1. Introduction: The Goals of Human Behavioral Ecology
Human behavioral ecology (HBE) emerged as a coherent area of intellectual inquiry in the social sciences in the mid-1970s and early 1980s, primarily in the United States of America [
1,
2,
3,
4]. Its goal is to use principles of Darwinian natural selection to understand how people modify behaviors in response to variation in socio-ecological environments. The emphasis on explanations of variation sets HBE apart from related evolutionary social sciences, such as evolutionary psychology and sociobiology, which have historically been more interested in explaining behavioral universals. This emphasis has yielded important, contextually grounded findings that are both theoretically innovative [
5] and potentially significant in application [
6]. Methodologically, HBE aims for holism and nuance [
7], striving to draw generalizable inferences that are well situated in local socio-ecological contexts. In recent years, HBE has gained traction as a driving framework for understanding aspects of human behavioral and life history variation [
8], including exciting advances that marry a traditional focus on explanations of Darwinian function with proximate mechanisms that enable those functions to be carried out [
9,
10]. HBE’s historical emphasis on immersive fieldwork has similarly been augmented by more recent studies involving laboratory and experimental work, computational studies, and re-purposing of secondary data. The dominance of small-scale societies as sites for work has given way to the incorporation of a much broader range of societies [
11,
12], including industrialized ones, as practitioners of HBE (HBEers) have sought to understand the relevance of HBE in environments that bear limited resemblance to those of our evolutionary past [
5,
13]. In this article, we review the discipline of HBE and describe its applications in China, past, present, and future. We view its expansion into China with great enthusiasm, as an innovative means by which international partnerships and co-produced models of human variation can augment the expansive work on Chinese societies. This article is intended as a resource for novice HBEers; HBEers who wish to understand the current landscape of HBE as applied in China should skip to the relevant section (“Current Applications of HBE in Chinese contexts”).
Human behavioral ecology’s primary aim is to understand, using optimality frameworks, how natural selection has shaped people’s behavior [
8,
14]. A typical way for an HBEer to approach a question is to ask, given a person’s unique characteristics and constraints, how costs and benefits (
i.e., tradeoffs) of different behaviors are navigated in ways that maximize “
reproductive success” or “
fitness”—
i.e., genetic representation in future generations (terms in bold may be found in the Supplementary Glossary). HBEers anticipate that,
on average, people will adopt behaviors (“
strategies”) with the highest
net benefit,
i.e., the set of behaviors that maximizes reproductive success. For example, an HBEer might ask why women marry polygynously versus monogamously by generating models about how access to resources is affected by unions with a single versus an already-married partner [
15]. They could ask why babies are close together or far apart by modeling the effects of birth spacing on physical activity and testing them against real-world data [
16]. HBE invokes two critical assumptions in asking questions and testing hypotheses: first, reproductive competition is a fundamental driver of variation in behaviors and
phenotypes [
10,
17]. In other words, behavior that leads to more reproductive success in a given environment will become common over time. Second, HBEers expect humans to be flexible in responding to variation in environments. What maximizes reproductive success for one person may or may not maximize reproductive success for another. Costs and benefits for any given individual are also expected to differ across social and ecological contexts. The best decision for a male may not be the best for a female; the best decision in an environment of abundance may differ from one in an environment of scarcity. Moreover, people who make decisions are not presumed to be conscious in the attempt to maximize fitness; nor are they necessarily consciously weighing minute costs and benefits of any given set of alternatives. Rather, HBE begins with the premise that people generally make decisions that maximize fitness because it is what natural selection would have produced from prior rounds of competition.
To understand how natural selection may have resulted in contemporary behavioral variation, HBEers engage in a variety of studies that might be called typical for the discipline. Many initial studies surrounded the efficiency of foraging behavior and were motivated by quantitative optimal foraging theory [
18]. Such studies often involved an HBEer residing within a community whose foraging practices included a relatively heavy emphasis on hunting or fishing [
19]. The scholar would then painstakingly track what prey local hunters encountered, targeted, and acquired to see whether hunting preferences and returns were consistent with models and predictions based on, e.g., prey choice or patch choice. If hunters were routinely found to skip items modeled to be low ranking based on low caloric value or high processing costs, then hunting patterns would be deemed consistent with optimal foraging theory. If not, then initial hypotheses would be rejected, subsequently refined, and tested against existing or new data. Other HBE studies have built on optimality thinking to test ideas related to, e.g., whether daughter- or son-biased investments make sense under different social and ecological circumstances [
20], what sorts of individuals are elected (or insert themselves) into positions of leadership [
21], the outcomes of status and wealth inequality for health and well-being [
22], who is most likely to care for children under different circumstances [
23], the roles of older children and grandparents [
24], and so on. One of the primary means by which HBE has historically distinguished itself from allied disciplines is in its empirical emphasis on first-hand data and immersive fieldwork [
7,
13], though this is less dominant as HBE nears its fiftieth year of existence.
Human behavioral ecology is one of three closely related “styles” [
25] of investigating the evolution of variation in human (and sometimes non-human) behavior. Evolutionary psychology (EP) and cultural evolutionary theory (CET, aka dual inheritance theory, gene-culture coevolution) offer complementary, but distinct, frameworks for understanding the capacity for Darwinian natural selection to “select for” (
i.e., maintain) a given behavioral variant. The styles are easily conflated [
3,
26], but differ in crucial ways that affect both what each style attempts to explain and the assumptions that guide attempted explanations. Briefly [
25], as discussed above, HBE begins with the premise that contemporary behaviors are, on average, likely to be well suited to modern environments. Historically ignoring specific mechanisms that support behavioral adaptations and related constraints (
i.e., accepting the “
phenotypic gambit 1” [
28], HBE assumes that natural selection can act on general-purpose cognitive structures that are sufficient to allow for reasonably accurate weighing of costs and benefits of various behaviors across a range of potential environments. It emphasizes socio-ecological variation as a primary (but by no means sole) determinant of human behavioral variation.
Evolutionary psychology, by contrast, emphasizes longer-term, genetically-driven evolutionary processes that are more likely to lead to
mismatch between contemporary environments and “Stone Age minds” [
29]. In terms of mechanisms, EP is more likely to assume that more specific psychological modules have been selected for, each undergirding a specific behavioral variant. There is less flexibility envisaged by the EP perspective than by HBE, as EP typically views psychological mechanisms as genetically “hard-wired”. Compared to HBE, EP perspectives tend to be more “universalist”; socio-ecological variation is relatively downplayed in terms of causal significance, because psychological mechanisms are adapted to long-term and relatively stable
environments of evolutionary adaptedness [
30]. Finally,
cultural evolutionary theory (CET) emphasizes the potential for transmission of behaviors via cultural means (
i.e., social learning) to alter predicted patterns of behaviors, as well as their fit with socio-ecological environments. Previously dubbed dual-inheritance theory (DIT), cultural evolutionary theory essentially adapts models from population genetics based on vertical (
i.e., parent-to-child) transmission to incorporate non-genetic modes of transmission, including horizontal (peer-to-peer) and oblique (non-parental individual from parental generation to individual in a subsequent generation), as well as dynamic feedback between evolutionary processes and socio-environmental change [
31]. Relative to HBE and EP, CET is more circumspect about whether behaviors are likely to be adaptive—while general-purpose learning mechanisms are thought to be adaptive, on average, this can often lead to
maladaptive behaviors [
32,
33]. Although CET has become more empirical in recent years [
34,
35], it still draws more heavily on mathematical modeling and purely conceptual tests of hypotheses than either of its sister disciplines.
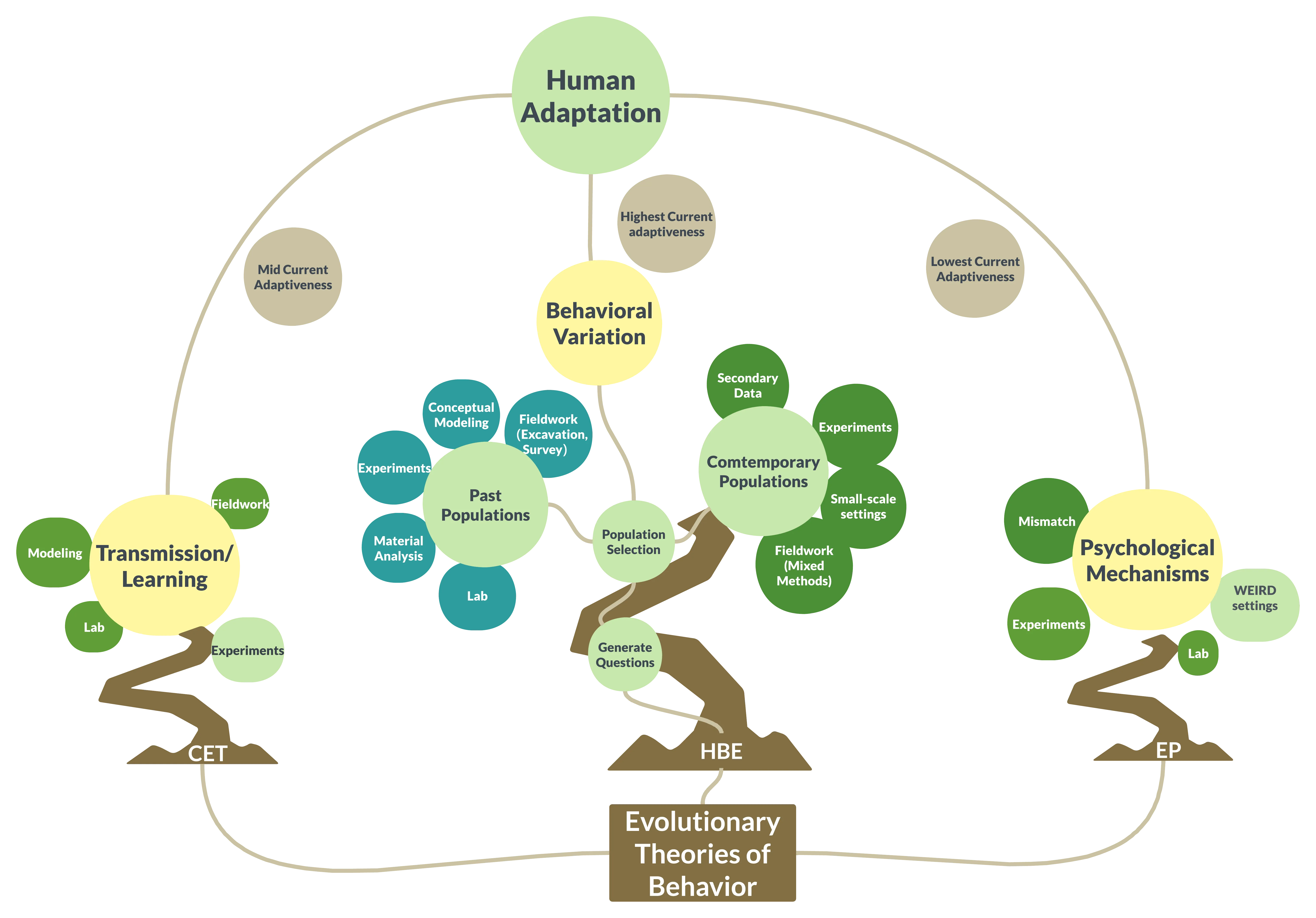
([
25], with permission) summarizes these differences.
.
Summary of the ‘three styles’ of studying human behavior. After Smith [25], with permission. Note that this describes average tendencies and historical emphases. Current trends in each discipline are variable and more convergent than this table implies.
|
Human Behavioral Ecology |
Evolutionary Psychology |
Cultural Evolutionary Theory |
| Explanatory outcome |
Behavior |
Psychological mechanisms |
Transmission of cultural information |
| Key constraints |
Ecological, phenotypical (e.g., gender, resources) |
Cognitive, genetic |
Information, socio-structural (e.g., presence/absence of cultural models) |
| Temporal scale of adaptive change |
Short-term (phenotypic) |
Long-term (genetic) |
Medium-term (cultural) |
| Expected current adaptiveness |
Highest |
Lowest |
Intermediate |
| Hypothesis generation |
Optimality and ESS models, ideally [36]; informal inference |
Informal inference |
Population modeling; dynamical modeling |
| Hypothesis testing |
Mixed methods; historical emphasis on quantitative ethnography |
Surveys, lab experiments |
Mathematical modeling & simulation |
| Pet topics: |
Subsistence, cooperation, reproduction |
Mating, parenting, sex differences |
Large-scale cooperation, population divergence, maladaptation |
We illustrate the comparison among these sister styles using a concrete, but hypothetical example, of how each might approach the question of modern food choice in
Box 1.
.
Three styles applied to understanding contemporary food choice.
How does someone decide what to eat? Whether to pursue kudu when hunting, a nut while foraging, or an apple in the grocery store, humans continually evaluate whether the benefits reaped from a particular food choice merit the costs of pursuit and processing. While there is significant overlap between the three styles of evolutionary behavioral analysis [37], each style is likely to emphasize different dilemmas or tradeoffs experienced when choosing what to eat. This box builds on existing scholarship to offer hypothetical approaches to understanding what drives food choice when starting within a given style.
HBE, as described in the main text, leverages optimal foraging theory to make predictions regarding food choice. Extant scholarship focuses on hunting and gathering scenarios [18,38,39], but there is no reason optimal foraging theory couldn’t be extended to food choice in urban or transitional subsistence settings and in consideration of common, if unfortunate problems associated with food deserts and lack of access to affordable, healthy foods. To assess optimal food choice, an HBEer would attempt to clarify all relevant costs and benefits, including costs associated with food search, acquisition, processing, and consumption and benefits associated with energy capture and nutrient density. In essence, the HBEer would attempt to describe the food choice landscape in terms of the return and encounter rates of a given food item and then model the decision of whether to handle the item or continue searching for something with higher profitability. In principle, the costs and benefits could be modeled separately based on gender or other demographic factors and tested against empirical data, though we are aware of few studies that engage these complications.
Food choice has received less attention in evolutionary psychology except where it intersects with other, more common foci, including mate choice and status enhancement. In classic evolutionary psychology, food choice decisions would likely be modeled from a universalist framework that anticipates individuals choosing foods based on evolutionary predispositions built upon millennia of adaptation to prevailing environments. If food choice, per se, is an uncommon topic in evolutionary psychology, the underlying logic of “mismatch” predominates in evolutionary medicine, which views much of contemporary metabolic dysfunction as a result of outdated psychological and biological mechanisms that lead people to prefer carbohydrate-dense, salty, and fatty foods. Another classic example motivated by universalist and mismatch framing is nausea and vomiting during pregnancy, which many socio-biologists believed was a mechanism for avoiding the intake of toxic, teratogenic substances during pregnancy, although there has been considerable debate over the adequacy of evidence supporting these views [40]. Similar arguments underlie ideas surrounding disgust, which is supposed to motivate avoidance of potentially harmful foods [41,42,43,44]. Finally, eating disorders are the target of ongoing evolutionary psychology and medicine investigations [45], with significant emphasis on the potential of status enhancing psychological mechanisms to go “haywire” and produce extremes of calorie restriction that would not have been common in ancestral environments.
Cultural Evolutionary Theory (CET) might address food choice by modeling transmission of food preferences [46], including vertically from parent to child [47], or through peer-to-peer (i.e., horizontal) or other forms of transmission. It might also incorporate specific biases affecting the transmission process, such as “prestige” biases, which would result in learners attending to models that have higher prestige, “content” biases that might predispose learners to weigh fatty or salty foods more heavily in their food choice algorithms, or “conformist” biases that result in learners choosing foods that are also chosen commonly by others in their community. The emphasis in CET does not exclude the adaptive (i.e., fitness) relevance of food choice, but focuses on learning mechanisms that are presumed to be adaptive in general, if not in any given instance of food choice.
A full view of food choice could incorporate aspects of all three styles to understand how variation in individual and socio-environmental circumstances impact food choice (HBE), whether and how such choices are motivated by psychological and biological mechanisms that were honed in evolutionarily relevant environments, and how, mechanistically, preferences and choices are transmitted from one individual to the next. Extending the food choice analogy to other behaviors and norms of interest, as reviewed in this article, reveals the considerable potential for integration and nuanced knowledge-generation that can arise through partnerships with Chinese scholars with backgrounds in relevant fields (biology, economics, decision-theory, anthropology, psychology, etc.).
2. The History of Human Behavioral Ecology and What Human Behavioral Ecology Is Not
HBE emerged as a formal discipline in the 1980s from a marriage of animal behavioral ecology and cultural anthropology. However, it has deeper intellectual roots going back to the 1950s and 1960s in cultural ecology and ethology [
10]. Cultural ecology was formalized by Julian Steward [
48] to describe how societies adapt to their environments. He posited that much cultural variation was due to differences in social and physical environments, which influenced, but did not determine, aspects of social organization and other human institutions. Indeed, cultural ecology was seen by some as a compromise between environmental determinists, who saw human behavior as dictated by their environments, and possibilists, who saw human behaviors as flexible, subject to cultural diffusion, and modulated by technology [
49]. Ethology, which can be traced back to Darwin, was popularized by Konrad Lorenz and Niko Tinbergen, who focused on instinctual behaviors in animals and the possible benefits they conveyed in terms of survival and reproduction [
50]. Ethology explicitly embraces a Darwinian perspective, positing that human-environment interactions are best studied by applying evolutionary theory to behavior in natural settings, aligning it with other tenets of HBE and filling a gap in cultural ecology [
51].
The contextualized adaptiveness of behavior and life history have long been foregrounded by HBE, distinguishing it from related social evolutionary sciences. Indeed, the idea that human behaviors differed across distinct groups in response to specific environments and regardless of genetic relatedness was first articulated in anthropology in an edited volume in 1979 [
52]. The volume simultaneously embraced relatively unnuanced sociobiological hypotheses and biological rather than anthropological frameworks, leaving room for even more contextualized approaches [
53]. Key players in the development of the nascent field of HBE include Irven DeVore, then at Harvard, Napoleon Chagnon and William Irons at Northwestern University, Richard Alexander at the University of Michigan, and Eric Charnov and Kristen Hawkes at the University of Utah. These pioneering HBEers were among the first to explore issues related to gender, status differentiation, and alloparental contributions to childcare in settings that differed markedly from the student populations favored by evolutionary psychology and sociobiology. From those points forward, HBE began to distinguish itself from closely adjacent frameworks of evolutionary psychology and cultural evolutionary theory [
54], with its emphasis on using neo-Darwinian principles of selection to understand human behavioral variation rather than universals (like evolutionary psychology).
Key models and theories that remain prominent today were being developed and tested by the 1970s. Kin selection theory [
55], posits that reproductive success can be gained both directly through one’s own reproduction and indirectly through the reproduction of those with whom an individual shares genes, focused primarily on food sharing and cooperative production.
Optimal foraging theory [
56] offered models to describe how predators searched for and processed food to yield the greatest net payoffs.
Parental investment theory [
57] modeled how parents allocate resources across different children, including assessments of the optimality of birth spacing [
58] and how the amount and timing of parental investments might vary according to social and environmental constraints.
Life history theory [
59] is premised on the
principle of allocation [
60], which stipulates that effort (time, energy, resources) allocated to one domain (for example, enhancing one’s own survival and somatic maintenance) cannot be allocated to another (for example, reproduction), and that these trade-offs are subject to
optimization [
61]. All such theories and models emphasize the means by which humans flexibly adjust to prevailing socioecological circumstances and individual constraints [
25].
As alluded to in the foregoing material, HBE has made important contributions to the evolutionary social sciences, but this hasn’t been without controversy and misconceptions of its aims and inferences. Good disciplinary hygiene demands that students and practitioners of HBE anticipate and militate against its misuse. As an evolutionary framework, HBE carries all the baggage that attends evolutionary frameworks more generally [
62,
63], including fallacious applications of scientific inferences to moral positions and actions (
i.e., the “
naturalistic fallacy”) [
64], which have led to (false) claims that HBEers are socially conservative [
65], and/or genetic or ecological determinists who fail to recognize the importance of human agency [
10,
66]. For example, HBE research has described the evolutionary underpinnings of variable marriage systems, from monogamy [
67,
68], to polygyny [
69], to polyandry [
70]. HBE practitioners have sometimes been called on to use these findings to weigh in on the appropriateness of legislation affecting whom one is allowed to marry [
71], presumably because lawmakers are interested in justifying legislation in light of what is “natural” and/or “healthy”. This is obviously risky business—HBE has helped to understand the significant variation in human marital practices across cultures [
72], and, to a lesser extent, relationships between marital practices and well-being [
73,
74]. These studies—and HBE more generally—emphasize the context-specificity of behavioral and health outcomes, challenging simplistic applications of HBE to regional or national policy and legislation. On the other hand, most HBEers, at least historically, have adhered to a philosophy of science that emphasizes objectivity and distance between oneself and one’s research. This has arguably led HBEers to be less comfortable than other social scientists to wade into important political and legislative debates, when HBE offers precisely the type of nuance that would help to make more informed decisions [
6]. We urge both new and veteran HBEers to deliberate carefully when disseminating information and any resultant viewpoints so that, regardless of how deeply one engages with the public aspect of their research [
75,
76], they do so in ways that acknowledge the potential for misrepresentation and misuse [
77].
HBE must also be deliberate in describing its relationships to other frameworks, both in terms of drawing connections to related areas (e.g.,
evolutionary demography [
78,
79], economic anthropology, comparative ethnology, sociology, biology, comparative phylogenetics, economics, and so on) and in clearly dispelling misunderstandings of HBE principles and assumptions [
80]. For example, one common misunderstanding results from the conflation of HBE with “unilineal evolutionism”.
Unilineal evolutionism is a framework that was globally common at the turn of the 20th century that envisioned evolution as a linear, hierarchical process, whereby a population evolved from a lesser state to a superior state, as evolutionary processes refined features (including behavioral) into ever more adaptive forms [
81,
82,
83]. In anthropology, it is unfortunately common for non-HBEers to conflate HBE with unilineal evolutionist frameworks [
84,
85,
86,
87] that categorize populations on a linear continuum from less to more evolved. However, HBE has little in common with unilineal evolutionary perspectives [
7,
82]; indeed, Darwinian frameworks view evolution as tree-like, rather than ladder-like; changes to traits and behaviors can move in any direction and do not necessarily proceed in a linear or ratchet-like fashion, even if
sometimes rachet-like evolution is apparent [
81]. HBEers accordingly do not categorize populations as more versus less evolved; rather, they seek to understand whether and how natural selection has led behaviors to adjust to prevailing socio-ecological circumstances. In sum, it behooves HBEers to recognize the potential for reflexive reactions to the loaded history associated with evolutionary frameworks and to point toward the ways through which HBE supplements and complements other ways of knowing to enhance fundamental knowledge and/ or improve welfare [
88,
89].
3. Traditional Methods of Human Behavioral Ecology
HBE’s focus on individual variation and context specificity is supported by a diverse set of methods, ranging from intensive ethnographic fieldwork with a single population, from analysis of secondary data, to, albeit less commonly, use of modeling and/ or experimental methods [
90]. Indeed, relative to many of the quantitative social sciences, HBE stands out for its emphasis on “methodological individualism”
2 [
14,
91]—
i.e., explicit methodological attention to behavioral decisions that are specific to an individual’s circumstances. Methods typically follow the use of the hypothetico-deductive method, which generates testable predictions using models that attempt to “capture the basic elements of a situation, sacrificing detail and nuance for clarity and generalizability” [
10]. HBEers rarely rely on a single method; rather, most research projects involve mixed methods, including both qualitative and quantitative tools. Qualitative methods are essential for identifying
emic perspectives on behaviors and socio-ecological contexts, interpreting quantitative data, and otherwise describing scenarios that are difficult to capture with quantitative data; quantitative methods provide robust tests of hypotheses that guide HBE research [
7]. We provide examples of the most common data collection and analysis tools, recognizing that the methodological world is extremely broad [
93] and evolving in ways that we cannot capture here.
Historically, HBE has and continues to draw most of its inferences from primary data painstakingly collected in non-WEIRD (
i.e., Western, Educated, Industrialized, Rich, and Democratic; see [
94] settings. Following early work by, e.g., Lee and DeVore with !Kung hunters and gatherers in Africa, [
95], HBE has now expanded its fieldwork to all six inhabited continents and both industrialized and non-industrialized contexts [
5]. A typical HBE study begins with a question and/or hypothesis or hypotheses that are sufficiently rooted within the local context. For example, Siobhán Cully’s (formerly Mattison) work with patrilineal and matrilineal Mosuo in China began with a question surrounding the evolutionary logic of daughter- versus son-preference in different socio-ecological contexts. Following Holden et al. [
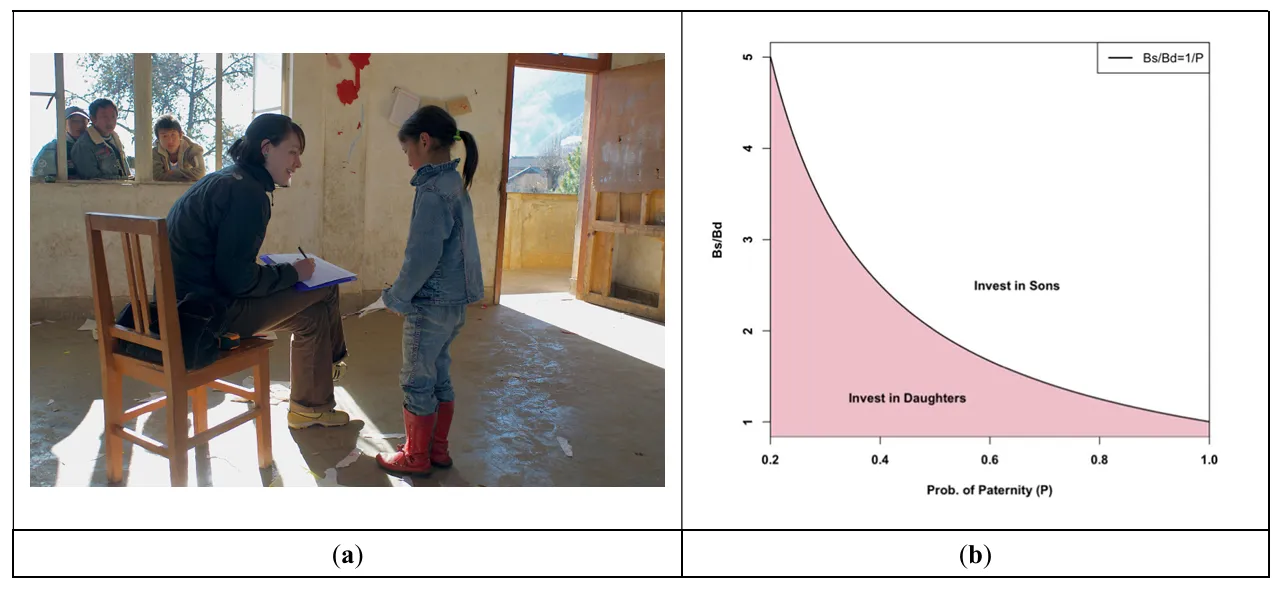
20], she hypothesized that resources that did not lead to disproportionate reproductive success through males would be transmitted matrilineally. She tested this with demographic surveys of over 200 households in both matrilineal and patrilineal communities, asking people to describe their resource base (e.g., land holdings, income, and other forms of wealth), in addition to other demographic variables and reproductive histories so that she could test whether women’s or men’s fertility had a greater relationship with wealth (
) [
82,
96]. Other common primary data collection tools include name generator questionnaires for social network analysis. For example, Ruizhe Liu’s work with matrilineal Mosuo people focuses on whether market integration leads to the dissolution of familial networks as kin are replaced with non-kin [
97]. She joins these data with socio-demographic data that describe market variables such as distance to market, education, majority language acquisition and so on, finding that, contrary to expectations, at intermediate levels of market integration, kin continue to dominate social networks [
97,
98]. Questionnaires, structured interviews, and semi-structured interviews often complement core instruments like socio-demographic surveys and reproductive histories. These are highly tailored to specific interests and might ask about educational outcomes (e.g., in relation to adoption status [
99]), the timing of initiation rituals [
100], or anything else that is of specific interest to researchers. Quantitative observational methods [
101,
102] can be useful for testing hypotheses (e.g., measuring parental investment in terms of breastfeeding bouts or duration [
103]) and for validating reported findings and otherwise self- or other-reported data.
Secondary data analysis has long been part of the methodological tool kit for HBE [
104,
105] and has become more common as secondary data sources have proliferated and analytical software and training have improved [
13,
106,
107]. Indeed, there are several clear benefits to secondary data analysis, including: secondary data create efficiencies in science in terms of data collection effort; they often provide large sample sizes that allow for more nuanced statistical analysis than is possible with smaller sample sizes; they are typically attended by thorough meta-data that facilitate interpretation of variables; they may have longitudinal designs that facilitate causal analysis not possible with cross-sectional data; similarly, they can cover much larger geographic areas than are possible in the typical HBE study design (and funding level). Secondary data are not always archived in places that make them easy to find; common sources for human behavioral ecologists include: parish registers, sometimes collected locally and opportunistically (with appropriate approvals!) in conjunction with primary data analysis; household registers; and large demographic and health data sets such as the Demographic and Health Survey [
108]. Such data sets have provided insights that are simply not possible with typical cross-sectional, small-scale research design associated with primary data collection, often challenging results that find evidence for adaptation in cross-section, but not longitudinally or with more sophisticated statistical controls [
105,
109]. There are also well-established pitfalls to the use of secondary data, however [
90]. The costs and benefits of any method or data set should be evaluated with reference to research objectives: for HBE, the ability to link individual data across a number of variables and with understanding of the local socio-ecological context is paramount.
. (<b>a</b>) Siobhán Cully (then Mattison) engaging in fieldwork for her dissertation, which focused on investments in daughters and sons among Mosuo people in Southwest China. (<b>b</b>) Visual depiction of Holden et al.’s [
20] model describing when it makes sense to invest in daughters versus sons based on paternity certainty (<i>x</i> axis) and the effect of a resource on sons’ (Bs) versus daughters’ (Bd) reproduction. Photo: PM Mattison.
Finally, among the more common methods used by HBEers, mathematical modeling using optimization or game theoretic frameworks has an important role to play in guiding data collection and can be used to inform empirical tests and conceptual refinements of HBE hypotheses. Optimization models are well developed in fields such as microeconomics, decision theory, and operations research and have long been widespread in evolutionary biology [
18]. In HBE, they follow directly from the principles of natural selection, which favors locally advantageous adaptations, leading to an increasingly better fit with environments over time [
10]. When human behavioral ecologists develop optimization models, they carefully weigh the potential costs and benefits of a set of behavioral alternatives and attempt to predict, given individual characteristics and local environmental constraints, which behavior is likely to yield the highest fitness under a given set of circumstances. Optimization models must describe a specific currency (typically a proxy for reproductive success), a goal (e.g., maximizing efficiency), and any constraints impacting alternative decisions [
18]. Perhaps the most developed tests using optimization models, as well as some of the earliest, fall under optimal foraging theory. Most optimal foraging models attempt to identify what foraging choices will lead to the greatest net rate of return in terms of energy or nutrients per unit of foraging time [
18]. Broad model classes include prey choice or diet breadth models, patch choice and time allocation models, and models that attempt to determine the optimal group size for foraging or hunting bouts [
18]. For example, Jeremy Koster used optimal foraging models to predict how the use of dogs would affect prey choice in Nicaraguan foraging populations, finding that dogs increase encounter and return rates for foragers [
19]. Cody Ross and Bruce Winterhalder expanded on optimal foraging models and accurately predicted that active foraging replaced sit-and-wait foraging as metabolic search costs decreased with the introduction of the horse to the Great Plains and the snowmobile to Arctic foraging communities [
110]. Numerous other optimization models have guided HBE research, including models predicting birth spacing based on the efficiencies associated with carrying children of different ages [
16], models predicting when to marry polygynously versus monogamously [
15], and many more [
8]. Indeed, optimization logic underlies all HBE hypothesis generation, whether or not the logic is modeled in a mathematically explicit way.
In the end, HBE is a theoretical framework that helps to generate questions, hypotheses, and predictions and to interpret findings in light of underlying principles that are either implicitly or explicitly linked to optimization thinking. Some methods are more common, some better suited to those objectives, but HBE does not prescribe or proscribe any specific method or methodology. Indeed, like many other field-based disciplines, HBE had to adjust to the global pandemic, taking field-based methods online and using related platforms (e.g., MTurk, video-conferencing) to collect data remotely. Contemporary HBE uses numerous methods that are borrowed from related fields, including pile sorts, vignette studies, economic games, bio-specimen collection, and other “lab in the field” methodologies. Key considerations in choosing methods are question- and context-specific and are likely to continue evolving as HBE engages more with other fields and a range of geographic contexts and temporal contexts.
4. Areas of Interest in HBE as Illustrated via the Example of Gendered Division of Labor
Human behavioral ecology is a useful framework for understanding variation in all areas of human behavior and life history. A glance at any textbook in evolutionary behavioral sciences provides a good sense of the breadth of the field. The most recent and authoritative text was just published in 2024: Jeremy Koster et al.’s edited volume: Human Behavioral Ecology [
8]. Until this book was published, Smith and Winterhalder’s Evolutionary Ecology and Human Behavior [
14] was the leading text for graduate students of behavioral ecology, while various other texts have been used for undergraduate classrooms [
3,
111,
112]. From these tables of contents emerge the following common areas of interest for behavioral ecologists: foraging, subsistence, cooperation, gendered division of labor, status, hierarchy, and inequality, reproduction, and parenting, and cooperative care. Allied fields of demography, cultural evolution, and evolutionary psychology are of enduring interest to HBEers. Intersections with traditional human biology—a field that is more applied and mechanistic in orientation [
113]—have increased in recent years as behavioral ecologists have sought to understand intersections between behavior and health [
89]. For the sake of brevity, we describe here a single issue—the gendered division of labor—as it intersects with numerous topical areas of interest to behavioral ecologists. Readers who wish to go more in-depth in a given area are referred to the aforementioned textbooks as good starting points for exploration.
An enduring debate in human behavioral ecology surrounds the extent to which men versus women have driven the unique suite of adaptations that distinguish humans from related species. The human “adaptive complex” refers to this collection of traits: humans are distinct from other primates in having long lifespans that include a post-reproductive period, lengthy development, large brains relative to body size, their use of tools, cooperative subsistence and childrearing of numerous multiple different-aged dependents within nuclear family units, central place foraging, and reliance on intergenerational transfers of knowledge and resources. What lies at the center of these traits? What is the key transition made by humans that links all of these exceptionalities together? One argument holds that men’s hunting [
114] and the provisioning of women [
115] are the key to this evolutionary puzzle [
115]. With the increase in meat and protein in human diets, the gut size decreased while brain size and complexity increased [
116]. Men’s hunting focused on the large, risky game, which they shared with others in camp, including offering the choicest parts to their female partners. Women who exchanged fidelity for access to meat, experienced shorter birth intervals and correspondingly higher fertility, out-reproducing women without access to this key dietary staple (
i.e., meat). The most prominent alternative to the male provisioning hypothesis—the grandmother hypothesis—posits that grandmothers have played a more central role in driving human evolution [
117]. According to this view, grandmothers produce the surpluses that enable higher fertility, slower life histories, and so on, by provisioning daughters and grandchildren with tubers, which are comparably reliable, calorie-dense, and far less risky as a source of nourishment. Further, because of their comparably long lives, grandmothers are argued to be in a stronger position to provide critical information on social and environmental factors that facilitate intergenerational local adaptations.
These two alternatives—the male provisioning hypothesis and the grandmother hypothesis—are presented fairly starkly in this overview. The data that have been brought to bear on them are far-ranging and have not resulted in consensus, especially surrounding the extent to which men versus women
initiated the transition toward the contemporary human adaptive complex [
118]. Broader views have demonstrated that, at least within contemporary environments, numerous caregivers emerge as important across a range of environments: mothers are the most consistent, followed by maternal grandmothers, but fathers and siblings often play meaningful roles [
119]. This has led to a more general, “cooperative breeding” model [
120] of “pooled [household] energy” [
121] that emphasizes sharing and cooperation as general strategies for household subsistence, with
who provides caring being more context- and household-specific [
122]. Regardless of where the data ultimately fall, the example of gendered division of labor illustrates intersections with numerous topical areas of focus in HBE: time allocation and social network data surrounding foraging and cooperation reveal how dietary needs are actually met [
102]; associations between help and reproduction provide critical tests of hypotheses surrounding the pace of childbearing (and its effects on child welfare) [
123]; gendered divisions of labor can also help to understand the roots of wealth inequalities and hierarchy [
124,
125]; the dynamics of marriage and parenting are clearly tied to provisioning and subsistence dynamics. Indeed, a major strength of HBE is its interest in integrating topics and methods to generate nuanced interpretations of societally important variation, from gender bias [
126,
127], to gendered contributions to households [
128], to inequality [
129] and health [
22], to climate change [
130] and many more.
5. Archaeological Applications
HBE provides flexible, expanding sets of models that can be tested with archaeological data [
131]. Indeed, archaeological applications of HBE emerged in the 1980s as HBE was gaining ground as a way of understanding forager behavior [
132,
133,
134]. Accordingly, optimal foraging theory has had some of the deepest impact on the field of archaeology, where it has been used to understand choices and changes in diet through time, as foragers have economized their choices in different climatic and ecological environments, with an emphasis on population-dependent increases in diet breadth and the intensified use of some food types [
135,
136]. Repeated studies have shown that the fundamental microeconomic principles underlying the diet breadth model are sound and that predictions about food choice are useful and sometimes counterintuitive [
133]. Optimal foraging models can thus offer important explanations for human and non-human animal population dynamics over a range of landscapes, with deep-time explanations complementing ethnographic work in ways that offer holistic insights concerning human-animal interfaces.
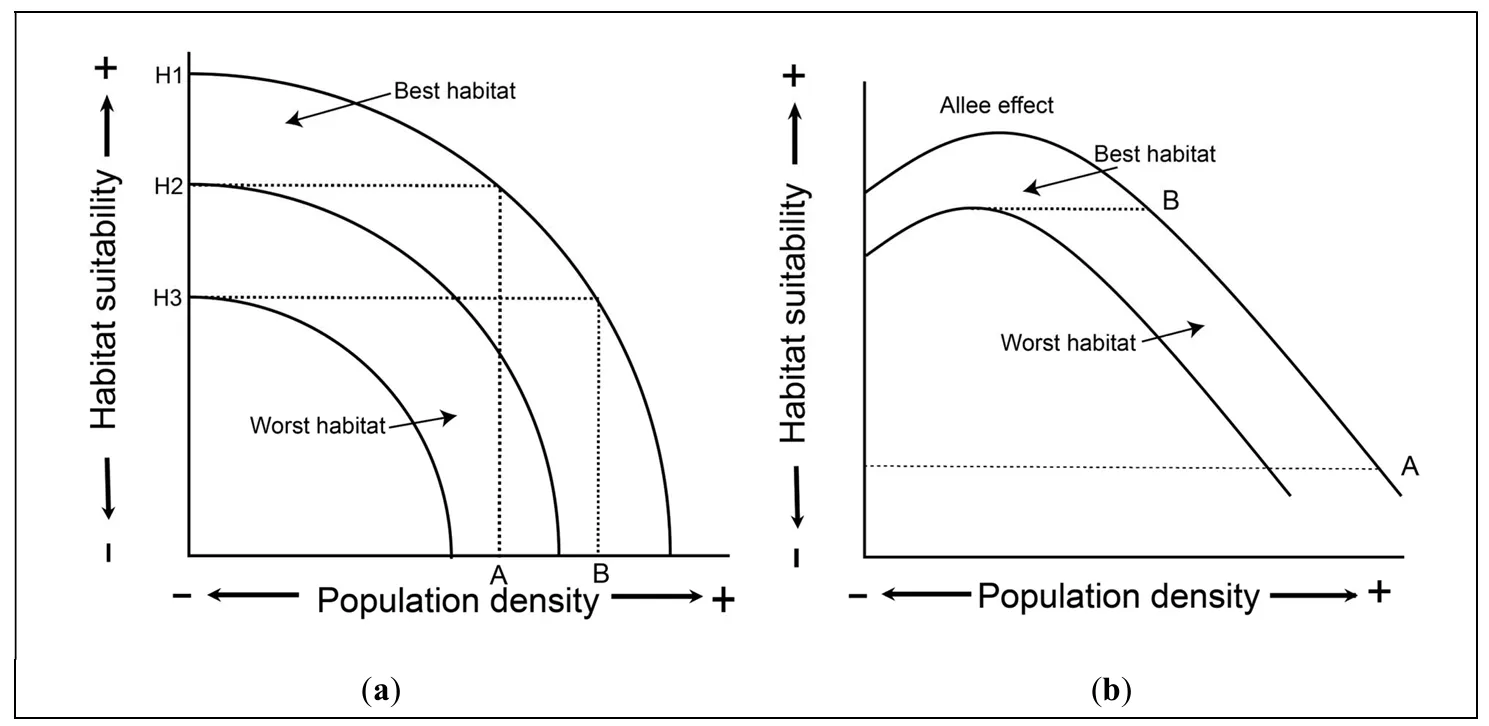
Clearly, resource abundance, including the presence or absence of prey, is an important consideration for how people are distributed across landscapes, a perennial interest in archaeology. Originally designed for bird populations in animal ecology [
137], the
ideal free distribution (IFD) provides a potential explanatory framework for predicting when individuals will move from a more to a less desirable resource patch. In essence, this framework sees patch quality as density-dependent, such that individuals should prefer to reside in higher-quality patches with relatively low population density [
131]. Because individuals are all considered competitors with equal ability and equal access to resources, the first individuals to settle in a habitat are predicted to locate themselves in the best available patch. As the population density of that patch increases due to in-migration or increased reproductive success of its members, the suitability of that patch drops. When patch quality declines to that of the next lowest-ranked patch, new immigrants or children of earlier habitants spill over into those patches. Many applications of the IFD in archaeology focus on early human migrations and specifically emphasize the colonization of islands, with their circumscribed areas and reliance on seafaring [
138]. They are relevant for a wide range of contexts, such as the Pleistocene rock art at Serranía de la Lindosa, Colombia (
), which has been linked to identity formation and territoriality of early colonizing foragers of the northern Amazon and may reflect patches with the highest ranking in food and/or other resources [
139]. IFD models assume that the effects of density on suitability are constant across habitats, and predict that the most suitable habitats will be occupied first and will maintain the highest population density [
134].
The relationship between population density and fitness in the IFD is not monotonic (
). Indeed, an interesting variant of the IFD that has been tested with archaeological data is the
Allee effect, a density-dependent phenomenon in which population growth or individual components of fitness increase as population density increases. At very low population densities, survival rates may be depressed because of difficulties in finding appropriate mates [
140,
141]. Increasing density can improve habitat suitability for subsequent arrivals. Subsistence systems may be affected by positive economies of scale, where scale is a consequence of density. Population increases can directly affect the density of desirable resources through some behaviors, such as maintaining forest cover in the early stages of succession, or through the application of technological innovations. For foragers and agriculturalists, habitat suitability of more marginal areas could be improved after initial colonization through the clearance of forest edges for hunting or, the establishment of fields for farming, or other types of intensification such as terraces or raised fields. Humans can also engage in collative actions to capture, use, and defend resources [
142].
Generally, these models predict there can be increased benefits leading to higher fitness for co-occupation by conspecifics up to some density threshold. This increase in suitability accompanying the addition of each individual to the habitat establishes an optimal group size [
143]. Once the density threshold is exceeded, the patch quality will still decline, which is consistent with the general predictions of the IFD.
. Pleistocene rock art in the Colombian Amazon. Las Dantas panel at Cerro Azul, Sierra de la Lindosa depicting a large mammal interpreted as a now extinct giant sloth. These images suggest early concepts of territoriality, patch selection and the formation of social identities around high-quality resources. Photo: KM Prufer.
. The Ideal Free Distribution (<b>a</b>) and Ideal Despotic Distribution (<b>b</b>). In the IFD (<b>a</b>), initial settlers populate the highest-ranked habitat. Continued growth (in situ or external immigration) increases density and population growth eventually spills over into lower-ranked habitats. With an Allee effect (<b>b</b>), the suitability of habitat B increases with each migrant drawn from habitat A, leaving the latter with lower density. If peak suitability/ density habitat B is higher than A, the Allee effect conceivably will empty habitat A for a period, as individuals seeking a higher quality habitat relocate to B. Taken to a logical conclusion, individuals might abandon areas once considered highly suitable during a period of change such as when agricultural subsistence strategies replaced those reliance on hunting and foraging [
138].
A more complicated variant of the IFD in archaeology is the
ideal despotic distribution (IDD) [
141], which models how differential access to resources alters how individuals are distributed over landscapes. The IFD applies to scenarios where there is interference among competitors of unequal abilities, or when superior competitors (dominants) can establish territories and defend them. This can protect dominants from density-dependent habitat deterioration as dominants more successfully defend better resource opportunities than inferior competitors, who are pushed to poorer habitats [
138]. A despot arises then, when one or more individuals in the population is able to monopolize a disproportionately large share of the factor or factors determining habitat suitability [
144]. Unconstrained and equal resource access in the IFD gives way to control and unequal access and defensibility in the IDD [
145]. The relevant suitability factors may be as diverse as the size of territory, resource and environmental amenities, access to reproductive partners and labor, or socio-economic advantages stemming, for instance, from contiguity with a desirable trade route or even a religious shrine [
146]. While differential control over such resources is a condition for social stratification and political control [
147], this is a narrower definition of despotism than that adopted by social scientists, including some archaeologists who more typically follow Wittfogel [
148] in implying to some degree an absolutist, managerial state. Flexible HBE models like the IDD provide nuanced approaches for understanding why individuals subject themselves to subordinate statuses, drawing on key HBE concepts of optimization and cost-benefit tradeoffs. Subordinates weigh the costs of subjecting themselves to the rule of others and the benefits of protection and group membership against the costs of migration. Dominants balance their interests in controlling and defending a disproportionate share of resources with the need to provide concessions to subordinates to prevent them from migrating. Along with other HBE models, there are relatively few limits on what kinds of variables one might accommodate in the IFD and its Allee and IDD variants [
134].
Models like the IFD and IDD have frequently been applied to foragers and early agriculturalists, but are increasingly being applied to other economic modes in the past, including large agricultural societies [
146], transhumant pastoralism [
149], and to model resource competition at high population densities [
150]. An important consideration of these studies has been the behavior of dominant despots in relation to subordinates. In the IDD model, subordinates will remain in an oppressive environment, one in which dominants claim part of subordinates’ production, if the dominants make concessions sufficient to outweigh the costs and uncertainties of emigration [
151]. Those concessions can include access to subsistence and other material resources, but can also be construed as the benefits and security of group membership. Examples might be access to market economies and goods and services, promises of collective security, redistribution of goods as a form of reciprocity, and access to public events to reinforce group identity. If elites can elevate local suitability—by providing amenities such as (a) solutions to coordination problems, (b) better defense, or (c) access to sacred sites and ritual opportunities—they may encourage in-migration. Even subordination may be advantageous if the alternative is worse.
For early agricultural societies, HBE models have been applied to understand population dynamics under varying conditions related to food production, which is a determinant of age-specific rates of birth and death. Modeling the relationships between food and space limitations on population dynamics in past societies by invoking mechanisms of hunger and well-being provides frameworks for understanding long-term processes of expansion and recession of human populations [
152]. The growth rate of a population determines the time until population is limited by space [
153]. This limitation introduces a tradeoff between population density and well-being, so population effects become more important than the direct effects on the environment. In natural fertility, agrarian societies modeling phases of population growth in newly settled habitats reveal distinct relationships between the phase of surplus food production and vital rates, transitions characterized by food shortages resulting in increased mortality and lessened fertility, and Malthusian intervals where vital rates and quality of life are depressed [
154]. The applications of these models to archaeological societies support the view that relatively low long-term population growth rates of prehistoric human populations, which are conditioned by carrying capacities, may have been a consequence in part of boom-bust cycles varying scales [
155].
6. Current Applications of HBE in Chinese Contexts
As described above, HBE’s historical emphasis on small-scale societies led to limited geographic representation in terms of study populations. China does not have any contemporary foraging communities and was not part of early HBE studies. Furthermore, HBE scholars are predominantly housed in English-speaking institutions, limiting the reach of HBE as a theoretical and conceptual area of scholarship for non-English-speaking countries. We are hopeful that China will be a focus of ongoing HBE partnership and research. China’s extensive history, vast territory, and diverse natural and climatic conditions yield a rich cultural tapestry that is ripe for understanding how “local ecologies nurture distinct cultures (一方水土养育一方文化)”. There is significant variation in Chinese populations in the realm of kinship, subsistence, cooperative norms, and so on, which we know very little about from an HBE perspective. Indeed, opportunities for comparative work that extend insights into Chinese contexts are numerous. Further, engaging in ethical partnerships with study communities is critical both in its own right and for the production of good science [
156]. There are many gains to be made from applying HBE to studies of Chinese populations, especially if this is done in close collaboration with Chinese scholars and local population experts. At present, the existing application of HBE in China is focused on two communities: matrilineal (and patrilineal) Mosuo and ethnic Tibetans. We review these studies and suggest future directions to expand into new regions and timescales of analysis.
Matrilineal Mosuo (aka Moso, Na, Naze) people—a population of around 50,000 residing on the border of Sichuan and Yunnan Provinces—are famous to anthropologists and social scientists due to a relatively uncommon form of reproductive union known in English as “walking marriage”, in Chinese as
zou hun (走婚), and in the local Naru language as
tisese, literally—walking back and forth. Traditionally, this form of union is open and non-binding: a man visits a woman in her bedroom at night and leaves (sometimes furtively) the next morning; children born of the union reside with their mother and belong to their mother’s lineage. Men and women remain members of their natal households, to which they contribute most of their labor. Men’s authority is over their sister’s children rather than over their own and inheritance is matrilineal [
157,
158,
159,
160,
161,
162,
163]. This relatively uncommon form of kinship has generated significant interest in and beyond anthropology. For HBEers, it renewed interest in classic questions surrounding why men would allocate time and energy to their sisters’ children instead of their own, when it might make sense for parents to favor daughters over sons, and how post-“marital” residence patterns may impact or reflect reproductive conflict.
Siobhán Cully (then Mattison) and her team of researchers were the first human behavioral ecologists to work in China. Beginning in 2006, and in collaboration with local and national Chinese institutions (Yunnan Academy of Social Sciences, Yunnan University, and Fudan University), Cully resided with Mosuo families in both matrilineal areas abutting Lugu Lake and patrilineal areas further afield [
82,
158,
164,
165], collecting data via interviews, socio-demographic questionnaires, social network instruments, anthropometric devices, and dried blood spots. Over numerous field seasons spanning almost 20 years, results have generally offered support for HBE hypotheses, though not in simplistic ways. For example, the team leveraged variation in market integration that has been ongoing since the 1990s [
161] to offer refined means for creating metrics of market integration that are tailored to local circumstances [
166]. They have also shown that matrilineal norms and social structures may buffer against rising material inequality [
125] and decrease hypertension and inflammation [
167] relative to Mosuo patrilineal counterparts. These gender “reversals” were also apparent in social support networks [
168], providing one possible means by which health inequities narrow. Earlier results offered partial support for hypotheses suggesting that subsistence is tied to gender-biased inheritance [
82], with men contributing to their own children when economic circumstances favored more intensive parental investment [
169,
170]. Results from other social sciences generally support these trends, revealing “gender reversals” in economic behavior, including Mosuo women being less generous than Mosuo men, in contrast to patrilineal Yi, who show the opposite pattern [
171]. The gender gap in risk aversion is also smaller among Mosuo than among Yi, consistent with Mosuo women taking on roles that are traditionally ascribed to men [
172]. Psychological studies further bolster the differences between Mosuo family structures and other family structures in China [
173,
174].
Mosuo kinship and social structure have elicited interest in broader patterns of cooperation and allocare. Consistent with the normative emphasis on sharing among kin, researchers have shown that Mosuo social networks reflect a strong emphasis on genetic relatedness as an underlying principle for allocating labor across households, even in the absence of reciprocity [
175]. Interestingly, while Mosuo societies frequently emphasize household harmony and generalized reciprocity, scholars have also demonstrated reproductive conflict among sisters co-residing in a household, where fitness decreases as the number of co-resident sisters increases [
157,
176]. Reproductive conflict also appears to incentivize men to perform more labor in their natal households, in order to avoid directing investments toward partners’ sisters’ children [
177,
178]. Post-marital residence obviously affects and reflects gender-biased cooperation and conflict, as shown in several studies among Mosuo and related Chinese populations. For example, duo-locality is posited to decrease incentives for large-scale cooperation due to increased reproductive conflict in the home, a pattern reflected in economic games played across a range of communities with differing levels of dispersal in China [
179]. Female workloads appear to be positively impacted by co-residence near natal kin, however: Although almost always higher than male workloads, female workloads are relatively lower in matri- and duo-local contexts compared to contexts in which women disperse from their natal homes and experience lesser bargaining power [
180]. Similarly, whereas the normative emphasis on caretaking and authority lies with the maternal uncle [
174,
181], others have emphasized variable roles for fathers in direct and indirect care [
169,
170,
174].
The only other population to have received significant attention from human behavioral ecologists in China to date are Tibetans. Tibetan populations also reveal a range of kinship structures, including norms and institutions that are variably centered around women and girls, providing an interesting test case for questions surrounding how subsistence systems impact gender-biases in dispersal [
182] and how gender-biases in dispersal impact broader cooperation patterns [
179]. Such studies provide support for hypotheses that link agriculture with female-biased dispersal. Gender-biased dispersal is also hypothesized to contribute to male celibacy in Tibetan monks [
183], a societal feature that is often assumed to be maladaptive, yet associated with enhanced fitness for relatives of celibate individuals in Tibet [
184]. Indeed, variation in gender-biased dispersal has been linked to numerous fitness-related demographic outcomes, including ages at first and last birth [
185] and marital stability [
186], as well as who in the household helps to raise children. For example, grandpaternal investment has been shown to be significant, if not of the same quality as grandmaternal help, among Tibetan populations where women are more economically active [
187], reinforcing the flexibility of human family systems, and the potential for numerous caretakers to have positive effects on children, depending on broader social and environmental circumstances [
23]. Finally, a recent article points to shifts in family structures from female-biased to more gender-neutral as markets continue to alter the landscape of family decision-making in Tibet [
188], reinforcing the dynamic, adaptive nature of human kinship across a range of lifeways.
Although Mosuo and Tibetan populations have attracted the most research attention in recent anthropology literature, they are certainly not the only Chinese populations that would be of interest to HBEers [
189,
190,
191]. Indeed, the remarkable variation in subsistence, gender-biased dispersal, and cultural norms and institutions across China has significant potential for yielding new insights regarding human behavioral evolution. Moreover, anthropology is not the only field where HBE theory has been applied in China; other disciplines, such as sociology, demography, and psychology, have used associated evolutionary theoretical frameworks to study contemporary human reproduction and fitness [
192,
193,
194,
195,
196,
197,
198], pointing to increasing potential not just for HBE applications in Chinese contexts, but for applications that are also increasingly convergent.
7. Chinese Archaeological Behavioral Ecology
HBE archaeology in China is in its early phases but is not a blank slate [
199,
200]. There are papers in English focused on introducing HBE and its relevant models, particularly as applied to hunter-gatherer populations [
199], including discussion of optimal foraging theory [
199] and costly signaling theory [
200,
201], e.g., to explain burial ornamentation [
201]. While not always explicit in leveraging HBE as an explanatory framework, these contributions appeal to principles of adaptation in response to specific ecological and environmental conditions [
202]. Notably, much of this HBE archaeology research focuses on the origin of agriculture [
203,
204,
205], but there are numerous other questions that continue to elicit interest and debate in archaeology that could be fruitfully applied to Chinese settings, including questions surrounding variation in material inequality, the roles of women versus men in production and childcare, how cultural traits spread and evolve, how environment shapes culture, and so on.
provides a preliminary list of current and future applications that we think would be especially productive in the Chinese context.
.
Areas of research for potential investigation in Chinese archaeology.
| Theoretical Area |
Keywords |
Key Themes from Existing Applications in China |
Possible Archaeological Applications |
| Natural selection |
behavioral variation, adaptation, reproduction, market integration, context-dependency |
tourism/markets in rural areas, cultural diversity, life history variation, menopause, reproduction |
architecture, household demography, traditional ecological knowledge (TEK), ancestor lineage, iconography, symbolism, climate adaptations |
| Sexual selection |
gender roles, gendered division of labor, mate choice, parental investment, marriage |
gender roles, gendered division of labor, gendered parental biases; marital stability; celibacy |
gendered symbolism, gendered division of labor, gendered signaling materials, social networks, paleo-demography; gendered morbidity and mortality |
| Cooperation |
kin selection, conflict, reciprocity, altruism, allocare, mutualism, group size, ideal free distribution, ideal despotic distribution, reproductive skew |
kinship structure, cooperation, reciprocity |
religion, exchange, inter-group relations; warfare; tribal conflict; social complexity |
| Cultural evolution |
learning, cultural transmission |
population migration, religion |
behavior transmission, social learning, social complexity, technology/ material evolution, symbolism, identity, ethnogenesis |
| Subsistence |
adaptation, survival, fitness, hunter-gatherers, forager, Neolithic Revolution, pastoralism, horticulture, agriculture, polyculture, production systems, complexity, stratification and hierarchy, inequality |
agriculture, paleolithic-neolithic transition, lithic technology, zooarchaeology |
subsistence transition; nutrition; bioarchaeology; paleodemography; transhumance; health; diet quality, population dynamics; climate change; ecological anthropology |
8. So How Do I Start an HBE Project in China (or Anywhere)?
As the reader can tell, HBE has significant potential for use in China, yet has only been minimally deployed in the Chinese context. We are optimistic, however, that these beginnings are evidence of a strong foundation for expanding HBE scholarship in China. To continue this work, research teams must craft new projects with care, ensuring that questions are appropriately framed and assumptions made explicit, ideally with clear framing of sets of alternatives [
206] that are rooted in optimality frameworks [
36], thereby avoiding common pitfalls that create a sense of “just-so” story telling [
207,
208]. Without a clear set of alternatives being specified, any relationship between a behavior and some proxy of fertility may be spurious and claims for natural selection potentially untenable [
36]. Researchers must also ensure that questions and assumptions are appropriate for the population or context within which they are being posed. In some cases, populations are selected prior to questions being developed. This is justifiable, not only because questions can be precisely tailored to contexts and population priorities, but also because it can also facilitate longitudinal follow-up and deeply contextualized data for researchers who remain in place for long periods of time (e.g., more than forty years for Nicholas Blurton-Jones and Hadza foragers of Africa [
209]). Objectivity, systematicity, and triangulation are paramount in HBE studies; where populations or data set selection precedes the development of questions and hypotheses, researchers are wise to take additional precautions to ensure that data collection and analysis produce unbiased results.
Once the research team has identified questions, hypotheses, assumptions, and what data (primary versus secondary) are appropriate to apply to them, methods must be carefully crafted to ensure that variables are clearly defined and operationalized, that data collection is defined in a way that is conducive to reproducibility, validity, and reliability, and that analysis is appropriate for the data and questions or hypotheses driving data collection. To identify the specifics of this procedure is beyond the scope of this review, but we recommend that readers peruse methods handbooks [
93] and attend to methods employed as described in primary scholarship that inform the development of the researchers’ projects. We strongly recommend pilot work ahead of formal study [
210], including mock-up analyses of any final data that are anticipated to result from data collection methods. Finally, an important part of any research project involves the ethical considerations that govern the collection of data, confidentiality, population relations, and so on. These considerations are not universal across contexts (see, e.g., Hudson et al., 2020) [
211]; researchers are advised to work in collaboration with local partnering institutions and populations to ensure that stakeholders at every level are comfortable with the research design and implementation and dissemination of results [
156].
9. The Past and Future of HBE
Like every discipline, human behavioral ecology has experienced a mix of both successes and failures. While, to our knowledge, HBE has never endorsed eugenicist or deterministic frameworks, it could undoubtedly do more to clarify its position against such movements [
76], as HBE is commonly conflated with frameworks such as unilineal evolutionism, sociobiology, and evolutionary psychology, which have had more problematic histories [
63,
80]. Further, HBE remains at heart a reductionist framework, which sits uncomfortably with some anthropologists [
66]; while we would argue that HBE is deeply more contextualized than many of its sister frameworks, it never hurts for HBEers to carefully describe the ways that it may complement and supplement other frameworks. Indeed, the incisiveness of HBE means that it has been able to cut through many debates that would arguably be difficult to resolve using descriptive or atheoretical perspectives [
5]. For example, HBE has helped to understand the timing and pace of demographic transitions across the world [
212], why women take on roles that are sometimes considered “male” [
23,
167,
168,
213], why subsistence regimes result in differential fertility [
123], why inequality has become so dominant across the globe [
129], and how humans leverage cooperation to manage risk within and across households and communities [
214,
215,
216], including within the increasingly important context of global climate change [
130,
217].
At the same time, several aspects of HBE have faced criticism. Some are uncomfortable with the overemphasis on the current adaptiveness of traits [
80,
218]. Other scholars place varying degrees of emphasis on drift-like processes as sources of neutral behavioral variation, for example [
219], while others posit that many contemporary behaviors are due to constraints on behavioral evolution and do not require solutions centered on adaptation [
220]. Evolutionary psychologists are more likely to posit that current behaviors are maladaptive, due to mismatch or adaptive lag and are more concerned about the long-term evolutionary value of traits. We view such debates as resolvable through empirical and analytical hypothesis testing [
206] and through careful logic. In particular, within the Tinbergian “levels of causation/ analysis” framework [
208,
221,
222], HBE primarily attempts to explain the current functions of behaviors in terms of their effects on fitness in contemporary environments. Perspectives that are concerned with the evolutionary origins of a behavioral trait or the dynamics of how the behavior changes over time are often not in competition with explanations focused on current function [
207,
223]. Similarly, explanations of behavior that focus on mechanisms (e.g., hormonal, psychological,
etc. underpinnings) or ontogeny (development over the life course) are not usefully pitted against explanations focused on current function. As discussed below, there is certainly room for HBE to become more integrative in its focus; we caution simply that students of evolutionary behavioral paradigms should always seek to evaluate the soundness of theoretical debates and any underlying assumptions of incompatibility before wading in to specific debates.
Additional criticisms surround the appropriateness of using a
piecemeal approach to investigate behavior [
80]. The piecemeal approach breaks down complex, related behaviors, to analyze the potential functions of each [
25]. For example, rather than analyzing marital systems, an HBEer might attempt to understand, in turn, factors associated with preferences for specific kinds of partners, gender-biases in adolescent dispersal, arranged versus love marriages, and so on. This reductionist approach is obviously not fully realistic: it ignores correlations between traits, including tradeoffs, and suffers from difficulties in establishing causality. For example, we know that horticultural systems are often associated with matrilineal kinship structures [
82,
224,
225], in part due to women’s ability to manage horticultural resources without significant input from men [
170]. One could argue that it is a matrilineal kinship that causes horticulture as much as the reverse. Modeling and longitudinal evidence stack up in favor of subsistence changes preceding changes in kinship structures [
226,
227], so these criticisms, too, are addressable with increasing evidence and care to think through potential inter-correlations of traits and causal pathways [
34].
As the foregoing examples illustrate, HBE has been highly successful as a theoretical framework that is focused on understanding the current functions of behaviors that have evolved in ways that have increased people’s fit with their current environments. As HBE continues to grow as a discipline and to engage with other areas, we anticipate greater integration and a less singular focus on Tinbergian “current functions” [
223]. Moving beyond the “phenotypic gambit” is likely to facilitate a better understanding of the extent (and limits) of human adaptability, shedding light on the nature of genetic, physiologic, and socio-cultural constraints affecting decision-making, as well as on feedback between decision-making and other aspects of human biology. For example, understanding how transmitted culture may or may not constrain adaptive decision-making is increasingly of interest to scholars who want to know when behavior is likely to be adaptive versus maladaptive [
228]. In another example, Wander and colleagues hypothesized differential mechanisms for erythropoeisis among high-altitude Mosuo people in China, which may underlie differences in the expression of related health outcomes [
229]. If so, this could have implications for patterns of labor, including by gender, dietary decisions, and demography more generally. Understanding such interlinkages is extremely exciting, especially where it creates knowledge that may positively impact policy and livelihoods [
230]. Certainly, greater integration would bypass some of the criticisms of an overly piecemeal approach to understanding human behavior [
80] and is reflected in recent movements to strive to better understand the mechanisms that underlie behavior (
i.e., that “mechanisms matter”) [
9].
HBE’s historical focus on small-scale and hunting and gathering communities has yielded to interest in a much broader range of societies, including ones that are urbanized and market integrated [
5,
13,
166]. This is a positive move for the field, as the extent to which HBE can explain how people make decisions under the influence of global and local influences remains unclear. As HBE continues to expand its influence and to incorporate other frameworks into its own, we urge that it prioritize equitable global engagements [
156], including attending to the needs and priorities of populations with whom HBEers engage. As part of this, HBEers could consider running field schools that train both local and non-local scholars and otherwise developing long-term collaborations with partners who benefit from the research being performed.
Finally, there is growing interest in some corners of HBE in the need to revisit critically language and frameworks that may impede rather than enhance understanding of underlying concepts. Terms such as “reproductive value”, for example, are arguably imprecise and, for some, likely to evoke personal rather than intellectual responses. Similarly, “fitness” is mathematically well described [
231], but poorly operationalized in contemporary studies [
232], and again laden with implied meanings that are rejected by HBEers, but otherwise commonplace. As HBE works to extend its reach to engage new audiences, including in China, we have opportunities to revisit the language and frameworks that guide our research. Specifically, as terms are translated and explained in new languages, we may gain critical insights that help to shape a more holistic, specific, and inclusive field.
10. Conclusions
In conclusion, human behavioral ecology, now nearly fifty years old, has provided the evolutionary behavioral sciences with an important, testable framework for understanding variation in behavior and life history in ways that are nonetheless highly context-specific. It has expanded its focus from small-scale communities to every type of community, both in the present and in the past. Yet, its incorporation into scholarship and training outside of English-speaking contexts remains limited. This is a priority for the field as the world becomes even more global and as populations rightly demand more benefits from the work in which they participate. For scholars who wish to learn about HBE, there is now a thorough and accessible book [
10] that lays out the state of the field. We very much hope that scholars of and in China will carry this work forward in integrative and innovative ways, recognizing the need to mitigate harm, including in disseminating findings, and to establish trusting relationships that maximize benefits to participant communities and relevant stakeholders, including the broader public.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/512, Glossary of select terms.
Acknowledgments
We thank Nature Anthropology for inviting this submission, as it prompted us to think more deeply about our engagements, past, present, and future, in China. Lee Cronk and members of the Human Behavioral Ecology Lab at Rutgers University provided helpful feedback on this manuscript. The National Science Foundation (BCS 2215559, 1920812, 1461514) supported much of the work that informed the development of this manuscript. Thank you to peer reviewers for the time, energy, and expertise applied to improving this manuscript and, foremost, to our Chinese colleagues, collaborators, and participants who have been so generous in their time and spirit.
Author Contributions
Conceptualization, S.M.C.; Resources, S.M.C.; Writing—Original Draft Preparation, S.M.C., R.L., K.P.; Writing—Review & Editing, S.M.C., R.L., K.P.; Visualization, S.M.C., R.L., K.P.; Supervision, S.M.C.; Funding Acquisition, S.M.C., R.L.
Ethics Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Funding
The National Science Foundation (BCS 2215559, 1920812, 1461514) supported much of the work that informed the development of this manuscript.
Declaration of Competing Interest
Siobhán Cully is on the editorial board of
Nature Anthropology. The authors declare that they have no other known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Footnotes
- 1.
-
The phenotypic gambit refers to the assumption that genetic, physiologic, and other biological mechanisms do not impede evolutionary trajectories [27]. Adopting the phenotypic gambit has allowed behavioral ecologists to learn much about ecological drivers of human behaviors, but understandings that result with this assumption in place are necessarily limited. Indeed, there is significant interest in understanding the ways by which cultural transmission may constrain the evolution of human behavior by impacting the ways humans learn to behave [9].
- 2.
-
Methodological individualism in its strongest form suggests that all social phenomena should be explained solely in terms of individuals and their interactions [91]; HBE also considers the importance of constraints enacted through groups (e.g., norms and institutions), though it has been relatively quiet about the feedback between the two [92].
References
-
1.
Cronk L. Human Behavioral Ecology.
Annu. Rev. Anthropol. 1991,
20, 25–53.
[Google Scholar]
-
2.
Cronk L, Chagnon NA, Irons W. Adaptation and Human Behavior: An Anthropological Perspective; Aldine de Gruyter: Hawthorne, NY, USA, 2000.
-
3.
Laland KN, Brown GR. Sense and Nonsense: Evolutionary Perspectives on Human Behaviour, 2nd ed.; Oxford University Press: Oxford, UK, 2011.
-
4.
Winterhalder B, Smith EA. Analyzing adaptive strategies: human behavioral ecology at twenty-five.
Evol. Anthropol. 2000,
9, 51–72.
[Google Scholar]
-
5.
Nettle D, Gibson MA, Lawson DW, Sear R. Human behavioral ecology: Current research and future prospects.
Behav. Ecol. 2013,
24, 1031–1040.
[Google Scholar]
-
6.
Gibson MA, Lawson DW. Applied Evolutionary Anthropology: Darwinian Approaches to Contemporary World Issues; Springer Science & Business Media: ew York, NY, USA, 2014; Volume 1.
-
7.
Shenk MK, Mattison SM. The rebirth of kinship: evolutionary and quantitative approaches in the revitalization of a dying field.
Hum. Nat. 2011,
22, 1–15.
[Google Scholar]
-
8.
Koster J, Scelza B, Shenk MK. Human Behavioral Ecology; Cambridge University Press: Cambridge, UK, 2024; Volume 92.
-
9.
Borgerhoff Mulder M. Human behavioral ecology—necessary but not sufficient for the evolutionary analysis of human behavior.
Behav. Ecol. 2013,
24, 1042–1043.
[Google Scholar]
-
10.
Scelza BA, Koster JM, Shenk MK. Human behavioral ecology. In Human Behavioral Ecology, 1st ed.; Cambridge University Press: Cambridge, UK, 2024.
-
11.
Koster J, Lukas D, Nolin D, Power E, Alvergne A, Mace R, et al. Kinship ties across the lifespan in human communities.
Philos. Trans. R. Soc. B Biol. Sci. 2019,
374, 20180069.
[Google Scholar]
-
12.
Ross CT, Hooper PL, Smith JE, Jaeggi AV, Smith EA, Gavrilets S, et al. Reproductive inequality in humans and other mammals.
Proc. Natl. Acad. Sci. USA 2023,
120, e2220124120.
[Google Scholar]
-
13.
Mattison SM, Sear R. Modernizing evolutionary anthropology.
Hum. Nat. 2016,
27, 1–16.
[Google Scholar]
-
14.
Smith EA, Winterhalder B. Evolutionary Ecology and Human Behavior; Aldine de Gruyter: York, NY, USA, 1992.
-
15.
Borgerhoff Mulder M. Reproductive decisions. In Evolutionary Ecology and Human Behavior; Smith EA, Winterhalder B, Eds.; Aldine de Gruyter: New York, NY, USA, 1992; pp. 339–374.
-
16.
Blurton Jones N. Bushman birth spacing: A test for optimal interbirth intervals.
Ethol. Sociobiol. 1986,
7, 91–105.
[Google Scholar]
-
17.
Williams GC. Adaptation and Natural Selection: A Critique of Some Current Evolutionary Thought; Princeton University Press: Princeton, NJ, USA, 2008.
-
18.
Smith EA, Bettinger RL, Bishop CA, Blundell V, Cashdan E, Casimir MJ, et al. Anthropological Applications of Optimal Foraging Theory: A Critical Review [and Comments and Reply].
Curr. Anthropol. 1983,
24, 625–651.
[Google Scholar]
-
19.
Koster JM. Hunting with Dogs in Nicaragua: An Optimal Foraging Approach.
Curr. Anthropol. 2008,
49, 935–944.
[Google Scholar]
-
20.
Holden CJ, Sear R, Mace R. Matriliny as daughter-biased investment.
Evol. Hum. Behav. 2003,
24, 99–112.
[Google Scholar]
-
21.
Seabright E. An Empirical and Theoretical Analysis of Leadership in Two Egalitarian Horticultural Societies. Doctoral Dissertation, The University of New Mexico, Albuquerque, NM, USA, 2022.
-
22.
Jaeggi AV, Blackwell AD, von Rueden C, Trumble BC, Stieglitz J, Garcia AR, et al. Do wealth and inequality associate with health in a small-scale subsistence society?
eLife 2021,
10, e59437.
[Google Scholar]
-
23.
Starkweather KE, Shenk MK, McElreath R. Biological constraints and socioecological influences on women’s pursuit of risk and the sexual division of labor.
Evol. Hum. Sci. 2020,
2, e59.
[Google Scholar]
-
24.
Kramer KL. Children’s help and the pace of reproduction: cooperative breeding in humans.
Evol. Anthropol. 2005,
14, 224–237.
[Google Scholar]
-
25.
Smith EA. Three styles in the evolutionary study of human behavior. In Human Behavior and Adaptation: An Anthropological Perspective; Cronk L, Chagnon N, Irons W, Eds.; Aldine de Gruyter: Hawthorne, NY, USA, 2000; pp. 27–46.
-
26.
Mace R. Human behavioral ecology and its evil twin.
Behav. Ecol. 2014,
25, 443–449.
[Google Scholar]
-
27.
Rittschof CC, Robinson GE. Genomics: Moving behavioural ecology beyond the phenotypic gambit.
Anim. Behav. 2014,
92, 263–270.
[Google Scholar]
-
28.
Grafen A. Natural selection, kin selection and group selection. In Behavioural Ecology: An Evolutionary Approach; Wiley Blackwell: Hoboken, NJ, USA, 1986.
-
29.
Cosmides L, Tooby J. Evolutionary Psychology: A Primer; Center for Evolutionary Psychology: Santa Barbara, CA, USA, 1997.
-
30.
Irons W. Adaptively relevant environments versus the environment of evolutionary adaptedness.
Evol. Anthropol. Issues News Rev. 1998,
6, 194–204.
[Google Scholar]
-
31.
Ready E, Price MH. Human behavioral ecology and niche construction.
Evol. Anthropol. Issues News Rev. 2021,
30, 71–83.
[Google Scholar]
-
32.
Boyd R, Richerson PJ. Culture and the Evolutionary Process; University of Chicago Press: Chicago, IL, USA, 1985.
-
33.
Boyd R, Richerson PJ. Why does culture increase human adaptability?
Ethol. Sociobiol. 1995,
16, 125–143.
[Google Scholar]
-
34.
Deffner D, Fedorova N, Andrews J, McElreath R. Bridging theory and data: A computational workflow for cultural evolution.
Proc. Natl. Acad. Sci. USA 2024,
121, e2322887121.
[Google Scholar]
-
35.
Mesoudi A. Cultural Evolution: A Review of Theory, Findings and Controversies.
Evol. Biol. 2016,
43, 481–497.
[Google Scholar]
-
36.
McElreath R, Koster J. 17 The End of Human Behavioral Ecology.
Hum. Behav. Ecol. 2024,
92, 402.
[Google Scholar]
-
37.
Barrett HC. Evolutionary Psychology. In Human Behavioral Ecology; Koster J, Scelza B, Shenk MK, Eds.; Cambridge University Press: Cambridge, UK, 2024; pp. 380–401.
-
38.
Hawkes K, O’Connell J. On Optimal Foraging Models and Subsistence Transitions.
Curr. Anthropol. 1992,
33, 63–66.
[Google Scholar]
-
39.
Pyke GH, Pulliam HR, Charnov EL. Optimal Foraging: A Selective Review of Theory and Tests.
Q. Rev. Biol. 1977,
52, 137–154.
[Google Scholar]
-
40.
Emmott EH. Re-examining the adaptive function of nausea and vomiting in pregnancy.
Evol. Med. Public Health 2024,
12, 97–104.
[Google Scholar]
-
41.
Curtis V, Aunger R, Rabie T. Evidence that disgust evolved to protect from risk of disease.
Proc. R. Soc. Lond. B 2004,
271 (suppl_4), S131–S133.
[Google Scholar]
-
42.
Matchett G, Davey GC. A test of a disease-avoidance model of animal phobias.
Behav. Res. Ther. 1991,
29, 91–94.
[Google Scholar]
-
43.
Oaten M, Stevenson RJ, Case TI. Disgust as a disease-avoidance mechanism.
Psychol. Bull. 2009,
135, 303.
[Google Scholar]
-
44.
Tybur JM, Lieberman D, Kurzban R, DeScioli P. Disgust: evolved function and structure.
Psychol. Rev. 2013,
120, 65.
[Google Scholar]
-
45.
Faer LM, Hendriks A, Abed RT, Figueredo AJ. The evolutionary psychology of eating disorders: Female competition for mates or for status?
Psychol. Psychother. Theory Res. Pract. 2005,
78, 397–417.
[Google Scholar]
-
46.
Aunger R. Are Food Avoidances Maladaptive in the Ituri Forest of Zaire?
J. Anthropol. Res. 1994,
50, 277–310.
[Google Scholar]
-
47.
Henrich J, Henrich N. The evolution of cultural adaptations: Fijian food taboos protect against dangerous marine toxins.
Proc. R. Soc. B 2010,
277, 3715–3724.
[Google Scholar]
-
48.
Steward JH. Theory of Culture Change: The Methodology of Multilinear Evolution; University of Illinois Press: Champaign, IL, USA, 1955.
-
49.
Kroeber AL. Stimulus Diffusion.
Am. Anthropol. 1940,
42, 1–20.
[Google Scholar]
-
50.
Lorenz K. The Foundations of Ethology; Springer: Vienna, Austria, 1981.
-
51.
Scelza BA, Koster J. Behavioral ecology, human. In The International Encyclopedia of Anthropology; John Wiley & Sons, Ltd: Hoboken, NJ, USA, 2018; pp. 1–11.
-
52.
Chagnon NA, Irons W. Evolutionary Biology and Human Social Behavior: An Anthropological Perspective; Duxbury Press: North Scituate, MA, USA, 1979.
-
53.
Richerson PJ, Boyd R. Evolutionary Biology and Human Social Behavior: An Anthropological Perspective.
Hum. Ecol. 1980,
8, 68–72.
[Google Scholar]
-
54.
Creanza N, Kolodny O, Feldman MW. Cultural evolutionary theory: how culture evolves and why it matters.
Proc. Natl. Acad. Sci. USA 2017,
114, 7782–7789.
[Google Scholar]
-
55.
Hamilton WD. The genetical evolution of social behaviour, I. and II.
J. Theor. Biol. 1964,
7, 1–52.
[Google Scholar]
-
56.
MacArthur RH, Pianka ER. On Optimal Use of a Patchy Environment.
Am. Nat. 1966,
100, 603–609.
[Google Scholar]
-
57.
Trivers R. Parental investment and sexual selection. In Sexual Selection and the Descent of Man; Campbell B, Ed.; Aldine: Chicago, IL, USA, 1972; pp. 136–179.
-
58.
Blurton Jones NG, da Costa E. A suggested adaptive value of toddler night waking: Delaying the birth of the next sibling.
Ethol. Sociobiol. 1987,
8, 135–142.
[Google Scholar]
-
59.
Stearns SC. The Evolution of Life Histories; Oxford University Press: Oxford, UK, 1992.
-
60.
Hill K, Hurtado AM. Ache Life History: The Ecology and Demography of a Foraging People; Aldine de Gruyter: Hawthorne, NY, USA, 1996.
-
61.
Smith EA, Winterhalder B. Encyclopedia of Cognitive Science; University of California: Berkeley, CA, USA, 2006.
-
62.
Joseph S. Anthropological evolutionary ecology: A critique.
J. Ecol. Anthropol. 2000,
4, 6–30.
[Google Scholar]
-
63.
Smith EA, Mulder MB, Hill K. Controversies in the evolutionary social sciences: a guide for the perplexed.
Trends Ecol. Evol. 2001,
16, 128–135.
[Google Scholar]
-
64.
Smuts B. Feminism, the Naturalistic Fallacy, and Evolutionary Biology.
Politics Life Sci. 1992,
11, 174–176.
[Google Scholar]
-
65.
Lyle H, Smith E. How Conservative Are Evolutionary Anthropologists?
Hum. Nat. 2012,
23, 306–322.
[Google Scholar]
-
66.
Smith EA. Agency and adaptation: New directions in evolutionary anthropology.
Annu. Rev. Anthropol. 2013,
42, 103–120.
[Google Scholar]
-
67.
Fortunato L, Archetti M. Evolution of monogamous marriage by maximization of inclusive fitness.
J. Evol. Biol. 2009,
23, 149–156.
[Google Scholar]
-
68.
Henrich J, Boyd R, Richerson PJ. The puzzle of monogamous marriage.
Philos. Trans. R. Soc. B Biol. Sci. 2012,
367, 657–669.
[Google Scholar]
-
69.
Scelza BA. Perceptions of Polygyny: The Effects of Offspring and Other Kin on Co-Wife Satisfaction.
Biodemogr. Soc. Biol. 2015,
61, 98–110.
[Google Scholar]
-
70.
Starkweather KE, Hames R. A survey of non-classical polyandry.
Hum. Nat. 2012,
23, 149–172.
[Google Scholar]
-
71.
Luscombe B. I do, I do, I do, I do. Time Magazine, 6 August 2012. Available online: https://time.com/archive/6642208/i-do-i-do-i-do-i-do/ (accessed on 1 January 2024).
-
72.
Fortunato L. Evolution of Marriage Systems. In International Encyclopedia of the Social & Behavioral Sciences, 2nd ed.; Wright JD, Ed.; Elsevier: Oxford, UK, 2015; pp. 611–619.
-
73.
Lawson DW, James S, Ngadaya E, Ngowi B, Mfinanga SGM, Mulder MB. No evidence that polygynous marriage is a harmful cultural practice in northern Tanzania.
Proc. Natl. Acad. Sci. USA 2015,
112, 13827–13832.
[Google Scholar]
-
74.
Ross CT, Borgerhoff Mulder M, Oh S-Y, Bowles S, Beheim B, Bunce J, et al. Greater wealth inequality, less polygyny: rethinking the polygyny threshold model.
J. R. Soc. Interface 2018,
15, 20180035.
[Google Scholar]
-
75.
Fuentes A. More than Ethics: Changing Approaches to Research in Human Biology. In Human Biology; Wayne State University Press: Detroit, MI, USA, 2022.
-
76.
Fuentes A. We need better and more PopSci by scientists.
Science 2024,
385, eadq8026.
[Google Scholar]
-
77.
Sear R. Demography and the rise, apparent fall, and resurgence of eugenics.
Popul. Stud. 2021,
75 (Suppl. 1), 201–220.
[Google Scholar]
-
78.
Mattison Siobhán M, Shenk Mary K. Ecological Evolutionary Demography: Understanding Variation in Demographic Behavior. In Human Evolutionary Demography; Open Book Publishers: Cambridge, UK, 2023.
-
79.
Sear R, Mattison Siobhán M, Shenk Mary K. Demography. In Human Behavioral Ecology, 1st ed.; Koster J, Scelza BA, Shenk Mary K, Eds.; Cambridge University Press: Cambridge, UK, 2024; pp. 307–332.
-
80.
Brown GR, Lala KN. Sense and Nonsense: Evolutionary Perspectives on Human Behaviour, 3rd ed.; Oxford University Press: Oxford, UK, 2024.
-
81.
Currie TE, Mace R. Mode and tempo in the evolution of socio-political organization: Reconciling ‘Darwinian’ and ‘Spencerian’ evolutionary approaches in anthropology.
Philos. Trans. R. Soc. B Biol. Sci. 2011,
366, 1108–1117.
[Google Scholar]
-
82.
Mattison SM. Evolutionary contributions to solving the “matrilineal puzzle”: A test of Holden, Sear, and Mace’s model.
Hum. Nat. 2011,
22, 64–88.
[Google Scholar]
-
83.
Mesoudi A, Whiten A, Laland KN. Towards a unified science of cultural evolution.
Behav. Brain Sci. 2006,
29, 329–347.
[Google Scholar]
-
84.
Bachofen JJ. Das Mutterrecht: Eine Untersuchung Über Die Gynaikokratie Der Alten Welt Nach Ihrer Religiösen Und Rechtlichen Natur; Unveränderte Aufl. Basel: B. Schwabe: Basel, Switzerland, 1897.
-
85.
Engels F. The Origin of the Family, Private Property, and the State; International Publishers: New York, NY, USA, 1972.
-
86.
Maine HS. Ancient Law; J.M. Dent: London, UK, 1960.
-
87.
Morgan LH. Ancient Society; Belknap Press of Harvard University Press: Cambridge, UK, 1964.
-
88.
Lawson D, Uggla C. Family structure and health in the developing world: What can evolutionary anthropology contribute to population health science? In Applied Evolutionary Anthropology: Darwinian Approaches to Contemporary World Issues; Springer: New York, NY, USA, 2014; pp. 85–118.
-
89.
Wells JCK, Nesse RM, Sear R, Johnstone RA, Stearns SC. Evolutionary public health: introducing the concept.
Lancet 2017,
390, 500–509.
[Google Scholar]
-
90.
Mattison SM, Shenk MK. 6. Ecological Evolutionary Demography: Understanding Variation in Demographic Behaviour; 2024.
-
91.
Udehn L. The Changing Face of Methodological Individualism.
Annu. Rev. Sociol. 2002,
28, 479–507.
[Google Scholar]
-
92.
Currie TE, Campenni M, Flitton A, Njagi T, Ontiri E, Perret C, et al. The cultural evolution and ecology of institutions.
Philos. Trans. R. Soc. B Biol. Sci. 2021,
376, 20200047.
[Google Scholar]
-
93.
Bernard HR. Social Research Methods: Qualitative and Quantitative Approaches; Sage Publications, Incorporated: Thousand Oaks, CA, USA, 2012.
-
94.
Henrich J, Heine SJ, Norenzayan A. Most people are not WEIRD.
Nature 2010,
466, 29.
[Google Scholar]
-
95.
Lee R, DeVore I. Man the Hunter; Aldine De Gruyter: New York, NY, USA, 1968.
-
96.
Mattison SM. Economic Impacts of Tourism and Erosion of the Visiting System Among the Mosuo of Lugu Lake.
Asia Pac. J. Anthropol. 2010,
11, 159–176.
[Google Scholar]
-
97.
Liu R, Cully SM. Does Kin-based Support Decrease under Market Transition?
-
98.
Colleran H. Market integration reduces kin density in women’s ego-networks in rural Poland.
Nat. Commun. 2020,
11, 266.
[Google Scholar]
-
99.
Brandl E, Emmott EH, Mace R. Adoption, Fostering, and Parental Absence in Vanuatu.
Hum. Nat. 2023,
34, 422–455.
[Google Scholar]
-
100.
Scelza B. Father’s presence speeds the social and reproductive careers of sons.
Curr. Anthropol. 2010,
1, 295–303.
[Google Scholar]
-
101.
Borgerhoff Mulder M, Caro T. The use of quantitative observational techniques in anthropology.
Curr. Anthropol. 1985,
26, 323–335.
[Google Scholar]
-
102.
Hames R. Time allocation. In Evolutionary Ecology and Human Behavior; Smith EA, Winterhalder B, Eds.; Water de Gruyter: Hawthorne, NY, USA, 1992; pp. 203–236.
-
103.
Kushnick G. Parental supply and offspring demand amongst Karo Batak mothers and children.
J. Biosoc. Sci. 2009,
41, 183.
[Google Scholar]
-
104.
Boone JL. Parental investment, social subordination and population processes among the 15th and 16th century Portuguese nobility. In Human Reproductive Behaviour: A Darwinian Perspective; Betzig LL, Mulder MB, Turke PW, Eds.; Cambridge University Press: Cambridge, UK, 1988; pp. 201–219.
-
105.
Josephson SC. Status, reproductive success, and marrying polygynously.
Ethol. Sociobiol. 1993,
14, 391–396.
[Google Scholar]
-
106.
McElreath R. Statistical Rethinking: A Bayesian Course with Examples in R and Stan; CRC Press: Boca Raton, FL, USA, 2016.
-
107.
Stulp G, Sear R, Barrett L. The Reproductive Ecology of Industrial Societies, Part I.
Hum. Nat. 2016,
27, 422–444.
[Google Scholar]
-
108.
Lawson D, Gibson MA. Polygynous marriage and child health in sub-Saharan Africa: What is the evidence for harm?
Demogr. Res. 2018,
39, 177–208.
[Google Scholar]
-
109.
Lawson DW, Mace R. Trade-offs in modern parenting: A longitudinal study of sibling competition for parental care.
Evol. Hum. Behav. 2009,
30, 170–183.
[Google Scholar]
-
110.
Ross CT, Winterhalder B. Sit-and-wait versus active-search hunting: A behavioral ecological model of optimal search mode.
J. Theor. Biol. 2015,
387, 76–87.
[Google Scholar]
-
111.
Barrett L, Dunbar R, Lycett J. Human Evolutionary Psychology; Princeton University Press: Princeton, NJ, USA, 2002.
-
112.
Fuentes A. Evolution of Human Behavior; Oxford University Press: Oxford, UK, 2009.
-
113.
Blackwell A. Human Biology. In Human Behavioral Ecology, 1st ed.; Koster JM, Scelza BA, Shenk MK, Eds.; Cambridge University Press: Cambridge, UK, 2024.
-
114.
Washburn SL, Lancaster CS. The evolution of hunting. In Man the Hunter; Lee RB, DeVore I, Eds.; Aldine: Chicago, IL, USA, 1968; pp. 293–303.
-
115.
Kaplan H. A theory of fertility and parental investment in traditional and modern human societies.
Am. J. Phys. Anthropol. 1996,
39, 91–135.
[Google Scholar]
-
116.
Aiello LC, Wheeler P. The Expensive-Tissue Hypothesis: The Brain and the Digestive System in Human and Primate Evolution.
Curr. Anthropol. 1995,
36, 199–221.
[Google Scholar]
-
117.
Hawkes K, O’Connell JF, Jones NGB. Grandmothering, menopause and the evolution of human life histories.
Proc. Natl. Acad. Sci. USA 1998,
95, 1336–1339.
[Google Scholar]
-
118.
Alger I, Dridi S, Stieglitz J, Wilson ML. The evolution of early hominin food production and sharing.
Proc. Natl. Acad. Sci. USA 2023,
120, e2218096120.
[Google Scholar]
-
119.
Sear R, Mace R. Who keeps children alive? A review of the effects of kin on child survival.
Evol. Hum. Behav. 2008,
29, 1–18.
[Google Scholar]
-
120.
Mace R, Sear R. Are humans cooperative breeders? In Grandmotherhood : the evolutionary significance of the second half of female life (143-159), Voland E, Chasiotis A, Schiefenhövel W, Eds.; Rutgers University Press: New Brunswick, NJ, USA, 2005.
-
121.
Kramer KL, Ellison PT. Pooled energy budgets: resituating human energy allocation trade-offs.
Evol. Anthropol. 2010,
19, 136–147.
[Google Scholar]
-
122.
Mattison SM. Male-provisioning hypothesis. In Encyclopedia of Evolutionary Psychological Science; Weekes-Shackelford V, Shackelford TK, Weekes-Shackelford VA, Eds.; Springer International Publishing: Cham, Switzerland, 2016; pp. 1–6.
-
123.
Page AE, Ringen EJ, Koster J, Borgerhoff Mulder M, Kramer K, Shenk MK, et al. Women’s subsistence strategies predict fertility across cultures, but context matters.
Proc. Natl. Acad. Sci. USA 2024,
121, e2318181121.
[Google Scholar]
-
124.
Mattison SM, MacLaren N, Sum C-Y, Mattison PM, Liu R, Shenk MK, et al. Market integration, income inequality, and kinship system among the Mosuo of China.
Evol. Hum. Sci. 2023,
5, e4.
[Google Scholar]
-
125.
Mattison SM, Mattison PM, Beheim BA, Liu R, Blumenfield T, Sum CY, et al. Gender disparities in material and educational resources differ by kinship system.
Philos. Trans. R. Soc. B Biol. Sci. 2023,
378, 20220299.
[Google Scholar]
-
126.
Mace R. Biased parental investment and reproductive success in Gabbra pastoralists.
Behav. Ecol. Sociobiol. 1996,
38, 75–81.
[Google Scholar]
-
127.
Mattison SM, Beheim B, Chak B, Buston P. Offspring sex preferences among patrilineal and matrilineal Mosuo in Southwest China revealed by differences in parity progression.
R. Soc. Open Sci. 2016,
3, 160526.
[Google Scholar]
-
128.
Martin M, de la Mora AN, Valeggia C, Veile A. Can women hunt? Yes. Did women contribute much to human evolution through endurance hunting? Probably not.
Am. Anthropol. 2024,
126, 365–369.
[Google Scholar]
-
129.
Mattison SM, Smith EA, Shenk MK, Cochrane EE. The evolution of inequality.
Evol. Anthropol. 2016,
25, 184–199.
[Google Scholar]
-
130.
Jones JH, Ready E, Pisor AC. Want climate-change adaptation? Evolutionary theory can help.
Am. J. Hum. Biol. 2020,
33, e23539.
[Google Scholar]
-
131.
Kennett DJ. The Island Chumash: Behavioral Ecology of a Maritime Society; University of California Press: Berkeley, CA, USA, 2005.
-
132.
Winterhalder BP. Foraging Strategy Adaptations of the Boreal Forest Cree: An Evaluation of Theory and Models from Evolutionary Ecology; Cornell University: Ithaca, NY, USA, 1977.
-
133.
Winterhalder B. Diet choice, risk, and food sharing in a stochastic environment.
J. Anthropol. Archaeol. 1986,
5, 369–392.
[Google Scholar]
-
134.
Codding BF, Bird DW. Behavioral ecology and the future of archaeological science.
J. Archaeol. Sci. 2015,
56, 9–20.
[Google Scholar]
-
135.
Hildebrandt WR, Jones TL. Evolution of marine mammal hunting: A view from the California and Oregon coasts.
J. Anthropol. Archaeol. 1992,
11, 360–401.
[Google Scholar]
-
136.
Broughton JM. Resource Depression and Intensification during the Late Holocene, San Francisco Bay: Evidence from the Emeryville Shellmound Vertebrate Fauna; University of Washington: Seattle, WA, USA, 1995.
-
137.
Fretwell S, Lucas H. On the territorial behaviour and other factors influencing habitat distribution in birds.
I Theor. Dev. Acta Biotheor 1970,
19, 16–36. doi:10.1007.BF01601953.
[Google Scholar]
-
138.
Kennett D, Anderson A, Winterhalder B. The Ideal Free Distribution, Food Production, and the Colonization of Oceania; In Behavioral Ecology and the Transition to Agriculture; Report #: 12. 2006. Available online https://escholarship.org/uc/item/5fj6m74d (accessed on 1 January 2024).
-
139.
Iriarte J, Ziegler MJ, Outram AK, Robinson M, Roberts P, Aceituno FJ, et al. Ice Age megafauna rock art in the Colombian Amazon?
Philos. Trans. R. Soc. B 2022,
377, 20200496.
[Google Scholar]
-
140.
Allee WC, Park O, Emerson AE, Park T, Schmidt KP. Principles of Animal Ecology; Philadelphia, Saunders Co: Philadelphia, PA, USA, 1949.
-
141.
Sutherland WJ. From Individual Behaviour to Population Ecology; Oxford University Press: New York, NY, USA, 1996; Volume 11.
-
142.
Winterhalder B, Kennett DJ, Grote MN, Bartruff J. Ideal free settlement of California’s Northern Channel Islands.
J. Anthropol. Archaeol. 2010,
29, 469–490.
[Google Scholar]
-
143.
Weitzel EM, Codding BF. The ideal distribution model and archaeological settlement patterning.
Environ. Archaeol. 2022,
27, 349–356.
[Google Scholar]
-
144.
Summers K. The evolutionary ecology of despotism.
Evol. Hum. Behav. 2005,
26, 106–135.
[Google Scholar]
-
145.
Smith EA, Mattison SM, DiNapoli R. Institutionalized inequality in small-scale societies. In Institutional Dynamics and Organizational Complexity: How Social Rules Have Shaped the Evolution of Human Societies Throughout Human History; Open Access Book; Cultural Evolution Society: 2023.
-
146.
Prufer KM, Thompson AE, Meredith CR, Culleton BJ, Jordan JM, Ebert CE, et al. The Classic Period Maya transition from an ideal free to an ideal despotic settlement system at the polity of Uxbenká.
J. Anthropol. Archaeol. 2017,
45, 53–68.
[Google Scholar]
-
147.
D’Altroy TN, Earle TK, Browman DL, La Lone D, Moseley ME, Murra JV, et al. Staple Finance, Wealth Finance, and Storage in the Inka Political Economy [and Comments and Reply].
Curr. Anthropol. 1985,
26, 187–206.
[Google Scholar]
-
148.
Wittfogel KA. Oriental despotism: A comparative study of total power.
Sci. Soc. 1959,
23, 58–65.
[Google Scholar]
-
149.
McClure SB, Kennett DJ. Niche Construction and the Ideal Free Distribution: Partners in Characterizing Past Human-Environmental Dynamics. In Cultural Landscapes and Long-Term Human Ecology; Springer: Cham, Switzerland, 2024; pp. 147–164.
-
150.
Jazwa CS, Jazwa KA. Resource Competition and Settlement Distribution in Bronze Age Greece.
Hum. Ecol. 2022,
50, 399–418.
[Google Scholar]
-
151.
Bell AV, Winterhalder B. The Population Ecology of Despotism.
Hum. Nat. 2014,
25, 121–135.
[Google Scholar]
-
152.
Lee CT, Puleston CO, Tuljapurkar S. Population and prehistory III: Food-dependent demography in variable environments.
Theor. Popul. Biol. 2009,
76, 179–188.
[Google Scholar]
-
153.
Puleston CO, Tuljapurkar S. Population and prehistory II: Space-limited human populations in constant environments.
Theor. Popul. Biol. 2008,
74, 147–160.
[Google Scholar]
-
154.
Puleston C, Tuljapurkar S, Winterhalder B. The invisible cliff: abrupt imposition of Malthusian equilibrium in a natural-fertility, agrarian society.
PLoS ONE 2014,
9, e87541.
[Google Scholar]
-
155.
Shennan S, Sear R. Archaeology, demography and life history theory together can help us explain past and present population patterns.
Philos. Trans. R. Soc. B 2021,
376, 20190711.
[Google Scholar]
-
156.
Urassa M, Lawson DW, Wamoyi J, Gurmu E, Gibson MA, Madhivanan P, et al. Cross-cultural research must prioritize equitable collaboration.
Nat. Hum. Behav. 2021,
5, 668–671.
[Google Scholar]
-
157.
Ji T, Wu J-J, He Q-Q, Xu J-J, Mace R, Tao Y. Reproductive competition between females in the matrilineal Mosuo of southwestern China.
Philos. Trans. R. Soc. B Biol. Sci. 2013,
368, 20130081.
[Google Scholar]
-
158.
Mattison SM. Demystifying the Mosuo: The behavioral ecology of kinship and reproduction of China’s “last matriarchal society”. Ph.D. Dissertation Thesis, University of Washington, Seattle, WA, USA, 2010.
-
159.
Shih C-K. Quest for Harmony: The Moso Traditions of Sexual Union & Family Life; Stanford University Press: Stanford, CA, USA, 2010.
-
160.
Sum C-Y, Blumenfield T, Cully SM. Beyond romantic partnerships: Sese, gender egalitarianism, and kinship diversity in Mosuo society. In Gender in Cross-Cultural Perspective; Routledge: London, UK, 2024; pp. 175–190.
-
161.
Walsh ER. From Nü Guo to Nü’er Guo: Negotiating Desire in the Land of the Mosuo.
Mod. China 2005,
31, 448–486.
[Google Scholar]
-
162.
Walsh E. A Society without Fathers or Husbands: the Na of China (Review).
Am. Ethnol. 2008,
29, 1043–1045.
[Google Scholar]
-
163.
Yong JC, Li NP. Elucidating evolutionary principles with the traditional Mosuo: Adaptive benefits and origins of matriliny and “walking marriages.”
Cult. Evol. 2022,
19, 22–40.
[Google Scholar]
-
164.
Harrell S. Ways of Being Ethnic in Southwest China; University of Washington Press: Seattle, WA. USA, 2001.
-
165.
Mattison SM, Sum C-Y, Reynolds AZ, Baca, Gabrielle D, Blumenfield T, Niedbalski S, et al.
Using evolutionary theory to hypothesize a transition from patriliny to matriliny and back again among the ethnic Mosuo of Southwest China. Matrix A J. Matricul. Stud. 2021,
2, 90–117.
[Google Scholar]
-
166.
Mattison SM, Hare D, MacLaren NG, Reynolds AZ, Sum C-Y, Liu R, et al. Context Specificity of “Market Integration” among the Matrilineal Mosuo of Southwest China.
Curr. Anthropol. 2022,
63, 118–124.
[Google Scholar]
-
167.
Reynolds AZ, Wander K, Sum C-Y, Su M, Emery Thompson M, Hooper PL, et al. Matriliny reverses gender disparities in inflammation and hypertension among the Mosuo of China.
Proc. Natl. Acad. Sci. USA 2020,
117, 30324–30327.
[Google Scholar]
-
168.
Mattison SM, MacLaren NG, Liu R, Reynolds AZ, Baca GD, Mattison PM, et al. Gender Differences in Social Networks Based on Prevailing Kinship Norms in the Mosuo of China.
Soc. Sci. 2021,
10, 253.
[Google Scholar]
-
169.
Mattison SM, Scelza B, Blumenfield T. Paternal investment and the positive effects of fathers among the matrilineal Mosuo of Southwest China.
Am. Anthropol. 2014,
116, 591–610.
[Google Scholar]
-
170.
Mattison SM, Quinlan RJ, Hare D. The expendable male hypothesis.
Philos. Trans. R. Soc. B Biol. Sci. 2019,
374, 20180080.
[Google Scholar]
-
171.
Gong B, Yan H, Yang C-L. Gender differences in the dictator experiment: evidence from the matrilineal Mosuo and the patriarchal Yi.
Exp. Econ. 2015,
18, 302–313.
[Google Scholar]
-
172.
Gong B, Yang C-L. Gender differences in risk attitudes: Field experiments on the matrilineal Mosuo and the patriarchal Yi.
J. Econ. Behav. Organ. 2012,
83, 59–65.
[Google Scholar]
-
173.
Wang J, Endo T. Cyclical processing of maternal lineage and parent–child dynamics: the dual family relationship system in the matrilineal Mosuo society.
Curr. Psychol. 2025,
44, 1793–1804.
[Google Scholar]
-
174.
Xiao E, Jin J, Hong Z, Zhang J. The Relationship Between Children and Their Maternal Uncles: A Unique Parenting Mode in Mosuo Culture.
Front. Psychol. 2022,
13, 873137.
[Google Scholar]
-
175.
Thomas MG, Ji T, Wu J, He Q, Tao Y, Mace R. Kinship underlies costly cooperation in Mosuo villages.
R. Soc. Open Sci. 2018,
5, 171535.
[Google Scholar]
-
176.
Shih C-K, Jenike MR. A cultural-historical perspective on the depressed fertility among the matrilineal Moso in Southwest China.
Hum. Ecol. 2002,
30, 21–47.
[Google Scholar]
-
177.
Kolodny O, Mattison SM, Creanza N. An Economic Solution for the Matrilineal Puzzle; Working Paper; 2019.
-
178.
He Q-Q, Rui J-W, Zhang L, Tao Y, Wu J-J, Mace R, et al. Communal breeding by women is associated with lower investment from husbands.
Evol. Hum. Sci. 2022,
4, e50.
[Google Scholar]
-
179.
Wu J-J, Ji T, He Q-Q, Du J, Mace R. Cooperation is related to dispersal patterns in Sino-Tibetan populations.
Nat. Commun. 2015,
6, 8693.
[Google Scholar]
-
180.
Chen Y, Ge E, Zhou L, Du J, Mace R. Sex inequality driven by dispersal.
Curr. Biol. 2023,
33, 464–473.
[Google Scholar]
-
181.
Shih C. Tisese and its anthropological significance: issues around the visiting sexual system among the Moso.
L’homme 2000,
154–155, 697–712.
[Google Scholar]
-
182.
Ji T, Zhang H, Pagel M, Mace R. A phylogenetic analysis of dispersal norms, descent and subsistence in Sino-Tibetans.
Evol. Hum. Behav. 2022,
43, 147–154.
[Google Scholar]
-
183.
Micheletti AJC, Mace R. Sibling competition and dispersal drive sex differences in religious celibacy.
Evol. Hum. Behav. 2024,
45, 144–152.
[Google Scholar]
-
184.
Micheletti AJC, Ge E, Zhou L, Chen Y, Zhang H, Du J, et al. Religious celibacy brings inclusive fitness benefits.
Proc. R. Soc. B Biol. Sci. 2022,
289, 20220965.
[Google Scholar]
-
185.
Du J, Huang Y, Bai P-P, Zhou L, Myers S, Page AE, et al. Post-marital residence patterns and the timing of reproduction: evidence from a matrilineal society.
Proc. R. Soc. B Biol. Sci. 2023,
290, 20230159.
[Google Scholar]
-
186.
Du J, Mace R. Marriage stability in a pastoralist society.
Behav. Ecol. 2019,
30, 1567–1574.
[Google Scholar]
-
187.
Du J, Page AE, Mace R. Grandpaternal care and child survival in a pastoralist society in western China.
Evol. Hum. Behav. 2022,
43, 358–366.
[Google Scholar]
-
188.
Huang Y, Bai P, Zhou L, Mace R, Du J. A rapid decline in gender bias relates to changes in subsistence practices over demographic changes in a formerly matrilineal community.
iScience 2025,
28, 111926.
[Google Scholar]
-
189.
Morais DB, Yarnal C, And ED, Dowler L. The impact of ethnic tourism on gender roles: A comparison between the Bai and the Mosuo of Yunnan province, PRC.
Asia Pac. J. Tour. Res. 2005,
10, 361–367.
[Google Scholar]
-
190.
Wang J, Lin Y, Gao L, Li X, He C, Ran M, et al. Menopause-Related Symptoms and Influencing Factors in Mosuo, Yi, and Han Middle-Aged Women in China.
Front. Psychol. 2022,
13, 763596.
[Google Scholar]
-
191.
Zhang Y, Zhao X, Leonhart R, Nadig M, Wang J, Zhao Y, et al. A Cross-Cultural Comparison of Climacteric Symptoms, Health-Seeking Behavior, and Attitudes towards Menopause Among Mosuo Women and Han Chinese Women in Yunnan, China.
Transcult Psychiatry 2019,
56, 287–301.
[Google Scholar]
-
192.
He Q, Pan Y, Sarangi S. Lineage-based heterogeneity and cooperative behavior in rural China.
J. Comp. Econ. 2018,
46, 248–269.
[Google Scholar]
-
193.
Hong X, Zhu W, Luo L. Non-parental care arrangements, parenting stress, and demand for infant-toddler care in China: Evidence from a national survey.
Front. Psychol. 2022,
12, 822104.
[Google Scholar]
-
194.
Ji C, Jin S, Wang H, Ye C. Estimating effects of cooperative membership on farmers’ safe production behaviors: Evidence from pig sector in China.
Food Policy 2019,
83, 231–245.
[Google Scholar]
-
195.
Luo L, Zuo Y, Xiong X. Discriminative Grandparental Investment in China.
Hum. Nat. 2024,
35, 21–42.
[Google Scholar]
-
196.
Schooling CM, Jiang C, Zhang W, Lam TH, Cheng KK, Leung GM. Size Does Matter: Adolescent Build and Male Reproductive Success in the Guangzhou Biobank Cohort Study.
Ann. Epidemiol. 2011,
21, 56–60.
[Google Scholar]
-
197.
Tsang EY, Lee PK. Raising Successful Offspring by Chinese Middle-Class Parents: A Sociocultural Approach to the Study of Class Reproduction in Urban China.
Asian J. Soc. Sci. 2016,
44, 165–186.
[Google Scholar]
-
198.
Zhang L, Liu J, Lummaa V. Intention to have a second child, family support and actual fertility behavior in current China: An evolutionary perspective.
Am. J. Hum. Biol. 2022,
34, e23669.
[Google Scholar]
-
199.
Yi M, Gao X, Bettinger R. Hunter-Gatherer Foraging Models: Review and Prospect in Paleolithic Archaeology.
Acta Anthropol. Sin. 2013,
32, 156–166.
[Google Scholar]
-
200.
Liu R, Chen S. Reflection and Prospective on the Application of Costly Signaling Theory in Archaeology of China.
Archaeol. North. Ethn. 2024, 16, 191–205.
[Google Scholar]
-
201.
Liu R, Zou G. Transmitted costly signaling theory (TCS): An evolutionary perspective of inequality transmission at Liangzhu culture sites, southern coastal China. 2023 Available online: https://www.researchgate.net/publication/372867837_Transmitted_costly_signaling_theory_TCS_an_evolutionary_perspective_of_inequality_transmission_at_Liangzhu_culture_sites_southern_coastal_China (accessed on 1 January 2024).
-
202.
Liu R, Liu H, Chen S. Alternative Adaptation Strategy during the Paleolithic–Neolithic Transition: Potential Use of Aquatic Resources in the Western Middle Yangtze Valley, China.
Quaternary 2020,
3, 28.
[Google Scholar]
-
203.
Kelly RL. New Discoveries and Theoretical Implications for the Last Foraging and First Farming in East Asia.
Quaternary 2021,
4, 40.
[Google Scholar]
-
204.
Yu P-L. Modeling Incipient Use of Neolithic Cultigens by Taiwanese Foragers: Perspectives from Niche Variation Theory, the Prey Choice Model, and the Ideal Free Distribution.
Quaternary 2020,
3, 26.
[Google Scholar]
-
205.
Yu P-L. Tempo and Mode of Neolithic Crop Adoption by Palaeolithic Hunter-Gatherers of Taiwan: Ethno-Archaeological and Behavioural Ecology Perspectives. In Hunter-Gatherers in Asia: From Prehistory to the Present; National Museum of Ethnology: Suita, Japan, 2021.
-
206.
Platt JR. Strong inference.
Science 1964,
146, 347–353.
[Google Scholar]
-
207.
Gould SJ, Lewontin RC. The spandrels of San Marco and the Panglossian paradigm: A critique of the adaptationist programme.
Proc. R. Soc. Lond. B Biol. Sci. 1979,
205, 581–598.
[Google Scholar]
-
208.
Reeve HK, Sherman PW. Adaptation and the goals of evolutionary research.
Q. Rev. Biol. 1993,
68, 1–32.
[Google Scholar]
-
209.
Blurton Jones N. Demography and Evolutionary Ecology of Hadza Hunter-Gatherers; Cambridge University Press: Cambridge, UK, 2016.
-
210.
Hruschka DJ, Munira S, Jesmin K, Hackman J, Tiokhin L. Learning from failures of protocol in cross-cultural research.
Proc. Natl. Acad. Sci. USA 2018,
115, 11428–11434.
[Google Scholar]
-
211.
Hudson M, Garrison NA, Sterling R, Caron NR, Fox K, Yracheta J, et al. Rights, interests and expectations: Indigenous perspectives on unrestricted access to genomic data.
Nat. Rev. Genet. 2020,
21, 377–384.
[Google Scholar]
-
212.
Shenk MK, Towner MC, Kress HC, Alam N. A model comparison approach shows stronger support for economic models of fertility decline.
Proc. Natl. Acad. Sci. USA 2013,
110, 8045–8050.
[Google Scholar]
-
213.
Bliege Bird R, Bird DW. Why Women Hunt: Risk and Contemporary Foraging in a Western Desert Aboriginal Community.
Curr. Anthropol. 2008,
49, 655–693.
[Google Scholar]
-
214.
Cronk L, Aktipis A. Design principles for risk-pooling systems.
Nat. Hum. Behav. 2021,
5, 825–833.
[Google Scholar]
-
215.
Glowacki L, Lew-Levy S. How small-scale societies achieve large-scale cooperation.
Curr. Opin. Psychol. 2022,
44, 44–48.
[Google Scholar]
-
216.
Gurven M, Jaeggi AV, von Rueden C, Hooper PL, Kaplan H. Does Market Integration Buffer Risk, Erode Traditional Sharing Practices and Increase Inequality? A Test among Bolivian Forager-Farmers.
Hum. Ecol. 2015,
43, 515–530.
[Google Scholar]
-
217.
Pisor AC, Jones JH. Do people manage climate risk through long-distance relationships?
Am. J. Hum. Biol. 2020,
33, e23525.
[Google Scholar]
-
218.
Symons D. On the Use and Misuse of Darwinism in the Study of Human Behavior; Oxford University Press: Oxford, UK, 1992.
-
219.
Chiaroni J, Underhill PA, Cavalli-Sforza LL. Y chromosome diversity, human expansion, drift, and cultural evolution.
Proc. Natl. Acad. Sci. USA 2009,
106, 20174–20179.
[Google Scholar]
-
220.
Rogers AR. Does biology constrain culture?
Am. Anthropol. 1988,
90, 819–831.
[Google Scholar]
-
221.
Sherman PW. The levels of analysis.
Anim. Behav. 1988,
36, 616–619.
[Google Scholar]
-
222.
Tinbergen N. On aims and methods of ethology.
Zeitschrift für Tierpsychologie 1963,
20, 410–433.
[Google Scholar]
-
223.
Bergman TJ, Beehner JC. Leveling with Tinbergen: Four levels simplified to causes and consequences.
Evol. Anthropol. Issues News Rev. 2021,
31, 12–19.
[Google Scholar]
-
224.
Aberle D. Matrilineal descent in cross-cultural perspective. In Matrilineal Kinship; Schneider DM, Gough K, Eds.; University of California Press: Berkeley, CA, USA, 1961; pp. 655–727.
-
225.
Surowiec A, Snyder K, Creanza N. A worldwide view of matriliny: Using genetic data to shed light on the evolution of human kinship systems.
Philos. Trans. R. Soc. B Biol. Sci. 2019,
374, 20180077.
[Google Scholar]
-
226.
Holden CJ, Mace R. Spread of cattle led to the loss of matrilineal descent in Africa: A coevolutionary analysis.
Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2003,
270, 2425–2433.
[Google Scholar]
-
227.
Jordan FM, Gray RD, Greenhill SJ, Mace R. Matrilocal residence is ancestral in Austronesian societies.
Proc. R. Soc. Ser. B 2009,
276, 1957–1964.
[Google Scholar]
-
228.
Mesoudi A. Cultural selection and biased transformation: two dynamics of cultural evolution.
Philos. Trans. R. Soc. B 2021,
376, 20200053.
[Google Scholar]
-
229.
Wander K, Su M, Mattison PM, Sum C-Y, Witt CC, Shenk MK, et al. High-altitude adaptations mitigate risk for hypertension and diabetes-associated anemia.
Am. J. Phys. Anthropol. 2020,
172, 156–164.
[Google Scholar]
-
230.
Nesse RM, Stearns SC. The great opportunity: Evolutionary applications to medicine and public health.
Evol. Appl. 2008,
1, 28–48.
[Google Scholar]
-
231.
Jones JH, Bliege Bird R. The Marginal Valuation of Fertility.
Evol. Hum. Behav. 2014,
35, 65–71.
[Google Scholar]
-
232.
Goodman A, Koupil I, Lawson DW. Low fertility increases descendant socioeconomic position but reduces long-term fitness in a modern post-industrial society.
Proc. R. Soc. B Biol. Sci. 2012,
279, 4342–4351.
[Google Scholar]