1. Introduction
The Okunev culture, primarily located in South Siberia, dates back to Afanasievo and Andronovo cultures [
1]. The Okunev people had advanced animal husbandry, metallurgy, and unique burial practices and were known for their mysterious carvings and art. Scholars believe these cultural elements reflect a mysterious spiritual world and belief system [
2,
3]. Okunev cultural elements have been found in Central Asia, North China, and the Indus Valley Civilization, indicating its broad influence on Eurasian Bronze Age cultures [
3]. However, the development and spread of the Okunev people remain unclear.
Ancient DNA research has offered insights into the origin and evolution of the Okunev culture. The ancient Okunev people can be seen as a blend of the indigenous Mongoloid population in South Siberia and a certain proportion of Indo-European- related ancestral components [
4]. Their maternal lines include East Eurasian haplogroups such as A, A8, A8a, A8a1, A10, C5, C5c, D4, D4b2a2a, and West Eurasian ones like H, U5, H15b1, H1bv1, H6a, H6a1b, J [
5,
6]. Their paternal lines consist of Q-L330-YP761, R1b-Z2103, Q1b-L330-BZ180, Q-L940, Q-M25, N-Tat [
5]. Scholars consider the presence of paternal line R1b-Z2103 as an indicator of the influence of the earlier Afanasievo culture, while N-Tat reflects the contemporaneous Uralic-speaking population’s impact [
4,
5]. Thus, the diverse branches of Q-M242 can be seen as the primary paternal type of the ancient Okunev people, with Q-L330-YP761 being the dominant one.
This paper will use modern-human sequences to explore the origin and evolution of the ancient Okunev people’s primary paternal lineage (Q-M242). The main aims are threefold. First, to track the flow of this lineage in later ancient populations. Second, to analyze the evolution and migration of the Okunev people’s descendants. Third, to identify the present-day descendants of the ancient Okunev people.
2. Materials and Methods
Samples and sequencing: Blood or saliva samples were collected from unrelated healthy male individuals from Chinese populations. All participants provided signed consent prior to participating in the study. Before signing the informed consent form, the contents of the form were clearly explained to the participants in either Chinese or their native language (like the language of minority). The study and sample collection procedures were reviewed and approved by the Medical Ethics Committee of Fudan University Approval NO.15137, adhering to the ethical principles of the 2013 Declaration of Helsinki by the World Medical Association.
Genomic DNA was extracted using the DP-318 Kit 128 (Tiangen Biotechnology, Beijing, China), following the manufacturer’s protocol. Firstly, routine Y-SNP tests were conducted to determine the general haplogroup of all male samples. DNA extracted from twenty-five Q-L330-YP761 male individuals’ biological samples was sent for next-generation Y-chromosome sequencing using the Illumina HiSeq2000 platform (Illumina, San Diego, CA, USA). Besides, the genome and information of twenty-five ancient individuals of haplogroup Q-L330-YP761 were extracted from previous publications [
4,
6,
7,
8,
9,
10,
11]. Also, the variant situation of 10 haplogroup samples was adopted from publicly available resources, like previous literature [
12] and the website of the human phylogenetic tree https://www.yfull.com/tree/Q-Y18330/ (accessed on 10 March 2025).
Data availability statement: Following the regulations of the Human Genetic Resources Administration of China (HGRAC), the raw sequence data reported in this paper are available at the request of the corresponding author.
Data analysis: In total, we summarized a final dataset of 60 samples, including 25 newly generated sequences (from the research group’s internal database, unpublished data, these individuals were preliminarily classified as haplogroup Q-L330-YP761), 25 sequences of ancient individuals sourced from published literature across distinct archaeological sites (detailed in Supplementary Table S1, all categorized as Q-L330-YP761), and 10 modern individuals from public available resources (all belonging to Q-L330-YP761 haplogroup). Read mapping and SNP calling from next-generation sequencing data were conducted using standard procedures (BWA and SAMtools) and the human reference genome sequence hg38 [
13,
14]. The final SNP list of 25 newly generated sequences was used to construct a phylogenetic tree and age estimation analyses. Bayesian evolutionary analyses were conducted using BEAST (v.2.4.3) [
15]. To calculate divergence times in the phylogenetic tree, a point mutation rate of 0.74 × 10
−9 per site per year was applied [
16], which was very similar to the rate of 0.78 × 10
−9 per site per year reported in analyses of the ancient genome of the 45,000-year-old Ust’Ishim male [
17]. We refer to the protocol used in our previous publications for data processing and Bayesian evolutionary analyses [
18,
19,
20]. Detailed sample information can be found in Supplementary Table S1.
In the next step, we placed the ancient available modern individuals onto the revised phylogenetic tree according to the variants of ancient genomes. Due to the low coverage of genomes of some ancient samples, the mutation situation of some SNPs of some samples is unclear. Therefore, some samples may be placed on a root location when they actually belong to a downstream sub-branch.
3. Results
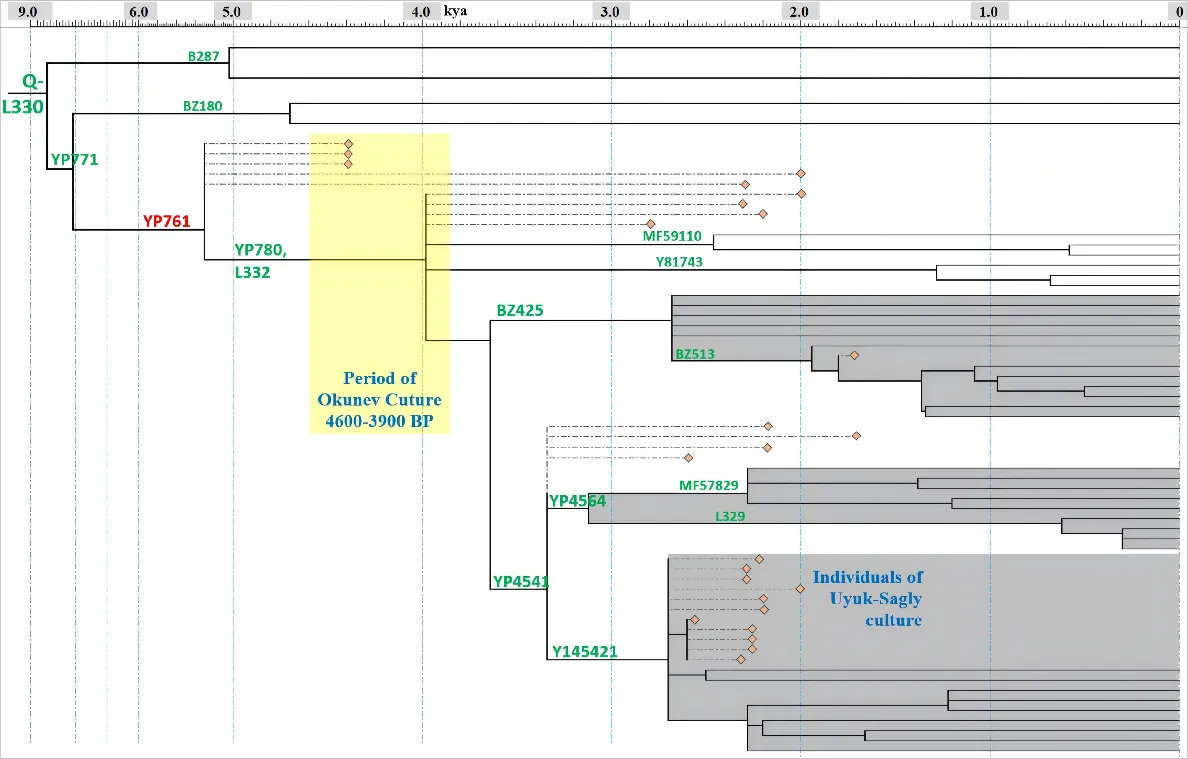
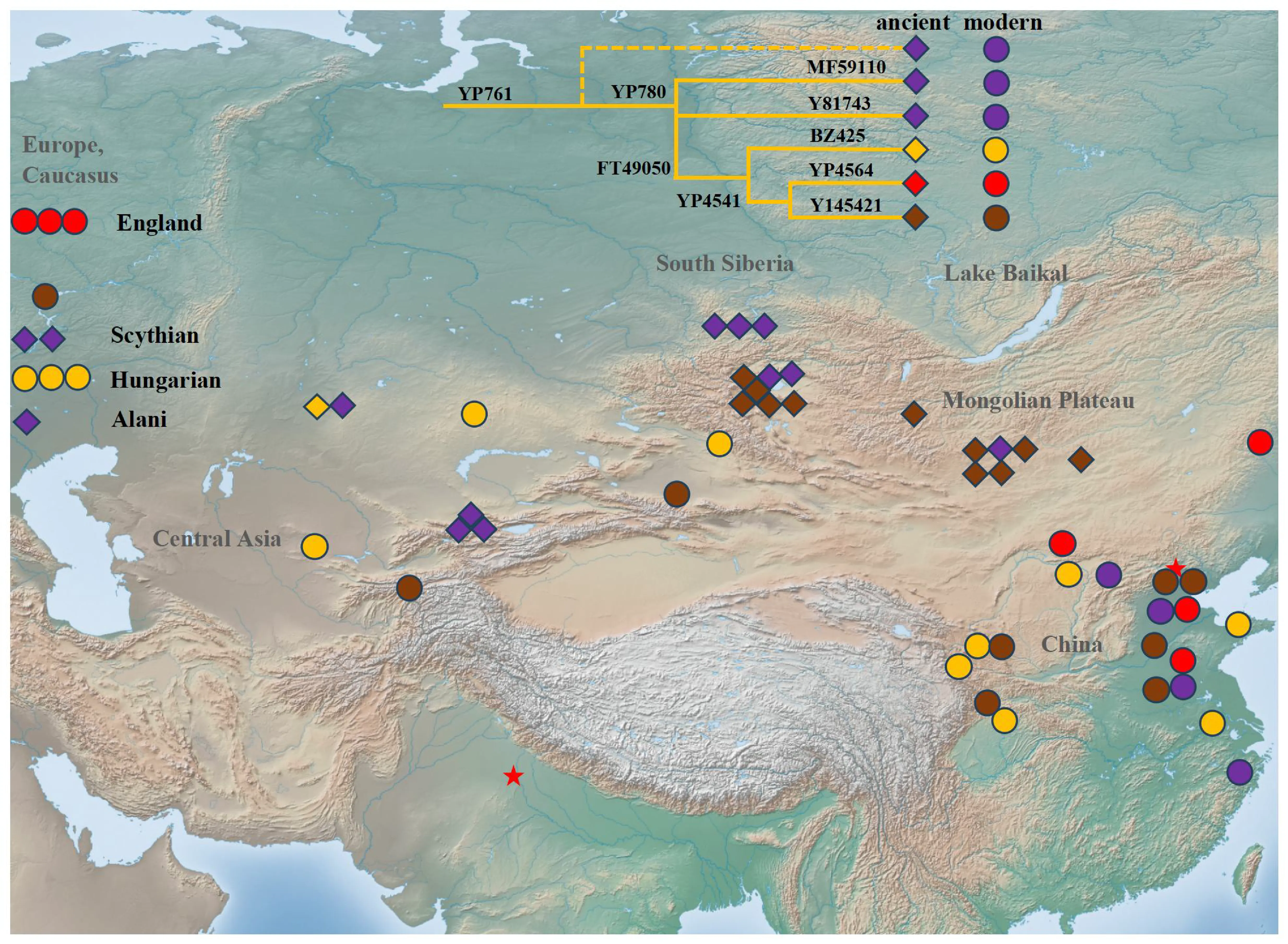
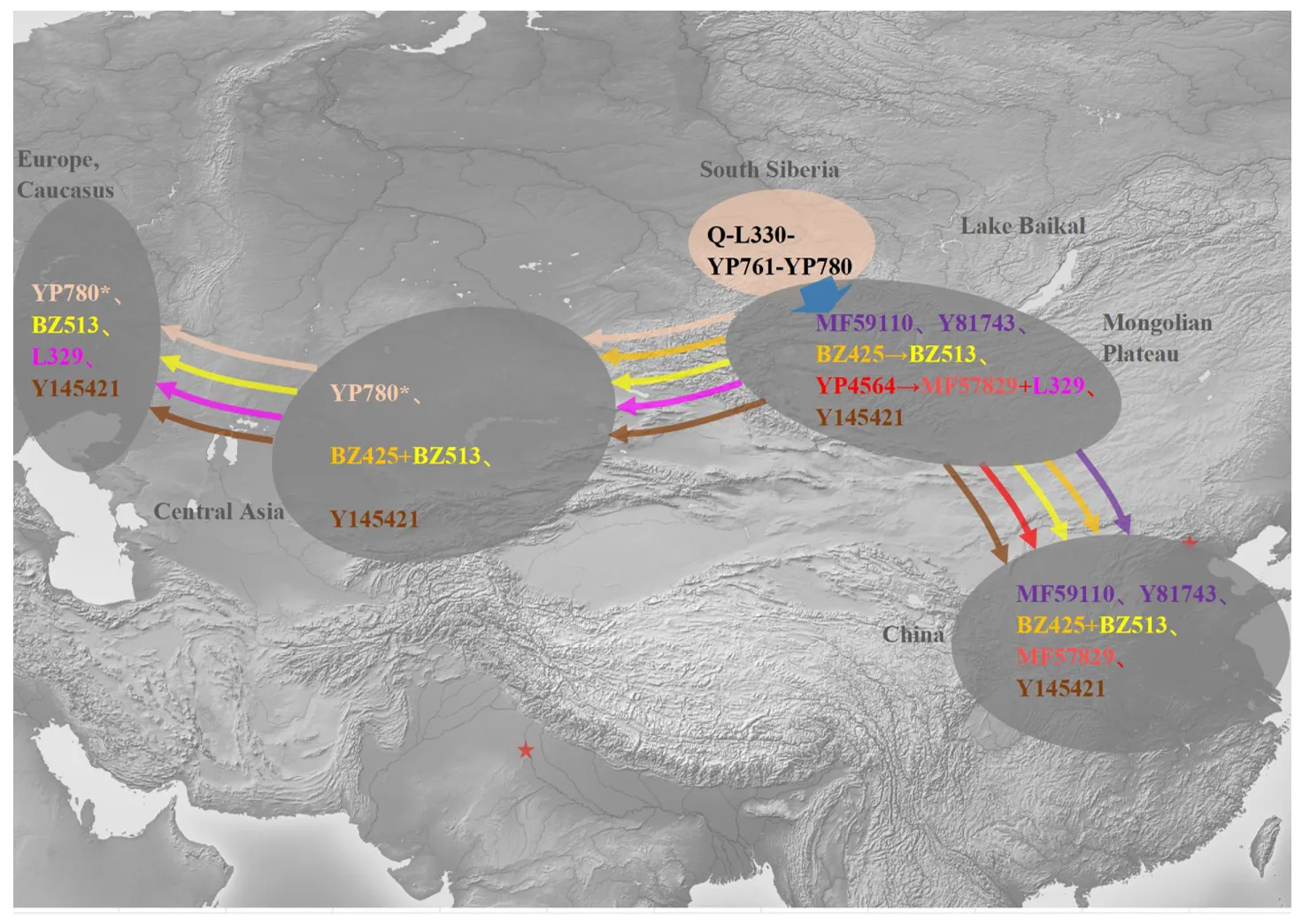
A schematic phylogenetic tree labeled ancient DNA is shown in . The detailed tree can be found in Supplementary Table S1. We presented clear distribution patterns for sub-branches of haplogroup Q-L330-YP761 across Eurasia (). The possible migration routes of sub-branches of haplogroup Q-L330-YP761 are shown in .
. Schematic diagram of revised phylogenetic tree of Q-L330-YP761. The paternal lineage tree of modern humans is depicted, with orange diamonds representing ancient samples. Dotted lines indicate their positions and corresponding ages on the tree.
. Schematic diagram of the location of Q-L330-YP761 individuals and the possible diffusion routes of major sub-branches. This legend illustrates a simplified paternal lineage tree of Q-L330-YP761, with distinct downstream lineages differentiated by color. Ancient samples are represented by diamonds, and modern samples by circles. To facilitate the identification of the spatial distribution of samples, we have marked the locations of Beijing and Delhi with red five-pointed stars and labeled some important geographical regions in grey text. The map includes most samples to show their distribution across Eurasia clearly. However, four groups lie outside the map’s range. They are labeled at its edge: three modern British individuals, three modern Hungarians, two ancient Scythian Culture individuals from Ukraine, and one Alan individual from Kyrgyzstan. Detailed information on all samples is provided in Supplementary Table S1.
Figure 3. Schematic diagram of the possible diffusion routes of major sub-branches of Q-L330-YP761. The figure shows the inferred migration routes of Q-L330-YP761 downstream lineages. Different colored arrows indicate possible migration routes of different lineages. To facilitate the identification of the spatial distribution of samples, we have marked the locations of Beijing and Delhi with red five-pointed stars and labeled some important geographical regions in grey text. In the figure, all haplotypes located within the same gray ellipse as YP780∗ are sub-lineages of YP780. Thus, YP780∗ denotes YP780 haplogroups excluding those in the grey ellipse.
The haplogroups provided by the original ancient DNA literature are general ones. Therefore, we re-analyzed the data and determined the most downstream sub-branch for ancient genomes (). Ancient individuals of the same haplogroup with Okunev samples were also summarized from public literature to trace the origin and descendants of Okunev lineages [
4,
5]. Haplogroup Q-L330-YP761 was dominant among remains from the core sites of Okunev culture. It was also the only paternal lineage found at the Tenisei Early Bronze Age site, which was proposed as the direct predecessor of Okunev culture. Therefore, we discussed this haplogroup in the next section.
.
list of revised Y-chromosome haplogroup of ancient DNA.
| ID |
Location |
Y
Haplogroup |
Mt
Haplogroup |
Year |
| RISE664 |
Russia |
Q-YP761 |
A8a1 |
4272 ybp |
| RISE662 |
Russia |
Q-YP761 |
H6a1b |
4063 ybp |
| RISE718 |
Russia |
Q-YP761 |
C5c |
4434 ybp |
| DA73 ERS2374338 |
Kyrgyzstan |
Q-YP761 |
G2a1 |
4000 ybp |
| UKR109 |
Ukraine |
Q-YP761 |
H5a1 |
2400 ybp |
| DA96 ERS2374351 |
Kyrgyzstan |
Q-L332 |
D4j5 |
2000 ybp |
| DA161 ERS2374384 |
Russia |
Q-L332 |
X2f |
2300 ybp |
| I6231 |
Mongolia |
Q-L332 |
F1b1b |
2300 ybp |
| IS2 |
Russia |
Q-L332 |
HV-CRS |
|
| DA221 ERS2374408 |
Kazakhstan |
Q-BZ425 |
J1b1a1 |
|
| ERR5211315 |
Mongolia |
Q-YP4547 |
|
2180 ybp |
| ALN003.A0101 ERS6072385 |
Kyrgyzstan |
Q-YP4547 |
U5a1g |
1700 ybp |
| I6230 |
Mongolia |
Q-YP4547 |
U2e1h |
2200 ybp |
| UKR113 |
Ukraine |
Q-YP4547 |
HV0a |
2600 ybp |
| ERR5211347 |
Mongolia |
Q-Y145452 |
|
2200 ybp |
| CHN003 |
Mongolia |
Q-Y145452 |
U4a1 |
2300 ybp |
| CHN007 |
Mongolia |
Q-Y145452 |
U2e1h |
Mongolia |
| DEL001 |
Mongolia |
Q-Y145452 |
D4j11 |
2000 ybp |
| SKT008 |
Mongolia |
Q-Y145452 |
Hz |
|
| I7029 |
Mongolia |
Q-Y145452 |
U4a1 |
2300 ybp |
| CHN016 |
Mongolia |
Q-FTA37701 |
A8a |
|
| ERR5211222 |
Mongolia |
Q-TY520082 |
|
2250 ybp |
| ERR5211316 |
Mongolia |
Q-FTA37701 |
|
2200 ybp |
| I6232 ERR5211317 |
Mongolia |
Q-TY520082 |
|
2250 ybp |
| I12970 |
Mongolia |
Q-TY520082 |
A8a |
2300 ybp |
Haplogroup R1b-Z2103 was found in two samples (Kh7, RISE675) from the Uybat V site in Khakasia, South Siberia [
4,
5]. This haplogroup is one of the dominant paternal lineages of earlier Afanasievo people from the same region [
5,
21,
22]. Therefore, it is clear that this paternal lineage, along with those west Eurasian mtDNA lineages, are results of admixture with local Afanasievo ancestry.
Haplogroup N-Tat was found in three samples (Kh12, Kh13, and Kh15) from the Itkol’II site and the Uybat-Charkov site in Khakasia, South Siberia. This haplogroup is one of the founder paternal lineages of Uralic-speaking populations and Turkic-speaking populations [
21,
22]. The duration of the Okunev culture (2500–1700 BC) was roughly concurrent with the Seima-Turbino phenomenon e (2200–1900 BC), which underwent significant expansion in the northern part of Eurasia during this period [
23]. Therefore, this lineage among Okunev people is also likely a result of admixture with surrounding Uralic-speaking populations [
24,
25].
One individual of haplogroup Q1b-L330-BZ180 was found from Syda 5, Tumen site in Russia, South Siberia. The lineage was dominant among Early Bronze Age remains from the Cis-Baikal region and later in many sites across the Mongolian Plateau [
8,
11,
26,
27]. The appearance of this lineage among Okunev people may result from admixture. Four individuals related to Q-L940-Y2679 were found at the Verkhni Askiz site in Russia South Siberia [
4]. They may belong to an extinct or a minor root lineage.
One individual of Q-M25-L712-YP832 was found in two samples (Kh7, RISE675) from the Uybat V site in Khakasia, South Siberia [
4]. The earliest sample of this haplogroup, a 7500-year-old individual FRS002, was found in the Altai-Sayan region, South Siberia [
28]. The haplogroup has also been found in large numbers among ancient Karasuk people [
4], ancient Xiongnu [
10], ancient Türk people, and ancient Avar people in Hungary [
8]. This paternal lineage is likely flourishing among nomadic populations across the Eurasian Steppe.
In general, the paternal gene pool of ancient Okunev people was formed as a result of the rising of lineages (like Q-L330-YP761, Q-L940-Y2679, and Q-M25-L712-YP832) and admixture from surrounding people (like R1b-Z2103, N-Tat, and Q1b-L330-BZ180).
3.2. Demographic History of Sub-Branches of Q-L330-YP761, the Major Paternal Lineage of Okunev People
Previous studies have found many individuals of haplogroup Q-L330-YP761 from remains of Okunev culture and other archaeological cultures from the central part of the Eurasian Steppe at a later age (, , and Table S1). The Okunev individuals’ variants suggested they are the direct predecessor of downstream sub-branch Q-L330-YP761-YP780, L332, which experienced continuous expansion about 4000 years ago. Samples of two root sub-linages, Y81743 and MF59110, were found among Han populations.
The revised phylogenetic tree suggested that sub-branch BZ425 is like a paternal lineage of ancient Xiongnu and Türk people. Samples of sub-lineage BZ513 were found among individuals from Kazakhstan, Hungary, and Chinese populations. The age of BZ513 (~2000 years ago) surprisingly coincides with the westward migration of ancient Xiongnu tribes.
The East-West dichotomous structure was also found among the topology of sub-lineage YP4564. Sub-lineage L329 was exclusively found in Europe while sub-lineage MF57829 was only found in China until now. This paternal is also likely the result of long-distance migration across the Eurasian Steppe.
Many Y145421 individuals were found among remains from the Chandman site of Uyuk-Sagly culture (900–300 BC) and several sites of early Xiongnu in the same region. According to the studies of archaeology and history, the northwest part of Mongolia and South Siberia became parts of the Xiongnu Empire after the rising of the Xiongnu people. Two modern individuals of this lineage were found in Tajik and Russia, suggesting a trace of westward migration.
4. Discussions
4.1. High Genetic Diversity among Ancient and Modern Populations from Central Part of Eurasian Steppe
Current aDNA research shows that Eurasian steppe populations during the Copper and Bronze Ages had highly diverse genetic lineages. In the late Paleolithic and Neolithic, NEA populations from the middle and lower Amur River diffused westward to the Baikal region and the central and eastern Mongolian Plateau [
8,
11]. Ancestral Uralic-speaking populations rapidly moved through the northern Eurasian taiga [
29]. Between the Altai and Baikal, ANE populations differentiated rapidly, producing many genetic lineages and descendant groups [
28]. These population movements and mixtures were very complex. Existing aDNA research has gradually revealed this demographic history [
8,
11,
28,
29]. This is reflected in the paternal genetic structure by the emergence of dozens of downstream lineages of C-L1373, Q-M242, and N-Tat and their extensive spread across northern Eurasia and the Eurasian steppe [
18,
21,
22,
30,
31].
During the Copper and Bronze Age, the eastward expansion of Indo-European-related groups brought significant population movements and mixtures. West and East Eurasian lineages blended in Central Asia, shaping the region’s genetic makeup. Strong nomadic groups emerged in the eastern Eurasian steppe after the mid-Bronze Age (around 3500 BP), later establishing nomadic empires and shifting the population’s direction westward. This study’s detailed analysis of the ancient Okunev people’s paternal genetic structure supports the rise and spread of local lineages in the eastern Eurasian steppe, offering a fine-grained example of Eurasian steppe populations’ migration and genetic evolution.
4.2. Research Approaches to Population Changes among Grassland Populations
The accumulation of aDNA studies has made the genetic structure of ancient archaeological cultures clearer over space and time. Meanwhile, the genetic makeup of modern populations has also become more evident. Therefore, integrating ancient and modern DNA research could be an effective way to explore the population history of Eurasian steppe peoples since ancient times.
In this paper, we explore the formation of the Okunev culture and the spread of its people. Ancient Okunev genetic lineages mainly flowed into later groups like the Xiongnu, Tiele, modern Chinese, and central-west Eurasians. Our high-resolution update of the Q-L330-YP761 phylogeography reveals multiple east-west differentiation patterns, strongly supporting the substantial westward migration of some genetic lineages from the eastern Eurasian steppe. However, ancient DNA data from northern China and northern South Asia are still scarce, and no Okunev-related genetic lineages have been identified in these regions so far. It remains unclear whether the influence of Okunev culture on these two regions originated from the spread of cultural factors or population movements. It remains unclear whether the influence of Okunev culture on these two regions is due to the spread of cultural factors or population mobility.
5. Conclusions
This research presents the largest dataset of ancient and modern sequences of the patrilineal haplogroup Q-L330-YP761 to date, the most revised phylogeny, and related evolutionary background information. The analysis in this paper shows that the patrilineal haplogroup Q-L330-YP761 originated in South Siberia, supporting the idea that the ancient Okunev people were direct descendants of the Upper Paleolithic Ancient Northern Eurasians (ANE). In subsequent stages, populations with diverse sub-branches of haplogroup Q-L330-YP761 as their main patrilineage were mainly active on the Mongolian Plateau, and they were likely part of the ancient Xiongnu, Turkic, and Uyghur peoples. From here, some of them spread to North China and eventually integrated into the Chinese population. Some other parts spread westward to Central Asia and Europe and participated in the formation of local populations in subsequent historical periods. The appearance of Okunev-like cultural elements in archaeological remains in the Mongolian Plateau, North China, India, Central Asia, and Eastern Europe may be related to the spread of the patrilineal haplogroup Q-L330-YP761. For the first time, we have provided a complete description of the origin and evolution of patrilineal haplogroup Q-L330-YP761 and its related ancient and modern populations over several thousand years.
In summary, this study comprehensively analyzed ancient and modern DNA data related to the paternal formation process of ancient Okunev people and their descendants. We provided a clear scenario for the rise of the Okunev people and the demographic history of the descendants’ lineages of the Okunev people in the late history period. This study’s data and analysis results will assist in further future research on the history of nomadic populations across the Eurasian Steppe.
Supplementary Materials
The following supporting information can be found at: https://www.sciepublish.com/article/pii/504, Supplementary Table S1. The detailed patrilineal phylogenetic tree of haplogroup Q-L330-YP761.
Acknowledgments
We thank all donors for providing DNA samples and/or DNA sequences.
Author Contributions
L.L.: Conceptualization, Methodology, Software, Formal analysis, Visualization, Writing—Original draft, Writing—Review & Editing. S.X.: Writing—Original draft, Validation, Investigation. H.L.: Data curation. L.W.: Data curation, Writing—Review & Editing, Supervision. H.Y.: Data curation, Writing—Review & Editing, Supervision, Project administration, Funding acquisition.
Ethics Statement
The study and sample collection procedures were reviewed and approved by the Medical Ethics Committee of Fudan University Approval NO.15137 (approved on 2 March 2015), adhering to the ethical principles of the 2013 Declaration of Helsinki by the World Medical Association.
Informed Consent Statement
Informed consent was obtained from all participants in the study.
Data Availability Statement
Following the regulations of the Human Genetic Resources Administration of China (HGRAC), the raw data reported in this paper are available upon reasonable request from the corresponding author.
Funding
This work was supported by the Annual Open Project of the Key Research Institute for Humanities and Social Sciences at Inner Universities of Inner Mongolia Autonomous Region (Grant No. KFSM-SDSK0305).
Declaration of Competing Interest
The authors report no conflicts of interest.
References
-
1.
Svyatko SV, Mallory JP, Murphy EM, Polyakov AV, Reimer PJ, Schulting RJ. New radiocarbon dates and a review of the chronology of prehistoric populations from the Minusinsk Basin, southern Siberia, Russia.
Radiocarbon 2009,
51, 243–273.
[Google Scholar]
-
2.
Outram AK, Polyakov A, Gromov A, Moiseyev V, Weber AW, Bazaliiskii VI, et al. Archaeological supplement B to Damgaard et al. 2018: Discussion of the archaeology of Central Asian and East Asian Neolithic to Bronze Age Hunter-Gatherers and Early Pastoralists, Including Consideration of Horse Domestication. 2018. Available online: https://ore.exeter.ac.uk/repository/handle/10871/32792 (accessed on 17 April 2025).
-
3.
Zhang C, Festa M. Review of Okunev Culture.
Res. China's Front. Archaeol. 2023,
01, 328–344. (In Chinese)
[Google Scholar]
-
4.
de Barros Damgaard P, Martiniano R, Kamm J, Moreno-Mayar JV, Kroonen G, Peyrot M, et al. The first horse herders and the impact of early Bronze Age steppe expansions into Asia.
Science 2018,
360, eaar7711.
[Google Scholar]
-
5.
Hollard C, Zvénigorosky V, Kovalev A, Kiryushin Y, Tishkin A, Lazaretov I, et al. New genetic evidence of affinities and discontinuities between bronze age Siberian populations.
Am. J. Phys. Anthropol. 2018,
167, 97–107.
[Google Scholar]
-
6.
Unterländer M, Palstra F, Lazaridis I, Pilipenko A, Hofmanová Z, Groß M, et al. Ancestry and demography and descendants of Iron Age nomads of the Eurasian Steppe.
Nat. Commun. 2017,
8, 14615.
[Google Scholar]
-
7.
Saag L, Utevska O, Zadnikov S, Shramko I, Gorbenko K, Bandrivskyi M, et al. North Pontic crossroads: Mobility in Ukraine from the Bronze Age to the early modern period.
Sci. Adv. 2025,
11, eadr0695.
[Google Scholar]
-
8.
Wang CC, Yeh HY, Popov AN, Zhang HQ, Matsumura H, Sirak K, et al. Genomic insights into the formation of human populations in East Asia.
Nature 2021,
591, 413–419.
[Google Scholar]
-
9.
Gnecchi-Ruscone GA, Khussainova E, Kahbatkyzy N, Musralina L, Spyrou MA, Bianco RA, et al. Ancient genomic time transect from the Central Asian Steppe unravels the history of the Scythians.
Sci. Adv. 2021,
7, eabe4414.
[Google Scholar]
-
10.
Damgaard PDB, Marchi N, Rasmussen S, Peyrot M, Renaud G, Korneliussen T, et al. 137 ancient human genomes from across the Eurasian steppes.
Nature 2018,
557, 369–374.
[Google Scholar]
-
11.
Jeong C, Wang K, Wilkin S, Taylor WTT, Miller BK, Bemmann JH, et al. A dynamic 6000-year genetic history of Eurasia’s Eastern Steppe.
Cell 2020,
183, 890–904.e29.
[Google Scholar]
-
12.
Zhabagin M, Wei LH, Sabitov Z, Ma PC, Sun J, Dyussenova Z, et al. Ancient components and recent expansion in the Eurasian heartland: insights into the revised phylogeny of Y-chromosomes from Central Asia.
Genes 2022,
13, 1776.
[Google Scholar]
-
13.
Li H, Durbin R. Fast and accurate long-read alignment with Burrows–Wheeler transform.
Bioinformatics 2010,
26, 589–595.
[Google Scholar]
-
14.
Li H, Durbin R. Fast and accurate short read alignment with Burrows–Wheeler transform.
Bioinformatics 2009,
25, 1754–1760.
[Google Scholar]
-
15.
Bouckaert R, Heled J, Kühnert D, Vaughan T, Wu CH, Xie D, et al. BEAST 2: A software platform for Bayesian evolutionary analysis.
PLoS Comput. Biol. 2014,
10, e1003537.
[Google Scholar]
-
16.
Karmin M, Saag L, Vicente M, Sayres MAW, Järve M, Talas UG, et al. A recent bottleneck of Y chromosome diversity coincides with a global change in culture.
Genome Res. 2015,
25, 459–466.
[Google Scholar]
-
17.
Fu Q, Li H, Moorjani P, Jay F, Slepchenko SM, Bondarev AA, et al. Genome sequence of a 45,000-year-old modern human from western Siberia.
Nature 2014,
514, 445–449.
[Google Scholar]
-
18.
Sun J, Ma PC, Cheng HZ, Wang CZ, Li YL, Cui YQ, et al. Post‐last glacial maximum expansion of Y‐chromosome haplogroup C2a‐L1373 in northern Asia and its implications for the origin of Native Americans.
Am. J. Phys. Anthropol. 2021,
174, 363–374.
[Google Scholar]
-
19.
Sun J, Li YX, Ma PC, Yan S, Cheng HZ, Fan ZQ, et al. Shared paternal ancestry of Han, Tai‐Kadai‐speaking, and Austronesian‐speaking populations as revealed by the high resolution phylogeny of O1a‐M119 and distribution of its sub‐lineages within China.
Am. J. Phys. Anthropol. 2021,
174, 686–700.
[Google Scholar]
-
20.
Sun J, Wei LH, Wang LX, Huang YZ, Yan S, Cheng HZ, et al. Paternal gene pool of Malays in Southeast Asia and its applications for the early expansion of Austronesians.
Am. J. Hum. Biol. 2021,
33, e23486.
[Google Scholar]
-
21.
Lan H. Y chromosome Clues to the East Asian Origins of Turki and Uralic populations.
Commun. Contemp. Anthropol. 2007,
1. doi:10.4236/COCA.2007.11016.
[Google Scholar]
-
22.
Rootsi S, Zhivotovsky LA, Baldovič M, Kayser M, Kutuev IA, Khusainova R, et al. A counter-clockwise northern route of the Y-chromosome haplogroup N from Southeast Asia towards Europe.
Eur. J. Hum. Genet. 2007,
15, 204–211.
[Google Scholar]
-
23.
Childebayeva A, Fricke F, Rohrlach AB, Huang L, Schiffels S, Vesakoski O, et al. Bronze age Northern Eurasian genetics in the context of development of metallurgy and Siberian ancestry.
Commun. Biol. 2024,
7, 723.
[Google Scholar]
-
24.
Grünthal R, Heyd V, Holopainen S, Janhunen JA, Khanina O, Miestamo M, et al. Drastic demographic events triggered the Uralic spread.
Diachronica 2022,
39, 490–524.
[Google Scholar]
-
25.
Asko P. Location of the Uralic proto-language in the Kama River Valley and the Uralic speakers’ Expansion east and west with the ‘Sejma-Turbino transcultural phenomenon’ 2200-1900 BC.
Архе. Евра. Степ. 2022,
2, 258–277.
[Google Scholar]
-
26.
Moussa NM, McKenzie HG, Bazaliiskii VI, Goriunova OI, Bamforth F, Weber AW. Insights into Lake Baikal's ancient populations based on genetic evidence from the Early Neolithic Shamanka II and Early Bronze Age Kurma XI cemeteries.
Archaeol. Res. Asia 2021,
25, 100238.
[Google Scholar]
-
27.
Hollard C, Keyser C, Giscard PH, Tsagaan T, Bayarkhuu N, Bemmann J, et al. Strong genetic admixture in the Altai at the Middle Bronze Age revealed by uniparental and ancestry informative markers.
Forensic Sci. Int. Genet. 2014,
12, 199–207.
[Google Scholar]
-
28.
Wang K, Yu H, Radzevičiūtė R, Kiryushin YF, Tishkin AA, Frolov YV, et al. Middle Holocene Siberian genomes reveal highly connected gene pools throughout North Asia.
Curr. Biol. 2023,
33, 423–433.e5.
[Google Scholar]
-
29.
Zeng TC, Vyazov LA, Kim A, Flegontov P, Sirak K, Maier R, et al. Postglacial genomes from foragers across Northern Eurasia reveal prehistoric mobility associated with the spread of the Uralic and Yeniseian languages.
Biorxiv 2023. doi:10.1101/2023.10.01.560332.
[Google Scholar]
-
30.
Huang YZ, Pamjav H, Flegontov P, Stenzl V, Wen SQ, Tong XZ, et al. Dispersals of the Siberian Y-chromosome haplogroup Q in Eurasia.
Mol. Genet. Genom. 2018,
293, 107–117.
[Google Scholar]
-
31.
Ilumäe AM, Post H, Flores R, Karmin M, Sahakyan H, Mondal M, et al. Phylogenetic history of patrilineages rare in northern and eastern Europe from large-scale re-sequencing of human Y-chromosomes.
Eur. J. Hum. Genet. 2021,
29, 1510–1519.
[Google Scholar]