The efficiency of industrial production was highly dependent on the support of a large number of raw materials, and the same is true for biological production. The global bio-manufactured product market keeps expanding, indirectly increasing the demand for developing various carbon sources. Currently, the carbon sources used in biological production are mainly food grain-derived C6 compounds, such as glucose, starch, and molasses [
1,
2]. However, the global food security situation remains severe. According to a report from the Food and Agriculture Organization (FAO) of the United Nations, since 2019, the global prevalence of food insecurity has been increasing. By 2023, 733 million people worldwide were facing hunger, and 2.33 billion people were in a state of moderate or severe food insecurity [
3]. With the continued population growth, the principle of “not competing with humans for food” in biological production remains increasingly important. Against this backdrop, using C2 substances (such as ethylene glycol, ethanol, and acetic acid) as carbon sources for biological production has attracted more and more attention. According to the Web of Science database, about 50 papers on this topic have been published in the past three years.
C2 substances have numerous sources, including cellulose and waste from food production [
4,
5,
6]. These features enable C2 substances to be obtained without relying on food-related materials. For example, through the hydrolysis of polyethylene terephthalate (PET) plastics, ethylene glycol can be efficiently produced together with terephthalic acid and other valuable chemical units [
7,
8]. In addition, with the advancement of electrochemical technology, the electrochemical or photo-electrochemical reduction of CO
2 has been applied to convert it to ethylene glycol, ethanol and acetic acid [
9,
10,
11]. Therefore, using C2 substances as carbon sources for biological production not only avoids the situation of “competing with humans for food” but also offers significant benefits for environmental protection and carbon neutrality. Moreover, compared to C6 sugars, C2 substances have a shorter pathway to enter central metabolism and can achieve less loss of carbon atoms. Theoretically, their utilization efficiency is higher than that of sugars. Using C2 substances for bioproduction is also more cost-effective. As shown on the online platform (Chemical Product Price Database: Compare and Download Prices Online—ECHEMI), recently, acetic acid (99%, Shandong Baiding Chemical Co., Ltd., Qingdao, China) is about 300–350 $/ton. Ethanol (99%, First Chems Co., Ltd., Xi’an, China) is about 360–380 $/ton. Ethylene glycol (99%, First Chems Co., Ltd., Xi’an, China) is about 360–380 $/ton, while glucose (powder, Wuhan Gongchen International Trading Co., Ltd., Wuhan, China) is about 500–550 $/ton, much expensive than C2 substances. And considering that C6 feedstocks cost two carbon atoms lost in the glycolysis, using C6 feedstocks to produce acetyl-CoA consumes an additional 50% of the feedstock compared to C2 feedstocks, equal to an additional 250–275 $/ton. Although not all products are derivatives of acetyl-CoA, certain pathways can fix CO
2 when the acetyl-CoA produced by C2 substances is replenished into the gluconeogenesis pathway, thereby obtaining free carbon atoms. From this perspective, using C2 substances for bioproduction is still more cost-effective. Additionally, the products achieved using C2 feedstocks show comparable yields with those using C6 feedstocks. However, since C2 substances are not commonly used by most microorganisms in nature, using them as carbon sources requires adaptive modification or engineering of the host microorganisms.
In this review, we summarize the acquisition methods of the three C2 substances—ethylene glycol, ethanol, and acetic acid—as well as the metabolic pathways required for biological production using these substances. We also exemplify some of the recent achievements and present our perspectives on using them in this area.
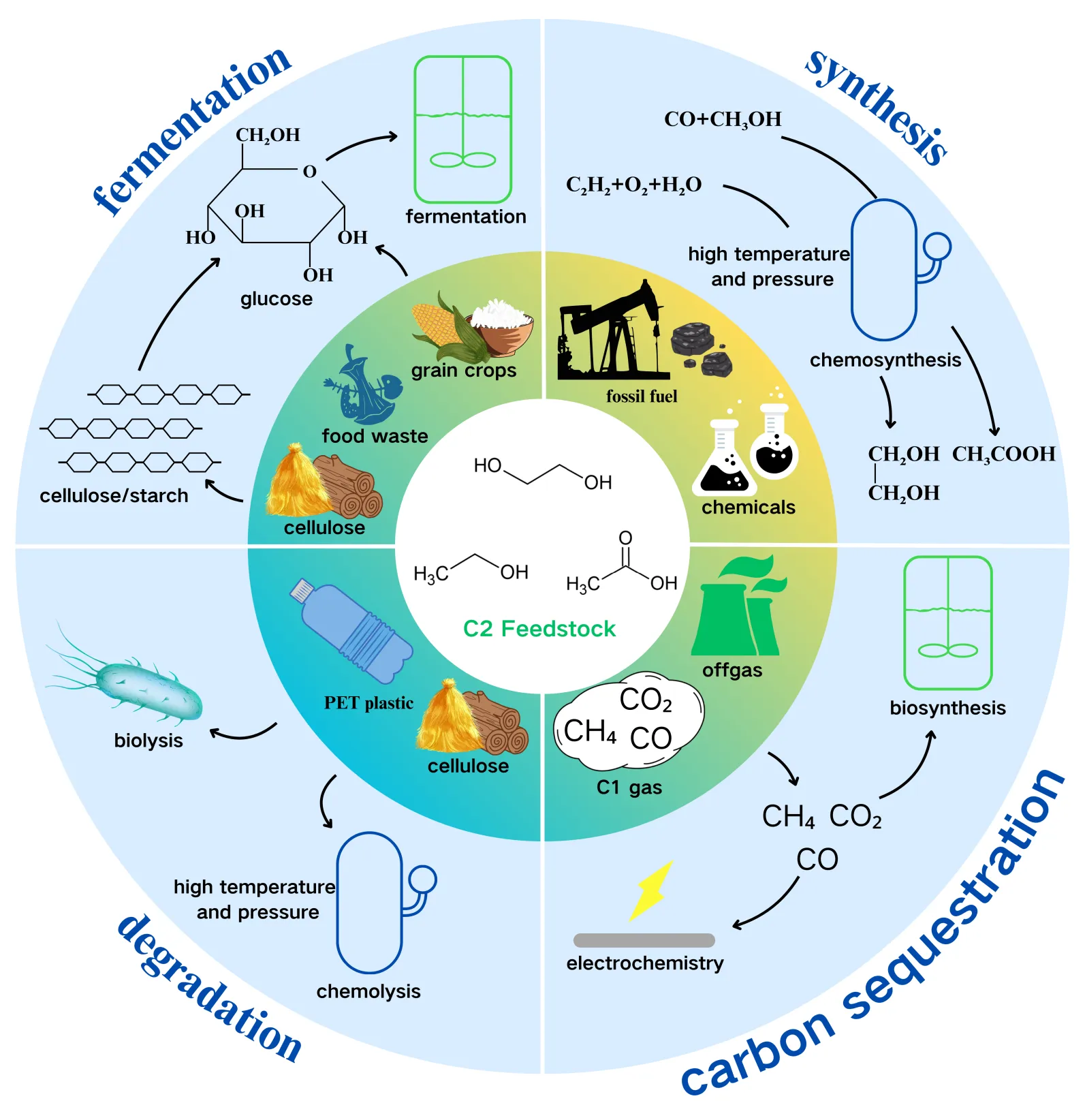
C2 substances are widely available from kinds of sources. In addition to being obtained through fermentation using carbon sources from traditional food crops, C2 substances can also be derived through conventional chemical, electrochemical, and emerging biological methods (as shown in
). Here, we introduce the sources of the three C2 substances and present some perspectives on each.
. Sources of C2 substances.
Ethylene glycol is a bulk chemical with an annual production as high as 20 million tons. It is primarily used as an antifreeze and to synthesize PET plastics [
12]. Currently, ethylene glycol is mainly derived from ethylene. Ethylene can be converted to glycol through a two-step chemical or one-step electrochemical reaction [
13]. In the two-step chemical process, ethylene is first oxidized to ethylene oxide under high temperature and pressure (200–300 °C, 1–3 MPa) in the presence of a silver catalyst [
14]. Subsequently, ethylene oxide is hydrated at 200 °C to produce ethylene glycol. In the emerging electrochemical method, one molecule of ethylene reacts with two molecules of water, catalyzed by a noble metal catalyst (e.g., palladium), to produce ethylene glycol [
13]. However, as a petroleum-derived product, ethylene-derived ethylene glycol has certain disadvantages in terms of sustainability. Therefore, finding green feedstocks for ethylene glycol production is becoming increasingly important. Wang et al. [
15] developed a method to produce ethylene glycol from cellulose. In this method, cellulose is subjected to hydrogenolysis in water under high-pressure hydrogen (6–10 MPa) at 120–150 °C for 0.5–4 h, using a tungsten-based catalyst. This process converts cellulose into ethylene glycol with minimal by-products, and the yield of EG can achieve a rate greater than 60%.
With the growing awareness of plastic pollution and environmental protection, degrading PET plastics to produce ethylene glycol has gained much attention. One example involves glycolyzing PET with ethylene glycol first using a titanium catalyst at 220 °C for 12–18 h to produce bis(2-hydroxyethyl) terephthalate (BHET) [
16]. Subsequently, a series of enzymatic reactions in engineered
Pseudomonas putida degrade BHET stepwise into β-ketoadipate, releasing two molecules of ethylene glycol per molecule of BHET [
17]. The theoretical ratio of consumed to released ethylene glycol is 1:2, resulting in a net production of ethylene glycol. Compared to the above-mentioned absolute chemical methods, the biocatalysis-coupled method [
17] exhibits a significant advantage: the reaction conditions are more moderate and convenient for direct use in the subsequent biological production processes. More importantly, the raw feedstock used in this method is PET plastic, a major environmental concern today. Using this method, the PET plastic can be recycled when producing EG rather than deposited as environmental waste. Therefore, this method reveals significant environmental benefits.
2.2. Sources of Ethanol
Ethanol is a traditional C2 chemical with a wide range of applications in both food and industrial sectors. Unlike the other two C2 compounds, the bio-production of ethanol is highly mature and widely applied. Therefore, the focus here is primarily on the bioproduction methods of ethanol. Currently, most ethanol fermentation feedstocks still come from food materials such as cereal crops and sugar-producing crops [
18,
19], which conflicts with the principle of “not competing with humans for food”. In a traditional fermentation process of ethanol, using diluted molasses from sugarcane can produce ethanol at a concentration of 7–8% (
v/
v) [
20,
21]. And in high-density fermentation processes, ethanol yields can reach 12–14% (
v/
v), or even exceed 15% (
v/
v) [
22,
23].
To overcome the use of food materials for ethanol fermentation, various raw feedstocks, such as by-products or waste from food production [
24], or lignocellulosic biomass [
25], have been explored based on the cell factory in the production processes. In the lignocellulosic biomass-based process, the key is to break down the recalcitrant cellulose into substances that yeast can easily utilize. Therefore, pretreatment to decompose lignocellulosic biomass is crucial. The commonly used methods mainly include acid hydrolysis under heat [
26,
27] and organic solvent treatment [
28]. However, several concerns, such as the low efficiency of pretreatment, high cost, and environmental effects, limit the wide application in most plants. After addressing the technical and economic difficulties, these methods will be adopted increasingly in industrial applications.
For subsequent bioproduction using ethanol as a carbon source, the separation and purification steps are necessary and important to obtain ethanol without impurities. In the mostly used distillation process, the culture medium is heated to 78–80 °C, and then the ethanol vapor is condensed in a condenser to obtain relatively pure liquid ethanol. Through this method, a purity of 96% (
v/
v) can be achieved [
20].
2.3. Sources of Acetic Acid
As a commonly used bulk C2 chemical in several industries, acetic acid has a wide range of sources [
2]. By 2020, the global acetic acid market had reached 16.3 million tons and is expected to reach 19.6 million tons by 2027 [
29]. In current industrial production, acetic acid is mostly produced from petrochemical feedstocks such as methanol, acetaldehyde, ethylene, and
n-butane [
30,
31]. The most widely used method is the classical chemical synthesis from methanol and carbon monoxide, which was first proposed in 1913 [
32]. Nowadays, various metal catalysts for the reaction have been designed to improve the catalytic efficiency under [
33,
34]. However, this chemical method still has several issues. First, the high-temperature (150–200 ℃ [
31]) and high-pressure (30–50 atm [
31]) conditions require significant energy input and impose higher safety requirements on the reaction vessels. Additionally, the reactant conditions for chemical synthesis are not suitable for direct biological utilization, since they usually have extreme pH and include toxic substances such as formaldehyde, as well as toxic catalysts like tungsten and uranium [
34], so it is necessary to separate acetic acid from the reaction system before bioproduction.
In addition, the biological production of acetic acid has been developed using various raw feedstocks, such as sugars, glycerol, lignocellulosic biomass, waste materials, and even C1 gases [
35]. The traditional and mostly used acetic acid bioproduction is the fermentation of sugars by yeast or acetic acid bacteria [
36]. However, the acetic acid produced in this way is usually intended for the food industry and is not cost-effective as a carbon source for the production of other chemicals.
Recently, the abundant non-food biomass and C1 gases exhibit great potential as an alternative in the chemical production of acetic acid. When using lignocellulose biomass as the raw material, the commonly adopted method is pyrolysis. In this process, the material is decomposed at high temperatures in the absence of oxygen, resulting in a mixture of solid biochar, gaseous syngas, and liquid pyrolysis oil. Among these, the pyrolysis oil contains a significant number of organic acids, with acetic acid being the most abundant ingredient [
37]. The pyrolysis temperature has a considerable impact on the composition ratio of the pyrolysis oil [
38], and it is generally believed that the highest yield of organic acids is achieved at 350 °C. The relatively pure acetic acid can be separated and purified from pyrolysis oil through a series of methods [
39]. By this way, the concentration of acetic acid can reach up to 20 g/L. It is worth noting that using the pyrolysis oil mixture directly as a carbon source for production can even bring a high yield, as the other substances can serve as additional carbon sources to assist biological production. In terms of producing acetic acid from C1 gases, using certain special microorganisms for production is also an excellent option, in addition to the emerging electrochemical methods. Hu et al. [
40] designed a specialized reactor called bubble column bioreactor, and used this device to cultivate the anaerobic acetogenic bacterium
Moorella thermoacetica supplied with a mixture of CO and CO
2 gases in a certain ratio. The maximum yield of 31 g/L was achieved for acetic acid from those C1 gases. This study not only reveals a new method for acetic acid production but, more importantly, breaks away from the dependence on petrochemicals for industrial acetic acid production, contributing significantly to carbon neutrality.
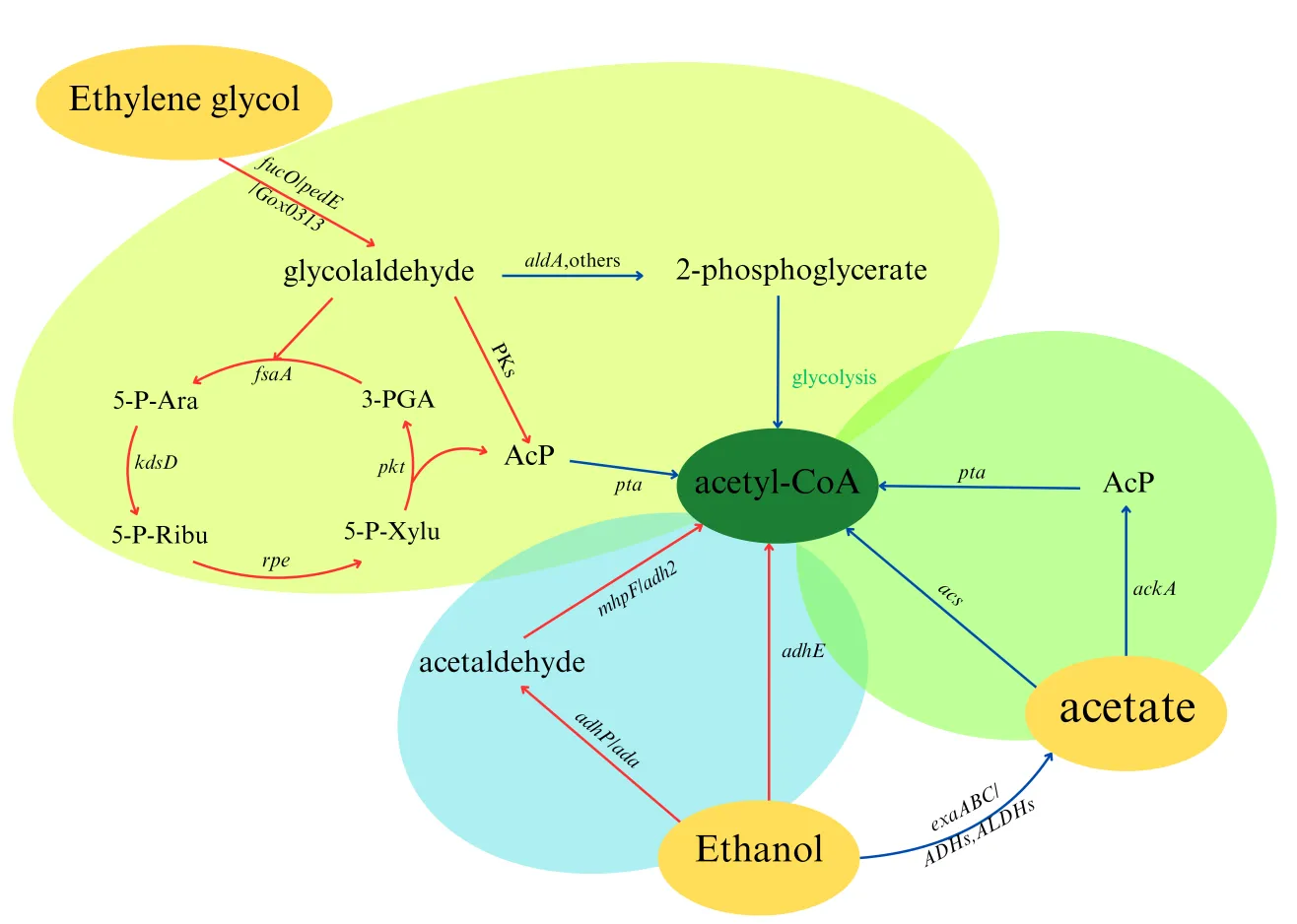
The conversion of C2 substances to acetyl-CoA involves relatively fewer steps and is often devoid of carbon loss. As a key substance in central carbon metabolism, acetyl-CoA can enter the tricarboxylic acid (TCA) cycle downstream and the gluconeogenesis pathway upstream. In biological production, most products can be further metabolized from the TCA cycle products and glycolysis products. Therefore, converting C2 substances to acetyl-CoA is the first and most important step for the utilization of C2 substances in biological systems. In this part, we will introduce the pathways of converting C2 substances to acetyl-CoA (as shown in
) and then list some examples of production using C2 substances.
. Pathways of C2 substances utilization. The blue arrows represent endogenous metabolic pathways, while the red arrows indicate artificially constructed or engineered metabolic pathways. The text on the arrows denotes the genes encoding the enzymes of the pathways.
As an unconventional C2 carbon source, ethylene glycol can’t be utilized for growth by most conventional strains such as
Escherichia coli. However,
E. coli possesses endogenous pathways that can channel glycolaldehyde into the tricarboxylic acid (TCA) cycle. Therefore, the key to converting ethylene glycol into acetyl-CoA lies in transforming ethylene glycol into glycolaldehyde. In some studies, it has been found that overexpression of the
fucO gene, which encodes 1,2-propanediol reductase in
E. coli, can enhance the utilization efficiency of ethylene glycol by
E. coli [
41]. This enzyme is Fe²⁺-dependent and loses activity during aerobic growth. When the point mutations I7L and L8V are introduced, the enzyme can function under aerobic conditions [
42].
In some other organisms, natural pathways for ethylene glycol utilization do exist. For example, it was found that
Pseudomonas putida JM37 (isolated from soil rich in m-xylene) can naturally use ethylene glycol as a carbon source to grow, while the wild-type strain KT2440 can’t. Proteomic analysis discovered a series of genes associated with ethylene glycol utilization, among which the
pedE gene encodes a PQQ-dependent alcohol dehydrogenase (pyrroloquinoline quinone-dependent alcohol dehydrogenase, PQQ-ADH) that can convert ethylene glycol to glycolaldehyde [
43].
Rhodococcus jostii RHA1 can also grow using ethylene glycol. Genomic analysis identified the
egaA gene, which encodes a mycofactocin (MFT)-associated dehydrogenase, as the key to ethylene glycol utilization, and it can also catalyze the oxidation of ethylene glycol to glycolaldehyde [
44]. Additionally, in a study on
Gluconobacter oxydans, the enzyme expressed by the
Gox0313 gene was found to convert ethylene glycol to glycolaldehyde [
45]. In this work, the
pedE,
fucOI7L,L8V, and
Gox0313 genes were introduced into
E. coli to compare their abilities to utilize ethylene glycol, respectively. The results showed that the
Gox0313 gene was significantly more effective than the other two. When using ethylene glycol as the sole carbon source,
E. coli overexpressing the
Gox0313 gene reached a cell concentration of OD
600~6 in 48 h and remained relatively stable, even with the production of glycolaldehyde [
45].
After converting ethylene glycol to glycolaldehyde, for conventional engineered strains of
E. coli, the endogenous pathway mainly involves the conversion of glycolaldehyde to glyoxylate via the genes
aldA and
glcDEF. Subsequently, two molecules of glyoxylate are condensed into tartronate semialdehyde by tartronate semialdehyde synthase encoded by the
gcl gene, with the release of one molecule of CO
2. Tartronate semialdehyde is then converted to 2-phosphoglycerate via GlxRK and GarK, entering the glycolytic pathway and ultimately being converted to acetyl-CoA, with another molecule of CO
2 being released. Therefore, in the endogenous pathway, the carbon utilization rate for converting glycolaldehyde to acetyl-CoA is only 50%. For a bioproduction process, such a high carbon loss is clearly uneconomical, so finding a pathway with higher carbon utilization or even no carbon loss is crucial. Many studies have focused on this direction. In one work, researchers overexpressed the endogenous
fsaA,
kdsD, and
rpe genes from
E. coli and the
pkt gene from
Clostridium acetobutylicum in an
E. coli with the endogenous glycolaldehyde utilization gene
aldA knocked out [
46]. This artificial cycle can convert glycolaldehyde to acetyl-CoA without carbon loss. It firstly fixes glycolaldehyde with glyceraldehyde-3-phosphate to form arabitol-5-phosphate, which is then converted to xylulose-5-phosphate and back to glyceraldehyde-3-phosphate with the release of acetyl phosphate. The acetyl phosphate is then converted to acetyl-CoA by the endogenous
pta gene in
E. coli. However, the metabolic efficiency was very low for this cyclic pathway. Isotope tracing revealed that only 16% of the target metabolites, mevalonate and acetyl-CoA, originated from glycolaldehyde. This is far from sufficient for using glycolaldehyde, or even ethylene glycol, as a carbon source for microbial growth or production.
In another study, researchers proposed a novel approach for glycolaldehyde utilization [
47]. They aimed to identify an enzyme that could convert glycolaldehyde directly to acetyl phosphate in a single step, thereby significantly shortening the pathway from glycolaldehyde to acetyl-CoA. Although the natural enzyme that catalyzes this reaction was not reported, some enzymes, known as phosphoketolases (PKs), can produce acetyl phosphate from fructose-6-phosphate or xylulose-5-phosphate [
48]. Therefore, researchers focused on these enzymes and selected eight PKs for experimental verification following a phylogenetic tree construction using 111 bacterial species. An enzyme, BaXfspk, was identified as having high efficiency in converting glycolaldehyde to acetyl phosphate. By introducing this system into
E. coli, the ability of
E. coli to grow using glycolaldehyde was significantly enhanced, reaching a biomass yield of 0.681 ± 0.028 gCDW/g (g cell dry weight per g glycolaldehyde) [
47]. However, research on this topic is very limited, and others are still using the endogenous glycolaldehyde conversion pathways in
E. coli [
41,
45,
49,
50].
In summary, using ethylene glycol as a carbon source for microbial growth or production mainly involves integrating the oxidation of ethylene glycol and the utilization of glycolaldehyde. In previous studies, engineered
E. coli strains could grow or produce target products using ethylene glycol, with yields higher than those achieved using glucose as a carbon source [
50]. It is particularly notable that the endogenous glycolaldehyde utilization pathway in
E. coli has a carbon utilization efficiency of only 50%. This suggests that there is still a long way to go in optimizing the use of ethylene glycol as a carbon source for bioproduction, and it also holds great potential and significant development value. It is also worth noting that the pathways for ethylene glycol utilization share many similarities with those for ethanol utilization. This implies that future work could leverage bioinformatics tools to explore ethanol metabolic pathways, particularly by identifying more enzymes that could be used in the utilization of ethylene glycol.
3.2. Ethanol to Acetyl-CoA
Ethanol, as a common C2 metabolite, exists with a natural utilization pathway as a carbon source. Currently, many researchers are attempting to use ethanol as a carbon source for bioproduction, including fatty acid ethyl esters(biodiesel), squalene, and acetyl-CoA derived drugs [
51,
52,
53,
54]. The pathways they use can be roughly divided into two types: directly converting ethanol to acetyl-CoA, or first converting ethanol to acetic acid and then converting it to acetyl-CoA via the acetic acid utilization pathway. In this section, we mainly introduce the pathways for the direct conversion of ethanol to acetyl-CoA or the conversion to acetic acid, while the conversion of acetic acid to acetyl-CoA will be separately introduced in the next section.
In
E. coli, there is a natural pathway for converting ethanol to acetyl-CoA. This pathway is mediated by a bifunctional enzyme, AdhE, which can convert acetyl-CoA to ethanol via acetaldehyde in two steps [
55]. Although the reaction catalyzed by this enzyme is reversible, in the actual growth of
E. coli, due to various reasons, ethanol cannot often be converted to acetyl-CoA via this pathway, but it has been found that the AdhE mutant AdhE
A267T/E568K can enhance the ability of
E. coli to utilize ethanol under aerobic conditions [
56]. In a recent study, another ethanol utilization pathway in
E. coli has been discovered. In this pathway, ethanol is first converted to acetaldehyde by alcohol dehydrogenase AdhP, and then converted to acetyl-CoA by the mutant of acetaldehyde dehydrogenase MhpF [
57]. In another study comparing these two metabolic pathways, it was found that the pathway using Adh
EA267T/E568K not only significantly increased the growth rate of
E. coli in a medium with ethanol as the sole carbon source, but also resulted in a high yield of 3-hydroxypropionic acid (3-HP) [
58]. However, the production method in this study is whole-cell catalysis. Although this approach of separating bacterial growth from the production process can effectively improve the production efficiency of the product, the bacterial growth process still relies on traditional carbon sources, so there is room for further improvement in this study, such as increasing the effect of the ethanol utilization pathway and comprehensively regulating central metabolic network. In a subsequent work of this group, they used ethanol as a carbon source in the fermentation process. The yield of the target product isopropanol, increased to 67.45% of the maximum theoretical yield, which is higher than that using glucose as a carbon source [
59], possibly because ethanol induced the expression of endogenous genes related to product synthesis.
In addition to modifying the existing pathways in
E. coli to utilize ethanol, attempts have also been made to construct artificial ethanol utilization pathways. Researchers combined the aldehyde dehydrogenase from
Dickeya zeae (encoded by the
ada gene) and the alcohol dehydrogenase from
Saccharomyces cerevisiae (encoded by the
adh2 gene) to construct a metabolic pathway in
E. coli that converts ethanol to acetyl-CoA via acetaldehyde in two steps. Using this pathway, the researchers achieved the growth of
E. coli with ethanol as the sole carbon source and obtained the corresponding acetyl-CoA derived products [
60]. In the commonly used engineered strain
S. cerevisiae, the pathway for growth using ethanol as a carbon source is naturally present. Therefore, by artificially regulating the endogenous pathways and the expression of regulatory genes in yeast to increase the production of acetyl-CoA or reduce its consumption in non-target pathways, the ability of yeast to utilize ethanol for growth and production can be effectively enhanced at the same time [
53,
61,
62,
63].
In other microorganisms, ethanol is first converted to acetic acid and then enters the endogenous acetic acid utilization pathway to further transform to acetyl-CoA. In this process, alcohol dehydrogenase (ADH) and aldehyde dehydrogenase (ALDH) play key roles. These two enzymes are present in many species and often rely on NAD⁺ or NADP⁺ as proton acceptors [
64,
65,
66]. A novel oxidative pathway (ExaABC) derived from
Pseudomonas aeruginosa has been identified, which relies on a cytochrome to oxidize ethanol [
67]. However, converting ethanol to acetic acid before utilization significantly lengthens the metabolic pathway compared to direct conversion to acetyl-CoA. Therefore, such pathways are often not cost-effective and are rarely used in production studies. Of course, even for pathways that directly convert ethanol to acetyl-CoA, the most critical step is the oxidation of ethanol. The enzymes in these pathways can serve as alternative options, providing researchers with new ideas. It is important to note that these ethanol oxidation pathways may generate reducing power in the form of NADH or NADPH, or may not generate reducing power at all. While this difference may not significantly impact normal microbial growth, it becomes crucial when ethanol is used as the primary or sole carbon source for growth and production. Therefore, these different ethanol oxidation pathways and their enzymes can be selected by researchers to meet their specific needs for generating NADH, NADPH, or no reducing power, thereby better matching the production of various target compounds.
3.3. Acetic Acid to Acetyl-CoA
Acetic acid can be produced naturally by many microorganisms. There are also natural metabolic pathways for utilizing acetic acid as a carbon source, which is why research on acetic acid utilization pathways is more extensive and in-depth compared to the utilization of other C2 compounds.
In
E. coli, there are two main pathways for acetyl-CoA utilization: the ACS pathway and the ACKA-PTA pathway. The ACS pathway is catalyzed by a single enzyme, acetyl-CoA synthetase (Acs). Under the catalysis of Acs, acetic acid first binds with ATP to form acetyl-AMP, releasing one molecule of pyrophosphate. Subsequently, the AMP group is exchanged with the CoA group to produce acetyl-CoA [
68,
69]. In the ACKA-PTA pathway, two enzymes are involved: acetate kinase (AckA) and phosphotransacetylase (Pta). In this pathway, acetic acid is first catalyzed by AckA to form acetyl phosphate and ADP, and then acetyl phosphate is converted to acetyl-CoA by Pta. Both pathways require an intermediate product to convert acetic acid to acetyl-CoA, but there is a significant difference between them. In the ACS pathway, the formation of pyrophosphate during the conversion of acetic acid to acetyl-AMP makes the pathway irreversible, as pyrophosphate is rapidly removed in the cell. In contrast, the ACKA-PTA pathway lacks such a constraint and is reversible. Additionally, the affinity of Acs for acetic acid is higher than that of AckA (Km values of 0.2 mM for Acs and 7 mM for AckA [
70]), making Acs more suitable for acetic acid utilization. Overexpression of the ACS pathway has been shown to improve the growth of
E. coli in acetate-containing media, while overexpression of AckA and Pta can inhibit cell growth [
71]. This may be caused by the difference in whether the reaction is reversible.
Numerous studies have been conducted on the use of acetate as a carbon source for bioproduction. Since acetate is directly converted to acetyl-CoA upon assimilation, it has a natural advantage as a carbon source for the production of acetyl-CoA-derived products. For example, a strain of
E. coli was engineered to produce isopropanol from acetate, achieving a yield of 13.3 g/L in a bioreactor with a stable acetate concentration of 30 mM [
72]. In another study, 2.54 g/L of γ-aminobutyric acid was produced from 5.91 g/L of acetate [
73]. Both studies used the
E. coli W strain, which is known for its better acetate utilization compared to other common strains such as BL21 (DE3), W3110, and MG1655[
71]. The W strain was originally isolated from soil near Rutgers University in 1943 [
74] and has been found to have better tolerance under various stress conditions [
75,
76]. These studies suggest that the W strain may be a naturally optimized strain for acetate utilization, potentially eliminating the need for overexpression of acetate utilization pathways.
In addition to
E. coli, other microorganisms with high acetate tolerance and utilization capabilities have been identified, such as
Corynebacterium glutamicum [
77]. For instance, by introducing relevant pathway enzymes and regulating carbon flux-related genes,
C. glutamicum was able to produce itaconic acid from 20 g/L of acetate, achieving a yield of 116 mM, which is 35% of the maximum theoretical yield [
78]. This indicates that selecting naturally acetate-tolerant and -utilizing strains as hosts may achieve or even surpass the effects of artificially engineered common strains for acetate-based production.
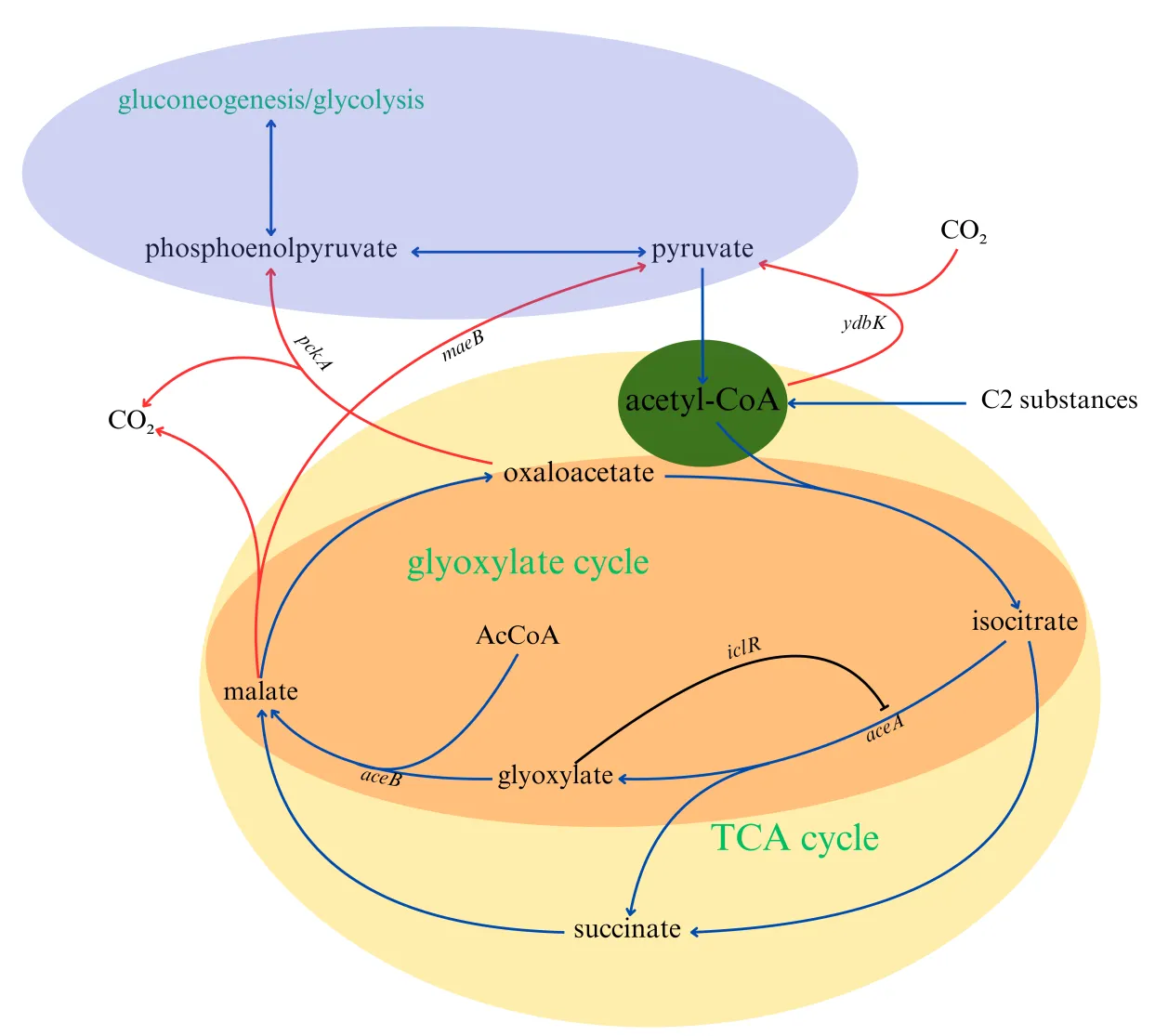
3.4. Acetyl-CoA back to Gluconeogenesis Pathway and Other Metabolic Regulation
C2 carbon sources can be more directly converted into acetyl-CoA, which is advantageous for bioproduction but also brings some challenges. Not all high-value compounds in biological production are derivatives of acetyl-CoA. For example, the production of various aromatic compounds in
E. coli requires the shikimate pathway, which begins with the condensation of phosphoenolpyruvate (PEP) and erythrose-4-phosphate (E4P). Clearly, PEP is an upstream metabolite of acetyl-CoA in glycolysis, while E4P is a product of the pentose phosphate pathway. Therefore, when producing such compounds, it is necessary to replenish acetyl-CoA into the gluconeogenesis pathway (as shown in
). This allows microorganisms to use C2 carbon sources to produce a wider variety of high-value products, rather than being limited to acetyl-CoA derivatives.
. The pathway for the incorporation of acetyl-CoA into gluconeogenesis. The blue arrows indicate the normal reactions in the pathway, the red arrows indicate the key reactions for the replenishment of acetyl-CoA, and the black arrows represent gene expression regulation.
It was discovered that overexpression of the
ydbK gene enhances the ability of
E. coli to utilize acetate for the production of non-acetyl-CoA derivatives [
79]. The
ydbK gene encodes pyruvate-ferredoxin oxidoreductase, which catalyzes the reversible reaction between pyruvate and acetyl-CoA with CO
2. Although overexpression of
ydbK effectively improved bacterial growth and product yield, subsequent research found that co-overexpression of
pckA and
maeB yielded even better results [
80]. The
pckA gene encodes phosphoenolpyruvate carboxykinase, which catalyzes the conversion of oxaloacetate to phosphoenolpyruvate. The
maeB gene encodes NADP⁺-dependent malate dehydrogenase, which catalyzes the conversion of malate to pyruvate. However, in this study, the
pckA and
maeB genes were overexpressed simultaneously, but neither was more effective than
ydbK. It is also worth noting that, unlike the other two genes, the pathway catalyzed by the enzyme encoded by
ydbK is involved in carbon fixation rather than carbon loss. Therefore, this pathway may potentially offer better performance. When ethylene glycol is used as a carbon source, if it is converted to glycolaldehyde and then assimilated via the endogenous pathway of
E. coli, it can directly enter the glycolysis/gluconeogenesis pathway in the form of 2-phosphoglycerate, thus eliminating the need for additional acetyl-CoA supplementation.
Among the other methods for acetyl-CoA replenishment, except for the overexpression of the
ydbK gene, the substrates used are derived from the TCA cycle. Withdrawing these substances from the TCA cycle may severely disrupt the normal function of the TCA cycle, thereby affecting bacterial growth. Therefore, the glyoxylate cycle becomes an attractive alternative. In the key reaction of the glyoxylate cycle, one molecule of isocitrate and one molecule of acetyl-CoA are converted into one molecule of malate and one molecule of succinate. Thus, without considering the conversion of substances outside the TCA cycle, the glyoxylate cycle can increase the number of substances within the TCA cycle. Therefore, appropriately regulating the expression levels of the glyoxylate cycle and the replenishment pathways can achieve the replenishment of acetyl-CoA into the gluconeogenesis pathway without compromising the stability of the TCA cycle. Some studies have also shown that using promoters with different strengths to regulate the intensity of these two pathways can effectively increase the yield of target products [
81,
82]. Additionally, the regulatory gene of the glyoxylate cycle, such as
iclR [
83], can be knocked out to enhance the carbon flux through the glyoxylate cycle. However, another study suggests that the knockout of the
iclR gene may reduce the carbon flux through the pentose phosphate pathway [
84], which could be disadvantageous for the shikimate pathway that relies on products from the pentose phosphate pathway. Therefore, directly regulating the expression of key enzyme genes in the glyoxylate cycle, such as
aceA [
85] or
icd, may be a better approach.
When ethylene glycol and ethanol are used as carbon sources, their conversion to acetyl-CoA requires oxidation reactions, which may lead to an increased demand for oxygen by the bacteria. To satisfy the O
2 demand, overexpression of oxygen-binding proteins such as vitreous hemoglobin (VHb) may be helpful [
86]. Additionally, it should be noted that these three C2 carbon sources can inhibit bacterial growth at high concentrations. In
E. coli, acetate at concentrations above 5 g/L can inhibit growth [
87], while ethanol shows an inhibitory effect just at a low concentration of 2% (
v/
v). There is less research on ethylene glycol, and the specific toxic concentration has not been clearly identified, but it may likely be similar to ethanol. These concentrations are too low for bioproduction. Therefore, to utilize these carbon sources for bioproduction, it is necessary to enhance the tolerance of the host strain to the carbon sources. To improve the tolerance of strains, many methods, such as adaptive laboratory evolution (ALE) or targeted tolerance-related modifications based on omics studies, have been developed [
88]. Given the limited understanding of global metabolism in biology, irrational adaptive evolution is a more commonly used and convenient method. The important aspect of adaptive evolution is the introduction of mutations at a desired degree. The atmospheric and room temperature plasma (ARTP) mutagenesis has been proved to be highly efficient for kinds of strains [
89]. Additionally, new methods such as targeted artificial DNA replisome (TADR) [
90], which introduce mutations in bacteria during growth, make adaptive evolution more efficient and sustainable.
To efficiently produce target products using C2 carbon sources, many other regulations are possibly involved, such as reducing the loss of key substances through side pathways [
91], regulating the ATP/ADP and NAD(P)
+/NAD(P)H ratios, and controlling specific metabolic pathways. These are more related to the specific target products and will not be elaborated on here.
Pathway engineering enables various host strains to take C2 compounds as carbon sources for growth and bioproduction.
exemplifies some high-value chemicals produced using those feedstocks.
The research on production using ethylene glycol has only just begun. Fewer studies have been reported on using it to produce more complex products. The most common product obtained from ethylene glycol at present is glycolic acid. The metabolic pathway from ethylene glycol to glycolic acid is relatively short and does not even go through acetyl-CoA. In one work, researchers used
E. coli to ferment ethylene glycol into glycolic acid in a bioreactor [
49]. After 112 h of continuous fed-batch fermentation, 10.4 g/L of glycolic acid was obtained. In a subsequent work, another group used
E. coli to produce 5.1 g/L of glycolic acid from a shake flask containing 10 g/L ethylene glycol over 96 h [
45]. Although this study appears to have a lower production efficiency, it should be noted that its production time is shorter. Moreover, under the conditions of production using shake flasks, the result of this study is more efficient than the work mentioned above. By introducing a plasmid expressing the ethylene glycol utilization pathway into a tyrosine-producing
E. coli MG1655 strain, 2 g/L of tyrosine was produced from a medium containing 10 g/L ethylene glycol [
50]. This provides a good solution for the future production of more complex products from ethylene glycol.
Compared with ethylene glycol, the research on using ethanol as a carbon source is relatively more extensive. The most common target product is the biofuel: fatty acid ethyl esters. Among the studies using ethanol to produce other high-value products, one introduced the ethanol utilization pathway and the 3-HP synthesis pathway into
E. coli, and 1.66 g/L of 3-HP was produced from a shake flask medium containing 10 g/L ethanol [
58]. Subsequently, by changing the production method from shake flask fermentation to whole-cell catalysis, the yield of 3-HP reached 13.17 g/L after two times of ethanol feeding. In another study, researchers engineered
E. coli with an introduced ethanol utilization pathway that used AdhE mutant as the pathway enzyme, achieving a polyhydroxybutyrate(PHB) yield of 3.12 g/L through fed-batch fermentation in a bioreactor over 54 h [
92]. Moreover, this study compared the efficiency of the strain in producing PHB using acetic acid, ethanol, and glucose as substrates, and ethanol achieved a PHB yield of 0.935 g/g, outperforming the other two substrates. This also highlights the advantages of ethanol as a carbon source.
Compared with the above two C2 compounds, acetic acid has a natural utilization pathway in
E. coli. Therefore, more research has been conducted on using acetic acid as a carbon source for production, and good results have been achieved. In one study, researchers used engineered
E. coli to produce threonine from acetate through a staged continuous fed-batch fermentation in a bioreactor, achieving a yield of 45.8 g/L after 61 h [
93]. In another work, researchers engineered a special strain
E. coli W to assimilate 38.7 g/L of acetate and produced 3.57 g/L of itaconic acid [
71]. Although the yield in this case was not high, it provided an idea: using a strain that naturally tolerates a specific carbon source for production with that carbon source. In addition to these relatively high-yield common products, more complex products can also be produced from acetate. Naringenin was produced from acetate with a relatively low yield of 122.12 mg/L for the first time [
94]. This work represents a valuable example of producing a complex compound from an unconventional carbon source.
.
Examples of production using C2 feedstocks.
| Host Strain |
Feedstock |
Product |
Cultivation Strategy |
Titers (g/L) |
Yield (mol/mol) |
References |
| E. coli MG1655 |
ethylene glycol |
glycolic acid |
flask |
5.1 |
0.612 |
[45] |
| E. coli MG1655 |
2,4-dihydroxybutyric acid |
flask |
0.8 |
0.15 |
[95] |
| E. coli MG1655 |
glycolic acid |
fed-batch |
10.4 |
0.652 |
[49] |
| E. coli MG1655 |
tyrosine |
flask |
2 |
/ |
[50] |
| E. coli MG1655 |
mevalonate |
flask |
1.6 |
/ |
[46] |
| Pichia naganishii AKU 4267 |
glycolic acid |
whole-cell catalysis |
35.3 |
0.26 |
[96] |
| Gluconabcter oxydans NL71 |
glycolic acid |
whole-cell catalysis |
110.5 |
0.944 |
[97] |
| E. coli W3110 |
ethanol |
PHB |
fed-batch |
3.12 |
/ |
[92] |
| E. coli MG1655 |
PHB |
flask |
1.1 |
/ |
[60] |
| E. coli BL21(DE3) |
prenol |
flask |
0.024 |
/ |
[60] |
| E. coli MG1655 |
3-hydroxypropionic acid |
flask |
1.66 |
0.123 |
[58] |
| E. coli MG1655 |
3-hydroxypropionic acid |
whole-cell catalysis |
13.17 |
0.291 |
[58] |
| E. coli MG1655 |
isopropanol |
flask |
4.41 |
0.337 |
[59] |
| Saccharomyces cerevisiae BY4741 |
itaconic acid |
flask |
0.142 |
/ |
[63] |
| Pseudomonas putida KT2440 |
PHB |
fed-batch |
9.3 |
0.049 |
[98] |
| Yarrowia lipolytica |
isocitric acid |
fed-batch |
90.5 |
0.185 |
[99] |
| E. coli W |
acetate |
itaconic acid |
fed-batch |
3.57 |
0.054 |
[71] |
| E. coli W |
isopropanol |
fed-batch |
13.3 |
0.196 |
[72] |
| E. coli W H-31 |
threonine |
fed-batch |
45.8 |
0.22 |
[93] |
| E. coli W3110 |
tyrosine |
flask |
0.7 |
0.032 |
[81] |
| E. coli W |
GABA |
flask |
2.54 |
0.342 |
[73] |
| E. coli BW25113 |
isobutanol |
flask |
0.157 |
/ |
[79] |
| E. coli BL21(DE3) |
naringenin |
flask |
0.122 |
/ |
[94] |
| Corynebacterium glutamicum |
itaconic acid |
flask |
5.01 |
0.116 |
[78] |
| Yarrowia lipolytica MTYL065 |
lipid |
semicontinuous |
115 |
/ |
[100] |
| Aspergilus oryzae |
malic acid |
batch |
8.62 |
0.125 |
[101] |
| Cupriavidus necator DSM 545 |
PHA |
fed-batch |
43 |
/ |
[102] |
D.-K.Y.: Writing—original draft. D.-Y.X.: Writing—editing. J.-F.L.: Writing—editing. P.X.: Writing—review & editing, Funding acquisition, Conceptualization. J.-Z.L.: Writing—review & editing, Supervision, Funding acquisition, Conceptualization.
Not applicable.
Not applicable.
This work was financially supported by National Key Research and Development Program of China (2024YFA0918000), Natural Science Foundation of Guangdong Province (305196996052), Guangdong S&T Program (2024B1111160007, 2024B1111130003), and Science and Technology Plan Project of Guangzhou (2024A04J3803).
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.