chips + LDPE treatments during the WVG period. The relative abundance of Saprotroph-Symbiotroph groups decreased under LDPE treatment in both periods, with additional decreases under wood chips + LDPE treatment in the WVG period. Symbiotroph groups decreased under LDPE treatment in both periods. Pathotroph groups increased by 70.19% and 111.88% respectively under LDPE and wood chips + LDPE treatments in the MVG period, and increased by 2.84% and 32.29% respectively under wood chips and wood chips + LDPE treatments in the WVG period. Pathotroph-Symbiotroph groups increased by 56.73% under LDPE treatment in the MVG period but decreased by 41.85% in the WVG period; adding wood chips reversed this trend.
. Relative abundances predicted by the top 20 functions of soil (<strong>a</strong>) bacterial and (<strong>b</strong>) fungal community under each treatment (C: control; W: wood chips; N: LDPE; WN: wood chips + LDPE. 8: vigorous season of maize growth; 4: vigorous season of wheat growth).
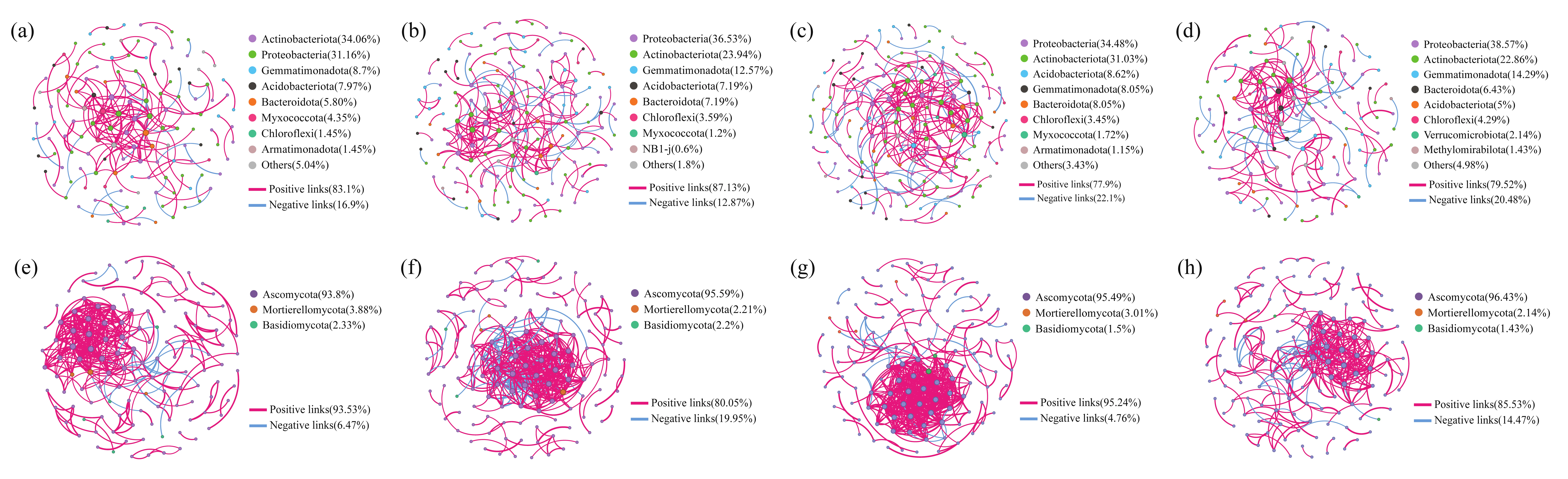
LDPE increased the number of nodes, links, average degree, and negative link ratio in bacterial and fungal networks (
,
). It decreased bacterial network density by 16.67% while increasing fungal network density and clustering coefficient by 14.63% and 12.67% respectively. Bacterial network modularity increased by 0.57%, whereas fungal network modularity decreased by 13.61%. Wood chips reduced total links, average degree, and graph density but increased average path length and modularity for both networks. They also increased the bacterial clustering coefficient by 22.52% while decreasing the fungal clustering coefficient by 23.16%, with opposite effects on the negative link ratio (
).
Figure 6. Co-occurrence network of bacteria and fungi under non-LDPE, LDPE, non-wood chips and wood chips. (<strong>a</strong>) bacteria under non-LDPE; (<strong>b</strong>) bacteria under LDPE; (<strong>c</strong>) bacteria under non-wood chips; (<strong>d</strong>) bacteria under wood chips; (<strong>e</strong>) fungi under non-LDPE; (<strong>f</strong>) fungi under LDPE; (<strong>g</strong>) fungi under non-wood chips; (<strong>h</strong>) fungi under wood chips (Non-LDPE: control and wood chips treatments; LDPE: LDPE and LDPE+ wood chips treatments; non-wood chips: control and LDPE treatments; wood chips: wood chips and LDPE+ wood chips treatments).
.
Topological index of bacterial and fungal co-occurrence networks.
|
|
Total Nodes |
Total Links |
Average Degree |
Average path Length |
Graph Density |
Clustering Coefficient |
Modularity |
| Bacteria |
Non-LDPE |
138 |
171 |
2.478 |
3.737 |
0.018 |
0.366 |
0.700 |
| LDPE |
167 |
213 |
2.551 |
3.961 |
0.015 |
0.333 |
0.704 |
| Non-wood chips |
174 |
276 |
3.172 |
4.146 |
0.018 |
0.302 |
0.657 |
| wood chips |
140 |
166 |
2.371 |
4.262 |
0.017 |
0.370 |
0.701 |
| Fungi |
Non-LDPE |
129 |
340 |
5.271 |
2.314 |
0.041 |
0.592 |
0.441 |
| LDPE |
136 |
436 |
6.412 |
2.803 |
0.047 |
0.667 |
0.381 |
| Non-wood chips |
133 |
441 |
6.632 |
2.271 |
0.05 |
0.721 |
0.318 |
| wood chips |
140 |
318 |
4.543 |
2.882 |
0.033 |
0.554 |
0.514 |
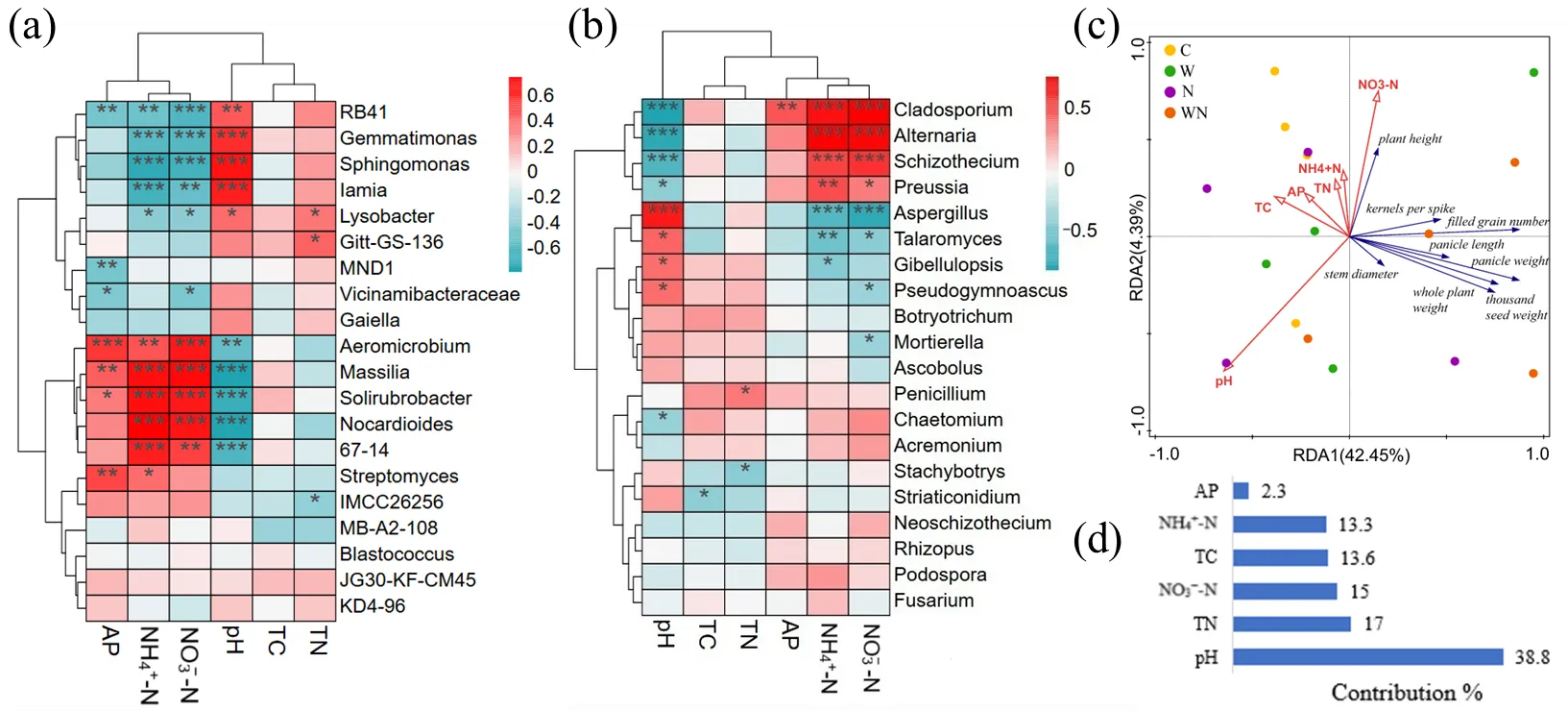
3.6. Factors Affecting Soil Microbial Community and Wheat Growth
Sphingomonas was significantly negatively correlated with soil NH
4+-N and NO
3−-N content and positively correlated with pH.
Nocardioides was the opposite (
a,
p < 0.05).
Vicinamibacteraceae was significantly negatively correlated with soil AP and NO
3−-N content (
a,
p < 0.05).
Cladosporium and
Alternaria were significantly negatively correlated with pH, and significantly negatively correlated with soil NH
4+-N and NO
3−-N content (
b,
p < 0.05).
Stachybotrys was significantly negatively correlated with TN content (
b,
p < 0.05).
. Heat map of correlations between the top 20 genera of soil (<strong>a</strong>) bacteria (<strong>b</strong>) fungi abundance and physicochemical properties. (<strong>c</strong>) Redundancy analysis and of soil properties and wheat growth traits. (<strong>d</strong>) The contribution rate of environmental factors. Significance levels: <sup>∗</sup> <em>p</em> < 0.05, <sup>∗∗</sup> <em>p</em> < 0.01, <sup>∗∗∗</sup> <em>p</em> < 0.001.
The relationship between soil properties and wheat growth traits was further evaluated by redundancy analysis. The interpretation rates of RDA1 and RDA2 axes were 42.45% and 4.39%, respectively (
c). The results showed that pH had the highest contribution rate to wheat growth, reaching 33.8% (
d). It was negatively correlated with all growth parameters (
c). The second most important factor was TN, which explained 17% of the variation (
d). TN was only positively correlated with plant height and negatively correlated with other growth parameters (
c).
4. Discussion
4.1. Wood Chips Improve the Fertility of MP-Contaminated Soil
In our research, LDPE film slightly increased soil pH (about 0.5%), which is consistent with other studies [
19]. It may be due to increased soil aeration and porosity when these MPs are added to the soil, altering the soil biota and thus affecting soil pH [
20]. The addition of wood chips can reduce the pH value (about 1.18%) because wood chips contain acidic substances such as crude tannic acid, and the decomposed organic matter can lower the soil pH [
21] We found that LDPE reduced soil TC, TN, NH
4+-N, and AP, while adding wood chips increased the content of these soil nutrients. Yu et al. (2021) also found that TP, TN, and K in soil aggregates decreased significantly after adding PE microplastics to soil [
22]. The hydrophobicity and high molecular weight of MPs enable them to adsorb nutrients and other contaminants, reducing their bioavailability to plants and soil organisms [
23]. Additionally, MPs can physically obstruct root surfaces and alter soil properties, further exacerbating the decrease in nutrient availability[
24]. Wood chips can improve soil fertility and release valuable nutrients over time, thus improving the soil environment and characteristics.
4.2. Wood Chips Mitigate the Harmful Effects of Microplastics on Plants
Our results showed that LDPE plastic residues negatively affect wheat growth, which has been discussed and reported in previous studies [
25]. The presence of mulch residues in soil can destroy plant growth through various mechanisms. One possible explanation may be that plastic residues in the soil change the soil properties and reduce the nutrient content of the soil, thereby inhibiting plant growth [
26]. It is clear from our study that the addition of wood chips had a positive effect on the growth of wheat (except for stem diameter) and mainly alleviated the damage caused by plastic residues. This is because, during the decomposition process, wood chips release some nutrients, such as N, P, K, and other elements, which are essential for wheat growth [
21]. RDA analysis also showed that pH affected wheat growth the most, and pH was only positively correlated with stem diameter and negatively correlated with other growth parameters. Generally, plants can grow well in neutral, weakly acidic, and weakly alkaline soils,
i.e., pH ranges from 6.5 to 7.5. This is because the nutrients required by plants are most available in this range, which is beneficial for plant uptake and utilization [
27]. The addition of wood chips reduced the pH of the alkaline soil in this study, which may also be why it promotes crop growth. Moreover, it may also be attributed to the wood chips increasing the relative abundance of beneficial microorganisms, such as
Trichoderma. Trichoderma can use direct biological control mechanisms to act on pathogens and indirect mechanisms to enhance plants [
28]. Studies have also shown that it can degrade plastics [
29].
4.3. Wood Chips Restore the Fungal Communities in MP-Contaminated Soil and Enhance Their Functional Capacity
The α diversity index of soil bacteria increased first and then decreased with the addition of LDPE-MP, while the α diversity index of fungi showed the opposite trend. As exogenous particles with hydrophobic surfaces, LDPE-MPs are highly likely to provide new niches for heterotrophic microbial activities [
30]. Bacteria grow much faster, enabling them to occupy organic resources more quickly [
31]. Over time, MPs will have a negative impact on the alpha diversity of bacteria in soil [
5], which may be due to the difference in growth factors between different bacterial species [
32]. This was also confirmed by the significant difference in bacterial communities between the control and LDPE treatments at the later stage of the experiment. Fungi have higher hydrophobicity and more effective extracellular enzymes than bacteria, allowing them to utilize hydrophobic compounds through oxidation and hydrolysis [
31]. It is generally believed that the ability of fungi to degrade MPs is generally higher than that of bacteria [
33]. We found that only the fungal community was significantly separated between the control and LDPE treatment in the early stage of the experiment. The fungal community may be more sensitive to MPs than bacteria, which is consistent with the results of other studies [
34]. In the early stage, fungi enriched some plastic-degrading bacteria, such as Aspergillus. Adding wood chips can improve the α diversity of fungal communities, not bacteria. This is because fungi play an important role in wood fiber degradation because they have enzymatic tools for degrading stubborn plant polymers [
35].
Nocardioides is a genus of Actinomycetes that can tolerate various low nutritional conditions. It can use a variety of organic materials (such as C source and N source) to degrade pollutants [
36], so it is increased by 6.38% under LDPE treatment. The relative abundance of
Georgenia increased under the treatment of wood chips in the MVG period. Although
Georgenia itself is not a plant pathogen, it may be involved in the pathogenic mechanism of plant-microorganism interaction [
37].
Cladosporium and
Alternaria were negatively correlated with pH, and the pH value decreased during the vigorous growth season of wheat, so these two fungi increased during the vigorous growth season of wheat. These fungal genera are well-known fungal pathogens [
38], but
Cladosporium,
Alternaria, and
Aspergillus are effective LDPE film degrading agents with the ability to degrade and modify traditional polymers [
39,
40,
41]. Therefore, MPs are considered a persistent host and potential carrier of fungal pathogens in the soil environment.
Hypoxylon is usually an ascomycete often found on rotten wood, and most have a strong ability to degrade cellulose and lignin, which explains why its relative abundance increased under wood chips treatment [
42]. The correlation heatmap showed that the relative abundance of
Stachybotrys was negatively correlated with TN, while LDPE treatment negatively correlated with TN content, which explained its increase under LDPE treatment.
Stachybotrys is a filamentous fungus that occurs in cellulose rich humid environments and supplements nutrients by degrading cellulose in decaying plants [
43]. At the same time,
Stachybotrys has been proven to be related to human diseases [
44].
The results indicated that LDPE treatment inhibited the relative abundance of C and N cycling functional groups, while wood chips addition promoted the relative abundance of most C and N cycling functional groups. These results are consistent with the results of our soil enzyme activity changes. The activity of P-related enzymes also showed such changes. This may be because changes in soil physical and chemical properties can affect plant physiological status, root characteristics, and nutrient availability, which can indirectly affect enzyme activity [
45]. The wood chips can provide organic amendments to increase organic matter (SOM) content, and soil enzyme activity was positively correlated with SOM content [
46]. The relative abundance of human pathogens and human pathogens pneumonia-related functional groups increased under all treatments, especially in the wood chips and wood chips + LDPE treatment, indicating that the addition of unfermented wood chips will produce animal parasites and human pathogens. The direct return of wood chips will continue to bring the eggs or diseases on the infected branches into the soil, increasing the incidence of pests and diseases [
47]. Previous studies have shown that MPs carry a large number of pathogenic microorganisms on their surfaces [
48]. In addition, plastic residues enrich some pathogenic genes and serve as a transport carrier for these pathogens, posing a threat to ecosystem security and human health [
49].
The Pathotroph-Saprotroph-Symbiotroph fungal group is dominant, likely due to the dominant genus
Fusarium and
Botryotrichum belonging to this trophic type.
Fusarium is the dominant fungal genus in the soil studied in this study because its members include several opportunistic potential plant pathogens [
50]. However, it can also degrade a variety of environmental pollutants (including PE fragments) and is also a key group of soil organic matter decomposition rate [
51]. Saprotroph fungi increased under wood chips and LDPE treatments because wood chips contain a large amount of lignin and cellulose, which has a more obvious effect on fungi related to the degradation of organic matter in soil. The increase in the number of saprophytic fungi in soil is conducive to the degradation of soil organic carbon, thereby increasing soil nutrient content [
52]. This was also confirmed by the enrichment of Trichoderma in this study. The enrichment of saprophytic bacteria in LDPE may be due to its promoting effect on plastic degradation, such as
Aspergillus. Li et al. (2023) also showed that PE MPs exposure promoted the growth of saprophytic bacteria [
53]. Pathotroph is a general term for pathogens that obtain nutrients by destroying host cells and attacking crops [
54]. Pathotroph is enriched under wood chips and LDPE treatments, such as
Stachybotrys and
Alternaria. This is consistent with bacterial function prediction results, indicating that wood chips and LDPE can increase pathogenic microorganisms. Symbiotroph obtains nutrients by exchanging resources with host cells. Symbiotroph decreased in both periods of LDPE treatment, consistent with the results that LDPE increased the competition of the microbial co-occurrence network.
4.4. LDPE Exposure Increased, But Wood Chips Addition Reduced the Complexity of the Bacterial and Fungal Co-Occurrence Network
Our results suggested that LDPE exposure increases the complexity of bacterial and fungal networks, reduces the interaction of bacterial members but makes the bacterial network more stable, makes the fungal network connection closer but unstable, and increases the competition between bacterial and fungal members. Habitat heterogeneity, niche differentiation, and environmental selection may affect the module [
55]. Ecological network theory predicts that communities of closely related species should be more vulnerable [
56]. Therefore, the disappearance of some species has less impact on the entire network, indicating that the network is more stable. LDPE addition increased the negative association of microbial co-occurrence networks, which may be due to limited resource competition. PE MPs is likely to be carbon and/or release additives and small molecule compounds, which in turn contributes to niche overlap between specific taxa in the microbial community [
3]. It showed that the addition of wood chips reduced the complexity of bacterial and fungal networks but increased stability. Wood chips can increase the tightness and cooperation of the bacterial network and make the fungal network looser and more competitive. Nutrient-rich wood input materials may lead to more complex symbiotic networks and stronger connectivity [
57]. In this study, compared with wood chips addition, non-wood chips addition reduced nutrients, which may help to form more complex soil microbial relationships to cope with limited resource challenges [
58]. The high modular value indicates a strong niche differentiation in the soil. This differentiation can be attributed to abundant plant resources and improved soil conditions, which create more diverse niches for microorganisms [
59]. Therefore, the increase in niche differentiation may lead to a decrease in microbial interactions in the soil [
60], which explains the low number of bacterial and fungal connections observed in this study.
5. Conclusions
Our study found that the application of wood chips promoted the activities of carbon, nitrogen, and phosphorus-related enzymes, which alleviated the inhibition of soil nutrient cycle processes by microplastics and improved the availability of soil nutrients, thus effectively promoting the growth of wheat in MPs-contaminated soil. Fungal communities might be more sensitive to MPs than bacteria, and MPs could negatively affect the α diversity of bacteria and fungi in soil. Adding wood chips increased the α diversity of fungal but not bacterial communities. Both LDPE and wood chips at the later stage treatment increased pathotrophic fungi. Wood chips could increase the number of saprophytic fungi that degrade microplastics. LDPE reduced symbiotroph fungi. LDPE exposure increased the complexity of the bacterial and fungal co-occurrence networks, making the bacterial co-occurrence network more stable and the co-occurrence fungal network less stable. The addition of wood chips reduced the complexity of bacterial and fungal co-occurrence networks but enhanced the stability. These findings broaden our understanding of soil properties, rhizosphere microbial community transformation characteristics, functions, and networks after microplastic pollution and wood chips modification, and provide a basis for using wood chips as a soil amendment for microplastic pollution in farmland. In the future, it is recommended that the potential of enhancing microplastic pollution mitigation through the pre-composting of wood chips prior to its application be investigated to minimize the risk of pathogen transmission.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/638, Table S1. Basic physical and chemical properties of soil; Table S2. Results of the permutation multivariate analysis of variance.
Acknowledgments
We thank Jing Wang, Biqi Bao and Guotao Du for their contributions to the laboratory work and Shixia Wan and Shijie Han for assisting with fieldwork.
Author Contributions
Conceptualization, J.Z. and F.R.; Methodology, J.Z.; Formal Analysis, Y.C. and F.R.; Investigation, Y.C., Z.S. and Y.W.; Data Curation, Y.C.; Writing—Original Draft Preparation, Y.C.; Writing—Review & Editing, H.L., M.X.; Visualization, Y.C.; Supervision, J.Z.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
This research received no external funding.
Funding
This study was financially supported by the National Natural Science Foundation of China (42273084) and Natural Science Foundation of Henan Province (242300421039).
Declaration of Competing Interest
The authors declare no conflict of interest in this paper.
References
-
1.
Xiang Y, Jiang L, Zhou Y, Luo Z, Zhi D, Yang J, et al. Microplastics and environmental pollutants: Key interaction and toxicology in aquatic and soil environments.
J. Hazard. Mater. 2022,
422, 126843.
[Google Scholar]
-
2.
Dioses-Salinas DC, Pizarro-Ortega CI, De-la-Torre GE. A methodological approach of the current literature on microplastic contamination in terrestrial environments: Current knowledge and baseline considerations.
Sci. Total Environ. 2020,
730, 139164.
[Google Scholar]
-
3.
Li K, Zhang M, Jia W, Xu L, Huang Y. Deciphering the effects of LDPE microplastic films on diversity, composition and co-occurrence network of soil fungal community.
Appl. Soil Ecol. 2023,
182, 104716.
[Google Scholar]
-
4.
Liu EK, He WQ, Yan CR. ‘White revolution’ to ‘white pollution’—agricultural plastic film mulch in China.
Environ. Res. Lett. 2014,
9, 091001.
[Google Scholar]
-
5.
Huang Y, Zhao Y, Wang J, Zhang M, Jia W, Qin X. LDPE microplastic films alter microbial community composition and enzymatic activities in soil.
Environ. Pollut. 2019,
254, 112983.
[Google Scholar]
-
6.
Zhu D, Ma J, Li G, Rillig MC, Zhu Y-G. Soil plastispheres as hotspots of antibiotic resistance genes and potential pathogens.
ISME J. 2022,
16, 521–532.
[Google Scholar]
-
7.
Fei Y, Huang S, Zhang H, Tong Y, Wen D, Xia X, et al. Response of soil enzyme activities and bacterial communities to the accumulation of microplastics in an acid cropped soil.
Sci. Total Environ. 2020,
707, 135634.
[Google Scholar]
-
8.
Wang J, Liu X, Li Y, Powell T, Wang X, Wang G, et al. Microplastics as contaminants in the soil environment: A mini-review.
Sci. Total Environ. 2019,
691, 848–857.
[Google Scholar]
-
9.
Guo W, Ye Z, Zhao Y, Lu Q, Shen B, Zhang X, et al. Effects of different microplastic types on soil physicochemical properties, enzyme activities, and bacterial communities.
Ecotoxicol. Environ. Saf. 2024,
286, 117219.
[Google Scholar]
-
10.
Yang W, Cheng P, Adams CA, Zhang S, Sun Y, Yu H, et al. Effects of microplastics on plant growth and arbuscular mycorrhizal fungal communities in a soil spiked with ZnO nanoparticles.
Soil Biol. Biochem. 2021,
155, 108179.
[Google Scholar]
-
11.
Li B, Huang S, Wang H, Liu M, Xue S, Tang D, et al. Effects of plastic particles on germination and growth of soybean (
Glycine max): A pot experiment under field condition.
Environ. Pollut. 2021,
272, 116418.
[Google Scholar]
-
12.
Zhuang Q-L, Yuan H-Y, Sun M, Deng H-G, Zama EF, Tao B-X, et al. Biochar-mediated remediation of low-density polyethylene microplastic-polluted soil-plant systems: Role of phosphorus and protist community responses.
J. Hazard. Mater. 2025,
486, 137076.
[Google Scholar]
-
13.
Fontana M, Johannes A, Zaccone C, Weisskopf P, Guillaume T, Bragazza L, et al. Improving crop nutrition, soil carbon storage and soil physical fertility using ramial wood chips.
Environ. Technol. Innov. 2023,
31, 103143.
[Google Scholar]
-
14.
Rössert S, Gosling E, Gandorfer M, Knoke T. Woodchips or potato chips? How enhancing soil carbon and reducing chemical inputs influence the allocation of cropland.
Agric. Syst. 2022,
198, 103372.
[Google Scholar]
-
15.
Spisak W, Chlebicki A, Wołowski K, Szar M, Kozak J, Kaszczyszyn M. Using wood chips for the protection of plants and soil from the harmful effects of road salt.
Eur. J. Wood Wood Prod. 2020,
78, 1209–1219.
[Google Scholar]
-
16.
Wang Y, Wang M, Wang Q, Wang T, Zhou Z, Mehling M, et al. Flowthrough Capture of Microplastics through Polyphenol-Mediated Interfacial Interactions on Wood Sawdust.
Adv. Mater. 2023,
35, 2301531.
[Google Scholar]
-
17.
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, et al. The SILVA ribosomal RNA gene database project: improved data processing and web-based tools.
Nucleic Acids Res. 2012,
41, D590–D596.
[Google Scholar]
-
18.
Kõljalg U, Nilsson RH, Abarenkov K, Tedersoo L, Taylor AFS, Bahram M, et al. Towards a unified paradigm for sequence-based identification of fungi.
Mol. Ecol. 2013,
22, 5271–5277.
[Google Scholar]
-
19.
Wang F, Zhang X, Zhang S, Zhang S, Sun Y. Interactions of microplastics and cadmium on plant growth and arbuscular mycorrhizal fungal communities in an agricultural soil.
Chemosphere 2020,
254, 126791.
[Google Scholar]
-
20.
Lozano YM, Lehnert T, Linck LT, Lehmann A, Rillig MC. Microplastic Shape, Polymer Type, and Concentration Affect Soil Properties and Plant Biomass.
Front. Plant Sci. 2021,
12, 616645.
[Google Scholar]
-
21.
Li Z, Schneider RL, Morreale SJ, Xie Y, Li C, Li J. Woody organic amendments for retaining soil water, improving soil properties and enhancing plant growth in desertified soils of Ningxia, China.
Geoderma 2018,
310, 143–152.
[Google Scholar]
-
22.
Yu H, Qi W, Cao X, Hu J, Li Y, Peng J, et al. Microplastic residues in wetland ecosystems: Do they truly threaten the plant-microbe-soil system?
Environ. Int. 2021,
156, 106708.
[Google Scholar]
-
23.
Mondol M, Angon PB, Roy A. Effects of microplastics on soil physical, chemical and biological properties.
Nat. Hazards Res. 2025,
5, 14–20.
[Google Scholar]
-
24.
Somanathan H, Sathasivam R, Sivaram S, Mariappan Kumaresan S, Muthuraman MS, Park SU. An update on polyethylene and biodegradable plastic mulch films and their impact on the environment.
Chemosphere 2022,
307, 135839.
[Google Scholar]
-
25.
Qi Y, Yang X, Pelaez AM, Huerta Lwanga E, Beriot N, Gertsen H, et al. Macro- and micro-plastics in soil-plant system: Effects of plastic mulch film residues on wheat (
Triticum aestivum) growth.
Sci. Total Environ. 2018,
645, 1048–1056.
[Google Scholar]
-
26.
Dewi SK, Han ZM, Bhat SA, Zhang F, Wei Y, Li F. Effect of plastic mulch residue on plant growth performance and soil properties.
Environ. Pollut. 2024,
343, 123254.
[Google Scholar]
-
27.
Neina D. The Role of Soil pH in Plant Nutrition and Soil Remediation.
Appl. Environ. Soil Sci. 2019,
2019, 5794869.
[Google Scholar]
-
28.
Woo SL, Hermosa R, Lorito M, Monte E.
Trichoderma: a multipurpose, plant-beneficial microorganism for eco-sustainable agriculture.
Nat. Rev. Microbiol. 2023,
21, 312–326.
[Google Scholar]
-
29.
Bernat P, Jasińska A, Niedziałkowska K, Słaba M, Różalska S, Paraszkiewicz K, et al. Adaptation of the metolachlor-degrading fungus
Trichoderma harzianum to the simultaneous presence of low-density polyethylene (LDPE) microplastics.
Ecotoxicol. Environ. Saf. 2023,
267, 115656.
[Google Scholar]
-
30.
Arias-Andres M, Kettner MT, Miki T, Grossart H-P. Microplastics: New substrates for heterotrophic activity contribute to altering organic matter cycles in aquatic ecosystems.
Sci. Total Environ. 2018,
635, 1152–1159.
[Google Scholar]
-
31.
Wang C, Kuzyakov Y. Mechanisms and implications of bacterial–fungal competition for soil resources.
ISME J. 2024,
18, wrae073.
[Google Scholar]
-
32.
Li H-Z, Zhu D, Lindhardt JH, Lin S-M, Ke X, Cui L. Long-Term Fertilization History Alters Effects of Microplastics on Soil Properties, Microbial Communities, and Functions in Diverse Farmland Ecosystem.
Environ. Sci. Technol. 2021,
55, 4658–4668.
[Google Scholar]
-
33.
Du H, Xie Y, Wang J. Microplastic degradation methods and corresponding degradation mechanism: Research status and future perspectives.
J. Hazard. Mater. 2021,
418, 126377.
[Google Scholar]
-
34.
Yuan Y, Zu M, Li R, Zuo J, Tao J. Soil properties, microbial diversity, and changes in the functionality of saline-alkali soil are driven by microplastics.
J. Hazard. Mater. 2023,
446, 130712.
[Google Scholar]
-
35.
Ran W, Lian J, Zhang J. Evaluation and modeling of fungi towards wood degradation.
Bioengineered 2022,
13, 3284–3299.
[Google Scholar]
-
36.
Tawfik A, Eraky M, Osman AI, Ai P, Zhou Z, Meng F, et al. Bioenergy production from chicken manure: A review.
Environ. Chem. Lett. 2023,
21, 2707–2727.
[Google Scholar]
-
37.
Patel PP, Rakhashiya PM, Thaker VS. Genomic analysis of novel phytopathogenic
Georgenia sp. strain SUB25.
Genom. Data 2015,
5, 320–322.
[Google Scholar]
-
38.
Gkoutselis G, Rohrbach S, Harjes J, Obst M, Brachmann A, Horn MA, et al. Microplastics accumulate fungal pathogens in terrestrial ecosystems.
Sci. Rep. 2021,
11, 13214.
[Google Scholar]
-
39.
Tamoor M, Samak NA, Jia Y, Mushtaq MU, Sher H, Bibi M, et al. Potential Use of Microbial Enzymes for the Conversion of Plastic Waste Into Value-Added Products: A Viable Solution.
Front. Microbiol. 2021,
12, 777727.
[Google Scholar]
-
40.
Sathiyabama M, Boomija RV, Sathiyamoorthy T, Mathivanan N, Balaji R. Mycodegradation of low-density polyethylene by
Cladosporium sphaerospermum, isolated from platisphere.
Sci. Rep. 2024,
14, 8351.
[Google Scholar]
-
41.
Verma N, Gupta S. Assessment of LDPE degrading potential
Aspergillus species isolated from municipal landfill sites of Agra.
SN Appl. Sci. 2019,
1, 701.
[Google Scholar]
-
42.
Ma H, Song Z, Pan X, Li Y, Yang Z, Qu Z. Multi-Gene Phylogeny and Taxonomy of
Hypoxylon (Hypoxylaceae, Ascomycota) from China.
Diversity 2022,
14, 37.
[Google Scholar]
-
43.
Lombard L, Houbraken J, Decock C, Samson RA, Meijer M, Réblová M, et al. Generic hyper-diversity in
Stachybotriaceae.
Persoonia 2016,
36, 156–246.
[Google Scholar]
-
44.
Terr AI.
Stachybotrys: Relevance to human disease.
Ann. Allergy Asthma Immunol. 2001,
87, 57–63.
[Google Scholar]
-
45.
de Souza Machado AA, Lau CW, Kloas W, Bergmann J, Bachelier JB, Faltin E, et al. Microplastics Can Change Soil Properties and Affect Plant Performance.
Environ. Sci. Technol. 2019,
53, 6044–6052.
[Google Scholar]
-
46.
Wu L, Ma H, Zhao Q, Zhang S, Wei W, Ding X. Changes in soil bacterial community and enzyme activity under five years straw returning in paddy soil.
Eur. J. Soil Biol. 2020,
100, 103215.
[Google Scholar]
-
47.
Callan BE, Leal I, Islam A, Foord B. Analysis of Canadian wood pellets for the presence of living phytopathogenic fungi and pinewood nematode.
EPPO Bull. 2018,
48, 245–253.
[Google Scholar]
-
48.
Tavşanoğlu ÜN, Akca G, Pekmez T, Başaran Kankılıç G, Çırak T, Çağan AS, et al. Increasing microplastics pollution: An emerging vector for potentially pathogenic bacteria in the environment.
Water Res. 2025,
274, 123142.
[Google Scholar]
-
49.
Amaral-Zettler LA, Zettler ER, Mincer TJ. Ecology of the plastisphere.
Nat. Rev. Microbiol. 2020,
18, 139–151.
[Google Scholar]
-
50.
Summerell BA, Laurence MH, Liew ECY, Leslie JF. Biogeography and phylogeography of
Fusarium: A review.
Fungal Divers. 2010,
44, 3–13.
[Google Scholar]
-
51.
Xiao D, He X, Wang G, Xu X, Hu Y, Chen X, et al. Network analysis reveals bacterial and fungal keystone taxa involved in straw and soil organic matter mineralization.
Appl. Soil Ecol. 2022,
173, 104395.
[Google Scholar]
-
52.
Marschner P, Umar S, Baumann K. The microbial community composition changes rapidly in the early stages of decomposition of wheat residue.
Soil Biol. Biochem. 2011,
43, 445–451.
[Google Scholar]
-
53.
Crowther TW, Boddy L, Hefin Jones T. Functional and ecological consequences of saprotrophic fungus–grazer interactions.
ISME J. 2012,
6, 1992–2001.
[Google Scholar]
-
54.
Schmidt R, Mitchell J, Scow K. Cover cropping and no-till increase diversity and symbiotroph:saprotroph ratios of soil fungal communities.
Soil Biol. Biochem. 2019,
129, 99–109.
[Google Scholar]
-
55.
Li K, Xu L, Bai X, Zhang G, Zhang M, Huang Y. Differential fungal assemblages and functions between the plastisphere of biodegradable and conventional microplastics in farmland.
Sci. Total Environ. 2024,
906, 167478.
[Google Scholar]
-
56.
Widder S, Besemer K, Singer GA, Ceola S, Bertuzzo E, Quince C, et al. Fluvial network organization imprints on microbial co-occurrence networks.
Proc. Natl. Acad. Sci. USA 2014,
111, 12799–12804.
[Google Scholar]
-
57.
Zhu P, Li Y, Gao Y, Yin M, Wu Y, Liu L, et al. Insight into the effect of nitrogen-rich substrates on the community structure and the co-occurrence network of thermophiles during lignocellulose-based composting.
Bioresour. Technol. 2021,
319, 124111.
[Google Scholar]
-
58.
Che R, Liu D, Qin J, Wang F, Wang W, Xu Z, et al. Increased litter input significantly changed the total and active microbial communities in degraded grassland soils.
J. Soils Sediments 2020,
20, 2804–2816.
[Google Scholar]
-
59.
Lin Q, Li L, Adams JM, Heděnec P, Tu B, Li C, et al. Nutrient resource availability mediates niche differentiation and temporal co-occurrence of soil bacterial communities.
Appl. Soil Ecol. 2021,
163, 103965.
[Google Scholar]
-
60.
Wang F, Li Z, Fu B, Lü Y, Liu G, Wang D, et al. Short-Term Grazing Exclusion Alters Soil Bacterial Co-occurrence Patterns Rather Than Community Diversity or Composition in Temperate Grasslands.
Front. Microbiol. 2022,
13, 824192.
[Google Scholar]